Embed Size (px)
Citation preview

Estudo de abrigos para peixes a jusante de centrais hidroeléctricas
Modelação numérica
Rui Tiago Pessoa Zabelo Matos de Almeida
Dissertação para obtenção do Grau de Mestre em
Engenharia Civil
Orientador: Professor Doutor António Alberto do Nascimento Pinheiro
Orientador: Doutora Isabel Maria Bento de Matos Boavida
Júri
Presidente: Professor Doutor António Alexandre Trigo Teixeira
Orientador: Professor Doutor António Alberto do Nascimento Pinheiro
Vogal: Professor Doutor Jorge de Saldanha Gonçalves Matos
Vogal: Doutor José Maria Horta e Costa Silva Santos
Junho 2014


Às minhas avós


i
RESUMO
Quando uma central hidroeléctrica opera em regime de hydropeaking, o caudal natural do curso de
água é periodicamente alterado por descargas fortes, eventualmente de curta duração, resultantes da
produção de hidroelectricidade durante os períodos de consumo de ponta. Estes eventos criam
frequentemente um forte stress para os peixes, em todos os estágios de vida, interferindo assim com
o bem-estar das populações piscícolas e com as condições ecológicas do rio a jusante da central
hidroeléctrica. A criação de abrigos laterais para peixes tem sido apontada como uma das mais
promissoras medidas de mitigação para proteger as espécies piscícolas dos caudais de ponta. Desta
forma, é crucial determinar critérios de dimensionamento para os abrigos laterais, de forma a
aumentar a disponibilidade de habitat. Sendo cada vez mais aplicados em estudos de modelação de
habitat, os modelos hidrodinâmicos 2D são uma ferramenta poderosa para perceber as interacções
entre o escoamento do rio e os padrões hidráulicos numa escala de microhabitat. Quando acoplados
com um modelo biológico de habitat que represente os diferentes requisitos de habitat, estes modelos
permitem uma melhor percepção da influência dos caudais de ponta nos peixes, enquanto estimam a
abundância relativa de habitats. No presente estudo, são comparados resultados obtidos numa
instalação experimental com as simulações de habitat efectuadas com o modelo River2D. Foram
testadas diferentes configurações de abrigo observando a taxa de presença no abrigo da truta juvenil,
bem como da sua localização no interior do mesmo. Foram utilizadas as curvas de preferência da
truta-de-rio (Salmo trutta fario) e do barbo-comum (Luciobarbus bocagei), nos estágios juvenil e
adulto, e foram integradas essas curvas com os valores de profundidade do escoamento e velocidade
obtidos com o River2D, para estimar a disponibilidade de habitat para as diferentes configurações de
abrigos para peixes. Os resultados para a truta juvenil estão razoavelmente de acordo com as
principais conclusões do trabalho experimental. Simulações numéricas semelhantes foram também
desenvolvidas num trecho de rio, com o objectivo de analisar problemas de dimensionamento e
implantação dos abrigos e de avaliar a sua eficiência num curso de água natural, tendo sido obtidas
melhorias em termos de disponibilidade de habitat.
Palavras-chave: Modelação de habitat, abrigos laterais, hydropeaking, truta, barbo.

ii

iii
ABSTRACT
Wherever a hydropower station operates under a hydropeaking regime, the stream base flow is
periodically changed by strong and eventually short-duration discharge fluctuations resulting from
hydropower production during peak electricity consumption periods. These events often place an
intolerable stress on fish over all life-stages, thus interfering with the population success and the
ecological condition of the river reach downstream of the powerhouse. The creation of lateral fish
shelters has been pointed out as one of the most promising mitigation measure to protect fish species
from peak flows. Therefore is crucial to determine design guidelines for the fish shelters in order to
increase habitat availability. Being more and more applied in habitat studies, the 2D hydrodynamic
numerical models are a powerful tool to understand the interactions between river flow and the
hydraulic patterns at a microhabitat scale. When coupled with a biological model of habitat
representing different habitat requirements, these models allow a better understanding on the
influence of peak flows over fish occurrence while estimating the relative abundance of habitats. In this
study, results from an experimental facility are compared with habitat simulations carried out with the
River2D. Different shelter configurations were tested in the facility by analysing the shelter global
frequentation rate of juvenile brown trout as well as their preference location inside the shelter. The
habitat requirements curves of the brown trout (Salmo trutta) and Iberian barbel (Luciobarbus bocagei)
were used, both for juvenile and adults, and integrated with the depth and velocity results obtained
with River2D to estimate the habitat availability for different shelters design configurations. Results for
the juvenile trout reasonably agree with the experimental major findings. Similar simulations were also
developed in a river reach, with the purpose of analysing the design problems and shelter construction
and evaluating their efficiency in a natural stream, which conducted to increases in terms of habitat
availability.
Keywords: Habitat modelling, lateral shelters, hydropeaking, trout, barbel.

iv

v
AGRADECIMENTOS
Apesar de ser um trabalho individual, a presente dissertação não teria sido concluída sem o apoio de
uma série de pessoas. Nesta página ficam os meus sinceros agradecimentos.
Ao Professor Doutor António Pinheiro e à Doutora Isabel Boavida, orientadores científicos desta
dissertação, expresso o meu agradecimento não só pelo rigor exigido ao longo deste trabalho, mas
também pela liberdade que me deram para o efectuar, dando-me a escolha do rumo a seguir. Um
muito obrigado pela total disponibilidade demonstrada e por me terem ajudado a dar os primeiros
passos na modelação numérica e na ecohidráulica.
Aos meus pais e avô, por me colocarem sempre em primeiro lugar. Um dia retribuirei, com toda a
certeza. Este trabalho também é fruto do vosso esforço.
À minha madrinha, padrinho e primos pelo apoio incondicional e entusiamo demonstrado durante o
meu percurso académico.
À Bárbara, por todo o apoio e pela inesgotável paciência.
A um grupo de amigos onde nenhum interesse pessoal se sobrepõe a nada. Ao João Fabião, ao
Manuel Guerra, ao João Sá Madeira, ao Francisco Ceia, ao Romeu Rosa, à Maria Gomes, ao João
Correia, ao João Delgado, ao Bruno Cossermelli, ao Francisco do Vale e ao Tiago Pereira o meu
sincero obrigado, pois foram anos bem passados, tanto dentro como fora do Técnico. Tenho a
certeza que o futuro nos reserva mais do mesmo.
Aos verdadeiros amigos que fiz durante a minha passagem pelo Fórum Civil. Não serão esquecidos.
Aos meus companheiros de equipa, pelas muitas vitórias e bons momentos vividos fora de campo.
Em especial ao Nuno Requeijão, ao João Pireza, ao Ricardo Raimundo, ao António Sequeira e por
fim, ao Pedro Pinto, pelos “nossos” dezassete anos de carreira.
Para terminar, um agradecimento especial às minhas avós, pois sei que gostariam de estar presentes
neste momento da minha vida. Guardo todas as memórias com saudade.

vi

vii
PUBLICAÇÕES
Os seguintes artigos foram publicados no seguimento da presente dissertação:
(i) Almeida, R., Boavida, I., Pinheiro, A. 2014. Abrigos laterais para peixes a jusante de
centrais hidroeléctricas – modelação numérica da disponibilidade de habitat. Actas do 12º
Congresso da Água, 5 a 8 de Março de 2014, Lisboa.
(ii) Almeida, R., Boavida, I., Pinheiro, A. 2014. Assesment of trout habitat suitability fish
shelters downstream a hydropower dam. Procedimentos do 10th International Symposium
on Ecohydraulics, 23 a 27 de Junho de 2014, Trondheim, Noruega.
(iii) Almeida, R., Boavida, I., Pinheiro, A. 2014. Habitat numerical modelling to assess fish
shelter design under hydropeaking conditions. 7th River Flow – International Conference
on Fluvial Hydraulics, 3 a 5 de Setembro, Lausanne, Suiça. Submetido.

viii

ix
ÍNDICE
1 Introdução ........................................................................................................... 1
1.1 Enquadramento ....................................................................................................................... 1
1.2 Objectivos ................................................................................................................................ 2
1.3 Estrutura da dissertação .......................................................................................................... 2
2 Síntese de conhecimentos ................................................................................ 5
2.1 Hydropeaking........................................................................................................................... 5
2.1.1 Caracterização e condicionamentos de estudo .................................................................. 5
2.1.2 Consequências .................................................................................................................... 6
2.1.3 Medidas de mitigação .......................................................................................................... 7
2.2 Modelos hidrodinâmicos de habitat ......................................................................................... 9
2.2.1 Considerações gerais .......................................................................................................... 9
2.2.2 Habitat Suitability Criteria e Habitat Suitability Index ........................................................ 11
2.2.3 River2D .............................................................................................................................. 14
2.2.4 Casimir Fish 2D ................................................................................................................. 15
2.3 Espécies estudadas .............................................................................................................. 16
2.3.1 Truta .................................................................................................................................. 16
2.3.2 Barbo ................................................................................................................................. 18
3 Modelação numérica ........................................................................................ 21
3.1 Calibração do modelo numérico ............................................................................................ 21
3.1.1 Considerações gerais ........................................................................................................ 21
3.1.2 Instalação experimental ..................................................................................................... 21
3.2.1.1 Introdução .................................................................................................................. 21
3.2.1.2 Configurações testadas ............................................................................................. 22
3.2.1.3 Conclusões ................................................................................................................ 23
3.2.2 Modelo numérico ............................................................................................................... 24
3.2.2.1 Introdução .................................................................................................................. 24
3.2.2.2 Condições de simulação ........................................................................................... 24
3.3 Análise de resultados ............................................................................................................ 24

x
4 Análise de novas configurações para os abrigos laterais ............................ 29
4.2 Configurações criadas ........................................................................................................... 29
4.3 Análise de resultados ............................................................................................................ 31
4.3.1 Valores de HHS e WUA para as configurações criadas ................................................... 31
4.3.2 Análise dos resultados obtidos para as trutas ................................................................... 37
4.3.2.1 Considerações gerais ................................................................................................ 37
4.3.2.2 Conclusões ................................................................................................................ 50
4.3.3 Análise dos resultados obtidos para os barbos ................................................................. 51
4.3.3.1 Considerações Gerais ............................................................................................... 51
4.3.3.2 Conclusões ................................................................................................................ 57
5 Aplicação dos abrigos a um curso de água natural – rio Ocreza ................ 59
5.1 Introdução .............................................................................................................................. 59
5.2 Disponibilidade de habitat no rio Ocreza ............................................................................... 59
5.3 Abrigos utilizados .................................................................................................................. 61
5.4 Resultados ............................................................................................................................. 63
5.4.1 Valores de WUA no rio Ocreza ......................................................................................... 63
5.4.2 Disponibilidade de habitat nos abrigos .............................................................................. 69
5.4.3 Conclusões ........................................................................................................................ 71
6 Conclusões e recomendações ........................................................................ 77
6.1 Conclusões ............................................................................................................................ 77
6.2 Recomendações .................................................................................................................... 80
Bibliografia .............................................................................................................. 81
Anexos........................................................................................................................ I
Anexo A – Malhas de elementos finitos. Configurações C ................................................................... I
Anexo B – Malhas de elementos finitos. Novas configurações .......................................................... III

xi
ÍNDICE DE FIGURAS
Figura 1.1 - Evolução da produção de energia eléctrica a partir de energias renováveis. Ano de 2013
provisório (Fonte: DGEG, 2013) .............................................................................................................. 1
Figura 2.1 - Parâmetros utilizados para classificar rápidas flutuações do escoamento ......................... 5
Figura 2.2 - Esquema de bacia de compensação (Schweizer et al., 2009) ............................................ 8
Figura 2.3 - Esquema de ensaio laboratorial realizado por Meile (2008) com o objectivo de estudar a
influência das macro-rugosidades no escoamento ................................................................................. 8
Figura 2.4 - Esquema de instalação laboratorial de abrigo para peixes - planta (Ribi et al., 2009) ....... 9
Figura 2.5 - Truta-de-rio (Filipe Ribeiro) ................................................................................................ 16
Figura 2.6 - Riffle (Bain and Stevenson, 1999) ..................................................................................... 16
Figura 2.7 - Curvas de preferência - Truta Juvenil (Inverno-Primavera) .............................................. 17
Figura 2.8 - Curvas de preferência - Truta Juvenil (Verão-Outono) ..................................................... 17
Figura 2.9 - Curvas de preferência - Truta Adulta (Inverno-Primavera) ............................................... 17
Figura 2.10 - Curvas de preferência - Truta Adulta (Verão-Outono) .................................................... 18
Figura 2.11 - Barbo-comum (DGRF) ..................................................................................................... 18
Figura 2.12 - Curvas de preferência - Barbo Juvenil ............................................................................ 19
Figura 2.13 - Curvas de preferência - Barbo Adulto ............................................................................. 19
Figura 3.1 - Esquema em planta da instalação experimental com obstáculo na zona do abrigo......... 21
Figura 3.2 - Esquema 3D da instalação laboratorial (vista de montante) ............................................. 21
Figura 3.3 - Área de referência (a tracejado) das configurações testadas ........................................... 22
Figura 3.4 - Configurações testadas (adaptado de Ribi (2011)) ........................................................... 23
Figura 3.5 - Taxas de presença observadas nas configurações testadas na instalação experimental 23
Figura 3.6 - Malha de elementos finitos. Configuração C8 (River2D) .................................................. 24
Figura 3.7 - Área analisada para determinar o valor de HHS ............................................................... 25
Figura 3.8 - Relação entre a taxa de presença e o HHS - Trutas juvenis (Inverno-Primavera) ........... 25
Figura 3.9 - Relação taxa de presença - HHS para as espécies estudadas. VO – Verão-Outono; IP –
Inverno Primavera. ................................................................................................................................ 27
Figura 4.1 – Novas configurações para o abrigo .................................................................................. 29
Figura 4.2 - Profundidades (m) na configuração C0 ............................................................................. 31
Figura 4.3 - Distribuição de DSI - Truta Juvenil – Inverno-Primavera .................................................. 32
Figura 4.4 - Distribuição de DSI - Truta Adulta – Inverno-Primavera ................................................... 32
Figura 4.5 - Distribuição de DSI - Truta Juvenil – Verão-Outono.......................................................... 33
Figura 4.6 - Distribuição de DSI - Truta Adulta – Verão-Outono........................................................... 33
Figura 4.7 - Distribuição de DSI - Barbo Juvenil ................................................................................... 33
Figura 4.8 - Distribuição de DSI - Barbo Adulto .................................................................................... 34
Figura 4.9 - Ganhos/perdas de HHS em relação à configuração C0 – Trutas (IP – Inverno Primavera;
VO – Verão-Outono) .............................................................................................................................. 36
Figura 4.10 - Ganhos/perdas de HHS em relação à configuração C0 - Barbos ................................... 36
Figura 4.11 - Configuração A ................................................................................................................ 38

xii
Figura 4.12 - Distribuição de velocidades de escoamento (m/s) na configuração A ............................ 38
Figura 4.13 - Distribuição de VSI na configuração A - Truta Juvenil (Inverno-Primavera) ................... 39
Figura 4.14 - Distribuição de CSI na configuração A - Truta Juvenil (Inverno-Primavera) ................... 39
Figura 4.15 - Distribuição de VSI na configuração A - Truta Adulta (Verão-Outono) ........................... 40
Figura 4.16 - Distribuição de CSI na configuração A - Truta Adulta (Verão-Outono) ........................... 40
Figura 4.17 - Configuração B ................................................................................................................ 41
Figura 4.18 - Distribuição de velocidades de escoamento (m/s) na configuração B ............................ 41
Figura 4.19 - Distribuição de VSI na configuração B - Truta Juvenil (Inverno-Primavera) ................... 42
Figura 4.20 - Distribuição de CSI na configuração B - Truta Juvenil (Inverno-Primavera) ................... 42
Figura 4.21 - Distribuição de VSI na configuração B - Truta Adulta (Inverno-Primavera) .................... 43
Figura 4.22 - Distribuição de CSI na configuração B - Truta Adulta (Inverno-Primavera) .................... 43
Figura 4.23 - Distribuição de VSI na configuração B - Truta Adulta (Verão-Outono) ........................... 44
Figura 4.24 - Distribuição de CSI na configuração B - Truta Adulta (Verão-Outono) ........................... 44
Figura 4.25 - Configuração H ................................................................................................................ 45
Figura 4.26 - Distribuição de velocidades de escoamento (m/s) na configuração H ............................ 45
Figura 4.27 - Distribuição de VSI na configuração H - Truta Juvenil (Verão-Outono) .......................... 46
Figura 4.28 - Distribuição de CSI na configuração H - Truta Juvenil (Verão-Outono) .......................... 46
Figura 4.29 - Configuração J ................................................................................................................. 47
Figura 4.30 - Distribuição de velocidades de escoamento (m/s) na configuração J ............................ 47
Figura 4.31 - Distribuição de VSI na configuração J - Truta Juvenil (Verão-Outono) ........................... 48
Figura 4.32 - Distribuição de CSI na configuração J - Truta Juvenil (Verão-Outono) .......................... 48
Figura 4.33 - Configuração K ................................................................................................................ 49
Figura 4.34 - Distribuição de velocidades de escoamento (m/s) na configuração K ............................ 49
Figura 4.35 - Distribuição de VSI na configuração K - Truta Adulta (Inverno-Primavera) .................... 50
Figura 4.36 - Distribuição de CSI na configuração K - Truta Adulta (Inverno-Primavera) .................... 50
Figura 4.37 - Configuração C ................................................................................................................ 51
Figura 4.38 - Distribuição de velocidades de escoamento (m/s) na configuração C ............................ 52
Figura 4.39 - Distribuição de VSI na configuração C - Barbo Juvenil ................................................... 52
Figura 4.40 - Distribuição de CSI na configuração C - Barbo Juvenil ................................................... 53
Figura 4.41 - Configuração D ................................................................................................................ 53
Figura 4.42 - Distribuição de velocidades de escoamento (m/s) na configuração D ............................ 54
Figura 4.43 - Distribuição de VSI na configuração D - Barbo Juvenil ................................................... 54
Figura 4.44 - Distribuição de CSI na configuração D - Barbo Juvenil ................................................... 54
Figura 4.45 - Distribuição de VSI na configuração H - Barbo Juvenil ................................................... 55
Figura 4.46 - Distribuição de CSI na configuração H - Barbo Juvenil ................................................... 55
Figura 4.47 - Distribuição de VSI na configuração J - Barbo Adulto ..................................................... 56
Figura 4.48 - Distribuição de CSI na configuração J - Barbo Adulto .................................................... 56
Figura 4.49 - Distribuição de VSI na configuração K - Barbo Adulto .................................................... 57
Figura 4.50 - Distribuição de CSI na configuração K - Barbo Adulto .................................................... 57
Figura 5.1 - Localização do rio Ocreza (Boavida, 2012) ....................................................................... 59

xiii
Figura 5.2 - Curvas Caudal - WUA - Área molhada - HHS para o Barbo no rio Ocreza ...................... 60
Figura 5.3 - Abrigo utilizado no rio Ocreza ............................................................................................ 61
Figura 5.4 - Localização dos abrigos (1,2 e 3) (River2D) ..................................................................... 62
Figura 5.5 - Esquema 3D do abrigo ...................................................................................................... 63
Figura 5.6 - Curva Caudal - WUA com abrigos (Barbo Juvenil) ........................................................... 63
Figura 5.7 - Curva Caudal - WUA com abrigos (Barbo Adulto) ............................................................ 64
Figura 5.8 - Profundidade (m) para os para os caudais de 50, 60 e 80 m3/s ....................................... 65
Figura 5.9 - Velocidade do escoamento (m/s) para os caudais de 50, 60 e 80 m3/s ........................... 66
Figura 5.10 - Distribuição de CSI (Barbo Juvenil) no rio Ocreza .......................................................... 67
Figura 5.11 - Distribuição de CSI (Barbo Adulto) no rio Ocreza ........................................................... 68
Figura 6.1- Curvas de preferência para as trutas (esquerda – juvenil, direita – adulta) (as curvas de
preferência do rio Lima estão assinaladas a vermelho) (adaptado de Vismara et al., 2001) ............... 78

xiv

xv
ÍNDICE DE TABELAS
Tabela 2.1 - Medidas de mitigação e impactes associados (+ impacte positivo, - impacte negativo) .. 9
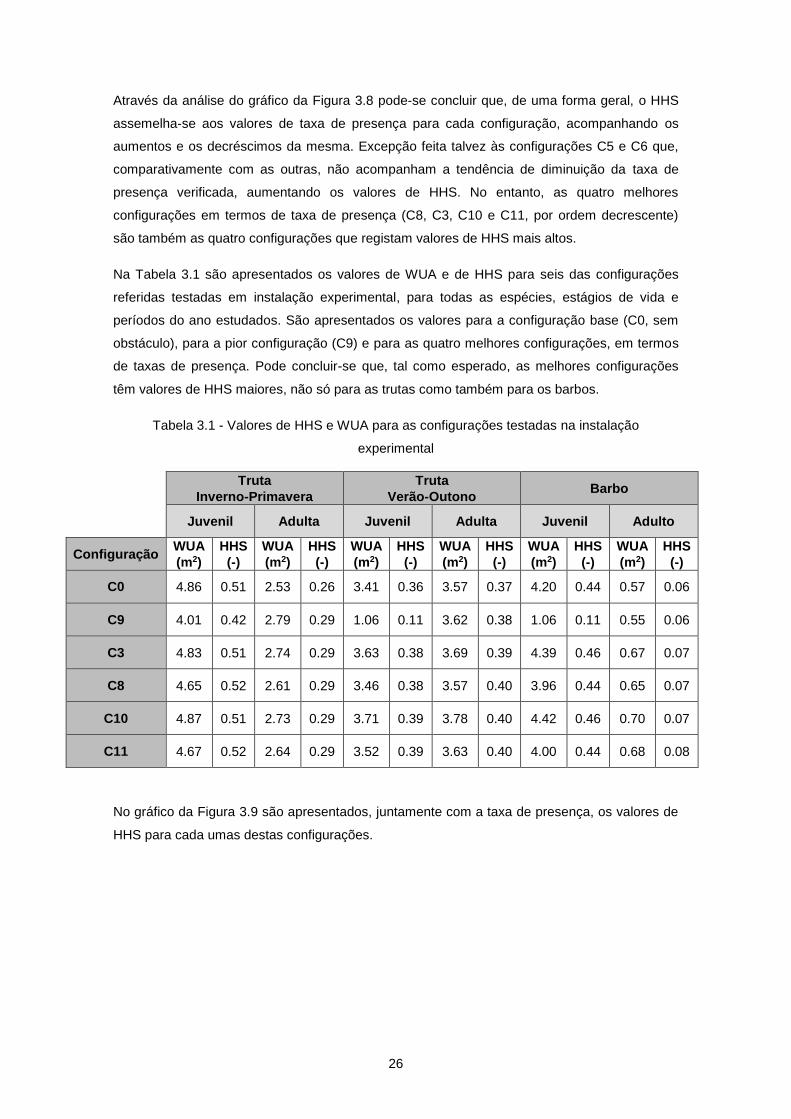
Tabela 3.1 - Valores de HHS e WUA para as configurações testadas na instalação experimental..... 26
Tabela 4.1 - Descrição das novas configurações testadas ................................................................... 30
Tabela 4.2 - Valores aproximados de HSI para as alturas de escoamento no intervalo considerado –
Rio Lima................................................................................................................................................. 31
Tabela 4.3 - Valores de HHS e de WUA para as configurações testadas ............................................ 35
Tabela 5.1 - Cota de soleira dos abrigos .............................................................................................. 62
Tabela 5.2 - Distribuição de CSI no Abrigo 1 ........................................................................................ 69
Tabela 5.3 - Distribuição de CSI no Abrigo 2 ........................................................................................ 70
Tabela 5.4 - Distribuição de CSI no Abrigo 3 ........................................................................................ 70
Tabela 5.5 - Distribuição de DSI no Abrigo 1 ........................................................................................ 71
Tabela 5.6 - Distribuição de DSI no Abrigo 2 ........................................................................................ 72
Tabela 5.7 - Distribuição de DSI no Abrigo 3 ........................................................................................ 72
Tabela 5.8 - Distribuição de VSI no Abrigo 1 ........................................................................................ 73
Tabela 5.9 - Distribuição de VSI no Abrigo 2 ........................................................................................ 74
Tabela 5.10 - Distribuição de VSI no Abrigo 3 ...................................................................................... 74
Tabela 6.1 - Melhores configurações e ganhos de HHS em relação à configuração C0 ..................... 79

xvi

xvii
SIGLAS
CFD Computational Fluid Dynamics
CISI Channel Index Suitability Index
CSI Combined Suitability Index
DSI Depth Suitability Index
DGEG Direcção Geral de Energia e Geologia
GPS Global Positioning System
HHS Hydraulic Habitat Index
HSC Habitat Suitability Criteria
HSI Habitat Suitability Index
IFIM Instream Flow Incremental Methodology
PHABSIM Physical Habitat Simulation
REN21 Renewable Energy Policy Network for the 21st Century
VSI Velocity Suitability Index
WUA Weighted Usable Area

xviii

xix
SIMBOLOGIA
𝝆 Massa volúmica da água
𝝉𝒊𝒋 Tensão exercida no plano perpendicular a i na direção j
Ai Área de influência do nó i
g Aceleração da gravidade
h Altura do escoamento
Ks Coeficiente de rugosidade absoluta
n Número de nós
qi Caudal na direcção i
S0i Declive do leito do canal na direcção i
Sfi Perda de carga unitária
t Tempo
U Velocidade média do escoamento na direcção x
V Velocidade média do escoamento na direcção y

xx
1
1 INTRODUÇÃO
1.1 ENQUADRAMENTO
A energia hídrica ou hidroenergia é uma das fontes de energia renováveis mais antigas da
humanidade, tendo sido inicialmente utilizada para fins agrícolas e industriais. À medida que as
necessidades energéticas das populações têm vindo a aumentar, a utilização da energia
hídrica tem-se concentrado na produção de energia hidroeléctrica, a qual tem adquirido
importância crescente no mercado energético a nível global.
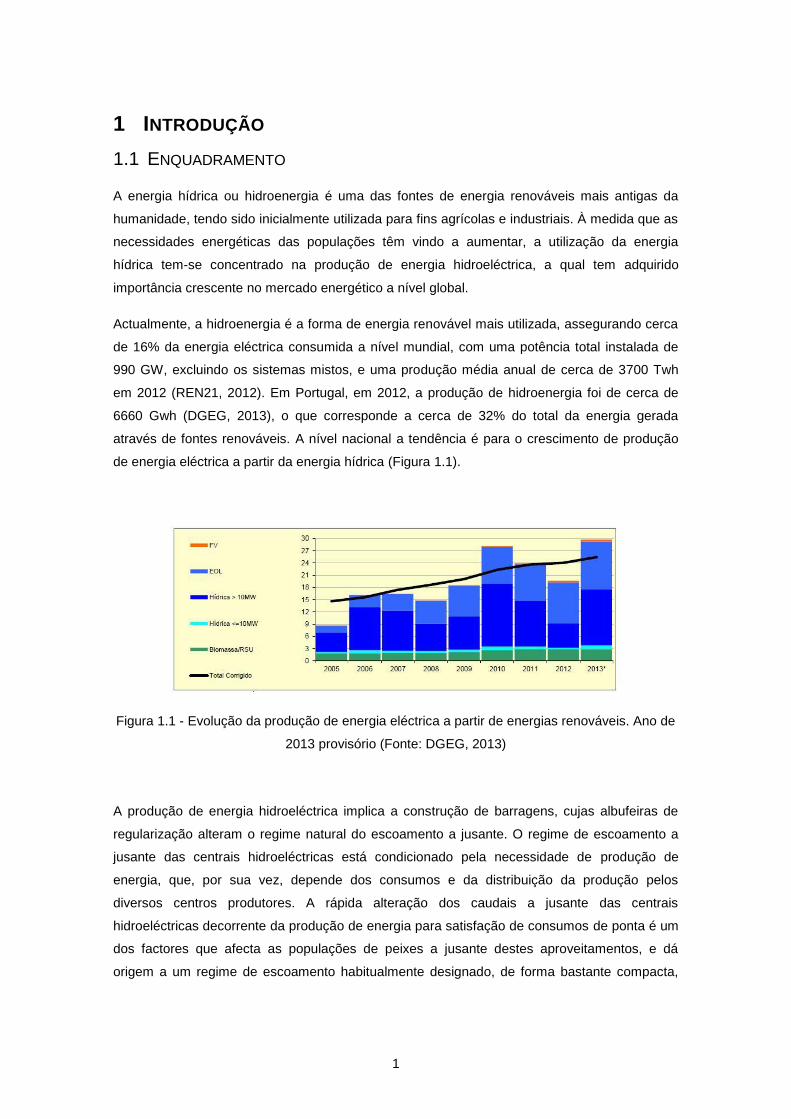
Actualmente, a hidroenergia é a forma de energia renovável mais utilizada, assegurando cerca
de 16% da energia eléctrica consumida a nível mundial, com uma potência total instalada de
990 GW, excluindo os sistemas mistos, e uma produção média anual de cerca de 3700 Twh
em 2012 (REN21, 2012). Em Portugal, em 2012, a produção de hidroenergia foi de cerca de
6660 Gwh (DGEG, 2013), o que corresponde a cerca de 32% do total da energia gerada
através de fontes renováveis. A nível nacional a tendência é para o crescimento de produção
de energia eléctrica a partir da energia hídrica (Figura 1.1).
Figura 1.1 - Evolução da produção de energia eléctrica a partir de energias renováveis. Ano de
2013 provisório (Fonte: DGEG, 2013)
A produção de energia hidroeléctrica implica a construção de barragens, cujas albufeiras de
regularização alteram o regime natural do escoamento a jusante. O regime de escoamento a
jusante das centrais hidroeléctricas está condicionado pela necessidade de produção de
energia, que, por sua vez, depende dos consumos e da distribuição da produção pelos
diversos centros produtores. A rápida alteração dos caudais a jusante das centrais
hidroeléctricas decorrente da produção de energia para satisfação de consumos de ponta é um
dos factores que afecta as populações de peixes a jusante destes aproveitamentos, e dá
origem a um regime de escoamento habitualmente designado, de forma bastante compacta,

2
por hydropeaking, na terminologia anglo-saxónica. Este é o termo que se adoptará na presente
dissertação.
As variações rápidas de caudal, e as consequentes variações de nível, afectam o ecossistema
aquático, sendo um dos factores de maior stress, em particular, para a fauna piscícola (Scruton
et al., 2008). De forma a mitigar os impactes sentidos por estas populações, os abrigos laterais
para peixes são apontados como uma das mais promissoras medidas de mitigação estruturais
para o hydropeaking. Devido à dificuldade de execução de ensaios em protótipos, a
conjugação de ensaios em instalações experimentais com simulações numéricas dos regimes
de escoamento e das condições de habitat é o modo mais viável para testar a eficiência dos
abrigos.
Com o objectivo de avaliar a disponibilidade de habitat dos abrigos laterais para peixes, Ribi
(2011) realizou, no âmbito de uma dissertação de doutoramento, uma série de ensaios
experimentais cujos resultados são comparados com os obtidos através de simulações
numéricas. Validando o modelo numérico com base nos referidos ensaios, analisar-se-ão
geometrias alternativas para os abrigos testados por Ribi (2011), de forma a aumentar a
disponibilidade de habitat dos mesmos e ainda para extrapolar a sua utilização num curso de
água natural.
1.2 OBJECTIVOS
O presente estudo tem como objectivo principal aferir a eficiência de abrigos laterais para
peixes a jusante de centrais hidroeléctricas que funcionem em regime de hydropeaking. Para
concretizar o referido objectivo será necessário:
1. Efectuar a modelação numérica 2D dos escoamentos num canal rectangular com um
abrigo lateral para peixes, com diferentes configurações, em conformidade com o
testado por Ribi (2011);
2. Utilizar curvas de preferência de habitat, de forma a estudar a disponibilidade de
habitat proporcionada por cada tipo de abrigo para as diferentes espécies e estágios de
desenvolvimento;
3. Comparar os resultados das simulações numéricas e da instalação experimental, de
modo a verificar a validade das mesmas;
4. Estudar formas de abrigo alternativas às estudadas por Ribi (2011);
5. Estudar as condições de aplicação e a eficiência destes abrigos laterais para peixes
quando aplicados a um curso de água natural.
1.3 ESTRUTURA DA DISSERTAÇÃO
A presente dissertação encontra-se dividida em 6 capítulos. No primeiro, de introdução,
apresenta-se o enquadramento do tema e enunciam-se os objectivos do estudo. No capítulo 2
é apresentado o conceito de hydropeaking e uma introdução à modelação hidrodinâmica

3
bidimensional e de habitat, sendo referidos alguns modelos computacionais presentemente
utilizados para o efeito, e são descritas as espécies piscícolas consideradas no estudo. No
capítulo 3 descreve-se a calibração do modelo e apresentam-se a comparação entre
resultados experimentais e de simulações numéricas. No capítulo 4 são apresentadas novas
configurações para os abrigos, que foram desenvolvidas com o objectivo de aumentar a
disponibilidade de habitat nos mesmos. No capítulo 5 são apresentadas as simulações
correspondentes à aplicação dos abrigos laterais a um curso de água natural, o rio Ocreza, e é
analisada a variação da disponibilidade de habitat no troço analisado. Por fim, no último
capítulo, são apresentadas as conclusões gerais desta dissertação e algumas recomendações
para trabalhos futuros.

4

5
2 SÍNTESE DE CONHECIMENTOS
2.1 HYDROPEAKING
2.1.1 CARACTERIZAÇÃO E CONDICIONAMENTOS DE ESTUDO
As centrais hidroeléctricas com albufeira de armazenamento destinadas à produção de
electricidade são responsáveis pelo fenómeno de hydropeaking (Ribi et al., 2009). A alteração
do regime natural de escoamento, causado pelos picos de produção de energia hidroeléctrica,
difere substancialmente dos regimes de cheias naturais, quer na frequência, quer na variação
temporal do caudal, quer na magnitude. Este fenómeno pode assim ser definido como a
variação rápida do caudal escoado num troço de um curso de água imediatamente a jusante
duma central hidroeléctrica. Estas variações abruptas e frequentes de caudal afectam o
ecossistema aquático a jusante, com especial incidência na fauna piscícola. As elevadas
velocidades de escoamento podem levar à morte dos peixes inclusive (Jungwirth et al., 2003) e
pelas reduções rápidas de habitat, que podem implicar a retenção dos peixes em zonas que,
por redução do nível, ficam isoladas do leito onde ocorre o escoamento.
Na Figura 2.1 são representados os parâmetros utilizados para classificar as rápidas flutuações
do caudal: razão entre caudal máximo e o caudal mínimo, taxa de variação da altura de
escoamento e taxa de variação do caudal.
A razão entre o caudal máximo e o caudal mínimo é obtida através da divisão do maior
valor de caudal registado pelo caudal de base. Este parâmetro pode ser também
utilizado para classificar cheias.
A taxa de variação da altura de escoamento é representada pela taxa de variação do
nível da água ao longo do tempo. Este rácio pode ser maior quando o caudal aumenta
devido a fenómenos de precipitação intensa.
A taxa de variação do caudal é o tempo decorrido entre dois picos no hidrograma. A
frequência com que estes picos ocorrem também pode ser um parâmetro de
caracterização das flutuações do escoamento.
Seria de esperar uma maior frequência nos períodos da manhã e do final do dia, quando as
necessidades de electricidade são maiores. No entanto, como consequência das alterações
existentes na utilização de energias renováveis, estes padrões têm sofrido mudanças nos
últimos anos.
Figura 2.1 - Parâmetros utilizados para classificar rápidas flutuações do escoamento
(Harby et al., 2013)

6
Os estudos em protótipo dos regimes de hydropeaking não são fáceis de realizar, dadas as
implicações que têm na operação das centrais hidroeléctricas, e consequentemente, na
produção de energia, pelo que, alternativamente, se recorre a ensaios em instalações
experimentais. Estes ensaios permitem mais facilmente analisar determinados aspectos
específicos de forma sistemática, desde que as condições sejam próximas das reais. A
reprodução de condições morfológicas naturais (como por exemplo o substrato, o escoamento
subterrâneo e a dinâmica de sedimentos) é difícil (Harby et al., 2013).
De forma a obter resultados válidos e conseguir reproduzir o regime de hydropeaking
aconselha-se assim a utilização de estudos em protótipo conjugada com simulações
numéricas.
2.1.2 CONSEQUÊNCIAS
As características físicas num rio podem ser variadas, nomeadamente no que diz respeito às
condições hidráulicas, temperatura, gelo, visibilidade, cobertura, substrato e erosão. No
entanto, as alterações introduzidas pelo homem nos cursos de água naturais podem levar a
severas alterações ambientais no rio e a criar condições insuportáveis para os organismos que
lá habitam. Assim, os impactes causados pelo hydropeaking num rio podem ser divididos em
três grupos: impactes morfológicos, na qualidade da água e biológicos.
Dentro dos impactes morfológicos no rio o denominador comum é a erosão. Este fenómeno
afecta não só as margens e leito do rio, como potencia a existência de sedimentos em
suspensão na coluna de água. Com uma diminuição da altura de escoamento podem ser
criadas zonas secas junto às margens. No que diz respeito à qualidade da água, o
hydropeaking pode criar alterações ao nível da temperatura da água, na sua turbidez e
aumentar a vegetação existente na massa de água.
Em termos de impactes biológicos, sentidos de uma forma directa pelos peixes, pode referir-se
a possibilidade de existirem peixes encalhados (i.e. que não conseguiram regressar a tempo ao
centro do canal durante a diminuição do caudal após o turbinamento), o aumento do stress
sentido pelos mesmos devido às constantes variações de caudal, a necessidade dos peixes
utilizarem mais energia para lidar com as elevadas velocidades de escoamento causando a
sua fadiga, a redução de áreas disponíveis para reprodução e o possível arrastamento pela
corrente, com especial incidência nos estágios juvenis.
Em rios de cabeceira, como nos Alpes por exemplo, a salmo trutta fario é uma das espécies
que mais sofre com este regime. Os leitos de desova correm o risco de ficarem secos e o
habitat junto às margens para as populações juvenis pode desaparecer (Liebig et al., 1998).
Sem um refúgio apropriado e com a necessidade de um maior uso de energia por parte dos
peixes para sobreviver, torna-se difícil a sua sobrevivência durante o Inverno.

7
2.1.3 MEDIDAS DE MITIGAÇÃO
Dentro das medidas de mitigação podem ser definidos dois grupos: medidas operacionais e
medidas estruturais.
As medidas operacionais dizem respeito à operação da central hidroeléctrica, e têm como
principais objectivos diminuir o caudal de ponta (ou pico) e diminuir a taxa de variação da altura
de escoamento. A melhor medida para evitar os impactes negativos do hydropeaking é alterar
o caudal restituído de uma forma tão lenta que permita aos organismos adaptarem-se às novas
condições de escoamento no rio, a jusante da central hidroeléctrica. Como medida adicional, e
se possível, pode ser útil aumentar ou diminuir o caudal escoado de uma forma lenta que
funcione como “sinal de alerta” para os peixes e restantes organismos. Estas medidas estão
directamente relacionadas com as válvulas de seccionamento e turbinas existentes nas
centrais, pelo que podem ser mais difíceis de executar.
Existem alguns riscos associados às medidas operacionais, como por exemplo:
A possibilidade de existirem peixes encalhados no leito do rio quando o hydropeaking
ocorre após um longo período de caudais elevados e quando a redução do caudal é
feita de forma abrupta;
A redução do caudal feita durante os períodos diurnos é mais preocupante uma vez
que os peixes tendem a refugiar-se visualmente no substrato ou junto a raízes e outros
detritos, podendo ficar encalhados nesses refúgios após a paragem das turbinas. No
Inverno, esta situação tende a ser mais alarmante devido aos baixos níveis energéticos
das espécies;
A possibilidade de efectuar manobras de fecho que coloquem em causa a segurança
da central hidroeléctrica, devido aos regimes transitórios.
Clarke et al. (2008) sugere três medidas de mitigação para as operações hidráulicas:
Assegurar que o caudal de base que ocorre num rio é suficiente para manter um
ecossistema sustentável;
O aumento do caudal deve mimetizar um aumento natural do caudal, como nas cheias
de pequena dimensão;
A taxa de variação de caudal deve ser ajustada aos comportamentos diurnos das
espécies existentes.
As medidas estruturais estão relacionadas com as alterações morfológicas do rio de forma a
melhorar as condições de habitat para os peixes. Estas alterações têm como base a criação de
estruturas artificiais.
Uma das opções a considerar pode passar por evitar a restituição do caudal turbinado para o
rio, utilizando um lago ou um canal paralelo ao rio para o efeito. Outra abordagem pode ser a
de uma restituição controlada do caudal turbinado com o objectivo de tornar o regime de

8
escoamento próximo do natural, utilizando uma bacia de compensação. Por fim, pode ainda ser
feita uma melhoria das condições morfológicas do rio ou construir abrigos para os peixes nas
margens do rio.
A criação de uma bacia de compensação pode criar um espaço para fins de lazer ou ainda
permitir que a água seja turbinada novamente, caso a queda útil se revele favorável para a
produção de energia. Com a criação desta bacia, parte do caudal turbinado seria restituído
directamente no rio (aproximando a descarga a um regime natural), sendo o caudal excedente
restituído na bacia. Na Figura 2.2 pode ser observado um esquema de uma possível solução
com bacia de compensação.
Figura 2.2 - Esquema de bacia de compensação (Schweizer et al., 2009)
Com o objectivo de melhorar as condições morfológicas do rio podem ser criadas macro-
rugosidades nas margens do rio (Figura 2.3), que aumentam a resistência ao escoamento e
criam condições de abrigo.
Figura 2.3 - Esquema de ensaio laboratorial realizado por Meile (2008) com o objectivo de
estudar a influência das macro-rugosidades no escoamento
De forma a criar melhores condições de habitat para a fauna piscícola podem ser construídos
abrigos para os mesmos. Estes abrigos são instalados ou construídos nas margens do rio,
podendo ter diversas configurações, com o objectivo de criar padrões de velocidade atractivos
para as espécies que nele habitam. Com a introdução de obstáculos na zona de entrada do
abrigo é possível melhorar as condições hidráulicas do escoamento aumentando a efectividade

9
do mesmo. Na Figura 2.4 é possível observar um esquema de uma instalação laboratorial com
o objectivo de estudar a efectividade de um abrigo lateral para peixes.
Figura 2.4 - Esquema de instalação laboratorial de abrigo para peixes - planta (Ribi et al., 2009)
As medidas de mitigação teriam, preferencialmente, impactes positivos tanto nos rios como nas
espécies que lá habitam. No entanto nem todas as medidas têm só impactes positivos, pelo
que é necessário analisar também os impactes negativos que uma possível medida possa ter.
Na Tabela 2.1 é feito um resumo de possíveis medidas de mitigação e impactes associados.
Tabela 2.1 - Medidas de mitigação e impactes associados (+ impacte positivo, - impacte
negativo)
Medida de mitigação
Diminuição gradual do
caudal restituído
Aumento gradual do
caudal restituído
Bacia de compensação
Abrigos para
peixes
Impactes
Morfológicos Zonas secas junto às margens + +
-
Aumento da erosão
+ +
Qualidade da água
Alteração na temperatura da água + + + +
Alterações na turbidez + +
Biológicos
Encalhamento +
+ +
Fadiga + + + +
Arrastamento
+
+ *adaptado de Harby et al. (2013)
2.2 MODELOS HIDRODINÂMICOS DE HABITAT
2.2.1 CONSIDERAÇÕES GERAIS
As características dos escoamentos em canais naturais é influenciada por diversos factores,
como por exemplo a existência de sedimentos, as características topográficas e a
permeabilidade do leito. O escoamento interage com os sedimentos e com as características
topográficas do rio criando complexos padrões de escoamento que variam não só no espaço

10
mas também no tempo (Crowder et al., 2000). Estas interacções entre o escoamento e as
características topográficas revelam-se fundamentais para compreender e determinar a
adequabilidade de habitat existente num rio. Os modelos hidrodinâmicos, quando utilizados de
forma isolada, nada dizem quanto à adequabilidade de um determinado habitat para uma certa
espécie. Desta forma, é necessário juntar a estes modelos um modelo biológico de selecção de
habitat, como por exemplo o Habitat Suitability Criteria (HSC) (Bovee, 1982).
As alterações provocadas pelo homem no regime natural de escoamento podem danificar ou
destruir totalmente os habitats existentes num rio sendo que, sempre que se regista alguma
alteração ao escoamento, são efectuados estudos para determinar as condições de habitat
disponíveis. Para efectuar estes estudos a Dinâmica de Fluídos Computacional (CFD –
Computational Fluid Dynamics) tornou-se uma técnica fundamental para fornecer previsões
quantitativas não só temporais como espaciais das características do escoamento em cursos
de água naturais (Leclerc et al., 1995). É importante determinar as variações espaciais do
escoamento devido ao seu papel vital na determinação do tipo, qualidade e quantidade de
habitat disponível num curso de água.
Os métodos para previsão das características do escoamento podem ser divididos em quatro
categorias: (1) modelos tridimensionais (3D), (2) modelos bidimensionais (2D), (3) modelos
unidimensionais (1D) e (4) modelos não numéricos. Os três primeiros são baseados em
informação espacial do escoamento em três, duas e numa só dimensão, respectivamente,
sendo que o último inclui medidas e observações feitas no local. Tal como o nome indica, os
modelos 3D permitem determinar as propriedades do escoamento em três dimensões,
longitudinal (em direcção a jusante – eixo x), transversal (eixo y) e vertical (eixo z). Os modelos
2D determinam as propriedades do escoamento nas direcções longitudinal e transversal, sendo
que os modelos 1D apenas permitem determinar estas mesmas propriedades na direcção
longitudinal.
Hoje em dia a utilização de modelos 2D é mais vantajosa que a utilização de modelos 3D,
devido às capacidades computacionais necessárias não serem tão exigentes. A utilização
destes modelos bidimensionais é considerada suficiente para a maior parte das utilizações
existentes na ecohidráulica (Rodi et al., 1981) e tem também a vantagem de poder ser utilizada
numa escala comparável à escala do microhabitat dos peixes.
Os modelos 1D de modelação de escoamento analisam um troço de rio dividindo-o em células
(ou subsecções) em que cada uma delas tem um único valor de profundidade do escoamento e
velocidade de escoamento associado (Bovee, 1978). Assim, qualquer variação espacial no
escoamento, como os gradientes de velocidade que se propagam na direcção transversal não
podem ser modelados por estes modelos. Consequentemente, a utilização de modelos 2D em
estudos de habitat pode ser superior em relação à utilização de modelos 1D em diversos
aspectos (Bovee,1996). A grande vantagem dos modelos bidimensionais está relacionada com
a possibilidade de quantificar as variações espaciais do escoamento que influenciam o habitat

11
disponível, sem exigir as capacidades computacionais dos modelos 3D. De forma a explorar
totalmente as capacidades dos modelos 2D, os modelos hidráulicos precisam que as malhas
utilizadas sejam adequadas para reproduzir de forma exacta a topografia do leito que
determina as condições de habitat disponíveis para as espécies em estudo (Crowder et al.,
2000).
A utilização de modelos 2D para prever as características de escoamento importantes para o
habitat consiste primariamente em 3 passos (Crowder et al., 2000): (1) determinar o tipo e a
escala dos padrões de escoamento importantes para o estudo; (2) determinar a batimetria do
local a uma escala adequada que permita ao modelo reproduzir as variações espaciais do
escoamento relevantes para o estudo; e (3) refinar a malha do modelo a um nível que forneça
uma solução com uma resolução aceitável.
2.2.2 HABITAT SUITABILITY CRITERIA E HABITAT SUITABILITY INDEX
Os modelos hidrodinâmicos, quando utilizados de forma isolada, nada dizem quanto à
adequabilidade de um determinado habitat para uma certa espécie. Desta forma é necessário
juntar a estes modelos um modelo biológico de selecção de habitat, o Habitat Suitability Criteria
(HSC) (Bovee, 1982).
O HSC converte as características hidráulicas do escoamento e do curso de água em índices
de adequabilidade de habitat, o chamado Índice de Adequabilidade de Habitat (Habitat
Suitability Index - HSI). O HSI representa a adequabilidade de um determinado habitat para
uma certa espécie que pode ainda ser descrita em termos de estágio de vida (juvenil, adulto) e
ser influenciada pelo período do ano. O HSI resume a preferência das espécies relativamente
aos factores abióticos, sendo os mais importantes a profundidade (Geist et al., 2000), a
velocidade do escoamento (Kynard et al., 2000), a composição do substrato (Knapp et al.,
1999) e a cobertura (Vadas et al., 2001). A velocidade do escoamento influencia a
disponibilidade de alimentos, os comportamentos de locomoção dos peixes e o seu
metabolismo (por exemplo, a respiração). A profundidade tem influência na temperatura da
água, funciona também como refúgio, afecta a disposição espacial das espécies,
nomeadamente na distância existente entre predadores e presas, podendo também ser um
factor limitador dos movimentos migratórios. O substrato é constituído pelas partículas
existentes no rio, sejam elas orgânicas ou inorgânicas, que podem servir de suporte para uma
série de actividades dos peixes, como a sua reprodução, refúgio de predadores ou de caudais
que sejam menos favoráveis. A cobertura afecta também a facilidade que os peixes têm de
encontrar refúgio e manterem-se a salvo de outros predadores. O HSI toma valores entre 0,0 e
1,0 que correspondem, respectivamente, à mínima e à máxima preferência para uma dada
espécie.
Os factores físicos acima descritos (i.e. profundidade, velocidade do escoamento, substrato e
cobertura) são variáveis abióticas que influenciam directamente a distribuição dos peixes. O
valor de HSI para estes factores pode ser determinado através do rácio da percentagem de

12
utilização (número de indivíduos observados que usam um determinado intervalo de uma
variável) com a disponibilidade desse intervalo de valores (percentagem da área do rio com
estas características). Em suma, isto significa que dada uma igual possibilidade de escolha
para os indivíduos existentes num curso de água, eles irão escolher áreas específicas que lhes
são mais favoráveis, pondo de parte outras.
Utilizar valores de HSI previamente determinados para um determinado local, em outras
regiões de características diferentes (i.e. hidrológicas, biológicas, geográficas, entre outras)
pode levantar uma série de problemas (Freeman et al., 1997). Por exemplo, os valores de HSI
podem ser influenciados pela disponibilidade de alimento, pelas relações entre espécies e pela
disponibilidade de habitat (Fausch et al., 1986). Assim deve-se, sempre que possível,
determinar os valores de HSI para o local em estudo.
O conjunto de valores de HSI determinados relativos a uma série de factores abióticos para
uma determinada espécie pode ser definido como Habitat Suitability Curve ou Curva de
Preferência de Habitat. As curvas de preferência de habitat são determinadas em diferentes
troços de rio natural ou semi-natural da mesma região através de electrofishing (pesca
eléctrica), prática que quando feita de forma correcta não traz qualquer dano permanente para
os peixes. Normalmente é feita por uma equipa de duas pessoas – uma com o eléctrodo e a
outra com uma rede para retirar o peixe da água. O troço de rio é percorrido em zig-zag de
jusante para montante para assegurar uma correcta recolha de dados. Introduzindo o eléctrodo
na água é criada uma corrente eléctrica que perturba os peixes que são então recolhidos
através da rede. É identificada a espécie e o peixe é medido por forma a aferir o seu estágio de
vida, sendo depois devolvidos à água mais para jusante do que o local onde foram recolhidos,
de forma a evitar uma segunda recolha do mesmo indivíduo. Na zona onde o peixe foi
encontrado é deixado um chumbo com uma identificação, para que posteriormente possam aí
ser medidas as variáveis abióticas – profundidade, velocidade de escoamento e substrato. De
forma a aferir sobre a disponibilidade de habitat existente naquele troço, são seleccionados um
conjunto de pontos ao longo de diferentes secções transversais (15 a 25) onde se procede à
medição das referidas variáveis abióticas. Os valores registados das variáveis abióticas são
depois divididos em classes e são construídos os respectivos histogramas de frequência de
uso (i.e. número de peixes encontrado em determinada classe de valores) e de disponibilidade
(i.e. número de pontos medidos em determinada classe de valores). O HSI é calculado tendo
em conta o quociente entre o uso e a disponibilidade, sendo depois normalizados, dividindo
pelo valor máximo, para que o valor mínimo seja 0 e o máximo 1.
As curvas de preferência de habitat têm algumas limitações, podendo não reflectir de forma
totalmente correcta o que acontece na realidade. Por exemplo, não é possível concluir que
para uma determinada espécie e estágio de vida uma velocidade de 0,3 m/s é favorável e uma
velocidade de 0,31 m/s já não é favorável. De certa forma pode referir-se que em ecologia as
transições não são nítidas, mas graduais (Mathews, 1998). Outra característica das curvas de
preferência de habitat é que consideram a independência entre factores abióticos.

13
O habitat não depende de uma só variável mas sim de um conjunto das mesmas, combinando-
se entre si, estabelecendo assim a Adequabilidade Combinada de Habitat (Combined
Suitability Index - CSI). O CSI pode ser determinado multiplicando o valor de HSI das variáveis
envolvidas, admitindo assim uma independência entre as variáveis analisadas (2.2.1).
𝐶𝑆𝐼 = 𝐷𝑆𝐼 × 𝑉𝑆𝐼 × 𝐶𝐼𝑆𝐼
(2.2.1)
Onde 𝐷𝑆𝐼 representa o Depth Suitability Index (profundidade do escoamento), 𝑉𝑆𝐼 o Velocity
Suitability Index (velocidade) e 𝐶𝐼𝑆𝐼 o Channel Index Suitability Index que pode ser relativo à
influência do substrato ou da cobertura. Se qualquer destes parâmetros for igual a zero, conduz
automaticamente a um valor de CSI semelhante. Esta foi a abordagem utilizada nesta
dissertação. O valor de CISI foi considerado 1 para qualquer uma das opções, anulando assim
o efeito do substrato e/ou cobertura.
Outra abordagem para o cálculo do CSI é a média geométrica (2.2.2) (Korman et al., 1994).
Este método reduz o peso de índices HSI mais elevados comparativamente com os restantes.
Por exemplo, tendo um valor de DSI e VSI de 0,1 e um CISI de 0,9, a média geométrica dará
um valor mais alto para o CSI do que o produto dos três parâmetros, atenuando assim a
influência do parâmetro com preferência mais elevada.
𝐶𝑆𝐼 = √𝐷𝑆𝐼 × 𝑉𝑆𝐼 × 𝐶𝐼𝑆𝐼3
(2.2.2)
Após determinados os valores de CSI é possível determinar a Superfície Ponderada Útil
(Weighted Usable Area - WUA). A WUA é a área (m2) que pode eventualmente ser utilizada por
uma determinada espécie e é calculada através da multiplicação da área de influência de cada
nó da malha pelo valor de CSI do mesmo:
𝑊𝑈𝐴 = ∑ 𝐶𝑆𝐼𝑖 ∗ 𝐴𝑖
𝑛
𝑖=1
(2.2.3)
Onde 𝐴𝑖 é a área de influência do nó i e 𝑛 é o número de nós.
A WUA depende do caudal existente pois influencia directamente as velocidades e
profundidades de escoamento num rio. A validade deste parâmetro tem sido discutida, pelo
que a Disponibilidade Hidráulica de Habitat (Hydraulic Habitat Suitability – HHS) (Stalnaker et
al., 1995) pode revelar-se um índice mais adequado para quantificar o habitat existente.
O HHS resulta da divisão da WUA pela área ocupada pelo escoamento, eliminando assim a
influência da área molhada. Desta forma é possível comparar diferentes troços de rio, avaliar
cenários de caudal e aferir sobre diferentes configurações de abrigo de uma forma mais
precisa. O índice HHS varia tal como os anteriores entre 0,0 e 1,0.

14
2.2.3 RIVER2D
O modelo River 2D (Steffler, 2000) foi um dos modelos utilizados neste estudo para determinar
o habitat disponível. É um modelo numérico de elementos finitos que resolve as equações de
conservação da massa e de conservação da quantidade de movimento, considerando as
componentes horizontais da velocidade (segundo x e y) e os respectivos valores médios em
profundidade (Steffler et al., 2002).
O princípio de conservação da massa é expresso pela equação (2.2.4).
𝜕ℎ
𝜕𝑡+
𝜕𝑞𝑥
𝜕𝑥+
𝜕𝑞𝑦
𝜕𝑦= 0 (2.2.4)
As equações de conservação da quantidade de movimento são:
𝜕𝑞𝑥
𝜕𝑡+
𝜕
𝜕𝑥(𝑈𝑞𝑥) +
𝜕
𝜕𝑦(𝑉𝑞𝑥) +
𝑔
2
𝜕
𝜕𝑥ℎ2 = 𝑔ℎ(𝑆0𝑥 − 𝑆𝑓𝑥) +
1
𝜌(
𝜕
𝜕𝑥(ℎ𝜏𝑥𝑥)) +
1
𝜌(
𝜕
𝜕𝑦(ℎ𝜏𝑥𝑦))
(2.2.5)
𝜕𝑞𝑥
𝜕𝑡+
𝜕
𝜕𝑥(𝑈𝑞𝑦) +
𝜕
𝜕𝑦(𝑉𝑞𝑦) +
𝑔
2
𝜕
𝜕𝑦ℎ2 = 𝑔ℎ(𝑆0𝑥 − 𝑆𝑓𝑦) +
1
𝜌(
𝜕
𝜕𝑥(ℎ𝜏𝑦𝑥)) +
1
𝜌(
𝜕
𝜕𝑦(ℎ𝜏𝑦𝑦))
(2.2.6)
Em que:
ℎ - profundidade;
𝑞𝑥 e 𝑞𝑦- caudais na direcção x e y, respectivamente;
𝑡 – tempo;
𝑈 e 𝑉 – velocidades médias do escoamento na direcção x e y;
𝑔 – aceleração da gravidade;
𝑆0𝑖 – declive do leito do canal na direcção i;
𝑆𝑓𝑖 – perda de carga unitária;
𝜌 – massa volúmica da água;
𝜏𝑖𝑗 – tensão exercida no plano perpendicular a i na direcção j.
Os dados de entrada do modelo River2D são a caracterização topográfica do leito do rio, os
valores da rugosidade do leito do rio, as condições de fronteira e as condições iniciais de
escoamento.
A caracterização topográfica têm uma importância fundamental no rigor dos resultados obtidos,
pelo que deve ser suficientemente detalhada para reproduzir com rigor o leito do curso de água
e, consequentemente, as condições de habitat. Depois de recolhidos os dados, é aconselhada
a criação de um modelo digital do terreno antes de ser utilizado no modelo 2D, por forma a
verificar a adequação da topografia utilizada.

15
A rugosidade no leito do rio, em cada nó da malha, é caracterizada pelo coeficiente de
rugosidade absoluta, Ks. Uma observação do material constituinte do leito e das margens é
suficiente para estabelecer uma aproximação de valores para o parâmetro Ks. A calibração do
modelo consiste no ajuste das alturas de escoamento medidas em campo com as simuladas
através da alteração de valores de rugosidade absoluta. Valores de rugosidade invulgares
podem significar a existência de um mau levantamento topográfico.
As condições de fronteira do modelo River2D são o valor de caudal numa secção de entrada
(i.e. inflow) e de uma altura de escoamento na secção de saída (i.e. outflow). É aconselhado
que estas duas secções estejam suficientemente afastadas da área específica em estudo para
eliminar a influência do “efeito fronteira” nos resultados obtidos. O “efeito fronteira” pode ser
traduzido como uma incerteza associada aos resultados obtidos nessas secções. As condições
iniciais de escoamento são importantes pois servem de aproximação inicial no processo
iterativo de resolução do problema.
A criação da malha de elementos finitos é o passo mais importante na utilização destes
modelos. É necessário criar uma malha que represente o melhor possível a topografia do leito
do rio. Nós mais próximos uns dos outros nas áreas de maior interesse, alteração do
espaçamento entre nós de uma forma gradual e regularidade dos elementos criados são
considerações importantes.
A regularidade dos triângulos criados pode ser traduzida pelo QI – Mesh Quality Index. O QI
traduz o grau de semelhança entre os triângulos gerados pela malha de elementos finitos. É
assim definido pelo quociente entre a área circunscrita de cada triângulo e pela área
circunscrita de um triângulo equilátero. O River2D permite suavizar a malha, ajustando de
forma automática os triângulos de forma a aumentar o QI da mesma. A optimização da malha
de elementos finitos passa pela adição de linhas de quebra, eliminado ou adicionando nós,
entre outras.
Os resultados fornecidos pelo River2D são as duas componentes horizontais de velocidade e a
profundidade, que podem ser obtidas em cada ponto do domínio. As distribuições de
velocidade na vertical são assumidas como uniformes e a distribuição de pressões é
considerada hidrostática. Efeitos tridimensionais como as correntes secundárias em canais não
rectilíneos não são incluídos. Também as forças de Coriolis e do vento, que podem ter
influência em grandes massas de água, como lagos e estuários, não são consideradas.
2.2.4 CASIMIR FISH 2D
O Casimir Fish 2D (Schneider et al., 2010) foi outro dos modelos utilizados neste estudo. Com
origem na Universidade de Estugarda, o Casimir (Computer Aided Simulation System for
Instream Flow Requirements) foi desenvolvido no início da década de 90 e sofreu várias
alterações até permitir a modelação da disponibilidade de habitat.

16
O modelo Casimir utiliza como dados de entrada as variações temporais e espaciais das
profundidades, velocidades de escoamento e tipos de substrato e permite calcular a
disponibilidade de habitat através das curvas de preferência ou da lógica fuzzy.
Este modelo permite também utilizar como dados de entrada as simulações já feitas com
outros modelos computacionais, como o River2D.
2.3 ESPÉCIES ESTUDADAS
2.3.1 TRUTA
A truta-de-rio (Salmo truta fario) (Figura 2.5) é uma espécie que surge em rios com águas frias
e oxigenadas, que são característicos do norte de Portugal (Valente, 1993). Prefere rios com
velocidades elevadas, baixas profundidades e uma boa qualidade de água. De uma forma
geral podem ser encontradas em riffles (zonas de águas menos profundas e de elevadas
velocidades de escoamento) (Figura 2.6).
As trutas-fário têm um tamanho médio menor que 50 cm, tamanho máximo igual a 100 cm e
podem viver até 15 anos. De uma forma geral alimentam-se das presas que descem o rio,
podendo mesmo as adultas alimentarem-se de outras espécies piscícolas como o barbo, a
boga, ou até outras trutas.
Figura 2.5 - Truta-de-rio (Filipe Ribeiro)
Figura 2.6 - Riffle (Bain and Stevenson, 1999)
Neste estudo a análise feita a esta espécie subdividiu-se em dois estágios (adulto e juvenil) e
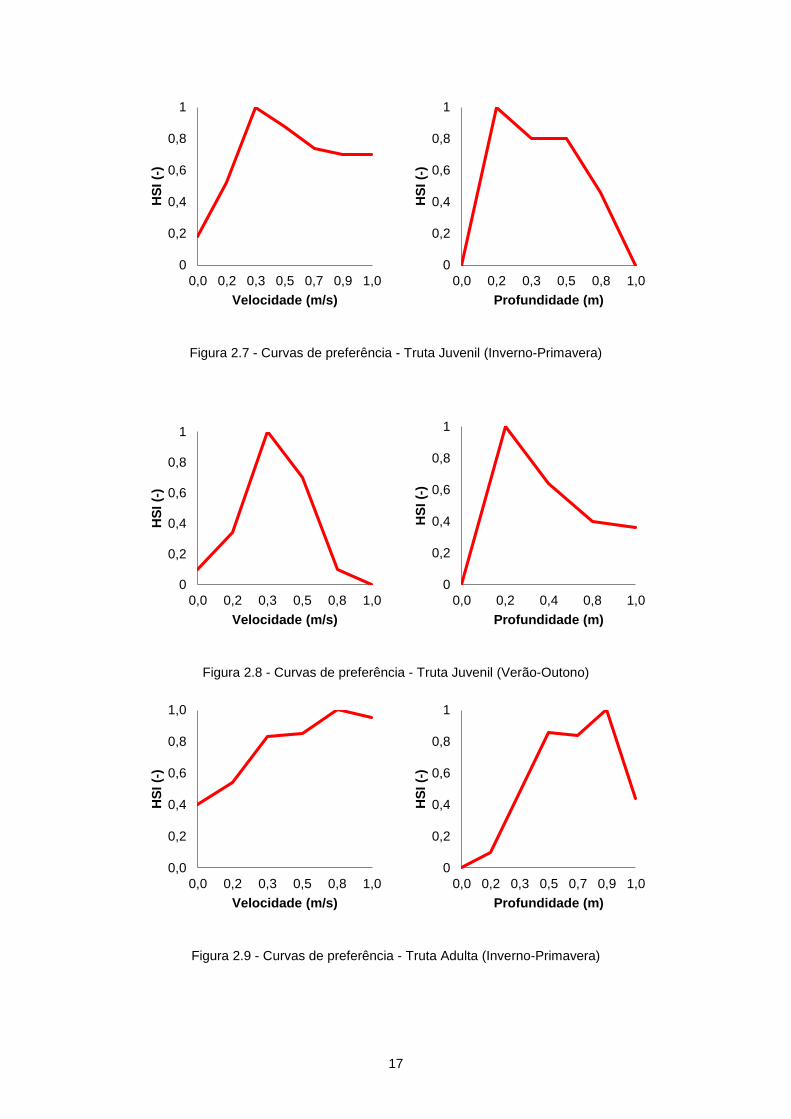
dois períodos do ano (Inverno-Primavera e Verão-Outono). As curvas de preferência desta
espécie utilizadas neste estudo foram determinadas para o rio Lima (Cortes et al., 2000) e
podem ser observadas da Figura 2.7 até à Figura 2.10.
17
Figura 2.7 - Curvas de preferência - Truta Juvenil (Inverno-Primavera)
Figura 2.8 - Curvas de preferência - Truta Juvenil (Verão-Outono)
Figura 2.9 - Curvas de preferência - Truta Adulta (Inverno-Primavera)
0
0,2
0,4
0,6
0,8
1
0,0 0,2 0,3 0,5 0,7 0,9 1,0
HS
I (-
)
Velocidade (m/s)
0
0,2
0,4
0,6
0,8
1
0,0 0,2 0,3 0,5 0,8 1,0
HS
I (-
)
Profundidade (m)
0
0,2
0,4
0,6
0,8
1
0,0 0,2 0,3 0,5 0,8 1,0
HS
I (-
)
Velocidade (m/s)
0
0,2
0,4
0,6
0,8
1
0,0 0,2 0,4 0,8 1,0
HS
I (-
)
Profundidade (m)
0,0
0,2
0,4
0,6
0,8
1,0
0,0 0,2 0,3 0,5 0,8 1,0
HS
I (-
)
Velocidade (m/s)
0
0,2
0,4
0,6
0,8
1
0,0 0,2 0,3 0,5 0,7 0,9 1,0
HS
I (-
)
Profundidade (m)

18
Figura 2.10 - Curvas de preferência - Truta Adulta (Verão-Outono)
Importa referir que a preferência desta espécie relativa ao substrato não é apresentada aqui
uma vez que se considerou uma preferência máxima (igual a 1) para qualquer classe de
substrato, conforme foi referido.
2.3.2 BARBO
O barbo-comum (Luciobarbus bocagei) (Figura 2.11) pode ser encontrado nos troços médios e
inferiores dos rios. Prefere zonas com pouca ou moderada velocidade de corrente,
exceptuando na época de reprodução. Em termos de habitat, o barbo adulto tem preferência
por troços mais profundos, com mais oxigénio e substrato fino. Em contrapartida, os juvenis
preferem zonas com alguma profundidade, desde que próximas da margem e sem corrente,
evitando zonas onde a luz solar não incida. Tem uma dimensão média menor que 50 cm, tal
como as trutas, e pode viver até 13 anos. É encontrado com maior frequência nas zonas
Centro e Norte de Portugal (Carta Piscícola, 2014).
Neste estudo os barbos foram divididos em dois estágios de vida – adultos e juvenis, sendo a
estação do ano analisada o Verão.
Figura 2.11 - Barbo-comum (DGRF)
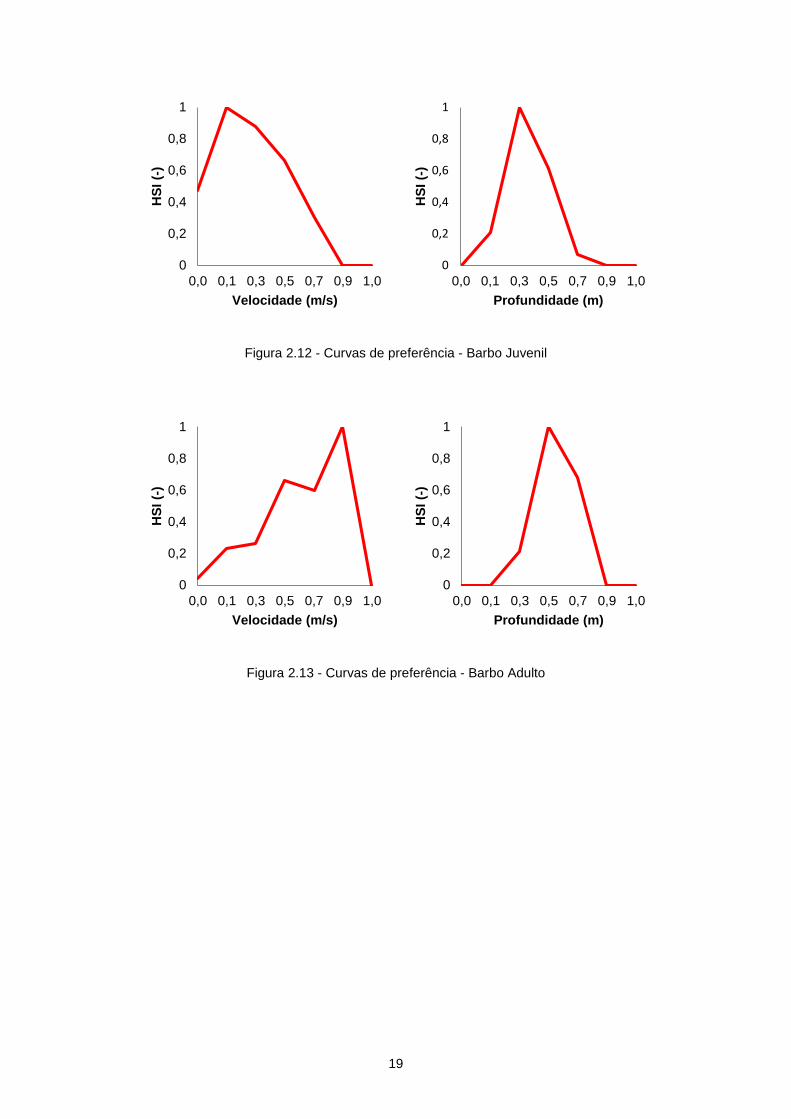
As curvas de preferência para o barbo utilizadas neste estudo foram determinadas para o rio
Ocreza (Boavida et al., 2013) e podem ser observadas na Figura 2.12 e na Figura 2.13.
0
0,2
0,4
0,6
0,8
1
0,0 0,2 0,3 0,5 0,7 0,9 1,0
HS
I (-
)
Velocidade (m/s)
0
0,2
0,4
0,6
0,8
1
0,0 0,2 0,3 0,7 1,0
HS
I (-
)
Profundidade (m)
19
Figura 2.12 - Curvas de preferência - Barbo Juvenil
Figura 2.13 - Curvas de preferência - Barbo Adulto
0
0,2
0,4
0,6
0,8
1
0,0 0,1 0,3 0,5 0,7 0,9 1,0
HS
I (-
)
Velocidade (m/s)
0
0,2
0,4
0,6
0,8
1
0,0 0,1 0,3 0,5 0,7 0,9 1,0
HS
I (-
)
Profundidade (m)
0
0,2
0,4
0,6
0,8
1
0,0 0,1 0,3 0,5 0,7 0,9 1,0
HS
I (-
)
Velocidade (m/s)
0
0,2
0,4
0,6
0,8
1
0,0 0,1 0,3 0,5 0,7 0,9 1,0
HS
I (-
)
Profundidade (m)

20

21
3 MODELAÇÃO NUMÉRICA
3.1 CALIBRAÇÃO DO MODELO NUMÉRICO
3.1.1 CONSIDERAÇÕES GERAIS
De forma a calibrar ou validar um modelo numérico é necessário ter como referência uma
situação aproximadamente real. Assim, utilizou-se a instalação experimental de Ribi (2011)
como base para este estudo. Foram analisados os resultados das simulações físicas e
posteriormente comparados com os das simulações computacionais.
3.1.2 INSTALAÇÃO EXPERIMENTAL
3.2.1.1 Introdução
O canal testado por Ribi (2011) e utilizado neste estudo é um canal de secção rectangular com
cerca de 12 m de comprimento e 1,20 m de largura, com um abrigo lateral na margem direita
de dimensões 2x1,2 m. O leito do canal é constituído essencialmente por seixo grosso e o
caudal testado foi de 0,22 m3/s. Na Figura 3.1 apresenta-se um esquema em planta da
instalação experimental e na Figura 3.2 um esquema tridimensional do canal. Mais detalhes
sobre a instalação experimental podem ser encontrados em Ribi (2011).
Figura 3.1 - Esquema em planta da instalação experimental com obstáculo na zona do abrigo.
Figura 3.2 - Esquema 3D da instalação laboratorial (vista de montante)

22
No início de cada ensaio foi imposto um caudal de 0,02 m3/s. Em seguida, introduziram-se os
peixes e aumentou-se o caudal para 0,22 m3/s durante um período de 3 h. Durante esse
período, a cada 20 min, a posição dos peixes no canal foi registada visualmente.
Foram testadas doze configurações para o abrigo lateral; uma configuração base (C0) sem
qualquer obstáculo no abrigo e onze configurações com recurso a obstáculos formados por
placas verticais com 0,03 m de espessura, inseridas no fundo do canal e atingindo uma cota
superior à da superfície livre do escoamento. Os obstáculos tiveram como objectivo criar
campos de velocidades favoráveis para os peixes, atraindo-os para o interior do abrigo. De
referir que nestes ensaios experimentais, realizados na Primavera, apenas foram utilizadas
trutas juvenis. As trutas juvenis foram capturadas através de pesca eléctrica e cada um dos
peixes foi sujeito em média a cerca de 18 horas de ensaios. Mais detalhes sobre os ensaios
experimentais podem ser encontrados em Ribi (2011).
3.2.1.2 Configurações testadas
O obstáculo no abrigo tomou várias posições e formas dentro de uma área de referência de
1,16 m2, representada na Figura 3.3. As configurações testadas, podem ser observadas na
Figura 3.4.
Figura 3.3 - Área de referência (a tracejado) das configurações testadas
(adaptado de Ribi (2011))
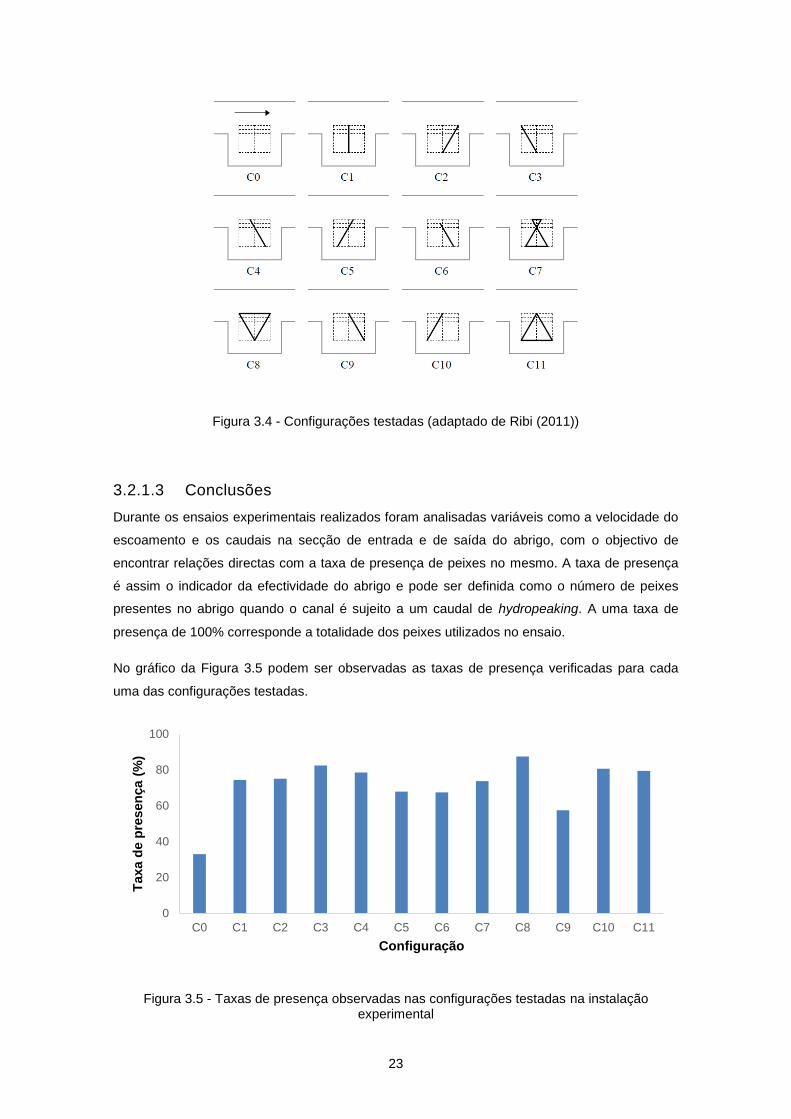
23
Figura 3.4 - Configurações testadas (adaptado de Ribi (2011))
3.2.1.3 Conclusões
Durante os ensaios experimentais realizados foram analisadas variáveis como a velocidade do
escoamento e os caudais na secção de entrada e de saída do abrigo, com o objectivo de
encontrar relações directas com a taxa de presença de peixes no mesmo. A taxa de presença
é assim o indicador da efectividade do abrigo e pode ser definida como o número de peixes
presentes no abrigo quando o canal é sujeito a um caudal de hydropeaking. A uma taxa de
presença de 100% corresponde a totalidade dos peixes utilizados no ensaio.
No gráfico da Figura 3.5 podem ser observadas as taxas de presença verificadas para cada
uma das configurações testadas.
Figura 3.5 - Taxas de presença observadas nas configurações testadas na instalação experimental
0
20
40
60
80
100
C0 C1 C2 C3 C4 C5 C6 C7 C8 C9 C10 C11
Taxa d
e p
resen
ça (
%)
Configuração

24
Concluiu-se assim que as quatro configurações mais eficientes (isto é, com uma taxa de
presença mais elevada) foram, por ordem decrescente, as configurações C8, C3, C10 e C11.
Importa também referir a configuração com piores resultados verificados, a configuração C9.
3.2.2 MODELO NUMÉRICO
3.2.2.1 Introdução
De forma a poder concluir que este modelo numérico permite representar de uma forma
correcta a disponibilidade de habitat existente para os peixes é necessário comparar os
resultados obtidos por este meio com os obtidos através da instalação experimental. Sendo os
resultados da instalação experimental expressos em taxas de presença, seria de esperar que
as configurações associadas a taxas maiores tivessem maiores valores de HHS no modelo
numérico.
3.2.2.2 Condições de simulação
Em termos hidráulicos (caudais e alturas de escoamento) as condições de simulação foram
idênticas às da instalação experimental. Foi assim simulado um caudal de 0,22 m3/s, sendo a
altura do escoamento na secção de jusante do canal de 0,24 m. Do ponto de vista
computacional é necessário descrever a malha de elementos finitos utilizada.
Utilizou-se uma malha semelhante para todas as configurações testadas, com um
espaçamento de 0,10 m entre nós. O número de nós existentes varia, sendo directamente
influenciado pela configuração do obstáculo a testar. Uma das malhas utilizadas pode ser
observada na Figura 3.6
Figura 3.6 - Malha de elementos finitos. Configuração C8 (River2D)
As características de cada malha utilizada podem ser observadas no Anexo A – Malhas de
elementos finitos. Configurações C.
3.3 ANÁLISE DE RESULTADOS
Após as simulações com recurso ao River2D foram obtidos os campos de velocidade e
distribuições de CSI para cada caso testado. Assim foi possível calcular o valor de WUA
disponível em cada uma destas configurações. Dividindo este valor pela área total ocupada
pelo escoamento obteve-se então o índice HHS. De referir que a área analisada (Figura 3.7) é
delimitada pela secção imediatamente a montante do abrigo até à secção mais a jusante do
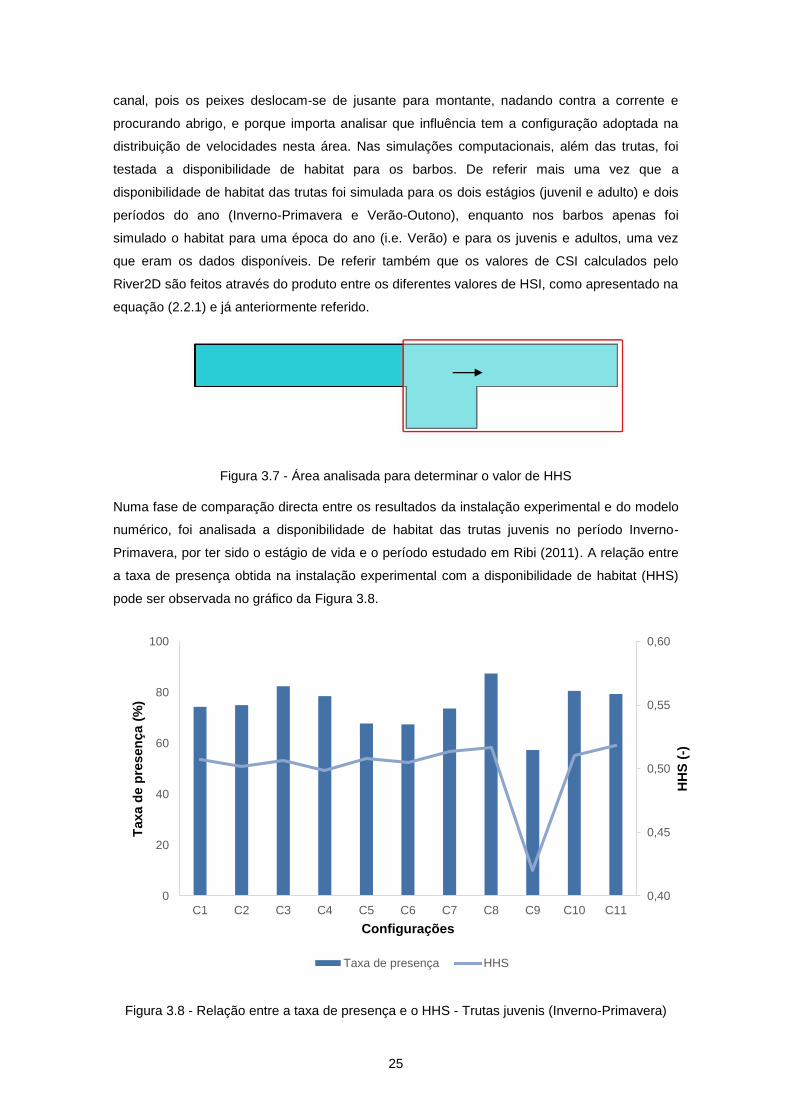
25
canal, pois os peixes deslocam-se de jusante para montante, nadando contra a corrente e
procurando abrigo, e porque importa analisar que influência tem a configuração adoptada na
distribuição de velocidades nesta área. Nas simulações computacionais, além das trutas, foi
testada a disponibilidade de habitat para os barbos. De referir mais uma vez que a
disponibilidade de habitat das trutas foi simulada para os dois estágios (juvenil e adulto) e dois
períodos do ano (Inverno-Primavera e Verão-Outono), enquanto nos barbos apenas foi
simulado o habitat para uma época do ano (i.e. Verão) e para os juvenis e adultos, uma vez
que eram os dados disponíveis. De referir também que os valores de CSI calculados pelo
River2D são feitos através do produto entre os diferentes valores de HSI, como apresentado na
equação (2.2.1) e já anteriormente referido.
Figura 3.7 - Área analisada para determinar o valor de HHS
Numa fase de comparação directa entre os resultados da instalação experimental e do modelo
numérico, foi analisada a disponibilidade de habitat das trutas juvenis no período Inverno-
Primavera, por ter sido o estágio de vida e o período estudado em Ribi (2011). A relação entre
a taxa de presença obtida na instalação experimental com a disponibilidade de habitat (HHS)
pode ser observada no gráfico da Figura 3.8.
Figura 3.8 - Relação entre a taxa de presença e o HHS - Trutas juvenis (Inverno-Primavera)
0,40
0,45
0,50
0,55
0,60
0
20
40
60
80
100
C1 C2 C3 C4 C5 C6 C7 C8 C9 C10 C11
HH
S (
-)
Taxa d
e p
resen
ça (
%)
Configurações
Taxa de presença HHS
26
Através da análise do gráfico da Figura 3.8 pode-se concluir que, de uma forma geral, o HHS
assemelha-se aos valores de taxa de presença para cada configuração, acompanhando os
aumentos e os decréscimos da mesma. Excepção feita talvez às configurações C5 e C6 que,
comparativamente com as outras, não acompanham a tendência de diminuição da taxa de
presença verificada, aumentando os valores de HHS. No entanto, as quatro melhores
configurações em termos de taxa de presença (C8, C3, C10 e C11, por ordem decrescente)
são também as quatro configurações que registam valores de HHS mais altos.
Na Tabela 3.1 são apresentados os valores de WUA e de HHS para seis das configurações
referidas testadas em instalação experimental, para todas as espécies, estágios de vida e
períodos do ano estudados. São apresentados os valores para a configuração base (C0, sem
obstáculo), para a pior configuração (C9) e para as quatro melhores configurações, em termos
de taxas de presença. Pode concluir-se que, tal como esperado, as melhores configurações
têm valores de HHS maiores, não só para as trutas como também para os barbos.
Tabela 3.1 - Valores de HHS e WUA para as configurações testadas na instalação
experimental
Truta
Inverno-Primavera
Truta
Verão-Outono Barbo
Juvenil Adulta Juvenil Adulta Juvenil Adulto
Configuração WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
C0 4.86 0.51 2.53 0.26 3.41 0.36 3.57 0.37 4.20 0.44 0.57 0.06
C9 4.01 0.42 2.79 0.29 1.06 0.11 3.62 0.38 1.06 0.11 0.55 0.06
C3 4.83 0.51 2.74 0.29 3.63 0.38 3.69 0.39 4.39 0.46 0.67 0.07
C8 4.65 0.52 2.61 0.29 3.46 0.38 3.57 0.40 3.96 0.44 0.65 0.07
C10 4.87 0.51 2.73 0.29 3.71 0.39 3.78 0.40 4.42 0.46 0.70 0.07
C11 4.67 0.52 2.64 0.29 3.52 0.39 3.63 0.40 4.00 0.44 0.68 0.08
No gráfico da Figura 3.9 são apresentados, juntamente com a taxa de presença, os valores de
HHS para cada umas destas configurações.
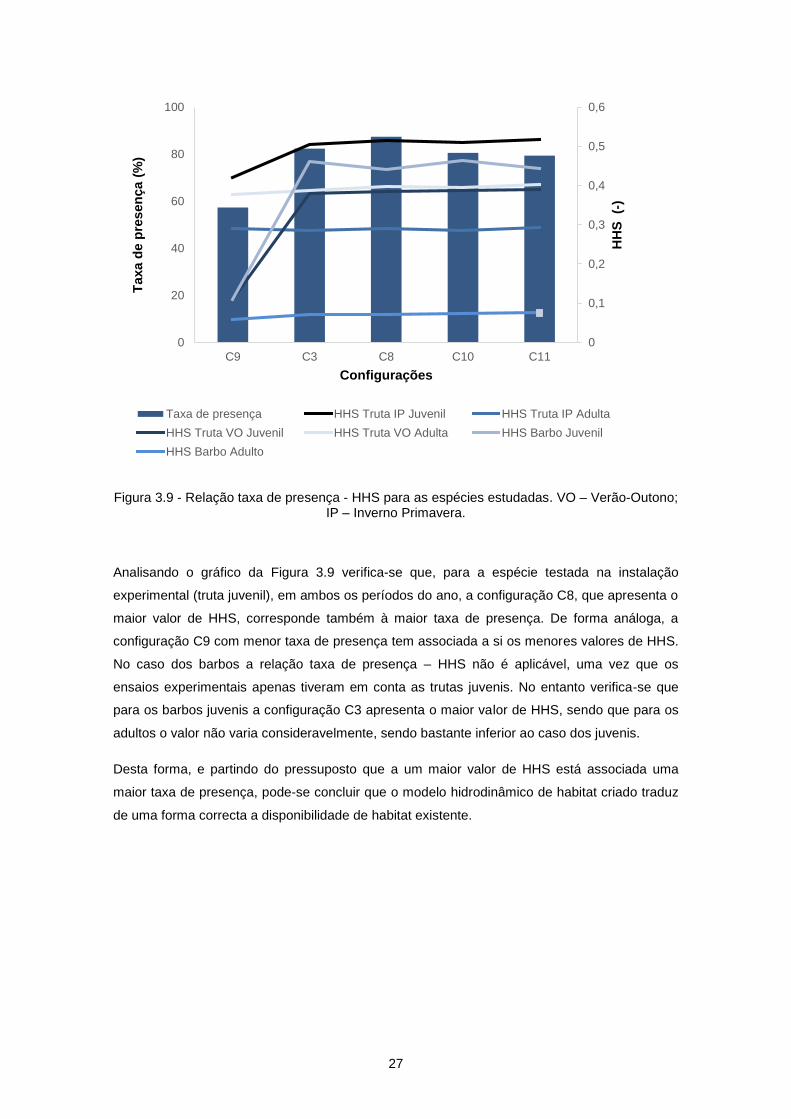
27
Figura 3.9 - Relação taxa de presença - HHS para as espécies estudadas. VO – Verão-Outono; IP – Inverno Primavera.
Analisando o gráfico da Figura 3.9 verifica-se que, para a espécie testada na instalação
experimental (truta juvenil), em ambos os períodos do ano, a configuração C8, que apresenta o
maior valor de HHS, corresponde também à maior taxa de presença. De forma análoga, a
configuração C9 com menor taxa de presença tem associada a si os menores valores de HHS.
No caso dos barbos a relação taxa de presença – HHS não é aplicável, uma vez que os
ensaios experimentais apenas tiveram em conta as trutas juvenis. No entanto verifica-se que
para os barbos juvenis a configuração C3 apresenta o maior valor de HHS, sendo que para os
adultos o valor não varia consideravelmente, sendo bastante inferior ao caso dos juvenis.
Desta forma, e partindo do pressuposto que a um maior valor de HHS está associada uma
maior taxa de presença, pode-se concluir que o modelo hidrodinâmico de habitat criado traduz
de uma forma correcta a disponibilidade de habitat existente.
0
0,1
0,2
0,3
0,4
0,5
0,6
0
20
40
60
80
100
C9 C3 C8 C10 C11
HH
S
(-)
Taxa d
e p
resen
ça (
%)
Configurações
Taxa de presença HHS Truta IP Juvenil HHS Truta IP Adulta
HHS Truta VO Juvenil HHS Truta VO Adulta HHS Barbo Juvenil
HHS Barbo Adulto

28

29
4 ANÁLISE DE NOVAS CONFIGURAÇÕES PARA OS ABRIGOS
LATERAIS
4.2 CONFIGURAÇÕES CRIADAS
De forma a procurar aumentar os valores de HHS para o abrigo, criaram-se novas
configurações para o obstáculo e para o próprio abrigo. Foram criadas onze configurações,
designadas de A a K. As configurações criadas podem ser observadas na Figura 4.1.
Figura 4.1 – Novas configurações para o abrigo
Tomando sempre como referência a área a tracejado cujas dimensões já foram apresentadas
na Figura 3.3, procedeu-se a alterações na configuração do obstáculo, tendo por base as 4
configurações C com melhores resultados: C8, C3, C10 e C11. Nas configurações C e D as
dimensões do abrigo foram alteradas. Na Tabela 4.1 é apresentada uma breve descrição das
configurações bem como dos motivos que levaram à sua projecção. No Anexo B são descritas
as malhas de elementos finitos utilizadas nestas configurações.
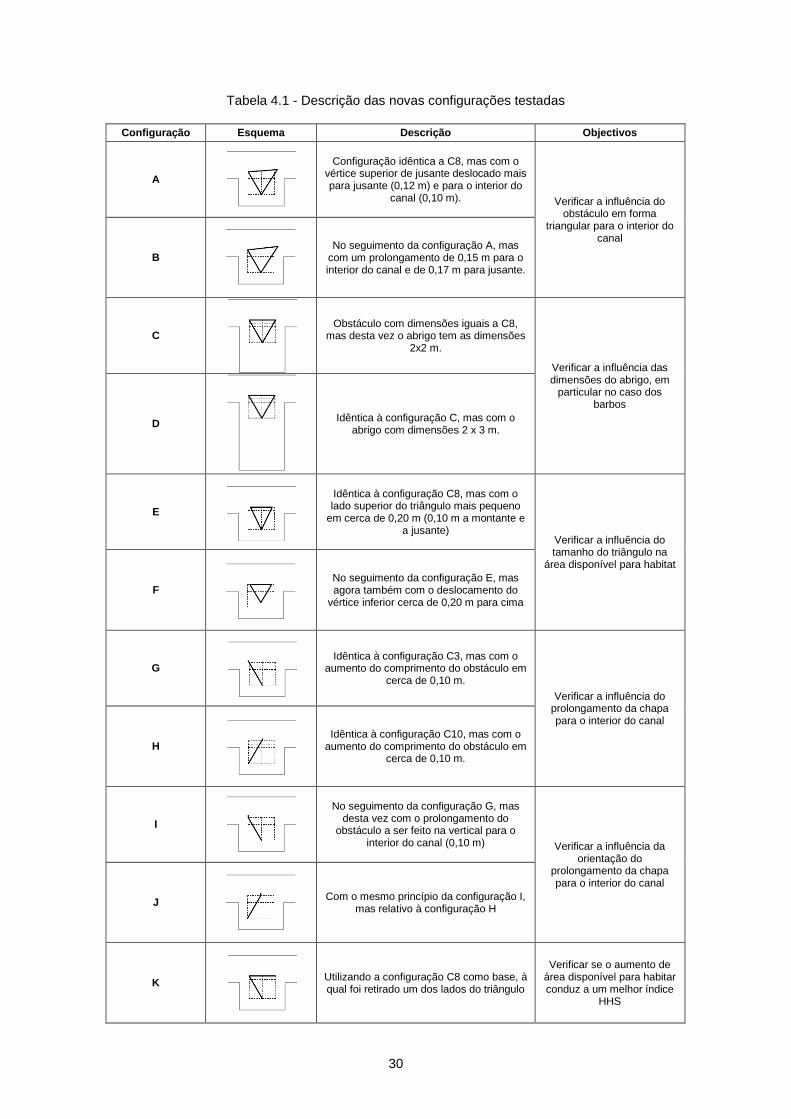
30
Tabela 4.1 - Descrição das novas configurações testadas
Configuração Esquema Descrição Objectivos
A
Configuração idêntica a C8, mas com o vértice superior de jusante deslocado mais para jusante (0,12 m) e para o interior do
canal (0,10 m). Verificar a influência do obstáculo em forma
triangular para o interior do canal
B
No seguimento da configuração A, mas com um prolongamento de 0,15 m para o interior do canal e de 0,17 m para jusante.
C
Obstáculo com dimensões iguais a C8, mas desta vez o abrigo tem as dimensões
2x2 m.
Verificar a influência das dimensões do abrigo, em
particular no caso dos barbos
D
Idêntica à configuração C, mas com o abrigo com dimensões 2 x 3 m.
E
Idêntica à configuração C8, mas com o lado superior do triângulo mais pequeno
em cerca de 0,20 m (0,10 m a montante e a jusante)
Verificar a influência do tamanho do triângulo na
área disponível para habitat
F
No seguimento da configuração E, mas agora também com o deslocamento do
vértice inferior cerca de 0,20 m para cima
G
Idêntica à configuração C3, mas com o aumento do comprimento do obstáculo em
cerca de 0,10 m.
Verificar a influência do prolongamento da chapa para o interior do canal
H
Idêntica à configuração C10, mas com o aumento do comprimento do obstáculo em
cerca de 0,10 m.
I
No seguimento da configuração G, mas desta vez com o prolongamento do
obstáculo a ser feito na vertical para o interior do canal (0,10 m) Verificar a influência da
orientação do prolongamento da chapa para o interior do canal
J
Com o mesmo princípio da configuração I, mas relativo à configuração H
K
Utilizando a configuração C8 como base, à qual foi retirado um dos lados do triângulo
Verificar se o aumento de área disponível para habitar conduz a um melhor índice
HHS

31
4.3 ANÁLISE DE RESULTADOS
4.3.1 VALORES DE HHS E WUA PARA AS CONFIGURAÇÕES CRIADAS
Nas configurações estudadas, e sendo o caudal constante em todas as simulações, a
profundidade não varia consideravelmente no troço analisado (do início do abrigo até ao final
do canal, a jusante) como se pode observar na Figura 4.2. Tendo em conta essa evolução
pode ser tomada como referência a configuração C0 (sem qualquer obstáculo no abrigo).
Desta forma pode concluir-se que o principal factor que afectou o CSI foi a velocidade do
escoamento (traduzido no índice VSI), visto que a influência do substrato também foi ignorada
(ou seja, atribuída a preferência máxima (HSI=1,0) a todo o canal).
Figura 4.2 - Profundidades (m) na configuração C0
No troço estudado a profundidade varia entre 0,27 e 0,24 m. Os valores de HSI aproximados
para as alturas de escoamento compreendidas nesse intervalo relativos às espécies analisadas
encontram-se na Tabela 4.2.
Tabela 4.2 - Valores aproximados de HSI para as alturas de escoamento no intervalo
considerado – Rio Lima
Intervalo de valores HSI (-) para profundidades entre
0,24 e 0,27 m (aproximado)
Truta
Inverno Primavera
Juvenil 0,48 a 1,00
Adulta 0,10 a 0,48
Verão Outono
Juvenil 0,82 a 1,00
Adulta 0,42 a 0,90
Barbo
Juvenil 0,60 a 1,00
Adulto 0,10 a 0,21
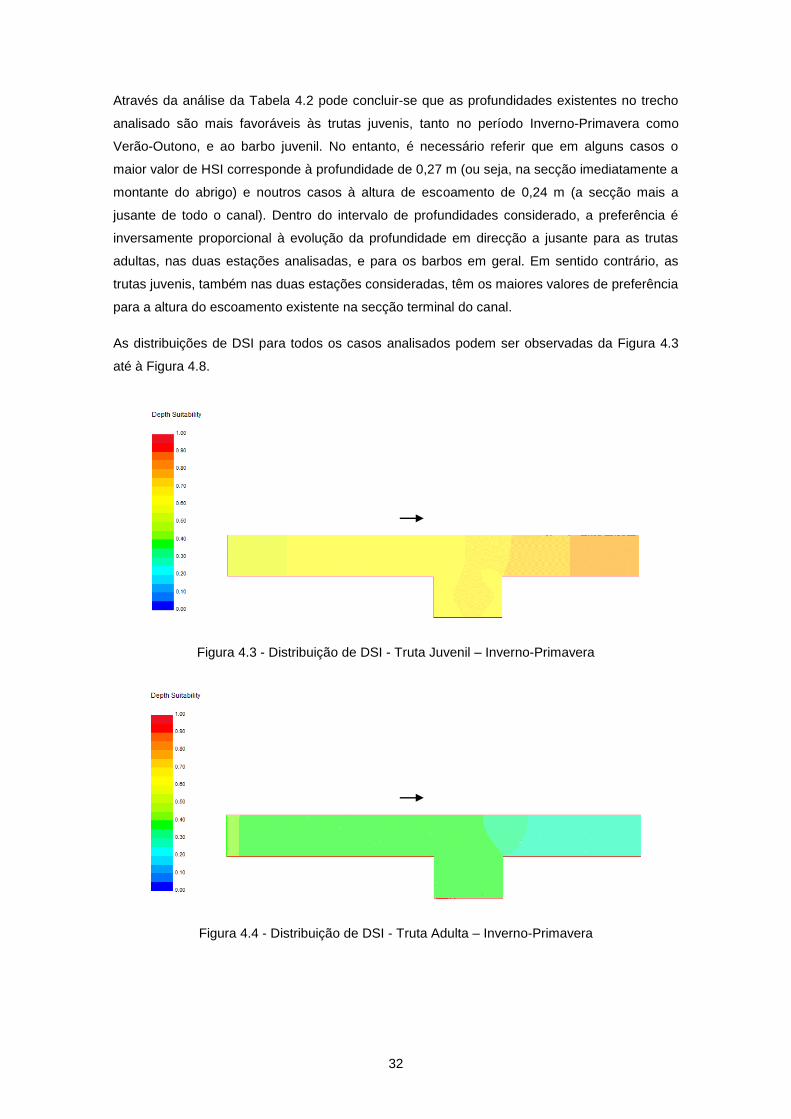
32
Através da análise da Tabela 4.2 pode concluir-se que as profundidades existentes no trecho
analisado são mais favoráveis às trutas juvenis, tanto no período Inverno-Primavera como
Verão-Outono, e ao barbo juvenil. No entanto, é necessário referir que em alguns casos o
maior valor de HSI corresponde à profundidade de 0,27 m (ou seja, na secção imediatamente a
montante do abrigo) e noutros casos à altura de escoamento de 0,24 m (a secção mais a
jusante de todo o canal). Dentro do intervalo de profundidades considerado, a preferência é
inversamente proporcional à evolução da profundidade em direcção a jusante para as trutas
adultas, nas duas estações analisadas, e para os barbos em geral. Em sentido contrário, as
trutas juvenis, também nas duas estações consideradas, têm os maiores valores de preferência
para a altura do escoamento existente na secção terminal do canal.
As distribuições de DSI para todos os casos analisados podem ser observadas da Figura 4.3
até à Figura 4.8.
Figura 4.3 - Distribuição de DSI - Truta Juvenil – Inverno-Primavera
Figura 4.4 - Distribuição de DSI - Truta Adulta – Inverno-Primavera
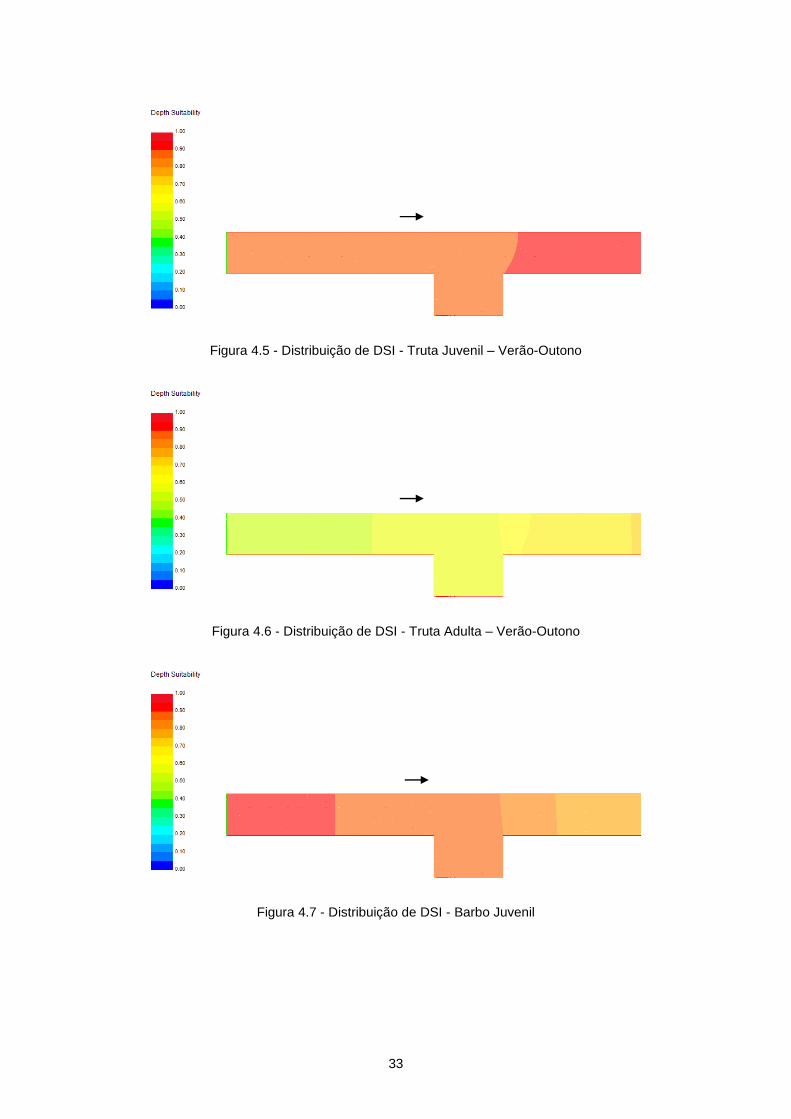
33
Figura 4.5 - Distribuição de DSI - Truta Juvenil – Verão-Outono
Figura 4.6 - Distribuição de DSI - Truta Adulta – Verão-Outono
Figura 4.7 - Distribuição de DSI - Barbo Juvenil

34
Figura 4.8 - Distribuição de DSI - Barbo Adulto
A consideração de valores aproximados para o HSI na Tabela 4.2 pode parecer imprecisa, mas
decorre da discretização das curvas de preferência utilizadas. Por exemplo, são descritos
valores de HSI para profundidades de 0,20 e 0,30 m, não sendo apresentado o valor específico
de HSI para profundidades de 0,24 e 0,27 m. Também não é possível assumir uma linearidade
dos valores de HSI dentro desse intervalo de profundidades. No entanto, para uma análise
comparativa entre estas duas secções esta aproximação é válida, pois importa apenas
perceber como varia a preferência das espécies em termos de profundidade ao longo do troço
analisado; percebendo se as espécies se sentem mais confortáveis a montante ou a jusante.
Uma análise idêntica em termos de velocidades terá de ser mais exaustiva visto que a
existência de obstáculos no abrigo altera as distribuições de velocidades desde o abrigo até à
secção de jusante. Assim, e percebendo qualitativamente como varia a preferência em termos
de profundidade, a importância da velocidade será analisada com maior pormenor de seguida
nas configurações que obtiveram melhores valores de HHS, estando implícita a sua
importância nos valores de CSI obtidos. De certa forma pode concluir-se que as diferentes
configurações para o abrigo afectam mais a velocidade do escoamento do que a profundidade
e que os diferentes resultados obtidos têm como principal factor o VSI.
Os resultados dos valores de WUA e de HHS são apresentados na Tabela 4.3. A negrito são
assinalados os maiores valores de HHS para cada uma das situações.
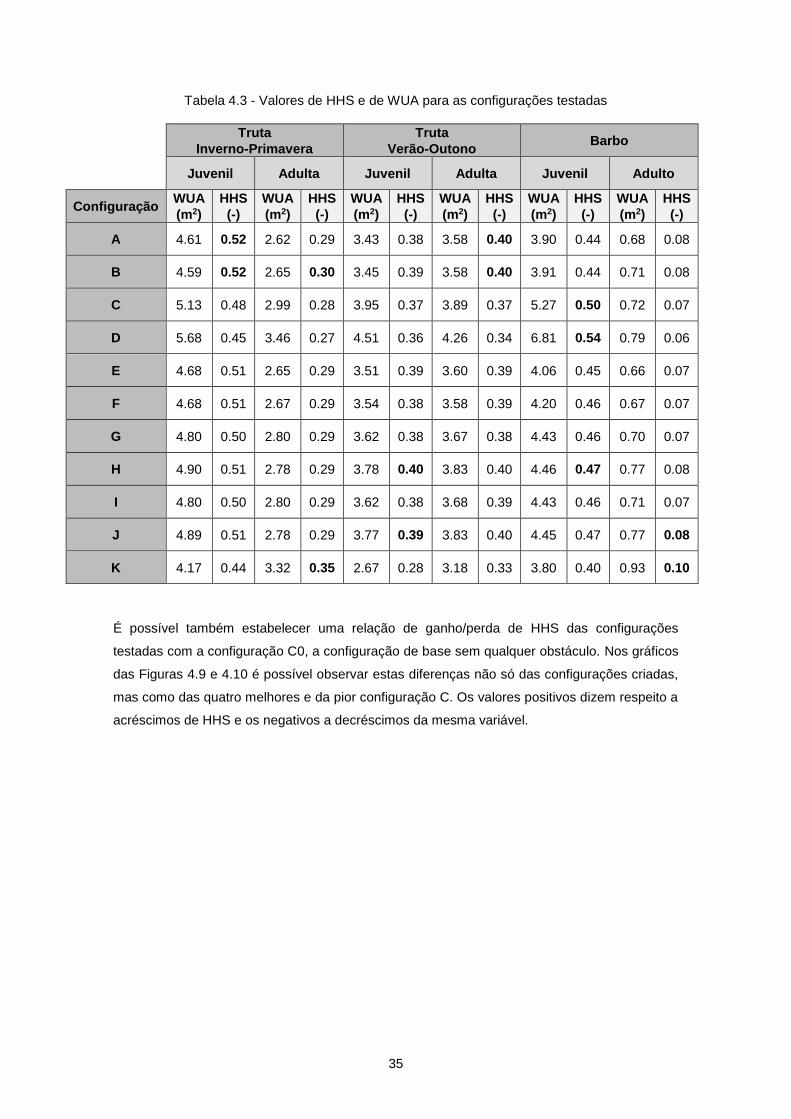
35
Tabela 4.3 - Valores de HHS e de WUA para as configurações testadas
Truta
Inverno-Primavera
Truta
Verão-Outono Barbo
Juvenil Adulta Juvenil Adulta Juvenil Adulto
Configuração WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
WUA
(m2)
HHS
(-)
A 4.61 0.52 2.62 0.29 3.43 0.38 3.58 0.40 3.90 0.44 0.68 0.08
B 4.59 0.52 2.65 0.30 3.45 0.39 3.58 0.40 3.91 0.44 0.71 0.08
C 5.13 0.48 2.99 0.28 3.95 0.37 3.89 0.37 5.27 0.50 0.72 0.07
D 5.68 0.45 3.46 0.27 4.51 0.36 4.26 0.34 6.81 0.54 0.79 0.06
E 4.68 0.51 2.65 0.29 3.51 0.39 3.60 0.39 4.06 0.45 0.66 0.07
F 4.68 0.51 2.67 0.29 3.54 0.38 3.58 0.39 4.20 0.46 0.67 0.07
G 4.80 0.50 2.80 0.29 3.62 0.38 3.67 0.38 4.43 0.46 0.70 0.07
H 4.90 0.51 2.78 0.29 3.78 0.40 3.83 0.40 4.46 0.47 0.77 0.08
I 4.80 0.50 2.80 0.29 3.62 0.38 3.68 0.39 4.43 0.46 0.71 0.07
J 4.89 0.51 2.78 0.29 3.77 0.39 3.83 0.40 4.45 0.47 0.77 0.08
K 4.17 0.44 3.32 0.35 2.67 0.28 3.18 0.33 3.80 0.40 0.93 0.10
É possível também estabelecer uma relação de ganho/perda de HHS das configurações
testadas com a configuração C0, a configuração de base sem qualquer obstáculo. Nos gráficos
das Figuras 4.9 e 4.10 é possível observar estas diferenças não só das configurações criadas,
mas como das quatro melhores e da pior configuração C. Os valores positivos dizem respeito a
acréscimos de HHS e os negativos a decréscimos da mesma variável.
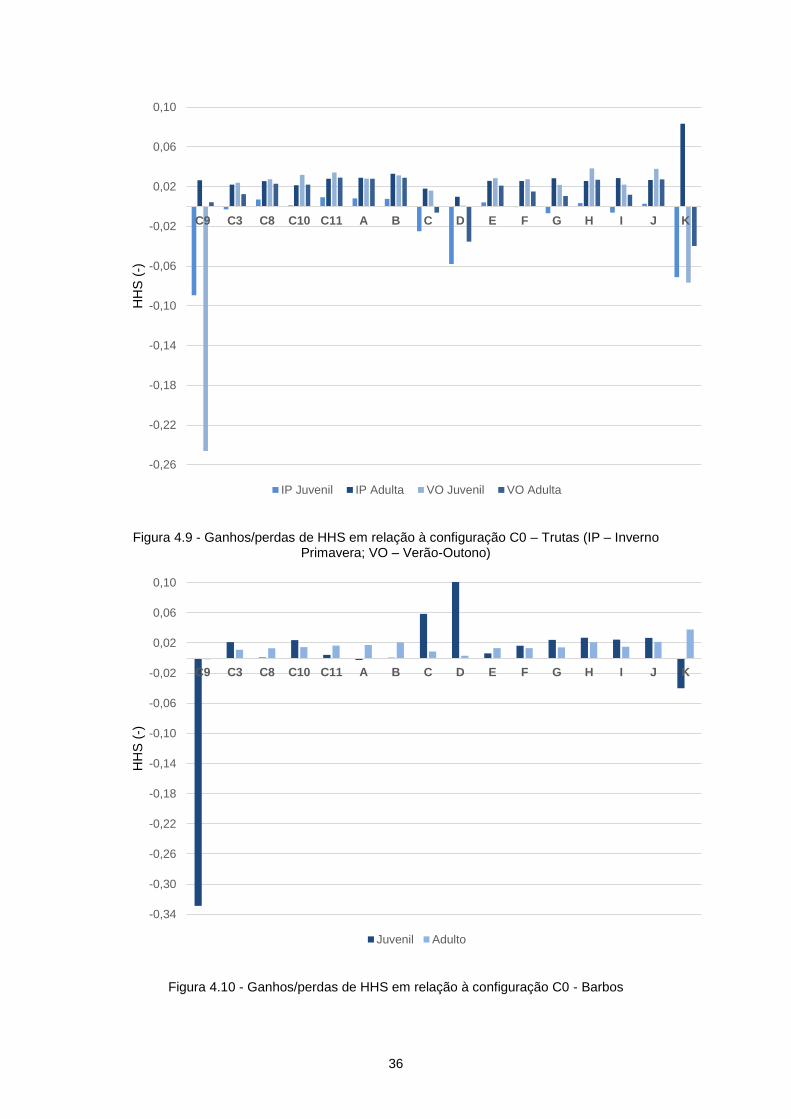
36
Figura 4.9 - Ganhos/perdas de HHS em relação à configuração C0 – Trutas (IP – Inverno Primavera; VO – Verão-Outono)
Figura 4.10 - Ganhos/perdas de HHS em relação à configuração C0 - Barbos
-0,26
-0,22
-0,18
-0,14
-0,10
-0,06
-0,02
0,02
0,06
0,10
C9 C3 C8 C10 C11 A B C D E F G H I J K
HH
S (
-)
IP Juvenil IP Adulta VO Juvenil VO Adulta
-0,34
-0,30
-0,26
-0,22
-0,18
-0,14
-0,10
-0,06
-0,02
0,02
0,06
0,10
C9 C3 C8 C10 C11 A B C D E F G H I J K
HH
S (
-)
Juvenil Adulto

37
Relativamente às trutas a configuração K apresenta o maior acréscimo de HHS de todas as
configurações, mas apenas para o caso das trutas adultas no período de Inverno-Primavera.
Esta mesma configuração resulta num decréscimo significativo de HHS para os restantes
estágios/períodos do ano. As configurações com maiores acréscimos nos quatro casos
relativos às trutas são as configurações A,B, H e J. De forma oposta a configuração C9 é
aquela que apresenta o maior decréscimo, no caso da truta juvenil – Verão-Outono.
No caso dos barbos a configuração C9 é também a que apresenta maiores decréscimos de
HHS, sendo as que apresentam maiores aumentos de HHS as configurações C,D,H e J. A
configuração K apresenta melhorias para os adultos, mas no caso dos juvenis os resultados
são mais desfavoráveis.
De seguida serão apresentados os resultados obtidos, divididos entre trutas e barbos. Em cada
uma das espécies serão apresentadas as configurações com melhores resultados de HHS e
será feita uma análise do CSI em cada caso.
4.3.2 ANÁLISE DOS RESULTADOS OBTIDOS PARA AS TRUTAS
4.3.2.1 Considerações gerais
As configurações que revelaram melhores resultados para esta espécie foram as configurações
A, B, H, J e K. Embora cada configuração tenha tido melhores resultados em diferentes
estágios ou estações do ano, as configurações A e B revelaram-se as melhores para um maior
número de casos, tendo obtido os maiores valores de HHS para as trutas juvenis no período
Inverno-Primavera e para as adultas no período Verão-Outono. A configuração B teve ainda o
valor mais elevado de HHS considerando a truta adulta no período Inverno-Primavera.
1. Configuração A
A configuração A (Figura 4.11), baseada na configuração C8, é caracterizada por ter o vértice
mais a jusante do triângulo ligeiramente deslocado para o interior do canal, comparativamente
com o vértice mais a montante.
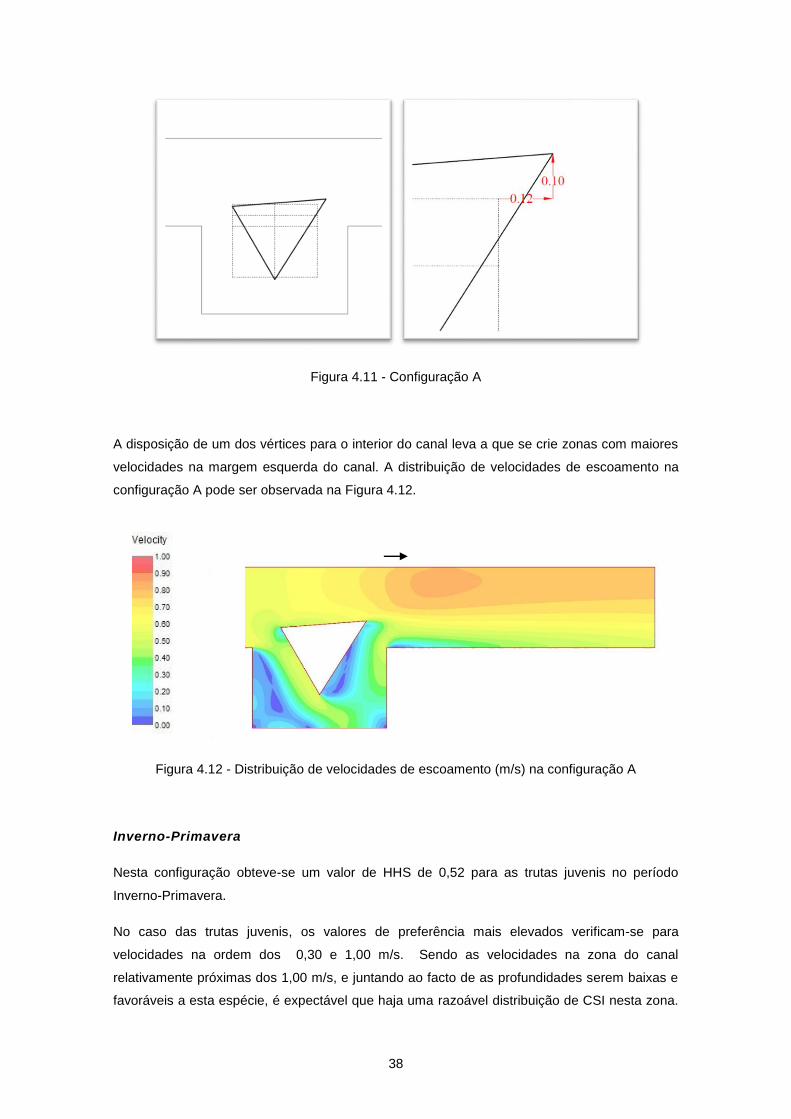
38
Figura 4.11 - Configuração A
A disposição de um dos vértices para o interior do canal leva a que se crie zonas com maiores
velocidades na margem esquerda do canal. A distribuição de velocidades de escoamento na
configuração A pode ser observada na Figura 4.12.
Figura 4.12 - Distribuição de velocidades de escoamento (m/s) na configuração A
Inverno-Primavera
Nesta configuração obteve-se um valor de HHS de 0,52 para as trutas juvenis no período
Inverno-Primavera.
No caso das trutas juvenis, os valores de preferência mais elevados verificam-se para
velocidades na ordem dos 0,30 e 1,00 m/s. Sendo as velocidades na zona do canal
relativamente próximas dos 1,00 m/s, e juntando ao facto de as profundidades serem baixas e
favoráveis a esta espécie, é expectável que haja uma razoável distribuição de CSI nesta zona.
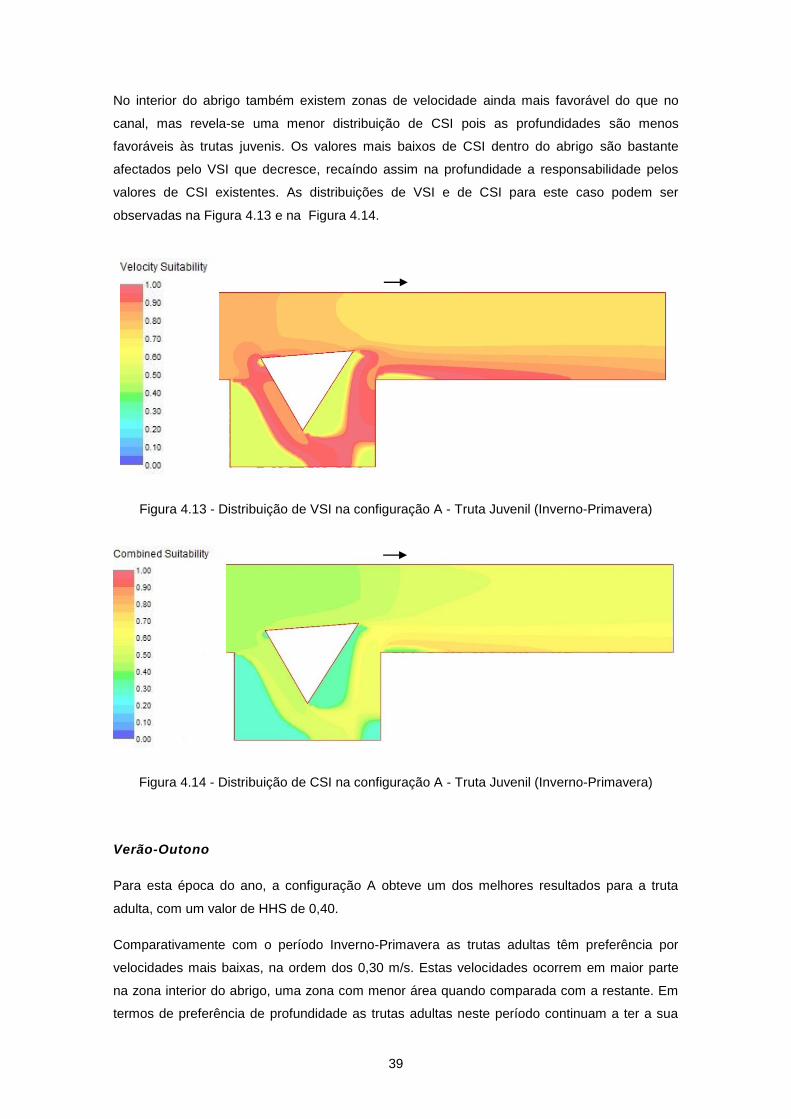
39
No interior do abrigo também existem zonas de velocidade ainda mais favorável do que no
canal, mas revela-se uma menor distribuição de CSI pois as profundidades são menos
favoráveis às trutas juvenis. Os valores mais baixos de CSI dentro do abrigo são bastante
afectados pelo VSI que decresce, recaíndo assim na profundidade a responsabilidade pelos
valores de CSI existentes. As distribuições de VSI e de CSI para este caso podem ser
observadas na Figura 4.13 e na Figura 4.14.
Figura 4.13 - Distribuição de VSI na configuração A - Truta Juvenil (Inverno-Primavera)
Figura 4.14 - Distribuição de CSI na configuração A - Truta Juvenil (Inverno-Primavera)
Verão-Outono
Para esta época do ano, a configuração A obteve um dos melhores resultados para a truta
adulta, com um valor de HHS de 0,40.
Comparativamente com o período Inverno-Primavera as trutas adultas têm preferência por
velocidades mais baixas, na ordem dos 0,30 m/s. Estas velocidades ocorrem em maior parte
na zona interior do abrigo, uma zona com menor área quando comparada com a restante. Em
termos de preferência de profundidade as trutas adultas neste período continuam a ter a sua
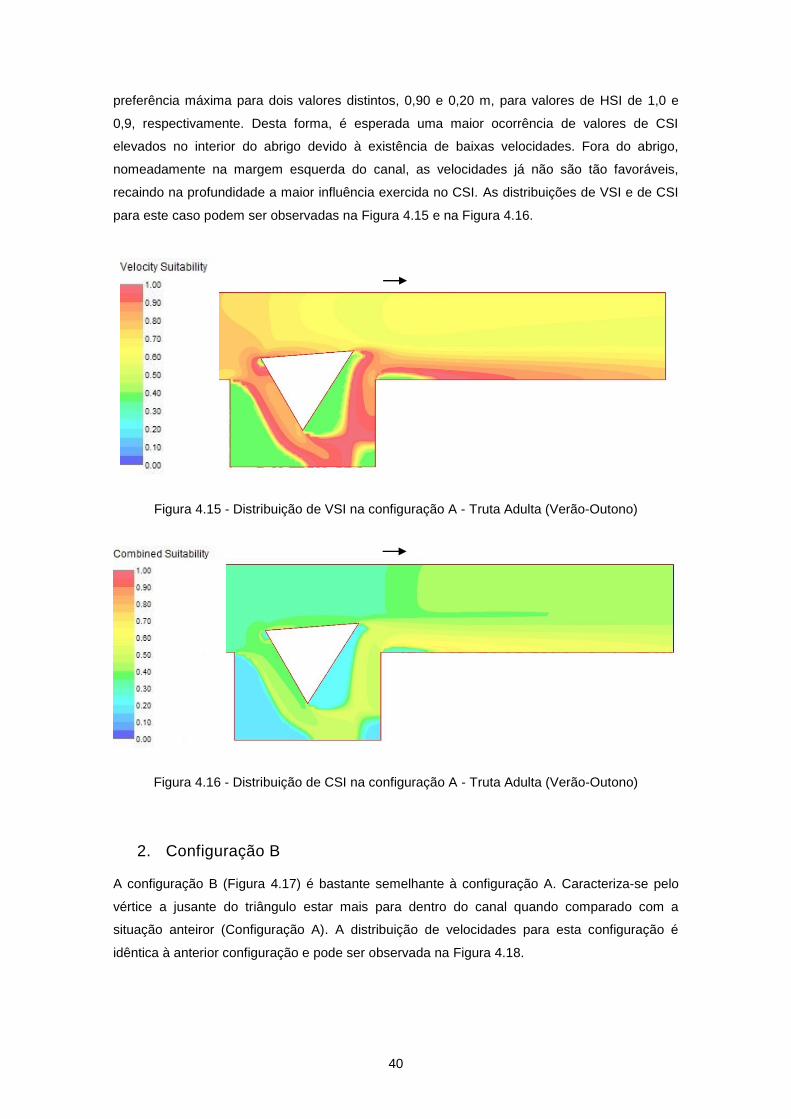
40
preferência máxima para dois valores distintos, 0,90 e 0,20 m, para valores de HSI de 1,0 e
0,9, respectivamente. Desta forma, é esperada uma maior ocorrência de valores de CSI
elevados no interior do abrigo devido à existência de baixas velocidades. Fora do abrigo,
nomeadamente na margem esquerda do canal, as velocidades já não são tão favoráveis,
recaindo na profundidade a maior influência exercida no CSI. As distribuições de VSI e de CSI
para este caso podem ser observadas na Figura 4.15 e na Figura 4.16.
Figura 4.15 - Distribuição de VSI na configuração A - Truta Adulta (Verão-Outono)
Figura 4.16 - Distribuição de CSI na configuração A - Truta Adulta (Verão-Outono)
2. Configuração B
A configuração B (Figura 4.17) é bastante semelhante à configuração A. Caracteriza-se pelo
vértice a jusante do triângulo estar mais para dentro do canal quando comparado com a
situação anteiror (Configuração A). A distribuição de velocidades para esta configuração é
idêntica à anterior configuração e pode ser observada na Figura 4.18.
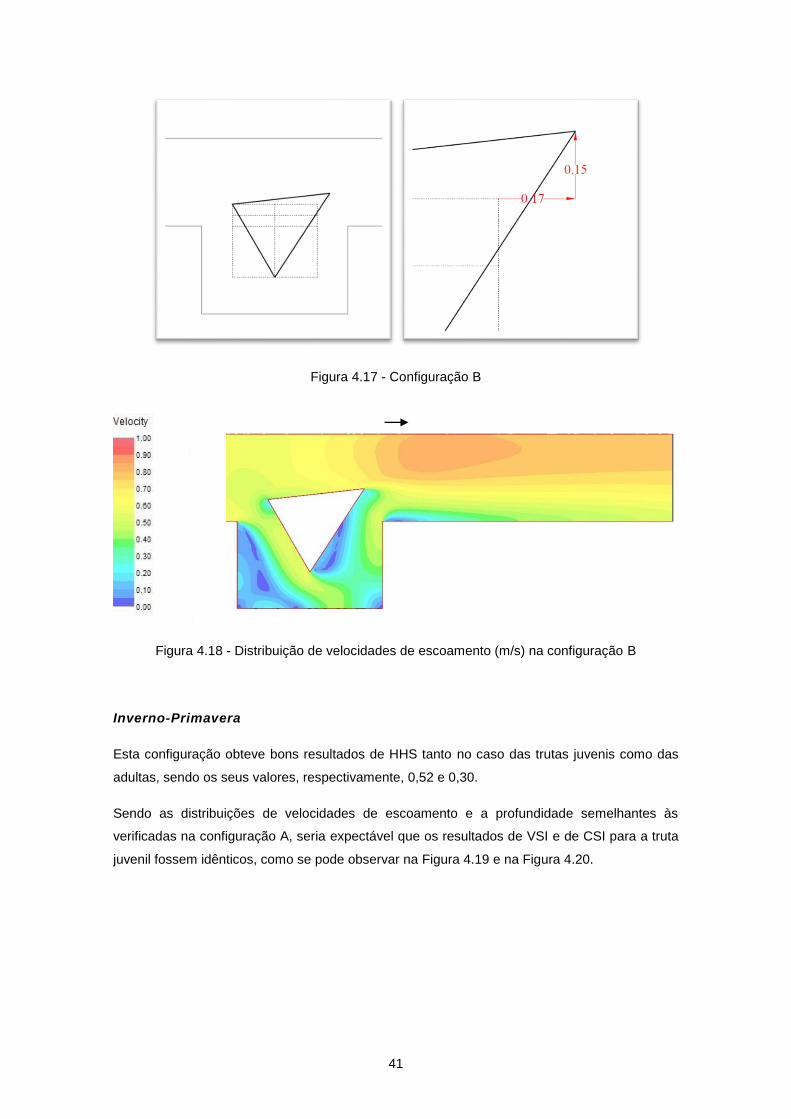
41
Figura 4.17 - Configuração B
Figura 4.18 - Distribuição de velocidades de escoamento (m/s) na configuração B
Inverno-Primavera
Esta configuração obteve bons resultados de HHS tanto no caso das trutas juvenis como das
adultas, sendo os seus valores, respectivamente, 0,52 e 0,30.
Sendo as distribuições de velocidades de escoamento e a profundidade semelhantes às
verificadas na configuração A, seria expectável que os resultados de VSI e de CSI para a truta
juvenil fossem idênticos, como se pode observar na Figura 4.19 e na Figura 4.20.
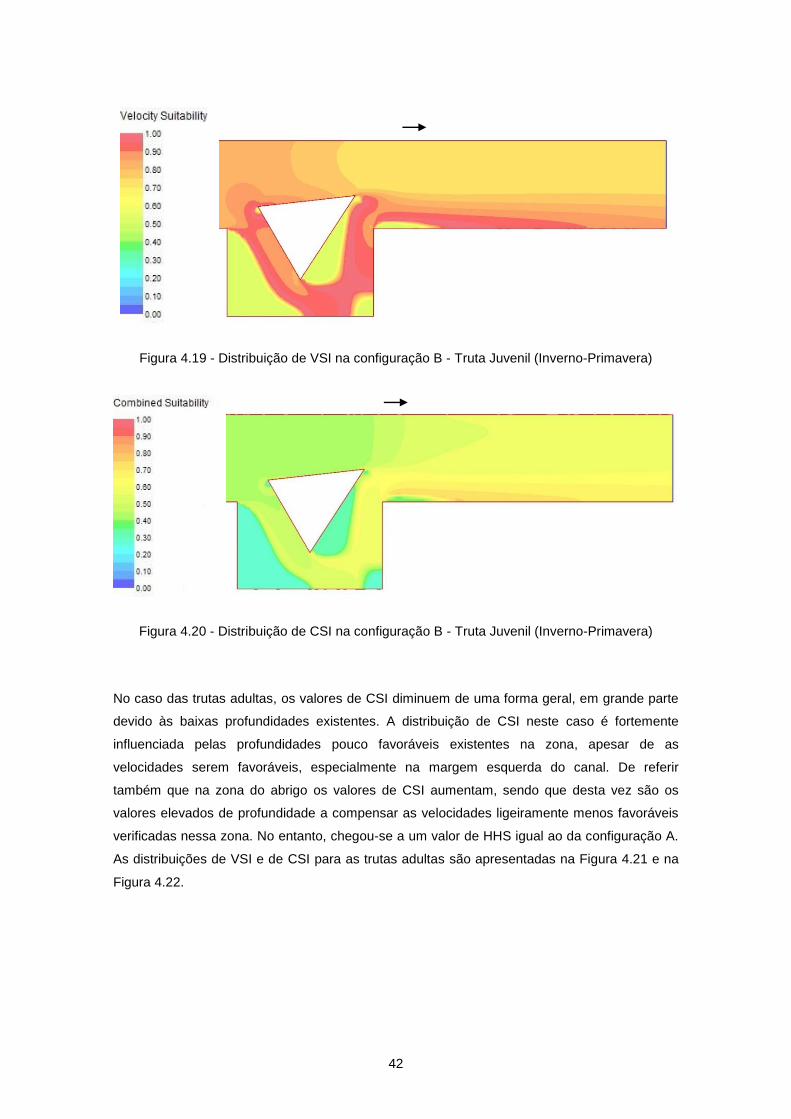
42
Figura 4.19 - Distribuição de VSI na configuração B - Truta Juvenil (Inverno-Primavera)
Figura 4.20 - Distribuição de CSI na configuração B - Truta Juvenil (Inverno-Primavera)
No caso das trutas adultas, os valores de CSI diminuem de uma forma geral, em grande parte
devido às baixas profundidades existentes. A distribuição de CSI neste caso é fortemente
influenciada pelas profundidades pouco favoráveis existentes na zona, apesar de as
velocidades serem favoráveis, especialmente na margem esquerda do canal. De referir
também que na zona do abrigo os valores de CSI aumentam, sendo que desta vez são os
valores elevados de profundidade a compensar as velocidades ligeiramente menos favoráveis
verificadas nessa zona. No entanto, chegou-se a um valor de HHS igual ao da configuração A.
As distribuições de VSI e de CSI para as trutas adultas são apresentadas na Figura 4.21 e na
Figura 4.22.
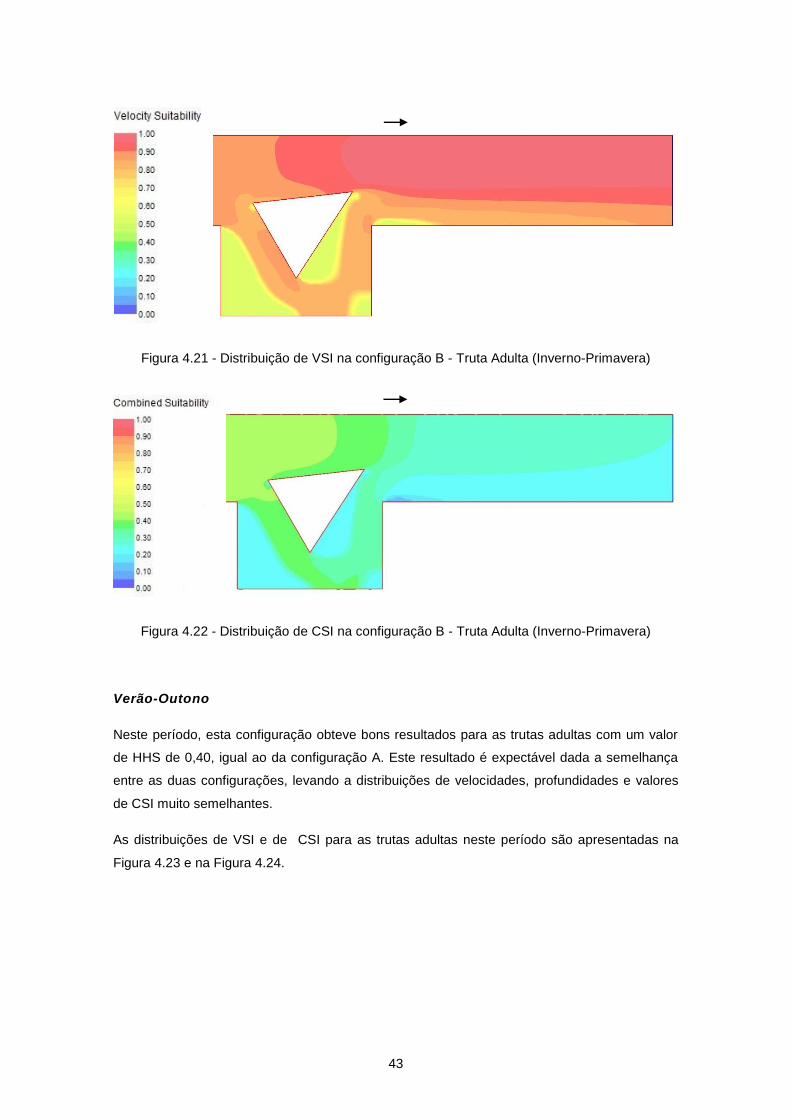
43
Figura 4.21 - Distribuição de VSI na configuração B - Truta Adulta (Inverno-Primavera)
Figura 4.22 - Distribuição de CSI na configuração B - Truta Adulta (Inverno-Primavera)
Verão-Outono
Neste período, esta configuração obteve bons resultados para as trutas adultas com um valor
de HHS de 0,40, igual ao da configuração A. Este resultado é expectável dada a semelhança
entre as duas configurações, levando a distribuições de velocidades, profundidades e valores
de CSI muito semelhantes.
As distribuições de VSI e de CSI para as trutas adultas neste período são apresentadas na
Figura 4.23 e na Figura 4.24.
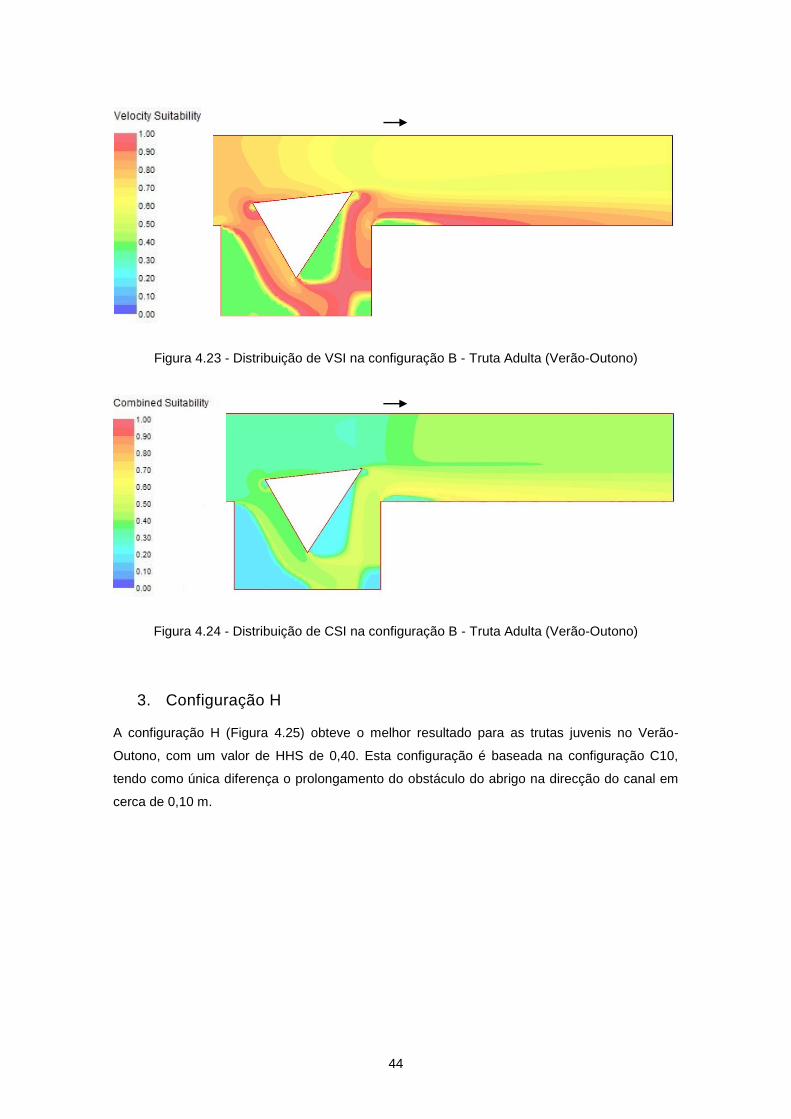
44
Figura 4.23 - Distribuição de VSI na configuração B - Truta Adulta (Verão-Outono)
Figura 4.24 - Distribuição de CSI na configuração B - Truta Adulta (Verão-Outono)
3. Configuração H
A configuração H (Figura 4.25) obteve o melhor resultado para as trutas juvenis no Verão-
Outono, com um valor de HHS de 0,40. Esta configuração é baseada na configuração C10,
tendo como única diferença o prolongamento do obstáculo do abrigo na direcção do canal em
cerca de 0,10 m.
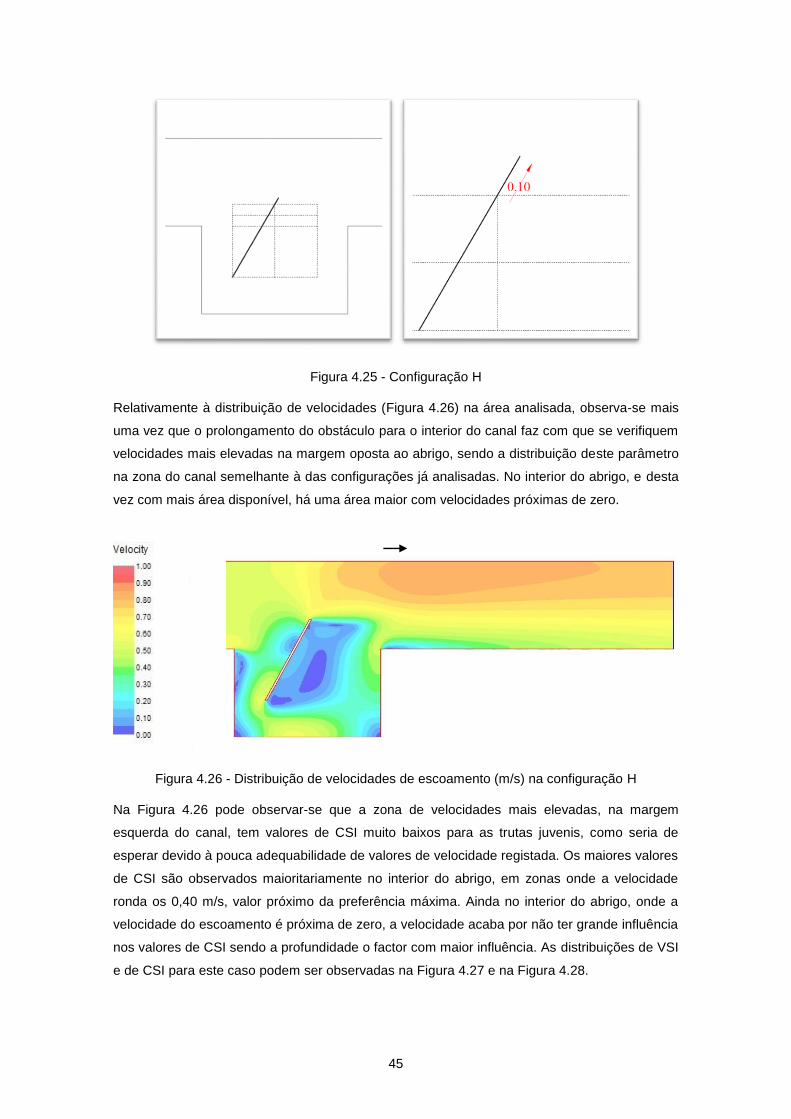
45
Figura 4.25 - Configuração H
Relativamente à distribuição de velocidades (Figura 4.26) na área analisada, observa-se mais
uma vez que o prolongamento do obstáculo para o interior do canal faz com que se verifiquem
velocidades mais elevadas na margem oposta ao abrigo, sendo a distribuição deste parâmetro
na zona do canal semelhante à das configurações já analisadas. No interior do abrigo, e desta
vez com mais área disponível, há uma área maior com velocidades próximas de zero.
Figura 4.26 - Distribuição de velocidades de escoamento (m/s) na configuração H
Na Figura 4.26 pode observar-se que a zona de velocidades mais elevadas, na margem
esquerda do canal, tem valores de CSI muito baixos para as trutas juvenis, como seria de
esperar devido à pouca adequabilidade de valores de velocidade registada. Os maiores valores
de CSI são observados maioritariamente no interior do abrigo, em zonas onde a velocidade
ronda os 0,40 m/s, valor próximo da preferência máxima. Ainda no interior do abrigo, onde a
velocidade do escoamento é próxima de zero, a velocidade acaba por não ter grande influência
nos valores de CSI sendo a profundidade o factor com maior influência. As distribuições de VSI
e de CSI para este caso podem ser observadas na Figura 4.27 e na Figura 4.28.
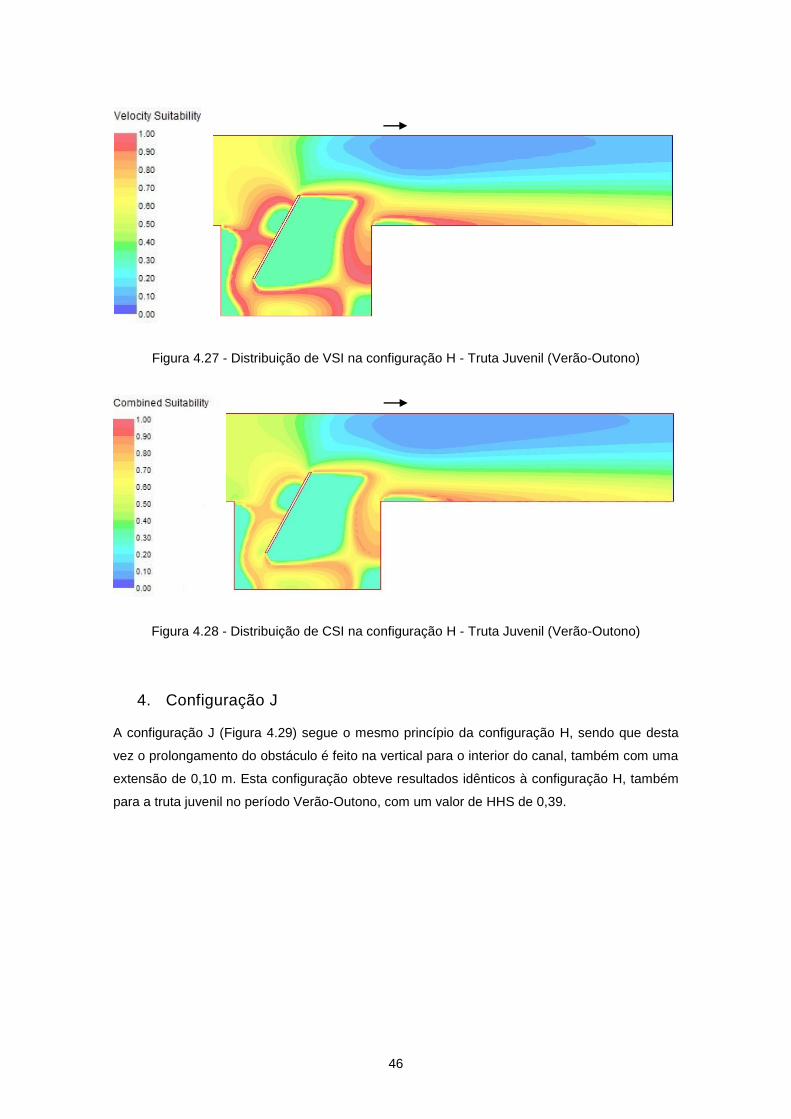
46
Figura 4.27 - Distribuição de VSI na configuração H - Truta Juvenil (Verão-Outono)
Figura 4.28 - Distribuição de CSI na configuração H - Truta Juvenil (Verão-Outono)
4. Configuração J
A configuração J (Figura 4.29) segue o mesmo princípio da configuração H, sendo que desta
vez o prolongamento do obstáculo é feito na vertical para o interior do canal, também com uma
extensão de 0,10 m. Esta configuração obteve resultados idênticos à configuração H, também
para a truta juvenil no período Verão-Outono, com um valor de HHS de 0,39.
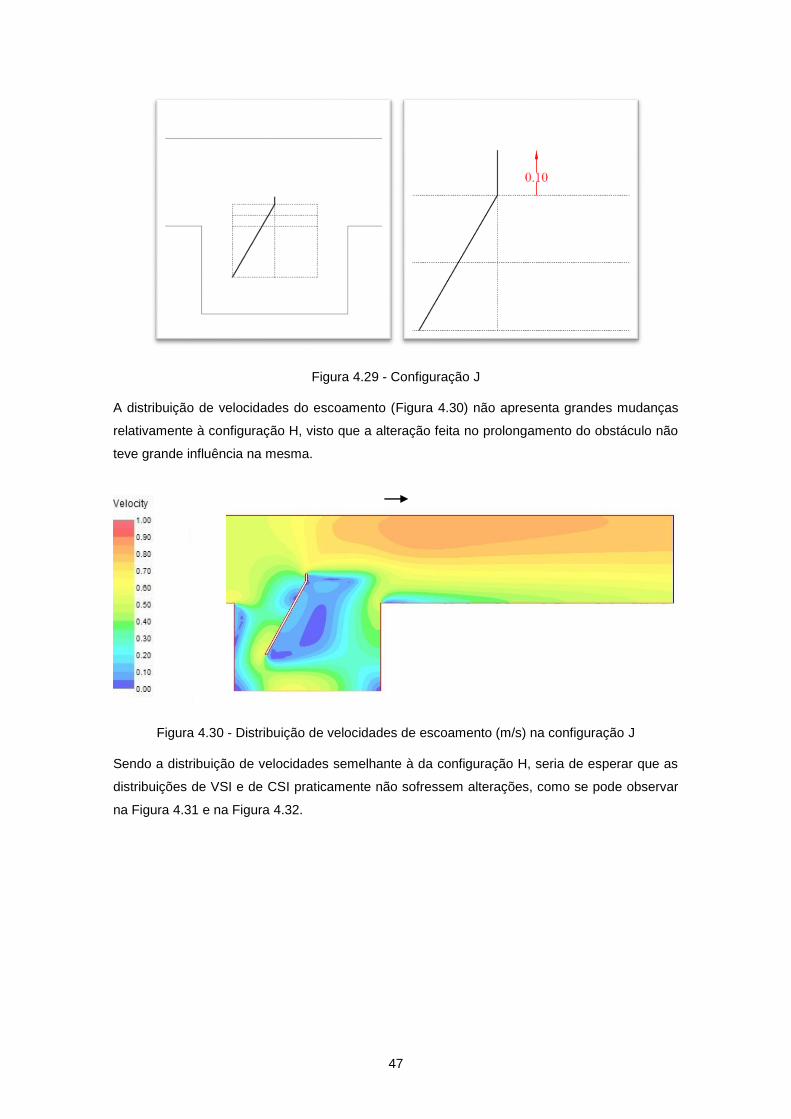
47
Figura 4.29 - Configuração J
A distribuição de velocidades do escoamento (Figura 4.30) não apresenta grandes mudanças
relativamente à configuração H, visto que a alteração feita no prolongamento do obstáculo não
teve grande influência na mesma.
Figura 4.30 - Distribuição de velocidades de escoamento (m/s) na configuração J
Sendo a distribuição de velocidades semelhante à da configuração H, seria de esperar que as
distribuições de VSI e de CSI praticamente não sofressem alterações, como se pode observar
na Figura 4.31 e na Figura 4.32.
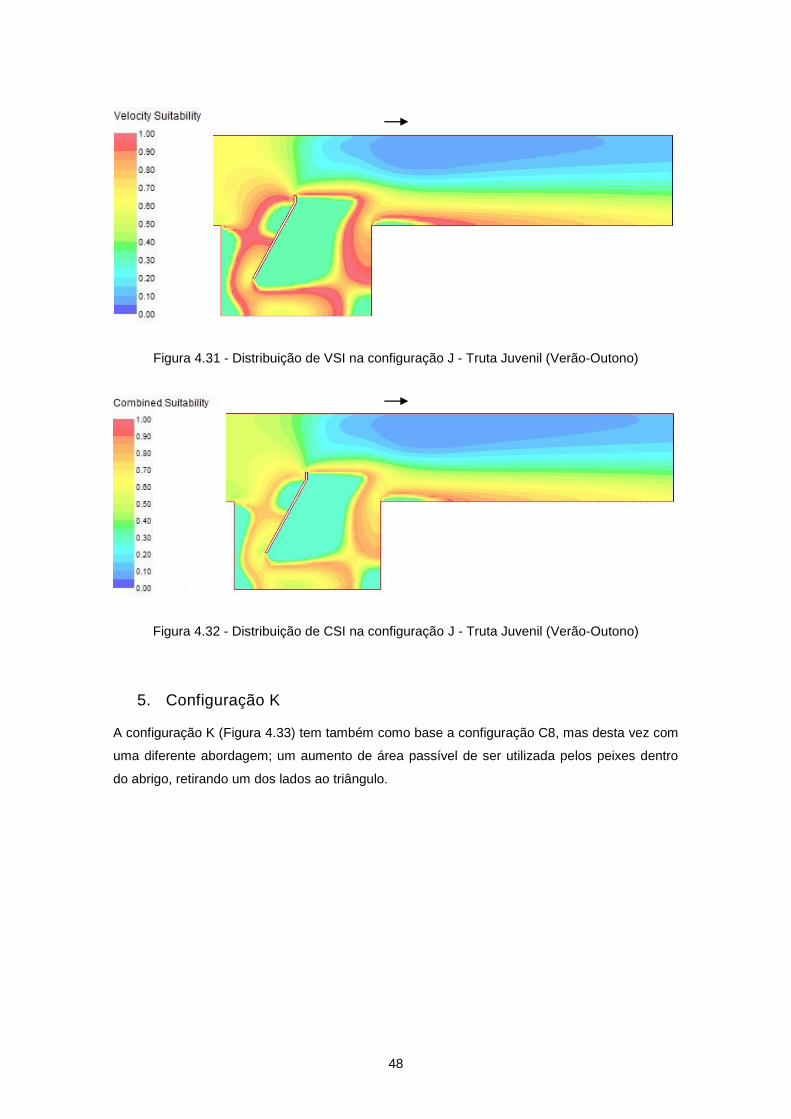
48
Figura 4.31 - Distribuição de VSI na configuração J - Truta Juvenil (Verão-Outono)
Figura 4.32 - Distribuição de CSI na configuração J - Truta Juvenil (Verão-Outono)
5. Configuração K
A configuração K (Figura 4.33) tem também como base a configuração C8, mas desta vez com
uma diferente abordagem; um aumento de área passível de ser utilizada pelos peixes dentro
do abrigo, retirando um dos lados ao triângulo.
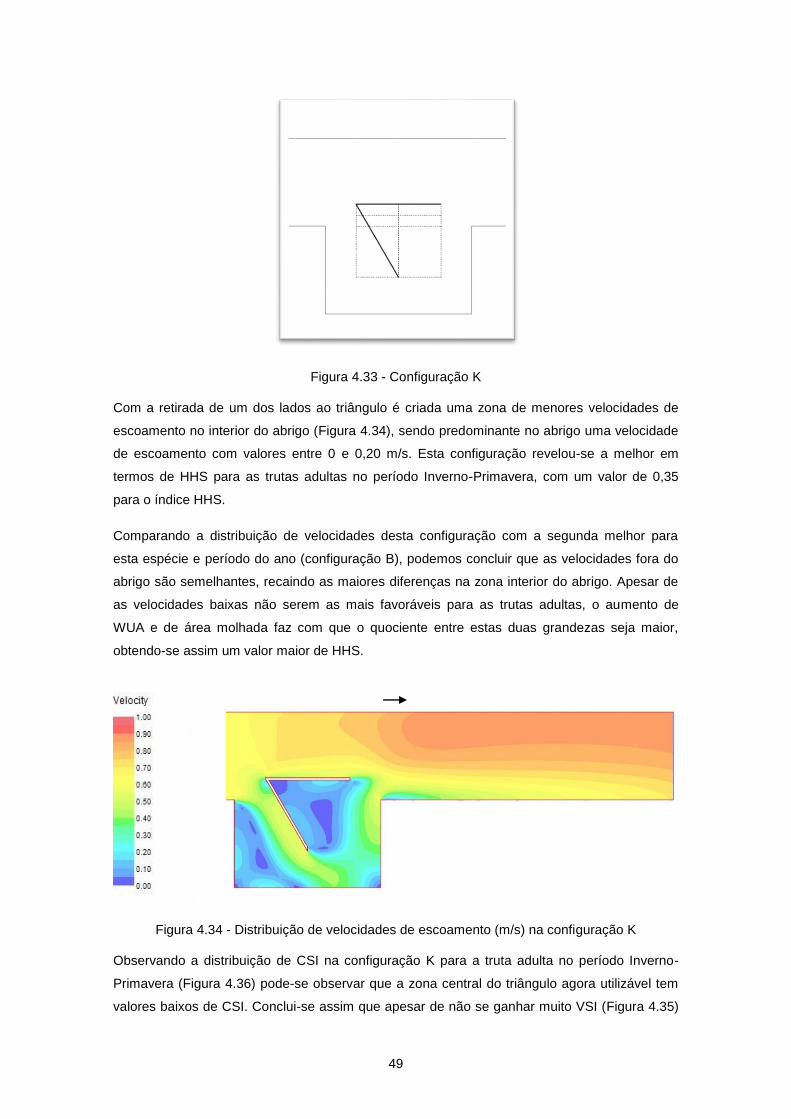
49
Figura 4.33 - Configuração K
Com a retirada de um dos lados ao triângulo é criada uma zona de menores velocidades de
escoamento no interior do abrigo (Figura 4.34), sendo predominante no abrigo uma velocidade
de escoamento com valores entre 0 e 0,20 m/s. Esta configuração revelou-se a melhor em
termos de HHS para as trutas adultas no período Inverno-Primavera, com um valor de 0,35
para o índice HHS.
Comparando a distribuição de velocidades desta configuração com a segunda melhor para
esta espécie e período do ano (configuração B), podemos concluir que as velocidades fora do
abrigo são semelhantes, recaindo as maiores diferenças na zona interior do abrigo. Apesar de
as velocidades baixas não serem as mais favoráveis para as trutas adultas, o aumento de
WUA e de área molhada faz com que o quociente entre estas duas grandezas seja maior,
obtendo-se assim um valor maior de HHS.
Figura 4.34 - Distribuição de velocidades de escoamento (m/s) na configuração K
Observando a distribuição de CSI na configuração K para a truta adulta no período Inverno-
Primavera (Figura 4.36) pode-se observar que a zona central do triângulo agora utilizável tem
valores baixos de CSI. Conclui-se assim que apesar de não se ganhar muito VSI (Figura 4.35)
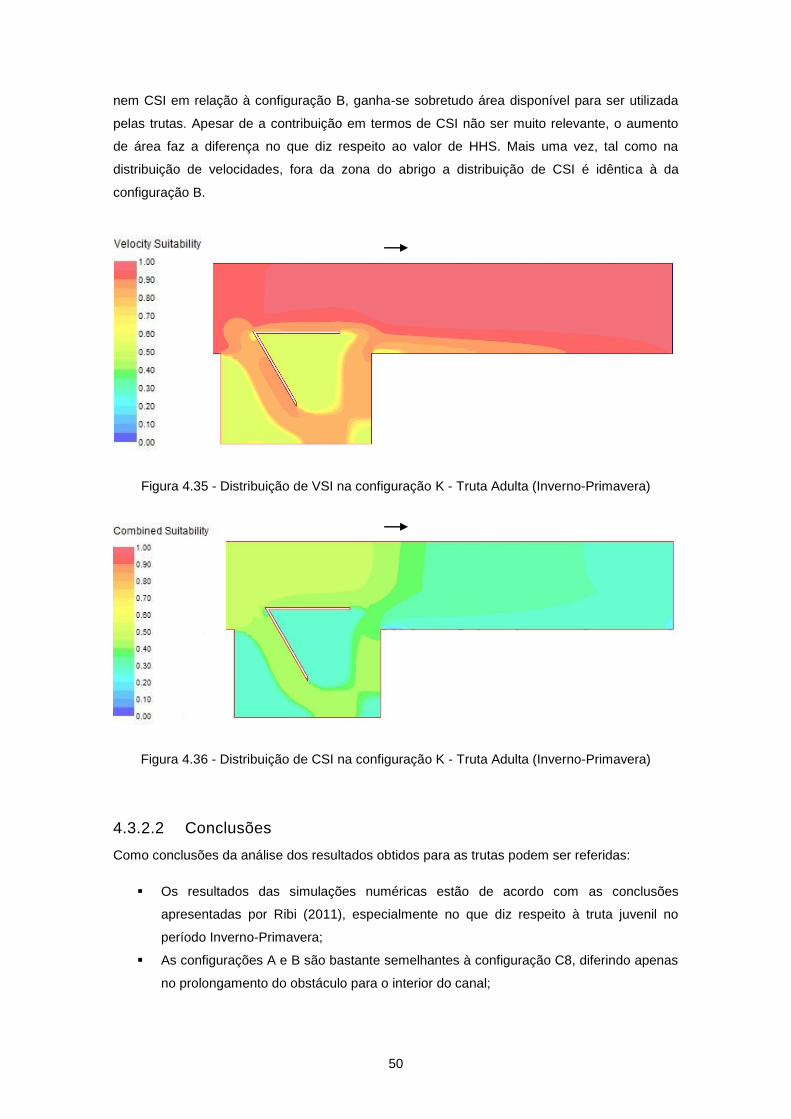
50
nem CSI em relação à configuração B, ganha-se sobretudo área disponível para ser utilizada
pelas trutas. Apesar de a contribuição em termos de CSI não ser muito relevante, o aumento
de área faz a diferença no que diz respeito ao valor de HHS. Mais uma vez, tal como na
distribuição de velocidades, fora da zona do abrigo a distribuição de CSI é idêntica à da
configuração B.
Figura 4.35 - Distribuição de VSI na configuração K - Truta Adulta (Inverno-Primavera)
Figura 4.36 - Distribuição de CSI na configuração K - Truta Adulta (Inverno-Primavera)
4.3.2.2 Conclusões
Como conclusões da análise dos resultados obtidos para as trutas podem ser referidas:
Os resultados das simulações numéricas estão de acordo com as conclusões
apresentadas por Ribi (2011), especialmente no que diz respeito à truta juvenil no
período Inverno-Primavera;
As configurações A e B são bastante semelhantes à configuração C8, diferindo apenas
no prolongamento do obstáculo para o interior do canal;

51
Mesmo nos estágios onde estas duas configurações não foram as melhores (truta
adulta no Inverno-Primavera no caso da configuração A, e truta juvenil no Verão-
Outono em ambas) os valores de HHS estão muito próximos dos valores mais
elevados para qualquer alternativa;
O prolongamento do obstáculo para o interior do canal tem um impacto positivo nos
valores de HHS.
4.3.3 ANÁLISE DOS RESULTADOS OBTIDOS PARA OS BARBOS
4.3.3.1 Considerações Gerais
Os valores mais elevados de HHS para os barbos foram obtidos nas configurações C, D e H
para os juvenis e J e K para os adultos. Verificou-se uma acentuada diferença nos valores de
HHS entre os estágios juvenil e adulto, devido essencialmente à escassez de profundidades
favoráveis aos barbos adultos.
1. Configuração C
A configuração C é semelhante à configuração C8 no que diz respeito à forma e dimensões do
obstáculo, sendo as alterações efectuadas nas dimensões do abrigo. O abrigo tem dimensões
2x2 m, assumindo assim uma forma quadrada (Figura 4.37). Nesta configuração obteve-se,
para os barbos juvenis, um valor de HHS de 0,50.
Figura 4.37 - Configuração C
Observando a distribuição de velocidades do escoamento (Figura 4.38) é fácil perceber o
porquê dos bons resultados desta configuração para o barbo juvenil. A criação de uma zona de
velocidades baixas no interior do abrigo associada às profundidades registadas no mesmo
local é bastante favorável aos barbos juvenis (Figura 4.39).
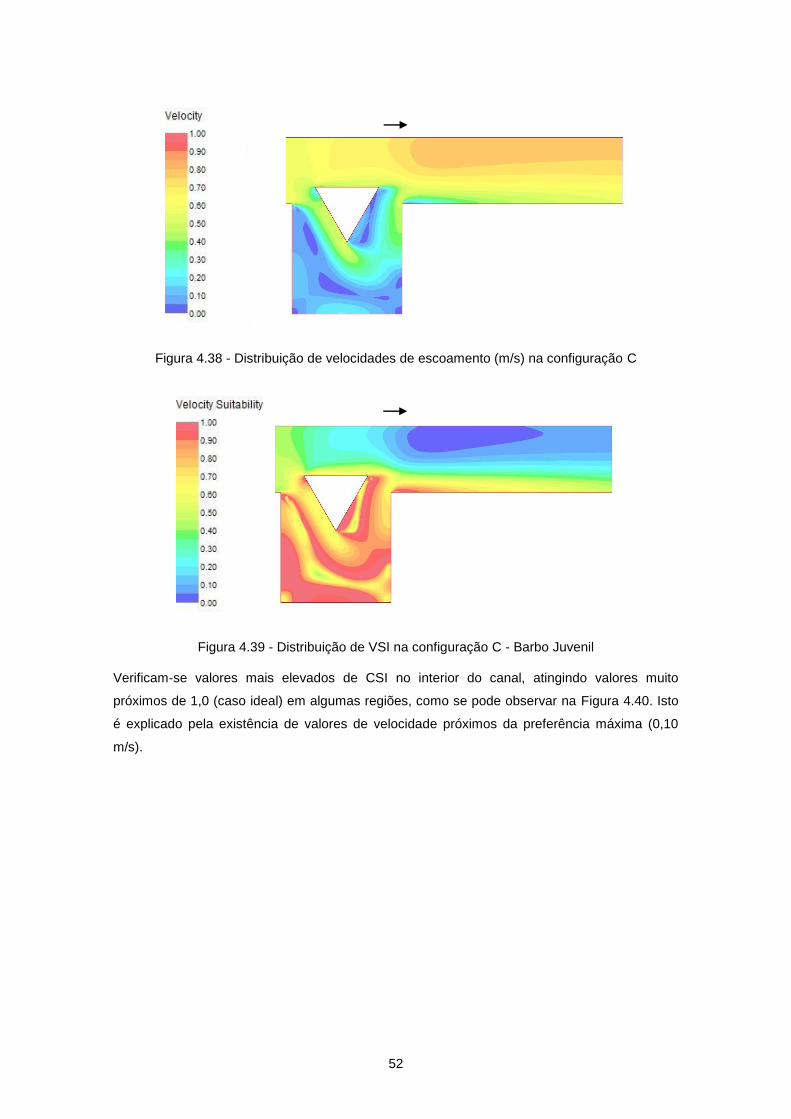
52
Figura 4.38 - Distribuição de velocidades de escoamento (m/s) na configuração C
Figura 4.39 - Distribuição de VSI na configuração C - Barbo Juvenil
Verificam-se valores mais elevados de CSI no interior do canal, atingindo valores muito
próximos de 1,0 (caso ideal) em algumas regiões, como se pode observar na Figura 4.40. Isto
é explicado pela existência de valores de velocidade próximos da preferência máxima (0,10
m/s).
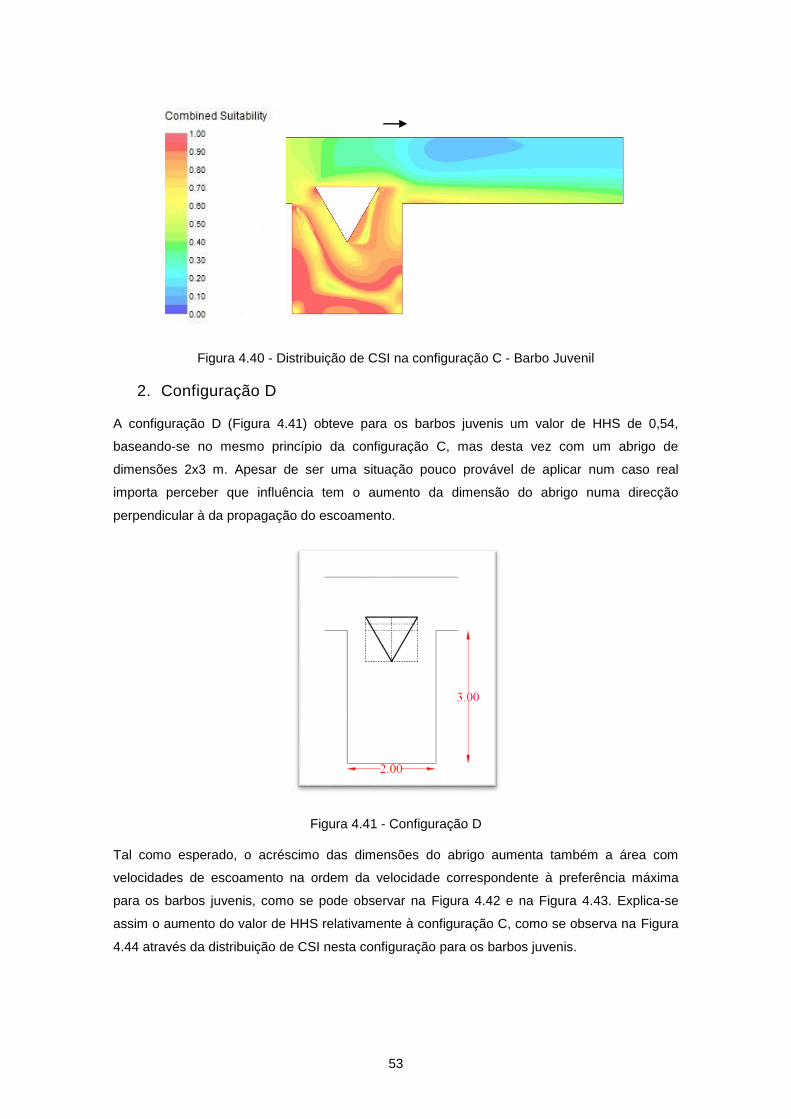
53
Figura 4.40 - Distribuição de CSI na configuração C - Barbo Juvenil
2. Configuração D
A configuração D (Figura 4.41) obteve para os barbos juvenis um valor de HHS de 0,54,
baseando-se no mesmo princípio da configuração C, mas desta vez com um abrigo de
dimensões 2x3 m. Apesar de ser uma situação pouco provável de aplicar num caso real
importa perceber que influência tem o aumento da dimensão do abrigo numa direcção
perpendicular à da propagação do escoamento.
Figura 4.41 - Configuração D
Tal como esperado, o acréscimo das dimensões do abrigo aumenta também a área com
velocidades de escoamento na ordem da velocidade correspondente à preferência máxima
para os barbos juvenis, como se pode observar na Figura 4.42 e na Figura 4.43. Explica-se
assim o aumento do valor de HHS relativamente à configuração C, como se observa na Figura
4.44 através da distribuição de CSI nesta configuração para os barbos juvenis.
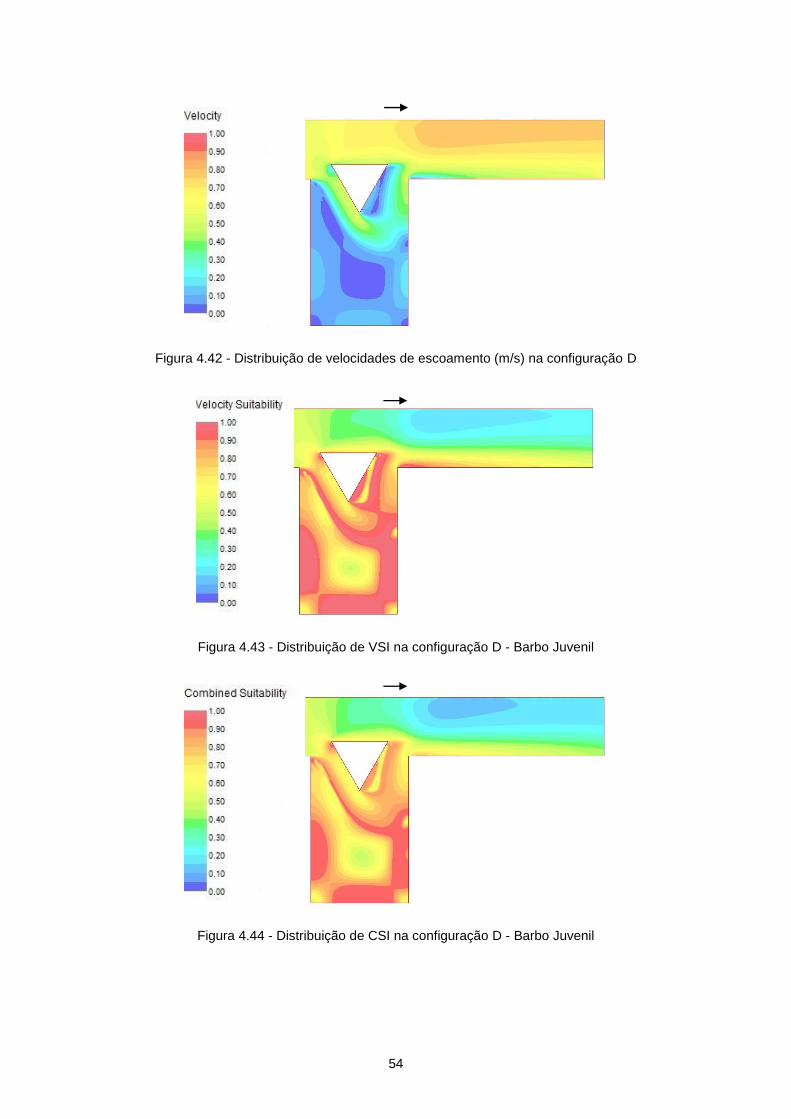
54
Figura 4.42 - Distribuição de velocidades de escoamento (m/s) na configuração D
Figura 4.43 - Distribuição de VSI na configuração D - Barbo Juvenil
Figura 4.44 - Distribuição de CSI na configuração D - Barbo Juvenil
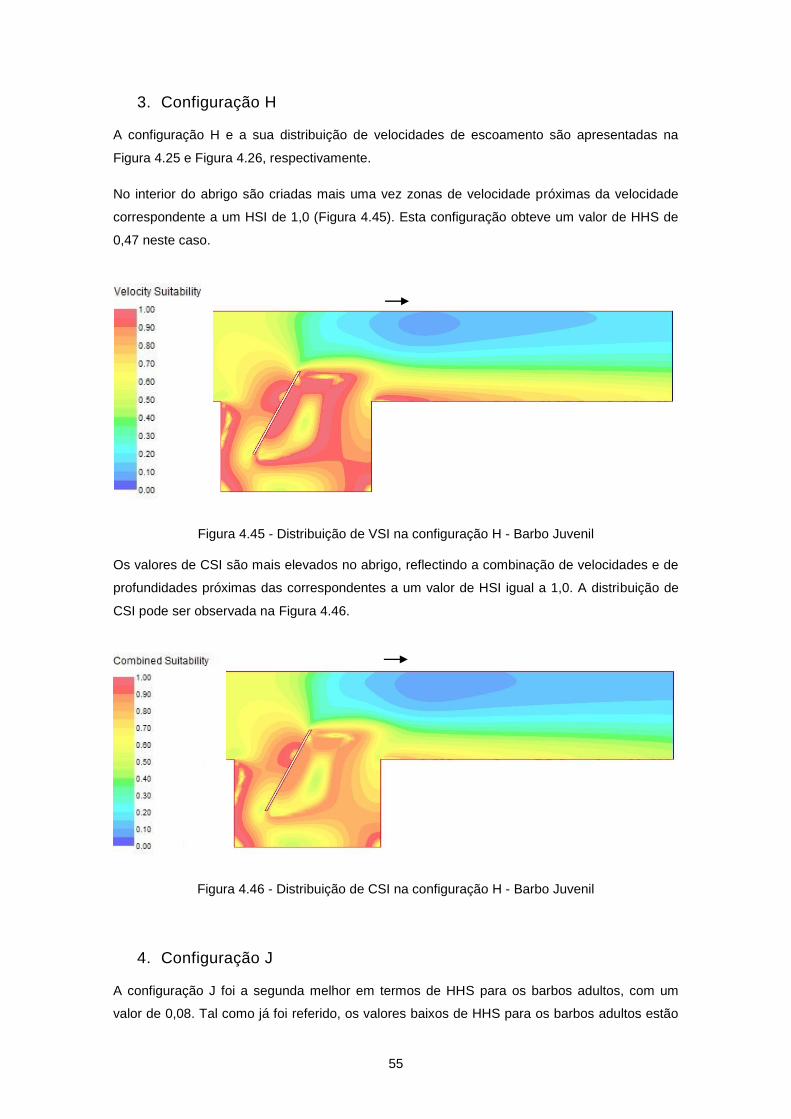
55
3. Configuração H
A configuração H e a sua distribuição de velocidades de escoamento são apresentadas na
Figura 4.25 e Figura 4.26, respectivamente.
No interior do abrigo são criadas mais uma vez zonas de velocidade próximas da velocidade
correspondente a um HSI de 1,0 (Figura 4.45). Esta configuração obteve um valor de HHS de
0,47 neste caso.
Figura 4.45 - Distribuição de VSI na configuração H - Barbo Juvenil
Os valores de CSI são mais elevados no abrigo, reflectindo a combinação de velocidades e de
profundidades próximas das correspondentes a um valor de HSI igual a 1,0. A distribuição de
CSI pode ser observada na Figura 4.46.
Figura 4.46 - Distribuição de CSI na configuração H - Barbo Juvenil
4. Configuração J
A configuração J foi a segunda melhor em termos de HHS para os barbos adultos, com um
valor de 0,08. Tal como já foi referido, os valores baixos de HHS para os barbos adultos estão
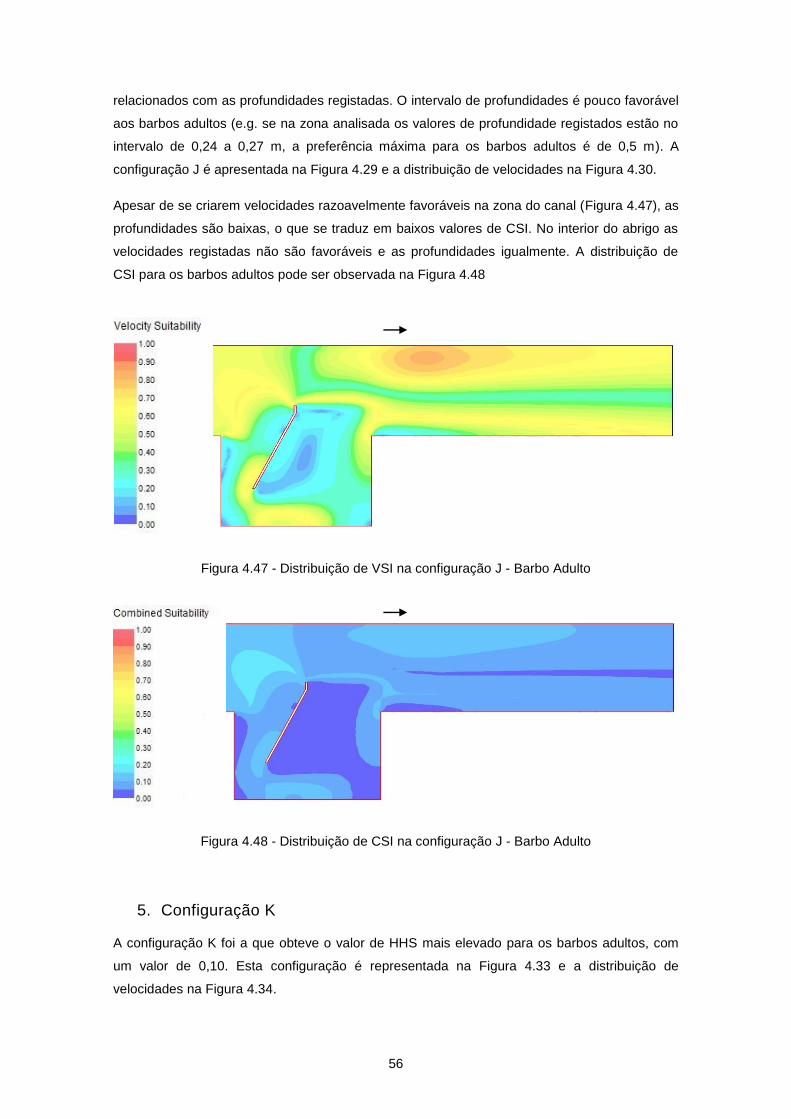
56
relacionados com as profundidades registadas. O intervalo de profundidades é pouco favorável
aos barbos adultos (e.g. se na zona analisada os valores de profundidade registados estão no
intervalo de 0,24 a 0,27 m, a preferência máxima para os barbos adultos é de 0,5 m). A
configuração J é apresentada na Figura 4.29 e a distribuição de velocidades na Figura 4.30.
Apesar de se criarem velocidades razoavelmente favoráveis na zona do canal (Figura 4.47), as
profundidades são baixas, o que se traduz em baixos valores de CSI. No interior do abrigo as
velocidades registadas não são favoráveis e as profundidades igualmente. A distribuição de
CSI para os barbos adultos pode ser observada na Figura 4.48
Figura 4.47 - Distribuição de VSI na configuração J - Barbo Adulto
Figura 4.48 - Distribuição de CSI na configuração J - Barbo Adulto
5. Configuração K
A configuração K foi a que obteve o valor de HHS mais elevado para os barbos adultos, com
um valor de 0,10. Esta configuração é representada na Figura 4.33 e a distribuição de
velocidades na Figura 4.34.
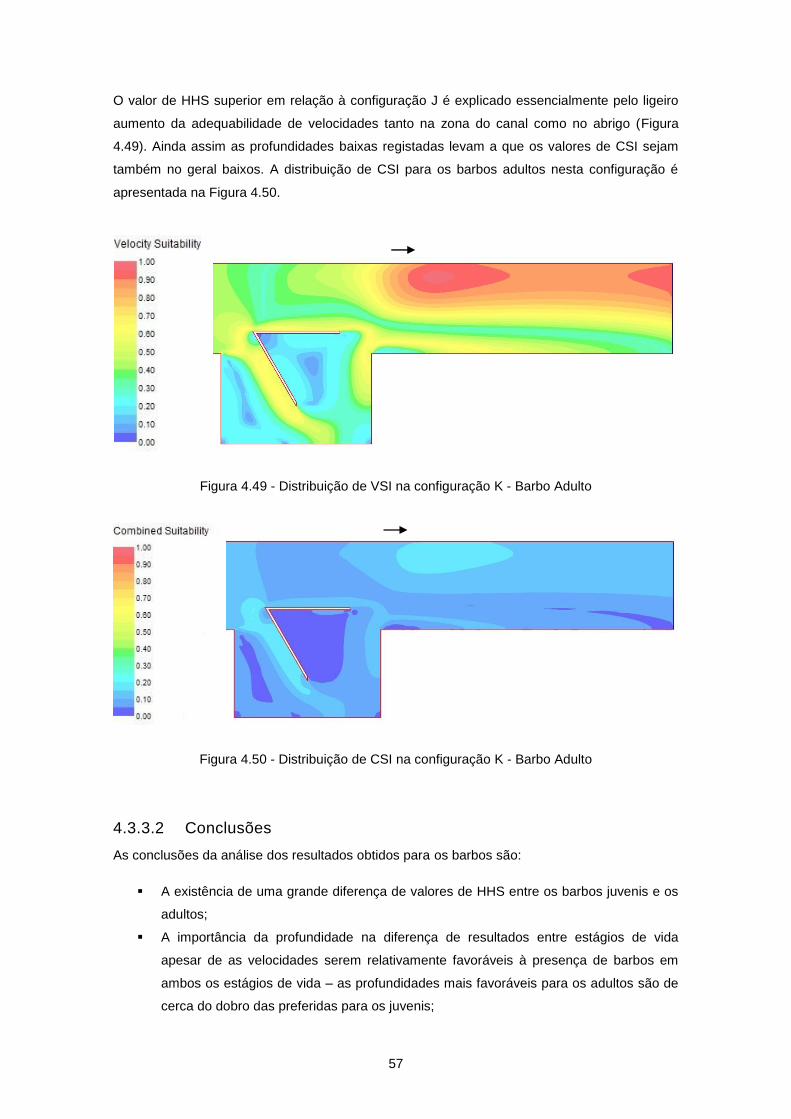
57
O valor de HHS superior em relação à configuração J é explicado essencialmente pelo ligeiro
aumento da adequabilidade de velocidades tanto na zona do canal como no abrigo (Figura
4.49). Ainda assim as profundidades baixas registadas levam a que os valores de CSI sejam
também no geral baixos. A distribuição de CSI para os barbos adultos nesta configuração é
apresentada na Figura 4.50.
Figura 4.49 - Distribuição de VSI na configuração K - Barbo Adulto
Figura 4.50 - Distribuição de CSI na configuração K - Barbo Adulto
4.3.3.2 Conclusões
As conclusões da análise dos resultados obtidos para os barbos são:
A existência de uma grande diferença de valores de HHS entre os barbos juvenis e os
adultos;
A importância da profundidade na diferença de resultados entre estágios de vida
apesar de as velocidades serem relativamente favoráveis à presença de barbos em
ambos os estágios de vida – as profundidades mais favoráveis para os adultos são de
cerca do dobro das preferidas para os juvenis;

58
As profundidades, a variarem entre 0,24 e 0,27 m na área em estudo, dificultam a
obtenção de valores elevados de CSI e consequentemente de valores elevados HHS
para os barbos adultos.

59
5 APLICAÇÃO DOS ABRIGOS A UM CURSO DE ÁGUA
NATURAL – RIO OCREZA
5.1 INTRODUÇÃO
Apesar de as configurações testadas permitirem um aumento do HHS do canal analisado, o
verdadeiro propósito dos abrigos para peixes é a sua aplicação num curso de água natural. A
influência da topografia do leito e das margens difere da dos casos estudados previamente,
razão pela qual se aplicaram os abrigos a um rio. O caso de estudo escolhido foi o rio Ocreza.
O rio Ocreza é um dos maiores afluentes da margem direita do rio Tejo, com cerca de 80 km de
extensão (Figura 5.1). Da sua população piscícola nativa realçasse o barbo comum
(Luciobarbus bocagei), espécie escolhida para testar a eficiência dos abrigos.
Figura 5.1 - Localização do rio Ocreza (Boavida, 2012)
O troço analisado do rio Ocreza tem cerca de 400 m de comprimento e localiza-se a montante
do regolfo da barragem de Pracana e a jusante da povoação de Foz do Cobrão.
5.2 DISPONIBILIDADE DE HABITAT NO RIO OCREZA
Com o objectivo de avaliar a eficiência dos abrigos para os barbos foi necessário seleccionar
um caudal para o qual os abrigos entrariam em funcionamento; um caudal para o qual a cota
da superfície do livre do escoamento fosse tal que permitisse aos peixes a entrada no abrigo.
Desta forma, enquanto a superfície livre do escoamento não atingisse um determinado valor, o
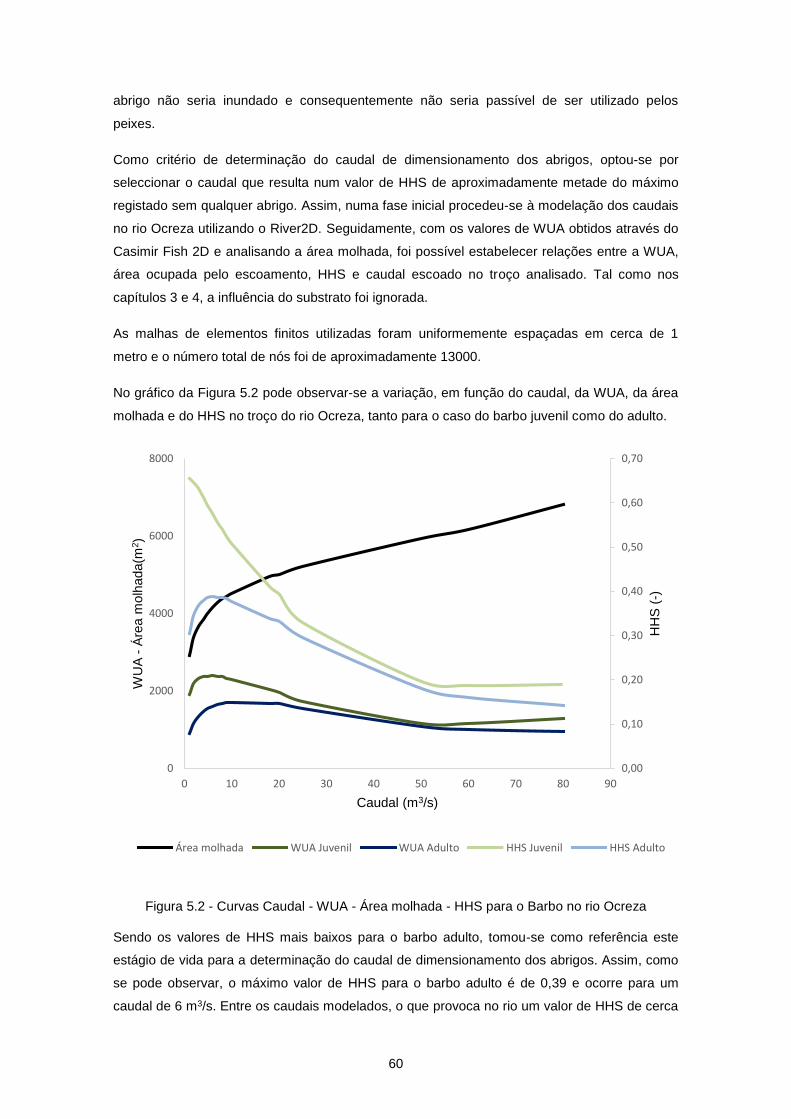
60
abrigo não seria inundado e consequentemente não seria passível de ser utilizado pelos
peixes.
Como critério de determinação do caudal de dimensionamento dos abrigos, optou-se por
seleccionar o caudal que resulta num valor de HHS de aproximadamente metade do máximo
registado sem qualquer abrigo. Assim, numa fase inicial procedeu-se à modelação dos caudais
no rio Ocreza utilizando o River2D. Seguidamente, com os valores de WUA obtidos através do
Casimir Fish 2D e analisando a área molhada, foi possível estabelecer relações entre a WUA,
área ocupada pelo escoamento, HHS e caudal escoado no troço analisado. Tal como nos
capítulos 3 e 4, a influência do substrato foi ignorada.
As malhas de elementos finitos utilizadas foram uniformemente espaçadas em cerca de 1
metro e o número total de nós foi de aproximadamente 13000.
No gráfico da Figura 5.2 pode observar-se a variação, em função do caudal, da WUA, da área
molhada e do HHS no troço do rio Ocreza, tanto para o caso do barbo juvenil como do adulto.
Figura 5.2 - Curvas Caudal - WUA - Área molhada - HHS para o Barbo no rio Ocreza
Sendo os valores de HHS mais baixos para o barbo adulto, tomou-se como referência este
estágio de vida para a determinação do caudal de dimensionamento dos abrigos. Assim, como
se pode observar, o máximo valor de HHS para o barbo adulto é de 0,39 e ocorre para um
caudal de 6 m3/s. Entre os caudais modelados, o que provoca no rio um valor de HHS de cerca
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0
2000
4000
6000
8000
0 10 20 30 40 50 60 70 80 90
HH
S (
-)
WU
A -
Áre
a m
olh
ada(m
2)
Caudal (m3/s)
Área molhada WUA Juvenil WUA Adulto HHS Juvenil HHS Adulto

61
de metade do máximo verificado, é o de 50 m3/s com um valor de HHS de 0,18. Desta forma
escolheu-se este valor de caudal como referência para implantação dos abrigos.
5.3 ABRIGOS UTILIZADOS
Com o objectivo de não restringir os abrigos a um estágio de vida, optou-se por testar uma
nova configuração, resultado da conjugação de duas configurações previamente testadas em
4.2. Assim, juntando as características mais determinantes dos abrigos para os juvenis e para
os adultos, chegou-se à configuração apresentada na Figura 5.3.
Figura 5.3 - Abrigo utilizado no rio Ocreza
Esta configuração surge na conjugação de duas características:
O prolongamento do abrigo na direcção perpendicular à do escoamento, que cria
velocidades reduzidas, mais favoráveis aos barbos juvenis;
A escolha da configuração K para o obstáculo do abrigo, configuração esta que obteve
os valores mais elevados de HHS para o barbo adulto.
Tanto as velocidades como as preferências mais favoráveis são relativamente diferentes para
os juvenis e para os adultos.
Como critério utilizado para as dimensões deste abrigo, e tratando-se de um troço de rio com
um comprimento muito maior que os casos anteriormente estudados, optou-se por aumentar as
dimensões do abrigo face ao testado em 4.2 para uma área de 4 x 6 m2. Foram simulados três
abrigos, em funcionamento para o mesmo caudal de 50 m3/s. A localização dos abrigos (1, 2 e
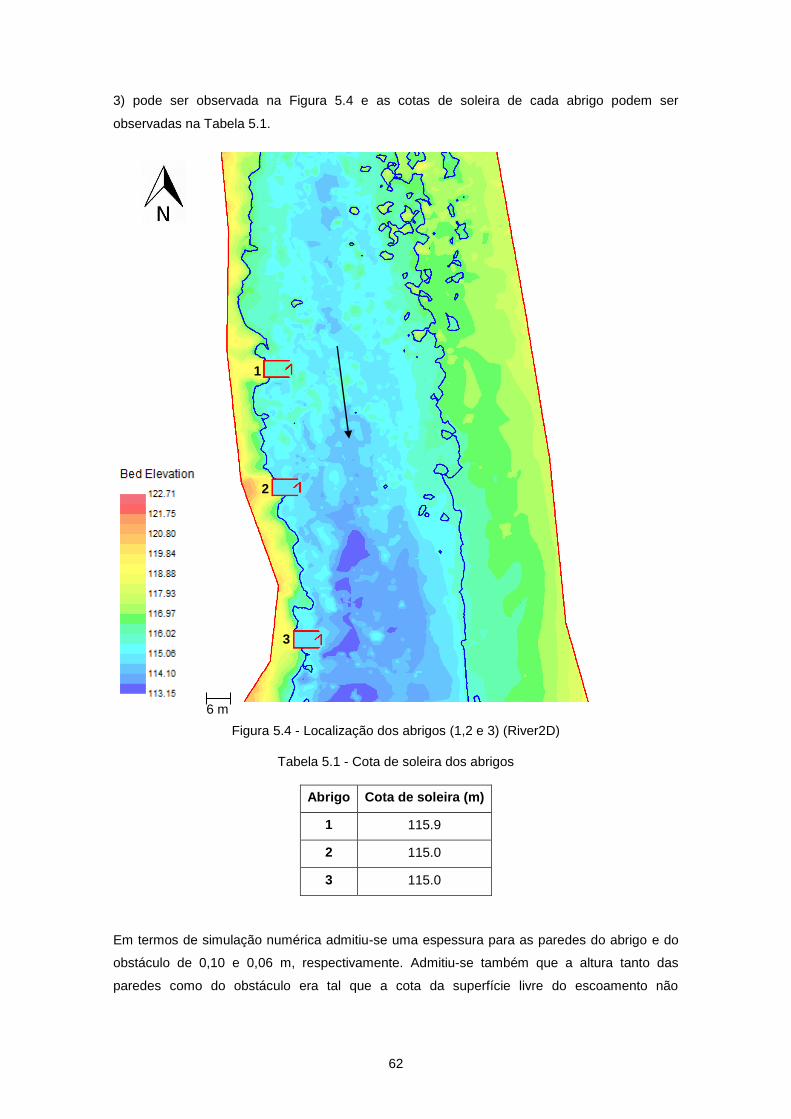
62
3) pode ser observada na Figura 5.4 e as cotas de soleira de cada abrigo podem ser
observadas na Tabela 5.1.
Figura 5.4 - Localização dos abrigos (1,2 e 3) (River2D)
Tabela 5.1 - Cota de soleira dos abrigos
Abrigo Cota de soleira (m)
1 115.9
2 115.0
3 115.0
Em termos de simulação numérica admitiu-se uma espessura para as paredes do abrigo e do
obstáculo de 0,10 e 0,06 m, respectivamente. Admitiu-se também que a altura tanto das
paredes como do obstáculo era tal que a cota da superfície livre do escoamento não
1
2
3
6 m
63
ultrapassou a cota superior dos mesmos. Um esquema 3D do abrigo pode ser observado na
Figura 5.5.
Figura 5.5 - Esquema 3D do abrigo
A implantação dos abrigos foi feita tendo em conta várias considerações:
A necessidade de posicionar o abrigo a uma cota que permitisse a entrada de água no
abrigo, com uma profundidade favorável para os barbos: entre 0,2 e 0,4 m.
A criação de um campo de velocidades favorável de forma a atrair os barbos para o
interior do abrigo.
5.4 RESULTADOS
5.4.1 VALORES DE WUA NO RIO OCREZA
Nos gráficos da Figura 5.6 e da Figura 5.7 pode ser observada a variação da WUA com o
caudal, para os dois casos, com e sem abrigos, a partir dos 50 m3/s e até aos 80 m3/s.
Figura 5.6 - Curva Caudal - WUA com abrigos (Barbo Juvenil)
0
400
800
1200
1600
2000
50 60 80
WU
A (
m2)
Caudal (m3/s)
Sem abrigos Com abrigos

64
Figura 5.7 - Curva Caudal - WUA com abrigos (Barbo Adulto)
Como se pode observar nos gráficos anteriores, a existência de abrigos laterais aumenta os
valores de WUA tanto para o barbo juvenil como para o adulto em cerca de 200 e 600 m2,
respectivamente. Os abrigos, além de criarem uma zona de habitat favorável para as espécies
em análise no interior dos mesmos, criam também condições favoráveis, especialmente de
velocidade, na proximidade dos mesmos.
Apesar de ter sido utilizado como referência o valor de caudal de 50 m3/s, o aumento do valor
de WUA só se começa a fazer sentir para um caudal de 60 m3/s.
De forma a perceber como evolui espacialmente a disponibilidade de habitat é necessário fazer
uma análise em termos de profundidade, velocidade do escoamento e CSI, tanto do estágio
juvenil como adulto, no troço analisado.
Na Figura 5.8 podem ser observadas as profundidades verificadas para os caudais de 50, 60 e
80 m3/s. Como seria de esperar, com o aumento do caudal a profundidade aumenta no troço
analisado.
0
400
800
1200
1600
2000
50 60 80
WU
A (
m2)
Caudal (m3/s)
Sem abrigos Com abrigos
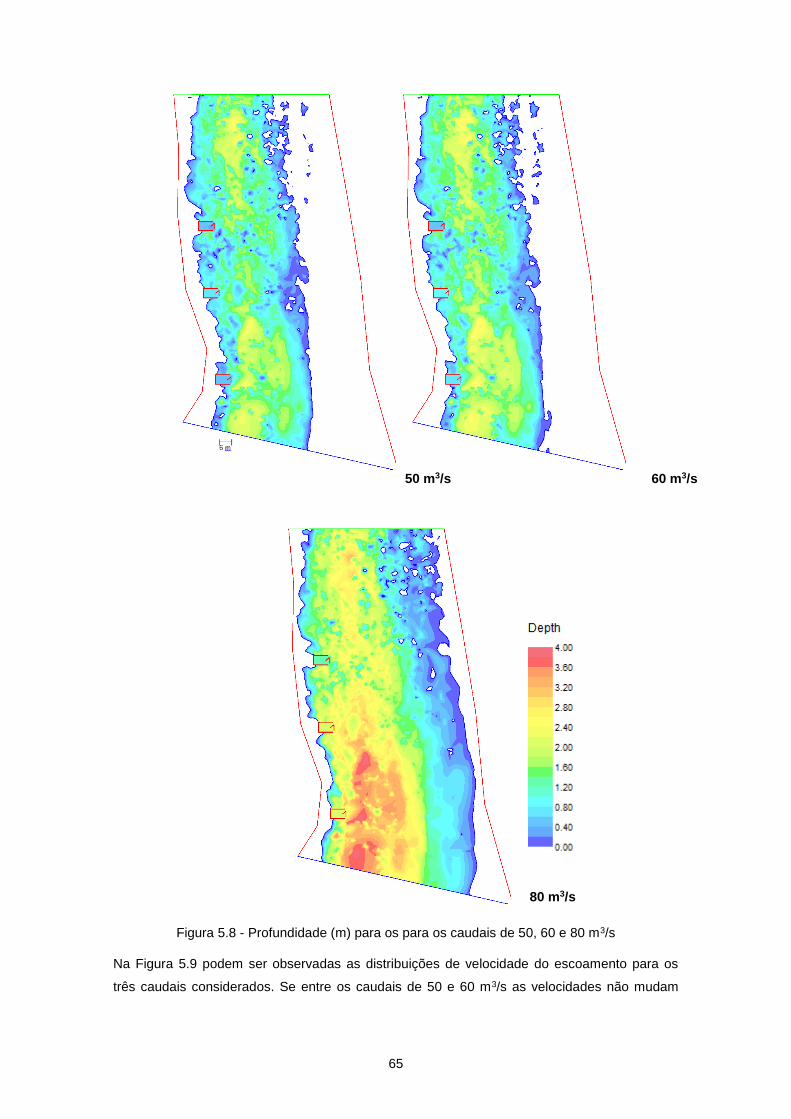
65
Figura 5.8 - Profundidade (m) para os para os caudais de 50, 60 e 80 m3/s
Na Figura 5.9 podem ser observadas as distribuições de velocidade do escoamento para os
três caudais considerados. Se entre os caudais de 50 e 60 m3/s as velocidades não mudam
80 m3/s
50 m3/s 60 m3/s
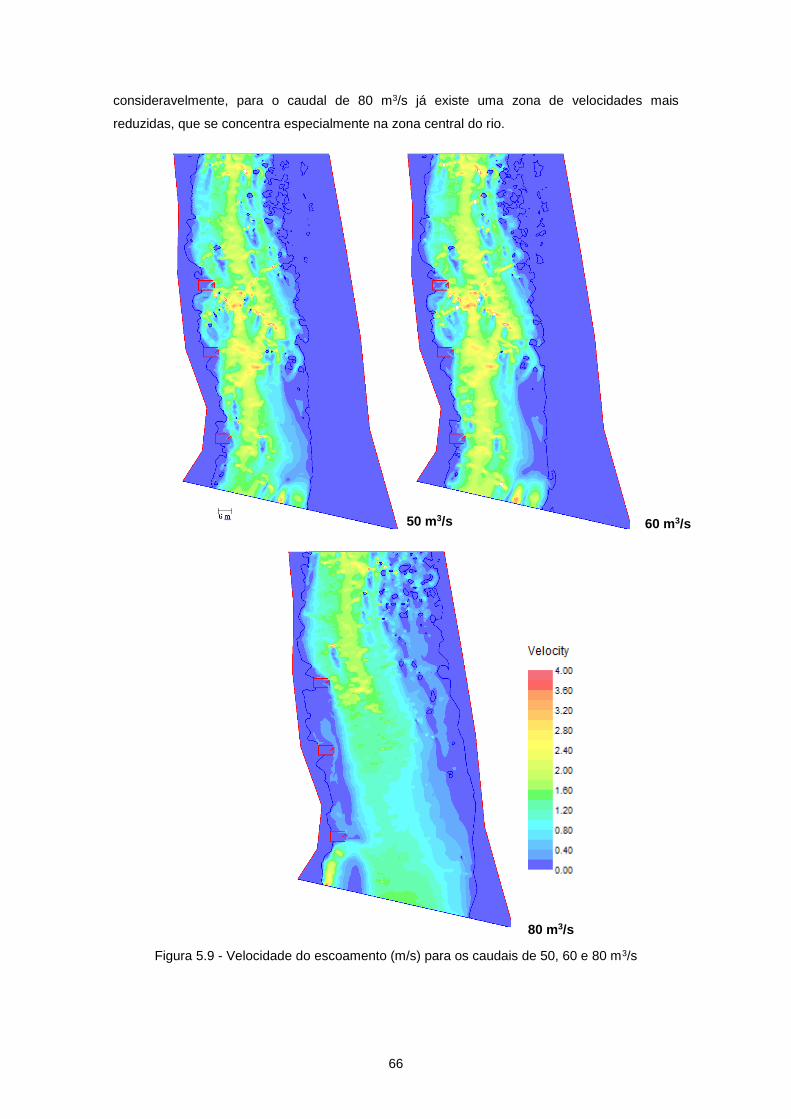
66
consideravelmente, para o caudal de 80 m3/s já existe uma zona de velocidades mais
reduzidas, que se concentra especialmente na zona central do rio.
Figura 5.9 - Velocidade do escoamento (m/s) para os caudais de 50, 60 e 80 m3/s
50 m3/s 60 m3/s
80 m3/s
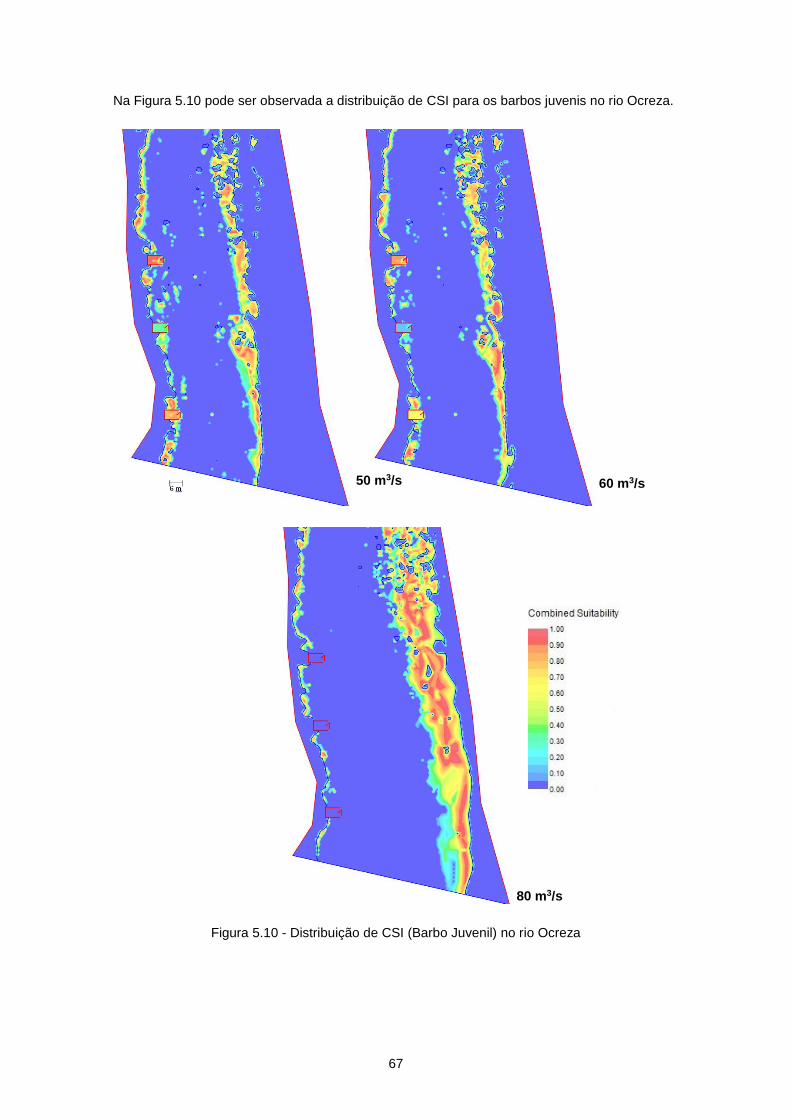
67
Na Figura 5.10 pode ser observada a distribuição de CSI para os barbos juvenis no rio Ocreza.
Figura 5.10 - Distribuição de CSI (Barbo Juvenil) no rio Ocreza
60 m3/s 50 m3/s
80 m3/s
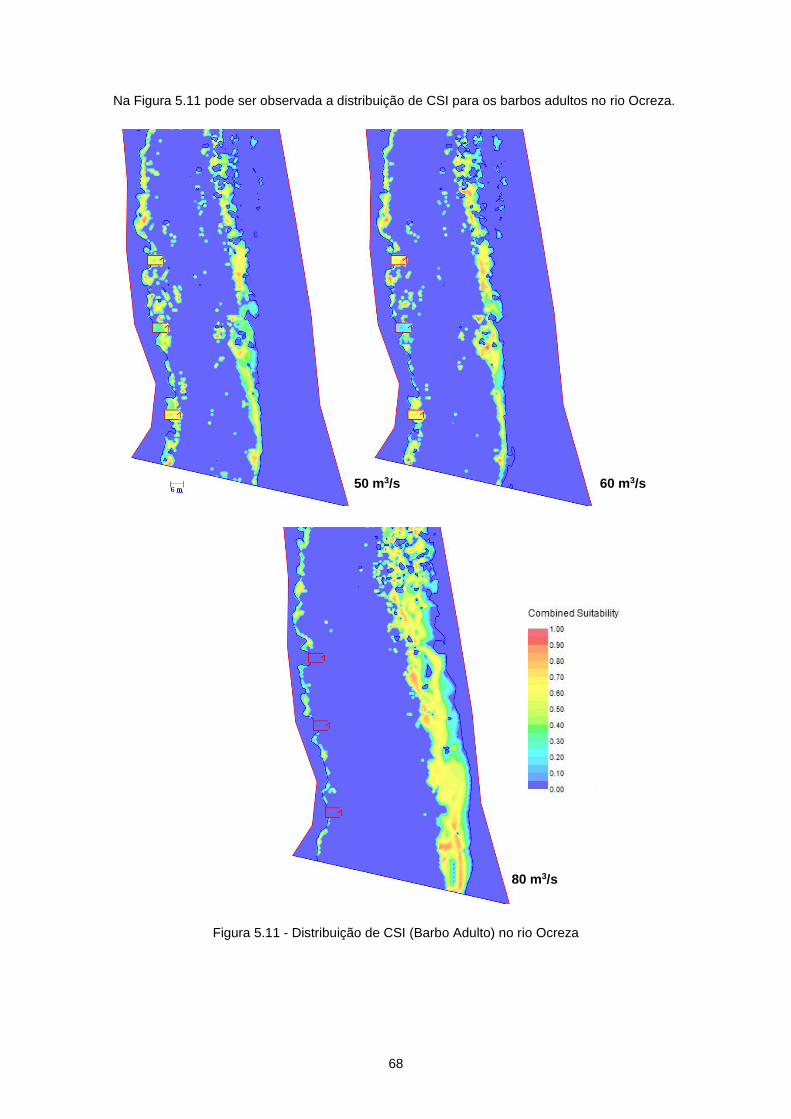
68
Na Figura 5.11 pode ser observada a distribuição de CSI para os barbos adultos no rio Ocreza.
Figura 5.11 - Distribuição de CSI (Barbo Adulto) no rio Ocreza
60 m3/s 50 m3/s
80 m3/s
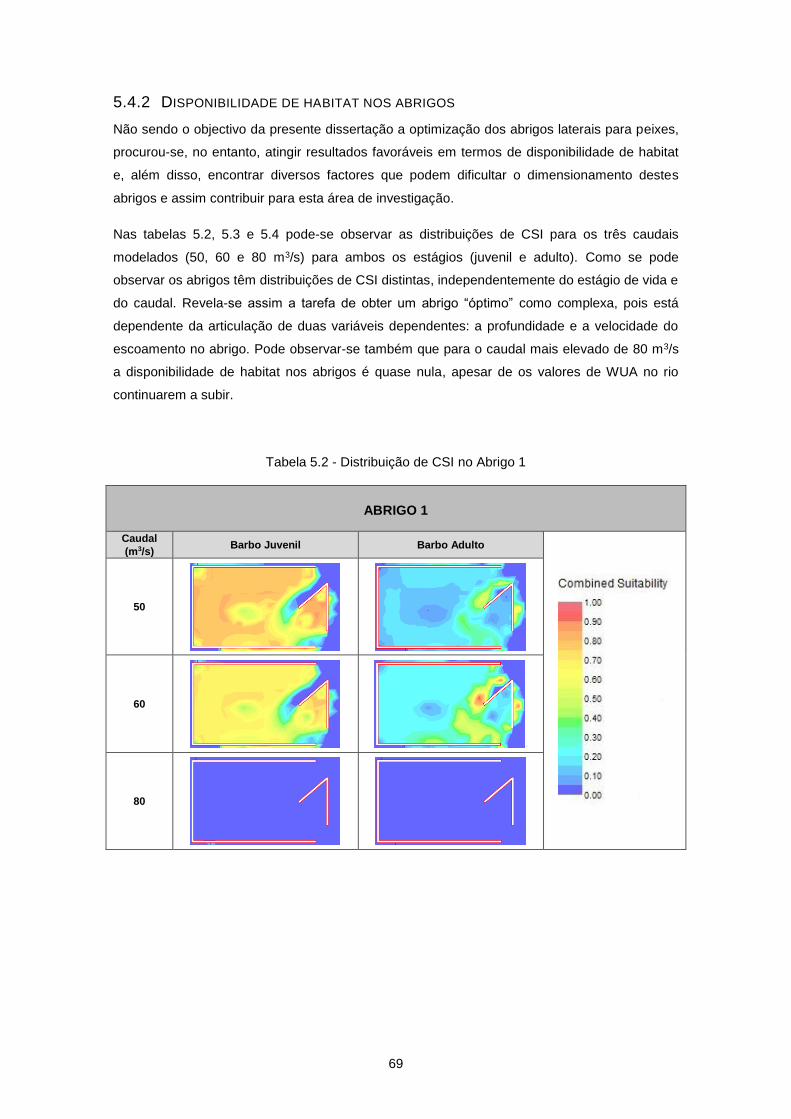
69
5.4.2 DISPONIBILIDADE DE HABITAT NOS ABRIGOS
Não sendo o objectivo da presente dissertação a optimização dos abrigos laterais para peixes,
procurou-se, no entanto, atingir resultados favoráveis em termos de disponibilidade de habitat
e, além disso, encontrar diversos factores que podem dificultar o dimensionamento destes
abrigos e assim contribuir para esta área de investigação.
Nas tabelas 5.2, 5.3 e 5.4 pode-se observar as distribuições de CSI para os três caudais
modelados (50, 60 e 80 m3/s) para ambos os estágios (juvenil e adulto). Como se pode
observar os abrigos têm distribuições de CSI distintas, independentemente do estágio de vida e
do caudal. Revela-se assim a tarefa de obter um abrigo “óptimo” como complexa, pois está
dependente da articulação de duas variáveis dependentes: a profundidade e a velocidade do
escoamento no abrigo. Pode observar-se também que para o caudal mais elevado de 80 m3/s
a disponibilidade de habitat nos abrigos é quase nula, apesar de os valores de WUA no rio
continuarem a subir.
Tabela 5.2 - Distribuição de CSI no Abrigo 1
ABRIGO 1
Caudal
(m3/s) Barbo Juvenil Barbo Adulto
50
60
~
80
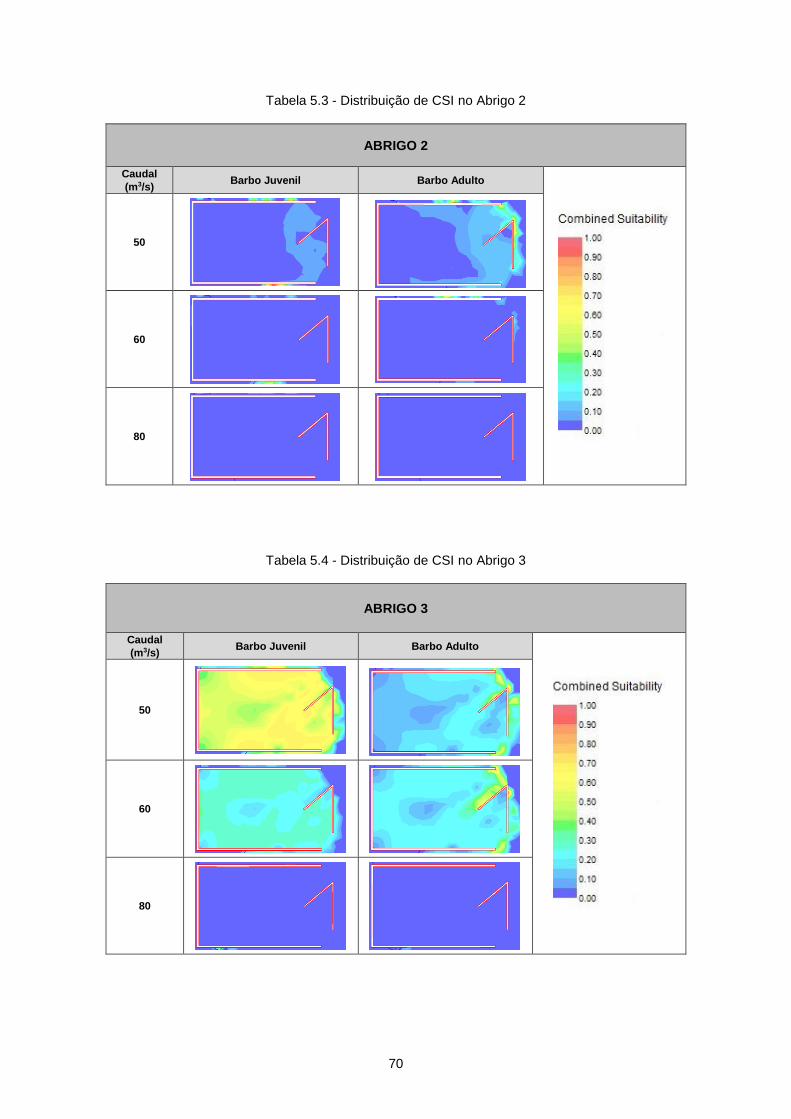
70
Tabela 5.3 - Distribuição de CSI no Abrigo 2
ABRIGO 2
Caudal
(m3/s) Barbo Juvenil Barbo Adulto
50
60
80
Tabela 5.4 - Distribuição de CSI no Abrigo 3
ABRIGO 3
Caudal
(m3/s) Barbo Juvenil Barbo Adulto
50
60
80
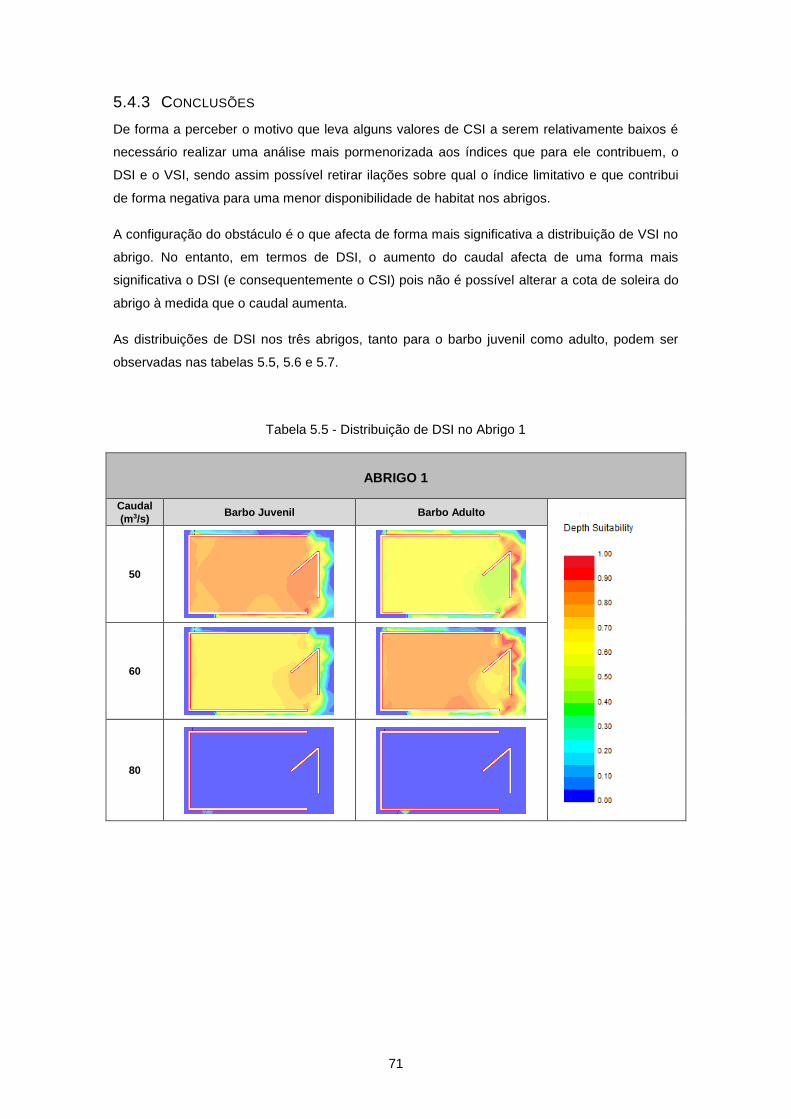
71
5.4.3 CONCLUSÕES
De forma a perceber o motivo que leva alguns valores de CSI a serem relativamente baixos é
necessário realizar uma análise mais pormenorizada aos índices que para ele contribuem, o
DSI e o VSI, sendo assim possível retirar ilações sobre qual o índice limitativo e que contribui
de forma negativa para uma menor disponibilidade de habitat nos abrigos.
A configuração do obstáculo é o que afecta de forma mais significativa a distribuição de VSI no
abrigo. No entanto, em termos de DSI, o aumento do caudal afecta de uma forma mais
significativa o DSI (e consequentemente o CSI) pois não é possível alterar a cota de soleira do
abrigo à medida que o caudal aumenta.
As distribuições de DSI nos três abrigos, tanto para o barbo juvenil como adulto, podem ser
observadas nas tabelas 5.5, 5.6 e 5.7.
Tabela 5.5 - Distribuição de DSI no Abrigo 1
ABRIGO 1
Caudal
(m3/s) Barbo Juvenil Barbo Adulto
50
60
80
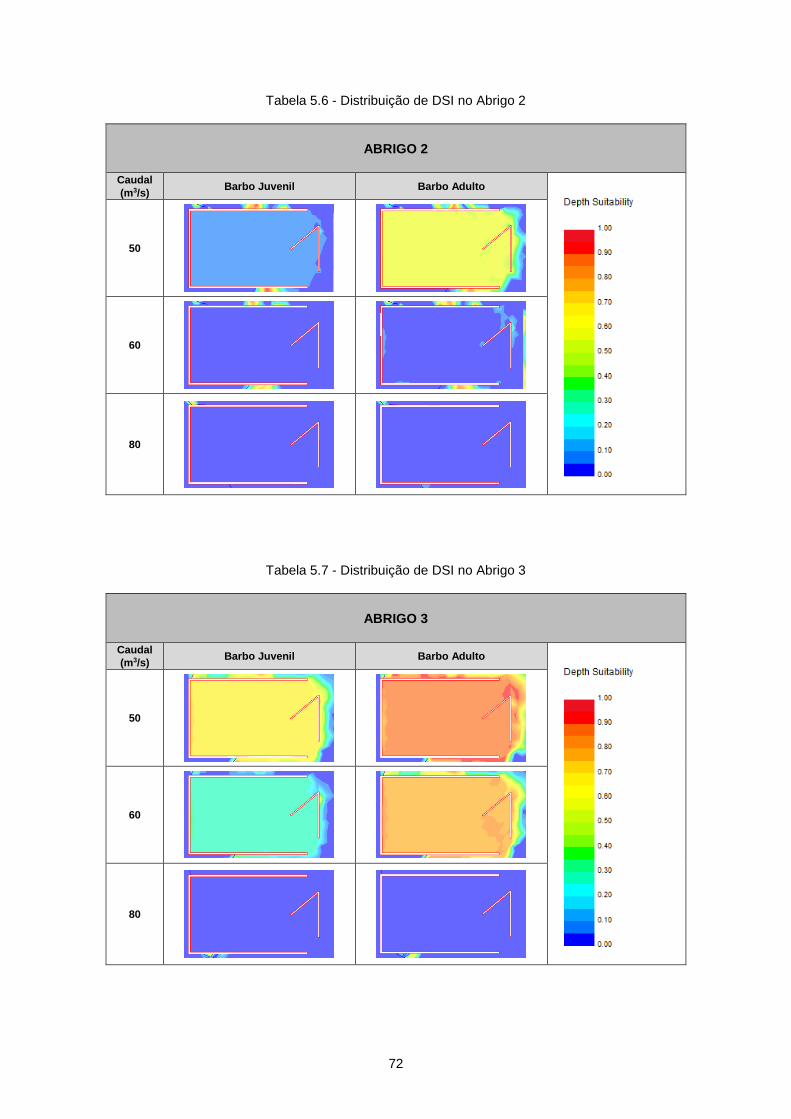
72
Tabela 5.6 - Distribuição de DSI no Abrigo 2
ABRIGO 2
Caudal
(m3/s) Barbo Juvenil Barbo Adulto
50
60
80
Tabela 5.7 - Distribuição de DSI no Abrigo 3
ABRIGO 3
Caudal
(m3/s) Barbo Juvenil Barbo Adulto
50
60
80
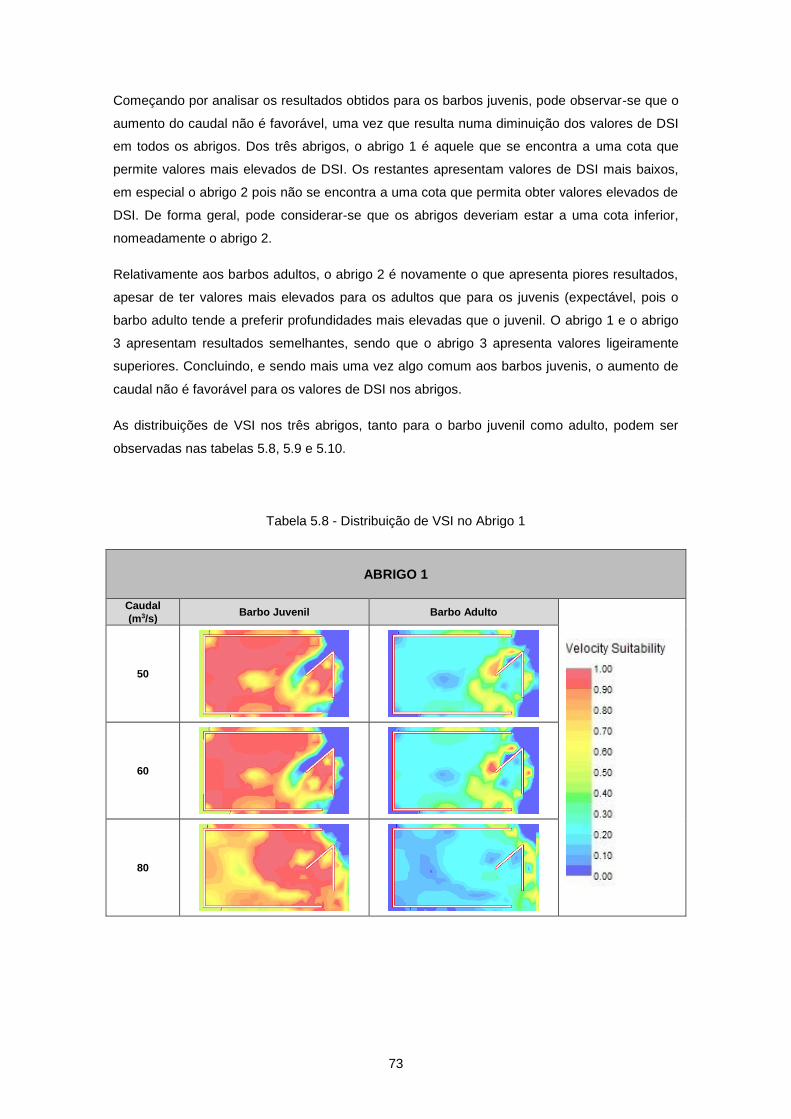
73
Começando por analisar os resultados obtidos para os barbos juvenis, pode observar-se que o
aumento do caudal não é favorável, uma vez que resulta numa diminuição dos valores de DSI
em todos os abrigos. Dos três abrigos, o abrigo 1 é aquele que se encontra a uma cota que
permite valores mais elevados de DSI. Os restantes apresentam valores de DSI mais baixos,
em especial o abrigo 2 pois não se encontra a uma cota que permita obter valores elevados de
DSI. De forma geral, pode considerar-se que os abrigos deveriam estar a uma cota inferior,
nomeadamente o abrigo 2.
Relativamente aos barbos adultos, o abrigo 2 é novamente o que apresenta piores resultados,
apesar de ter valores mais elevados para os adultos que para os juvenis (expectável, pois o
barbo adulto tende a preferir profundidades mais elevadas que o juvenil. O abrigo 1 e o abrigo
3 apresentam resultados semelhantes, sendo que o abrigo 3 apresenta valores ligeiramente
superiores. Concluindo, e sendo mais uma vez algo comum aos barbos juvenis, o aumento de
caudal não é favorável para os valores de DSI nos abrigos.
As distribuições de VSI nos três abrigos, tanto para o barbo juvenil como adulto, podem ser
observadas nas tabelas 5.8, 5.9 e 5.10.
Tabela 5.8 - Distribuição de VSI no Abrigo 1
ABRIGO 1
Caudal
(m3/s) Barbo Juvenil Barbo Adulto
50
60
80
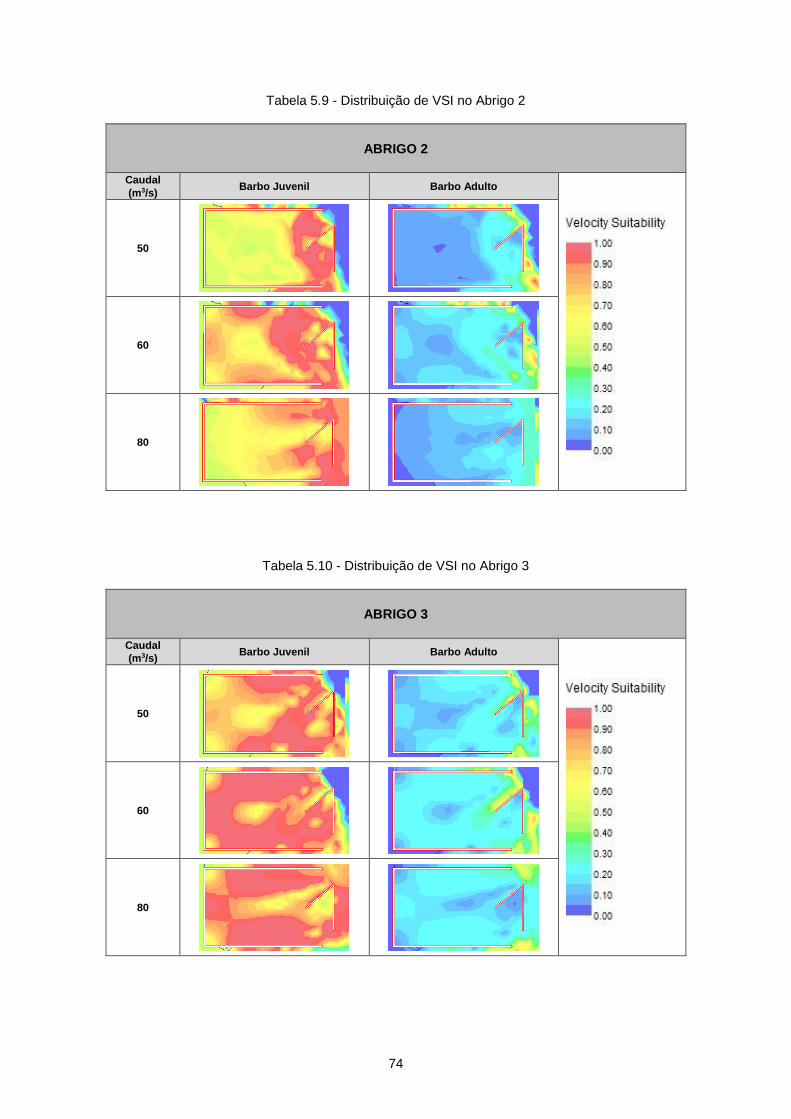
74
Tabela 5.9 - Distribuição de VSI no Abrigo 2
ABRIGO 2
Caudal
(m3/s) Barbo Juvenil Barbo Adulto
50
60
80
Tabela 5.10 - Distribuição de VSI no Abrigo 3
ABRIGO 3
Caudal
(m3/s) Barbo Juvenil Barbo Adulto
50
60
80

75
Apesar de a configuração escolhida para o abrigo ter sido feita com o objectivo de combinar as
características mais favoráveis para os barbos juvenis e adultos, as profundidades verificadas
no abrigo acabam por influenciar também a velocidade e, consequentemente, os valores de
VSI verificados.
Os valores de VSI são elevados para os abrigos 1 e 3, sendo inferiores no abrigo 2, no caso
dos barbos juvenis. De referir, como já foi observado anteriormente, que o abrigo 2 é aquele
que apresenta piores valores de DSI para ambos os estágios de vida. No barbo adulto os
valores de VSI são relativamente mais baixos, o que seria expectável face ao que acontece
para os juvenis, pois os valores de preferência máxima em termos de velocidade diferem
bastante entre estágios de vida: cerca de 0,1 m/s e 0,9 m/s para os juvenis e para os adultos,
respectivamente.
Tendo em conta esta análise pode concluir-se que:
A utilização de uma única configuração de abrigo para uma espécie só é possível
quando as preferências em termos de velocidade para ambos os estágios são
semelhantes;
Em função do caudal, a profundidade no abrigo altera-se, influenciando também os
valores de velocidade, alterando desta forma a disponibilidade de habitat;
A colocação de abrigos a cotas significativamente diferentes pode ser favorável,
funcionando assim cada abrigo para um caudal diferente.

76

77
6 CONCLUSÕES E RECOMENDAÇÕES
6.1 CONCLUSÕES
A presente dissertação aborda a modelação hidrodinâmica de habitat de abrigos laterais para
peixes, tendo como primeira fase a calibração do modelo numérico através da comparação de
resultados com os obtidos numa instalação experimental no âmbito de uma dissertação de
doutoramento na École Polythecnique Fédérale de Lausanne. O modelo aplicado foi o River2D,
cujo funcionamento é brevemente explicado nesta dissertação.
Integrando curvas de preferência de habitat, foi possível comparar os resultados numéricos
com os obtidos em ensaios experimentais, traduzidos em termos de taxa de presença dos
peixes nos abrigos. Verificou-se uma boa correlação entre os resultados obtidos nos ensaios
experimentais e nos modelos numéricos. Ainda assim, e tendo como referência as
configurações testadas experimentalmente (configurações C), o habitat foi simulado para
outras espécies, estágios de vida e períodos do ano, de forma a encontrar uma relação entre
espécies.
As diferenças nas preferências de velocidade entre a espécie utilizada nos ensaios
experimentais (truta juvenil) e as estabelecidas para o rio Lima poderão ajudar a explicar os
resultados discordantes. Diversos autores referem a importância de desenvolver curvas de
preferência de habitat do local em estudo, pois as necessidades de habitat dependem não só
do estágio de vida mas também do tipo de rio. Vismara et al. (2001) encontrou diferenças entre
curvas de preferência obtidas em regiões diferentes, o que vem reforçar a necessidade de
utilização de curvas de preferência do local em estudo (Moyle et al., 1985; Orth, 1987), para o
correcto dimensionamento dos abrigos laterais para peixes.
De facto, existem diferenças entre as preferências da truta do rio Lima e as estudadas por
Vismara et al. (2001). Se para preferências em termos de velocidade das trutas juvenis os
resultados são similares (i.e., maior VSI ocorre para velocidades baixas), o mesmo não
acontece para as adultas. A truta do rio Lima aparenta suportar melhor as elevadas
velocidades típicas de um rio montanhoso. Em termos de preferência, no que diz respeito às
baixas profundidades de escoamento, os resultados entre curvas são semelhantes. Contudo,
notam-se grandes discrepâncias para profundidades elevadas, o que pode ser causado pelas
diferentes morfologias dos rios onde as curvas foram desenvolvidas. No caso da instalação
experimental, as profundidades verificadas (0,24 a 0,27 m) não são favoráveis para as trutas
adultas, no que resulta em valores mais baixos nas simulações de habitat (HHS). Isto explica a
baixa disponibilidade de habitat para os adultos. Na Figura 6.1 podem ser observadas
diferentes curvas de preferência de diferentes regiões, juntamente com as curvas de
preferência da truta do rio Lima.
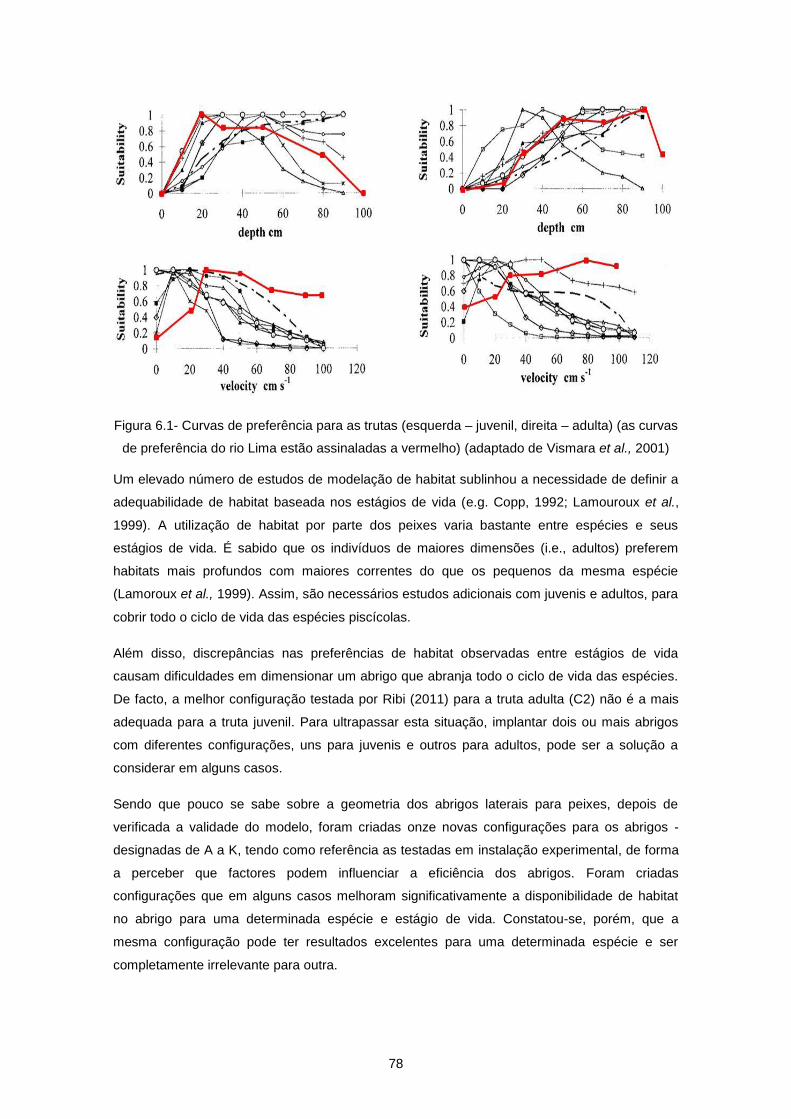
78
Figura 6.1- Curvas de preferência para as trutas (esquerda – juvenil, direita – adulta) (as curvas
de preferência do rio Lima estão assinaladas a vermelho) (adaptado de Vismara et al., 2001)
Um elevado número de estudos de modelação de habitat sublinhou a necessidade de definir a
adequabilidade de habitat baseada nos estágios de vida (e.g. Copp, 1992; Lamouroux et al.,
1999). A utilização de habitat por parte dos peixes varia bastante entre espécies e seus
estágios de vida. É sabido que os indivíduos de maiores dimensões (i.e., adultos) preferem
habitats mais profundos com maiores correntes do que os pequenos da mesma espécie
(Lamoroux et al., 1999). Assim, são necessários estudos adicionais com juvenis e adultos, para
cobrir todo o ciclo de vida das espécies piscícolas.
Além disso, discrepâncias nas preferências de habitat observadas entre estágios de vida
causam dificuldades em dimensionar um abrigo que abranja todo o ciclo de vida das espécies.
De facto, a melhor configuração testada por Ribi (2011) para a truta adulta (C2) não é a mais
adequada para a truta juvenil. Para ultrapassar esta situação, implantar dois ou mais abrigos
com diferentes configurações, uns para juvenis e outros para adultos, pode ser a solução a
considerar em alguns casos.
Sendo que pouco se sabe sobre a geometria dos abrigos laterais para peixes, depois de
verificada a validade do modelo, foram criadas onze novas configurações para os abrigos -
designadas de A a K, tendo como referência as testadas em instalação experimental, de forma
a perceber que factores podem influenciar a eficiência dos abrigos. Foram criadas
configurações que em alguns casos melhoram significativamente a disponibilidade de habitat
no abrigo para uma determinada espécie e estágio de vida. Constatou-se, porém, que a
mesma configuração pode ter resultados excelentes para uma determinada espécie e ser
completamente irrelevante para outra.
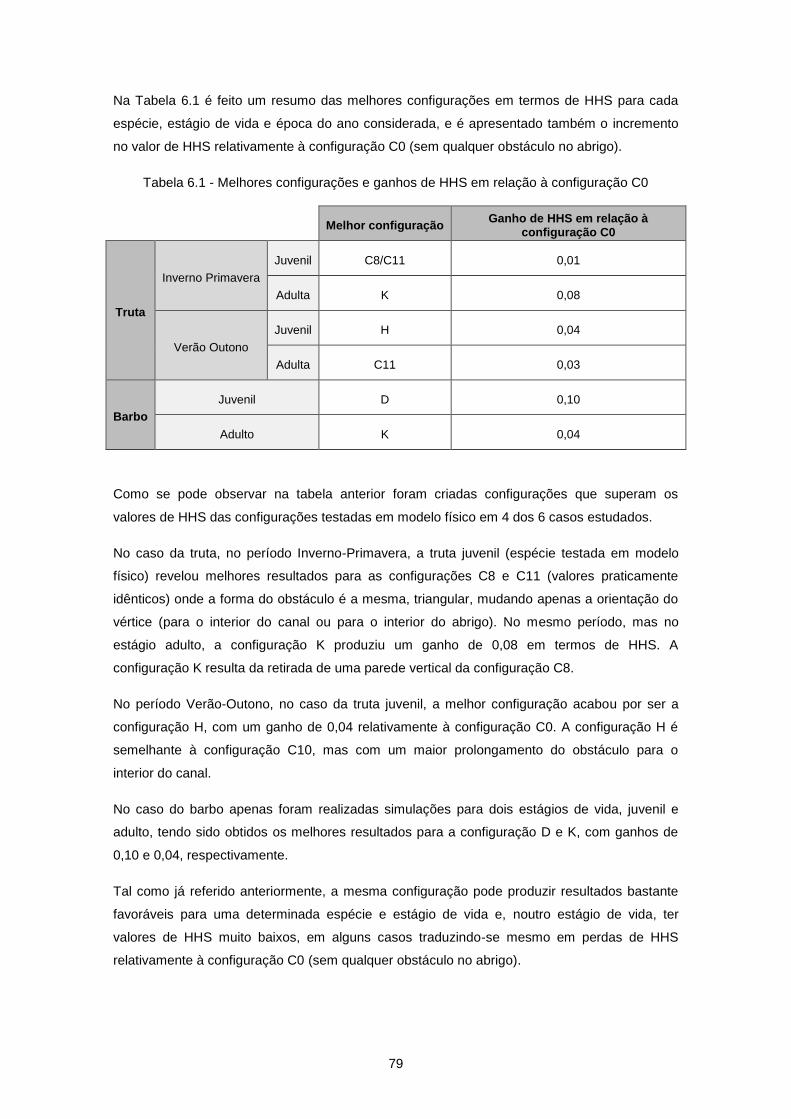
79
Na Tabela 6.1 é feito um resumo das melhores configurações em termos de HHS para cada
espécie, estágio de vida e época do ano considerada, e é apresentado também o incremento
no valor de HHS relativamente à configuração C0 (sem qualquer obstáculo no abrigo).
Tabela 6.1 - Melhores configurações e ganhos de HHS em relação à configuração C0
Melhor configuração
Ganho de HHS em relação à configuração C0
Truta
Inverno Primavera
Juvenil C8/C11 0,01
Adulta K 0,08
Verão Outono
Juvenil H 0,04
Adulta C11 0,03
Barbo
Juvenil D 0,10
Adulto K 0,04
Como se pode observar na tabela anterior foram criadas configurações que superam os
valores de HHS das configurações testadas em modelo físico em 4 dos 6 casos estudados.
No caso da truta, no período Inverno-Primavera, a truta juvenil (espécie testada em modelo
físico) revelou melhores resultados para as configurações C8 e C11 (valores praticamente
idênticos) onde a forma do obstáculo é a mesma, triangular, mudando apenas a orientação do
vértice (para o interior do canal ou para o interior do abrigo). No mesmo período, mas no
estágio adulto, a configuração K produziu um ganho de 0,08 em termos de HHS. A
configuração K resulta da retirada de uma parede vertical da configuração C8.
No período Verão-Outono, no caso da truta juvenil, a melhor configuração acabou por ser a
configuração H, com um ganho de 0,04 relativamente à configuração C0. A configuração H é
semelhante à configuração C10, mas com um maior prolongamento do obstáculo para o
interior do canal.
No caso do barbo apenas foram realizadas simulações para dois estágios de vida, juvenil e
adulto, tendo sido obtidos os melhores resultados para a configuração D e K, com ganhos de
0,10 e 0,04, respectivamente.
Tal como já referido anteriormente, a mesma configuração pode produzir resultados bastante
favoráveis para uma determinada espécie e estágio de vida e, noutro estágio de vida, ter
valores de HHS muito baixos, em alguns casos traduzindo-se mesmo em perdas de HHS
relativamente à configuração C0 (sem qualquer obstáculo no abrigo).

80
De forma a simular a aplicação destes abrigos, aplicaram-se os abrigos laterais ao rio Ocreza.
Simulou-se o habitat para a espécie mais relevante neste rio, o barbo, e utilizando o mesmo
tipo de abrigo para ambos os estágios de vida. Para determinar a disponibilidade de habitat no
rio Ocreza foi utilizado o modelo Casimir Fish 2D. A utilização do mesmo tipo de abrigo para os
diferentes estágios de vida revelou-se algo desapropriada, sendo que a colocação de todos os
abrigos aproximadamente à mesma cota também se revelou incorrecta. No entanto, e apesar
de para caudais elevados a disponibilidade de habitat nos abrigos ser quase nula, a WUA
aumenta de uma forma geral ao longo do rio, conseguindo-se ganhos de cerca de 200 e 600
m2, respectivamente para o barbo juvenil e adulto, utilizando três abrigos com cerca de 24 m2.
6.2 RECOMENDAÇÕES
Presentemente pouco se conhece relativamente a abrigos laterais para peixes especialmente
no que diz respeito à(s) configuração(ões) a adoptar. Assim, como recomendações para
estudos futuros tem-se:
Testar diferentes configurações para os abrigos laterais, bem como a sua geometria;
Estudar a influência de abrigos em diferentes margens, não só para a mesma espécie
e estágio de vida mas também para casos diferentes;
Estudar a relação entre a existência de dois abrigos na mesma margem e a distância
óptima entre eles, pois um abrigo afecta as condições de habitat a jusante. Se a
jusante deste primeiro abrigo existir outro, importa perceber qual a disponibilidade de
habitat do segundo abrigo;
Estudar a colocação de abrigos a diferentes cotas num curso de água natural. A
existência de abrigos a cotas diferentes (e para caudais diferentes) pode revelar-se
benéfica;
Estudar a utilização de um abrigo com profundidade variável na sua
Estudar os padrões de escoamento decorrentes da morfologia do rio em estudo;
Criar condições, para os caudais mais frequentes, para que a velocidade de
preferência máxima possa coincidir com a profundidade de preferência máxima;
Utilizar, juntamente com os modelos hidrodinâmicos de habitat, um modelo físico;
Construir abrigo lateral para peixes e acompanhar a sua evolução e taxa de presença
em condições de hydropeaking de forma a validar os ensaios em instalação
experimental e as simulações numéricas.

81
BIBLIOGRAFIA
Bain, M.B., Stevenson, N.J. 1999. Aquatic Habitat Assessment. American Fisheries Society.
Bethesda. Maryland. 216 pp.
Berg, O.K., Arnekleiv, J.V., Lohrmann, A. 2006. The influence of hydroelectric power generation
on the body composition of juvenile Atlantic salmon. River Research and Applications 22: 993-
1008.
Boavida, I. 2012. Ecohydraulic modelling for river restoration in Mediterrean-type streams,
Instituto Superior Técnico, Universidade Técnica de Lisboa.
Boavida, I., Santos, J.M., Katopodis, C., Pinheiro, A.N., Ferreira, M.T. 2013. Uncertainty in
predicting the fish-response to two-dimensional habitat modelling using field data. River
Research and Applications, 29(9): 1164-1174.
Bovee, K.D. 1978. The incremental method of habitat potential for coolwater species with
management implications. American Fisheries Society Special Publication 11, pp. 340-346.
Bovee, K.D. 1982. A guide to stream habitat analysis using the instream flow incremental
methodology, Instream Flow Information Paper 12. USDI Fish and Wildlife Service. Washington,
248 pp.
Bovee, K.D. 1996. Perspectives on two-dimensional river habitat models: the PHABSIM
Experience. In: Leclerc, M., Capra, H., Valentin, S., Boudreault, A., Côté, Y., Eds. Ecohydraulics
2000 Proceedings of Second International Symposium on Habitat Hydraulics B. INRS-Eau &
FQSA, IAHR/AIRH, pp. B149-B162.
Bruno, M.C., Siviglia, A. 2012. Assessing impacts of dam operations - interdisciplinary
approaches for sustainable regulated river management. River Research and Applications 28:
675-677.
Carta Piscícola. www.cartapiscicola.org. Janeiro de 2014.
Clarke, K.D., Pratt, T.C., Randall, R.G., Scuton, D.A., Smokorowski, K.E. 2008. Validation of the
flow management pathway: effects of altered flow on fish habitat and fishes downstream from a
hydropower dam. Canadian Technical Report of Fisheries and Aquatic Sciences 2784.
Cortes, R., Ferreira, M.,; Santos, J., Godinho, F., Diogo, R., Albuquerque, A., Fernandes, F.,
Ribeiro, P., Oliveira, S., Lopes, L., Motta, A., Oliveira, D., Tavares, I., Saraiva, Z. 2000. Estudo
Experimental para a definição do caudal ecológico do rio Lima. Vila Real, Universidade de
Trás-os-Montes e Alto Douro, Instituto Superior de Agronomia e Instituto de Conservação da
Natureza.

82
Copp, G.H., 1992. Comparative microhabitat use of cyprinid larvae and juveniles in a lotic
floodplain channel. Environmental Biologic of Fishes 33, 181–193.
Crowder, D.W., Diplas, P. 2000. Using two-dimensional hydrodynamic models at scales of
ecological importance. Journal of Hydrology 230: 172-191. Gesist, D.R., Jones, J., Murray, C.J.,
Dauble, D.D. 2000. Suitability criteria analyzed at the spatial scale of red clusters improved
estimates of fall Chinook salmon spawning habitat use in the Hanford Reach, Columbia River.
Canadian Journal of Fisheries and Aquatic Sciences 57: 1636-1646.
EC. 2000. Council Directive for a legislative frame and actions for the water policy, 2000/60/EC,
Official Journal of the EC 22/12/2000.
DGEG. 2013. Renováveis – Estatísticas rápidas. n.º 106.
Fausch, K.D., White, R.J., 1986. Competition among juveniles of coho salmon, brook trout, and
brown trout in a laboratory stream, and implications for Great Lakes tributaries. Transactions of
the American Fisheries Society. 115, 363–381.
Freeman, M.C., Bowen, Z.H., Crance, J.H., 1997. Transferability of habitat suitability criteria for
fishes in warmwater streams. North American Journal of Fisheries Management 17, 20–31.
Geist, D.R., Jones, J., Murray, C.J., Dauble, D.D. 2000. Suitability criteria analyzed at the
spatial scale of red clusters improved estimates of fall Chinook salmon spawning habitat use in
the Hanford Reach, Columbia River. Canadian Journal of Fisheries and Aquatic Sciences 57:
1636-1646.
Ghanem, A., Steffler, P., Hicks, F., Katopodis, C. 1996. Two dimensional hydraulic simulation of
physical habitat conditions in flowing streams. Regulated Rivers: Resources and Management
12: 185-200. Harby, A., Noack, M. 2013. Rapid Flow Fluctuations and Impacts on Fish and the
Aquatic Ecosystem. Ecohydraulics – an integrated approach. Wiley Blackwell.
Harby, A., Noack, M. 2013. Rapid Flow Fluctuations and Impacts on Fish and the Aquatic
Ecossystem. Ecohydraulics – an integrated approach, Wiley Blackwell.
Jungwirth, M., Haidvogel, G., Moog, O., Muhar, S., Schmutz, S. 2003. Angewandte
Fischökologie an Fliessgewässern, UTB Verlag Wien.
Junk, W.J., Bayley, P.B., Sparks, R.E. 1989. The flood pulse concept in river floodplain
systems. Canadian Journal of Fisheries and Aquatic Science, 106.: 110-127.
Knapp, R.A., Preisler, H.K.1999. Is it possible to predict habitat use by spawning salmonds? A
test using California golden trout (Oncorhynchus mykiss aguabonita). Canadian Journal of Fish
and Aquatic Sciences 56. 1576 – 1584.

83
Korman, J., Perrin, C.J., Lekstrum, T. 1994. A Guide for the Selection of Standard Methods for
Quantifying Sportfish Habitat Capability and Suitability in Streams and Lakes of British
Columbia. Report to B.C. Environment Fisheries Branch, Vancouver: British Columbia.
Kynard, B., Horgan, M., Kieffer, M. 2000. Habitat used by shortnose sturgeon in two
Massachusetts rivers, with notes on estuarine Atlantic sturgeon: a hierarchical approach.
Transactions of the American Fisheries Society 129: 487 – 503.
Lamouroux, N., Olivier, J.M., Persat, H., Pouilly, M., Souchon, Y., Statzner, B., 1999 Predicting
community characteristics from habitat conditions: fluvial fish and hydraulics. Freshwater
Biology 42, 275–299.
Leclerc, M., Boudreault, A., Bechara, J.A., Corfa, G. 1995. Two-dimensional hydrodynamic
modeling: a neglected tool in the instream flow incremental methodology. Transactions of the
American Fisheries Society 123: 645-662.
Liebig, H., Lim, P., Belaud, A. 1998. Influence of basic flow and hydropeaking duration on the
drift of post-emergent fry of brown trout: experiments on a semi-natural stream. Bulettin
Français de la Peche et de la Pisciculture (350-51): 337-347.
Matthews, W.J. 1998. Patterns in Freshwater Fish Ecology. Chapman and Hall. New York.
Meile, T. 2008. Influence of Macro-Roughness of Walls on Steady and Unsteady Flow in a Channel, École Polytechinque Fédérale de Lausanne. Milhous, R.T., Updike, M.A., Schneider, D.M. 1989. Physical habitat simulation system reference manual – version II. U.S. Fish and Wildlife Service Biological Report 89(16): 403. Milhous, R.T. 1999. History, theory use and limitations of the physical habitat simulation system,
Proceedings of the 3rd International Symposium on Ecohhydraulics, CD-ROM published by Utah
State University Extension, Logan, Utah.
Moog, O. 1993. Quantification of daily peak hydropower effects on aquatic fauna and
management do minimize environmental impacts. Regulated Rivers: Research & Management
8(1-2): 5-14.
Moyle, P.B., Baltz, D.M., 1985. Microhabitat use by an assemblage of California stream fishes:
developing criteria for instream flow determinations. Transactions of the American Fisheries
Society. 114, 695-704.
Nagrodski, A., Raby, G.D., Hasler, C.T., Taylor, M.K., Cooke, S.J. 2012. Fish stranding in
freshwater systems: sources, consequences, and mitigation. Journal of Environmental
Management 103: 133-141
Orth, D.J., 1987. Ecological considerations in the development and application of instream flow-
habitat models. Regulated Rivers: Research and Management. 1, 171–181

84
REN 21. 2012. Renewables 2012 Global Status Report.
Ribi, J.-M., Steffen, K., Boillat, J.-L., Peter, A., Schleiss, A.J. 2009. Influence of geometry fish
shelters in river banks on their attractiveness for fishes during hydropeaking, Proceedings of the
33rd Congress of IAHR, Vancouver, Canada, August 9-14, ISBN: 978-94-90365-01-1. pp. 6074-
6081.
Ribi, J.-M., Boillat, J.-L., Peter, A., Schleiss, A.J. 2010. Fish behaviour during hydropeaking in a
channel equipped with a lateral shelter. Proceedings of the 8th International Symposium on
Ecohydraulics (ISE,2010), COEX, Seoul, Korea, September 12-16 2010, p.675-682.
Ribi, J.-M. 2011. Etude expérimentale de refuges à poissons aménagés dans les berges de
rivières soumises aux éclusées hydroélectriques, École Polytechnique Fédérale de Lausanne.
Rodi, W., Pavlovic, R.N., Srivatsa, S.K. 1981. Prediction of flow and polluant spreading in rivers.
In Fisher, H.B. ed. Transport Models for Inland and Coastal Waters, Academic Press, New
York, NY, p. 542.
Saltveit, S.J., Halleraker, J.H., Arnekleiv, J.V., Harby, A. 2001. Field experiments on stranding in
juvenile Atlantic salmon (Salmo salar) and brown trout (Salmo trutta) during rapid flow
decreases caused by hydropeaking. Regulated Rivers: Research & Management 17: 609-622.
Schneider, M., Noack, M., Gebler, T., Kopecki, I. 2010. Handbook for the Habitat Simulation
Model Casimir – Module: Casimir-Fish Base Version. Institut für Wasserbau, Universität
Stuttgart.
Schweizer, S.,Neuner, J., Ursin, M., Tscholl, H., Meyer, M. 2009. Trotz Ausbau der
Kraftwerksleistung – deutlich geringere Pegelschwankungen inder Hasliaare.
Scruton, D.A., Pennell, C., Ollerhead, L.M.N., Alfredson, K., Stickler, M., Harby, A., Robertson,
M., Clarke, K.D., LeDrew, L.J. 2008. A synopsis of 'hydropeaking' studies on the response of
juvenile Atlantic salmon to experimental flow alteration. Hydrobiologia 609: 263-275
Stalnaker, C., Lamb, B. L., Henriksen, J., Bovee, K., Bartholow, J. 1995. The Instream Flow
Incremental Methodology – A primer for IFIM, Biological Report 29, National Biological Service,
US Department of the Interior, 45 pp.
Steffler, P. 2000. Software River2D. Two Dimensional Depth Averaged Finite Element
Hydrodynamic Model. University of Alberta, Canada.
Steffler, P. 2002. Bed topography file editor. User's manual. University of Alberta, Canada.
Steffler, P., Blackburn, J. 2002. Two-dimensional depth averaged model of river hydrodynamics
and fish habitat. Introduction to depth averaged modeling and user’s manual. Edmonton,
Alberta, Canada, University of Alberta.

85
Tonina, D., Jorde, K. 2013. Hydraulic Modelling Approaches for Ecohydraulic Studies: 3D, 2D,
1D and Non-Numerical Models. Ecohydraulics – an integrated approach, Wiley Blackwell.
Tuhtan, J.A., Noack, M., Wieprecht, S. 2012. Estimating stranding risk due to hydropeaking for
juvenile european grayling considering river morphology. Journal of Civil Engineering 16: 197-
206.
Vadas, R.I., Orth, D.J. 2001. Formulation of habitat suitability models for stream fish guilds: do
the standart methods work? Transactions of the American Fisheries Society 130: 217 – 235.
Valente, A.C. 1993. Biologia e Dinâmica das populações de truta de rio da bacia hidrográfica do
Lima. Tese de Doutoramento. Faculdade de Ciências do Porto. Porto.
Vilizzi, L., Copp, G.H. 2005. An analysis of 0+ barbel (Barbus barbus) response to discharge
fluctuations in a flume. River Research and Applications 21: 421-438.
Vismara R, Azzellino A, Bosi R, Crosa G, Gentili G. 2001. Habitat suitability curves for brown
trout (Salmo truttafario L.) in the River Adda, Northern Italy: comparing univariate and
multivariate approaches. Regulated Rivers: Research and Management 17: 37–50.
Waddle, T., Steffler, P. 2002. Mesh generation program for River 2D two dimensional depth
averaged finite element. Introduction to mesh generation and user’s manual. U.S. Geological
Survey

86
I
ANEXOS
ANEXO A – MALHAS DE ELEMENTOS FINITOS. CONFIGURAÇÕES C
Configuração Boundary Nodes (m) Uniform Fill (m) Ângulo (º) QI (-) Número de nós
C0 0,10 0,05 0 0.37 8594
C1 0,10 0,05 0 0.34 8638
C2 0,10 0,05 0 0.35 8689
C3 0,10 0,05 0 0.33 8672
C4 0,10 0,05 0 0.32 8670
C5 0,10 0,05 0 0.38 8686
C6 0,10 0,05 0 0.36 8676
C7 0,10 0,05 0 0.40 8579
C8 0,10 0,05 0 0.38 8512
C9 0,10 0,05 0 0.31 8721
C10 0,10 0,05 0 0.31 8700
C11 0,10 0,05 0 0.37 8520

II
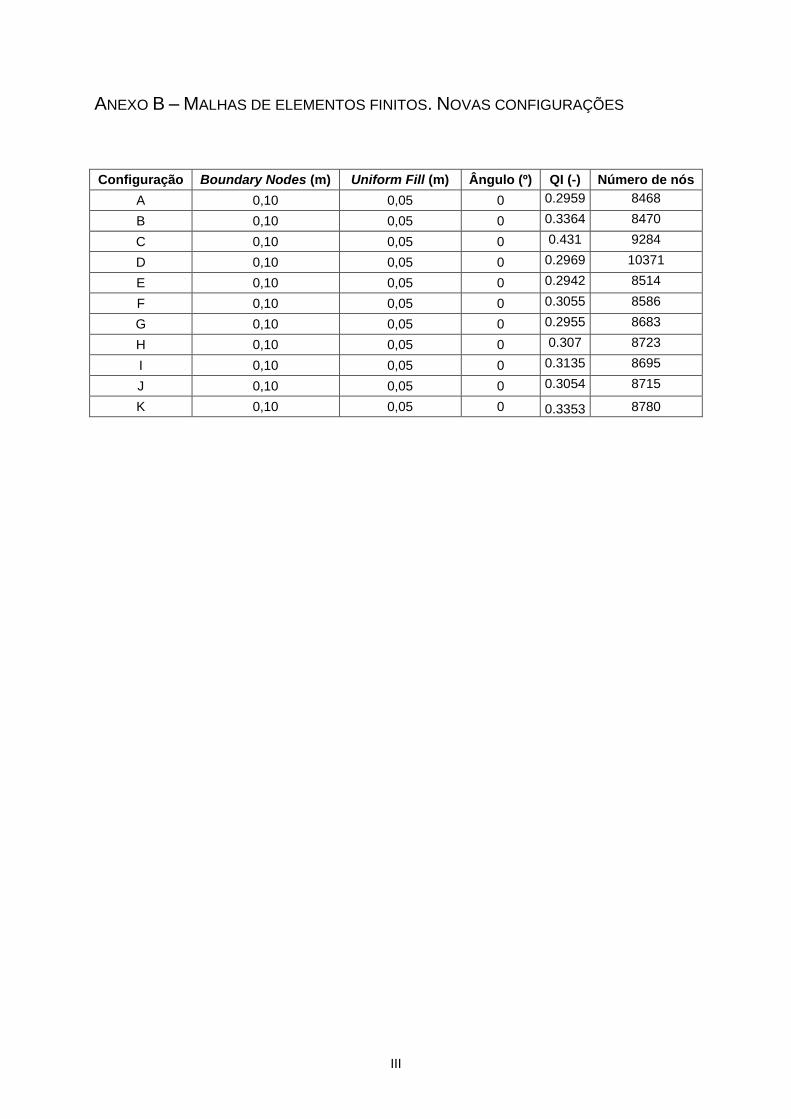
III
ANEXO B – MALHAS DE ELEMENTOS FINITOS. NOVAS CONFIGURAÇÕES
Configuração Boundary Nodes (m) Uniform Fill (m) Ângulo (º) QI (-) Número de nós
A 0,10 0,05 0 0.2959 8468
B 0,10 0,05 0 0.3364 8470
C 0,10 0,05 0 0.431 9284
D 0,10 0,05 0 0.2969 10371
E 0,10 0,05 0 0.2942 8514
F 0,10 0,05 0 0.3055 8586
G 0,10 0,05 0 0.2955 8683
H 0,10 0,05 0 0.307 8723
I 0,10 0,05 0 0.3135 8695
J 0,10 0,05 0 0.3054 8715
K 0,10 0,05 0 0.3353 8780

IV