EVOLUTION OF THE EYE - Smith Lab › ... › slides-Swamynathan_evolution.pdf · The division is based largely on embryological differences. Echinoderms, chordates, molluscs,\ഠannelids
Evolution can be described as a process of surviving unpredictable events in often unpredictable ways. This becomes much more evident when you consider the diversity of eye types in animals and their evolutionary history.
1860- The eye to this day gives me a cold shudder, but when I think of the fine known gradations, my reason tells me I ought to conquer the cold shudder.
1872- To suppose that the eye with all its inimitable contrivances for adjusting the focus to different distances, for admitting different amounts of light, and for the correction of spherical and chromatic aberration, could have been formed by natural selection, seems, I freely confess, absurd in the highest degree. When it was first said that the sun stood still and the world turned round, the common sense of mankind declared the doctrine false; but the old saying of Vox populi, vox Dei, (the voice of the people is the voice of God) as every philosopher knows, cannot be trusted in science. Reason tells me, that if numerous gradations from a simple and imperfect eye to one complex and perfect can be shown to exist, each grade being useful to its possessor, as is certainly the case; if further, the eye ever varies and the variations be inherited, as is likewise certainly the case; and if such variations should be useful to any animal under changing conditions of life, then the difficulty of believing that a perfect and complex eye could be formed by natural selection, though insuperable by our imagination, should not be considered as subversive of the theory.
Charles Darwin on evolution of eyes
A working definition for an eye
Eye is an organ that provides spatial vision
What is Spatial Vision?
Ability to detect an image by comparing light intensities in different directions
Eyes first appeared in early metazoans Insects, crustaceans, annelid worms, molluscs
Presenter
Presentation Notes
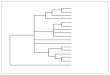
Origin of vertebrates. The origin of vertebrates, over a time-scale from roughly 700 to 400 million years ago (Mya), with timings of the branchings taken from a reconciliation of fossil and molecular data by Erwin et al. (2011). The red curve indicates our direct ancestors, beginning with early metazoans, and the numbers along the curve denote major branchings that are significant to the evolution of the vertebrate eye. ①: Cnidarians branched off around 700 Mya, and since then our ancestors have been bilaterally symmetric (bilaterians). ②: Protostomes branched off around 665 Mya, and since then our ancestors have been termed deuterostomes. ③, ④: By the time that cephalochordates and then tunicates diverged (around 650 and 600 Mya), our ancestors possessed a notochord, and are referred to as chordates.⑤: By the time that the ancestors of lampreys diverged around 500 Mya, they and our own ancestors were vertebrates. It has recently been confirmed that the ‘2R’ two rounds of whole-genome duplication occurred prior to that divergence of ancestral lampreys. Subsequently (by around 420 Mya) our own ancestors evolved jaws and hence became jawed vertebrates (gnathostomes). At the base of the tree are the Cnidarians (Hydra, jellyfish) with only two tissue layers, then the flatworms with three layers but no coelom (body cavity). All the later phyla have body cavity. There are two main branches, the deuterostomes which led to the strafish and the vertebrates, and the protostomes which gave rise to the molluscs, annelids and the arthropods. The division is based largely on embryological differences. Echinoderms, chordates, molluscs, annelids and arthropods are all phyla- the major divisions of the animal kingdom. Vertebrates and the three arthropod groups are subphyla, and the four vertebrate groups are classes, as are the three mollusc groups.
Chronology of animal locomotory abilities, and visual function
Presenter
Presentation Notes
A plausible chronology for animal locomotory abilities, visual function and the occurrence of master control genes (herePax-6). The need for fast locomotion and vision are likely to have originated during the Cambrian explosion, whereas master control genes in general would have been needed much earlier. Bilaterally symmetric animals with prominent eyes appear suddenly in the fossil record from the Cambrian explosion (which lasted less than 10 million years. One of the hypotheses for "causes" of Cambrian explosion, the "Light Switch" theory of Andrew Parker, holds that the evolution of eyes initiated an arms race that led to a rapid spate of evolution.
Eyes and vision : Unity in diversity
An optical system that can discriminate the direction of light to within a few degrees is present in only 6 of the 35 animal phyla. Yet, they account for 96% of all animal species
Presenter
Presentation Notes
An optical system that can discriminate the direction of light to within a few degrees is present in only 6 of the 35 animal phyla. Yet, they account for 96% of all animal species. Given the variety of eyes found throughout the animal kingdom, evolutionary biologists once thought eyes had evolved independently dozens or even hundreds of times. Thanks to DNA sequencing and other molecular tools, we know today that modern eyes are built from many of the same genes. Ancient "toolkit" genes (such as opsins, Pax, and Otx) first evolved in a primitive ancestor that gave rise to all animals with eyes. These genes have been preserved throughout evolution, and today we still find them at work in all types of eyes. The diversity of modern animal eyes is the result of refinements and specializations built on top of this basic genetic framework. Not all eye features are built using the same genes. Lenses, for example, are refinements that arose when different genes were recruited to perform a similar task in different organisms. Similarly, shielding pigments arose from a variety of genes. Molecular analysis has shown that many genes in the eye, whether shared among organisms or not, had other functions first. They were later recruited to take on second jobs.
Diversity of eye types in the animal kingdom
Eyes
Simple (Non-Compound) Eyes Compound Eyes
1. Pit Eyes (Found in ~85% of phyla) 2. Spherical Lensed Eye (Gastropods and annelids) 3. Multiple Lenses in the Optical Path (Copepods) 4. Refractive Cornea (Terrestrial vertebrates and birds) 5. Reflector Eyes (Scallops, Spookfish)
6. Apposition Eyes (Most common, seen in arthropods, annelids & horseshoe crab) 7. Refracting Superposition (Nocturnal insects) 8. Reflecting Superposition (Long bodied crustaceans like shrimp) 9. Parabolic Superposition (combines features of superposition and apposition eyes; functions by refracting light, then using a parabolic mirror to focus the image. Mayflies)
(A number of lenses above photoreceptors ) (Single-Chambered Eyes; One concave photoreceptive surface)
Presenter
Presentation Notes
There are many different eye layouts—indeed every technological method of capturing an optical image commonly used by human beings, with the exceptions of zoom and Fresnel lenses, occur in nature. Eye types can be categorized into "simple eyes", with one concave photoreceptive surface, and "compound eyes", which comprise a number of individual lenses laid out on a convex surface. Note that "simple" does not imply a reduced level of complexity or acuity. Indeed, any eye type can be adapted for almost any behavior or environment. The only limitations specific to eye types are that of resolution—the physics of compound eyes prevents them from achieving a resolution better than 1°. Also, superposition eyes can achieve greater sensitivity than apposition eyes, so are better suited to dark-dwelling creatures. Eyes also fall into two groups on the basis of their photoreceptor's cellular construction, with the photoreceptor cells either being cilliated (as in the vertebrates) or rhabdomeric. These two groups are not monophyletic; the cnidaria also possess cilliated cells, and some annelids possess both. Non-compound or Simple eyes: Simple eyes are rather ubiquitous, and lens-bearing eyes have evolved at least seven times in vertebrates, cephalopods, annelids, crustaceans and cubozoa. Pit eyes: Pit eyes, also known as stemma, are eye-spots which may be set into a pit to reduce the angles of light that enters and affects the eyespot, to allow the organism to deduce the angle of incoming light. Found in about 85% of phyla, these basic forms were probably the precursors to more advanced types of "simple eye". They are small, comprising up to about 100 cells covering about 100 µm. The directionality can be improved by reducing the size of the aperture, by incorporating a reflective layer behind the receptor cells, or by filling the pit with a refractile material. Pit vipers have developed pits that function as eyes by sensing thermal infra-red radiation, in addition to their optical wavelength eyes like those of other vertebrates. Spherical lensed eye: The resolution of pit eyes can be greatly improved by incorporating a material with a higher refractive index to form a lens, which may greatly reduce the blur radius encountered—hence increasing the resolution obtainable. The most basic form, seen in some gastropods and annelids, consists of a lens of one refractive index. A far sharper image can be obtained using materials with a high refractive index, decreasing to the edges; this decreases the focal length and thus allows a sharp image to form on the retina. This also allows a larger aperture for a given sharpness of image, allowing more light to enter the lens; and a flatter lens, reducing spherical aberration. Such an inhomogeneous lens is necessary in order for the focal length to drop from about 4 times the lens radius, to 2.5 radii. Heterogeneous eyes have evolved at least nine times: four or more times in gastropods, once in the copepods, once in the annelids, once in the cephalopods, and once in the chitons, which have aragonite lenses. No aquatic organisms possess homogeneous lenses; presumably the evolutionary pressure for a heterogeneous lens is great enough for this stage to be quickly "outgrown". This eye creates an image that is sharp enough that motion of the eye can cause significant blurring. To minimize the effect of eye motion while the animal moves, most such eyes have stabilizing eye muscles. The ocelli of insects bear a simple lens, but their focal point always lies behind the retina; consequently they can never form a sharp image. This capitulates the function of the eye. Ocelli (pit-type eyes of arthropods) blur the image across the whole retina, and are consequently excellent at responding to rapid changes in light intensity across the whole visual field; this fast response is further accelerated by the large nerve bundles which rush the information to the brain. Focusing the image would also cause the sun's image to be focused on a few receptors, with the possibility of damage under the intense light; shielding the receptors would block out some light and thus reduce their sensitivity. This fast response has led to suggestions that the ocelli of insects are used mainly in flight, because they can be used to detect sudden changes in which way is up (because light, especially UV light which is absorbed by vegetation, usually comes from above). Multiple lenses: Some marine organisms bear more than one lens; for instance the copepod Pontella has three. The outer has a parabolic surface, countering the effects of spherical aberration while allowing a sharp image to be formed. Another copepod, Copilia, has two lenses in each eye, arranged like those in a telescope. Such arrangements are rare and poorly understood, but represent an interesting alternative construction. An interesting use of multiple lenses is seen in some hunters such as eagles and jumping spiders, which have a refractive cornea: these have a negative lens, enlarging the observed image by up to 50% over the receptor cells, thus increasing their optical resolution. Refractive cornea: In the eyes of most mammals, birds, reptiles, and most other terrestrial vertebrates (along with spiders and some insect larvae) the vitreous fluid has a higher refractive index than the air. In general, the lens is not spherical. Spherical lenses produce spherical aberration. In refractive corneas, the lens tissue is corrected with inhomogeneous lens material (see Luneburg lens), or with an aspheric shape. Flattening the lens has a disadvantage; the quality of vision is diminished away from the main line of focus. Thus, animals that have evolved with a wide field-of-view often have eyes that make use of an inhomogeneous lens. As mentioned above, a refractive cornea is only useful out of water; in water, there is little difference in refractive index between the vitreous fluid and the surrounding water. Hence creatures that have returned to the water – penguins and seals, for example – lose their highly curved cornea and return to lens-based vision. An alternative solution, borne by some divers, is to have a very strongly focusing cornea. Reflector eyes: An alternative to a lens is to line the inside of the eye with "mirrors", and reflect the image to focus at a central point. The nature of these eyes means that if one were to peer into the pupil of an eye, one would see the same image that the organism would see, reflected back out. Many small organisms such as rotifers, copepods and platyhelminths use such organs, but these are too small to produce usable images.[1] Some larger organisms, such as scallops, also use reflector eyes. The scallop Pecten has up to 100 millimetre-scale reflector eyes fringing the edge of its shell. It detects moving objects as they pass successive lenses.[1] There is at least one vertebrate, the spookfish, whose eyes include reflective optics for focusing of light. Each of the two eyes of a spookfish collects light from both above and below; the light coming from above is focused by a lens, while that coming from below, by a curved mirror composed of many layers of small reflective plates made of guanine crystals. Compound eyes: An image of a house fly compound eye surface by using scanning electron microscope. Arthropods such as Calliphora vomitoria fly have compound eyes. A compound eye may consist of thousands of individual photoreceptor units or ommatidia (ommatidium, singular). The image perceived is a combination of inputs from the numerous ommatidia (individual "eye units"), which are located on a convex surface, thus pointing in slightly different directions. Compared with simple eyes, compound eyes possess a very large view angle, and can detect fast movement and, in some cases, the polarisation of light. (Even the trained human eye can determine the orientation of polarized light which manifests in a phenomenon called Haidinger's brush.) Because the individual lenses are so small, the effects of diffraction impose a limit on the possible resolution that can be obtained (assuming that they do not function as phased arrays). This can only be countered by increasing lens size and number. To see with a resolution comparable to our simple eyes, humans would require very large compound eyes, around 11 m in radius. Compound eyes fall into two groups: apposition eyes, which form multiple inverted images, and superposition eyes, which form a single erect image. Compound eyes are common in arthropods, and are also present in annelids and some bivalved molluscs. Compound eyes, in arthropods at least, grow at their margins by the addition of new ommatidia. Apposition eyes: Apposition eyes are the most common form of eyes, and are presumably the ancestral form of compound eyes. They are found in all arthropod groups, although they may have evolved more than once within this phylum. Some annelids and bivalves also have apposition eyes. They are also possessed by Limulus, the horseshoe crab, and there are suggestions that other chelicerates developed their simple eyes by reduction from a compound starting point. (Some caterpillars appear to have evolved compound eyes from simple eyes in the opposite fashion.) Apposition eyes work by gathering a number of images, one from each eye, and combining them in the brain, with each eye typically contributing a single point of information. The typical apposition eye has a lens focusing light from one direction on the rhabdom, while light from other directions is absorbed by the dark wall of the ommatidium. In the other kind of apposition eye, found in the Strepsiptera, lenses are not fused to one another, and each forms an entire image; these images are combined in the brain. This is called the schizochroal compound eye or the neural superposition eye. Because images are combined additively, this arrangement allows vision under lower light levels. Superposition eyes: The second type is named the superposition eye. The superposition eye is divided into three types; the refracting, the reflecting and the parabolic superposition eye. The refracting superposition eye has a gap between the lens and the rhabdom, and no side wall. Each lens takes light at an angle to its axis and reflects it to the same angle on the other side. The result is an image at half the radius of the eye, which is where the tips of the rhabdoms are. This type of compound eye is normally found in nocturnal insects because it can create images up to 1000 times brighter than equivalent apposition eyes, though at the cost of reduced resolution. In the parabolic superposition compound eye type, seen in arthropods such as mayflies, the parabolic surfaces of the inside of each facet focus light from a reflector to a sensor array. Long-bodied decapod crustaceans such as shrimp, prawns, crayfish and lobsters are alone in having reflecting superposition eyes, which also have a transparent gap but use corner mirrors instead of lenses. Parabolic superposition: This eye type functions by refracting light, then using a parabolic mirror to focus the image; it combines features of superposition and apposition eyes.
Different types of eyes in multicellular organisms
Pit Eye
Compound Eye
Aquatic Lens Eye
Apposition Compound Eye
Corneal Lens
Refracting Superposition
Eye
Reflecting Superposition
Eye
Single chambered mirror eye
Presenter
Presentation Notes
Major optical types of eyes found in multicellular organisms, forming images using shadow (a,b), refracting devices (c-f), and reflectors/mirrors (g,h). The photoreceptors are shown stippled. A, Pit eye, forerunner of other single chambered eyes shown in c, d and g. Found in planarians, annelids, molluscs and in Nautilus. B, Basic compound eye, in which each receptor is shielded from its neighbor by a simple pigment tube. Found in bivalve molluscs C, Aquatic lens eye ,found in fish and cephalopod molluscs D, Corneal lens eye found in terrestrial vertebrates, some spiders and insect larvae E, Apposition compound eye, found in diurnal insects and crustaceans. Each receptor cluster has its own lens. F, Refracting superposition compound eye found in animals from dim environments (e.g., moths and krill). Here, many lenses contribute to the image at each point on the retina. G, Single chambered eye where the image is produced by a concave mirror. Found in scallops H, Reflecting superposition eye. Similar to f, except that the lenses are replaced by mirrors. Found in decapod shrimps and lobsters.
• Clusters of photoreceptor proteins. Earliest predecessors of the eye.
• Can only sense ambient brightness, sufficient for synchronization of circadian rhythms.
• Insufficient for vision, as they cannot distinguish shapes or the direction of light.
• Found in nearly all animal groups, and are common among unicellular organisms including euglena.
Eyespots in unicellular organisms: Early predecessors of the eye
Euglena Eyespot
“Eyespots“ comprised of photoreceptor cell(s) surrounded by pigmented cells have evolved independently about 40-65 times.
Eyespots provide a basic sense of the direction and intensity of light
Eyespots in a planarian
Eyespots in multicellular organisms
Eyespots in a limpet
Pinhole camera eye in a Nautilus
Pinhole camera eyes
Presenter
Presentation Notes
The "pinhole camera eye" developed as the eye pit deepened into a cup, then a chamber. By reducing the size of the opening, the organism achieved true imaging, allowing for fine directional sensing and even some shape-sensing. Eyes of this nature are currently found in the nautilus. Lacking a cornea or lens, they provide poor resolution and dim imaging, but are still, for the purpose of vision, a major improvement over the early eyepatches.
Compound eyes in arthropods
Presenter
Presentation Notes
The compound eyes of arthropods like insects, crustaceans and millipedes[1] are composed of units called ommatidia (singular: ommatidium). An ommatidium contains a cluster of photoreceptor cells surrounded by support cells and pigment cells. The outer part of the ommatidium is overlaid with a transparent cornea. Each ommatidium is innervated by one axon and thus provides the brain with one picture element. The brain forms an image from these independent picture elements. The number of ommatidia in the eye depends upon the type of insect and ranges from just a handful in the primitive Archaeognatha and Thysanura to around 30 thousand in largerAnisoptera dragonflies and in some Sphingidae moths. The cornea and pseudocone form the outer ten percent of the length of the ommatidium. The inner 90% of the ommatidium contains 6 to 9 (depending on the species) long and thin photoreceptor cells in the case of some butterflies often abbreviated "R cells" in literature and often numbered, e.g. R1 through R9.These "R cells" tightly pack the ommatidium. The portion of the R cells at the central axis of the ommatidium collectively form a light guide, a transparent tube, called the rhabdom. In certain flies, the rhabdom has separated into seven independent rhabdomeres. This has required the rewiring of the eye such that each ommatidium now has seven axons leading from it. The advantage to this arrangement is that it increases the number of picture elements by a factor of seven, without increasing the number of ommatidia. Since an image from the compound eye is created from the independent picture elements produced by ommatidia, it is important for the ommatidia to react only to that part of the scene directly in front of it. To prevent light entering at an angle from being detected by the ommatidium it entered, or by any of the neighboring ommatidia, six pigment cells are present. The pigment cells line the outside of each ommatidium. Each pigment cell is situated at the apex of the hexagons and thus lines the outside of three ommatidia. Light entering at an angle passes through the thin cross-section of the photoreceptor cell, with only a tiny chance of exciting it, and is absorbed by the pigment cell, before it can enter a neighboring ommatidium. In many species, in low-light situations, the pigment is withdrawn, so that light entering the eye might be detected by any of several ommatidia. This enhances light detection but lowers resolution. The size of the ommatidia varies according to species, but ranges from 5 to 50 micrometres. The rhabdoms within them may cross-section at least as small as 1.x micrometres, the category of "small" being assigned in some cross-species studies to those under 2 micrometers. Naively, microlens arrays can be seen as a biomimetic analogy of ommatidia. Structure of the Drosophila eye. (A)Scanning electron microscope image of a Drosophila melanogaster head. The compound eye (pseudocolored in red) is composed of ~800 unit eyes, the ommatidia, each covered by its own lens (visible on the surface of the eye) that focuses light on the photoreceptors below. The third segments of the antennae (a) and the maxillary palps (m) are covered with sensilla, which are hair-like structures that house the olfactory receptor neurons. (B)Schematic side view of an ommatidium (unit eye). Under the lens (top) reside eight photoreceptor neurons (PRs R1-R8; only four are shown, colored). Each PR has a rod-like, subcellular specialization, the rhabdomere (darker colors), loaded with Rhodopsin proteins, which detect photons. Rhabdomeres of ‘outer’ PRs (green, only two of six are shown) span the entire length of the ommatidium. The rhabdomeres of the ‘inner’ PRs are located above each other, with R7 (violet) distal to R8 (red), such that they share a common light path and face the same point in space. (C)Schematic of a section through an ommatidium at the level indicated by the dashed line in B. Rhabdomeres (colored circles) belonging to the ‘outer’ PRs (R1-R6, green) form a trapezoid configuration, whereas the rhabdomere of the ‘inner’ PRs (R8 in this section, in red) is in the center of the trapezoid. (D)Image corresponding to the schematic in C showing expression of Rh1 (green) in R1-R6 PRs and Rh6 (red) in R8. (E)A longitudinal view of rhabdomeres within a single ommatidium. The ‘inner’ rhabdomeres contain Rh3 (blue) and Rh5 (turquoise) belonging to R7 and R8 PRs, respectively. The ‘outer’ rhabdomeres (brown) were visualized using an actin counterstain. The image shown in A was obtained with permission from Clouds Hill Imaging. Schematics shown in B and C were adapted with permission (Cagan and Ready, 1989).
Horseshoe crabs have two primary compound eyes and seven secondary simple eyes. Two of the secondary eyes are on the underside
Median Simple Eye
Lateral Compound Eye Lateral
Simple Eye
Horseshoe Crabs: Both simple and compound eyes
Presenter
Presentation Notes
The sizes of compound eyes with human like resolution
Presenter
Presentation Notes
The sizes of compound eyes with human like resolution. Left: A compound eye with 1 minute of resolution everywhere. Right: Compound eye with 1 minute resolution in the fovea, but falling off with eccentricity as in the human eye. (Kirschfeld, 1976).
Wolf spider has eight simple eyes, two main eyes at the front and six smaller secondary eyes. The main eyes form images. The secondary eyes detect peripheral movement
Simple corneal eyes in arachnids
Simple (single-chambered) lens eye in vertebrates
Lens eyes: Also seen in cephalopods, annelids, jellies, copepods…
Swamynathan et al 2003. FASEB J. 17: 1996-2005
Multiple eye types in each branch of evolutionary tree Single Chambered
Compound Eyes
All Eyes Sense Light
Presenter
Presentation Notes
Each branch of the evolutionary tree includes multiple eye types, and their distribution suggests that each type must have evolved several times independently. Single-chambered eyes are outlined in rectangles and compound eyes in ovals. The color of the outline is red for eyes that form images using only shadow, blue for eyes that use refracting devices such as lenses or corneas, and green for eyes that use mirrors.
Landscape of eye evolution O
ptic
al Q
ualit
y
Evolutionary Distance
“Climbing the hills is straightforward but going from one hill top to another is near impossible.” -Mike Land
Presenter
Presentation Notes
A possible landscape of eye evolution. “Climbing the hills is straightforward but going from one hill top to another is near impossible.”
Key innovations in eye evolution
Screening Pigment
Membrane Stacking
Focusing Optics
Photopigment Regeneration
Presenter
Presentation Notes
Key innovations in eye evolution. Directional photoreception is assumed to have evolved from nondirectional monitoring of ambient luminance by a cell duplication event and an opsin gene duplication leading to a receptor opsin and photoisomerase pair of proteins for efficient chromophore regeneration. This was followed by the introduction of screening pigment and soon also by membrane stacking to allow for better contrast discrimination, increased speed and more directional photoreception. Multiple receptor cells would then allow for true spatial vision and the scanning mode of operation could be abandoned. The single-chambered and compound eyes would have to evolve independently from directional ocelli. To collect enough photons for spatial vision with higher resolution, lenses must be introduced, but the new high-resolution tasks are expected to add to rather than replace the older low-resolution tasks.
Opsins: Key Molecules for Vision
First critical part of the puzzle in building an eye : Photosensitive molecule that can be regenerated
Presenter
Presentation Notes
Opsins lie embedded in the outer membrane of the cell, where retinal (grey molecule in the middle) can trap photons. Opsins trap photons with a small molecule in the heart of their architecture, called retinal. In its resting state retinal has a bent and twisted tail. But as soon as light strikes retinal, its tail unbends. This molecular stretching exercise forces the opsin to change shape as well. The opsin is now activated and eventually will cause a nearby nerve to fire, which will relay its message to the brain. The visual cycle is the biological conversion of a photon into an electrical signal in the retina. This process occurs via G-protein coupled receptors called opsins which contain the chromophore 11-cis retinal. 11-cis retinal is covalently linked to the opsin receptor via Schiff base forming retinylidene protein. When struck by a photon, 11-cis retinal undergoes photoisomerization to all-trans retinal which changes the conformation of the opsin GPCR leading to signal transduction cascades which causes closure of cyclic GMP-gated cation channel, and hyperpolarization of the photoreceptor cell. Following isomerization and release from the opsin protein, all-trans retinal is reduced to all-trans retinol and travels back to the retinal pigment epithelium to be "recharged". It is first esterified by lecithin retinol acyltransferase (LRAT) and then converted to 11-cis retinol by the isomerohydrolase RPE65. The isomerase activity of RPE65 has been shown; it is still uncertain whether it also acts as hydrolase. Finally, it is oxidized to 11-cis retinal before traveling back to the rod outer segment where it is again conjugated to an opsin to form new, functional visual pigment (rhodopsin).
Phylogeny of Opsins
Presenter
Presentation Notes
Origin of opsins, and their possible association with membrane type. Hypothesized duplications of ancestral opsin and its precursors, and suggested association with membrane type. An ancient GPCR (related to extant vertebrate melatonin receptors) duplicated, and its ligand became retinaldehyde, which bound non-covalently; this is denoted as ‘Retinaldehyde receptor’. After the divergence of the amoeba-like placozoans (w711 Mya), this GPCR evolved a lysine residue in its seventh transmembrane segment and a negatively charged residue (counterion) so that retinaldehyde bound covalently via a protonated Schiff base linkage; this form is denoted ‘Ancestral opsin’. Within a relatively short interval (prior to the divergence of cnidarians, w700 Mya), this opsin duplicated twice, giving rise to three major families of opsins: C-opsins, R-opsins, and RGR/Go-opsins. It is proposed that these three opsins preferentially associated with ciliary membrane, microvillar membrane, and the membranes of intracellular organelles, respectively.
Phylogeny of Opsins
Presenter
Presentation Notes
Light-sensing opsins belong to a family of proteins (G-protein coupled receptors, or GPCRs, a.k.a., "serpentine" proteins because they snake in and out of plasma membranes) found in all eukaryotes, and are involved in sensing external stimuli (such as chemical signals). In the earliest ancestor of animals, a GPCR apparently mutated and became able to sense a different type of stimulus: LIGHT. This early protein eventually developed into an opsin. opsins further differentiated in animals into different types of opsins sensing light in different tissues. They may only later have been recruited to sense light in actual light-sensing organs (eyes). All animal eyes share a homologous type of visual pigment, but the various specializations for accommodating large amounts of visual pigment and for transducing the information into electric signals appear not to be homologous. Instead, they are parallel solutions to a common problem of sensitivity and speed of visual receptor cells. The different types of receptor cells have subsequently been recruited independently in a number of instances to produce imaging eyes. There are striking similarities between independently evolved eyes in the chain of control genes that orchestrate eye development. This indicates that the control genes are ancient and were originally associated with opsin transcription and sensory or neural fate in a common ancestor. The levels of homology tells an interesting story about animal evolution. The common ancestor must have been a simple organism which used light intensity to control its behavior. Elaborations to obtain some directionality, high sensitivity and response speed may have independently resulted in different receptor types and visual organs. This evolutionary process must have been mostly finished in the Precambrian, and the results were used by animals which had more complex responses to light. The evolution of macroscopic and swiftly mobile animals in the early Cambrian led to a strong selection for improved spatial vision which produced the first proper eyes with imaging optics. This was a parallel process in several animal phyla and the superficial similarities between separate evolutionary lines is due to design constraints. Eyes of different phyla can thus be said to be both homologous and non-homologous, depending on the level of organization considered.
Second part of the puzzle: Spatiotemporal regulation of opsin gene expression
Drosophila Twin of Eyeless Drosophila Pax6 in Xenopus misexpression in legs
Presenter
Presentation Notes
(A and B) Misexpression of Pax6 family members is sufficient to induce ectopic eye structures in flies and frogs. (A) Drosophila: ectopic eyes on the legs of an adult (arrowheads) induced by the targeted misexpression of the fly Pax6 homolog, twin of eyeless (from Gehring, 2005). (B) Xenopus: ectopic eye in an embryo injected with Pax6 RNA. Side view of a stage 48 embryo. Arrow denotes endogenous eye. Arrowheads mark the ectopic eye cup (white arrowhead) and lens (black arrowhead) (Chow et al., 1999).
Genetic pathway that specifies eyefield is conserved in arthropods and vertebrates
Presenter
Presentation Notes
Conservation in the genetic pathway that regulates eye formation in flies and vertebrates. (C) Simplified version of the gene regulatory network controlling eye specification in Drosophila. For a more detailed summary see Kumar (2008). The nuclear factors depicted in this circuit are both necessary and sufficient for eye determination in the fly. The homologs of genes highlighted in red are also required for eye development in vertebrates. Note that the Rx gene, which sits atop the genetic cascade for eye specification in vertebrates, is not required for fly eye development. toy, twin of eyeless; ey, eyeless; so, sine oculis; eya, eyes absent; dac, dachshund; eyg, eyegone.
Diverse roles of Pax transcription factors during eye evolution
Presenter
Presentation Notes
Diverse roles of Pax transcription factors during eye evolution. All eyes require an assembly of photoreceptor cells (grey) and shielding pigment cells (black). An almost universal use of Pax genes in eye formation throughout the animal kingdom argues for their ancient role in generation of a prototypical eye as a photoreceptor/pigment cell combination (top). The bipartite ‘Paxcentric model’ [35] proposed independent functions of paired domain and homeodomain of Pax transcription factors in regulating either photoreceptor or pigment cell programs. Lens-containing eyes have evolved independently multiple times. Pax transcription factors are likely involved in the divergent morphogenesis of various types of lens-containing eyes. Convergent evolution of Pax regulatory elements was a driving force behind the recruitment of lens crystallin genes and eye ‘morphogenesis’ genes. ‘Morphogenesis’ genes are the ones required for eye development in any given animal. Note that opsin genes are homologous across phylogeny, whereas lens crystallins are encoded by structurally unrelated genes in different species
Evolution of eyes reflects a central idea in evolutionary theory-the diversity and unity of life
Evolution of Pax6-related genes Evolution of eye structures
Presenter
Presentation Notes
Each organism has a unique evolutionary history that sets it apart from other lineages — but it also shares a large portion of that history with other organisms. The original Pax gene — the ancestral version of eyeless, mouse Pax6, and the eye-building genes of different animal lineages— probably evolved more than 500 million years ago. As its descendants evolved, the basic eye-building gene was modified in different ways in the different lineages, giving rise to the diversity of Pax6-like genes seen in modern animals.
Analogous eyes; Homologous genes
Presenter
Presentation Notes
It's a bit like taking several similar on/off switches and using one to turn on the bathroom light, one to turn on the garbage disposal— the same switch can be co-opted for slightly different jobs, all requiring a switch. 1. More than 500 million years ago, the ancestor of most modern animals evolved the "switch" — the ancestral version of modern Pax6 and its cousins. In that organism, the gene may have helped build the simplest type of eye (one photoreceptor cell and one pigment cell) or helped build a light sensitive protein. 2. That ancestor then passed the basic gene to all of its descendants. 3. Some of its descendants began living in an environment in which more sensitive light perception was advantageous. In those organisms, natural selection favored any individuals that happened to use the gene to build more complex and sensitive eyes. Over time, these organisms evolved one type of complex eye which was switched on by a descendant of the original gene. Meanwhile, other animals living in a different environment also found fine-tuned light perception to be advantageous. Over time, these organisms evolved their own type of complex eye which was also switched on by a different descendant of the original gene.
One origin or many?
• Much of the genetic machinery employed in eye development is common to all eyed organisms, which suggest one origin from a common ancestor that utilized some form of light-sensitive machinery – even if it lacked a dedicated optical organ. Shared traits common to all light-sensitive organs include the opsins family of photo-receptive proteins.
• However, even photoreceptor cells may have evolved more than once from molecularly
similar chemoreceptors, and photosensitive cells probably existed long before the Cambrian explosion. Higher-level similarities – such as the use of crystallins in the independently derived cephalopod and vertebrate lenses – reflect the co-option of a protein from a more fundamental role to a new function within the eye
Lens-containing eyes evolved sporadically throughout the animal kingdom
Presenter
Presentation Notes
Re-occurrence of lens-containing eyes throughout the animal kingdom. A diagram of main animal phyla indicating the presence of lens-containing eyes or eyes without a lens (in the adult or in the larval stage) within each phylum. An eye is defined here as an organ of spatial vision with a minimum requirement of a single pigment cell and at least one photoreceptor cell
Different proteins serve as crystallins in diverse phyla
Presenter
Presentation Notes
A schematic diagram illustrating the eye morphology, crystallin variability and refractive index (n) in the center of the lens of selected model animals
Distribution of photoreceptor cell types and screening pigments in different phyla
Rhabdomeric Ancestral Ciliary (Microvilli-based)
Presenter
Presentation Notes
Basic Types of Photoreceptors. Rhabdomeric and ciliary photoreceptors and their homologies. Conservation of cell polarity and topology between Drosophila rhabdomeric photoreceptor (left) and vertebrate rod photoreceptor (right). The centre image pair, in top and side view, depicts an ancestral (or an immature) photoreceptor, which can evolve (or develop) into either of the two illustrated final forms simply by different modes of expansion of the apical membrane (pink); that apical membrane is separated from the baso-lateral membrane (yellow) by a zonula adherens (ZA, brown). r, rhabdomere; s, fly stalk; os, outer segment; cc, connecting cilium; is, inner segment; ELM, external limiting membrane; N, nucleus. Distribution of screening pigments: A schematic diagram of distribution of screening pigments in different animal phyla. The photoreceptor cell type screened by particular pigments is depicted on the left side of the box of each phylum. There is no apparent correlation between the photoreceptor cell type and shielding pigment used. Note the unknown type of photoreceptor and unique deployment of haemoglobin as a shielding pigment of nematode Mermis, caused probably by independent origin of the eye in this organism. See electronic supplementary material for further details and references.
Polyphyletic, Monophyletic, Convergent Divergent
• Independent evolution of similar features in species of different lineages
• Creates analogous structures with similar form or function, that were not present in their last common ancestor
Vertebrates, cephalopods, and cnidaria possess camera eyes, while their last common ancestor had a simple photoreceptive spot. Progressive refinement of this structure led to the advanced camera eye. The similarity of these structures, despite their complexity, illustrates how some biological challenges have an optimal solution, and suggests polyphyletic, convergent evolution of eyes. However, conservation of opsins and the transcription factors regulating their expression suggests monophyletic evolution of eyes.
• Accumulation of differences between groups leading to new speciation.
• Similarity is due to the common origin, such as divergence from a common ancestral structure or function
Evolution of Eyes: Convergent, or Divergent?
Presenter
Presentation Notes
Vertebrates, cephalopods, and cnidaria possess camera eyes, while their last common ancestor had a simple photoreceptive spot. Progressive refinement of this structure led to the advanced camera eye. The similarity of these structures, despite their complexity, illustrates how some biological challenges have an optimal solution, and suggests polyphyletic, convergent evolution of eyes. However, conservation of opsins and the transcription factors regulating their expression suggests monophyletic evolution of eyes.
Convergent evolution of analogous structures
Presenter
Presentation Notes
The phylogeny is based on anatomical and genetic studies and shows the evolutionary relationships among animals with eyes. Notice how closely related sharks and mice are! And even though sharks and squid both have eyes adapted for seeing in the ocean, they are not very closely related, and their eyes are analogous. Along the same lines, squid eyes and mouse eyes look similar and have similar layouts, but those similarities are analogies that evolved through convergent evolution. Although the camera eye of the octopus is very similar to that of humans, phylogenetic and embryological analyses have suggested that their camera eyes have been acquired independently. A larger number of conserved genes and their similar gene expression may be responsible for the convergent evolution of the camera eye
Different solutions for one problem suggest multiple origins
Vertebrate Eye Octopus Eye Built Upside down Built right way out
Photoreceptors Transducing nerves
Transducing nerves Photoreceptors
Optic nerve Optic nerve
Presenter
Presentation Notes
The vertebrate eye is built "backwards and upside down", requiring "photons of light to travel through the cornea, lens, aqueous fluid, blood vessels, ganglion cells, amacrine cells, horizontal cells, and bipolar cells before they reach the light-sensitive ciliary photoreceptors that transduce the light signal into neural impulses, which are then sent to the visual cortex at the back of the brain for processing into meaningful patterns." The camera eyes of cephalopods, in contrast, are constructed the "right way out", with the nerves attached to the rear of the microvillar photoreceptors in the retina. This means that they do not have a blind spot. This difference may be accounted for by the developmental origins of eyes; in cephalopods they develop as an invagination of the head surface ectoderm, whereas in vertebrates they originate as an extension of the brain. The camera-type eye of cephalopods (octopus, squid, cuttle-fish etc.) has previously been considered to represent a paradigm for convergent evolution. However, this notion has to be modified in the light of the discovery of a highly conserved Pax6 gene in squids which shares 67% amino acid sequence identity with vertebrates. In the PD, squid Pax6 shows 91–95% sequence identity with its homologs in vertebrates, Drosophila, Nemertine, and sea urchins, whereas the HD shows 90–98% identity with Pax6 from these species. Ectopic expression of squid Pax6 cDNA in Drosophila induces the formation of ectopic eyes. These findings indicate that the cephalopod eye like all other bilaterian eyes is specified by the Pax6 gene and has originated monophyletically and subsequently evolved by divergent, parallel, and convergent evolution. Comparative genomics between the camera-type eyes of octopuses and humans show that of 1052 nonredundant genes which are expressed in the octopus eye 729 (=69%) are in common with the human eye, which is a surprisingly large fraction. Of these 1052 genes, 1019 are already found in the last common ancestor of bilateria, and 875 out of 1052 are conserved between humans and octopus. By comparing three species of cephalopods with cameratype eyes with molluscs with pin-hole eyes (Nautilus) and mirror-eyes (scallops); 5707 cephalopod camera eye-specific candidate genes (5707) were selected for further analysis. From the 1571 genes found in common between cephalopod and vertebrate cameratype eyes, 156 genes were identified which were under positive selection. These studies suggest that both changes in gene expression and in the primary structure of the respective proteins (through positive selection) have contributed to the evolution of the cephalopod camera-type eye
General scheme of eye evolution
Presenter
Presentation Notes
General scheme of eye evolution. The first step in eye evolution is the evolution of a light receptor molecule which in all metazoa is rhodopsin. In the most ancestral metazoa, the sponges, a single Pax gene, but no opsin gene has been found. In the larva of the box jellyfish Tripedalia, a unicellular photoreceptor has been described. The adult jellyfish forms complex lens eye with ciliary photoreceptor cells, which form under control of PaxB, a putative precursor of Pax6. However, the eyes of the hydrozoan jellyfish Cladonema are controlled by PaxA. Walter Gehring proposes that the prototypic eye consisting of just two cells a photoreceptor cell and a pigment cell originated from a unicellular photoreceptor by a first step of cell differentiation. This cellular differentiation led to formation of a photoreceptor cell and a pigment cell under the genetic control by Pax6 and Mitf, respectively. As true innovations are rare in evolution all the more complex eye-types arose monophyletically from one of these Darwinian prototypes leading to a large diversity of eye-types by divergence, parallel evolution, and convergence.
Modern eyes evolved by division of labor
• The number of cell types in an eye has gone up concomitantly with the increasing complexity of evolving eye types
• The number of protein-coding genes representing cellular eye-related functions has not increased at a similar pace.
Division of labor, or functional segregation of cell types resulting in differential distribution of cellular functions between sister cell types
Pace of eye evolution
Presenter
Presentation Notes
A patch of light sensitive epithelium can be gradually turned into a perfectly focused camera-type eye if there is a continuous selection for improved spatial vision. A theoretical model based on conservative assumptions about selection pressure and the amount of variation in natural populations suggest that the whole sequence can be accomplished amazingly fast, in less than 400,000 generations. The number of generations is also given between each of the consecutive intermediates that are drawn in the figure. The starting point is a flat piece of epithelium with an outer protective layer, an intermediate layer of receptor cells, and a bottom layer of pigment cells. The first half of the sequence is the formation of a pigment up eye. When this principle cannot be improved any further, a lens gradually evolves.
• The eye, defined as an organ of spatial vision, is present in a fraction of the animal phyla • Most of the eyes we see today arose during Cambrian Explosion about 530 Mya • Despite their separate evolutionary origins, even analogous eyes (like those of sharks and
squid) have basic similarities. All eyes, wherever they evolved on the tree of life, sense light and are used by organisms to interact with their environments. Many analogous eyes share similar cell types — and those cells contain similar light-sensing molecules
• The photopigment rhodopsin and certain genes controlling eye development appear to have been present in a common metazoan ancestor, as they are homologous in all eyes
• Though structural evidence suggests that the eyes evolved independently many times, the small number of photoreceptor cell types suggests that they predate the eyes themselves
• Conservation of the genetic network of transcription factors required to specify eyefield suggests monophyletic origin of the eyes followed by a series of specializations
• It is estimated that the evolution of an advanced fish eye from a patch of photosensitive tissue could occur in less than 400,000 generations, providing an explanation for the large diversity of eye types in animals
• Cell-type functional segregation, the differential distribution of cellular functions between sister cell types may have played a major role in eye evolution