Embed Size (px)
Citation preview

JOURNAL OP FERMENTATION AND BIOENGINEERING Vol. 83, No. 4, 321-327. 1997
Fatty Acid Specificity of Rhizopus delemar Lipase in Acidolysis YUJI SHIMADA,‘* AK10 SUGIHARA,’ HIROFUMI NAKANO,’ TOSHIHIRO NAGAO,’
MASAHARU SUENAGA,2 SEIICHI NAKAI,2 AND YOSHIO TOMINAGA’ Osaka Municipal Technical Research Institute, I-6-50 Morinomiya, Joto-ku, Osaka 536 and Yashiro Co. Ltd., l-5-9
Kitahorie, N&hi-ku, Osaka 550,2 Japan
Received 5 November 1996IAccepted 24 January 1997
We investigated the fatty acid specificity of 1,lpositional specific Rhizopus deZemar lipase in acidolysis using a randomly interesterified oil, of which the constituent fatty acid contents were approximately the same, as a substrate. Acidolysis was carried out at 3O“C with shaking in a reaction mixture containing caprylic acid (CA)/the randomly interesterilied oil (2 : 1, w/w) and the lipase immobilized on a ceramic carrier (4% of reaction mixture). The activity on each fatty acid was experimentally determined as the ratio of the content of the fatty acid exchanged for CA to that of the fatty acid existing at the l(3)-position of the substrate oil, and regarded as the fatty acid specificity in acidolysis. The fatty acid specif~ity in hydrolysis was investigated by analysis of the composition of fatty acids liberated from the same randomly interesteri5ed oil. The lipase acted strongly on myristic, pahnitic, palmitoleic, steak, oleic, linoleic, and a-linolenic acids, and moderately on arachidonic and eicosapentaenoic acids. The activities on these fatty acids relative to that on oleic acid were almost the same in acidolysis and hydrolysis. On the other hand, the lipase acted moderately on r-linolenic and docosahexaenoic acids in the hydrolysis, but only very weakly on these fatty acids in the early stage of acidolysis. This shows that the fatty acid specificity in acidolysis is stricter than that in hydrolysis. Since the kinetics of acidolysis of borage oil, arachidonic acid-containing oil (TGA-25), and tuna oil could be easily predicted from the fatty acid specificity in acidolysis, it was concluded that the speciiicity was accurately evaluated. In addition, the fatty acids at the l(3)-position of borage oil and TGA-25 were completely exchanged for CA in a total of three cycles of the reaction, and the contents of fatty acids other than CA in the transesteri5ed oils agreed with that at the 2-position of the oils. Therefore, it was shown that repeated acidolysis using the immobilized 1,3- specific lipase could be applied to regiospecific analysis.
[Key words: lipase, fatty acid specificity, acidolysis, Rhizopus delemar, immobilized enzyme, structured lipid, y-linolenic acid, arachidonic acid]
The physiological functions of polyunsaturated fatty acids (PUFA) have drawn attention in the last decade (l-3), and methods of processing PUFA-containing oils are desired. However, PUFA are very unstable against heat and oxidation. Thus lipase-catalyzed reactions are considered promising for use in such methods because they proceed efficiently at ordinary temperature and pres- sure and under a nitrogen stream.
It is well known that lipases catalyze hydrolysis, es- terification and transesterification (4). Generally, efficient high-level processing of oils and fats can be achieved by use of one of these reactions and maximal advantage taken of the enzyme characteristics (especially fatty acid and positional specificities). For example, PUFA-rich oil can be produced by hydrolysis with a lipase that acts only weakly on PUFA (named selective hydrolysis) (S- 8), and y-linolenic acid (18 : 3n -6, GLA) and docosahex- aenoic acid (22 : 6n - 3, DHA) were purified as free fatty acids by esterification with a lipase which acts only very weakly on these fatty acids (selective esterification) (9, 10). In addition, cocoa-like fat (1,3-stearoyl-2-oleoyl-gly- cerol and 1,3-behenoyl-2-oleoyl-glycerol) (11, 12) and high- absorptive structured lipids (13-15) can be produced using an immobilized 1,3-positional specific lipase. For
* Corresponding author. Abbreviations: AA, arachidonic acid; ALA, a-linolenic acid; CA,
caprylic acid; DHA, docosahexaenoic acid; EPA, eicosapentaenoic acid; GLA, y-linolenic acid; LA, linoleic acid; MA, myristic acid; OA, oleic acid; PA, palmitic acid; POA, palmitoleic acid; PUFA, polyun- saturated fatty acids; SA, steak acid.
the construction of these reaction systems and for pre- diction of the reaction kinetics, the fatty acid specificity of the respective lipase must be evaluated accurately.
The fatty acid specificities of lipases have been investi- gated mainly by measurement of rates of hydrolysis of simple triglycerides or methyl (ethyl) esters (16), or by analysis of the composition of fatty acids liberated from a natural oil used as a substrate (17). The former me- thod is not precise because of the differences between the physical states of the substrates, liquid and solid, at the reaction temperature. While the latter method is free of this problem, the substrate specificities of 1,3-position- al specific lipases cannot be determined accurately be- cause fatty acids are not homogeneously distributed in triglyceride molecules in natural oils. Therefore, we devel- oped a new method of investigating fatty acid specificity in hydrolysis with a randomly interesterified oil as the substrate (18). The specificity can be evaluated by analy- sis of the composition of fatty acids liberated from the oil in the early stage of the reaction, and the results are useful for screening for a lipase suitable for enrichment of a desired PUFA by selective hydrolysis, and for prediction of the reaction kinetics (18, 19).
It was recently suggested that the fatty acid specifici- ties of a lipase in hydrolysis, esterification and trans- esterification are slightly different (10, 14, 20). However, the fatty acid specificity in esterification and transesterifi- cation could not be precisely investigated because the hy- drolysis occurred even in a reaction mixture containing only a trace amount of water. We developed a method of producing structured lipids containing DHA, linoleic acid
321

322 SHIMADA ET AL. J. FERMENT. BIOENG.,
(18 : 2n-6, LA) and a-linolenic acid (18 : 3n-3, ALA) from tuna, safflower and linseed oils, respectively, using the im- mobilized Rhizopus delemar lipase (14, 15). In these reac- tions, hydrolysis was completely repressed and only trans- esterification (acidolysis) proceeded efficiently. In this paper, we describe a new method for investigating fatty acid specificity in acidolysis, and show that the results are useful for the prediction of the actual reaction kinetics.
MATERIALS AND METHODS
Lipases R. delemar lipase (Ta-lipase, 120,000 U/g), and Candida rugosa lipase (Lipase-OF, 36O,OOOU/g) were gifts from Tanabe Seiyaku Co. Ltd. (Osaka), and Meito Sangyo Co. Ltd. (Aichi), respectively. The Rhizo- pus lipase was immobilized on a ceramic carrier SM-10, a gift from NGK Insulators Ltd. (Aichi), as described in our previous paper (14). After the ceramic carrier (30g) was suspended in 120ml of 10% lipase solution, cold acetone (-SO’C) was added gradually with stirring, and then the precipitate was dried in vacua.
Oils and fatty acid Borage oil and arachidonic acid (20 : 4n-6, AA)-containing oil (TGA-25; single cell oil from Mortiellera alpina) were obtained from Nihon Syn- thetic Chemical Industry Co. Ltd. (Osaka) and from Suntory Ltd. (Osaka), respectively. Sardine oil (eicosa- pentaenoic acid, 20 : 5n-3, EPA, 18%) and tuna oil @HA, 30%), and SOS fat (1,3-stearoyl-2-oleoyl-glycerol; purity 69%) were gifts from Maruha Corp. (Tokyo), and Fuji Oil Co. Ltd. (Osaka), respectively. Linseed oil, trimyristin and caprylic acid (8 : 0, CA) were purchased from Yamakei Sangyo Co. Ltd. (Osaka), Tokyo Kasei Kogyo Co. Ltd. (Tokyo) and Wako Pure Chemical Industries Ltd. (Osaka), respectively.
Preparation of GLA- and AA-rich triglycerides AA- rich oil was prepared according to our previous paper (19). The mixture containing 4g of TGA-25, 4g of water and 720U of Candida lipase was stirred (500 rpm) at 35°C for 16 h. After the reaction, glycerides were extracted with n-hexane under alkaline conditions as described previously (14). GLA-rich oil was prepared from borage oil by the same method, except that 350U of the lipase was used. Because the resulting oils con- tained partial glycerides at a content of about 20X, triglycerides were purified by silica gel 60 (Merck, Darm- stadt, Germany) column chromatography using benzene as a elution solvent (7).
Preparation of randomly interesterilied oil The ar- rangement of fatty acids in the triglyceride molecules was randomized according to the conventional inter- esterification method (19). Triglycerides (10 g AA-rich oil, 10 g GLA-rich oil, 8 g linseed oil, 20 g sardine oil, 8 g tuna oil, 5 g SOS fat, and 3 g trimyristin) were mixed to make the contents of the constituent fatty acids as equal as possible, and then the mixture was inter- esterified by incubation at 55°C with stirring in the pres- ence of 1.5% Na-methylate for 30 min. The resulting oil was extracted with 200ml of n-hexane after addition of 50ml of water, and then the solvent was removed by evaporation. No partial glycerides were generated, and the fatty acid composition of the oil after the reaction was not different from that before the reaction.
Reactions Acidolysis was carried out in a screw- capped vessel according to a method described elsewhere (14). The immobilized lipase (4% of reaction mixture) was activated by shaking (140 oscillations/min) at 30°C
for 2 d in the oil/CA mixture (1 : 2, w/w) containing 2% water. Since the activated lipase catalyzes acidolysis and hydrolysis simultaneously, it was shaken in a fresh oil/ CA (1 : 2, w/w) mixture without water under the same conditions. After this pretreatment was performed a total of three times, the resulting enzyme was used for the acidolysis, which was conducted under the same con- ditions as those of the pretreatment.
Hydrolysis of the randomly interesterified oil with the Rhizopus lipase was carried out at 30°C with stirring (500 rpm) in a mixture of 1 g of oil and 3 g of water. After the reaction, 10ml of ethanol was added, and the acid value was measured by titration with 0.2N KOH. The extent of hydrolysis was calculated from the acid value of the reaction mixture and the saponification value of the substrate oil.
Analysis Lipase activity was measured by titration of fatty acids liberated from olive oil (Wako Pure Chemi- cal) with 0.05 N KOH, as described previously (21). The reaction was carried out at 30°C for 30min with stirring at 500 rpm. One unit (U) of lipase activity was defined as the amount of the enzyme that liberated 1 pmol of fatty acid per minute.
Glycerides were extracted with n-hexane under alkaline conditions as described previously (14), and free fatty acids in the aqueous phase were extracted with n-hexane after adjustment of the pH of aqueous phase to less than 2 by addition of 4N HCl (7). Fatty acids in glycer- ides were methylated using Na-methylate, and free fatty acids were esterified with gaseous HCl-methanol. These methyl esters were analyzed by use of a Hewlett-Packard 5890 plus gas chromatograph (Avondale, PA, USA) con- nected to a DB-23 capillary column (0.25 mm x 30m, J&W Scientific, Forsom, CA, USA) as described previous- 1Y (14).
Regiospecific analysis of triglycerides was carried out by Grignard degradation with ally1 magnesium bromide, followed by isolation and analysis of the 1,3-diglyceride fraction (22). Triglycerides were analyzed on two oc- tadecyl silica (ODS) columns (4.6 x 150 mm, Wakosil- 113C18 HG, Wako) connected to a high-performance liquid chromatography (HPLC) system (LC-9A; Shirnadzu Co., Kyoto). The sample was eluted with a mixture of acetone/acetonitrile (1 : 1, v/v) at a flow rate of 0.4 ml/min and 4O”C, and detected with a refractometer.
RESULTS
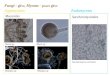
Acidolysis of randomly interesterified oil with CA The fatty acid contents of the randomly interesterified oil prepared as described in Materials and Methods were approximately the same except for that of palmitoleic acid (16 : In- 9, POA) (Table 1). The regiospecific analy- sis by Grignard degradation of the oil showed that the fatty acids were homogeneously distributed in the triglyceride molecules. The acidolysis of the oil with CA was conducted at 30°C using the immobilized Rhizopus lipase, and the time course is shown in Fig. 1. The acidolysis extent for each constituent fatty acid (A& was expressed relative to the content of the fatty acid ex- isting at the l(3)-position in the randomly interesterified oil, according to the equation
&iio= {(Fsub-%,,s)/(2/3 xF,,)) x 100 (1) where Fsub and F,,, are the contents (mol%) of a par- ticular fatty acid in the glycerides before and after the
VOL. 83, 1997 FATTY ACID SPECIFICITY OF RHIZOPUS LIPASE 323
TABLE I. Fatty acid composition of randomly interesterified oil
Fatty acid Content (mol%) Fatty acid Content
(moI%) 14 : 0 9.1 18 : 3n-6 8.0 16: 0 10.1 18 : 3n-3 8.4 16: 1 3.4 20 : 4 6.9 18 : 0 7.7 20 : 5 6.4 18: 1 13.4 22 : 6 6.4 18:2 8.3
reaction, respectively. The lipase acted very strongly on POA, oleic acid (18 : ln-9, OA), ALA (Fig. IB), myris- tic acid (14 : 0, MA), palmitic acid (16 : 0, PA) and LA (data not shown). AA and EPA were exchanged for CA at a constant rate from the early stage of the reaction (Fig. 1C). GLA and DHA were exchanged for CA after the exchanges of fatty acids which were good substrates reached a steady state, and the lipase acted on GLA more strongly than on DHA (Fig. 1D). The acidolysis- catalyzing activity of the Rhizopus lipase on each fatty acid was calculated based on the initial rate, and the results are shown in Table 2.
The Rhizopus lipase acted very weakly on fatty acids of which the carbon chain length was shorter than six carbon atoms, but acted strongly on not only CA but also capric acid. Since the randomly interesterified oil did not contain capric acid, the acidolysis was conducted using capric acid as a substrate, and the fatty acid specificity was evaluated. The results were the same as those obtained using CA, suggesting that the fatty acid specificity was not affected by the fatty acid used as a substrate (data not shown).
Hydrolysis of randomly interesterified oil It is known that lipase catalyzes esterification and trans- esterification even in a hydrolysis reaction system con- taining a large amount of water (7, 23). For avoidance of the effects of these simultaneous reactions on hydroly-
$100 g 80 A
3 60
.c
zz
40 5 20 e 2 O
i;:-j 0 10 20 30 40 50
s5 100 $ 80 +s a, 60 5 40 0 0 20
2 O 0 1020304050
100
D
80 60 40 20 0 u 0 10 20 30 40 50
Time(h) Time(h)
FIG. 1. Time course of the acidolysis of randomly interesterified oil with caprylic acid (CA). (A) Acidolysis extent expressed as the amount of CA incorporated into triglycerides. The amount incorpo- rated is expressed relative to that at the l(3)-position. (B) PaImitoleic (0 ), oleic (0 ) and a-linolenic ( 0 ) acids exchanged for CA. (C) Arachidonic (0 ) and eicosapentaenoic (0) acids exchanged for CA. (D) r-Linolenic (0) and docosahexaenoic (0) acids exchanged for CA. The acidolysis extent for each constituent fatty acid was calcu- lated using Eq. 1.
TABLE 2. Fatty acid specificity of Rhizopus lipase in acidolysis of randomly interesterified oil
Fatty acid Relative activity (%)
Fatty acid Relative activity (%)
14 : 0 105 18 : 3n-6 0 16:0 97 18 : 3n-3 106 16: 1 117 20 : 4 21 18:0 88 20 : 5 26 18: 1 100 22 : 6 0 18~2 89
The activity on each fatty acid is expressed relative to that on oleic acid.
sis, the fatty acid specificity of the lipase in hydrolysis was analyzed on the basis of the initial rate.
When the reaction mixtures of 1 g of randomly inter- esterified oil, 3 g of water, and 100, 150, or 200 U of the lipase were incubated with stirring (500rpm) at 30°C for 30 min, the hydrolysis extents were 8.6, 12.5, and 16.9X, respectively. The hydrolysis extent depended completely on the enzyme amount, showing that the simultaneous esterification and transesterification were negligible. The hydrolysis activity on each fatty acid (Ohio) was expressed according to the equation
A hio = (FdFg~y) X 100 (2)
where Fea and Fgly are the contents (mol%) of a particu- lar fatty acid in free fatty acids after the hydrolysis and in the substrate oil, respectively.
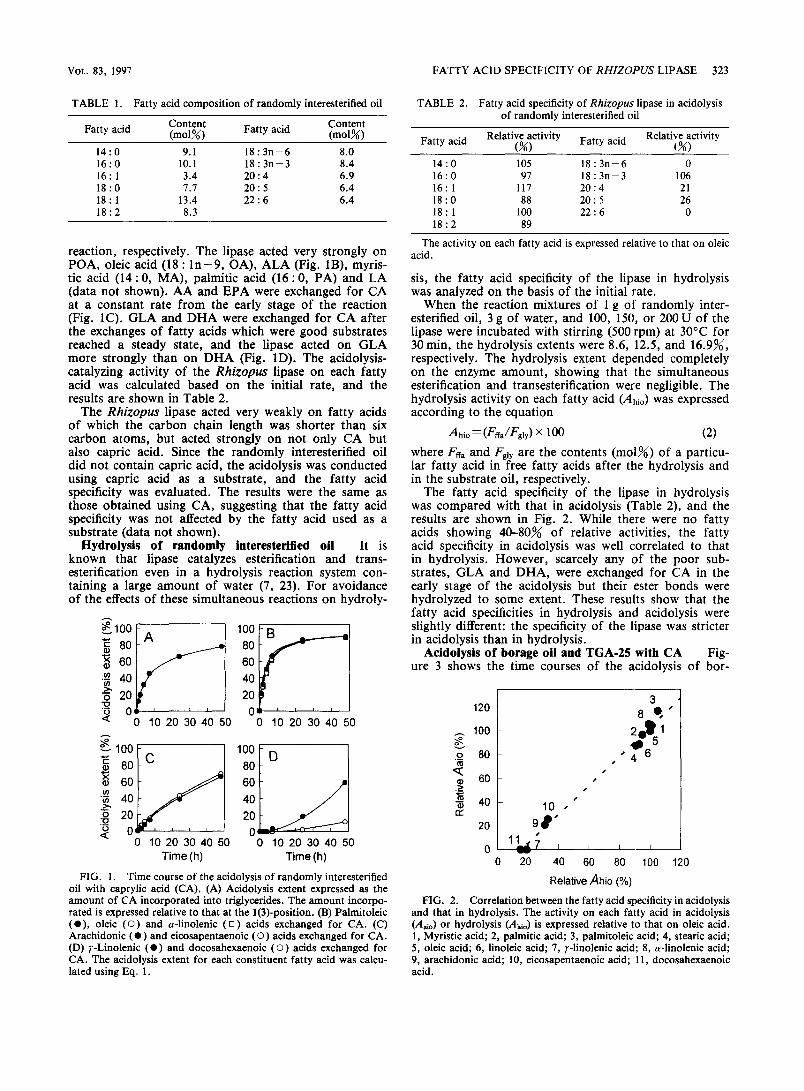
The fatty acid specificity of the lipase in hydrolysis was compared with that in acidolysis (Table 2), and the results are shown in Fig. 2. While there were no fatty acids showing 40-80X of relative activities, the fatty acid specificity in acidolysis was well correlated to that in hydrolysis. However, scarcely any of the poor sub- strates, GLA and DHA, were exchanged for CA in the early stage of the acidolysis but their ester bonds were hydrolyzed to some extent. These results show that the fatty acid specificities in hydrolysis and acidolysis were slightly different: the specificity of the lipase was stricter in acidolysis than in hydrolysis.
Acidolysis of borage oil and TGA-25 with CA Fig- ure 3 shows the time courses of the acidolysis of bor-
3 8 9'
2.91 .(+6
,'4 6 ,
, ,
1
1
J 0 20 40 60 80 100 120
Relative Ohio (%)
FIG. 2. Correlation between the fatty acid specificity in acidolysis and that in hydrolysis. The activity on each fatty acid in acidolysis (A& or hydrolysis (Ai,J is expressed relative to that on oleic acid. 1, Myristic acid; 2, palmitic acid; 3, pahnitoleic acid; 4, stearic acid; 5, oleic acid; 6, linoleic acid; 7, r-Iinolenic acid; 8, a-linolenic acid; 9, arachidonic acid; 10, eicosapentaenoic acid; 11, docosahexaenoic acid.
324 SHIMADA ET AL. J. FERMENT. BIOENG.,
60 I I-
0 20 40 60
0 20 40 60 Time (h)
FIG. 3. Time courses of acidolysis of borage oil (A) and arachi- donic acid-containing oil (TGA-25) (B) with caprylic acid. Symbols: ??, caprylic acid; o , palmitic acid; ??, oleic acid; ??, linoleic acid; + , r-linolenic acid; A, arachidonic acid.
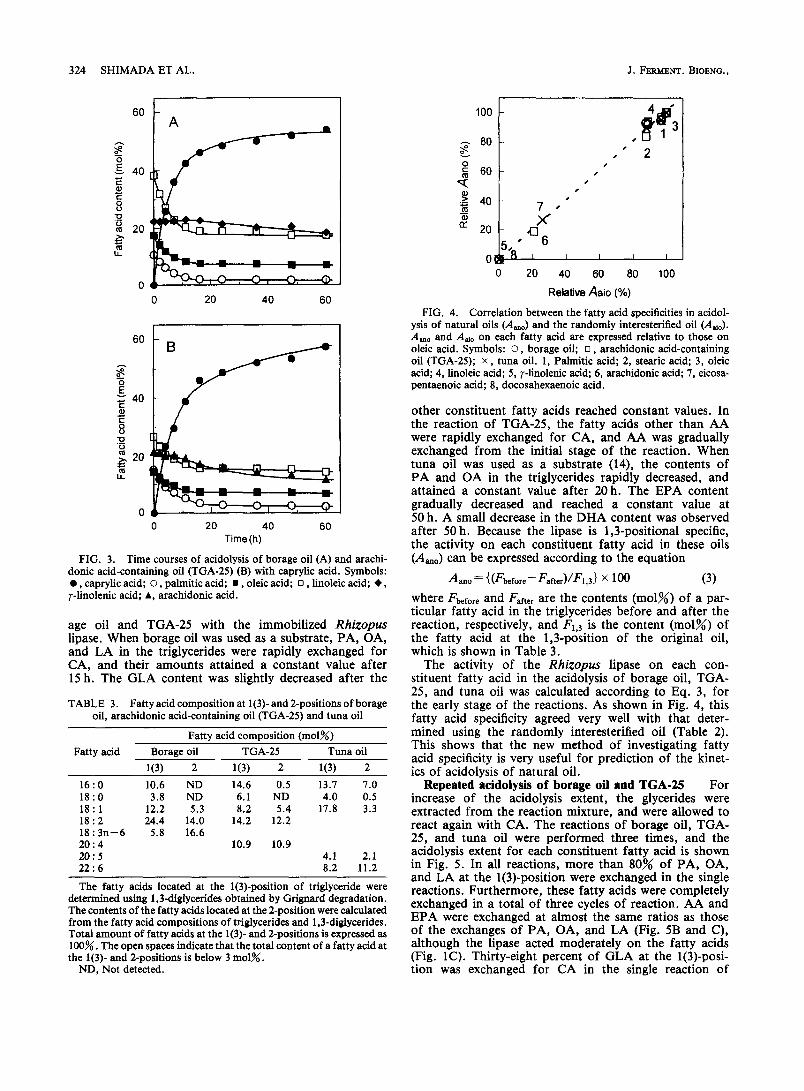
age oil and TGA-25 with the immobilized Rhizopus lipase. When borage oil was used as a substrate, PA, OA, and LA in the triglycerides were rapidly exchanged for CA, and their amounts attained a constant value after 15 h. The GLA content was slightly decreased after the
TABLE 3. Fatty acid composition at l(3)- and 2-positions of borage oil, arachidonic acid-containing oil (TGA-25) and tuna oil
Fatty acid composition (mol%) Fatty acid Borage oil TGA-25 Tuna oil
l(3) 2 l(3) 2 l(3) 2 16:0 10.6 ND 14.6 0.5 13.7 7.0 18:O 3.8 ND 6.1 ND 4.0 0.5 18: 1 12.2 5.3 8.2 5.4 17.8 3.3 18:2 24.4 14.0 14.2 12.2 18 : 3n-6 5.8 16.6 20 : 4 10.9 10.9 20 : 5 4.1 2.1 22 : 6 8.2 11.2
The fatty acids located at the l(3)-position of triglyceride were determined using 1,3-diglycerides obtained by Grignard degradation. The contents of the fatty acids located at the 2-position were calculated from the fatty acid compositions of triglycerides and 1,3_diglycerides. Total amount of fatty acids at the l(3)- and 2-positions is expressed as 100%. The open spaces indicate that the total content of a fatty acid at the l(3)- and 2-positions is below 3 mol%.
ND, Not detected.
100 - 4f$ ‘;; 80
- d 8’
% , 2
; 60 - , ,
q , 9 ,
g 40 -
d 20- 5, ’ Q6
ol$” ’ I 1 I I
0 20 40 60 80 100
Relative Aaio (%)
FIG. 4. Correlation between the fatty acid specificities in acidol- ysis of natural oils (A,,) and the randomly interesterified oil (A&. A,, and Ati, on each fatty acid are expressed relative to those on oleic acid. Symbols: 0, borage oil; ??, arachidonic acid-containing oil (TGA-25); X , tuna oil. 1, Pahnitic acid; 2, stearic acid; 3, oleic acid; 4, linoleic acid; 5, r-linolenic acid; 6, arachidonic acid; 7, eicosa- pentaenoic acid; 8, docosahexaenoic acid.
other constituent fatty acids reached constant values. In the reaction of TGA-25, the fatty acids other than AA were rapidly exchanged for CA, and AA was gradually exchanged from the initial stage of the reaction. When tuna oil was used as a substrate (14), the contents of PA and OA in the triglycerides rapidly decreased, and attained a constant value after 20 h. The EPA content gradually decreased and reached a constant value at 50 h. A small decrease in the DHA content was observed after 50 h. Because the lipase is 1,3-positional specific, the activity on each constituent fatty acid in these oils (A,,) can be expressed according to the equation
A*ll,= {(Fbefore-F*fter)/F~,s} x 100 (3) where Fbefore and Fafter are the contents (mol%) of a par- ticular fatty acid in the triglycerides before and after the reaction, respectively, and FlP3 is the content (mol%) of the fatty acid at the 1,3-position of the original oil, which is shown in Table 3.
The activity of the Rhizopus lipase on each con- stituent fatty acid in the acidolysis of borage oil, TGA- 25, and tuna oil was calculated according to Eq. 3, for the early stage of the reactions. As shown in Fig. 4, this fatty acid specificity agreed very well with that deter- mined using the randomly interesterified oil (Table 2). This shows that the new method of investigating fatty acid specificity is very useful for prediction of the kinet- ics of acidolysis of natural oil.
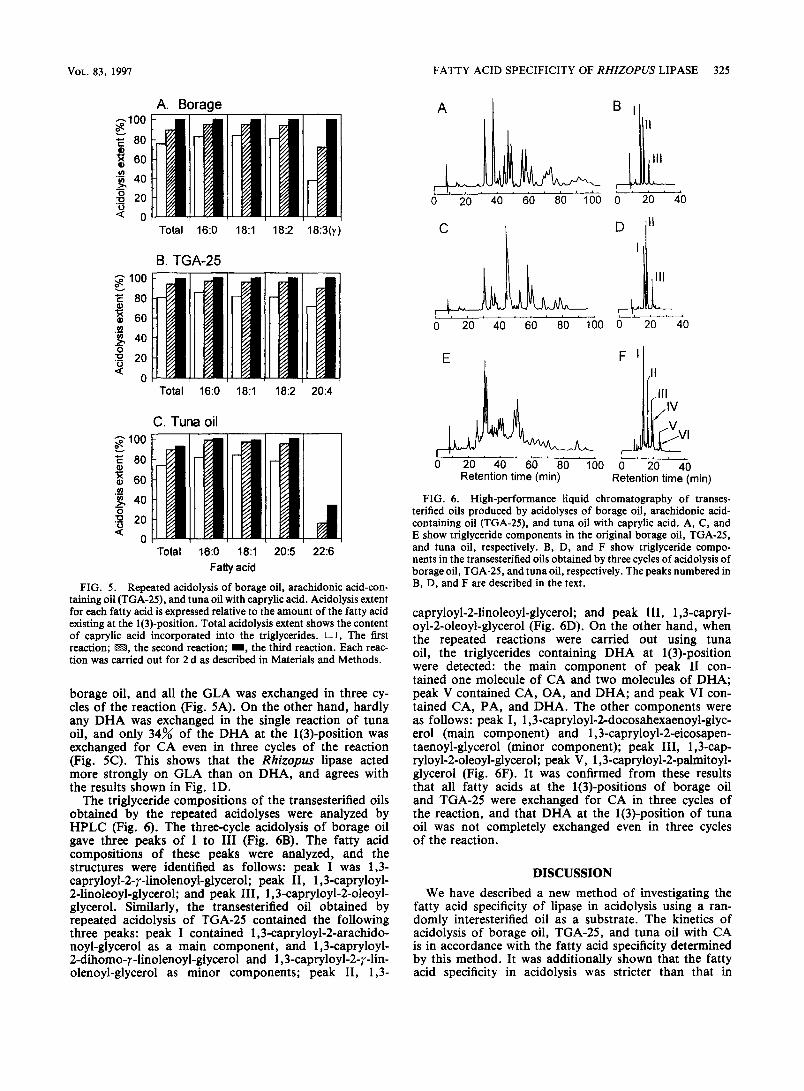
Repeated acidolysis of borage oil and TGA-25 For increase of the acidolysis extent, the glycerides were extracted from the reaction mixture, and were allowed to react again with CA. The reactions of borage oil, TGA- 25, and tuna oil were performed three times, and the acidolysis extent for each constituent fatty acid is shown in Fig. 5. In all reactions, more than 80% of PA, OA, and LA at the l(3)-position were exchanged in the single reactions. Furthermore, these fatty acids were completely exchanged in a total of three cycles of reaction. AA and EPA were exchanged at almost the same ratios as those of the exchanges of PA, OA, and LA (Fig. 5B and C), although the lipase acted moderately on the fatty acids (Fig. 1C). Thirty-eight percent of GLA at the l(3)-posi- tion was exchanged for CA in the single reaction of
VOL. 83, 1997
A. Boraae
FATTY ACID SPECIFICITY OF RHIZOPUS LIPASE 325
Total 16:0 181 18:2 18:3(y)
B. TGA-25 gg 100
iz 60
8 60 .B p 40
g 8
20
0 Total 16:0 18:l 18:2 20:4
C. Tuna oil 2100
k 80
8 60
$ 40
i 20 Y
0 Total 16:O 18:l 205 22:6
Fatty acid
FIG. 5. Repeated acidolysis of borage oil, arachidonic acid-con- taining oil (TGA-25), and tuna oil with caprylic acid. Acidolysis extent for each fatty acid is expressed relative to the amount of the fatty acid existing at the l(3)-position. Total acidolysis extent shows the content of caprylic acid incorporated into the triglycerides. 0, The first reaction; @, the second reaction; I, the third reaction. Each reac- tion was carried out for 2 d as described in Materials and Methods.
borage oil, and all the GLA was exchanged in three cy- cles of the reaction (Fig. 5A). On the other hand, hardly any DHA was exchanged in the single reaction of tuna oil, and only 34% of the DHA at the l(3)-position was exchanged for CA even in three cycles of the reaction (Fig. 5C). This shows that the Rhizopus lipase acted more strongly on GLA than on DHA, and agrees with the results shown in Fig. 1D.
The triglyceride compositions of the transesterified oils obtained by the repeated acidolyses were analyzed by HPLC (Fig. 6). The three-cycle acidolysis of borage oil gave three peaks of I to III (Fig. 6B). The fatty acid compositions of these peaks were analyzed, and the structures were identified as follows: peak I was 1,3- capryloyl-2-r-linolenoyl-glycerol; peak II, 1,3-capryloyl- 2-linoleoyl-glycerol; and peak III, 1,3-capryloyl-2-oleoyl- glycerol. Similarly, the transesterified oil obtained by repeated acidolysis of TGA-25 contained the following three peaks: peak I contained 1,3-capryloyl-2-arachido- noyl-glycerol as a main component, and 1,3-capryloyl- 2-dihomo-r-linolenoyl-glycerol and 1,3-capryloyl-2-y-lin- olenoyl-glycerol as minor components; peak II, 1,3-
0 20 40 60 80
iJ 20 40 60 80 100 I 0 L’. 20 . 40 I
F ’
,‘I
0 20 40 60 80 100 0 20 40 Retention time (min) Retention time (min)
FIG. 6. High-performance liquid chromatography of transes- terified oils produced by acidolyses of borage oil, arachidonic acid- containing oil (TGA-25), and tuna oil with caprylic acid. A, C, and E show triglyceride components in the original borage oil, TGA-25, and tuna oil, respectively. B, D, and F show triglyceride compo- nents in the transesterified oils obtained by three cycles of acidolysis of borage oil, TGA-25, and tuna oil, respectively. The peaks numbered in B, D, and F are described in the text.
capryloyl-2-linoleoyl-glycerol; and peak III, 1,3-capryl- oyl-2-oleoyl-glycerol (Fig. 6D). On the other hand, when the repeated reactions were carried out using tuna oil, the triglycerides containing DHA at l(3)-position were detected: the main component of peak II con- tained one molecule of CA and two molecules of DHA; peak V contained CA, OA, and DHA; and peak VI con- tained CA, PA, and DHA. The other components were as follows: peak I, 1,3-capryloyl-2-docosahexaenoyl-glyc- erol (main component) and 1,3-capryloyl-2-eicosapen- taenoyl-glycerol (minor component); peak III, 1,3-cap- ryloyl-2-oleoyl-glycerol; peak V, 1,3-capryloyl-2-pahnitoyl- glycerol (Fig. 6F). It was confirmed from these results that all fatty acids at the l(3)-positions of borage oil and TGA-25 were exchanged for CA in three cycles of the reaction, and that DHA at the l(3)-position of tuna oil was not completely exchanged even in three cycles of the reaction.
DISCUSSION We have described a new method of investigating the
fatty acid specificity of lipase in acidolysis using a ran- domly interesterified oil as a substrate. The kinetics of acidolysis of borage oil, TGA-25, and tuna oil with CA is in accordance with the fatty acid specificity determined by this method. It was additionally shown that the fatty acid specificity in acidolysis was stricter than that in

326 SHIMADA ET AL. J. FERMENT. BIOENG.,
hydrolysis. Precision processing of PUFA-containing oils may be enabled by use of advantage of a small differ- ence in fatty acid specificities in hydrolysis and acidoly- sis.
Application to regiospecilic analysis Regiospecific analysis of triglycerides in natural oils has been conduct- ed by two methods. One is the method of analyzing the fatty acid composition of 1,3-diglycerides or 2- monoglycerides obtained by Grignard degradation (22), which was used in this work (Table 3), and the other is the method of analyzing the fatty acid composition of 1,3-diglycerides or free fatty acids obtained by the hydrolysis of an oil with a free 1,3-positional specific lipase (23, 24). When this enzymatic method is used, the products in the early stage of the reaction must be ana- lyzed because the esterification and transesterification oc- cur simultaneously during the hydrolysis. However, the constituent fatty acids are liberated from the oil accord- ing to the fatty acid specificity of the lipase (Table 2). Thus the erroneous result that the fatty acids, on which the lipase acts strongly, exist at the l(3)-position is ob- tained.
In our reaction system with the immobilized lipase, the simultaneous hydrolysis did not occur, and only acidolysis proceeded efficiently (14, 15). In addition, the fatty acids other than DHA at the 1,3-position were com- pletely exchanged for CA in three cycles of the reaction (Fig. 3). Actually, the 2-position fatty acid compositions of borage oil and TGA-25 agree with the compositions (except for CA) of the transesterified oils obtained by their repeated reaction, respectively. Furthermore, when the acidolyses of safflower and linseed oils were per- formed three times, the fatty acid compositions of the transesterified oils thus obtained were found to agree with their composition at the 2-positions of the original oils (15). These results indicate that the acidolysis with the immobilized 1,3-specific lipase can be applied to regiospecific analysis, although it cannot be applied to oils containing a fatty acid, like DHA, on which the li- pase acts only very weakly.
Structured lipids containing GLA and AA It has been suggested that GLA supplementation could be use for modulation of immune and inflammatory responses and treatment of related diseases (3). Actually, borage oil is used as a material of functional foods for treat- ment of atopic eczema in Japan, and is added to formu- las fed to premature infants in Europe (25). AA is a fat- ty acid of potential pharmaceutical value because of a precursor of local hormones involved in the AA cascade (2). High-absorptive oils, structured lipids, containing these fatty acids are desired as medicines, clinical nutri- tion, health foods and components of cosmetics, and can be produced using the immobilized Rhizopus lipase (Figs. 3, 5, and 6), which is permitted for food process- ing. Furthermore, structured lipids containing DHA and essential fatty acids could also be produced by means of this acidolysis system, as reported previously (14, 15). Our findings show that this reaction system is useful for the production of structured lipids containing functional fatty acids.
REFERENCES
1. Fischer, S.: Dietary polyunsaturated fatty acids and eicosanoid formation in humans, p. 169-198. In Paoletti, R. and Kritchev- sky, D. (ed.), Advances in lipid research, vol. 23. Academic
Press, New York (1989). 2. Johnston, P. V.: Dietary fat, eicosanoids, and immunity, p.
103-141. In Paoletti, R. and Kritchevsky, D. (ed.), Advances in lipid research, vol. 21. Academic Press, New York (1985).
3. Wu, D. and Meydaai, S. N.: Z-Linolenic acid and immune func- tion, p. 106-117. In Huang, Y.-S. and Mills, D. E. (ed.), r- Linolenic acid: metabolism and its roles in nutrition and medi- cine. AOCS Press, Illinois (1996).
4. Yamane, T.: Enzyme technology for the lipids industry: an engineering overview. J. Am. Oil Chem. Sot., 64, 1657-1661 (1987).
5. Hoshino, T., Yamane, T., and Shimizu, S.: Selective hydroly- sis of fish oil by lipase to concentrate n-3 polyunsaturated fatty acids. Agric. Biol. Chem., 54, 1459-1467 (1990).
6. Shimada, Y., Maruyama, K., Okazaki, S., Nakamura, M., Sugihara, A., and Tominaga, Y.: Enrichment of polyunsaturat- ed fatty acids with Geotrichum candidum lipase. J. Am. Oil Chem. Sot., 71, 951-954 (1994).
7. Shimada, Y., Maruyama, K., Nakamura, M., Nakayama, S., Sugihara, A., and Tominaga, Y.: Selective hydrolysis of poly- unsaturated fatty acid-containing oil with Geotrichum candidum lipase. J. Am. Oil Chem. Sot., 72, 1577-1581 (1995).
8. Tanaka, Y., Hirano, J., and Funada, T.: Concentration of docosahexaenoic acid in glyceride by hydrolysis of fish oil with Candida cylindracea lipase. J. Am. Oil Chem. Sot., 69, 1210- 1214 (1992).
9. Hill, M. J., Kiewitt, I., and Mukherjee, K. D.: Enzymatic frac- tion of fatty acids: enrichment of r-linolenic acid and docosa- hexaenoic acid by selective esterification catalyzed by lipase. J. Am. Oil Chem. Sot., 67, 561-564 (1990).
10. Shimada, Y., Sugihara, A., Nakano, H., Kuramoto, T., Nagao, T., Gemba, M., and Tominaga, Y.: Purification of docosa- hexaenoic acid by selective ester&cation of fatty acids from tuna oil with Rhizopus delemar lipase. J. Am. Oil Chem. Sot., 74 (1997). (in press)
11. Yokozeki, K., Yamanaka, S., Takinami, K., Hirose, Y., Tan&a, A., Sonomoto, K., and Fokui, S.: Application of immobi- lized lipase to regio-specific interesterification of triglyceride in organic solvent. Eur. J. Appl. Microbial. Biotechnol., 14, l-5 (1982).
12. Sridhar, R., Lakshminarayana, G., and Kaimal, T. N. B.: Modification of selected indian vegetable fats into cocoa butter substitutes by lipase-catalyzed ester interchange. J. Am. Oil Chem. Sot., 68, 726730 (1991).
13. Shieh, C.-J., Akoh, C. C., and Koehler, P. E.: Four-factor response surface optimization of the enzymatic modification of triolein to structured lipids. J. Am. Oil Chem. Sot., 72, 619- 623 (1995).
14. Shiiada, Y., Sugihara, A., Maruyama, K., Nagao, T., Nakayama, S., Nakano, H., and Tominaga, Y.: Production of structured lipid containing docosahexaenoic and caprylic acids using immobilized Rhizopus delemar lipase. J. Ferment. Bioeng., 81, 299-303 (1996).
15. Shimada, Y., Sugihara, A., Nakano, H., Yokota, T., Nagao, T., Komemushi, S., and Tominaga, Y.: Production of struc- tured lipids containing essential fatty acids by immobilized Rhizopus delemar lipase. J. Am. Oil Chem. Sot., 73, 1415- 1420 (1996).
16. Iwai, M., Okumura, S., and Tsujisaka, Y.: The comparison of the properties of two lipases -from Penicillium cyclopium Westring. Agric. Biol. Chem., 39, 1063-1070 (1975).
17. Matori, M., Kusida, Y., Adachi, S., and Ota, Y.: Illustration fatty acid specificity in lipase hydrolysis of natural oils and fats. Yukagaku, 41, 347-348 (1992). (in Japanese)
18. Shimada, Y., Sugihara, A., Nagao, T., Kitai, S., Nakamura, M., Gemba, M., and Tominaga, Y.: Evaluation of fatty acid specificity using interesterified oil. Kagaku to Kogyo, 69, 95-99 (1995). (in Japanese)
19. Shimada, Y., Suglhara, A., Maruyama, K., Nagao, T., Nakayama, S., Nakano, H., and Tominaga, Y.: Enrichment of arachidonic acid: selective hydrolysis of a single-cell oil from Mortierelfa with Candida cylindracea lipase. J. Am. Oil Chem. Sot., 72, 1323-1327 (1995).

VOL. 83, 1997
20. Foglia, T. A. and Sonnet, P. E.: Fatty acid selectivity of li- Riemenschneider, R. W.: Pancreatic lipase hydrolysis of pases: r-linolenic acid from borage oil. J. Am. Oil Chem. Sot., triglycerides by semimicro technique. J. Am. Oil Chem. Sot., 72, 417-420 (1995). 41, 693-696 (1964).
21.
22.
Sugihara, A., Ueahima, M., Skimada, Y., Tsunasaws, S., and Tominaga, Y.: Purification and characterization of a novel ther- mostable lipase from Pseudomonas cepuciu. J. Biochem., 112, 598-603 (1992). Becker, C. C., Rosenquist, A., and Helmer, G.: Regiospecific analysis of triacylglycerols using ally1 magnesium bromide. Lipids, 28, 147-149 (1993).
23. Luddy, F. E., Barford, R. A., Herb, S. F., Magidman, P., and
24.
25.
FATTY ACID SPECIFICITY OF RHIZOPUS LIPASE 327
iwai, M., Tsujisaka, Y., Okumura, S., and Katsumoto, H.: Application of microbial lipase to the determination of the fatty acid distribution in natural fats. Yukaaaku. 29. 587-591 (1980). (in Japanese)
_
Crazier, G. L., Fleith, M., and Secretin, M.-C.: y-Linolenic acid in infant formula, p. 246-251. In Huang, Y.-S. and Mills, D. E. (ed.), r-Linolenic acid: metabolism and its roles in nutri- tion and medicine. AOCS Press, Illinois (1996).