Embed Size (px)
Citation preview

FAUNAof AUSTRALIA
34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
M.M. BRYDEN
1

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
2

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
TERMINOLOGY
The language of biology is vital in imparting knowledge and exchanging ideas.The language must be precise, because confusion can (and does) arise ifmultiple terms are used to describe a structure or physiological process.Veterinary anatomists have attempted to overcome misconceptions in biologydue to confusion in terms by developing a standard set of names for structures(Nomina Anatomica Veterinaria and Nomina Histologica 1983). Adoption ofappropriate terms from that source for the Eutheria generally would bebeneficial to biologists, and at the risk of offending those who fear some of theirfavourite terms are threatened, I shall follow the standard terms of the Nomina,or their English equivalents, in this chapter.
EXTERNAL BODY FORM
Eutheria in Australia range in size from the smallest bats, with body weight of 5g or less, to the large whales, with body weights exceeding 100 tonnes.
Although bearing the basic characters that distinguish mammals generally,namely the presence of hair and mammary glands, the external form ofAustralian Eutheria is very variable (Fig. 34.1). This is related partly to theenvironment in which they live, which varies from totally aquatic to arid desert.Small rodents have a dense pelage, whereas the totally aquatic mammals(dugongs and whales) have little or no hair on the general body surface.Mammals that need to conserve body heat, notably the aquatic mammals, havefusiform bodies and small appendages, presenting to the environment arelatively small body surface area per unit weight. This advantage is augmentedin many species by their very large size. Animals that inhabit hot environments,such as the camelids, have slender bodies and long limbs and, therefore, arelatively large surface area.
Figure 34.1 The shape of the head of fourAustralian eutherian mammals. (© ABRS)
[M. Thompson]
3

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The form of the limbs varies greatly. Most terrestrial species are digitigrade, buthave varying degrees of reduction in the number of digits. The rodents have fivedigits on each foot, and the digits of the hind feet are webbed in the aquatic rats.The artiodactyls (bovids, suids, cervids and camelids) and the perissodactyls(equids) have two digits and one digit on each foot, respectively. In the flyingmammals, the bats, the limbs are modified greatly to function as wings. In theaquatic mammals the limbs are adapted for swimming and, in some species, aregreatly modified. The hind limbs are absent in totally aquatic mammals.
The head (Fig. 34.1) is highly variable in form, from long and narrow to shortand broad. In some of the cervids and bovids it bears large antlers or horns. Inmany of the microchiropterids the ears and nose leaf are complex. There is along beak and rounded melon in the dolphins.
Not all eutherian mammals have a discernible tail, but some component alwaysremains. Even in humans, who have no tail, there are four small vertebrae thatconstitute the coccyx, the rudiment of a tail. The tail is long, short or absent inbats, whereas it is long and slender and acts as a thermoregulatory organ in somerodents. It bears long hairs that aid its use as a fly-whisk in the Equidae, whereasit is greatly modified to form the powerful swimming organ of the whales. Inrabbits and many artiodactyls the white underside is shown as a warning ofdanger to others.
The shape of the body is often used as a means of communication in the socialand sexual life of mammals, serving to produce appropriate responses in otherindividuals. Special shapes are often associated with the recognition of the sexesand in some instances this is served by body size. The most extreme example ofthis is seen in elephant seals, males of which are up to ten times larger thanfemales. A direct relationship between sexual disparity in size and polygyny hasbeen postulated. Some animals deliberately alter their shape and apparent size toexpress fear or aggression, for example the arching of the back and piloerectionin the cat.
THE INTEGUMENT
The skin (Fig. 34.2) marks the interface between the organism and itsenvironment. The skin of mammals has special features, not found in non-mammalian vertebrates, connected mainly with temperature control and thefunction of the skin as a sense organ. Investigation of the differences in the skinsof various mammals shows how closely the activities of the body are related tothe conditions of the environment: it varies in texture, colour, scent andtemperature among other features. This conformity with the environment ismaintained by control on time scales between the few seconds required fortemperature regulation and the many generations for selection of a new set ofgenes that determine coat colour.
The Epidermis and its Appendages
The outer layer of skin, the epidermis, is a non-vascular stratified squamousepithelium. It consists of a variable number of layers of cells, the inner of whichare active, continually producing cells that pass towards the surface andgradually undergo a process of cornification. The thickness of the epidermisvaries according to species and the region of the body, being greatest in thoseareas that are subjected to wear, for example the footpads of rodents, felids andcanids. The protein keratin is the distinctive component of the epidermis and itsderivatives. It is virtually insoluble in water and, therefore, forms a protectiveouter covering for the body. The process of keratinisation is not peculiar tomammals, but it is highly developed in them and in special parts of the body itgives rise to hairs, claws (or nails), hooves or horns.
4

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The hair follicles and the glands closely associated with them, the sweat glandsand sebaceous glands, are derivatives of the epidermis that project inwards intothe deeper layer, the dermis. Each hair consists of a rod of elongated keratinisedcells, forming a thread, usually with a central medulla. The mesenchyme at thebase of a developing hair forms a vascular papilla and the epidermal cells abovethis continue to produce keratin, thus providing the constantly growing base ofthe hair.
When large and dense, the hairs serve to retain heat. The associated sebaceousglands keep them and the skin surface oily and hence, waterproofed. They alsoare provided with nerve fibres which wrap around the base of the hair follicle. Innearly all mammals highly sensitive vibrissae, which are elongated andstiffened, have developed around the snout and make the animals sensitive tovery light touch or even to small changes of air-pressure.
Each sebaceous gland consists of a few short sacs, the alveoli, opening into thehair follicle. The secretory cells become converted into the substance sebum,which keeps the hair and the adjacent skin surface greasy. Attached to each hairis a small bundle of unstriated muscle fibres, the arrector pili, in most species.
The sweat glands are tubular and highly variable in form. They are reduced insemi-aquatic, and absent in totally aquatic mammals and are confined only tocertain regions of the body in some species. The secretory cells in the basal partof the gland are surrounded by myoepithelial cells that aid in expelling thesecretion.
The tips of the digits of mammals are nearly always covered with hardenedplates of keratin, which in their simplest form, claws, are similar to those ofreptiles and birds. The coronary border (vallum) of the claw fits into the spaceunder the ungual crest of the distal phalanx. This relationship is hidden by theskin of the claw fold. Dorsally, this fold is a modification of the hairy skin whichis free from hair on one side and fused to the horn of the claw. As the horny
Figure 34.2 Diagram of the skin, showing some of the accessory structures -the hairs, sweat and sebaceous glands - and the smooth muscle associatedwith the hairs (arrector pili). Some sensory nerve fibres are shown. Notshown are the autonomic motor nerve fibres that supply the sweat glands,arrector pili and the walls of blood vessels in the skin. Magnification \X 400(© ABRS) [S. Collin]
Encapsulatednerve ending
Epidermis
Freenerveending
Sensornervefibres
Sweat glandHair
Dermis
Sebaceousgland
Arrector pili
5

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
material is produced and grows out, it is covered by a thin stratum tectoriumwhich adheres to the proximal part of the claw. A furrow along the palmar orplantar surface of the claw separates it from the foot pad in a similar manner.
The horns found in various mammals also are formed of keratin, produced byareas of skin overlying prolongations of the bones of the skull. The antlers ofCervidae, however, are purely bony.
Small scales occur on certain mammals, for example on the tail of rats. Cyclicepidermal cellular proliferation is a vertebrate characteristic and probablyfollows an autonomous rhythm regulated through neurochemical systems, inresponse to environmental stimuli and related to adaptive factors.
Periodic shedding of the outer epidermis, the moult, seems to be related moreclosely to seasonal factors in mammals and birds than in fish, amphibians andreptiles. In mammals this shedding includes the hairs. The regular moulting andhair replacement results in a seasonal variation in coat quality in most species.
The Integument and Temperature Regulation
Hair tends to trap a layer of air which insulates the body, reducing heat exchangewith the environment. The air-retaining capacity is increased in some animals bya roughening of the surfaces of the hairs (examples are the wool of sheep, somegoats and rabbits) causing them to stick together. The hairs are not all of thesame kind. In many species long stiff hairs are found mingled with soft fur. Aspointed out above, the hair of many mammals changes according to the time ofyear so as to provide a thicker covering in winter than in summer.
Not all mammals use an air layer for the retention of heat. The body of whales,sea cows and seals is covered with a thick layer of fat, often called the blubber,which acts as an effective insulating blanket immediately beneath the skin.
The integument has mechanisms for losing as well as retaining body heat. Someof the sweat glands produce a watery solution, which serves to cool the bodythrough evaporation. Some mammals simulate this action behaviourally bywetting the skin periodically, for example, pigs in wallows and seals on a sunnybeach. The amount of heat lost from the skin surface is also controlled byregulating the flow of blood through the capillaries and arterio-venousanastomoses. Capillary blood flow is controlled locally, whereas flow througharterio-venous anastomoses is controlled centrally in the spinal cord and thehypothalamus. The surface of the lungs also provides a means of heat loss:animals such as dogs and sheep lose excess heat by panting.
Scent Glands
Scent-producing glands are important in that they provide the mechanism forrecognition of members of the species, especially in social mammals, and forbringing the sexes together. Although many mammals have good eyesight, theyrely mainly upon smell for recognition. There are probably characteristicdifferences of scent between individuals.
Scent glands occur in the skin of various parts of the body, for example theinfraorbital, interdigital and inguinal regions of certain artiodactyls, thesubmental and carpal regions of suids, the anal and caudal regions of canids.Some bat species have a glandular throat pouch, although its function isunknown. The characteristic smell of the rabbit is believed to be due to a pair ofinguinal glands. The scents produced by these glands may be left on trees andother objects where they serve to mark the territory of an individual.
6

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
Mammary Glands
These glands are characteristic of mammals and serve to nourish their young,which are born in a relatively immature and dependent state. They are pairedglands that form in the embryo along two mammary lines, extending from theaxilla to the inguinal region on either side of the midline. Mammary glands mayarise anywhere along these lines. The number formed and their location varieswith the species. In the resting state, they consist of a branching system of ducts.During pregnancy the epithelium of these ducts divides actively to producesecretory alveoli at their ends and, after birth of the young, the alveoli producethe milk.
The functioning of the mammary glands is dependent upon the interplay ofmultiple and complex nervous and endocrine factors. The growth of the ductsystem appears to depend primarily upon oestrogen, but for completedevelopment of the alveoli both oestrogen and progesterone are required. Thehypophysis also has a direct effect upon mammary growth through its secretionof prolactin and somatotropin. To obtain full morphological development of thegland comparable to that obtained in late pregnancy, prolactin, progesterone,oestrogen, somatotropin and adrenal corticoids are all believed to be necessary.
It is thought that when the inhibiting levels of circulating progesterone andoestrogen fall abruptly at the end of pregnancy, the increased output of prolactinby the hypophysis and the secretion of adrenal cortical steroids bring about milksecretion from the fully developed mammary gland.
The continued secretion of prolactin by the hypophysis appears to be necessaryfor the maintenance of lactation. The secretion of normal levels of prolactin byintact animals is dependent upon a neurohormonal reflex, in which the periodicsensory stimulus of sucking acts upon the hypothalamus to promote the releaseof prolactin.
Removal of milk from the glands depends mainly upon the stimulus of sucking,which acts via the hypothalamus to cause release of the hormone oxytocin fromthe neurohypophysis. This, in turn, stimulates the myoepithelial cells of thegland to contract, ejecting the accumulated milk from the alveoli and fine ductsinto the large ducts and sinuses of the gland.
The Dermis
The tissues of the dermis impart important properties to the skin as a whole. Thedermis forms a tough, flexible, elastic covering over the surface of the body.Immediately deep to it the hypodermis contains a large amount of fat in someanimals, for example the aquatic mammals, the pig and man. The dermiscontains almost all the nerve endings of the skin and serves to support the bloodvessels that extend to the epidermis. It plays a large part in the defence againstbacterial invasion.
The Musculature of the Skin
The skin contains unstriated (or smooth) muscle fibres which permit a certaindegree of movement. In addition to the arrector pili muscles, smooth muscleoccurs in the nipples and scrotum. In most mammals there are extensive striatedmuscles known collectively as the cutaneous muscles. Formerly termed thepanniculus carnosus, they occur in the superficial fascia beneath the skin,closely associated with the dermis. They are attached to the dermis and areanchored to the subcutaneous fascia rather than to bone, and their contractioncauses movement of the skin.
7

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
General
Since the colour of the skin and its appendages serves usually for concealmentrather than for recognition or stimulation as occurs in birds, the colours of theskin and hair of mammals tend to be relatively dull. The majority of mammalsare colour blind. Recognition and stimulation are accomplished mostly by scentor visual cues other than colour. Many mammals have coats of different coloursin different seasons. Deer become darker in winter, lighter and dappled in thesummer. Some Arctic species become white in winter, but Antarctic species donot, reflecting the absence of terrestrial predators in the Antarctic region.Changes in the basic colour patterns involve alteration of the hereditary make-up by variation and natural selection.
It is clear from the foregoing that the skin is far more than a passive covering forthe body. It plays a part in rapid adjustments such as temperature regulation, aswell as in the slow adjustments by which a species is kept in balance with itsenvironment. In addition to its roles in temperature regulation, protectionagainst abrasion and infection, camouflage, the synthesis of vitamins and thesecretion of sebum, sweat, milk and scents, it is an extensive sense organ thatplays an important part in maintaining an animal’s awareness of its surroundingsand the presence of various stimuli.
THE SKELETON
The bony skeleton (Fig. 34.3) provides structural support and protection as wellas levers for muscular action. It functions as a store of minerals and fat and is asite for blood cell formation. Bone is vital living tissue, supplied with bloodvessels and nerves, that is metabolically active such that it may be altered inshape, size and position by mechanical or biochemical demands.
Figure 34.3 Comparison of the skeleton of a blue whale and a dog. Thebones of the forelimb have been modified to form a paddle in the whale.Almost all the bones of the hind limb have disappeared. The caudal region isgreatly developed to form a propulsive organ. (© ABRS) [M. Thompson]
5 m
120 mm
8

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
Living bone consists essentially of connective tissue fibres impregnated withmineral substances, making it almost equally resistant to compression andtension, while giving it some elasticity. The form and structure of any bone areadapted to its function of support and of resisting mechanical stresses. The drybones of an articulated skeleton give an impression of stability and tend to beregarded as forming a rigid inorganic framework, uninfluenced by the factorswhich so readily mould and modify the softer structures of the body. In fact,during growth and in adult life, each bone is continually being modified tomaintain these functions as the stresses to which it is subjected change.Alteration of shape and structure is brought about by changes in the rates ofdeposition and absorption of bone substance. The factors controlling theseprocesses are complex and not only involve activity by various cell types, butare also affected by more general hormonal, metabolic and nutritional changes.
Parts of the skeleton are made of cartilage, which is either a temporaryformation later replaced by bone, as in the epiphyses of the long bones, orpermanent and persists throughout life, as in the skeleton of the external ear andportions of the nose. Cartilage is neither as hard nor as strong as bone, but it ismuch more flexible.
The investigation of skeletal structures plays an important part in the study ofmammalian evolution. Skeletal parts are resistant to decay in manycircumstances and remain as fossils which provide the major means offollowing the course of phylogenesis.
To provide movement in an otherwise rigid framework and to allow for growth,the bony skeleton is divided into separate parts joined, as a rule, by tissueswhich are sufficiently flexible to allow movement to take place. Where thebones can move freely on each other, a joint space is present and the structure isreferred to as a synovial joint. Where less movement occurs, the joint is made byfibrous connective tissue or cartilage, or both. The union may be made completeby fusion of the bones so that movement or further growth between them isprevented, as tends to occur in the joints (sutures) of the skull vault.
Movements at joints are controlled and produced mainly by muscles which areattached to the bones (and in some instances the cartilages), but the bones areheld together by all the tissues which surround them. Bone and cartilage areeach enclosed in a dense layer of fibrous tissue, the periosteum andperichondrium, respectively, and this sleeve runs from bone to bone,constituting the fibrous capsules of joints. Here it may be thickened to formconnecting bands or ligaments which, together with the muscles and other softtissues, maintain the continuity of the skeleton and transmit tensile stresses.Compression stresses demand something more solid and the ends of bonesusually are modified in shape and enlarged to provide a good bearing surfacewhich distributes the pressure in any normal position.
Axial Skeleton
The skull reflects the several major functions of the head. Shape is determinedby genetic factors that determine such features as the size of the brain and theorgans of special sense (nose, eyes and ears), the size and form of the jaws andteeth and the details of the musculature attached to the skull. The skull consistsof bony capsules whose shapes correspond to the organs they contain, notablythe brain, eyes, nose and mouth. The irregular shape of the skull is due partly tothe multiplicity of the various capsules and passages and partly to the presenceof protruberances for muscle attachments.
There is a large number of small skull bones in non-mammalian vertebrates, buta relatively small number of large bones in mammals. A large part of the skulldevelops in cartilage as the chondrocranium: the skull base, parts of the sidewalls and capsules surrounding the nasal and auditory organs. In mammals,
9

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
nearly all the chondrocranium ossifies to form cartilage bones. Early inembryonic development, membrane bones are laid down, particularly over thetop of the brain, and most fuse with the cartilage bones.
The bones that make up the skull in eutherian mammals is consistent, althoughsome variation can occur, for example, absence of the lachrymal bone in seals.The greatest degree of skull specialisation is seen in the whales, particularly thetoothed whales, which shows telescoping and bilateral asymmetry withelongation of the rostrum and foreshortening and overlapping of the cranialbones. Paired frontal, parietal and occipital bones and often a small medianinterparietal form the vault of the skull (calvaria). The bones of the calvariacontinue down over the sides and articulate with other bones. The presphenoidforms a small section of the back of the orbit, and caudal to this is the temporalwing of the sphenoid and the squamous part of the temporal. The latter iscontinuous ventrally with the petrous part of the temporal. The caudal end of thecranial cavity is formed by the occipital bone. The floor of the cranial cavity,extending rostrally from the foramen magnum, is formed by the basal part of theoccipital and the basisphenoid with its sella turcica which houses thehypophysis. The floor is continued to its rostral extent by the presphenoid andthat portion of the ethmoid that forms the cribriform plate, through which passthe fibres of the olfactory nerve.
The inside of the cranial cavity is moulded approximately to the form of thebrain, whereas externally the surface is irregular where muscles are attached.Caudally, the skull is flattened to form a nuchal surface for muscle attachment.Prominences project ventrally from the skull, notably the mastoid process of thetemporal for attachment of the sternomastoid muscle and the jugular process ofthe occipital for attachment of muscles of the tongue and hyoid. The ridges andprocesses on the skull are formed partly as a response to the stresses applied tothe bones by muscular action.
The cranium completely encloses the brain and is pierced at its base by foraminafor the passage of cranial nerves and blood vessels.
The nasal cavities are formed dorsally by the nasal bones and lateral cartilages,laterally by the maxillary and incisive (premaxillary) bones; the last two aretooth-bearing bones. Palatine processes of the incisives and maxillaries form thehard palate that makes up the rostral part of the floor of the nasal cavity. Thisbony palate is continued caudally by the palatine bones, then the pterygoidprocess of the basisphenoid. The medial wall is made up of a caudal bony part,formed by the perpendicular plate of the ethmoid above and the vomer below,and a rostral septal cartilage.
A system of spaces, the paranasal sinuses, communicates with the nasal cavity.These sinuses are present in several bones of the skull, notably the frontal,maxilla, sphenoid and ethmoid, forming a space between the outer and innertables of compact bone, which in many bones is occupied by spongy bone(diploë). The extent and degree of complexity of the paranasal sinuses is highlyvariable, from the large and complex frontal sinuses in some Artiodactyla to theabsence of sinuses in aquatic mammals.
The confines of the orbit are entirely bony in man, but incomplete in othermammals in which only the medial wall and part of the roof are osseous. Themedial wall is formed primarily by the orbital part of the frontal bone. Theorbital wing of the presphenoid forms the caudal part and contains the opticcanal. The lachrymal bone, when present, contributes to a small portion of themedial wall and contains the fossa for the lachrymal sac and the caudal orificefor the nasolachrymal canal. The medial aspect of the roof of the orbit is formedby the zygomatic process of the frontal bone; the lateral aspect in some speciesat the rim by the frontal process of the zygomatic bone (jugal), and in other
10

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
species by the orbital ligament. Laterally the eye is protected by the zygomaticarch, made by the zygomatic bone rostrally and the zygomatic process of thetemporal caudally.
The oral cavity has incomplete bony walls and is made larger or smaller by theactions on the temporomandibular joint of muscles that attach to the skull andmandible. The bony roof of the mouth is formed by the hard palate; the wallsand floor are musculoskeletal. The muscles are attached to the maxilla andincisive bone above, to the mandible below. The incisive bone houses the upperincisor teeth (which are absent in ruminants).
The upper premolars and molars are lodged in the alveoli of the maxilla, all thelower teeth in the alveoli of the mandible. The size and form of the teeth vary(Fig. 34.4) from the simple, conical (homodont) dentition of the toothed whalesto the heterodont and highly specialised dentition of some species, for example,the incisors of rodents and the cheek teeth of herbivores and some carnivores.The dentition is diphyodont in mammals, although in some the deciduous teethare reabsorbed partially in utero and the permanent teeth formed at birth.
Each tooth develops by secretion from ameloblasts in a thickening of theectoderm, the enamel organ, in harmony with the mesodermal dental papilla, theodontoblasts of which secrete the dentine. The tooth is thus an elevation with amesodermal core, covered by two types of hard material (enamel and dentine)and set in the alveolus (socket). A third material, cementum, may be added tothe surface of the tooth. In some herbivores it extends between the folds of thetooth so as to form part of the grinding surface.
B
C
D
E
F
A
enamel
dentine
cementation
pulp cavity
Figure 34.4 Diagrams of some mammalian teeth: A, canine tooth of acarnivore; B, sectorial (carnassial) tooth of a dog; C, lower cheek tooth of ahorse; D, cross-section of A; E, cross-section of C; F, the teeth of a crabeaterseal. The incisors and canines of this seal are unremarkable, but the cheekteeth are modified to function as a sieve to filter plankton. Note illustrationsnot to scale. (© ABRS) [B. Scott]
11

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
Enamel is harder than dentine or cementum and the different rates of wear ofthese materials ensures the maintenance of a rough grinding surface or a sharp,cutting edge.
The tooth narrows at the base, but remains open in most species to allow bloodvessels and nerves to enter the pulp cavity within the root. In teeth that growcontinually, the pulp is wide open below. Usually they do not have a completecovering of enamel and are either worn away continually, as the incisors ofrodents, or grow to a great size, as the tusks of pigs and dugongs.
In some species, parts of the tooth (dentine, cementum) appear laminated insections, the layers being laid down at regular intervals. This characteristic hasbeen used to estimate the age of individuals.
The hyoid bone of mammals (Fig. 34.10) consists typically of a body, situated inthe base of the tongue, and two pairs of cornua (horns). Additional ossificationsmay reach up to the ear region and in some mammals (including man) the distalend of the hyoid horn may fuse with the skull as a styloid process.
The vertebral column of quadrupedal mammals forms the main compressionmember of a complicated girder to support their weight. The number ofcomponents comprising the girder and its flexibility differs in differentmammals, but five regions can always be recognised: cervical, thoracic, lumbar,sacral and caudal (coccygeal). In the larger quadrupeds the vertebral columnforms a double cantilever girder and the majority of the weight is carried on theforelimbs, the hind limbs being used for pushing. In some small mammals, suchas the rabbit, the vertebral column is used in two ways: when the animal springsforward it forms a single girder, braced on the hind limbs; when standing still itis a double cantilever as in the larger quadrupeds. The vertebral elements extenddorsally from the bodies and function to protect the spinal cord.
The typical vertebra contains a body that acts as a compression strut. The bodiesdevelop from a larger middle part, corresponding to the diaphysis of long bones,with which the smaller vertebral epiphyses fuse cranially and caudally duringdevelopment. The terminal faces of the body are flat (acoelous vertebrae) and afibrocartilaginous disc joins one vertebra with another and acts as a cushionbetween the bodies. These symphyses allow limited movement, particularlybending and rotation, but this is limited by the interlocking articular processes(apophyses) of the vertebrae and by the several ligaments, particularly the dorsaland ventral longitudinal and supraspinous ligaments of the vertebral column.
The base of a neural arch arises from either side of the vertebral body. Above thespinal cord the two arches fuse to form the spinous process. At the base of thearch on either side there is a transverse process. Ventral to the articular processesthere are gaps, the intervertebral foramina, for the passage of the spinal nerves.The spinous processes are longest in the thoracic region. Within the thoracicregion they are longest over the withers, although their length and slope varygreatly in different mammals.
In quadrupeds the more cranial thoracic spinous processes slope caudally andthe lumbar spinous processes point cranially with one anticlinal vertebra almostvertical between. This represents the point between the two cantilever girderswith which the vertebral column has been compared.
The number of cervical vertebrae is remarkably constant in mammals; all but avery few exceptions have seven; even an apparently neckless whale and thelong-necked giraffe have seven cervicals. The number of dorsal vertebrae(thoracic and lumbar) tends to be rather constant within the limits of manymammalian families, but the ratio of thoracic to lumbar vertebrae is variable.There are three to six sacral vertebrae, some or all of which are fused together.The number of caudal vertebrae varies greatly, from three to as many as 50vertebrae.
12

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
In ancestral tetrapods ribs were borne by every vertebra from the atlas (firstcervical) to the base of the tail. Only one type of rib is present in extanttetrapods, which probably corresponds to the dorsal ribs of fishes. Only thethoracic vertebrae have discernible ribs in eutherian mammals, butembryological investigations have shown that the so-called transverse processesof the cervical vertebrae include short, fused, two-headed ribs. The transverseforamina that are characteristic of cervical vertebrae represent the gap betweencapitulum and tuberculum of the rib in each case. Ribs are present on all thethoracic vertebrae, the capitulum of each articulating with the body of either oneor two adjacent vertebrae, and the tuberculum with the transverse process of avertebra. Most ribs curve ventrally to reach the sternum through cartilaginoussegments, but the shorter, more caudal ‘floating ribs’ do not. The lumbarvertebrae have long transverse processes but no ribs, and caudal ribs are neverdeveloped. The lumbar vertebrae have large surfaces for muscular attachment.In addition to the broad dorsal spines and transverse processes there are alsomammillary processes to which are attached the tension members of the girder,the sacrospinalis muscles.
The tail of some mammals possesses a series of small elements, crescentic inend view, which are wedged ventrally in between successive vertebral bodies.These are the intercentra, to which are fused the haemal arches (chevrons),which extend down between the muscles of either side. The base of each haemalarch consists of a pair of processes that descends from the intercentrum toenclose a space in which lie the major blood vessels of the tail. The haemalarches are particularly well developed in whales, in which the two haemalprocesses of each arch join ventral to the vessels to give a V or Y shape as seenin end view.
The first two cervical vertebrae are modified and are given the specific namesatlas and axis. The condylar articulations of the atlas with the skull are double inmammals. The dens (odontoid process) of the axis is believed to include thebody of the atlas.
Appendicular Skeleton
The mammalian limbs provide support for the body and act as levers. They arestruts with an axial skeleton and differ markedly from the limbs of fishes, whichare flaps that project laterally from the body as fins. The hind limb of mammalsis attached to the vertebral column by the sacroiliac joint. The forelimb remainsrelatively free. The scapula is attached to the axial skeleton only by muscles.
The forelimb (Fig. 34.5) is thus more flexible than the hind limb and hasacquired functions other than support and locomotion, such as digging,collecting food and climbing. The ability to move the limb in various directionsis more important than firm attachment. This ability is developed conspicuouslyin man.
The thoracic (pectoral) girdle in fishes, and to a lesser extent in amphibians andreptiles, is a complex structure. The body of mammals is raised off the groundand the thoracic girdle has assumed the function of transmitting the weight ofthe body to the limb. In so doing it has become modified and simplified suchthat it consists of two elements: the scapula and clavicle, or in many species thescapula alone. The entire coracoid plate has disappeared, there is no cranialcoracoid and the coracoid has been reduced to a small process attached to thescapula. Even the scapula has changed radically. Instead of being a simple plate,it bears the scapular spine which separates supraspinous and infraspinous fossae.The acromion is fused to the base of the scapular spine. The glenoid cavity forarticulation with the head of the humerus is a relatively shallow cup comparedwith the deep acetabulum of the hip joint.
13

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The humerus is a typical long bone, with several roughenings andprotruberances where muscles are attached, for example, greater and lessertubercles, medial and lateral epicondyles.
The bones of the antebrachium, carpus and manus vary greatly with the use towhich the forelimb is put, but all mammals possess a radius and ulna whicharticulate with a proximal row of carpal bones, which in turn articulate with adistal row of carpals and these with the metacarpals. The ulna is reduced inmany species, particularly in its distal portion. The terminology for the carpals isat times confusing and has been simplified in the Nomina Anatomica Veterinaria(NAV) (Table 34.1).
Table 34.1 Comparison of simplified carpal terminology found in the Nomina AnatomicaVeterinaria (NAV) and that commonly found in the zoological literature.
NAV ZOOLOGICAL TERM
Proximal row
(Central) Central
Radial Scaphoid
Intermediate Lunate
Ulnar Triquetrum or Cuneiform
Accessory Pisiform
Distal row
Carpal I Trapezium
Carpal II Trapezoid
Carpal III Capitate or Magnum
Carpal IV Hamate or Unciform
Figure 34.5 The bones of the distal end of the forelimb of several mammals.1. distal end of radius (and ulna where present); 2. carpus; 3. metacarpus;4. phalanges. (© ABRS) [B. Scott]
1
2
3
4
dog pig ox horse whale
1
2
3
4
1
2
3
4
1
3
2
4
1
2
3
4&
14

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
Fusion of adjacent carpal bones occurs in some species. Movement at theintercarpal and carpometacarpal joints is very limited, but in most mammalsthere is considerable movement between the metacarpals and phalanges.
Five toes were present in the manus of ancestral mammals and are found inmany forms today. The first digit (pollex) is more or less opposable to the othersin arboreal forms and man, to give a grasping power. This digit is, however, oflittle use in terrestrial locomotion and is reduced or lost in most species. Furtherdigital reduction or modification occurs in a great variety of mammals.
The hind limb shows less tendency to modification than the forelimb for specialfunctions, because it is involved mainly in propulsion and support. The weightof the hind end of the body is carried by the hind limb in a quadrupedal animalwhen it is standing. Movement is produced by extension at the hip joint with thefoot fixed, when the entire limb is used as a lever. Propulsion also is effected bylengthening of the limb by its intrinsic muscles, acting especially at the knee andtarsus in plantigrade and at the knee in digitigrade types.
The pelvic girdle articulates with one or more sacral vertebrae at the sacroiliacjoint. A small amount of movement can occur at this joint, which is a synovialjoint reinforced by short, very strong sacroiliac ligaments.
Compared with non-mammalian vertebrates, marked changes associated withaltered limb posture and musculature have occurred in mammals. The ilium,which primitively extended mainly caudally, extends cranially to articulate withthe sacrum. The pubis and ischium have moved caudally. The primitivemammalian ilium is a rather slender rod, but in heavy-bodied ungulates (forexample, horse, cow) or bipeds (man) which possess powerful gluteal muscles,this bone is expanded considerably. These three bones that make up the pelvicgirdle, the ilium, ischium and pubis, enclose an aperture, the obturator foramen,which in life is closed by the obturator membrane except for a small portioncranially that allows the passage of the obturator nerve and vessels. Theobturator membrane gives attachment to internal and external obturator musclesinside and outside the pelvis.
The pelvic opening is broader in the female in many species, associated withviviparity and in some the symphysis which joins the girdle of either side(mainly between the pubes) is loosened under hormonal control at the time ofparturition.
The acetabulum is a deep cup into which the head of the femur fits. The jointcapsule is greatly thickened in some parts to form powerful ligaments whichlimit the movements of the joint mainly to the sagittal plane.
The lines of the trabeculae in the spongy bone of the proximal end of the femurare arranged to distribute weight downwards onto the tubular compact bone inthe body. The head is attached to the body of the bone by a long neck that holdsit out at a distance from the body and permits a swinging motion of the limb.There are protruberances where muscles attach (for example, major and minortrochanter, medial and lateral epicondyles) and, distally, condyles forarticulation with the tibia and a trochlea for articulation with the patella.
The tibia is the main supporting element of the crus and is always developedfully. On the cranial aspect at the proximal end is the tibial tuberosity forattachment of the quadriceps tendon. The fibula bears little weight, and inconsequence tends to undergo reduction. It persists in almost all mammals, butits ends may fuse with the tibia, or the shaft may be lost.
In the pes, the bones of the tarsus form a structure that is somewhat similar to thecarpus. There are three bones in the proximal row, one central element and fourelements in the distal row. The terminology tends to be confusing, as in thecarpus (Table 34.2). As in the carpus, adjacent elements may be fused.
15

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The description given for the digits of the manus applies in general to the pes,but the proximal bones are termed metatarsals.
Heterotopic Skeleton
Heterotopic bones occur in certain organs, generally replacing fibrousconnective tissue otherwise present there. Bone may form directly from suchtissues or pass through an intermediate cartilaginous stage.
Small cartilages or bones form in several tendons of the body in mammals, atpoints of potential friction. Termed sesamoids, these frequently occur in themanus or pes. The patella is an unusually large sesamoid structure.
The os penis (baculum) is formed in the fibrous tissue of the corpora cavernosain bats, rodents and carnivores, among other mammals. Its shape is the mosthighly varied of all bones. A homologous cartilaginous or bony os clitoridis ispresent in some species.
Ossa cordis develop in the fibrous skeleton of the heart of deer and bovids.
THE MUSCULATURE
Mammals possess an elaborate system for receiving information from theenvironment. This system is linked inextricably to correspondingly elaborateeffectors, the muscles which respond to motor signals received from the nervoussystem. The muscles can transfer static energy (derived from food and oxygen)into kinetic energy (movement) in response to nerve stimuli. The activity of thenervous system has little mode of expression other than the contraction ofmuscle fibres. The musculature constitutes one third to almost one half of themass of an individual mammal.
Muscle Function
Movement is achieved by an articulated framework, the skeletal parts, joined byties that can be varied in length, the skeletal muscles. The muscles serve to fix aswell as to move the skeletal parts. They can act as braces, holding the organs ofthe body in place.
Table 34.2 Comparison of simplified tarsal terminology found in the Nomina AnatomicaVeterinaria (NAV) and that commonly found in the zoological literature.
NAV ZOOLOGICAL TERM
Proximal row
Talus Astragalus
Calcaneus Calcaneum
Central
Central tarsal Navicular
Distal row
Tarsal I Medial or Internal Cuneiform
Tarsal II Intermediate or Middle Cuneiform
Tarsal III Lateral or External Cuneiform
Tarsal IV Cuboid
16

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
Muscles operate by exerting tension. If the ends of a muscle are fixed,contraction exerts a force along its length without any actual movementoccurring and is known as isometric contraction. If a muscle is allowed toshorten without increase of tension, the contraction is said to be isotonic. Inpractice the two types of contraction are unlikely ever to be fully separated.
The power of holding is developed most fully in the unstriated (smooth) muscleof the viscera. This muscle produces slow movements such as peristalsis.Unstriated muscle occurs in the wall of the gut, respiratory passages, urogenitalducts and blood vessels, in the skin and the iris. Myofibrils within the cells arethought to be the contractile material of unstriated muscle. They are doublyrefractile under the polarizing microscope, but show no sign of the alternatingisotropic and anisotropic transverse bands that are characteristic of themyofibrils of striated skeletal muscle.
Contraction of unstriated muscle may be initiated by autonomic nerve impulses,hormonal stimulation or local changes arising within the muscle itself.Stretching the muscle fibres can change the membrane potential and initiate awave of contraction, a local stimulus that is particularly important in thephysiology of the bladder, gastrointestinal tract and other hollow viscera.
Unstriated muscle in the walls of blood vessels behaves rather like striatedskeletal muscle in that its activity usually is initiated by motor nerve fibres andthere is little evidence of conduction between cellular units. In contrast, visceralunstriated muscle functionally resembles cardiac muscle to some extent: thecells behave like single muscular units and impulses are freely conducted fromcell to cell. Two forms of contraction occur in visceral smooth muscle, namelyrhythmic and tonic contraction, which appear to be independent; in addition,there is a continuous state of partial contraction known as muscle tone.
Cardiac muscle is found only in the heart wall. It shows cross striations similarto that seen in skeletal muscle, but in other respects cardiac muscle differsgreatly from skeletal muscle. The heart musculature is a continuous network ofdividing and recombining strands with prominent cross bands, intercalateddiscs, separating this network into short units. The heart beat is influenced byautonomic nerves, but the cardiac musculature is essentially autonomous.
The majority of movement in mammals is performed by the striated skeletalmuscles which also play an important part in posture. Their fibres are muchlarger than those of unstriated muscle and contain many nuclei. The mostobvious characteristic of skeletal muscle fibres (Fig. 34.6) is the transversestriations that make up the contractile units; a more delicate longitudinalstriation also is detectable within the fibre, attributable to the parallelarrangement of myofibrils.
Large numbers of parallel muscle fibres are grouped into fasciculi, visible to thenaked eye, which are arranged into various patterns to form several types ofmuscles - straplike, sheetlike, unipennate, bipennate, multipennate.
When a muscle contracts at its maximum power, it is capable of contracting toabout half its stretched length. The force of contraction of a given muscle at anytime depends on the number of fibres that are contracting. The contraction isinitiated by a nerve impulse travelling over a nerve axon to the muscle fibres.Each axon supplies several muscle fibres, the neuromuscular unit comprising amotor unit. The number of motor units functioning at any one time determinesthe activity of the muscle. The precision of movement is greatest in thosemuscles that have many motor units, each of which includes only a few musclefibres, for example the extrinsic muscles of the eyeball.
Pennate muscles, that is those with tendons throughout their length, are strongerthan straplike or sheetlike types because they are composed of many short,oblique fibres which have an additive effect upon the insertion. The straplike
17

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
and sheetlike types contain long fibres that are almost parallel with one anotherand with the long axis of the muscle. They have the ability to contract to agreater degree than pennate muscles, but with less strength.
The muscle fibres, the fasciculi and the entire muscle are each invested byconnective tissue (endomysium, perimysium and epimysium, respectively) thatforms a continuous stroma serving to bind together the contractile units andgroups of units and to integrate their action. It also permits a degree of freedomof movement between them such that each fibre is independent of adjacentfibres to some degree and each fasciculus can move independently ofneighbouring fasciculi. The connective tissue framework of muscles, therefore,is vital to normal muscle function. The epimysium of some muscles isindistinguishable from the more extensive investing fascia, a connective tissueforming fibrous membranes which separate organs and invest them. Fascia alsoforms extensive sheets, such as the extensive sheet of fascia that connects theskin with deeper structures. In relation to the muscles, sheets of dense fasciaprovide attachments, serve as elastic sheaths, form specialised retaining bands(retinacula) and fibrous sheaths for tendons and permit the gliding of one muscle
Figure 34.6 Diagrams showing the structure and mechanism of slidingfilaments in striated skeletal muscle: A, four within a single muscle cell athigh magnification; A band = anisotropic band; I band = isotropic band; B,the arrangement of the filaments of actin and myosin when the muscle cell isstretched (above) and contracted (below). (© ABRS) [M. Thompson]
A
B
18

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
on another. The development of fascia to form an extensive, highly elastic sheet,termed the tunica flava abdominis, is seen in the abdominal wall of largeherbivores such as the horse.
In the more distal parts of the limbs, especially in the lower limbs of humans,venous return is impeded by gravity and aided by muscular contraction. Themuscles are prevented by the fascia from swelling and bulging excessively withblood during contraction. The fascia serves as a resistant stocking and makesmuscular contraction more efficient in moving blood upward. This importantfunction of the fascia is enhanced by its attachment to the bones in some regionsto form fascial compartments that surround groups of muscles with similarfunction and nerve supply.
The attachment of muscles to bone or other tissue is usually by a long, cord-liketendon (sinew) or by a broad, relatively thin aponeurosis. Where they areattached to bone, the bundles of collagenous fibres fan out in the periosteum.Although some muscles appear to have an extensive, fleshy attachment to bonewith no interposing tendon or aponeurosis, there is always some connectiveelements between the muscle fibres and the attachment of the muscle.
The Contractile Mechanism
Detailed examination of the submicroscopic organisation of muscle by electronmicroscopy and X-ray diffraction has led to a new concept of the mechanism ofmuscle. contraction
The cross-banded pattern of striated skeletal muscle (Fig. 34.6) reflects thearrangement within the myofibrils of two sets of submicroscopic filaments,thicker myosin filaments and thinner actin filaments. The explanation of theirfunction necessitates some preliminary description. The fibrils bear cross-striations which tend to be in register within a muscle fibre, giving the entirefibre its distinctive cross-banded appearance under the light microscope. Eachfibril is made up of alternating light and dark portions. The former are onlyslightly bi-refringent, the isotropic or I bands, the latter are strongly bi-refringent and, therefore, are termed the anisotropic or A bands. Cutting acrossthe I band is a thin dark line, the Z line, and in the middle of the A band a thinclear line is sometimes seen, the H band.
When a muscle contracts, the thick and thin filaments maintain the same length,but slide past each other so that the ends of the actin filaments extend into the Aband, narrowing and ultimately obliterating the H band. As a consequence of thedeeper penetration of the A band by the actin filaments, the Z line is drawncloser to the ends of the adjacent A bands and there is an overall shortening ofthe myofibril. Calcium is required in the tissue fluid for muscular contractionand it is believed that neural excitation of the muscle fibre membrane isconducted inwards by tubules that are continuous with the membrane, causingthe endoplasmic reticulum to release calcium ions to the myofibrils andtriggering their contraction. When contraction is completed, calcium ions arerecaptured by the endoplasmic reticulum and relaxation occurs.
The energy required for contraction is provided ultimately from the glycogenabundant in muscle fibres. The rapid release of energy within the musclenecessary for contraction is effected by the conversion of adenosine triphosphate(ATP) to adenosine diphosphate (ADP). The energy for rebuilding ATP isderived from the oxidation of glycogen.
Muscle Action and Homeostasis
The chemical processes within functioning muscle result in the production ofenergy, some of which is in the form of heat. Mammals make use of this in thecontrol of body temperature. If cooled blood reaches the temperature-regulating
19

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
centres of the central nervous system, shivering, which consists of muscularcontraction with the production of heat, may result. Non-shiveringthermogenesis also may involve the muscles, when specific mechanisms forrelease of heat energy are involved in response to a fall in blood temperature.
Vigorous exercise helps to maintain body temperature in cold conditions by therelease of heat from working muscles.
Muscular Organisation
Several terms are used to describe muscles according to their action. Anextensor muscle tends to open out a joint, a flexor closes it. An adductor draws asegment toward the median plane of the body, an abductor does the reverse. Alevator raises while a depressor lowers the structure. Constrictor or sphinctermuscles surround orifices and tend to close them. Dilators do the opposite.
The two major trunk muscle groups (Fig. 34.7) are the epaxial and hypaxialgroups. The epaxial musculature, which in fishes is a massive column ofsegmented muscle lying lateral to the vertebrae, is in the main restricted to thechannel along the vertebral column between the spinous processes and thetransverse processes. It comprises three longitudinal divisions, with little of theoriginal metameric arrangement. Laterally is the iliocostalis system, medial towhich the longissimus system lies dorsal to the transverse processes of thevertebrae. Most medially, between the longissimus and the spinous processes, isthe transversospinalis system. In the lumbar region the iliocostalis andlongissimus unite into a powerful sacrospinalis. The epaxial musculature servesto extend the vertebral column, and to flex it laterally when acting on one side.Parts of it are capable of producing more subtle movements of rotation, flexionand extension between neighbouring vertebrae.
A
B
Figure 34.7 The superficial A, and deep B, musculature of the dog. (©ABRS) [M. Thompson]
20

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The hypaxial muscles of fishes lie ventral to the horizontal septum and extendaround the outer body wall to the ventral midline. They are reducedconsiderably in relative bulk in mammals. Some of them form muscle sheetsaround the belly and flanks and, aided by the rib cage, enclose and support thethoracic and abdominal cavities and their viscera. The hypaxial trunkmusculature includes a series of subvertebral muscles, ventral to the transverseprocesses and the ribs. They function broadly to flex the vertebral column and,with the epaxial muscles, laterally flex it when they act together on the one sideonly. Among these muscles are the psoas muscles, quadratus lumborum and thelongus colli. The hypaxial flank muscles follow the curve of the trunk from thetransverse processes of the vertebrae to the ventral region and include theexternal and internal abdominal oblique and transverse abdominal muscles.Ventrally, the rectus abdominis runs craniocaudally from the sternum and costalcartilages to the pelvis. Thin, transverse, tendinous inscriptions often are presentin the rectus, which may represent to some degree the original segmentalcondition of the muscle. The hypaxial muscles that attach the forelimb to theaxial skeleton form slings that suspend the body between the scapulae. Theymainly extend from the vertebral border of the scapula to the ribs. The muscleswhich form those slings are derivatives of the external oblique muscle. Themajor elements of the system spread out from the deep surface of the scapula ina fanwise manner to attach caudally to the ribs and cranially to the cervicalvertebrae. They include the serratus ventralis, levator scapulae and the moredorsally placed rhomboideus.
The development of the diaphragm is a characteristic of mammals. It is apartition, partly muscular and partly tendinous, separating the thoracic andabdominal cavities and is of major importance in respiration.
There is a basic pattern of six extrinsic muscles of the eyeball in vertebrates,namely the four rectus muscles (dorsal, ventral, medial and lateral) and the twooblique muscles (dorsal and ventral). As in the majority of tetrapods (birds andprimates are exceptions), there is a retractor bulbi which tends to pull the eyeballdeeper into its socket in almost all mammals. The levator palpebrae superioris isderived from the eyeball muscles, but attaches to the upper eyelid and, in somecases, to the plica semilunaris conjunctivae (third eyelid).
In the head, the facial skin muscles are commonly referred to as the muscles offacial expression. They have a special importance in the primates because theyplay a large part in social organization. The jaw arch musculature of mammalscomprises the muscles of mastication: the temporalis, masseter and medial andlateral pterygoids. Ventrally, a group of muscles attaches to the hyoid bone andacts as an elevator and depressor of the hyoid and, indirectly, the larynx. Themain depressor of the mandible is the digastric muscle.
The limb muscles of mammals are bulky and complex. Early in the developmentof the limb, a mass of premuscular tissue is formed on both the dorsal and theventral surfaces of the developing limb skeleton. These two masses areequivalent to the opposed dorsal and ventral muscle masses which raise andlower the paired fins in fishes. From the dorsal mass arise all the complexmuscles on the extensor surface of the limb and related muscles of the girdleregion. From the ventral mass arise the flexor muscles of the opposite limbsurface. The reader should refer to an anatomy text for a description of theindividual muscles of the limbs that are responsible for movement by flexion,extension, abduction, adduction, rotation, supination or pronation of the wholeor components of the limb. It should be borne in mind, however, that a givenmuscle almost never has a single function. The action of a muscle is restrainedand modified by several factors, such as the action of other muscles and whetherthe limb is fixed or swinging freely. The action of living muscles can bedetermined by electromyography.
21

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
THE BODY CAVITIES
The body cavities are large serous sacs, the pericardial, pleural and peritonealcavities. They are derived from one continuous cavity, the coelom, whosecranial end surrounds the heart during embryonic development to form thepericardial sac. This is connected caudally to the peritoneal sac by twopericardioperitoneal canals, one on either side of the midline. The peritoneal sacbecomes reduced by the developing abdominal viscera while thepericardioperitoneal canals accommodate the developing lungs and form thepleural sacs. These different parts of the coelom are soon separated off fromeach other. The large common cardinal veins carrying blood to the heart encirclethe openings from the pericardial sac to the pleural sacs and finally occludethem altogether. Slightly later, the developing diaphragm cuts off the pleuralsacs from the peritoneal cavity. In the male, a fourth subdivision of the coelom isformed by herniation of part of the peritoneal sac which surrounds the testicle inthe scrotum. This becomes separated off as a closed sac, the vaginal cavity orcavity of the tunica vaginalis.
The body cavities thus become, after development of the viscera, emptypotential cavities the lining membrane of which is reflected over the viscera.The viscera, thereby, become covered by a visceral layer of the serousmembrane which is continuous, often through mesenteries, with the parietallining layer which lines the body wall. This suspension of viscera into thepotential space of the cavities permits free movement of the viscera. The smoothsurface of the visceral layer of the serous membrane can glide freely over theopposed parietal layer, lubricated by a small amount of fluid secreted by theserous membrane. The serous membranes lining the pericardial and peritonealcavities and the pleural sacs are referred to as serous pericardium, peritoneumand pleura, respectively.
THE DIGESTIVE SYSTEM
Animals possess a mechanism that provides a ‘drive’ to seek food. The source ofthe food drive, or how it relates to the needs of the organism, are not wellunderstood, but in mammals it is strong indeed. In most species a pattern of foodintake at intervals, that varies with the species, develops during early postnatallife. Probably it is triggered partly by hunger contraction (movements of theempty stomach) and partly by complex memories within the central nervoussystem.
It is remarkable that animals seek out and obtain the raw materials they requirefor their particular metabolic needs. Chemoreceptors probably play a large partin finding the right food, olfaction acting as a distance receptor and taste servingto test the food at the point of intake. Other senses may play a part as well, suchas vision and, in bats and whales, sonar. Whether any given stimulus leads toacceptance or rejection of an object as food probably depends largely on pastexperience as recorded in the cortex.
Digestion
The features of the digestive system that are peculiar to mammals occur mainlyin the mouth, where the dry exterior meets the moist interior at themucocutaneous junction of the lips. The teeth, tongue and enzymes of the salivabegin the physical and chemical process of breaking up the food. The rest of thealimentary canal completes this process and absorbs from the products themolecules needed by the individual.
22

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
Chemical digestion is accomplished by the enzymes secreted by the alimentarycanal and its associated glands and the action of symbiotic micro-organismsliving within the gut. The high temperature of the body assists the fermentationprocess, which in herbivorous species is extensive and highly specialised inforestomach of ruminants and the large intestine of rabbits, horses and dugongs.Food is mixed and moved along the gut by rhythmic peristaltic movementsregulated by neural reflexes and chemical (hormonal) signalling. Gut motilityalso is influenced by centres in the medulla oblongata, hypothalamus andcerebral cortex.
The Mouth
Food is taken into the mouth in various ways, in herbivores for example, by thecutting action of the incisors as in the horse and rabbit or by the prehensileaction of the tongue as in cattle. Some herbivores have reduced anterior teeth,for example the ruminants, but the hard palate may be ridged such that the actionof a rough tongue against it aids in breaking up the food. Production of vastquantities of saliva in certain herbivores, particularly the ruminants, also assiststhe initial breakdown of the food. The ducts of three major salivary glands, someor all of which are present in mammals, open into the mouth. These are theparotid, mandibular and sublingual glands. The nature of their secretions differssomewhat. The tongue bears numerous papillae of various forms (filiform,fungiform, vallate, conical, lentiform, foliate, marginal) on its surface. Tastebuds occur in the epithelium of some papillae. The filiform papillae aredeveloped to function in grooming in the cat.
The food is held in the mouth and masticated at length by herbivores and may bemasticated more than once as in the ruminants. Carnivores tend to swallow thefood quickly, tearing and chewing only when it is necessary to render the foodportions small enough to swallow. The teeth have been considered in an earlierSection (see Section on Axial Skeleton).
The Oesophagus and Stomach
The oesophagus is a simple tube through which the food passes within a fewseconds. The oesophageal musculature is partially striated and partiallyunstriated. Nonetheless the food is pushed along its entire length by theinvoluntary process of peristalsis. Food reaches the oesophagus by the voluntaryact of swallowing, which involves raising the tongue and then contracting thelevator muscles of the soft palate and superior pharyngeal constrictor, thusclosing off the nasopharynx. The food passes through the oral and laryngealparts of the pharynx and into the oesophagus by the stripping action of thepharyngeal constrictor muscles and the simultaneous raising of the hyoid andclosing of the laryngeal aditus (Figs 34.8 & 34.9).
Portions of the stomach develop into enormous fermentation vats in theruminants (Fig 34.8). Although the rumen, reticulum and omasum often aretermed the forestomachs and are lined by stratified squamous epithelium like theoesophagus, they develop embryologically from the primitive stomach. Thefourth compartment, the abomasum, is similar morphologically and functionallyto the simple stomach of other mammals. Digestion of plant materials,particularly cellulose, occurs in the forestomachs, with breakdown to short chainfatty acids that form an important source of energy in herbivores. Considerablevolumes of fluids are reabsorbed through the wall of the omasum.
The glandular portion of the stomach has a lining of tall columnar mucus-secreting cells, folded to form gastric pits, into the bases of which open the ductsof glands. The histological structure and the secretion products differ in differentparts of the stomach. Some food, particularly liquid material, passes through the
23

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
stomach within minutes, while the more solid food may remain in the stomachfor up to four hours. In the glandular stomach the ingesta is acted upon by pepsinwhich in the acidic environment breaks down proteins into peptones andproteoses. These molecules are not absorbed until they have been further brokendown in the intestine. Little food is absorbed from the stomach, but under someconditions small molecules may be. Milk clots in the stomach, aided in newbornmammals particularly by the production of the enzyme rennin.
The secretion of the gastric glands is controlled partly by reflex action, whoseneural afferent pathway may be the sight or smell of food or the presence of foodin the mouth or stomach itself. In addition the presence of food in the stomachcauses the liberation into the blood of a hormone that stimulates secretion ofgastric juice.
The Small Intestine
Food passes from the stomach into the intestine in liquid condition (chyme).This passage is controlled by the pyloric sphincter between the stomach andduodenum. The reaction of this region is only faintly acidic, and in addition toenzyme digestion in the intestines, intestinal microflora continue the process offermentation. Although there are certain distinguishing features of the differentparts of the small intestine (duodenum, jejunum and ileum), the food is mixedwith the several enzymes responsible for breaking down all the chief classes offoodstuffs and absorption of the products begins along its length. Absorption iseither direct into the blood stream, when metabolites pass in the hepatic portalsystem to the liver, or into the lacteals when the products (mainly the products ofdigestion of fats) pass indirectly to the bloodstream via the lymphatics. Certainhormones are passed into the blood from the small intestine, although the exactcellular source of some of them is unknown. These hormones influence thefunctions of the pancreas, gall bladder, intestinal glands and the vasculature.
As the chyme enters the duodenum it is mixed with pancreatic juice, bile fromthe liver and secretions from the glands of the duodenal wall. The chyme isrendered less acid and its composition is altered rapidly by the digestion process.
In general the small intestine tends to be relatively longer in herbivores thancarnivores, presumably associated with fermentation of plant materials. Butthere are exceptions, as in the carnivorous Elephant Seal and Sperm Whalewhich have remarkably long intestines.
Figure 34.8 The appearance of the simple stomach of the dog comparedwith the complex stomach of the ox. Fermentation occurs mainly in the firstcompartment of the stomach of the ox. The first three compartments arereferred to collectively as the forestomachs, while the fourth compartment isglandular. 1 = rumen; 2 = reticulum; 3 = omasum; 4 = abomasum. (© ABRS)
[M. Thompson]
40 mm200 mm
24

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The Liver and Pancreas
These are large glandular structures whose prominent ducts empty theirsecretory products into the duodenum. The pancreas has both exocrine andendocrine components. The endocrine function is the production of insulin,important in glucose metabolism.
The secretion of the pancreatic juice, the exocrine component of pancreaticfunction, is controlled both by nervous and hormonal means. Food present in theduodenum stimulates the production of secretin and pancreozymin by theduodenum and these hormones in turn stimulate the release of pancreatic juice.Parasympathetic (vagal) stimulation also results in pancreatic secretion into theduodenum. Several digestive proenzymes are elaborated by the pancreaticalveoli, the chief derivatives being trypsin, amylase, lipase and the enzyme-likerennin.
The liver is a vital organ with many functions, which may be grouped under fourheadings: (a) those concerned with metabolism of the food and storage; (b)secretion and excretion; (c) activities concerned with composition of the blood;(d) protective and detoxicating activities. Its cells are primarily digestive infunction.
The actions of the hepatic cells on the food depend on the condition of theanimal at the time. For example, if amino acids are needed for building the body,they are passed directly into the general circulation to required sites. But in theadult, as a rule more amino acids are taken up from the intestine than are neededfor building, and the excess is de-aminated in the liver, the nitrogen beingconverted to urea which is excreted by the kidney, while the remaining parts ofthe molecules are available for energy production. Similarly, carbohydrate eithermay be passed on from the liver or converted into glycogen, in which form it isstored in the liver. Vitamins, particularly A and B, are also stored in the liver, asare enzymes, hormones and fat. Fat is usually stored in a masked form; it canarise from carbohydrates and be transformed into carbohydrates.
The secretory product of the liver is the bile. We may consider the columns ofhepatic cells as tubular glands, whose secretory products are released into thebile canaliculi. The latter join to form major bile channels that eventually unite
A
B
Figure 34.9 Theappearance of the largeintestine of a carnivore,the dog (A) and aherbivore, the horse (B).The horse has a simplestomach and the veryextensive large intestineacts as a fermentationchamber. (© ABRS)
[M. Thompson]
25

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
to form the hepatic duct. In some species there is a diverticulum off the mainhepatic duct, the cystic duct leading to the gallbladder, but a gallbladder islacking in the rodents, horses, certain ruminants and Cetacea.
Bile is released from the bile duct (the duct resulting from the joining of thehepatic and cystic ducts) into the duodenum. The bile salts aid in theemulsification of fats during intestinal digestion, thereby rendering the fatsvulnerable to the action of lipase, and are reabsorbed in part by the intestine andre-used. Bile pigments, the breakdown products of haemoglobin through theactivity of reticulo-endothelial cells, are excreted via the bile duct into theduodenum and eliminated in the faeces. Cholesterol, lecithin and fats also areexcreted via the bile.
Many substances that have a pronounced action when introduced into the body,such as alcohol and anaesthetics, are broken down in the liver.
The Large Intestine
The contents of the caecum and colon are nearly neutral and contain manymicro-organisms. Their action on the colonic contents, especially on proteins,produces a variety of putrefactive products. In simple-stomached herbivoressuch as the horse, rabbit and dugong, there are other micro-organisms in thisregion that play an important part in breaking up refractory foodstuffs,particularly complex carbohydrates, such as cellulose. An important function ofthe large intestine is the absorption of water from the gut contents.
THE RESPIRATORY SYSTEM
The high energy requirements of a mammal, resulting from the warm-bloodedcondition and high level of activity, are met ultimately by the oxidation offoodstuffs by oxygen taken in from the atmosphere. The apparatus of thethoracic cavity and respiratory tree ensures that a supply of oxygen from the airis available continually.
An oxygen debt may be accumulated for a short time during powerful musculareffort, but if the tissues are without oxygen for more than a few minutes theybecome impaired permanently.
Gaseous exchange between the air and the blood occurs in the terminal portionsof the respiratory tree, mainly the alveoli. This arrangement is made to functionby the enclosure of the lungs in a chamber, the thoracic cavity, whose walls canbe expanded by muscular contraction, causing air to be drawn in.
The Nasal Apparatus
The nasal passages (Fig. 34.10) function in olfaction, to moisten and warm theair before it passes to the lungs and to filter dust and other particles from the air.The nasal passages are modified greatly in the totally aquatic mammals that lackolfactory sense and have no special structures for moistening inhaled air. The airinspired immediately above the water surface in these species probably issaturated.
The olfactory function is considered later (see Section on Olfaction). Themoistening and warming of the air is accomplished by the extensive, richlyvascularised epithelium of the delicate nasal conchae. These are particularlylarge and intricate in polar mammals. The mucous membrane lining the nasalcavity and covering the conchae is a ciliated, mucus-secreting epithelium tofilter out small particles.
26

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
A further system of spaces communicates with the nasal cavity, the paranasalsinuses, whose lining mucous membrane is continuous with that of the nasalcavity. The function of these sinuses is not understood fully, but they areparticularly well developed in flying vertebrates, the birds and bats, but absentin aquatic mammals.
The Larynx and Phonation
The larynx functions as a vestibule to the trachea, as a valve preventing theentrance of food into the respiratory tree and in phonation. The laryngeal cavityis surrounded by a complex of cartilages. Associated muscles adjust thelaryngeal cartilages and open and close the glottis. The entrance to the larynxfrom the pharynx, the aditus, can be closed by the action of muscles and byfolding the epiglottis back over it.
Voice production is accomplished by the presence of a pair of vocal cords. Thesecords consist of ridges of mucous membrane containing an elastic ligament,stretched across the larynx. The two cords can be set in vibration by the passageof a current of expired air between them and their relative positions arecontrolled by the action of the intrinsic laryngeal muscles.
Trachea and Lungs
The trachea is continuous with the larynx, and consists of a tube whose walls aresupported by incomplete rings of cartilage that prevent collapse duringinspiration. At its caudal end the trachea divides into the bronchi. Division ofeach bronchus occurs and subsequent division leads to the bronchioles, fromwhich a system of irregular alveolar ducts leads to the terminal alveoli. Theeffect of this branching system of tubes is to allow air to penetrate to everyportion of the lung.
The walls of the alveolus are excessively thin. This allows a very closerelationship between the air in the alveolus and the blood in the capillaries of thealveolar walls.
Figure 34.10 Sagittal section through the head of a cat, showing the nasalconchae in the lateral wall of the nasal cavity and related structures. Scaleapproximates actual size. (© ABRS) [M. Thompson]
27

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The walls of the alveoli and bronchioles contain many elastic fibres, which givethem the power to contract during expiration. Thus, expiration is largely apassive process. The walls of the bronchioles also contain unstriated musclewhich receives motor innervation from the vagus and inhibitory sympatheticinnervation.
The trachea, bronchi and large bronchioles are lined by ciliated cells, which beatin such a way as to transport particulate matter towards the pharynx and protectthe respiratory tree.
Respiratory Exchange
Only a partial change of air in the lungs occurs with each breath. The lungs arenot emptied completely during each expiration, but much of the alveolar air isswept into the bronchioles and mixed with fresh air at the next inspiration. Thegases in the alveoli come into rapid equilibrium with the blood by diffusionacross the thin alveolar walls, as oxygen is taken up by the plasma and from thisby haemoglobin in the red blood cells, while carbon dioxide is given up to theblood.
Under the conditions in which haemoglobin occurs in the blood, it becomes 95%saturated with oxygen at the partial pressure of oxygen in alveolar air (100 mmmercury). The haemoglobin remains saturated to as much as 80% if the oxygentension is reduced to 50 mm Hg, but below this tension it begins to give upoxygen. This occurs normally in the capillaries. Here haemoglobin issurrounded by an environment poor in oxygen and it yields its oxygen to theplasma, then through the capillary wall to the tissues. The form of the oxygendissociation curve depends on the composition of the blood.
The uptake of oxygen in the lungs is often termed external respiration, incontrast to the process of oxidation within the tissues, internal or tissuerespiration. Tissue respiration takes place by a system of enzymatic changes,making the oxygen capable of undertaking chemical transformations that wouldnot otherwise occur under these conditions.
Control of Respiration
The regular rhythm of respiration is produced by the action of the respiratorycentre in the medulla oblongata. Afferents reach the centre via the vagus nervefrom receptors in the walls of the alveoli. Cells in the region of the respiratorycentre are sensitive to carbonic acid in the blood. Receptors in the aorta, carotidbodies and atria are sensitive to the pressure and oxygen content of the bloodand they influence respiration. Some vagal afferents influence the respiratorycentre directly.
There is, thus, an elaborate system of chemical and neural influences acting onthe respiratory centre. Other receptors are present that respond to tactile andchemical influences and effect the cough reflex.
THE CIRCULATORY SYSTEM
Mammals possess a closed circulatory system. Blood never leaves the vesselsthat contain it and, therefore, never comes into direct contact with the cells ofthe body as occurs in many non-vertebrate animals. The cells of the tissues arebathed in interstitial fluid and the blood serves the tissues by diffusion ofmaterials through the small, thin-walled capillaries. Blood is pumped by theheart, through the arteries to the capillaries, and returns to the heart through theveins.
28

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
To maintain a high and constant body temperature, mammals need to transportlarge quantities of fuel for combustion and oxygen with which to burn it, to thetissues. Circulation needs to be rapid and adjustable to meet the changing needsof the animal; it would be inefficient to maintain a maximum circulation ofblood to all parts of the body simultaneously.
Functions
As well as transporting fuel and oxygen to the cells (via the interstitial fluid),circulation is the means by which waste products from combustion are removedfrom the tissues. The tissues must be supplied constantly with oxygen from thelungs and food materials from the intestine or from storage and manufacturingcentres, notably the liver. Waste products must be removed continually, carbondioxide is taken to the lungs for removal, nitrogenous wastes and excessmetabolic water are excreted by the kidneys.
Another important function of the circulation is the maintenance of the steadyinternal environment necessary to permit efficient functioning of the cells.Uniform composition of the interstitial fluids is maintained by constantcirculation of liquid throughout the body. Circulation plays a vital role in themaintenance of a steady body temperature.
The circulatory system aids in the struggle against disease and in the repair ofinjuries. It carries substances (the hormones) produced in one part of the bodythat affect others.
Blood is a connective tissue, consisting of cellular components floating freely ina liquid matrix. The fluid fraction is plasma, whose salt content is approximatelythe same as that of the interstitial fluid. The plasma contains, in addition, theblood proteins albumin, globulin and fibrinogen. These proteins consist of largemolecules unable to pass through the capillary walls and whose presence(particularly the albumins) raises the osmotic pressure of the blood above that ofthe interstitial fluids, a point of importance in capillary function. Fibrinogenmakes up only a small percentage of the total blood protein, but it is importantwhen converted to fibrin in blood coagulation. One globulin type takes part inthe formation of the blood clot; other specific globulins are sensitive to ‘foreign’proteins and will, for example, cause the agglutination of red blood cells ofanother species. Certain globulins are active agents (antibodies) in protectionagainst disease.
The blood cells have various functions, the most obvious is the constanttransport of oxygen by the erythrocytes (red blood cells). This is achieved by thepresence of haemoglobin, a protein molecule which includes in its complexstructure four iron atoms, each capable of carrying an oxygen ion. Haemoglobinis also a factor in the transport of carbon dioxide from the tissues. The structureof the erythrocyte is unusual in mammals. It is round rather than oval (the camelis an exception) and the mature red blood cell lacks a nucleus, a fact thatpresumably increases the efficiency of oxygen carriage in the total mass of theerythrocytes.
The leukocytes, or white blood cells, are far fewer in number than theerythrocytes. Leukocytes are able to leave the blood stream to play an active rolein connective tissues. Two major groups of leukocytes are distinguishable, theagranulocytes and the granulocytes. The common forms of agranulocytes are thelymphocytes, present in all classes of vertebrates, which play an important rolein immunological protection of the organism. The other type of agranulocyte,the monocyte, may be merely a large lymphocyte.
29

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The granulocytes include acidophils (eosinophils), basophils and neutrophils.They seem to be essentially inert while in the blood stream, but are capable ofamoeboid movement and in this way move out of capillaries between theendothelial cells to accumulate rapidly in injured, inflamed or infected tissues.
A further cellular component of the blood is the thrombocytes (platelets), whichtake part in the process of blood clotting.
Haematopoiesis
Proliferation and differentiation of blood cells continues throughout life. Thelife of an individual blood cell is short, a few months at the most in the case oferythrocytes, days or even hours for leukocytes.
In mid-foetal life and almost up to the time of birth, the liver is concernedactively with haematopoiesis. Blood cells also are produced in the foetal spleen,which is actively haematopoietic in the later stages of foetal life. Towards theend of intrauterine life most haematopoietic activity shifts to the red marrow ofbones, although the agranulocytes continue to be produced by the spleen andlymph nodes throughout life. Much of the red marrow of the bones is replacedby fatty tissue (yellow marrow) during postnatal life, but in certain regions thecancellous tissue of the bones persists as actively haematopoietic tissuethroughout life, particularly the ribs, vertebrae, sternum, skull bones and pelvis.The yellow marrow may be replaced again by red marrow under exceptionalcircumstances, such as in certain types of anaemia. In extensive destruction ofthe bone marrow (for example, in myelofibrosis), the liver and spleen mayrevert to their former haematopoietic functions.
The thymus is a source of rapid production of lymphocytes during late foetaland early postnatal life (see Section on the Lymphoid Organs).
The Heart and Major Vessels
There is complete separation in the mammals of the systemic and pulmonarycirculation. By this means, oxygenated blood can be delivered to the capillariesat a far greater pressure than can be achieved in a plan such as that of the fish.There the blood passes first through the gill capillaries before passing to thecapillaries of the body, under the influence of a single pressure source in theheart. A particular advantage of the arrangement in mammals is that quick andsustained movements are possible because fuel and oxygen can be transportedefficiently to the muscles.
The separation of the systemic and pulmonary circulations has occurred byseptation of the heart to form four chambers and concurrent septation of theventral aorta. The pulmonary trunk, therefore, arises from the right ventricle,and the aorta arises from the left ventricle.
The arteries become progressively smaller as they branch in their peripheralcourse from the aorta and pulmonary arteries. The sum of the luminal diametersof all branches also increases progressively, so that the rate of blood flowthrough the peripheral vessels steadily diminishes as they get farther away fromthe heart.
The structure of the arterial wall varies to a considerable extent in relation to thesize of the vessel. In the larger arteries, elastic tissue predominates in the middlecoat. The elastic tissue permits them to expand with each cardiac systole andrecoil with each diastole, a pulsation which continues to drive the bloodperipherally. As the vessels get smaller, the blood flow is converted graduallyfrom an intermittent series of propulsions derived from the rhythmic contractionof the heart to a steady and continuous stream. No pulsation is evident in thecapillaries under normal conditions.
30

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The presence of unstriated muscle in the smaller arteries enables them, under theinfluence of the autonomic nervous system, to contract or dilate and so regulatethe distribution of blood to the areas they supply.
Veins commonly run with arteries. Medium arteries often are accompanied bytwo veins (venae comitantes), although in some parts of the body, the brain forexample, there is no close association between arteries and veins. Most veins arecharacterised by the presence of valves arranged at intervals along their courseand so disposed as to direct the blood flow towards the heart. The flow of bloodalong veins is primarily dependent on a pressure gradient, the pressuredecreasing from the periphery to the heart. This is the result of pressure changesrelated to the cardiac cycle and of the varying negative pressure in the pleuralsacs produced by respiration. Muscular action further assists the venouscirculation in peripheral vessels (see Section on Muscle Function).
In some regions of the body, particularly the limbs, veins course superficially inaddition to the axial veins accompanying the arteries. These superficial veinshave no accompanying artery, but anastomose freely with the deeper veins. Theyserve a thermoregulatory function by permitting heat radiation from the skinsurface when they are engorged with blood. Countercurrent heat exchange canoccur between the arteries and accompanying veins, serving a heat conservationfunction.
Supplementary to the venous system is a second series of vessels that returnsfluids from the tissues to the heart, the lymphatics. They are not connected withthe arteries, but arise from their own capillaries which end blindly. There is,thus, no arterial pressure behind the fluid in the lymphatics and flow in them issluggish. The intestinal lymphatics (lacteals) are important in the uptake of fatsfrom the intestine.
The lymphatics are most highly developed in tetrapods. The functionalexplanation is that because the blood circulates at a higher pressure than infishes, the entrance of tissue fluids into the capillaries is more difficult. Thelymphatics offer a relatively low pressure system of drainage into the great veinswhere the blood pressure is at its lowest.
Regulation of the Circulation
The rate of the heart beat varies greatly among mammals. Larger mammals havea lower rate than smaller ones. It also varies with ‘emotional’ and other factors,with activity and training. There are receptors in the heart, in the great bloodvessels and in the lungs which influence the rate of the heart beat.
Afferent impulses which arise in the aortic body and pass in the vagus result in alowering of blood pressure by action through a centre in the medulla oblongata.The aortic body is stimulated when the blood pressure is raised. Other receptors,the carotid body and carotid sinus, possess cells sensitive to changes of oxygenand carbon dioxide content of the blood as well as to pressure changes.
In the peripheral circulation, an elaborate system of instructions ensures thateach part of the body receives an adequate supply of blood according to itsfunctional state. When any tissue, such as muscle or intestine, becomes activethere are corresponding alterations in its blood supply produced through theautonomic nervous system under the overriding control of the central nervoussystem.
THE URINARY SYSTEM
The major functions of the urinary system are the regulation of the internalenvironment and the elimination of wastes. Although some products ofmetabolism such as lactic acid can be used again, others, such as urea, usually
31
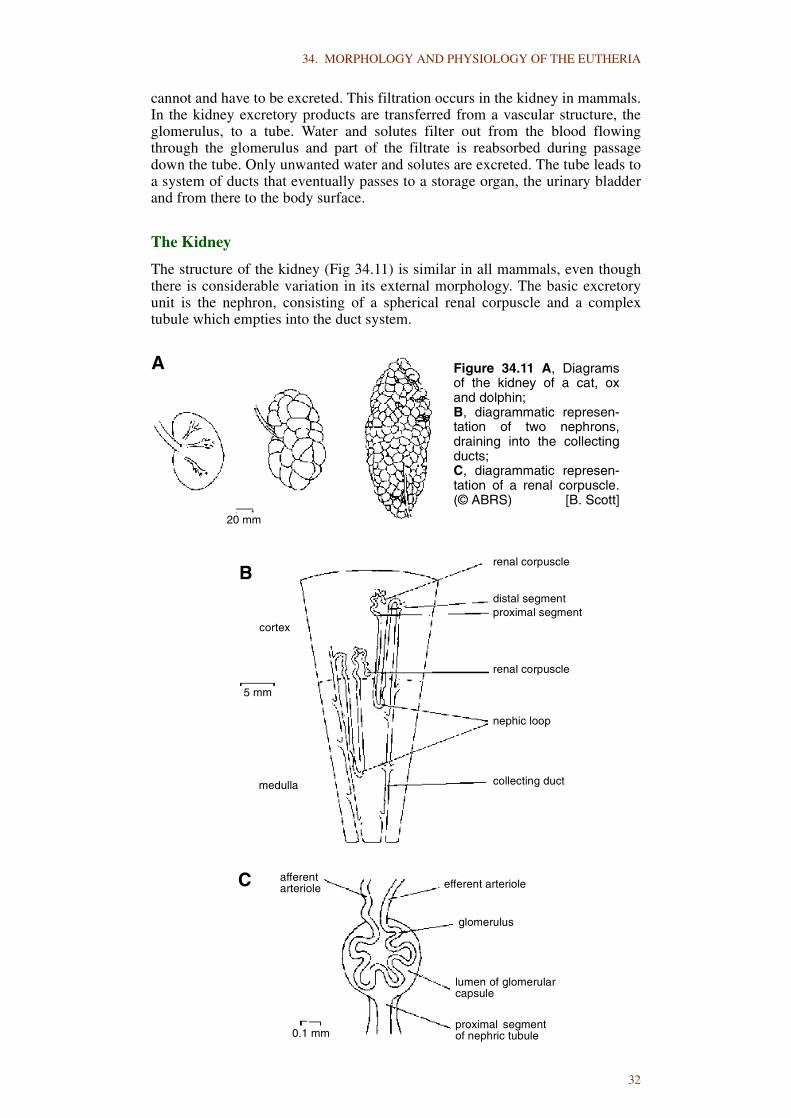
34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
cannot and have to be excreted. This filtration occurs in the kidney in mammals.In the kidney excretory products are transferred from a vascular structure, theglomerulus, to a tube. Water and solutes filter out from the blood flowingthrough the glomerulus and part of the filtrate is reabsorbed during passagedown the tube. Only unwanted water and solutes are excreted. The tube leads toa system of ducts that eventually passes to a storage organ, the urinary bladderand from there to the body surface.
The Kidney
The structure of the kidney (Fig 34.11) is similar in all mammals, even thoughthere is considerable variation in its external morphology. The basic excretoryunit is the nephron, consisting of a spherical renal corpuscle and a complextubule which empties into the duct system.
renal corpuscle
distal segmentproximal segment
renal corpuscle
nephic loop
collecting duct
cortex
medulla
5 mm
20 mm
efferent arterioleafferentarteriole
glomerulus
lumen of glomerularcapsule
proximal segmentof nephric tubule0.1 mm
A
B
C
Figure 34.11 A, Diagramsof the kidney of a cat, oxand dolphin; B, diagrammatic represen-tation of two nephrons,draining into the collectingducts; C, diagrammatic represen-tation of a renal corpuscle.(© ABRS) [B. Scott]
32

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The renal corpuscle consists of two parts, a glomerulus and a glomerularcapsule. The glomerulus is a compact cluster of small blood vessels; the capsuleis a proximal part of the tubule, whose shape is that of a double-layeredhemisphere. The inner part is closely adherent to the vessels of the glomerulus,so that only thin membranes separate the blood stream from the lumen betweenthe two parts of the capsule. This lumen is continuous with that of the renaltubule, a long structure with distinguishable proximal and distal segments.Along its course the tubule is bordered closely by a network of capillary vessels.Groups of cells adjacent to the glomerulus, referred to as the juxtaglomerularcells, produce the hormone renin, which raises arterial blood pressure.
Filtration and Reabsorption
The result of the activity of the renal corpuscle and the renal tubule is theproduction of urine. Urine is composed mainly of water, but contains to avariable degree other substances in solution. These include salt ions, particularlysodium, potassium, chloride and sulphate, and waste materials, mainly simplenitrogenous compounds generally in the form of urea.
The cells of the body require among other things an environment containing, insolution in the interstitial fluids surrounding them, appropriate amounts andproportions of specific simple salts. The maintenance of proper salt content inbody fluids demands a proper balance between intake and output. The renaltubules furnish the main means of eliminating excess salts, or of removingexcess amounts of liquid which tend to dilute the body fluid.
As blood flows through the glomerulus under pressure, a filtrate of it passesthrough the internal part of the renal capsule into the capsular lumen. Apart fromblood cells and large molecules such as the blood proteins, all other materialsincluding food materials such as glucose as well as wastes, pass readily throughinto the capsular lumen. The amount of fluid filtered is excessive. Duringpassage of this filtrate along the renal tubule, the volume is reduced greatly andthe composition changes as a result of reabsorption. Most of the water isreabsorbed and food materials (mainly glucose) are withdrawn in the proximalpart of the tubule. Salt is reabsorbed more distally. Further active secretion ofwaste substances occurs through the tubule cells into the urine.
Ureter and Bladder
The ureter and bladder have a muscular wall and a characteristic liningepithelium, the transitional epithelium. This type of epithelium presumablypermits a considerable increase in area when stretched, especially in the wall ofthe bladder. The bladder has thick coats of unstriated muscle. Continualrhythmical peristaltic contractions force the passage of urine along the uretersinto the bladder. The terminal portion passes obliquely through the bladder walland acts as a sphincter to prevent reflux. The proximal end of the ureter isexpanded to form the pelvis in some, but not all, mammals and portions of itabut against the kidney parenchyma to form the calyces. The renal collectingducts open into them.
The muscle fibres in the bladder wall run in many directions and are able, likeother unstriated muscle fibres, to adjust their length so that with distension of theorgan the pressure increases only slightly.
As the bladder fills with urine, receptors in the wall are stimulated andultimately by reflex action they produce the rhythmical contractions of themuscle which, together with relaxation of the sphincters of the urethra, lead tomicturition. The muscle is controlled by autonomic nerves, parasympathetic
33

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
stimulation resulting in contraction of the bladder muscle and relaxation of thesphincter; sympathetic stimulation has the opposite effects. Striated muscle inthe urogenital diaphragm serves as a sphincter under voluntary control.
Control of Excretion
The secretion of the adrenal cortex is necessary for the absorption of salts fromthe renal tubules. The amount of water reabsorbed is controlled largely by theinfluence of antidiuretic hormone (ADH) secreted by the neurohypophysis(posterior pituitary). The release of ADH is under the control of thehypothalamus, in which there are receptors sensitive to the degree of dilution ofthe blood. Excretory control is complex and is influenced by many otherendocrine and metabolic factors.
THE REPRODUCTIVE SYSTEM
The reproductive system includes the ovary and testis that produce the gametes,the ducts by which the gametes are carried to the exterior and the copulatoryorgans which aid in internal fertilization of the eggs. Important to thereproductive process are the secondary sexual characters which provide signalsto ensure that the sexes meet and pair, the mammary glands and specialbehaviour mechanisms for the care of the young. The whole reproductivesystem is coordinated by an elaborate system of chemical communication,centred upon the hypothalamo-hypophyseal axis.
The Male Reproductive System
The testis of mammals is peculiar in that in nearly all species it descends into thescrotal sac. In some mammals, particularly many rodents, the testes descend intothe scrotum during the reproductive period, but are withdrawn into theabdominal cavity at other times by the action of the external cremaster muscle.In other mammals (testiconda) the testes are retained permanently in the bodycavity. Some placental mammals appear to have been always testicond, whereasothers may have returned secondarily to this condition.
The testis consists of a number of long, convoluted seminiferous tubules whosewalls produce the spermatozoa. Throughout spermatogenesis the formativegerm cells remain attached to the nutritive Sertoli cells. The testis is subject toseasonal change, which in seasonal breeders may involve the cessation ofspermatogenesis during non-breeding, but it generally does not exhibit suchmarked differences between breeding and resting stages as does the ovary.
Between the seminiferous tubules of the testis there is a considerable amount ofconnective tissue in which are found collections of interstitial cells that producetestosterone.
Spermatogenesis is a cyclic phenomenon in most vertebrates, but theseminiferous tubules are permanent structures that produce successive crops ofspermatozoa and in most species change little throughout the breeding life of theindividual.
The spermatozoa of different mammals differ markedly in morphologicaldetails, but all have a head bearing the nucleus and acrosome and a flagellum(tail) consisting of a connecting piece and intermediate, principal and terminalparts.
The scrotum is derived from the scrotal sacs, paired pouches in the floor of theabdominal cavity lined with coelomic mesothelium. In eutherian mammals thesacs fuse ventro-caudal to the penis to form the scrotum.
34

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The spermatozoa are carried away by a system of tubes derived from themesonephric ducts. The seminiferous tubules open into the rete testis, fromwhich the spermatozoa pass into the efferent ducts, the first part of the long,coiled epididymis. The morphologically mature sperms become functionallymature in the epididymis. The tail of the epididymis is continuous with theductus deferens, a muscular tube that ascends through the inguinal canal andcurves medially within the pelvic cavity to open into the urethra, but just beforedoing so its expanded end, the ampulla, is joined by the duct from the seminalvesicle. The combined duct thus formed, the ejaculatory duct, pierces theprostate before opening into the urethra. Paired bulbo-urethral glands open intothe urethra below the urogenital diaphragm. The secretions of all these glandsare mixed with the spermatozoa during ejaculation and among other functionsserve to activate and nourish the sperms. Not all the accessory sex glands namedabove are present in all mammals and others, notably the rodents, have otheraccessory sex glands.
The penis consists of a tube containing three vascular erectile columns, thecorpora cavernosa, one on each side, and the corpus spongiosum ventrally. Theterminal part of the urethra is surrounded by the corpus spongiosum. Eachcorpus cavernosum is attached to the ischium of the same side. Erection isproduced by relaxation of the walls of the arteries that lead to spaces in theerectile tissues so that they become engorged with blood.
The Female Reproductive System
The female genital tract of the mammal is built on the plan of other vertebrates,being derived from a pair of paramesonephric ducts leading from the coelom tothe exterior.
The ovaries develop from the embryonic gonads. At the indifferent stage a seriesof primary sex cords is formed. These persist in the testis to form the convolutedseminiferous tubules, but degenerate in the developing ovary to be replaced bysecondary sex cords. The germ cells contained in these secondary sex cordsdivide repeatedly and develop eventually by the complex process of oogenesisinto definitive oocytes. Each oocyte becomes surrounded by a cluster of othercells derived from the sex cords to form a follicle, which is the major organ ofoestrogen production. As maturity is approached the follicle enlarges anddevelops a liquid-filled space in its interior.
Follicles ripen at the surface of the ovary, continuously in some mammals, butseasonally in others, and burst, releasing the oocyte into the surroundingcoelom. This is the process of ovulation. The oocyte is then taken up by thegenital tract. The follicular cavity that remains becomes filled with a yellowcellular mass, the corpus luteum, which secretes progesterone. Many moreoocytes are present in the ovary than are released by ovulation; a largeproportion fail to reach maturity and degenerate.
The ovaries are usually paired, but one or the other fails to reach full maturity insome species.
Since the mammalian ova are small, the upper part of the female genital tract,the uterine tube, is a slender structure. The portion that receives the oocyte fromthe coelom is enlarged to form a funnel-shaped end, the infundibulum. Thisbears finger-like processes (fimbria), and a ciliated epithelium, the action ofboth of which aid in directing the oocyte, surrounded by cells from the follicle,into the genital tract. Fertilization usually occurs in the upper portion of theuterine tube.
The uterus is a thick-walled muscular structure within which prenataldevelopment occurs. The uterine mucosa or endometrium, a vascularepithelium, is relatively thin during resting phases, but is thickened greatly at
35

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
times of reproductive activity. Its rhythms of growth and involution arecontrolled by ovarian hormones. In the most primitive mammalian situation thetwo uteri are separate and known as uterus duplex. This condition is present inmany rodents and bats. In most eutherians, however, the more caudal parts ofthe two uteri are fused to a greater or lesser extent to form a bicornuate uterusand in higher primates there is complete fusion to form a uterus simplex. Thecaudal end of the uterus narrows to form a thick-walled canal, the cervix, whichmay be paired (uterus duplex) or single (bicornuate uterus and uterus simplex).The whole uterus lies in a fold of peritoneum, the broad ligament.
The terminal portions of the two paramesonephric ducts invariably are fused ineutherians to form a vagina. This opens into the urogenital sinus, which in manyspecies appears like a continuation of the vagina. The exterior opening isprotected by the labia which enclose the major derivative of the urogenital sinus,the vestibule. The urethra opens into the vestibule, as do the ducts of the majorvestibular glands. Ventrally in the vestibule is the clitoris, which contains pairederectile corpora cavernosa as in the penis. Paired, erectile vestibular bulbs arepresent in the labia.
The Placenta and Fetal Membranes
The placenta which affords maternal nourishment of the developing embryo andfetus, is formed by a union of the uterine mucosa with the fetal membranes. Thelatter include the chorion, amnion and allantois, but the involvement of these inthe formation of the placenta varies greatly and influences to some extent thetype of placentation that occurs. The yolk sac plays a part in development of theplacenta in some species as well.
As the early conceptus (blastocyst) grows it may lie free in the uterine lumen,the entire surface of its trophoblast in contact with the uterine mucosa. In otherspecies the blastocyst remains attached only at the site of first contact, leaving afree surface projecting into the uterine lumen. In others, including man, theblastocyst is buried in the uterine mucosa and becomes surrounded by maternaltissues.
Intimate contact between part or all of the surface of the trophoblast with theuterine mucosa early in prenatal life results in the development of the placenta.On the embryonic side, blood vessels that develop in the chorion (that is, thetrophoblast plus its associated extraembryonic mesoderm) connect withembryonic vessels by contact with the wall either of the yolk sac or the allantois.
Certain areas of the surface of the trophoblast develop villi, which penetrate intothe uterine mucosa, thus increasing the absorptive surface of the chorion andproviding intimate contact between the fetal and maternal tissues. In somespecies, for example perissodactyls and the Pig, there develops a diffuseplacenta in which the chorionic villi are distributed nearly evenly over thesurface of the chorion. In a placenta multiplex (cotyledonary placenta) as seen inruminants, the chorionic villi are grouped in numerous patches, the cotyledons,which are separated by smooth chorion. Carnivores have a zonary placenta: thevilli are restricted to a band around the middle of the chorionic sac. A discoidalplacenta forms in primates, rodents and other species, where the villi are limitedto a disc-shaped area on the chorion.
The closeness of contact between the fetal and uterine parts of the placentavaries in different eutherian groups. With the outgrowth of villi from the surfaceof the chorion, corresponding crypt-like depressions develop in the uterineplacenta into which the villi become embedded. The most superficial cell layersof the uterine mucosa are destroyed by specific activity of the trophoblastic cellsand the nutrients thus freed (histotroph) conveyed to the embryo. This process isparticularly marked in some species and reduces the tissue remaining betweenthe uterine and fetal capillaries of the placenta. The number of layers remaining
36

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
between the two circulations has been used in a histophysiological classificationof placentas. Thus, there occur epitheliochorial, syndesmochorial, endothelio-chorial, haemochorial and haemoendothelial types of placentas.
A further classification of placentas is based on the loss of uterine mucosa(decidua) at the time of parturition. With deciduate placentas (carnivores,primates and others) there is loss of uterine mucosa at birth because of a moreintimate fusion of fetal and maternal tissues. In the simplest non-deciduateplacentas (horses, pigs, ruminants), maternal and fetal parts are merely inapposition and little or no maternal tissue is lost.
Control of Reproduction
Coupling of males and females is ensured by an elaborate system of signals,receptors and effector mechanisms. The mutual attraction of males and femalesfor the purpose of mating is brought about largely by olfactory signals in manymammals, but visual signals also play a large part and in some species auditorysignals are involved. Some of the secondary sexual characters serve to providesigns for recognition and attraction.
Mating behaviour is influenced by the internal condition of the individual,mainly by the action of the endocrine system on the nervous system. Seasonallybreeding mammals are sensitised by the hormones at certain times of the year,associated with environmental changes such as alteration of day length, so thatthey respond to stimulation by the secondary sexual characters of the oppositesex.
The production of the gametes, both oocytes and spermatozoa, is under intricateand finely-balanced hormonal control. Three protein hormones secreted by theadenohypophysis under hypothalamic control, the gonadotropins, regulateovarian functions. Follicle stimulating hormone (FSH) causes ovarian folliclesto grow and produce increased amounts of oestrogens. Luteinizing hormone(LH) causes ovulation and initiates the formation of the corpus luteum. Prolactinmay have a luteotropic function in some species. The ovary is capable ofproducing oestrogens from cells surrounding the follicles and progesterone fromthe corpus luteum. A third hormone, relaxin, may be produced by the ovary also.
The oestrogens produce oestrus. Small amounts of progesterone may also play arole in the induction of oestrus in some species. The oestrogenic hormonescontrol the development of all the female secondary sex characteristics and playa major role in development of the mammary gland. They also have major rolesin modifying production and rates of release of hypophyseal gonadotropins.
Progesterone is the most important hormone of pregnancy. Implantation cannotoccur in its absence; it acts synergistically with oestrogens to develop the uterineglands characteristic of the oestrous cycle and early pregnancy. Progesteroneplays a vital role in maintaining pregnancy. The placenta produces someprogesterone, in some species sufficient to maintain pregnancy in its latterstages in the absence of corpora lutea. This hormone is believed to beresponsible for inhibiting ovulation during pregnancy and pseudopregnancy.
The factors responsible for the initiation of parturition are not understood fully.Parturition is controlled both by hormonal influences and the degree ofdistension of the uterus. A constant relationship exists among litter-bearinganimals between the size of the uterus and the total size of its contents, despitevariations in litter size and the size of individuals making up the litter. Aremarkable series of hormonal changes occurs prior to and during parturitionand, although there seems to be variation among species, in general thesechanges include a rise or fall (depending on the species) of plasma progesteroneand oestrogen and a rise in prolactin and corticoids. Relaxin concentration
37

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
remains high until after parturition, but LH remains consistently low or isabsent. The rise or fall of progesterone is accompanied by a reciprocal fall orrise in oestrogens.
Implicated in triggering these abrupt hormonal changes, at least in certainspecies, is an increased output of fetal adrenal corticoids in response toincreased release of adrenocorticotrophic hormone (ACTH) from the foetaladenohypophysis. Prostaglandins and oxytocin seem to act directly on theuterine tunica muscularis (myometrium). Uterine contractions may be initiatedby prostaglandins and continued by the action of oxytocin during parturition.
Androgen secretion by the interstitial cells of the testis is controlled by theadenohypophysis, the hormone LH is the primary one involved. In addition,FSH stimulates the production by the Sertoli cells of an androgen-bindingprotein that is secreted into the testicular fluid. Prolactin also appears to have asynergistic effect with LH in maintaining testosterone production.Spermatogenesis and spermiogenesis in the seminiferous tubules are controlledby the gonadotropic hormones FSH and LH, either directly or indirectly,because normal spermatogenesis can no longer occur after hypophysectomy.
In most mammals, testicular temperature remains several degrees below bodytemperature. The scrotum helps to regulate testicular temperature, important inthe production of viable sperm, by two specialised mechanisms. The bloodentering the testis in the long, coiled testicular artery is precooled bycountercurrent heat exchange with the cooler venous blood returning in thepampiniform plexus. Temperature is regulated to some degree by contractionand relaxation of the striated external cremaster muscle and the unstriatedmuscle in the scrotal wall that constitutes the dartos. Exposure to cold results inthese muscles drawing the testis closer to the body wall where the temperature ishigher.
The accessory sex glands of the male are all dependent upon androgen for theirnormal functioning. Androgen also is responsible for the secondary sexcharacters in the male.
THE NERVOUS SYSTEM
There is an elaborate system of receptors from which information about theenvironment is transmitted to the spinal cord and brain. New information iscombined with information stored there and translated into action by effectororgans, especially the muscles. The nervous system, therefore, is the main agentregulating activity, its actions control almost every aspect of mammalianbehaviour.
The nervous system in vertebrates arises from a well-organised dorsal tubularstructure that enlarges cranially to form the brain. The remainder develops toform the spinal cord. These constitute the central nervous system. Runningoutward from the spinal cord and brain are numerous paired nerves along whichoccur clusters of nerve cells forming ganglia in specific regions. These nervesand ganglia constitute the peripheral nervous system.
The functional cellular units of the nervous system are the neurons, large cellsthat reside in the central nervous system and the ganglia, with long processes,the nerve fibres. The fibres include the dendrites which receive signals from theenvironment or from other neurons and transport them to the cell body and theaxons that carry signals away from the cell body. Neurons differ greatly inshape, especially in the length of the dendrites and axons and their method ofbranching. The neurons are able to conduct impulses because they have thecapacity to give discharges that serve as signals. This capacity depends on thefact that the surfaces of the cells are electrically-charged systems that aresensitive and readily discharge when a small change occurs near them. The
38

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
setting up of the charge by the cell depends upon chemical processes involvingexpenditure of energy and the discharge is probably initiated by molecularchanges at the surface followed by movements of ions. These movements set upelectrical currents that initiate discharge of neighbouring parts of the nerve fibreand thus make possible the transmission of signals.
The chemicals involved in the transmission process originate in the body of theneuron and flow outward to supply the various processes.
Nerve cells that are destroyed during postnatal life are not replaced, although thefibres may regenerate if the cell body remains intact.
Functionally, the most important part of a major nerve fibre is the neuronalprocess itself, the axis cylinder. This is wrapped almost always by a series ofcells which form the neurolemma. These cells are capable of producing a myelinsheath of fatty material surrounding the fibre.
The distance between the sensory organ (receptor) and a muscle or glandularstructure stimulated (effector) is never spanned by a single neuron in mammals.The action takes place through a chain of neurons, at least two and usually more.The point of transfer between one neuron and the next is the synapse. Minuteamounts of chemical material are produced at the terminal branches of the axonand diffuse across to stimulate the dendrite of the second neuron. The chemicalsubstance most generally concerned is acetylcholine, but there are manytransmitter substances, some known, some unknown. This is currently an area ofvery active research.
The simplest nerve action is the reflex arc, the type of ‘automatic’ reaction suchas the withdrawal of a paw that has touched a very hot object. A sensorystimulus is picked up from receptor cells and carried toward the central nervoussystem by a long dendrite that constitutes the afferent fibre. The cell body of thesensory neuron to which this fibre belongs lies in the dorsal root ganglion, at thelevel of the intervertebral foramen. The afferent fibre is continued as the axon ofthe neuron from the cell body to the central nervous system. Normally it doesnot synapse on a single successive neuron, but branches so that a stimulus maybe produced in a whole series of neurons. Conversely, each neuron with which itconnects may receive impulses from numerous afferent fibres, so thatconsiderable interplay between receptors and effectors may take place. In thissimplest reflex the neurons stimulated by incoming impulses may be efferent,usually motor neurons. Their cell bodies lie within the central nervous systemand long axons run from them to the effector organs (usually muscle fibres). Buteven a simple reflex arc is slightly more complex than this and is a three-neuronrather than a two-neuron chain. The afferent fibres entering the central nervoussystem synapse with neurons contained entirely within the central nervoussystem. These internuncial neurons send out branched axons that synapse withnumerous motor (efferent) cells. This further multiplies the number of possibleresponses which a sensory impulse may cause and, conversely, also increasesthe number of sensory impulses which might produce any given motor effect. Inthis case the effect on the motor neuron may be inhibitory rather thanstimulatory, tending to counter stimuli this cell may have received from othersources. Even at this relatively simple level of nervous activity, there are‘feedback’ mechanisms which increase complexity.
Spinal and Cranial Nerves
The so-called peripheral nervous system, which includes the spinal and cranialnerves, penetrates to almost every region of the body.
Typical spinal nerves are paired and present in every body segment. Each nervearises from the spinal cord by dorsal and ventral roots which unite and leave thevertebral canal as the spinal nerve before dividing into dorsal and ventral rami.
39

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The spinal nerve and its rami carry both afferent and efferent fibres, but the tworoots show a sharp division of function. The ventral roots convey mainlyefferent impulses and, therefore, are mainly motor, while the dorsal roots carryafferent fibres and thus are sensory. The cell bodies of the latter neuronsgenerally lie in the dorsal root ganglion.
Although the cranial nerves lack the regular arrangement of the spinal nerves,they subserve similar functions and convey afferent impulses from the head andneck (and from the trunk in the case of the vagus), as well as efferent impulses tothe muscles of the head, neck and in one case (the trapezius) the back. Unlikethe spinal nerves, however, the cranial nerves do not all carry all components.The first, second and eighth cranial nerves are special sensory nerves consistingonly of afferent fibres conveying to the central nervous system information onlyabout smell, sight and hearing and balance, respectively.
The processes of peripheral nerves grow out from the central nervous system orthe ganglion and reach their end organs: sensory structures, muscle fibres orgland cells. Although there is a degree of specific association in that efferentfibres do not attach to sensory structures nor afferent fibres to muscle cells, thereare no specific affinities. That is, a particular nerve will not always attach to agiven end organ.
Both afferent and efferent fibres can be divided functionally into two types,somatic and visceral. Somatic afferents carry impulses from the skin(exteroceptive) and sense organs of muscles and tendons (proprioceptive) to thecentral nervous system, whereas visceral afferents carry sensations from sensoryendings in the internal organs (enteroceptive). Somatic efferents are the motorfibres to the striated muscles, while visceral efferents carry stimuli primarily tothe unstriated muscles and glands in various regions. These four components arepresent in most spinal nerves, but this is not true of some cranial nerves. Thespecial somatic afferents have been mentioned above.
The Autonomic Nervous System
Whereas the somatic part of the nervous system controls those organs of thebody in contact with the outside world, the autonomic nervous system controlsthe internal workings of the body. Nonetheless, these outer (somatic) and inner(visceral) components of the nervous system work together in a coordinatedway. When, for example, the muscles of the trunk and limbs are activated by thesomatic nervous system, the autonomic system influences the heart and bloodvessels to adjust the flow of blood to the muscles. In this way, it ensures thatsufficient oxygen and fuel are carried in the blood to the muscles to permit theirincreased activity.
Sensory endings are present in the internal organs and blood vessels. Visceralafferent fibres pass from these to the central nervous system through autonomicnerves of either the sympathetic or the parasympathetic system.
The autonomic impulses which pass to the unstriated muscles and glands of thebody are carried from the central nervous system along a two-neuron chain ofefferent fibres, the visceral efferents. The first neuron, termed the preganglionicneuron, has its cell body in the central nervous system and its axon is typicallymyelinated. At some point along its course it enters a ganglion of the autonomicnervous system, where it synapses on a postganglionic neuron whose axoncompletes the passage to the end organ.
The autonomic nervous system comprises the sympathetic and parasympatheticsystems, which to a large extent are distinguished anatomically, physiologicallyand pharmacologically. Preganglionic outflow of the sympathetic system isconfined to the thoracic and lumbar parts of the spinal cord. Sympatheticstimulation tends to increase the activity of the animal, speed up the rate of heart
40

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
beating and increase the circulation, slow down digestive processes and, ingeneral, make it fit for ‘fright, flight or fight’. The tips of the nerve fibres releasenoradrenalin and adrenalin.
Preganglionic outflow of the parasympathetic system is from the brain and viacertain cranial nerves (particularly the vagus) and the sacral part of the spinalcord. The action of the parasympathetic system tends to slow down the activityof an animal in general but promotes digestion and a relative rest phase. Thefibres release acetylcholine at their ends.
The Spinal Cord
The spinal cord contains, as it did in the embryo, a central canal containing fluid,but in contrast to the embryonic structure it is relatively small in the adult, owingto the growth of nervous tissue surrounding it. The spinal cord is shorter than thevertebral column in mammals, so that in the caudal part of the vertebral canalthere is merely a series of nerves passing caudally from cord segments to thecorresponding intervertebral foramina.
Two distinct layers can be distinguished (even with the naked eye) in the cord, acentral area of grey matter surrounding the central canal and white mattersurrounding the grey matter.
The grey matter consists mainly of cell bodies of neurons, arranged into an H-shape. The dorsal and ventral projections of the ‘H’ are termed the dorsal andventral horns, respectively. Cell bodies of the efferent (motor) neurons of thespinal nerves are present in the ventral horns. Their axons typically pass out inthe ventral roots of successive spinal nerves. Most of the motor supply is tosomatic muscles, but in the thoracic and lumbar parts of the cord preganglionicsympathetic efferents are situated dorsolateral to the somatic motor neurons in alateral horn.
The dorsal horn is associated with the dorsal, sensory roots of the spinal nerves.Here is the seat of the cell bodies of association neurons through which afferentimpulses brought in from sense organs may be relayed and distributed.
The white matter is composed of myelinated fibres, including ascending anddescending fibres of sensory nerve cells, fibres which carry sensory stimulicranially to the brain and fibres returning from brain centres to act on motorneurons. Topographically, the white matter is subdivided into dorsal, lateral andventral funiculi, in which are found relatively discrete fibre tracts which carryinformation up and down the spinal cord.
The Brain
There is a considerable degree of concentration of command over bodilyfunctions in the brain of mammals. Intermediate neurons connecting the spinalcord with the brain tend to gather in functional centres (nuclei) in many parts ofthe brain and fibres with like connections tend to assemble into definite bundles(tracts). The nuclei range in size from microscopic clusters of cells to the entirecerebral cortex. In the nuclei afferent impulses may be correlated and integratedfor appropriate responses or motor mechanisms coordinated. At higher levelsthere may develop association centres whose activities are responsible for suchphenomena as memory, learning and consciousness.
A primitive system persists in mammals in the form of the reticular formation.Rather than discrete nuclei and tracts, this is a band of interlacing cells andfibres associated with the motor columns of the brain stem and cranial spinalcord. It is important in motor coordination and in carrying stimuli down from
41

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
higher brain centres to the motor centres of the medulla oblongata and the spinalcord. It also acts as an agent which tends to promote activation of the highercentres of the cerebral cortex.
The brain is largely bilaterally symmetrical and, therefore, cross connections arenecessary. These occur in commissures, fibre tracts which connectcorresponding regions of the two sides, and decussations of tracts.
In general, the complexity of the brain increases from caudal to rostral regions.The medulla oblongata and, to a lesser extent, the pons are structurallysomewhat similar to the spinal cord, with a series of nuclei of grey matterreminiscent of the grey horns of the cord, but their gross appearance is different.Dorsally, the central canal is greatly expanded to form the fourth ventricle, theroof of which is thin and membranous. Rostrally, it is covered by thecerebellum. The series of nuclei of the somatic motor column breaks down inthe pons and midbrain into several small nuclei related to movements of theeyes.
The series of motor and sensory nuclei of the medulla oblongata provide all theelements required for reflex circuits in the brain for the head region, with theexception of most of the special senses. In mammals, acoustic nuclei persist asdistinct centres for the vestibular and cochlear parts of the ear in the brain stemat about the level of junction of the pons and medulla oblongata, in the region ofthe acoustico-lateralis area of more primitive vertebrates.
The cerebellum, a large structure in mammals (particularly man), functions inthe coordination and regulation of motor activities and the maintenance ofposture. It acts in a passive, essentially reflex fashion in the maintenance ofequilibrium and body orientation. In addition, it plays a major positive role inlocomotion. Associated with these functions are the large, paired, convolutedcerebellar hemispheres which make up much of the bulk of the organ inmammals.
The midbrain and diencephalon (Fig. 34.12) show specialised functionalfeatures in mammals as in other vertebrates. Sensory and motor nuclei of moreposterior origin extend some distance into the midbrain, but such structures areabsent in the rostral midbrain and diencephalon. In general, the rostral brainstem centres have no direct connections outside the central nervous system otherthan via the optic nerve. The region of the midbrain and diencephalon,particularly the latter, is a principal relay station between lower brain areas andthe cerebrum. Most auditory and other somatic sensations that reach themidbrain are relayed by way of the thalamus to the cerebrum.
The tectum (roof) of the midbrain serves in a limited way as a centre for visualand auditory reflexes.
The tegmentum, the region of the midbrain between the tectum and the cruscerebri on each side, is essentially a rostral continuation of the motor areas of thepons and medulla oblongata. As such, it functions as a region in which variedstimuli from the diencephalon, tectum and cerebellum are coordinated andtransmitted to the motor nuclei of the pons, medulla and spinal cord.
The main importance of the hypothalamus is as a major integrative centre forvisceral functions. It has fibre connections with the autonomic centres of thebrain stem and cord. Stimuli from olfactory and taste organs as well assensations from various visceral structures pass to hypothalamic nuclei. Thehypothalamus controls the activity of the hypophysis to a considerable degreeand some hypothalamic nuclei secrete hormones which pass to the hypophysis.Functions such as thermoregulation, heart beat, respiratory rate, blood pressure,sleep and gut activities are controlled or influenced by the hypothalamus.
42

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
All sensory material (except olfaction) is relayed to the cerebral cortex via thedorsal thalamus. Other areas of the thalamus are associated with coordination ofmuscle function.
Much of the cerebrum of mammals is represented by the neopallium, a greatlyexpanded cortical area for correlation, association and learning. This areaassumes the greater part of the higher functions that in lower forms areconcentrated in the tectum or corpus striatum, gains a complete array of somaticsensory information through projection fibres from the thalamus and developsdirect motor paths to the motor columns of the brain stem and spinal cord. Amassive commissure, the corpus callosum, connects the left and right neopallialstructures, permitting both hemispheres to share memory and learning. Thesurface of the neopallium of eutherian mammals is thrown into folds (gyri),between which are grooves (sulci).
Supporting Elements of the Nervous System
Most of this Section on the nervous system has dealt with the neural elements ofthe nervous system, although the sheaths of nerve fibres have been mentioned.Other types of cells and other structures are developed as part of the nervoussystem to support the functioning units, the neurons.
In the central nervous system, cells known collectively as neuroglia serve toprotect and support the neural elements. In the ganglia, satellite cells perform asimilar function.
The brain and spinal cord, enclosed within the brain case and the vertebral canal,are wrapped by the meninges. These are the outer dura mater that lines the bonycavities, the arachnoid mater and the inner pia mater, the latter two separated bya fluid-filled subarachnoid space.
The mammalian nervous system develops as a hollow tube and this tubulararrangement persists in the adult. Within the brain are the ventricles (two lateral,third and fourth) and in the spinal cord a central canal. A clear liquid, thecerebrospinal fluid, is formed in the ventricles. It passes into the central canaland also through the foramina in the roof of the fourth ventricle into thesubarachnoid space, where it is reabsorbed into the venous system. Thecerebrospinal fluid is produced by special vascular structures, the choroidplexuses, that project into the ventricles. The membranes of the choroid
Figure 34.12 Medial view of a hemisected dog’s brain. The thalamus andhypothalamus together make up most of the diencephalon. Scaleapproximates actual size.(© ABRS) [M. Thompson]
43

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
plexuses and cerebral vessels serve as a barrier against introduction of foreignmaterials into the central nervous tissues. The cerebrospinal fluid further servesas a water-bed suspending brain and cord and protecting them from injury.
THE SPECIAL SENSES
Sense organs, which occur in various parts of the body, function to measureeither the intensity or the extent of change of conditions. The eye measuresintensity of light and its wave-length and also the distribution of patterns ofchange of light intensity, enabling the body to react to shapes. The skinmeasures mechanical movement and the ear is also a mechanoreceptor, able todetect the minute air movements we call sounds. The nose and tongue measurechemical concentrations, the skin measures temperature, and the vestibularapparatus measures changes in the position and orientation of the head.Although these various sense organs measure these changes of conditions, theydo not have the capacity to interpret them. Reception of such information is ofno value to the animal unless there is some channel of communication betweenthe receptor and the organs (muscles or glands) which should make theappropriate response. Such communication may be made by hormonal action,but in general it is made by the nervous system.
The Eye and Vision
Eyes operate by translating a change in the incident radiation into nerveimpulses in the fibres of the optic tract. These are then analysed by the brain toproduce appropriate reactions.
There are marked differences between the eyes and their neural connections ofdifferent mammals according to the type of information required. Nocturnalanimals, for example, require a high sensitivity which would be a distinctdisadvantage in very strong light. With few exceptions, however, the basic planof the eyeball is similar among species. The eyeball is roughly spherical. It issituated in the orbit and connected with the brain by a structure referred to as theoptic nerve, which is really a tract. Internally, there is a set of chambers filledwith watery or gelatinous fluids and toward the front of the eyeball lies the lens.The walls of the eyeball are formed of three layers: the inner retina, the choroidand the outer sclera. The retina includes the sensory part of the eye systemwhere visual stimuli are received and transferred to the brain via the optic nerve.The scleroid coat is modified rostrally to form the transparent cornea. The outerlayer of the retina is fused with the choroid and in most parts is pigmentedheavily. The pigment is black over much of the extent of the eyeball, but inmany species portions of it are coloured and act as a reflecting surface, thetapetum. Opposite the margins of the lens, the conjoined layers of retina andchoroid expand to form the ciliary body from which the lens is suspended.Rostral to this, these layers curve inward parallel to the lens to form the iris,which is incomplete centrally. The resulting central opening is the pupil.
The cornea and lens focus the light upon the retina, which receives the imageand transmits it to the brain. The iris acts as a diaphragm, under sympathetic andparasympathetic control, regulating the size of the pupil and, therefore, theamount of light entering the eye.
Accessory structures aid the efficient functioning of the eye. Ocular musclesmove the eyeball, lids serve to moisten and clean the corneal surface by closingat intervals and washing the secretion of the lachrymal glands across it. Landmammals have a duct leading from the medial corner of the eye to the nasalcavity. This drains the lachrymal fluid from the eye surface and has an accessoryfunction in moistening the nasal mucosa.
44

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The Ear, Hearing and Space Orientation
The apparatus of the inner ear shows a remarkably constant organization amongthe vertebrates. It contains receptors that respond to slight mechanicaldeformation and give information about the direction and frequency of incidentsound waves, the position of the head in relation to gravity and the angularacceleration of movement of the head in various directions.
The receptor systems of the ear are: the spiral organ within the cochlea formeasuring intensity and frequency of sound waves; the cristae within thesemicircular canals for response to rotational movements of the head in space;and the maculae of the utricle and saccule providing tonic gravitationalreceptors.
Sound reaches the receptors in the cochlea through the external and middle ears,which are developed solely for this purpose. Sound reception plays aparticularly important role in the lives of aquatic forms and considerablemodification of the external and middle ears of these are observed.
The auricle (pinna) is variable in shape and size among mammals and can bemoved to maximise sound reception and aid in detecting the direction fromwhich sound is coming. The remainder of the external ear consists of a tube heldopen by the presence of cartilage and bone in its walls. The tympanic membraneis stretched across the interior end of the tube and marks the outer wall of themiddle ear. Vibrations of the tympanic membrane, set up by sound waves, aretransmitted to the inner ear by the three auditory ossicles: the malleus, incus andstapes. The footplate of the stapes fits into a small opening leading to the innerear. There are synovial joints between the ossicles. The system ensures thatevery inward movement of the centre of the tympanic membrane produces amovement of about one-third the extent in the foot plate of the stapes. Thepressure at the stapes is several-fold greater than at the tympanic membrane. Theinner ear is protected, to some extent, against loud sounds by the presence oftwo tiny muscles attached to the ossicles which may act to decrease theamplitude of the vibrating systems.
Special use is made of the auditory system by certain bats and whales in thesystem known as echolocation or sonar. Sounds are produced as a series ofpulses and the echoes of these pulses provide auditory clues by which the animalnavigates among objects and detects prey.
The parts of the inner ear associated with three dimensional space orientationare universal in vertebrates. The three semicircular canals, set mutually at rightangles to one another, are tubes forming almost complete circles. They arisefrom a larger central sac, part of whose wall is evaginated to form the utricle.Each tube has an ampulla at one end, in which lies the very sensitive receptor.The utricle communicates with the saccule by a narrow duct and both containsensory receptors, the maculae.
Taste
Most of the simpler senses and some of the complex ones such as vision andsound perception, are responses to physical stimuli, but taste and smell areresponses to chemical stimulation.
The taste buds consist of barrel-like collections of elongated cells sunk withinthe epithelium. Taste buds are confined to the mouth and pharynx and, inmammals, are concentrated on the tongue, particularly certain papillae.
Much of what is casually thought of as taste is actually a smelling of mouthcontents and, in addition, the feel of food in the mouth.
45

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
Although all taste buds are similar in structure, their receptive capabilities differ.These differences lie in the responses of the taste buds to specific sorts ofmolecules or ions in solution in the mouth. Sensations of sour, salty, bitter andsweet can be distinguished in man; other mammals have similar discriminatorypowers.
Olfaction
Although the nose has become associated with breathing in mammals, itsprimary function is in olfaction. The whales, dolphins, dugongs and manateesare specialised to the extent that there is no olfactory mechanism at all. But,among the mammals and vertebrates in general, smell is in many ways the mostimportant of all senses.
Within the nasal cavity, particularly in its dorso-caudal region, is the olfactoryepithelium, which contains olfactory cells interposed with supporting elementsand mucous cells. The olfactory cells differ from other receptor organs inmammals in that they not only receive sensations, but transmit them along theirextensions to the olfactory bulb of the brain.
The olfactory cells are sensitive to minute amounts of chemical materials,derived from distant objects, which go into solution in the liquid covering theolfactory epithelium in the nasal cavity. The precise nature of olfaction is notwell understood. Smell discrimination may be due to the presence ofsubmicroscopic pockets of different shapes at the tips of the sensory cells andstimulation of a cell may be caused by the presence of a molecule whose shapeenables it to fit into one or another type of pocket.
Apparently closely associated with olfactory sense is the vomeronasal organ(Jacobson’s organ), which is an apparently functional organ in many groups ofmammals, but is absent in humans, many bats and various aquatic forms. It ispaired, lying in the rostral base of the nasal septum as a tubular pocket ofolfactory epithelium, partially enclosed by a scroll of cartilage. Eachvomeronasal organ opens rostrally into an incisive duct (or into the nasal cavityin many rodents). It is supplied by several nerves which pass through thecribriform plate of the ethmoid bone with the olfactory nerves and along themedial surface of the olfactory bulb to the accessory olfactory bulb.
The specific function of the vomeronasal organ is as yet unknown, althoughthere is evidence that it plays a role in sexual behaviour and kin recognition. Thebehavioural ‘lip curl’ or ‘Flehmen’ reaction observed in many mammals mayprovide mechanical assistance in clearing the passageways to the vomeronasalorgan and in aspirating odorants so they may reach the receptor sites of theorgan.
Receptors in the Skin and Viscera
The sensation of pain is believed to be produced by direct stimulation of the endfibres of sensory nerves. Such nerve endings, in the skin at least, appear to bealso sensitive to touch.
Many sensations are received by terminal corpuscles, specialised sensoryendings of variable size (mostly microscopic) and structure. They may bepresent in almost any part of the body, particularly the skin, muscles and tendonsand the viscera. Some five types of sensations may be distinguished inmammals: warmth, cold, pain, touch and pressure. Although it is difficult todetermine accurately the function of the different types of corpuscles, there isgood evidence now for the relationship of fine structure and function in some ofthem, notably the mechanoreceptors.
46

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
Complex nerve endings surrounding the bases of hairs, in particular thevibrissae, are important in touch reception. Proprioceptive sensations includethose obtained from sensory structures located in the striated skeletal musclesand tendons as neuromuscular and neurotendinous spindles. They convey to thecentral nervous system, via associated afferent nerve fibres, information aboutthe state of contraction of the muscles and the position in space of the variousparts of the body.
The receptors in the internal organs provide an essential feedback system thatinitiates and stops many actions, particularly in the young animal, before asystem for regulating rhythmic activity is fully established in the brain.
Receptors able to respond to distension probably are present in the bladder,uterus, stomach and other internal organs. Enteroceptors are present in thevascular system, important ones being in the arch of the aorta and the carotidsinus (sensitive to increased blood pressure) and the carotid body (sensitive toreduction of oxygen tension).
THE ENDOCRINE ORGANS
The endocrine organs, although they occur in many parts of the body, comprisean integrative system in which information and directives are transmitted in theblood stream by chemical messengers, the hormones, produced by the endocrineglands. The method of transmission by the endocrine system is slower than thatby the nervous system. Hormonal effects often are distributed broadly over thebody to a variety of tissues and organs, in contrast to the ‘pinpointing’ possiblein the nervous system.
Integrative organization of the body, thus, is effected by two systems, thenervous system and the endocrine system. The two are intimately interrelated,both morphologically and functionally. The nervous system may be powerfullyaffected, directly or indirectly, by hormones. Conversely, the hypophysis isinfluenced strongly by, and some of its hormones actually are produced bynuclei in, the hypothalamus. There is a complicated series of endocrineinterrelationships, which are extremely important in the integration of variousphysiological activities. Some of these are indicated in Fig. 34.13, whichemphasises the central role played by the adenohypophysis in the endocrinesystem. It, in turn, is influenced greatly by the hypothalamus, which produces avariety of releasing factors that stimulates the adenohypophysis to dispensevarious of its hormones in a specific manner. The central nervous system incertain ways influences the discharge of releasing factors, which then reach thetarget cells in the adenohypophysis via the blood stream, in vessels thatconstitute the hypothalamo-hypophyseal portal system. The releasing factors areproduced in secretory cells of the hypothalamus and released in the medianeminence at the base of the diencephalon. They are then picked up by theprimary plexus of the portal system and, passing to the sinusoids in the parsdistalis of the adenohypophysis, stimulate hormone production there.
In some systems, the secretion of a particular adenohypophyseal hormonestimulates increased hormone production by another endocrine gland. Theincreased level of that hormone then acts back on the hypothalamo-hypophysealaxis, resulting in a reduction in release of the adenohypophyseal hormone. Anexample of this type of interrelationship is shown in Fig. 34.14.
Hormones produced by the gastrointestinal tract and those secreted by the testesand ovary have been discussed previously in this chapter. Among its otherfunctions, the placenta synthesises hormones important in pregnancy whichcontinue or supplement activities of the hypophysis and ovaries.
47

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The Hypophysis Cerebri
The hypophysis cerebri is a dual structure, the two portions having differentembryological origins and functions. It lies in the hypophyseal fossa in the floorof the cranial cavity, immediately ventral to the hypothalamus.
The adenohypophysis lies rostral to the neurohypophysis and the bulk of it ismade up of branching cords of cells separated by sinusoids, the pars distalis. Sixtypes of cells are identifiable in the adenohypophysis, each responsible for therelease of one or more hormones.
The neurohypophysis is directly continuous with the hypothalamus. Thesecretions found in it are not produced there, but are neurosecretions producedby cells in the hypothalamus (particularly in the supraoptic and paraventricularnuclei) and released at the tips of their axons in the neurohypophysis.
Seven known hormones are secreted from the adenohypophysis, the first sixfrom the pars distalis. Follicle-stimulating hormone (FSH) stimulates the growthof ovarian follicles, promotes oestrogen secretion and ovulation and promotesspermatogenesis. Luteinizing hormone (LH) influences maturation of thegonads and production of sex hormones, acts in formation of corpora lutea andsecretion of progesterone in the ovary and stimulates the interstitial cells of thetestis, thus promoting the production of androgens and the maturation of sperm.Prolactin stimulates milk secretion and prolongs the functional life of the corpus
Figure 34.13 Some neurohormonal interactions inthe body, illustrating the central position of thehypothalamo-hypophyseal axis in the control ofendocrine function. This schema is a simplification ofa highly complex system of interactions. Not shownis that many hormones act back on the brain andhypophysis (see Fig. 34.14).
48

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
luteum. Adrenocorticotrophic hormone (ACTH) is vital for the normal activityof the adrenal cortex in secretion of hormones and exerts some metabolicinfluence in other ways. Thyroid stimulating hormone (TSH) is essential for thestimulation of the thyroid to form and release thyroid hormones. Growthhormone (GH) has a very broad influence on growth and metabolism in general,with marked influence on growth of skeleton and muscle, metabolism of fatsand carbohydrates and protein synthesis. It further acts to enhance the effect ofother hormones on the activity of the thyroid, adrenal cortex and reproductiveorgans. Intermedin, secreted by the pars intermedia, has no known function inmammals.
The secretions released from the neurohypophysis have two major functions,oxytocic and antidiuretic. The oxytocic effects are mainly contraction of theuterine muscle, mammary gland development and milk ejection after the birth ofyoung. Antidiuretic effects are associated principally with increasing bloodpressure through contraction of arterioles and controlling water output or intakethrough effect on the kidney.
The Adrenals
Adjacent to the kidneys at their cranial poles are the paired adrenals (suprarenals),consisting, in mammals, of distinct outer cortical and inner medullary layers. Bothare endocrine glands, but of different sorts. The medullary tissue is a modifiedpart of the nervous system, innervated by preganglionic autonomic nerve fibres,which on stimulation release into the blood stream two chemicals adrenalin andnoradrenalin. The cells of the adrenal cortex secrete a series of steroid hormoneswhich has widespread influence over bodily functions. In general, this influenceinvolves aiding the body to meet long-continued environmental stresses. This is in
Figure 34.14 Interrelationships of the hypothalamus, hypophysis and ovary.The organs within the ovary (follicles and corpus luteum) are stimulated byhypophyseal hormones and the products of those ovarian organs, oestrogenand progesterone, act in turn on the brain and hypophysis to influence therelease of hypophyseal hormones.
49

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
contrast to the function of the adrenal medulla, which functions in coping withmore immediate, short-term stresses. Three main types of cortical steroids may bedistinguished. The first includes cortisone, corticosterone and cortisol, which arepotent in affecting carbohydrate and protein metabolism. The second type, forexample, deoxycorticosterone, has major effects on salt and water metabolism.The third type is aldosterone, the most effective steroid in sodium and potassiummetabolism. With the exception of aldosterone, the secretion of these hormones isinfluenced by ACTH.
The Thyroid Gland
This gland is found in the neck of mammals, lying against the trachea. Ofpharyngeal origin, it migrates caudally to different levels in different species, sothat it may lie near the larynx or close to the cranial thoracic aperture. The glandis composed of numerous follicles bound together by connective tissue, eachfollicle consisting of a cuboidal epithelium of secretory cells surrounding acentral cavity that contains a colloid substance. In the colloid are found stores ofthyroglobulin from which, through hydrolysis, are formed the hormonesthyroxine and triiodothyronine - the former more abundant, but the latter moreactive.
Hormone secretion from the thyroid is controlled to a great extent by thehypophysis through the production of TSH, but a variety of environmentalstimuli may be influential as well.
Pale, parafollicular cells occur subjacent to the epithelial cells and areresponsible for production of the hormone calcitonin. Its secretion is inducedwhen the concentration of calcium in the blood becomes high. Its effect is todepress the level of blood calcium by suppressing reabsorption of bone.
The Parathyroid Glands
There are usually two pairs of parathyroids in mammals, either partly embeddedin the thyroid gland or beneath or just outside its capsule. They are very smalland consist of closely packed cells interspersed with numerous sinusoids.
The secretion of the parathyroid glands, through the release of parathormone,causes the withdrawal of calcium from bones into the blood. An increasingcalcium level in the plasma depresses parathyroid activity.
The function of the parathyroid glands and the parafollicular cells of the thyroidgland is to control the amount of calcium circulating in the blood. In performingthis function they cooperate with the vitamin D introduced with the food orsynthesised in the skin by the action of sunlight.
The amount of calcium in the interstitial fluids affects the surface properties ofevery cell, so it is obvious that the stabilisation of this ion is vitally important.
The Pancreatic Islands
Among the abundant lobules of the pancreas forming the exocrine part of thegland are isolated islands of cells, the pancreatic islands, which togetherconstitute an endocrine gland. They produce a specific protein, insulin. Insulinhas an important regulatory action on metabolism, particularly of carbohydrates,by increasing their utilisation by the tissues, moderating glucose formation inthe liver, facilitating the entry of glucose into certain types of cells andmaintaining the blood sugar level. Insulin secretion seems to be controlledmainly by the level of sugar in the blood passing through the pancreatic islands.The polypeptide glucagon also is produced in the pancreatic islands, but isformed in a different cell type from that secreting insulin. Glucagon offsets theinfluence of insulin. It affects sugar metabolism by breaking down glycogenstored in the liver so that the level of blood glucose is increased.
50

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
The Pineal Gland
The pineal gland (epiphysis cerebri) is in the dorsal part of the diencephalon. Itis extremely variable in size and shape in mammals, but absent in certainspecies, such as the dugong.
The pineal secretes the hormone melatonin, although it is not the only site ofproduction of this indole. Light impinging on the retina is believed to cause achange in the sympathetic output of the cranial cervical ganglion, which resultsin decreased production and release of melatonin. A daily rhythmic activity inmelatonin synthesis dependent on environmental lighting is observed inmammals. Melatonin has been shown to be inhibitory to the gonads.Considerable emphasis has been placed on the role of the pineal gland inmediating the influence of the photoperiod on the neuroendocrine-reproductiveaxis. For a time this was assumed to be the primary role of the gland, but its rolenow appears to be more general. As currently perceived, the function of thepineal gland in mammals is to serve as an intermediary between the externalenvironment, especially the photoperiod, and the organism as a whole.
THE LYMPHOID ORGANS
Certain regions or organs of the body contain accumulations of lymphoid tissue.These may be in simple organization in close relation to epithelial membranes,for example diffuse lymphoid tissue, solitary nodules, aggregate nodules,tonsils, or as more organised structures that comprise separate organs, forexample lymph node, spleen, thymus.
Lymphoid tissue consists of two primary elements: the reticular tissue and cellswhich are chiefly lymphocytes. The major functions of lymphoid tissue are theproduction of new lymphocytes and the phagocytic activities in the tissue of freeand fixed macrophages. The latter activity is directed against bacteria, spentblood cells and foreign materials associated with the filtration by the lymphoidtissue of tissue fluid, lymph or blood.
Diffuse lymphoid tissue occurs mainly in the tissue beneath the mucousmembrane of the alimentary and respiratory tracts. Aggregate nodules, oftencalled Peyer’s patches, are found in the intestinal lining. Three tonsillar groupsform a lymphoid ring about the entrance of the throat. The lymph nodes occur invarious parts of the body and are the only organs that act as filterers of lymph.As such, they tend to form groups, either many small or few large nodes(depending on the species), at foci of lymph drainage such as the proximal endsof the limbs, around the junction of the head and neck and in the mesenteries.Some species, notably the ruminants, possess haemal nodes, in addition to thelymph nodes, along the ventral side of the vertebrae.
The spleen is the largest of all lymphoid organs and is specialised for filteringblood. Its size is variable within and among mammals. It lies on the left side inclose relationship with the stomach and omentum. It is an elastic, controllablereservoir that is important in adjusting the volume of circulating blood tochanging needs. In particular, it stores red blood cells in its sinuses and red pulp.The spleen is an important haematopoietic organ, generating both lymphocytesand monocytes. In early foetal life it is, like other organs such as the liver, animportant source of all types of blood cells.
The thymus is a lymphoid organ found in the cranial thoracic region and in theyoung of some species extends the full length of the neck. Essential for thenormal development and maintenance of immunological competence, itbecomes relatively and often absolutely, smaller as postnatal growth proceeds.In addition, various protein substances have been isolated from the thymus andproposed as thymic hormones.
51

34. MORPHOLOGY AND PHYSIOLOGY OF THE EUTHERIA
LITERATURE CITED
Ariëns Kappers, C.U., Huber, G.C. & Crosby, E.C. (1960). The ComparativeAnatomy of the Nervous System of Vertebrates, Including Man. HafnerPublishing Co. : New York 3 vols xvii 1845 pp.
Basmajian, J.V. (1979). Muscles Alive. Their Functions Revealed byElectromyography. Williams & Wilkins : Baltimore 4th edn x 495 pp.
Bolk, L., Goppert, E., Kallius, E. & Lubosch, W. (1967). Handbook derVergleichenden Anatomie der Wirbeltiere. Urban & Schwarzenberg : Berlin 5Bands xvi 752 pp., x 1444 pp., xi 1018 pp., xii 1016 pp., xxii 1106 pp.
de Beer, G.R. (1937). The Development of the Vertebrate Skull. Clarendon Press: Oxford xxiv 552 pp. 143 pls
Ebashi, S., Maruyama, K. & Endo, M. (eds) (1980). Muscle Contraction. ItsRegulatory Mechanisms. Springer-Verlag : New York xvii 549 pp.
Flower, W.H. (1885). An Introduction to the Osteology of the Mammalia.MacMillan : London xi 382 pp.
Getty, R. (1975). Sisson and Grossman’s Anatomy of the Domestic Animals.W.B. Saunders : Philadelphia 5th edn 2 vols xxxv 2095 pp.
Handbook of Physiology (1959 to 1985). A critical, comprehensive presentationof physiological knowledge and concepts. Multiple sections, each dealingwith a body system; several volumes in each section. American PhysiologicalSociety : Bethesda
Kampmeier, O.F. (1969). Evolution and Comparative Morphology of theLymphatic System. C.C. Thomas : Illinois xiv 620 pp.
Kluge, A.G. (1977). Chordate Structure and Function. Macmillan : New York2nd edn x 628 pp.
Negus, V. (1958). The Comparative Anatomy and Physiology of the Nose andParanasal Sinuses. E. & S. Livingstone : London xx 402 pp.
Negus, V.E. (1962). The Comparative Anatomy and Physiology of the Larynx.Hafner : London xix 230 pp.
Nomina Anatomica Veterinaria. 3rd edition; Nomina Histologica. 2nd edition(1983). International Committee on Veterinary Gross AnatomicalNomenclature, World Association of Veterinary Anatomists : Ithaca 209 pp.;xii 60 pp.
Romer, A.S. (1970). The Vertebrate Body. W.B. Saunders : Philadelphia 4th ednviii 601 pp.
Slijper, E.J. (1946). Comparative biologic - anatomical investigations on thevertebral column and spinal musculature of mammals. Verhandelingen derKoninklijke nederlandsche Akademie van Wettenschappen (Tweede Sectie)42(5): 1-128
Swenson, M.J. (ed.) (1979). Duke’s Physiology of Domestic Animals. 9th ednCornell University Press : Ithaca x 914 pp.
Young, J.Z. (1962). The Life of Vertebrates. 2nd edn Clarendon Press : Oxford820 pp.
52





























