Embed Size (px)
Citation preview

Ferredoxin:NADPH oxidoreductase is recruited tothylakoids by binding to a polyproline type IIhelix in a pH-dependent mannerFerdinand Altea,b, Anna Stengela,c, J. Philipp Benza,c,1, Eike Petersena,c, Jürgen Solla,c, Michael Grolla,b,2, and Bettina Böltera,c,2
aMunich Center for Integrated Protein Science, Ludwig-Maximilians-Universität, 81377 Munich, Germany; bDepartment Chemie, Lehrstuhl für Biochemie,Technische Universität München, 85747 Garching, Germany; and cDepartment für Biologie I, Pflanzenbiochemie, Ludwig-Maximilians-UniversitätMünchen, 82152 Planegg-Martinsried, Germany
Edited by Bob B. Buchanan, University of California, Berkeley, CA, and approved September 14, 2010 (received for review June 30, 2010)
Ferredoxin:NADPH oxidoreductase (FNR) is a key enzyme of photo-synthetic electron transport required for generation of reductionequivalents. Recently, two proteins were found to be involvedin membrane-anchoring of FNR by specific interaction via a con-served Ser/Pro-rich motif: Tic62 and Trol. Our crystallographic studyreveals that the FNR-binding motif, which forms a polyproline typeII helix, induces self-assembly of two FNR monomers into a back-to-back dimer. Because binding occurs opposite to the FNR activesites, its activity is not affected by the interaction. Surface plasmonresonance analyses disclose a high affinity of FNR to the bindingmotif, which is strongly increased under acidic conditions. The pHof the chloroplast stroma changes dependent on the light condi-tions from neutral to slightly acidic in complete darkness or to alka-line at saturating light conditions. Recruiting of FNR to thethylakoids could therefore represent a regulatory mechanism toadapt FNR availability/activity to photosynthetic electron flow.
polyproline helix ∣ proline recognition domain ∣ protein–proteininteraction ∣ Tic62
The generation of reducing power is crucial for all biosyntheticprocesses within chloroplasts. The main source of reduction
equivalents is light-driven photosynthesis. The final step ofphotosynthetic linear electron transport is subsequently catalyzedby the ubiquitous key enzyme ferredoxin:NADPH oxidoreduc-tase (FNR, EC 1.18.1.2.), which transfers two electrons fromreduced Fd via its cofactor FAD to NADPþ. It thereby generatesthe reduction equivalents (NADPH) required for multiple meta-bolic pathways.
FNR is a soluble protein, however it was also found associatedwith thylakoid membranes (1). Several potential FNR-bindingpartners have been discussed, which might be involved in mem-brane attachment of FNR. Nevertheless, no conclusive biochem-ical evidence for a direct interaction with FNR was provided forany of them. It was demonstrated recently that FNR interactsdirectly and specifically with a conserved amino acid motif foundin two different proteins, leading to membrane association ofFNR into high-molecular-weight complexes. One of these pro-teins, Tic62, was originally identified as a member of the Ticcomplex (translocon at the inner envelope of chloroplasts) in-volved in redox-regulation of protein import into chloroplasts(2, 3). Tic62 was found to represent a major FNR interactionpartner at the thylakoids, and binding to Tic62 clearly increasesthe stability of FNR (4). The specific interaction with FNR ismediated by a conserved sequence motif rich in proline andserine residues, located in the C terminus of Tic62 (2, 3). Inter-estingly, this motif is restricted to Tic62 proteins from vascularplants (5), and it occurs in different numbers dependent onthe respective plants species. Additionally, a second protein con-taining the Ser/Pro-rich FNR-binding motif has been recentlyidentified in Arabidopsis thaliana and named Trol (thylakoid rho-danese-like protein). It represents an integral membrane proteinlocated in the thylakoids and contains a single FNR-interacting
domain at the C terminus, which was shown to act as an FNRanchor similar to Tic62 (6).
To date, the functional relevance as well as structural charac-teristics of FNR membrane recruitment by interaction withTic62/Trol remain elusive. Thus, in the present study, we analyzedthe specific interaction of FNR with a synthetic peptide, repre-senting the conserved binding motif [henceforth called theFNR-membrane-recruiting-motif (FNR-MRM)] found in Tic62and Trol, at atomic resolution. We determined the crystal struc-ture of the FNR:peptide complex at 1.7 Å and used Surface Plas-mon Resonance (SPR) analysis to characterize the biophysicalproperties of the interaction in more detail. Our structural datareveal that the FNR-MRM induces self-assembly of two FNRmolecules. FNR activity measurements showed that the perfor-mance of the complex-bound FNR remains unchanged comparedto the free enzyme. Finally, SPR analyses reveal high bindingaffinity of the two FNR enzymes to the peptide, which is signifi-cantly increased under acidic conditions.
ResultsFNR Membrane Recruitment is Mediated by a Highly ConservedSequence Motif. We first characterized the FNR-binding domainby database analysis. A stretch of 58 amino acids from the peaTic62 C terminus (amino acids 354–411) containing the Ser/Pro-rich FNR-MRM was used as a template for a search in allnonredundant databases via tblastn (7). The conserved motifwas found exclusively in vascular plants, either belonging toTic62 proteins (5) or to Trol homologs (Fig. 1C and Fig. S1A).In all Tic62 proteins identified to date at least two FNR-interact-ing repeats are present. In contrast, almost all Trol proteinscomprise only one FNR-MRM, except the monocotyledonsOryzasativa, Zea mays, and Triticum aestivum, which contain two FNR-binding repeats. Thus, the FNR-MRM is restricted to vascularplants.
Alignment of all conserved FNR-MRMs applying the Clus-talW2 program (8) revealed the presence of a “core motif” of19 amino acids located in the distal C terminus (Fig. 1C andFig. S1B). Intriguingly, an alternating pattern of hydrophobic/
Author contributions: F.A., J.S., M.G., and B.B. designed research; F.A., A.S., J.P.B., and E.P.performed research; F.A., A.S., J.P.B., J.S., M.G., and B.B. analyzed data; and F.A., M.G., andB.B. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The atomic coordinates have been deposited in the ResearchCollaboratory for Structural Bioinformatics Protein Data Bank, www.pdb.org (PDB ID code3MHP).1Present address: Energy Biosciences Institute, University of California, 130 CalvinLaboratory, Berkeley, CA 94720.
2To whom correspondence may be addressed. E-mail: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1009124107/-/DCSupplemental.
19260–19265 ∣ PNAS ∣ November 9, 2010 ∣ vol. 107 ∣ no. 45 www.pnas.org/cgi/doi/10.1073/pnas.1009124107
Dow
nloa
ded
by g
uest
on
Aug
ust 6
, 202
0

hydrophilic residues can be observed in this strictly conserved re-gion. Conservation is less pronounced in the first 40 amino acids,though an accumulation of Ser and Pro residues is still obvious.Most of the amino acid residues in the FNR-MRM are invariableand hence might be essential for FNR-binding. Consequently, weused a synthetic peptide representing the first FNR-binding re-peat from Pisum sativum Tic62 (PsTic62), including the strictlyconserved core motif (KTEQPLSPYTAYDDLKPPSSPSPTKPS)for further analysis.
Overall Structure of the Complex. Here we report the crystal struc-ture of FNR in complex with the aforementioned peptide. Themature part of PsFNR was cocrystallized with the peptide andthe structure was determined by molecular replacement (9) usingPsFNR [Protein Data Bank (PDB) ID: 1QG0] as a search model.The dataset was refined to 1.7 Å resolution (Table 1). The twelveN-terminal amino acids of both FNR subunits (termed FNR Aand FNR B, respectively) as well as a loop of seven amino acids(Thr167–Val173; numbers according to full-length PsFNR) inFNR B were structurally distorted. The geometry of the wholeTic62 peptide is well defined in the electron density map, exceptthe C-terminal serine. The structure discloses that the FNR:pep-tide complex consists of two FNR molecules, which are bound toone peptide (Fig. 1A). Hereby, the peptide is sandwiched by twoFNR monomers, with the two subunits arranged back-to-backin a way that the two FAD-molecules face opposite sides of thecomplex and are solvent exposed. Because both FNR monomersare turned by 180°, the central peptide constitutes an imaginaryrotational C2-symmetry axis. Thus, the redox centers of bothFADs are accessible for interaction with Fd (10). Even thoughprotein–protein interactions often go hand in hand with consid-erable structural rearrangements of the molecules involved, sucha rearrangement is not observed in the presented case. Compar-ing the structure of the FNRmonomers in the complex and in thefree state, the molecules appear like rigid bodies, not showing
Fig. 1. Structures of the FNR:Tic62-peptide complex using Molscript (27) andBobscript (28). (A) Stereo view of the two FNR-subunits, shown as gray ribbonplots and denoted as FNR A and FNR B, respectively. The Tic62 peptide,colored in green, and both FAD cofactors, colored in yellow, are shown asball-and-stick models. N and C termini of the peptide are numbered accord-ing to the full-length Tic62 protein. (B) Stereo view of the 2Fo-Fc electrondensity map (countered at 1σ) for the Tic62 peptide in the complex interface,in which the Tic62 peptide has been omitted for phasing. FNRs are repre-sented in coils and amino acid residues of both FNR subunits complexingthe Tic62 peptide are depicted as sticks. Hydrogen bonds and a salt bridgeare indicated by black dashed lines. Here, the peptide amino acids arenumbered from 1 to 27. (see also Table S1). (C) Alignment of the C terminiof Tic62/Trol proteins from different vascular plants containing the FNR-MRM. The red frame indicates the strictly conserved core-motif. Ps, Pisum sa-tivum: At, Arabidopsis thaliana: Os,Oryza sativa: Pt, Populus trichocarpa: OsJ,Oryza sativa subsp. japonica. The OsJ_04356 sequence is related to a putativeuncharacterized protein. (see also Fig. S1A)
Table 1. Data collection and refinement statistics regarding theFNR:peptide complex
FNR:Tic62 peptide
Crystal parametersSpace group P1Cell dimensions
a, b, c, Å 47.5; 48.8; 71.3α, β, γ, ° 106.6; 97.0; 91.8
Molecules per AU* 2 FNR; Tic62 peptideData collectionBeam line Cu Kα
Wavelength, Å 1.5418Resolution range,† Å 65–1.7 (1.8–1.7)Unique reflections‡ 66902Completeness, %† 99.9 (99.5)Redundancy 10.9 (3.9)Rmerge,
†, § % 3.5 (23.6)I∕σðIÞ† 23.6 (3.3)RefinementResolution, Å 15–1.7Rwork∕Rfree
¶ 0.138∕0.189No. atoms
Protein 4861FAD ligands 106Water 513
B factors 16.6rms deviations∥
Bond lengths, Å 0.024Bond angles, ° 1.826
Ramachandran,** % 99.5∕0.5∕0.0PDB accession code 3MHP
*Asymmetric unit.†Values in parenthesis of resolution range, completeness, Rmerge and I∕σ (I)correspond to the last resolution shell.
‡Friedel pairs were treated as identical reflections.§RmergeðIÞ ¼ ΣhklΣj j½IðhklÞj − IðhklÞ�j∕½ΣhklIhkl�, where IðhklÞj is the jthmeasurement of the intensity of reflection hkl and hIðhklÞi is theaverage intensity.
¶R ¼ ΣhkljjFobsj − jFcalcjj∕ΣhkljFobsj, where Rfree is calculated without asigma cutoff for a randomly chosen 5% of reflections, which werenot used for structure refinement, and Rwork is calculated for theremaining reflections (30).
∥Deviations from ideal bond lengths/angles (31).**Number of residues in favored region/allowed region/outlier region.
Alte et al. PNAS ∣ November 9, 2010 ∣ vol. 107 ∣ no. 45 ∣ 19261
BIOCH
EMISTR
Y
Dow
nloa
ded
by g
uest
on
Aug
ust 6
, 202
0

significant changes in the trace of the backbone, as depicted bybackbone superposition of the two FNR-monomers in the com-plex with the backbone of the free FNR (PDB ID 1QG0)(Fig. S2).
The FNR-domain interacting with the peptide is composedof a twisted three-stranded antiparallel β-sheet (β1–β3), two as-sociated loops, and two flanking α-helices (α1 and α2) (Fig. S3).The domain forms a shallow “trough-shaped” surface groove,which is occupied by the peptide without causing structural rear-rangements upon ligand binding. Most interactions betweenthe FNR-domain and the peptide originate from the two helicesα1 and α2, whereas only few residues of the β-sheet and theassociated loops are involved in complex formation. Helix α2protrudes into the interior of FNR, thus only the bottom windingand partially the adjacent loop interact with the peptide. The pep-tide itself does not adopt defined regular secondary structuressuch as α-helices or β-strands (Fig. 1A). At first sight the traceof its backbone consists of a random series of turns and loopswithout any apparent symmetric features. Hence, it is rathersurprising that the symmetry of the whole complex is generatedby a putative asymmetric core module.
The FNR-MRM Forms a Polyproline Helix That Interacts with a ProlineRecognition Domain on FNR. The conserved FNR-MRM containsabout 25% prolines, which suggests a protein–protein interactionmediated by a polyproline motif. It is known that polyprolineligands feature the conformation of a polyproline type II (PPII)helix when bound to the target protein (11, 12). However, thisligand type has not been identified in FNR-interacting proteinsto date. Interestingly, PPII helices contain an inherent twofoldrotational pseudosymmetry, thus, the respective binding domainof putative interaction partners can catch the PPII ligand in twopossible orientations (13). In the FNR:peptide crystal structure, aPPII conformation is indeed found in distinct peptide sections(Fig. 2). A structural overlay of a PPII helix with the peptideuncovered three segments that adopt a PPII helical conformation(segment 1, Lys1–Pro5; segment 2, Pro8–Pro18; segment 3,Pro21–Pro26). The axes of all three segments point to differentdirections, and the segments, on their part, are separated fromeach other by two amino acids. Interestingly, segment 2 (Pro8–Pro18) is intercepted by a loop, which consists of the residuesThr10–Leu15. These loop residues contribute to multipleinteractions with both FNR subunits. To our knowledge such aconformation has not yet been reported for polyproline ligands.
The peptide-binding FNR domain exhibits a unique proline-recognition-domain-fold (PRD), as domain superpositioningdoes not reveal structural similarity to any of the so far examinedPRDs (e.g., SH3, profilin) (11).
The Peptide Generates a Vast Surface Contact Area.We next analyzedthe contact area of both FNRmolecules involved in dimerization.Interestingly, the presence of the peptide leads to an enormousincrease of the contact region. Without the peptide, the theore-tical contact surface between the two FNR subunits is restrictedto 380 Å2, as calculated in silico on the protein interfaces,surfaces, and assemblies service at European Bioinformatics In-stitute Web site (14). This area is only formed by a small patchcomprising the amino acids 35–40, 57, 99–104, 135–137 of bothFNR subunits and a salt bridge between Glu215 (FNR A) andLys182 (FNR B) (Fig. 3B). Nevertheless, a large cavity remainsbetween the two FNRs due to steric interference. In the complexstructure, this cavity is filled by the peptide, which enlarges thecontact surface area to 2070 Å2 in total (Fig. 3A). This inducesthe assembly and stabilizes the ternary complex. Thus, althoughthe peptide is asymmetric with respect to its primary sequence, itstill acts as a symmetric dimerization module for the two FNRsubunits.
FNR Displays a pH-Dependent High Affinity to the MRM. In additionto solving the crystal structure of the FNR:peptide complex, wedetermined the affinity of the binding partners for each other by
Fig. 2. Structural overlay of the Tic62 peptide with three PPII helices. OnlyPro-side chains are shown. The backbone dihedral angles of the PPII helicesare φ ¼ −75° and Ψ ¼ 145°. The dihedral angles of the first five amino acidsof the Tic62 peptide (segment 1, Lys1—Pro5) are in the respective range andthe backbone widely matches the backbone of PPII helix 1. The three prolineresidues 8, 17, and 18 together with Tyr9 and Lys16 of the FNR-MRM forma second axis of a PPII helix (PPII helix 2; segment 2, Pro8–Pro18).The thirdsegment (Pro21–Pro26) in the FNR-MRM is indicated by PPII helix 3.
Fig. 3. Surface representation of the FNR:Tic62-peptide complex usingGRASP software (29). Stereo view of both FNR subunits in the complexwith (A) and without (B) the Tic62 peptide, which is colored in green. TheFNR-MRM fills a large cavity between the two FNR-molecules, thereby signif-icantly expanding the contact surface.
19262 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1009124107 Alte et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 6
, 202
0

SPR (15). To analyze binding of the FNR-MRM to FNR, we usedthe synthetic peptide as well as the whole C terminus of PsTic62(¼R3), comprising all three binding motives for the interactionstudies (2). The FNR-MRM peptide or R3 were coupled to aCM5 chip (16). After activation of the chip, binding of the respec-tive ligands and blocking of unspecific binding sites, FNR wasinjected at pH 8. An equilibrium dissociation constant KD of2.75� 0.33 μM was calculated for the peptide and FNR atpH 8 (Table 2 and Fig. S4A) which represents a high affinity,although it has to be noted that the observed association ratewas quite slow. The dissociation constant of R3 and FNR atpH 8 was similar to that of the peptide, but association and dis-sociation rates were about five times faster (Table 2 and Fig. S4B).This observation indicates that the number of FNR-binding re-peats in Tic62 proteins influences the speed of association/disso-ciation but not the general affinity for FNR. This can also bededuced from the association curves, which show no cooperativebinding effect for R3 compared to the peptide. We furthermoreanalyzed potential physiological effects on the binding by lower-ing the pH of the binding reaction to six. Intriguingly, the affinityof FNR to R3 significantly increased. Especially the associationoccurred about 30 times faster at pH 6 compared to pH 8, whichled to an equilibrium dissociation constant differing by a factor ofapproximately 80 (0.041� 0.027 μM) (Fig. S4C). Thus, our SPRresults demonstrate that FNR has a high affinity to the bindingmotif found in Tic62 and Trol, which is strongly increased atacidic conditions.
Next, we studied the activity of the complex-bound FNR com-pared to the free enzyme. FNR activity was measured employinga cytochrome c (Cyt c) reduction assay (4) with varying amountsof FNR (0.5–2 μg total protein) at pH 8 and pH 6, respectively(Fig. S5). R3 alone as control exhibited no activity. For both con-ditions, free FNR displayed a similar performance compared tocomplex-bound FNR, allowing the conclusion that FNR/R3 hasthe same activity as FNR alone. Lowering the pH in the assayresulted in a slightly lower general activity and no differencesin FNR activity of the free and the complexed protein have beenobserved under this condition as well. These data indicate thatthe Tic62-peptide functions as a FNR-recruiting motif that leadsto dimerization of the FNR while leaving its activity unchanged,which conforms to the crystallographic results.
DiscussionIdentifying FNR-binding proteins at the thylakoid membrane is acentral research focus, because it is still controversial how thesoluble FNR becomes attached to thylakoids. Recently, a Ser/Pro-rich domain has been identified in Tic62 and Trol thatspecifically interacts with FNR and is required for its membraneattachment (4). Here, we structurally and functionally character-ized this FNR-MRM in molecular detail. Previous data revealedthat the FNR-MRM of both Tic62 and Trol exclusively exists invascular plants (5) (Fig. 1C and Fig. S1A). Thus, membranetethering of FNR by this motif seems to be a recent evolutionaryinvention. Although FNR is commonly distributed among allthree domains of life, the membrane recruiting componentsTic62 and Trol are restricted to plants possessing a vascularsystem.
The FNR-MRM(s) found in Tic62 and Trol are characterizedby a highly homologous core domain of 19 amino acids (Fig. 1C
and Fig. S1B). Variability increases upstream of the core region,though accumulation of Ser and Pro residues is still evident.Whereas most Trol proteins comprise a single FNR-bindingdomain, its number varies to a higher extent in Tic62 proteins.Interestingly, the binding affinity to FNR did not change signifi-cantly when constructs comprising one or three FNR-interactingmotifs were analyzed (Fig. S4). This indicates that binding to eachdomain occurs independently of the other motifs and excludescooperative binding effects.
To date, a variety of FNR structures are available, either show-ing FNR alone or in complex with its substrate NADPH and/orFd. The present study resulted in a structure of FNR togetherwith a nonsubstrate binding partner, i.e., the conserved motiffound in Tic62 and Trol. In an earlier study, a hydrophobic cavityopposite to the active site was described for FNR (17). Becauseexposure of such a feature is highly unusual in a native protein, itwas speculated that this hydrophobic region could function inmembrane attachment. Several lines of evidence now suggest thatFNR is tethered to the membrane by binding to Tic62 (and Trol)rather than by direct membrane attachment in a pH-dependentmanner (18), for example thylakoid binding of FNR is signifi-cantly disturbed in the absence of Tic62 (or Trol) (4, 6).
Interestingly, the FNR-MRM was found to bind opposite tothe active site of both FNR molecules forming a back-to-backdimer. As found for the complex structures of FNR bound toNADPH and Fd, dimerization of FNR upon interactionwith the peptide does not induce structural rearrangements(Fig. S2A). Even the side chains at the interface exhibit only slightconformational rearrangements (Fig. S2B) and thus the FNRmo-lecules can be regarded as rigid bodies. Hence, the Tic62 peptideseems to mediate self-assembly of the FNR dimer only by surfaceexpansion without influence on the active sites.
A striking feature of the FNR-binding motif is the formation ofa PPII-like helix (Fig. 2) upon complex formation. This type ofsecondary structure element has often been implicated in pro-tein–protein interactions in various cytosolic signal transductionpathways and was extensively characterized in SH3 domainproteins (11) which are present in all eukaryotes (19). TheFNR-MRM in Tic62 and Trol is the only known example of adomain forming the PPII structure found in chloroplasts. Typicalfor PPII domains, Tic62 was proposed to play a role in signalingevents (2–4) by sensing the chloroplast redox state and regulatingprotein import accordingly. FNR was discussed to participatein this process, although the molecular details of redox-relatedimport regulation remain elusive (3, 4, 20).
The structural results could be corroborated by enzymaticmeasurements, which showed that binding of the peptide hasno effect on the activity of FNR (Fig. S5). This clearly differenti-ates Tic62 binding to FNR from other PPII domains, which havebeen described to drastically influence the conformation and/oractivity of their interaction partners (21, 22). However, as theexperiments have been performed in vitro, it remains to beestablished if membrane attachment mediated by Tic62/Trol in-fluences the activity of FNR in vivo, as was proposed by anotherstudy (18).
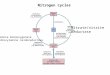
The most interesting property of the FNR-Tic62/Trol interac-tion is the clear difference of the affinity at acidic (pH 6;KD ≈ 0.04 μM) compared to alkaline (pH 8; KD ≈ 3 μM) condi-tions (Fig. S4). Both values indicate a high affinity of FNR and
Table 2. Overview of the association and dissociation constants measured by SPR
Sample name KD � SD, μM ka � SD, 1∕Ms kd � SD, 1∕s
Tic62 peptide, pH 8 2.75 ± 0.33 1.12 × 102 � 0.31 × 102 3.11 × 10−4 � 1.16 × 10−4
R3, pH 8 3.4 ± 1.33 6.18 × 102 � 3.55 × 102 1.31 × 10−3 � 3.75 × 10−4
R3, pH 6 0.041 ± 0.48 1.82 × 104 � 0.62 × 104 4.6 × 10−4 � 5.16 × 10−5
Standard deviations were calculated from at least three independent experiments.
Alte et al. PNAS ∣ November 9, 2010 ∣ vol. 107 ∣ no. 45 ∣ 19263
BIOCH
EMISTR
Y
Dow
nloa
ded
by g
uest
on
Aug
ust 6
, 202
0

Tic62/Trol, since the binding at pH 8 is similar to, e.g., the bindingstrength of a cochaperone to Hsp90 (23). It also reflects thedissociation equilibria of other PPII proteins with their sub-strates, which usually range between 1–10 μM (11). However,the increased binding strength at pH 6 is even in the range ofthe affinity of monoclonal antibodies for their antigens (24)and has not been described for interactions performed by PPIIproteins before. Thus, the interaction of FNR with Tic62/Trolseems to have a function distinct from mere signaling. Intrigu-ingly, the pH variations reflect differences of the chloroplast stro-ma between light and dark cycles (Fig. 4): During light phases,when photosynthetic activity is high, protons are transported intothe thylakoid lumen, leading to an alkaline stromal pH. By con-trast, when photosynthesis ceases during dark phases the stromalpH decreases again. Under these conditions, Tic62 is predomi-nantly associated with the thylakoid membrane where it recruitsFNR into stable high-molecular-weight complexes (4). Light cat-alyses photosynthetic electron flux, however light quantity canvary dramatically during the course of the day, therefore requir-ing constant adjustment of the light harvesting processes and theenzymatic reactions leading to the formation of NADPH as theend product of photochemistry. Changes in light quantities alterstromal pH and thus the amount of FNR bound to the thylakoids.Furthermore, the membrane attachment of FNR is influencedby the stromal redox state (i.e., the NADPþ∕NADPH ratio),which mimics variations in environmental conditions (3). There-fore, reversible attachment of FNR to the thylakoid membranevia Tic62/Trol provides an elegant way to store surplus molecules,not required when photosynthesis is less active or dormant.
Materials and MethodsProtein Purification. PsLFNR and R3 (C terminus of PsTic62; amino acidsVal346–Ser534) were purified via Ni-affinity chromatography as described(4). PsFNRL and R3 were further purified by size exclusion chromatographyapplying a Superdex200 column (GE Healthcare) in 10 mM Tris pH 8, 150 mMNaCl. The FNR/R3 complex was formed by overnight incubation at 4 °C andisolated by size exclusion chromatography applying a Superdex200 column inthe same buffer (Fig. S6).
Crystallization and Structure Determination. Crystals of pea FNR in complexwith a C-terminal peptide fragment (amino acids 383–409) of pea Tic62 weregrown by using hanging-drop vapor diffusion at 20 °C. The protein concen-tration used for crystallization was 37 mg∕mL. Crystals were obtained in
drops containing 2 μL of protein solution and 2 μL of reservoir solution(200 mM ammoniumcitrate and 20% PEG3350). Crystals were subsequentlyshock-frozen in a stream of nitrogen gas at 100 K (Oxford Cryo Systems) and adataset was collected inhouse using our CuKα-rotating anode. The FNR:Tic62complex crystallized in the triclinic space group P1 with cell parameters ofa ¼ 47.5, b ¼ 48.8, c ¼ 71.3 Å, and α ¼ 106.6°, β ¼ 97.0°, γ ¼ 91.8°, with40% solvent content (Table 1). Crystal structure analysis was performed bymolecular replacement using the program phaser (9) and coordinates ofFNR from Pisum sativum deposited at the Protein Data Bank (PDB ID1QG0) (25), yielding two FNR molecules and the Tic62 peptide in the asym-metric unit cell. Model building was performed with the graphic programMAIN. Most of the side chains of FNR could be identified unambiguously ex-cept the 1–13 N-terminal and the last seven C-terminal residues as well asresidues 113–121 in chain B. Refinement of the FNR-molecules resulted ina well-defined 2Fo-Fc electron density map for the Tic62 peptide, exceptfor the C-terminal serine. The model was refined by Refmac (26) usingconventional crystallographic rigid body, positional, and anisotropic tem-perature factor refinements with current crystallographic values of Rcryst
13.9%, Rfree 18.9%, rmsd bond length 0.024 Å, and rmsd bond angle1.83°. The current model of the native molecule in the asymmetric unit cellcomprises 4861 non-hydrogen atoms and 513 water molecules. The atomiccoordinates have been deposited at the Protein Data Bank, Research Colla-boratory for Structural Bioinformatics (PDB ID 3MHP).
SPR. To determine the individual affinities of R3 and the synthetic Tic62 pep-tide for FNR, each peptide was coupled to a CM5 chip (GE Healthcare) usingthe standard amine coupling procedure (16) after determining the optimalpH for binding. The peptides were coupled to flow cell 2 (FC2) of individualCM5 chips to a maximum density of 1,500 resonance units. Flow cell 1 (FC1)served as a reference and was treated equally without coupling of a ligand.Briefly, each chip was activated by injecting 70 μL of a 1∶1mixture of 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimidehydrochloride and N-hydroxysucci-nimide at a flow rate of 10 μL∕min followed immediately by injection ofthe peptide. Remaining active sites were blocked with 70 μL 1 M ethanola-mine. All experiments were performed on a BIAcoreX using 10 mM HepespH 8.0 (pH 6.0), 150 mM NaCl as running buffer, and evaluated using theBIAevalution software 4.1. Manual injections of at least five concentrationsof FNR were done in triplicate using 10 mMNaOH as a regeneration solution.The resulting referenced data (FC2-FC1) were fitted to a 1∶1 Langmuir modelof interaction. The model describes the interaction between an analyte A(FNR) and a ligand B forming a complex: Aþ B ⇌ AB. The association rateka½L∕ðmol · sÞ� corresponding to the number of complexes formed per second
in a molar solution is defined by the formula −d½AB�dt ¼ ka½A�½B�, where [A], [B],
and [AB] are the concentrations of A, B, and the AB complex. The dissociationrate constant kd (in s−1), corresponding to the portion of the complex that
dissociates in 1 s, is given by the formula −d½AB�dt ¼ kd½AB�.
Fig. 4. Schematic model of pH-dependent FNR-storage at the thylakoid membrane by Tic62. During dark periods, when photosynthesis is shut down, FNRstrongly interacts with the Tic62 C terminus due to an acidic stromal pH. Hence, FNR is attached to the thylakoid membrane (Left), where it is stored andstabilized until reactivation of photosynthesis. Low light quantities (indicated by a small, red arrow) induce a modest photosynthetic activity, resulting in ahigher stromal pH, which causes a partial release of FNR molecules into the stroma (Center). Under these conditions, possibly both FNR populations, at themembrane as well as in the stroma, contribute to the production of an adequate amount of reduction equivalents (NADPH). High light conditions (indicated bylarge, red arrows) evoke high photosynthetic activity, leading to an alkaline stroma, since protons are transported into the thylakoid lumen. Under theseconditions, FNR is predominantly found in a Tic62-free form, mediating the transfer of electrons from ferredoxin (Fd) to NADPþ (Right). For the sake of clarity,the role of Tic62 in the chloroplast redox shuttling system is not depicted.
19264 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1009124107 Alte et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 6
, 202
0

The equilibrium dissociation constant KD is calculated as KD ¼ ½A�½B�½AB� ¼ kd
ka.
Cyt c Reduction. Fd-dependent Cyt c reductase activity of free FNR, the FNR:R3complex, and R3 alone was determined as described previously (4) with slightmodifications: The total volume was 100 μL and the activity was monitoredfor 60 s.
Sequence Alignments. A sequence homology search (tblastn) was performedusing the PlantGDB database with the Tic62 protein sequence from Pisum
sativum (PsTic62) and the TROL protein sequence from Arabidopsis thaliana(AtTROL) as templates. The retrieved sequences weremanually inspected andthe C-terminal FNR-binding motifs aligned with the ClustalW2 program (8).
ACKNOWLEDGMENTS. This work was supported by Deutsche Forschungsge-meinschaft (SFB594) (to F.A., A.S., J.P.B., J.S., M.G., B.B.), Elitenetzwerk Bayern(J.P.B.), International Max-Planck Research School for Life Sciences (A.S.),Deutscher Akademischer Austausch Dienst (J.P.B.), and Studienstiftung desDeutschen Volkes (A.S.).
1. Shin M, Tagawa K, Arnon DI (1963) crystallization of ferredoxin-TPN reductase and itsrole in the photosynthetic apparatus of chloroplasts. Biochem Z 338:84–96.
2. Kuchler M, Decker S, Hormann F, Soll J, Heins L (2002) Protein import into chloroplastsinvolves redox-regulated proteins. EMBO J 21:6136–6145.
3. Stengel A, Benz P, Balsera M, Soll J, Bolter B (2008) TIC62 redox-regulated transloconcomposition and dynamics. J Biol Chem 283:6656–6667.
4. Benz JP, et al. (2009) Arabidopsis Tic62 and ferredoxin-NADP(H) oxidoreductase formlight-regulated complexes that are integrated into the chloroplast redox poise. PlantCell 21:3965–3983.
5. Balsera M, Stengel A, Soll J, Bolter B (2007) Tic62: A protein family frommetabolism toprotein translocation. BMC Evol Biol 7.
6. Juric S, et al. (2009) Tethering of ferredoxin: NADPþ oxidoreductase to thylakoidmembranes is mediated by novel chloroplast protein TROL. Plant J 60:783–794.
7. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment searchtool. J Mol Biol 215:403–410.
8. Larkin MA, et al. (2007) Clustal W and clustal X version 2.0. Bioinformatics23:2947–2948.
9. McCoy AJ, et al. (2007) Phaser crystallographic software. J Appl Crystallogr 40:658–674.10. Kurisu G, et al. (2001) Structure of the electron transfer complex between ferredoxin
and ferredoxin-NADPðþÞ reductase. Nat Struct Biol 8:117–121.11. Li SSC (2005) Specificity and versatility of SH3 and other proline-recognition
domains: Structural basis and implications for cellular signal transduction. BiochemJ 390:641–653.
12. Zarrinpar A, Bhattacharyya RP, Lim WA (2003) The structure and function of prolinerecognition domains. Sci STKE 2003:RE8.
13. Lim WA, Richards FM, Fox RO (1994) Structural determinants of peptide-bindingorientation and of sequence specifity in SH3 domains. Nature 372:375–379.
14. Krissinel E, Henrick K (2007) Inference of macromolecular assemblies from crystallinestate. J Mol Biol 372:774–797.
15. Hoa XD, Kirk AG, TabrizianM (2007) Towards integrated and sensitive surface plasmonresonance biosensors: A review of recent progress. Biosens Bioelectron 23:151–160.
16. Johnsson B, Lofas S, Lindquist G (1991) Immobilization of proteins to a carboxymethyl-dextran-modified gold surface for biospecific interaction analysis in surface plasmonresonance sensors. Anal Biochem 198:268–277.
17. Bruns CM, Karplus PA (1995) Refined crystal structure of spinach ferredoxin reductaseat 1.7 Angstrom resolution—oxidized, reduced and 2′-phospho-5′-AMP bound states.J Mol Biol 247:125–145.
18. Grzyb J, Gagos M, Gruszecki WI, Bojko M, Strzalka K (2008) Interaction offerredoxin: NADPðþÞ oxidoreductase with model membranes. Biochim Biophys Acta,Biomembr 1778:133–142.
19. Rubin GM, et al. (2000) Comparative genomics of the eukaryotes. Science287:2204–2215.
20. Stengel A, Benz JP, Buchanan BB, Soll J, Bolter B (2009) Preprotein import into chlor-oplasts via the Toc and Tic complexes is regulated by redox signals in Pisum sativum.Mol Plant 2:1181–1197.
21. Groemping Y, Rittinger K (2005) Activation and assembly of the NADPH oxidase: Astructural perspective. Biochem J 386:401–416.
22. Takeya R, Sumimoto H (2003) Fhos, a mammalian formin, directly binds to F-actinvia a region N-terminal to the FH1 domain and forms a homotypic complex via theFH2 domain to promote actin fiber formation. J Cell Sci 116:4567–4575.
23. Brinker A, et al. (2002) Ligand discrimination by TPR domains—relevance andselectivity of EEVD-recognition in Hsp70 center dot Hop center dot Hsp90 complexes.J Biol Chem 277:19265–19275.
24. Egler C, et al. (2009) Kinetic parameters of monoclonal antibodies ESH2, ESH4, ESH5,and ESH8 on coagulation factor VIII and their influence on factor VIII activity. J MolRecognit 22:301–306.
25. Deng Z, et al. (1999) A productive NADPðþÞ binding mode of ferredoxin-NADPðþÞ
reductase revealed by protein engineering and crystallographic studies.Nat Struct Biol6:847–853.
26. Vagin AA, et al. (2004) REFMAC5 dictionary: Organization of prior chemical knowl-edge and guidelines for its use. Acta Crystallogr, Sect D: Biol Crystallogr 60:2184–2195.
27. Kraulis PJ (1991) Molscript—a program to produce both detailed and schematic plotsof protein structures. J Appl Crystallogr 24:946–950.
28. Esnouf RM (1997) An extensively modified version of MolScript that includes greatlyenhanced coloring capabilities. J Mol Graphics Modell 15:132–134.
29. Nicholls A, Sharp KA, Honig B (1991) Protein folding and association—insights fromthe interfacial and thermodynamic properties of hydrocarbons. Proteins: Struct, Funct,Genet 11:281–296.
30. Brunger AT (1992) Free R-value—a novel statistical quantity for assessing the accuracyof crystal structures. Nature 355:472–475.
31. Engh RA, Huber R (1991) Accurate bond and angle parameters for X-ray proteinstructure refinement. Acta Crystallogr, Sect A: Found Crystallogr 47:392–400.
Alte et al. PNAS ∣ November 9, 2010 ∣ vol. 107 ∣ no. 45 ∣ 19265
BIOCH
EMISTR
Y
Dow
nloa
ded
by g
uest
on
Aug
ust 6
, 202
0