Embed Size (px)
Citation preview

J. Exp. Biol. (1969), 51, 407-430 4 0 7
With 3 plates and 5 text-figures
Printed in Great Britain
A FUNCTIONAL ANALYSIS OFTHE AQUATIC AND AERIAL RESPIRATORY MOVEMENTSOF AN AFRICAN LUNGFISH, PROTOPTERUS AETHIOPICUS,WITH REFERENCE TO THE EVOLUTION OF THE LUNG-
VENTILATION MECHANISM IN VERTEBRATES
BY B. R. McMAHON
Department of Zoology, University of Bristol*
(Received 30 December 1968)
Recent palaeontological evidence (Jarvik, 1967; White, 1966) indicates that theDipnoi should be regarded as a specialized group of fishes, which have been structur-ally different from the Rhipidistia (now thought to have included the prototetrapods)for as long as the fossil record can yet show. However, the homology of the dipnoanlung with that of the tetrapod, and the similarities of organization and developmentbetween living Dipnoi and Amphibia, might indicate a closer relationship between thetwo groups than can be revealed by palaeontological studies. The level of organizationseen in the living Dipnoi is not thought to be basically different from that of theDevonian forms and a study of the physiology of these animals must expand ourknowledge of the development of vertebrate physiological mechanisms. Since theDipnoi are adapted to a habitat very similar to that in which the first terrestrial verte-brates are thought to have evolved, and since they possess functional aerial and aquaticrespiratory systems, a study of respiratory function in these animals may help toelucidate the manner in which the ancestral tetrapods became able to colonize theterrestrial habitat.
Recently our knowledge of the mechanical basis of the respiratory pumps in fisheshas been very greatly increased by the introduction of modern techniques which allowthe accurate measurement of respiratory parameters with the minimum of disturbanceof the fish and the minimum distortion of the parameter under study. (Hughes &Shelton, 1958; Hughes 1960a, b; Saunders, 1961; Hughes & BaUintijn, 1965). Ventila-tion of the gills in teleosts (Hughes & Shelton, 1958; Hughes, i()6ob) and in elasmo-branchs (Hughes, 1960 a) has been shown to be achieved by the interaction of twopumps: a buccal pressure-pump forcing, and an opercular suction-pump drawing,water over the gills. That such a mechanism is basic in the vertebrates is indicated bythe presence of an essentially similar mechanism in the cyclostomes (Roberts, 1950).The involvement of a buccal force-pump in both air-breathing and water-breathing inthe amphibia has been demonstrated by Willem (1920 a, b, 1929) and the existence ofa buccal force-pump component has been described by Grigg (1965 a) and Johansen,Lenfant & Grigg (1967) in the breathing movements of the Australian lungfish Neo-ceratodus and by Jesse, Shub & Fishman (1968) in those of the African lungfishProtopterus sp. Bishop & Foxon (1968) have also demonstrated the importance of the
• Present address: Department of Biology, University of Calgary, Calgary, Alberta, Canada.

408 B. R. MCMAHON
buccal force-pump in both air-breathing and water-breathing in the South Americanlungfish Lepidosiren. The only other functional analysis of the mechanics of dipnoanrespiration was that of Dubois (1892) on air-breathing in the aestivating Protopterus.Szarski (1962) discusses possible mechanisms of ventilation in the Crossopterygii butquotes no experimental work. No complete analysis of the breathing mechanisms hasbeen published for any dipnoan and this paucity of information leaves a serious gapin our knowledge of the development of respiratory mechanisms in the vertebrates.
In the present study the mechanisms of aerial and aquatic respiratory movementsin Protopterus aethiopicus are described and compared, and the development of a lung-ventilation mechanism from the branchial irrigation mechanism of the water-breathingvertebrates is discussed. To avoid confusion in describing the movements of air andwater through the buccal area of air-breathing fishes, the movement of water acrossthe gills of these fishes will be termed 'irrigation' (Saunders, 1961) instead of thecommonly used 'ventilation* (Hughes & Shelton, 1958). In this account the term'ventilation' will be used only to describe the movement of air into and out of thelungs.Four main lines of approach were used:
(A) Anatomical studies of the head and branchial region.
(B) Analysis of X-ray cinematograph studies of the animal, during the respiratorycycles.
(C) Analysis of simultaneous recordings of the pressures developed in opercular,buccal and intra-pulmonary cavities.
(D) Analysis of the electromyograms, recorded from muscles active in the repira-tory cycles.
METH0D8
The animals used in this study were collected from the margins of Lake Victoria inUganda and transported to England by scheduled air services. They were maintainedin well-aerated water at a temperature of 25 ± i° C. The animals were fed on a diet ofox heart or liver, plus an occasional live fish when these were available. Animals werestarved for 1-2 days before any operative or experimental procedures were carried out.The animals were identified as Protopterus aethiopicus from the data presented inSterba (1962) and in Greenwood (1958).
Two methods were used to determine the gross structure of the head and branchialregion. Dissection revealed the position and attachments of the major muscular,ligamentous and skeletal elements, and this information was supplemented by the studyof thick sections cut transversely and longitudinally (using a circular saw) from thebodies of deep-frozen animals.
The pressures developed in buccal and opercular cavities were recorded by amethod adapted from Saunders (1961). A flexible plastic cannula (Portex PP. 160) wasintroduced into the buccal cavity by way of a hole drilled through the posterior snoutregion using a dental drill and handpiece. The size of the burr was chosen to producea hole that fitted the cannula tubing closely. The hole was drilled from a point mid-way between the snout and eyes and passed downward through the skull entering thebuccal cavity immediately posterior to the upper tooth-plate. The cannula was passed

Respiratory movements of an African lungfish 409
through the hole, out through the open mouth and a flange was formed on the proximalend. This end of the cannula was then drawn back into the buccal cavity until theflange fitted tightly against the buccal roof. A short length of close-fitting rubbertubing was slid down the cannula until it fitted closely against the outside of the head,thus anchoring the cannula in position. The opercular cannula was introduced throughthe roof of the opercular cavity by means of a large-bore hypodermic needle whichwas passed in through the tissues of the opercular roof and out through the opercularflap. The distal end of the cannula was forced into the bore of the needle and, as theneedle was carefully withdrawn, was carried out through the opercular roof. Theproximal end of this cannula had already been flanged and fitted with a fine rubberdam-washer. These were drawn up tight against the tissues of the opercular roof andsecured in position with a length of rubber tubing, as for the buccal cannula. Cannulaeinserted in this manner were well tolerated by the animals and remained patent andleak-free for a number of days at least, and in one case a patent buccal cannula wasretained for over 6 months.
It was planned to record the pressures from a number of different sites in eachcavity, but in fact the choice of sites was strictly limited. Satisfactory placement of theopercular cannula was only possible via the dorsal cavity wall. Implantation laterallythrough the operculum or through the opercular valve caused distortion of the oper-cular wall and impeded respiratory movements and flows. Implantation through theventral opercular region involved passing the cannula through muscle systems foundto be active during the respiratory cycles. Possible placement of the buccal cannulawas also found to be limited. Cannulation at the anterior end of the snout region intro-duced the cannula into the buccal area in front of the upper tooth-row, and thus infront of the buccal pressure-seal mechanism (discussed later), with the result that nopressure changes were recorded. Successful recordings were made from implantationsentering the buccal cavity immediately posterior to the tooth-row but implantationsa few millimetres further back often caused damage to the brain or associated structures.
In a limited number of experiments a cannula constructed from a No. 18 gaugehypodermic needle attached to length of flexible cannula tubing was inserted throughthe dorsal segmental muscle and into the anterior median sac of the lung. This methodof cannulation was hazardous, however, due to the extremely vascular nature of the lungwall, and it was discontinued after a minimum number of repeatable results had beenobtained (3 animals). Attempts were also made to cannulate the lung by means of acannula passing in through the glottis and pneumatic duct. This procedure failed,however, for the cannula appeared to irritate the walls of the duct and over-productionof mucus occurred blocking the duct, and eventually the cannula.
The insertion of all cannulae was performed under deep anaesthesia induced byimmersion of the animal in a 0-5—i-o% solution of tricaine methane-sulphonate(M.S. 222. Sandoz). Anaesthesia and operation times were kept as short as possible, asthe animals could not breathe during the anaesthesia and rapidly became anoxic.Changes in pressure were detected using either Sanborn 267 B or Devices pressure-transducer systems, and the outputs of the transducers were displayed and recordedusing either a pen recorder or an oscilloscope and camera. During the recording ofbuccal and opercular pressures, cannulae and transducers were flushed and filled withfreshly distilled water in order to minimize the risk of damping due to the formation

410 B. R. MCMAHON
of air bubbles in the system. During the recording of intra-pulmonary pressure bothtransducer and cannula were filled with moist air.
Electromyograms (EMG's) of the muscles of the head and branchial region wererecorded by a method modified from Hughes & Ballintijn (1965). Initially insulatedpins were used as recording electrodes but as it was feared that these might causemechanical damage to the muscle fibres moving about them, they were replaced bypairs of insulated stainless steel wires. Pairs of wires, cleaned and hooked at the tip,were threaded through a hypodermic needle. This needle was inserted through the skinto the desired site and was then withdrawn carefully, leaving the electrodes hookedinto the muscle. The potentials recorded from the muscles were amplified and thendisplayed on the separate channels of four-trace or six-trace, chopped-beam oscillo-scopes. Normally one or more pressure recordings would be displayed with theEMG's from a number of different muscles (Fig. 4). Electrodes implanted as abovewere well tolerated by the animals and did not appear to cause any mechanical damageor other impairment of the respiratory processes.
X-ray cinematograph studies were carried out using a Watson R 600 Generator anda Watson Windsor tube holder and stand. The X-ray image was received on a 9 in.Phillips image intensifier which was filmed by an Arriflex II BV 35 mm. cine camera.The animal was filmed under water in a chamber constructed from thin Perspexsheeting. The chamber was made just large enough to hold the fish so that the smallsize and the thinness of the walls helped to reduce the loss of contrast caused by thewater and Perspex surrounding the animal. Films were taken of animals displayingboth aquatic and aerial respiratory movements from lateral and antero-posterioraspects. Films were usually shot at 16 frames/sec, on Pan F film. Developed filmswere viewed on an X-ray film projector, »and for the purpose of analysis and displayselected frames were enlarged and printed on high-contrast paper. Various combina-tions of K.V. and mA. were used to try to increase the contrast of the resulting films,but a high degree of contrast was in fact never really achieved because of the scatteringof the X-ray beam by the plastic and water surrounding the animal. A rather higherdegree of contrast was obtained in 'still' photographs taken on Ilflex film.
During the pressure and electromyographic recordings the animals were containedin a Perspex tank fitted with a movable partition which could be adjusted so as tocontain the animal in as short a length of the tank as was desired. The animal wasgiven sufficient room to manoeuvre its way to the surface to breathe, but not enoughto enable it to swim forcefully. The tank was narrow but had sufficient room for theanimal to turn without difficulty, for if the tank was too narrow the animal struggledviolently to turn and removed cannulae and electrodes. The water in the tank wasstatic but well aerated, and was maintained at about 250 C. in a thermostatically con-trolled room. Usually the animal remained in a resting condition, only moving to cometo the surface to take air. Sudden movements on the part of the observer could, how-ever, startle the fish into violent swimming in which cannulae and electrodes were lost.To minimize this danger either sudden movements were avoided and/or the experi-mental chamber was shielded with polystyrene sheeting, except for a small observa-tion window.

Respiratory movements of an African lungfish 411
RESULTS
(a) Anatomical studies
X-ray photographic and dissection techniques were used to determine the grossskeletal and muscular anatomy of the head and branchial region (PI. 1; Text-fig. 1).A detailed anatomical description of the cranial muscles in Protopterus has beenpublished elsewhere (Edgeworth, 1935) and will not be repeated here, but a shortaccount of the positions and principal points of attachment of major muscular andskeletal elements is included with results of the EMG experiments below. Certain
n.ca.of>. —
p.pi.
R.M.—
c. -
n.c. —
'i -
it. P-o
—tr
CHy.
d.e.
LAIa.
/ 10. S.O.
LAI
w£Mh\\\
G.Th\ b.bCHy.
Id.e/
p.
K>Li •
•̂
sq.
'S
.
B
- i.w.
•j P.M.1
V.
I-c.r.
PI-
c. TAy/o.—_JS£a
s.o. nTTv^^
c.r. ^ ^
11
/.M.
> C.H/.
G.Th.
r,.Th.R.C.
c.r.
Fig. 1. Diagrammatic representations of head and branchial anatomy of Protoptenu to showposition and attachments of muscles and skeletal structure. A. lateral aspect. B. Sagittal sec-tion. C. ventral aspect.
Abbreviations: a.o.p., ant-orbital process; b.b., branchial bars; c, ceratohyal; ex., chondro-cranium; c.r., cranial rib; d.e., dermal ethmoid;/., pectoral fin;/./)., fronto-parietal; i.v., innerwall of cranium; l.d.e., lateral dermal ethmoid; /./., lower jaw; n.c., nasal capsule; o., opercular;p.g., pectoral girdle; p.pt., palato-pterygoid; s.o., sub-opercular; sq., squamosal; v., vertebralcolumn; A.M., anterior muscle of cranial ribs; CHy., constrictor hyoideus muscle; G.Th.,geniothoracicus muscle; I.M., intermandibularis muscle; L.Ma., levatores mandibulae muscle,ant.; L.Mp., levatores mandibulae muscle, post.; P.M., Post muscle of cranial rib; R.C., Rectuscervicis; 1-5, principal attachment points of the constrictor hyoideus.
muscles, found to be active in the respiratory cycles, have not apparently been de-scribed previously. As the homologies of the musculature in the Dipnoi are uncertainthese muscles will be referred to by terms indicating their position and attachmentsuntil they can be named with greater certainty than is possible at present. Themusculature of the head region of Protopterus is well developed, perhaps as a con-sequence of the evolution of the powerful suctorial predation methods shown in thisform. The majority of the muscles were found to be active in both feeding and

412 B. R. MCMAHON
respiratory movements but the level of electromyographic activity recorded duringrespiratory movements was usually less than during feeding. The nomenclature usedin this account is taken from Edgeworth (1935)-
The skull of Protopterus (PI. 1; Text-fig. 1) was found to differ greatly in structurefrom that of the teleost and elasmobranch skulls described by Ballintijn & Hughes(1965) and Hughes & Ballintijn (1965). Compared with the primitive bony fishes muchreduction and fusion of bone has occurred in Protopterus. This indicates that theskeletal couplings shown to be important in the respiratory mechanisms of the teleostand elasmobranch fishes by the above authors are likely to be of much less importancein Protopterus. This reduction of bone is particularly noticeable in the opercular regionwhere the opercular bones are very small and the opercular wall is mostly composedof muscle and elastic tissues. The jaw suspension is autostylic, hyomandibular,maxillary and premaxillary bones having been lost. The lower jaw is short but welldeveloped and articulates directly with the quadrate. The hyoid arch is reduced to aceratohyal element only. This is a •well developed arch of bone connected both to thecranium and to the dorsal flange of the lower jaw by means of ligaments. Ventrally itsupports the muscular tongue.
Two further skeletal elements were found to be concerned in the production of therespiratory movements. The pectoral girdle is a partly ossified arch underlying theposterior buccal cavity floor and the opercular cavity. Muscles from the ceratohyal,the branchial apparatus and the cranial ribs are attached to its surface. The cranial ribsare large ossified structures, peculiar to the Dipnoi, which articulate with the occipitalregion of the skull and are well anchored in the surrounding muscular tissues.
The ribs of Protopterus are not well developed (PI. 1) and appear to play no part inthe respiratory cycles.
(b) X-ray cinematograph studies
Still radiographs of the living animal show very clearly the outline of the lung andthe principal skeletal elements of the body. (PI. 1). Though the contrast was not high,X-ray cine films showed clearly the movements of air and of the skeletal elements ofthe anterior region during the respiratory cycles.
During the aquatic respiratory cycles the movements of both the ceratohyal andpectoral girdle arches could clearly be seen. Both arches were displaced upward andthen downward through the resting position during the course of one aquatic respira-tory cycle. It was hoped that films taken from the antero-postero (dorso-ventral) axiswould show movements of the opercular bones in a lateral plane, but sufficient con-trast was not developed to show this movement clearly.
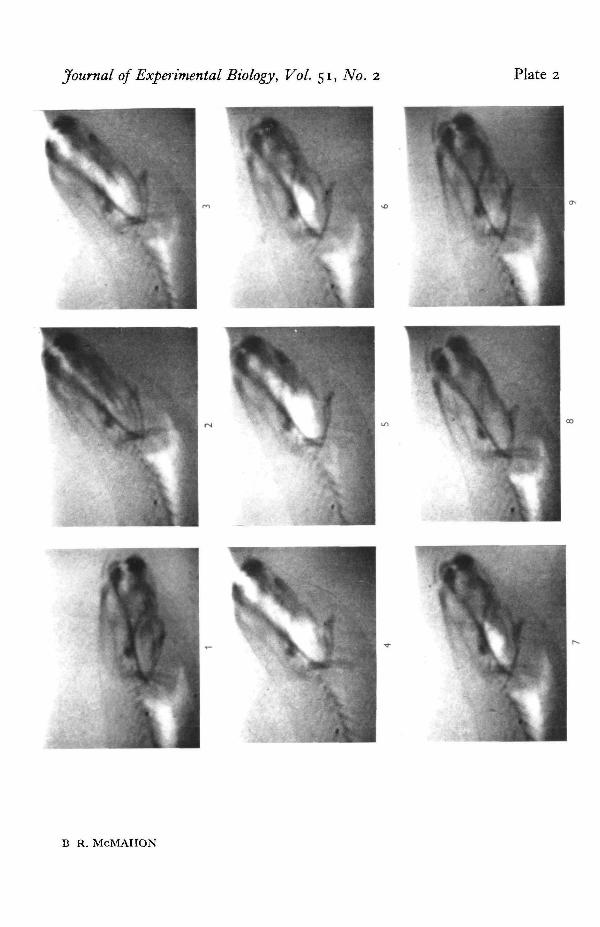
The principle events in an air-breathing cycle, as demonstrated by analysis of theX-ray films, are shown in PI. 2. The animal was first seen to flex the anterior part ofthe body so that the snout was thrust above the surface (PI. 2, 1-2). Animals in deeperwater do not show this head flexure but rise straight up to the surface. At the surfacethe mouth was opened and the buccal cavity expanded by a posterior-ventral deflexionof the ceratohyal and pectoral girdle elements which caused a downward displacementof the buccal cavity floor thus drawing air in through the open mouth (PI. 2, 2—4).Towards the end of this buccal cavity expansion phase the volume of the lung appearedto decrease considerably, indicating that pulmonary gas was passing from the lung

Respiratory movements of an African lungfish 413
and mixing with the atmospheric air in the buccal cavity at this time. (PI. 2 3-5). Atthe end of this expiratory phase the animal closed the mouth and lowered the headbelow the surface. An antero-dorsal movement of the buccal floor skeletal elementswas now seen, compressing the air contained in the buccal cavity and forcing it back,through the glottis and pneumatic duct, into the lungs (PI. 2 5-7). Following thisinspiratory phase the animal sank to the bottom of the container.
Expiratlorv Inspiration-10°
"S
"3
+10° I5
+20°
ITime (sec.)
Fig. 2. Movements of the skeletal elements of the buccal floor and angle of head (movement toand from surface) during the latter part of an air-breathing cycle. As plotted from and X-ray-cine film.
The movements of the skeletal elements of the buccal floor observed during theair-breathing cycle were similar to those seen in the aquatic cycles but were of greateramplitude and could be measured accurately. Text-fig. 2 shows the displacement ofthe skeletal elements of the head and branchial region plotted together with the degreeof flexion of the anterior part of the body for comparison. The plots commence as theanimal reached the surface and opened the mouth into the air. Initially all the elementsmoved together but the degree of movement of the pectoral girdle was greater andmore prolonged, producing considerable expansion of posterior buccal cavity. Move-ments of the cranial ribs were seen closely following those of the pectoral girdle buttheir exact function was not clear. The movements of the buccal structures thusexpanded the buccal cavity during the expiratory phase of lung ventilation. Duringinspiration the lower jaw moved upwards, closing the mouth. Following this anantero-dorsal movement of the ceratohyal was seen, raising the anterior part of thebuccal cavity floor and forcing the contained air back to the posterior part of the buccalcavity. Antero-dorsal movement of the pectoral girdle followed almost immediately,forcing the contained air back into the lung. Inspiration in this animal was thus seen toinvolve a buccal force-pump mechanism.
Expiration was seen to occur with the mouth open and atmospheric air entering thebuccal cavity. Some admixture of pulmonary and atmospheric air must then occur in

414 B. R. MCMAHON
the buccal cavity. However, the expulsion of gas is often forceful and an efficient tidalexchange during lung ventilation is indicated by the results of spirometry experiments(Jesse et al. 1968) and by the monitoring of pulmonary oxygen concentration (B. R.McMahon, 1969). In both cases 60-80% exchange of pulmonary gas is indicatedat each ventilation.
(c) Pressure studies
The pressures developed in buccal, opercular and intrapulmonary cavities weremonitored during both aquatic and aerial respiratory cycles. All recordings were madefrom non-anaesthetized, free-swimming animals. Considerable differences were seenbetween the pressure waveforms recorded during air-breathing and water-breathing(Text-fig. 3).
fi.pMm..Sec
10
JUL. mm. Hg
AIR AIR AIR
B
O.P.
Sec.
B.P.
+-T 10 mm. Hg
0 mm. Hg
Fig. 3. Recording of pressures developed in the buccal and opercular cavities during aerial andaquatic respiratory movements. A. Slow recording to show variation in amplitude of aquaticwaveforms between air-breathing movements. B. Fast recording to show details of pressurewaveforms from aerial and aquatic respiratory cycles.
BJ3., buccal pressure; OJ1., opercular pressure; Air, aerial respiratory cycle, I , calibration10 mm. Hg.; G., branchial respiratory cycle; L,, L», L,, stages of air breathing cycle.
(i) Pressures recorded during aquatic {branchial irrigatory) movements
Little difference was seen in the basic form of the pressure waveform recorded fromnine different animals Considerable variation occurred, however, in the amplitudeand frequency of the recorded waveforms even in the same animal during periods ofapparent rest. Such fluctuations could be correlated with variation of external orphysiological activity of the animal. Animals newly introduced into the experimentalchamber or recently disturbed by experimental procedures showed branchial irriga-tory movements of high frequency and amplitude. After a period of acclimatizationboth frequency and amplitude fell to a lower level, but would rise again if the animalwere disturbed, or subjected to certain forms of respiratory stress. Variations of both

Respiratory movements of an African lungfish 415
frequency and amplitude of the recorded waveforms were also observed in acclimatizedanimals apparently at rest, and these could often be associated with stages of the air-breathing cycles (Text-fig. 3 A). Immediately after an air breath both frequency andamplitude of the waveforms were high but decreased steadily until just before thenext air breath when they were seen to rise again.
During branchial irrigation, pressure changes were detected from the buccal andopercular cavities but never from the intra-pulmonary cavity. The waveforms recordedwere basically similar to those recorded by Hughes & Shelton (1958) and Hughes(1960 a, b) for teleost and elasmobranch fishes in that interacting buccal and opercularpressure changes combined to produce a differential pressure across the gill resistancetending to force water over the gills throughout most of the cycle. While the amplitudeof the pressure changes recorded from Protopterus was generally higher than thatobserved for other fishes, the opercular pressures were generally lower than thoserecorded from the buccal cavity. This suggests that the buccal force-pump is dominantin branchial irrigation in this animal. A similar condition is seen in the eel Congerconger (Hughes, 1960^).
(ii) Pressure waveforms recorded during aerial respiratory {lung ventilation) movements
The waveforms recorded during the air-breathing cycle were more complex thanthose of branchial irrigation. Three separate buccal and opercular waveforms co-incided with a single depression and subsequent re-establishment of the intra-pulmonary pressure, which was seen to remain positive throughout the rest of thecycle. Expiration was seen to occur between two of the waveforms, while inspirationoccurred simultaneously with a large pressure increase in both opercular and buccalcavities (Text-fig. 5B).
In both branchial and lung-ventilation cycles the phases of the pressure waveformsrecorded from the cavities could be related to specific events in the respiratory cycles,but as much additional information can be provided by the analysis of the results ofthe EMG experiments; the functional significance of the pressures will be discussedtogether with the results of the EMG studies later in this work.
(d) The analysis of activity in the respiratory muscles
The evidence from the X-ray and pressure studies outlined above indicates that thebasic mechanism of both aerial and aquatic respiratory cycles involves movements ofthe skeletal elements of the head and branchial region resulting in the development ofinternal pressures which serve to irrigate the gills or to ventilate the lungs. Recordingshave been made of the activity in the muscles of the head and branchial region, simul-taneously with recordings of the pressures developed (Text-figs. 4 and 5), to elucidatethe nature of the muscle system powering the respiratory pumps. The method ofrecording used here can give direct evidence of electrical activity only at the electrodesite, and thus monitors only a very small fraction of the possible motor units involvedin the activity of the whole muscle. For this reason it is imperative to examine therecordings taken from a large number of insertions, taken from different sites in thesame muscle, in more than one animal, before conclusions can be drawn as to theactivity of the muscle as a unit. The diagrammatic representation of the results seen
27 Exp. Biol. 51, 2

416 B. R. MCMAHON
in Text-figs. 5 A and B is compiled from recordings taken from a large number ofinsertions in six animals and indicates the period and relative strength of activity ofthe muscles concerned together with simultaneous recordings of respiratory pressuresand events. It must be further noted here that these are electrical signals and are onlyindirectly related to the degree of contraction produced by the muscle. Other factorssuch as the size of the muscle involved and the degree to which it is affected by theaction of other muscles of the system (and the state of skeletal couplings) may alsohave a considerable effect on the actual force transmitted by the contracting muscle.
0 25 In./sec.
6.P. I
0-50
0-25
10 mm. Hg
Fig. 4. Pressures developed in buccal, opercular and intrapulmonary cavities recorded withelectromyographic activity of the head and branchial muscles during aerial and aquatic respira-tory movements.
Abbreviations: D.P., differential pressure between buccal and opercular cavities; B.P.,pressure in buccal cacity; O.P., pressure in opercular cavity; I.P.P., intra-pulmonaxy pressure;g., aquatic respiratory movement (branchial irrigation); I. aerial respiratory movement (lungventilation).
Activity in: (i) rectus cervicis muscle, (a) geniothoracicus muscle, (3) levator mandibulaemuscles, (4) constrictor hyoideus muscle, (5) posterior muscle of cranial nb, (6) anterior muscleof cranial rib, (7) inter mandibularis muscle, (8) retractor mandibulae muscle, (9) retractoranguli oris muscle, I calibration = 10 mm. Hg.
The majority of the muscles of the head and branchial region were found to beactive in the respiratory cycles. Som'' muscles, notably those of the branchial basket,were too small or too difficult of access to be investigated here. Whereas it is probablethat they play a minor part in the fine control of the respiratory processes, it is con-

Respiratory movements of an African lungfish 417
tended here that they play no major role in the production of the respiratory move-ments. The muscles investigated are listed below and a short account given of theirpositions, principal points of attachment and the phase of the cycles in which they wereactive. The results are then incorporated into a functional analysis of the respiratorymechanisms of this animal.
The levatores mandibulae muscles
In development this is a single sheet of muscle which divides to form anterior andposterior sections. The levator mandibulae anterior is a very large muscle attachedcranially to the ventral side of the lateral dermal ethmoid and the supraorbital process,and to the dorsal side of the fronto-parietal. Ventrally it is inserted into Meckel'scartilage and into the posterior flange of the lower jaw. The posterior part is alsoattached to the posterior flange of the lower jaw and is inserted dorsally into thecranium and the squamosal. The position of these muscles is shown in diagrammaticform in Text-fig. 1 A.
No difference in the activity periods of the anterior and posterior parts of the musclecould be detected; both were active in aerial and aquatic respiratory movements butat a fairly low level of activity. (Higher levels were routinely recorded when theanimal' snapped' during feeding or attack.) It was concluded that because of the verylarge size of these muscles a low level of activity was sufficient to meet the needs ofthe respiratory system. Activity in this muscle occurred prior to and during thedevelopment of pressure in the buccal cavity. The action of the muscles would appearto lift the lower jaw, closing the mouth and beginning to raise the ceratohyal andinterconnected structures. By clamping the lower jaw tightly shut the action of thismuscle facilitates the development of the buccal pressure-seal mechanism, and alsoprofoundly affects the action of the buccal floor musculature. The action of thiscoupling will be discussed later.
The retractor anguK oris muscles
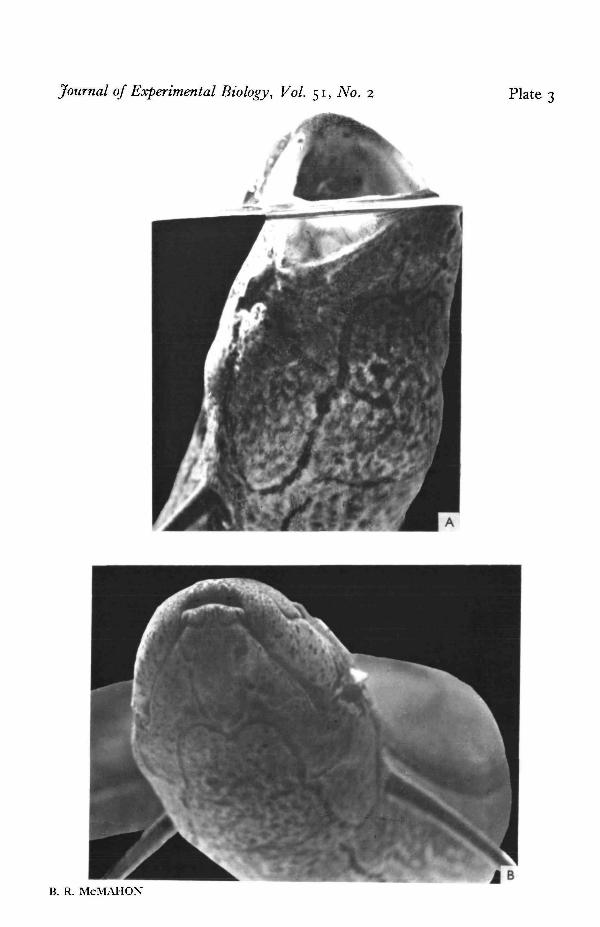
This muscle is inserted into the upper lip at the angle of the jaw and into thecranium dorsally. The upper lip overlaps the lower lip laterally producing a fold ofskin as shown in PI. 3. The activity of this muscle, which is concurrent with that of thelevator mandibulae, is thought to tighten this fold of skin, thus preventing any openingof the mouth and effecting a pressure seal laterally along the buccal cavity.
The retractor mandibulae muscles
Formed in development from the constrictor hyoideus muscle sheet, this musclein the adult is inserted via a ligament on to the posterior ventral part of the lower jaw.Dorsally it lies between the opercular bones and is connected to them. In this regionit overlies the constrictor hyoideus muscle sheet and abuts on to this muscle in theregion of the opercular wall (Text-fig. iA). Activity in the retractor mandibulaemuscle was always associated with maximal depression of the lower jaw as is seen inthe 'yawning movements' discussed below. Such maximal opening of the mouth wasrarely seen during the respiratory cycles but was occasionally seen, together with low-level activity in the retractor mandibulae muscle, in the expiratory phase of lungventilation.
37-a

418 B. R. MCMAHON
mm. Hg
P.P. +10
0
+10
0
-10
+10
0
-10+10
Mouth
Operc.
Phase
- 1 0Sec.
OpenShutShut
Open
W
UA.
R.A.O.
C.Th.
R.C.
IM.
KM.
P.M.
CJHy.
IfKMIIII
nun iamnt
wini i n
II M i l I
Fig. 5 A
Fig. 5. Combined data from pressure, electromyograms and observation experiments to show theinterrelation of muscular action and pressure development during the aquatic and aerialrespiratory cycles.
A. Aquatic cycle. Pressures and electromyograms recorded during the aquatic cycles.P.P. intra-pulmonary pressure. O.P. opercular pressure. B.P. buccal pressure. D.P. differentialpressure between buccal and opercular cavities.
Activity in: L.M. levator mandibulae muscle, R.A.O. retractor anguli oris muscle, G.Th.geniothoracicus muscle, /JVf. intermandibularis muscle, A.M. anterior muscle of cranial rib,P.M. posterior muscle of cranial rib, C.Hy. constrictor hyoideus muscle.
B. Pressures and electromyograms recorded during the movements of an air-breathing cycle.Abbreviations as in A.

Respiratory movements of an African lungfish 419
The mtermandibularis muscles
These small muscles are inserted on the inner surface of the lower jaw and con-nect in the mid-line (Text-fig. 1C). They were active during the development ofpressure in the buccal cavity and probably served both to tense the rami of the lowerjaw and to limit the movement of the overlying ceratohyal and associated structures.
P.P.
O.P.
°perc
LM. 11R.AO. "C.Th.R.C.I.M.AM.PM.
C.Hy.
•ma '111111I11
• • • M l• • • • I I*mmm
Hill
iiBan•
I I I I
1111 1 1 11 nirraui
Expiration Inspiration Inspiration
Fig. 5B. For legend see Fig. 5 A.
Irrigation
The geniothoracicus muscle
A single muscle connected by a broad ligament to the mid-ventral portion of thelower jaw and by loose ligamentous connexions to the ceratohyal. This muscle runsmid-ventrally to become inserted in the ventral segmental muscle in the pectoralregion. (Text-fig. 1A-C). The muscle is also connected by ligaments to the overlyingconstrictor hyoideus muscle sheet. Activity in this muscle was concurrent with that inthe rectus cervicis and intermandibularis muscles, and was associated with an increaseor decrease of pressure in the buccal cavity, depending on the action of the levatormandibulae muscles. The action of this coupling mechanism will be discussed later.
The rectus cervicis complex
This is a broad mass of muscular tissue containing several transverse inscriptionswhich forms the basis of the tissues of the tongue and is connected to the ceratohyalanteriorly and the pectoral girdle posteriorly (Text-fig. 1B). The recorded period ofactivity was similar to that of the geniothoracicus muscle. This muscle complex wasalso affected by the action of the levator mandibulae muscles.

420 B. R. MCMAHON
The constrictor hyoideus muscles
This is a broad muscle sheet surrounding the branchial region ventrolaterally, withattachments to almost all adjacent structures. Postero-laterally the muscle is attachedto the opercular bones and forms the wall of the opercular cavity. Postero-ventrallythe muscle is firmly attached to the skin along a line running ventrally from theopercular margin halfway to the ventral mid-line. From this point the muscle runsfree over the ventral muscles until it meets and fuses with its partner from the otherside. Anteriorly the muscle is loosely connected to the lower jaw by ligaments and isinserted along the ventral margin of the ceratohyal. Ventrally the muscle abuts on tothe intermandibularis muscle. Some of the attachments of this muscle sheet are shownin Text-fig, i A and C. The action of the muscle is complex and occurs at differentperiods in aquatic and aerial cycles, and will be discussed more fully later.
Un-named muscles
(1) The anterior muscles of the cranial ribs
This is a block of muscle connecting the pectoral girdle with the cranial ribs oneach side (Text-fig, i B), and forming part of the walls of the buccal cavity posteriorlyand the opercular cavity ventro-laterally. Activity in this muscle was seen at differenttimes in the aquatic and air-breathing cycles and was associated with both compressionand expansion of the buccal cavity.
(2) The posterior muscle of the cranial rib
Though very little modified from the surrounding segmental muscle, this musclewhich connects the cranial ribs with this segmental muscle (Text-fig. 1B) is alwaysfound to be active in the respiratory cycles. Its period of activity was similar to, butslightly preceding, that of the anterior cranial rib muscles.
Muscles of the glottis
These were described by Edgeworth (1935), but successful electrode implantationproved to be extremely difficult. One record, however, indicated the action of twomuscle systems in the glottis region, one continually active and the other active onlyduring ventilation of the lungs. This would suggest that the glottis was closed bymuscular activity except at the moment of lung ventilation when it was openedactively by dilator muscles. No confirmation of this was obtained but the existence ofdilator and constrictor muscle systems of the glottis were noted by Edgeworth (1935).Active closure of the glottis between ventilations was indicated by the occurrence ofa maintained pressure of 10-15 cm. of water in the lung.
Segmental muscles
Though EMG's were recorded from the flank muscles during swimming activity,no recordings were made which showed activity of dorsal or flank segmental musclewhich could be associated with respiratory movements.
In order to simplify subsequent discussion it is possible to divide the muscles dis-

Respiratory movements of an African lungfish 421
cussed into groups of muscles which were active at the same period of the cycle. Thesegroups are:
(a) Muscles involved in the closure of the lower jaw, including the levatoresmandibulae (L.M.) and the retractor anguli oris (R.A.O.).
(b) The muscles of the anterior buccal floor region, including the genio-thoracicus(G.Th), the rectis cervicis (R.C.) and the intermandibularis (I.M.).
(c) The muscle of the posterior buccal floor region, including the anterior andposterior muscles of the cranial ribs (A.M. and P.M.).
(d) The constrictor hyoideus muscle surrounding the buccal cavity ventrally andforming the muscular outer wall of the opercular cavity.
DISCUSSION
It is now possible to incorporate the results of the experiments in a functionalaccount of the mechanisms involved in the aerial and aquatic respiratory cycles.
(1) The aquatic respiratory cycle
Simultaneous recordings of the pressures developed in pulmonary, opercular andbuccal cavities, and the EMG recordings from the muscles active in the respiratorycycles are displayed in diagram form in Text-fig. 5 A. Differential pressures and theperiods of mouth and opercular closure are also indicated.
Activity was first recorded from the jaw-closing muscles (L.M. and RAO). Con-traction in these muscles raised the lower jaw, closing the mouth and pressure-sealingthe buccal cavity. Pressure experiments indicated that the tip of the tongue fitted intoa groove immediately posterior to the upper tooth-row as the mouth was closed, andprovided a further pressure seal anteriorly. A slight increase in both buccal andopercular pressures resulted from these movements.
Further increase of pressure was now observed, associated with activity in themuscles of the anterior buccal floor region. From purely anatomical evidence theactivity of these muscles should cause a depression of the skeletal elements of thebuccal floor and thus expansion of the buccal cavity, but here the muscles wereobviously active in the compression phase of the cycle. This apparent anomaly wasresolved when the effects of the continued activity in the jaw-dosing muscles wasconsidered. These powerful muscles were holding the lower jaw tightly shut. Lowerjaw, ceratohyal and pectoral girdle (the principal moving parts concerned with buccalfloor movement) were all interconnected by muscular and ligamentous tissues, andtherefore were fused in position by the immobility of the lower jaw. This couplingthus ensured that no depression of the buccal floor could occur at this time. Con-traction in these muscles must therefore have tensed the buccal floor structures,causing the upward deflexion seen in the X-ray studies, and the observed buccalcompression (Text-fig. 5 A, phase 2). Initially the pressure increased concurrently inboth cavities but later pressure increase was confined to the buccal cavity (phase 3).This produced a differential pressure between buccal and opercular cavities, whichcould have caused a respiratory water flow. In fact the opercular wall was seen to bedistended and the opercular flap forced open by a stream of exhalent water at this time.
As peak pressure was reached in the buccal cavity, activity ceased in the jaw-closing

422 B. R. MCMAHON
muscles. Activity could still be recorded from the buccal floor musculature (A.M. andP.M.). As the lower jaw was freed from the restraint of the jaw-closing muscles,activity in the buccal musculature could now draw down the buccal floor and expandthe buccal cavity. The buccal pressure falls sharply at this time, and as the buccalfloor is depressed the mouth opens slightly, causing water to enter the buccal cavityreplacing that formerly passed over the gills. Activity is now recorded from the con-strictor hyoideus muscle. Contraction in this muscle closed the opercular flap andmay have limited the expansion of the buccal cavity. A slight increase in opercularcavity pressure was often associated with the closure of the operculum. At this timea considerable negative differential existed between the pressure in the buccal andopercular cavities. It is possible that this temporarily induced a reverse flow of wateracross the gills. This back flow was unlikely to be large, because of the closure of theopercular flap and the small size of the opercular cavity.
At the end of the cycle the pressure in the buccal cavity began to rise. No activitywas seen in the musculature at this time and this movement was interpreted as beinga passive return of the skeletal elements of the buccal floor to the resting position. Asthe pressure in the buccal cavity rose, a pressure decrease occurred in the opercularcavity due to the elastic recoil of the operculum at the end of active closure. A second,smaller, positive differential pressure pulse thus occurred at this time which couldcause a second respiratory flow to occur.
Irrigation of the gills in Protopterus is thus seen to involve the action of two pumpsoperating on either side of the gills. The mechanism involved is similar to that de-scribed by Hughes and other authors for teleost and elasmobranch fishes. The basicpattern of ventilation in the dogfish, as described by Hughes & Ballintijn (1965),comprises three main phases: an active expiratory movement which causes distensionof the parabranchial cavity; a passive inspiratory movement; and finally a more activeinspiratory phase. A similar sequence is seen in Protopterus but no passive inspiratorystage is seen following expiration. Continued activity in the anterior branchial musclesand the release of the coupling effect are responsible for the initial movements ofinspiration.
By reason of the differences in the anatomy of these two forms it is very difficult tocompare the activity of specific muscles or muscular systems. However, the action ofthe adductor mandibulae of the dogfish and the levator mandibulae of the lungfishwould appear to be similar, as would the action of the constrictores hyoideus andbranchiales of the dogfish and the constrictor hyoideus of the lungfish. The action ofthe ventral (buccal floor) musculature appears to be less important in the dogfish thanin Protopterus, but generally the muscle systems involved are similar in basic pattern.Perhaps the greatest differences were found in the degree of involvement of thepectoral girdle, which is immobile in the dogfish.
The mechanism is also basically similar to that described by Ballintijn & Hughes(1965) for the trout. Here, though the anatomical differences are greater, the pressurewaveforms are still similar to those recorded from Protopterus but the method of theirproduction differs in detail. The most important differences are the absence of pectoralgirdle movement in the trout and the absence in Protopterus of the complex couplingsbetween the skeletal elements of the jaw suspension and operculum. The divergentmorphology of the two forms does in fact make comparison very difficult. It would be

Respiratory movements of an African lungfish 423
very interesting to record the mechanisms shown by anatomically (not phylogenetic-ally) intermediate forms such as the eels (the recorded pressure waves of Conger conger(Hughes, io,6o£) are rather similar to those recorded for Protopterus) in which boththe branchiostegal apparatus and the opercular suction pump are reduced in functionalimportance, and such as the Australian lungfish Neoceratodus, where the reduction ofthe opercular and hyomandibular bones is not so great as in Protopterus. A study ofthe respiratory mechanisms of the former animal would enable us to decide how muchof the pattern seen in Protopterus is typical of the Dipnoi, and how much is due to therather specialized nature of this lungfish. The basic pattern of the respiratory move-ments is also very similar to that exhibited by the early larval stages of the toad(Willem, 1920a, b).
A second type of aquatic movement was occasionally seen in Protopterus. The mouthwas opened maximally by activity in the buccal floor musculature and often in theretractor mandibulae muscles. The mouth was held open for periods of up to 1 min.and then allowed to close slowly. This movement was invariably followed by a typicalbranchial irrigatory movement. The movement was very similar to the yawning move-ments seen in many teleost fishes. In the teleost these movements have been demon-strated as being important in the depth-control mechanisms (McCutcheon, 1966), butsince they were often seen in the resting lungfish in the absence of depth change theirfunction in this animal remains obscure. If the response were to be respiratory (pul-monary) it would be of great interest, however, since this may have been the ' primitive'response which together with the air bladder in the teleost fishes has lost its respiratoryfunction and become concerned with the regulation of depth.
(2) The mechanism involved in the air-breathing movements
Pressure changes and electromyograms have also been recorded during a largenumber of air-breathing cycles. These results are expressed in diagram form in Text-fig. 5 B. At first sight the cycle seems very complex, including at least three differentstages. It is possible, however, to divide the cycle into three sub-cycles, each of whichcan be compared with a single aquatic cycle.
The first sub-cycle occurs as the animal approaches the surface prior to lungventilation. Activity was first recorded in the jaw-closing muscles, closing the mouthand pressure-sealing the buccal cavity. This activity was maintained and modifiedas was the subsequent action of the anterior buccal floor musculature deflecting thebuccal floor upwards, and compressing the water contained in the buccal cavity. Waterwas observed to be lost from the operculum at this time. Up to this stage no differencewas observed between this and a typical aquatic movement, but the animal was nowat the surface and the subsequent expansion phase occurred with the mouth openinginto the air. As air entered the mouth, pressure decreased very rapidly in the buccaland opercular cavities and activity ceased in the majority of the muscles. A decreasedlevel of activity in the rectus cervicis and posterior buccal floor musculature was suf-ficient to expand the buccal cavity as the incoming air offered little resistance to suchexpansion, thus no change in pressure was noted, though the expansion of the buccalcavity was almost maximal. Activity was also recorded from the constrictor hyoideusmuscle at this time, closing the operculum and preventing the entry of water frombelow.

424 B. R. MCMAHON
Towards the end of the expansion phase of this movement a decrease in pulmonarypressure was recorded. Pulmonary gas appeared to be held in the lungs under pressureby the action of the glottal sphincter muscles. These relaxed at this time allowing thecompressed gas to escape. This expiration is almost certainly aided by the naturalelasticity of the lung wall. Much smooth muscle has been described in the lungs ofProtopterus sp. (Poll, 1962) and Neoceratodus (Grigg, 1965 a) but it was not possible inthis study to show that active contraction of this muscle was associated with expiration.These records showed that expiration occurred during the expansion phase of the firstsub-cycle at a time when the mouth was open. In this way the lung was indirectly opento the atmosphere during expiration.
When the buccal cavity was fully expanded and the pulmonary pressure reducedthe next sub-cycle started immediately. No passive return of the skeletal elements ofthe buccal floor occurred between these two stages. Activity was immediately seen inthe jaw-closing muscles and a little later in the anterior buccal floor muscles, initiatinga buccal compression phase similar to that seen in aquatic cycles. However, concurrentactivity was also seen in the posterior buccal floor muscles which were now influencedby the coupling effect of the jaw-closing muscles and this became effective in the com-pression phase. This modification probably helped to produce the higher pressures(up to 20 mm./Hg.) apparently needed in lung ventilation. The resultant pressure riseoccurred in both cavities simultaneously and as the presence of air in the cavities oftenallowed the gill resistance to break down, a differential pressure between the twocavities was not always recorded.
Increased pulmonary pressure was also recorded at this time, indicating that thegas compressed in the buccal cavity was being forced into the lung. In some cases (asin Text-fig. 5B) a single inspiratory effort was not sufficient and a second, morepowerful, inspiratory stage followed the first. The stimulus which instigated a secondinspiratory stage would appear to have been pulmonary pressure, for in animals wherethe pulmonary cavity was experimentally kept at atmospheric pressure repeated in-spiratory efforts were seen and the animals remained at the surface; animals whichwere obliged to surface into mixtures containing no oxygen showed only a singleinspiration and then submerged often for quite long periods. A second expiration inone surface visit was never seen.
Following the final inspiration the expansion phase of that movement occurred afterthe head was submerged. The water thus drawn into the buccal cavity was passed overthe gills and lost from the operculum in a stage very similar to a typical irrigatorymovement. The movement is always present and is always more forceful than theusual aquatic respiratory movements. It is thought that its function is to expel thewater forcibly over the gills thus removing residual air from the system.
The inter-relation of aquatic and aerial respiratory mechanism
Though the pressure waveforms recorded during the aquatic and the aerial cycleswere very different in appearance, because of the different properties of the containedwater or gas respectively, examination of the EMG records and comparison with theevents as recorded by X-ray photography showed that the aerial cycle consisted of anumber of consecutive aquatic-type movements, each modified to serve a specificphase of lung ventilation.

Respiratory movements of an African lungfish 425
Relatively little modification is needed. The compression phase of the first move-ment is similar to those of typical aquatic cycles but serves an important function inremoving the water contained in the buccal cavity, which might otherwise enter thelungs and cause damage or drowning.
The subsequent expansion phase is prolonged, producing the maximal expansionof the buccal cavity necessary to encompass sufficient air to reinflate the lungs, andallowing expiration to occur while the mouth is open and the operculum closed. Thisfacilitated tidal exchange and prevented water entering through the opercular flapduring ventilation.
The compression phase of the second stage is modified to serve the function of lunginspiration. Essentially the air contained in the buccal cavity is compressed by a buccalforce-pump mechanism and forced through the pneumatic duct and into the lungs.The principal modification seen here is an alteration of the periods of muscular activityso that muscles normally active only in the expansion phase of the cycle (A.M., P.M.,C.Hy.) are now active earlier in the cycle, are affected by the lower jaw couplingmechanism, and are thus active in the compression phase. This modification producesgreater and more efficient compression as all the buccal floor muscles act together.The action of the constrictor hyoideus is particularly important at this time, for thecontraction of this muscle in the compression phase closes the opercular flap andprevents the loss of the highly compressed air through the operculum.
It is not known how these changes in the periods of muscular activity are effected.It is possible that as there is no passive return of the skeletal elements to a restingposition before this inspiratory compression begins (a passive return was described inthe aquatic cycles) and therefore the action of the powerful jaw-closing musculatureacts on a maximally expanded system. The upward movement of the lower jaw would,under these conditions, cause a stretching and tension in the buccal floor muscles andcould cause these to become active earlier than occurs in the aquatic cycle. This hypo-thesis would explain the difference between the aerial and aquatic compression phases,and would be a simple method of generating the high pressures needed for completegas transfer and thus efficient lung ventilation.
The expansion phase of the final inspiratory movement occurs as the animal sinksbelow the surface. The water thus drawn into the mouth is forced over the gills,flushing out any residual air which might interfere with further branchial irrigationor with buoyancy or postural regulation. This movement is very little modified fromthe normal aquatic cycle but is intimately associated with the air-breathing sequence.
The mechanism of lung ventilation in Protopterus is derived from a series of basicallyaquatic-type cycles, each of which is modified to serve a specific function in the air-breathing cycle. Basically, inspiration corresponds to the buccal compression phase ofthe branchial cycle while expiration occurs passively in the expansion phase of thecycle. Essentially the mechanism is based on, and easily derived from, a buccal-opercular-branchial pumping mechanism similar to that seen in all fishes (Hughes,19600,6; Hughes & Shelton, 1962) and in the amphibia (Willem, 1920a, b).
The evolution of the aerial respiratory mechanisms in the vertebrates
Of the many authors who have published observations on the form of the air breathin Protopterus, only one reports experimental work. Dubois (1892) examined the

426 B. R. MCMAHON
mechanism of air-breathing in aestivating Protopterus using a tambour fixed to themouth and levers attached to the body wall at various points. His account has neverbeen confirmed, but has provided the basis for a number of erroneous descriptions ofthe air-breathing mechanism found in this animal (Smith, 1931) and of schemes forthe evolution of aerial respiratory mechanisms in vertebrates (Willem, 1929). Byobservation through the open mouth and by the use of levers Dubois recognized thatthree stages occurred in the lung-ventilation cycle. He also observed that the glottiswas open during buccal expansion (just following the first compression of the air-breathing cycle in Text-fig. 5 B) and assumed that this was inspiration. He concludedthat as the mouth was open during inspiration then a buccal force-pump mechanism('deglutition') could not occur and that inspiration must occur by an aspiratorymechanism involving the use of the ribs. Dubois admitted that active dilation of thebody wall was an anatomically impossible solution but offered no real alternative. Inthis account it has been demonstrated that expiration, not inspiration, occurs at thistime and thus Dubois' assumptions are fundamentally incorrect. Willem, however(1929), accepted Dubois' main points and concluded that inspiration occurred partlyby the movements of the truncal ribs but mostly by the movements of the cranial ribs(which he equates with the abdominal movements of Dubois). In fact though move-ments of the cranial ribs are associated with the respiratory cycles (Text-fig. 2) thedirection of their movement is the opposite of that needed to substantiate Dubois' andWillems' account. No movements of the truncal ribs have been observed in the X-raystudies carried out on Protopterus and none were observed in a careful radiographicstudy on the air-breathing movements of Lepidosiren (Bishop & Foxon, 1968). It is inany case doubtful as to whether the small movements possible in the reduced truncalribs or in the cranial ribs could have more than a minor effect on pressure in the ex-tensive lung of Protopterus or Lepidosiren.
Szarski (1962) proposes another theory of the development of vertebrate lung-ventilation mechanisms. He quotes Schmalhausen (1957, 1958, not read by author)as deducing that the primitive air-breathing fishes utilized the hydrostatic pressure ofthe surrounding water as the lung-ventilation mechanism. Here, as the fish approachesthe surface at right angles, the differential hydrostatic pressure along its length isutilized to force the air from the lungs and out of the open mouth. The buccal cavityis then filled with air and the animal swims until the head is pointing downward; thedifferential hydrostatic pressure acting along the body is now imagined as forcing thebuccal air into the lung. While it is likely that the hydrostatic pressure could/have aminor beneficial effect on ventilation in a soft-bodied fish, it seems most unlikely thatsuch a mechanism could have provided efficient ventilation in the rigid-bodiedrhipidistian fishes. Schmalhausen, Szarski points out, considered that this mechanismwould be useless in air, and virtually useless in shallow water. Szarski maintains thatas this method was useless in air it was discontinued by the first amphibian animalswhich then relied on cutaneous exchange for both oxygen uptake and carbon dioxideexcretion. However, the habitat of the primitive amphibians is thought to have beenmostly in shallow, periodically hypoxic and hypercarbic water, with rare, essentialexcursions on to the land. Under tiiese conditions neither cutaneous exchange, northe lung ventilation method described by Szarski, would have been of use. It is morelikely that the cutaneous gas-exchanger was evolved because the primitive lung, though

Respiratory movements of an African lungfish 427
extremely efficient as an oxygen exchanger, was inefficient as a carbon dioxide excre-tion mechanism (Rahn, 1966), and this function, which was performed by the gills inthe air-breathing fish, was transferred to the skin as the gills lost function in theterrestrial habitat.
Finally it seems unnecessary to postulate the development of a new lung-ventilationmechanism when the existing buccal force-pump can be so easily modified to servethis purpose. Szarski argues that a mechanism by which the mouth and opercularcavity can be closed simultaneously is a prerequisite for the evolution of an aerialbuccal force-pump mechanism such as is seen in the recent amphibia. Indeed this isso, but such a mechanism is demonstrated here in the lungfish and must be presentin a great number of air-breathing teleosts. Szarski also considers that a ventilationmechanism where the atmospheric and pulmonary air are mixed in the buccal cavitywould be very inefficient, yet it has been shown that in Protopterus lung ventilation isvery efficient, as the tidal volume is 60-80% of the lung volume (Jesse et al. 1968;B. R. McMahon, 1969).
It is contended here that the primitive fresh-water fishes, in response to an occasion-ally hypoxic and hypercarbic environment, came to use the buccal force-pump,previously used only to power the branchial irrigatory flow, to pass oxygen-rich airfirst over the gills, secondly into respiratory pouches and finally into the developinglungs. The buccal force-pump, with only slight modification, could easily be used for thispurpose. This system is similar to that in use by recent amphibia except that the naresare not used. It is suggested that this method is most likely to have been that used by therhipidistian and prototetrapod forms. The use of the ribs, is shown not to be importantin the lungfishes and it is unlikely that it was of use in the proto-amphibian types. Theevolution of such an aspiratory mechanism is dependent on the possession of powerfuland movable rib structures. These were probably developed at a later stage in evolutionwhen the whole skeleton of the animal, including the ribs, was undergoing the massiveevolutionary changes needed to provide a new skeletal structure which could moreefficiently support the animal's mass on land. Such increase in rib area is seen in theearly amphibia, and it is possible that these animals had evolved the more efficientaspiratory ventilation mechanisms. We can then imagine that these may have becomeless important in the recent amphibia, together with adaptation to a more aquatichabitat and the reduction of the size of the ribs, but have progressively evolved in themore terrestrial vertebrates to eliminate the buccal pump completely.
If it is contended that the buccal force-pump mechanism seen in the Dipnoi isessentially similar to that seen in the rhipidistian fishes and Amphibia, it is necessaryto comment on the use of the nostrils in respiration. These are used in respiration onlyin the Amphibia, even though internal nostrils are found in both groups. It is nowgenerally recognized that the nares of the Dipnoi are not homologous with those ofthe rhipidistian tetrapod line (Thompson, 1964; Bertmar, 1965). Kerr (1932) thoughtthat the evolution of the nostril was an aid to more efficient olfaction and not torespiration, and it is argued here that such olfactory devices were evolved separatelyin the dipnoan and rhipidistian lines. It is not known at what stage the nares becameimplicated in respiration, nor is it clear what advantage is gained by their use in anaquatic animal. One of the more plausible theories is that the use of the nostrils wasof advantage in filling the buccal cavity with air in an air-breathing but primarily

428 B. R. MCMAHON
aquatic animal. The filling of the buccal cavity with air must have presented problemsto the early crossopterygian fishes for in these forms the jaws were extended and thegape very long. In these animals the head must have had to be raised high out of thewater for the buccal cavity to be filled with air. The use of the nostrils would haveallowed buccal filling with only the tip of the snout protruding above the surface. Inthe Dipnoi the gape of the jaws is much reduced, and buccal fining is further facilitatedby a fold of skin which acts as a pressure-seal in the compression phases but whichconsiderably reduces the gape when the mouth is opened to take air at the surface.(PI. 3). It is possible that these two different solutions to the problem developed veryearly in the evolution of the air-breathing forms and are preserved today in thesurviving dipnoan and amphibian forms. The use of the buccal force-pump hassimilarly been preserved in both lines. Though the presence of a hinge in the skull ofthe rhipidistian fishes is indicative of major structural differences separating themfrom the Dipnoi, it is unlikely to have seriously affected the respiratory mechanisms.Thompson (1967) has investigated the kinetics of possible movements between theanterior and posterior parts of the skull but is unable to make firm conclusions as to itsfunction. In the opinion of this author the limited degree of movement possible wasunlikely to have been responsible for the production of the respiratory movements,but the increased flexibility of the skull may have added to the efficiency of the buccal/opercular pumping mechanism.
In conclusion, the lung-ventilation mechanism in Protopterus has been shown toincorporate a buccal force-pump mechanism basically similar to that observed both inother fishes and in amphibians. The mechanism has been shown to consist of a seriesof aquatic-type movements, each of which is slightly modified to serve a specificfunction in the air-breathing cycle. Little modification is really needed. Expirationoccurs in the expansion phase of one movement and inspiration follows in the com-pression phase of the next. No movement of the ribs has been observed in this or otherlungfishes. It is concluded that the air-breathing mechanism exhibited by the earliesttetrapods is likely to have been basically similar to that exhibited by the modernDipnoi, and that the aspiratory method of inspiration observed in the fully terrestrialvertebrates was evolved later together with the increase in structural importance ofthe ribs.
SUMMARY
1. The anatomy of the head and branchial region of Protopterus has been studied bydissection and section techniques to show the relation between skeletal and muscularelements. X-ray cinematographic, pressure and electromyographic techniques havebeen used to show how the muscular and skeletal systems interact to produce therespiratory movements. The mechanisms involved in aquatic and aerial respiration inProtopterus have thus been elucidated.
2. The mechanisms of branchial irrigation has been shown to be basically similar to thatseen in teleost and elasmobranch fishes, and also similar to that seen in larval amphibia.
3. The aerial cycle is composed of a series of aquatic-type cycles, each of which ismodified slightly to serve a specific function in the aerial cycle. Inspiration occurs bya buccal force-pump mechanism. Expiration occurs by the release of compressedpulmonary gas, aided by the elasticity of the lung wall.

Respiratory movements of an African lungfish 429
4. In this animal the air-breathing mechanism is derived from the aquatic mech-anism. The modifications are relatively simple and produce an efficient ventilationmechanism.
5. No movements of the ribs can be seen associated with the respiratory cycles. Itis suggested that the aspiratory ventilation mechanisms were not present in the proto-tetrapods and were not evolved until a later, more fully terrestrial stage was reached.
6. The evidence suggests that the air-breathing mechanism of the tetrapods waspowered by a buccal force-pump mechanism which evolved directly from the aquaticsystem. The evolution of a new mechanism for lung ventilation in the prototetrapodsis considered unnecessary.
I am indebted to Professor G. M. Hughes, Department of Zoology, University ofBristol, in whose department and under whose supervision this work was conducted,and also to the Science Research Council, who provided financial support.
REFERENCES
ATZ, J. W. (195a). Narial breathing in fishes and the evolution of the internal nares. Q. Rev. Biol. vj,366-77.
BALIINTIJN, C. M. & HUGHES, G. M. (1965). The muscular basis of the respiratory pumps in the trout.J. exp. Biol. 43, 349-62.
BASMAJAN, J. Y. & STECKO, G. (1962). A new bi-polar electrode for electromyography. J. appl. Physiol.17, 849.
BBRTMAR, G. (1965). The olfactory organ and the upper lips in Dipnoi, a comparative embryologicalstudy. Acta Zool., Stockh. 46, 1-40.
BISHOP, I, R. & FOXON, G. E. H. (1968). The mechanism of breathing in the South American lungfishLeptdotiren paradoxa; radiological study. J. Zool., Lond. 154, 263-72.
DUBOIS, R. (1892). Du mechanisme respiratoire du Dipnoiques. Amis Soc. Lbvu Lyon, 36, 65.EDGEWORTH, F. H. (1935). The Cranial Muscles of Vertebrates. Cambridge University Press.GREENWOOD, P. A. H. (1958). The Fishes of Uganda. Kampala: The Uganda Society.GRICG, C. (1965 a). Studies of the Queensland lungfish Neoceratodus forsteri (Krefft). I. Anatomy,
histology and functioning of the lung. Aust. J. Zool. 13, 243-53.GRIGG, C. (1965 b). Studies of the Queensland lungfish Neoceratodus forsten (Krefft). III. Aerial respira-
tion in relation to habits. Aust. J. Zool. 13, 413-21.HUGHES, G. M. (1960a). The mechanism of gill ventilation in the dogfish & skate. J. exp. Biol. 37, 11-27.HUGHES, G. M. (10606). A comparative study of gill ventilation in marine teleosts. J. exp. Biol. yj,
28-45.HUGHES, G. M. (1961). Gill ventilation in fishes. Rep. Challenger. Soc. 3, No. 13.HUGHES, G. M. & BAmNTijN, C. M. (1965). The muscular basis of the respiratory pumps in the
Dogfish (Scyliorhinus canicula). J. exp. Biol. 43, 363-83.HUGHES, G. M. & SHHLTON, G. (1958). The mechanism of gill ventilation in three fresh water teleosts.
J. exp. Biol. 35, 807-23.HUGHES, G. M. & SHELTON, G. (1962). Respiratory mechanisms and their nervous control in fish.
Advanc. comp. Physiol. Btochem. 1, 275-364.JAQUET, M. (1898-9). Quoted in Edgeworth (1935).JARVIK, E. (1967). On the structure of the lower jaw in Dipnoans with a description of early Devonian
Dipnoan from Canada (Melanognathus canadensis). J. Linn. Soc. Zool. 47 (311) 155-85.JESSE, M. J., SHUB, C , FISHMAN, A. P. (1968). Lung and gill ventilation of the African lungfish. Resp.
Physiol. 3 (3) 267, 287.JOHANSEN, K., LENTANT, C. & GRIGG, C. (1967). Respiratory control in the lungfish Neoceratodus
forsteri (KREFFT). Comp. Biochem. Physiol. in, 835-54.KERR, J. G. (1932). Archaic fishes. Lepidosiren and Protopterus and their bearing on problems of
vertebrate morphology. Jena. 2. Naturtv. dfj, 419-433.MCCUTCHEON, F. W. (1966). Pressure sensitivity, reflexes, and buoyancy responses in Teleosts. Arnnt
Behav. 14, 204-17.MCMAHON, B. R. (1969). The relative efficiency of gaseous exchange across the lungs and gills of an
African lungfish, Protopterus aethiopicus. J. exp. Biol. (In the Press).POLL, N. (1962). XI fitude sur la structure adulte et la formation des sacs puhnonaires des Protopteres.
Aimls Mus. r. Afr. cent. Sir. Svo. 103, 131-71.

430 B. R. MCMAHON
RAHN, H. (1966). Aquatic gas exchange theory. Resp. phytiol. 1, 1-11.ROBERTS, T. D. M. (1950). The respiratory movements of the lamprey (Lampetra fluviatilis). Proc.
R Soc. Edin. B 64, 252-3.SAUNDERS, R. L. (1061). The irrigation of the gills in fishes. I. Studies of the mechanism of branchial
irrigation. Can. J. Zool. 39, 637-53.SCHMALHAUSEN (1957). Quoted in Szarski (1062).SCHMALHAUSEN (1958). Quoted in Szarski (1062).SMITH, H. W. (1931). Observations on the African lungfish ProtopUnu aetfaopicus and on the evolution
from water to land environments. Ecology ia, 164-81.STERBA, G. (1962). Fresh Water Fishes of the World. Translated by D. W. Tucker. London: Vista Books
(1963).SZARSKI, H. (1962). The origin of the amphibia. Q. Rev. Biol. 37, 180-241.THOMPSON, K. S. (1964). The nasal apparatus in Dipnoi with special reference to Protopterus. Proc.
zool. Soc. Lond. 145, 207-62.THOMPSON, K. S. (1967). Mechanisms of intercranial kinetics in fossil rhipidistian fishes (Cross-
opterygii) and their relatives. J. Linn. Soc. Zool. 46, 223-58.WHITE, E. (1966). A little on lungfishes. Proc. Liim. Soc. Zool. 177 (1) 1-10.WILLEM, V. (1920 a). Observation sur la respiration des Amphibiens. Bull. Acad. r. Belg. Cl. Sci. 6,
298-314.WILLEM, V. (19206). Observations sur la respiration des Amphibiens. Deuxieme note. Bull. Acad. r.
Belg. Cl. Sci. 6, 341-47.WILLEM, V. (1929). Les manoeuvres respiratoires chez les poissons et les amphibiens. Bull. Acad. r.
Belg. Cl. Sci. 15, 1-194.WIT 1 MI V. (1941). Synchronisme des mouvements respiratoires et des pulsations cardiaques chez les
poissons. Bull. Acad. r. Belg. Cl. Sci. vj, 49-64.
EXPLANATION OF PLATES
PLATE I
X-Ray photograph of a living Protopterus to show the skeleton of the head and branchial region and theextent of the lung.
PLATE 2
Selected stills from an X-Ray cine film of Protopterus during air-breathing to show the principal stagesin the air-breathing cycle. Explanation in text.
PLATE 3
Mechanism of reduction of gape in Protopterus
A. Mouth open during air-breathing. Skin flap reduces gape and enables buccal cavity to be maximallyexpanded without entry of water.B. Mouth closed. Skin flap concealed along margin of mouth. Acts as a pressure-seal during buccalcompression.

Journal of Experimental Biology, Vol. 51, No. 2 Plate 1
Lower jaw
Ceratohyal
Opercular
Cranium
Pectoral girdle
Cranial rib
Anteriormedian sacof the lung
Vertebralcolumn
Segmentalribs
Paired partof the lung
\
B. R. McMAHON {Facing p. 430)
Journal of Experimental Biology, Vol. 51, No. 2 Plate 2
B R. McMAHON
Journal of Experimental Biology, Vol. 51, No. 2 Plate 3
B. R. McMAHON





























