Embed Size (px)
Citation preview

71
C H A P T E R
3
DRUG METABOLISM
Frank J. Gonzalez and Robert H. Tukey
How Humans Cope with Exposure to Xenobiotics.
The ability of humans to metabolize and clear drugs is anatural process that involves the same enzymatic path-ways and transport systems that are utilized for normalmetabolism of dietary constituents. Humans come intocontact with scores of foreign chemicals or xenobiotics(substances foreign to the body) through exposure to envi-ronmental contaminants as well as in our diets. Fortunate-ly, humans have developed a means to rapidly eliminatexenobiotics so they do not cause harm. In fact, one of themost common sources of xenobiotics in the diet is fromplants that have many structurally diverse chemicals,some of which are associated with pigment productionand others that are actually toxins (called phytoallexins)that protect plants against predators. A common exampleis poisonous mushrooms that have many toxins that arelethal to mammals, including amanitin, gyromitrin, orella-nine, muscarine, ibotenic acid, muscimol, psilocybin, andcoprine. Animals must be able to metabolize and elimi-nate such chemicals in order to consume vegetation.While humans can now choose their dietary source, a typ-ical animal does not have this luxury and as a result issubject to its environment and the vegetation that exists inthat environment. Thus, the ability to metabolize unusualchemicals in plants and other food sources is critical forsurvival.
Drugs are considered xenobiotics and most are exten-sively metabolized in humans. It is worth noting thatmany drugs are derived from chemicals found in plants,some of which had been used in Chinese herbal medicinesfor thousands of years. Of the prescription drugs in usetoday for cancer treatment, many derive from plant spe-cies (
see
Chapter 51); investigating folklore claims led tothe discovery of most of these drugs. It is therefore notsurprising that animals utilize a means for disposing ofhuman-made drugs that mimics the disposition of chemi-
cals found in the diet. This capacity to metabolize xenobi-otics, while mostly beneficial, has made development ofdrugs very time consuming and costly due in large part to(1) interindividual variations in the capacity of humans tometabolize drugs, (2) drug-drug interactions, and (3) spe-cies differences in expression of enzymes that metabolizedrugs. The latter limits the use of animal models in drugdevelopment.
A large number of diverse enzymes have evolved inanimals that apparently only function to metabolize for-eign chemicals. As will be discussed below, there aresuch large differences among species in the ability tometabolize xenobiotics that animal models cannot berelied upon to predict how humans will metabolize a drug.Enzymes that metabolize xenobiotics have historicallybeen called drug-metabolizing enzymes, although they areinvolved in the metabolism of many foreign chemicals towhich humans are exposed. Dietary differences amongspecies during the course of evolution could account forthe marked species variation in the complexity of thedrug-metabolizing enzymes.
Today, most xenobiotics to which humans are exposedcome from sources that include environmental pollution,food additives, cosmetic products, agrochemicals, pro-cessed foods, and drugs. In general, these are lipophilicchemicals, that in the absence of metabolism would not beefficiently eliminated, and thus would accumulate in thebody, resulting in toxicity. With very few exceptions, allxenobiotics are subjected to one or multiple pathways thatconstitute the phase 1 and phase 2 enzymatic systems. Asa general paradigm, metabolism serves to convert thesehydrophobic chemicals into derivatives that can easily beeliminated through the urine or the bile.
In order to be accessible to cells and reach their sites ofaction, drugs generally must possess physical propertiesthat allow them to move down a concentration gradient

72
Section I / General Principles
into the cell. Thus, most drugs are hydrophobic, a proper-ty that allows entry through the lipid bilayers into cellswhere drugs interact with their target receptors or pro-teins. Entry into cells is facilitated by a large number oftransporters on the plasma membrane (
see
Chapter 2).This property of hydrophobicity would render drugs diffi-cult to eliminate, since in the absence of metabolism, theywould accumulate in fat and cellular phospholipid bilay-ers in cells. The xenobiotic-metabolizing enzymes convertdrugs and xenobiotics into compounds that are hydro-philic derivatives that are more easily eliminated throughexcretion into the aqueous compartments of the tissues.Thus, the process of drug metabolism that leads to elimi-nation plays a major role in diminishing the biologicalactivity of a drug. For example,
(S)-phenytoin
, an anticon-vulsant used in the treatment of epilepsy, is virtuallyinsoluble in water. Metabolism by the phase 1 cytochromeP450 isoenzymes (CYPs) followed by phase 2 uridinediphosphate-glucuronosyltransferase (UGT) enzymes pro-duces a metabolite that is highly water soluble and readilyeliminated from the body (Figure 3–1). Metabolism alsoterminates the biological activity of the drug. In the caseof phenytoin, metabolism also increases the molecularweight of the compound, which allows it to be eliminatedmore efficiently in the bile.
While xenobiotic-metabolizing enzymes are responsi-ble for facilitating the elimination of chemicals from thebody, paradoxically these same enzymes can also con-vert certain chemicals to highly reactive toxic and car-cinogenic metabolites. This occurs when an unstableintermediate is formed that has reactivity toward othercompounds found in the cell. Chemicals that can be con-verted by xenobiotic metabolism to cancer-causingderivatives are called carcinogens. Depending on thestructure of the chemical substrate, xenobiotic-metabo-lizing enzymes produce electrophilic metabolites thatcan react with nucleophilic cellular macromoleculessuch as DNA, RNA, and protein. This can cause celldeath and organ toxicity. Reaction of these electrophileswith DNA can sometimes result in cancer through themutation of genes such as oncogenes or tumor suppres-sor genes. It is generally believed that most human can-cers are due to exposure to chemical carcinogens. Thispotential for carcinogenic activity makes testing thesafety of drug candidates of vital importance. Testing forpotential cancer-causing activity is particularly criticalfor drugs that will be used for the treatment of chronicdiseases. Since each species has evolved a unique com-bination of xenobiotic-metabolizing enzymes, nonpri-mate rodent models cannot be solely used during drugdevelopment for testing the safety of new drug candi-
dates targeted for human diseases. Nevertheless, testingin rodent models such as mice and rats can usually iden-tify potential carcinogens.
The Phases of Drug Metabolism.
Xenobiotic metabo-lizing enzymes have historically been grouped into thephase 1 reactions, in which enzymes carry out oxidation,reduction, or hydrolytic reactions, and the phase 2 reac-tions, in which enzymes form a conjugate of the sub-strate (the phase 1 product) (Table 3–1). The phase 1enzymes lead to the introduction of what are called func-tional groups, resulting in a modification of the drug,such that it now carries an –OH, -COOH, -SH, -O- orNH
2
group. The addition of functional groups does littleto increase the water solubility of the drug, but can dra-
Figure 3–1.
Metabolism of phenytoin by phase 1 cyto-chrome P450 (CYP) and phase 2 uridine diphosphate-glucu-ronosyltransferase (UGT).
CYP facilitates 4-hydroxylation ofphenytoin to yield 5-(-4-hydroxyphenyl)-5-phenylhydantoin(HPPH). The hydroxy group serves as a substrate for UGT thatconjugates a molecule of glucuronic acid using UDP-glucuronicacid (UDP-GA) as a cofactor. This converts a very hydrophobicmolecule to a larger hydrophilic derivative that is eliminated
via
the bile.

Chapter 3 / Drug Metabolism
73
matically alter the biological properties of the drug.Phase 1 metabolism is classified as the functionalizationphase of drug metabolism; reactions carried out by phase1 enzymes usually lead to the inactivation of an activedrug. In certain instances, metabolism, usually the hydrol-ysis of an ester or amide linkage, results in bioactivationof a drug. Inactive drugs that undergo metabolism to anactive drug are called prodrugs. An example is the anti-tumor drug
cyclophosphamide
, which is bioactivated toa cell-killing electrophilic derivative (
see
Chapter 51)
.
Phase 2 enzymes facilitate the elimination of drugs andthe inactivation of electrophilic and potentially toxicmetabolites produced by oxidation. While many phase 1reactions result in the biological inactivation of the drug,phase 2 reactions produce a metabolite with improvedwater solubility and increased molecular weight, whichserves to facilitate the elimination of the drug from thetissue.
Superfamilies of evolutionarily related enzymes andreceptors are common in the mammalian genome; theenzyme systems responsible for drug metabolism are goodexamples. The phase 1 oxidation reactions are carried outby CYPs, flavin-containing monooxygenases (FMO), andepoxide hydrolases (EH). The CYPs and FMOs are com-posed of superfamilies of enzymes. Each superfamily con-tains multiple genes. The phase 2 enzymes include severalsuperfamilies of conjugating enzymes. Among the more
important are the glutathione-S-transferases (GST), UDP-glucuronosyltransferases (UGT), sulfotransferases (SULT),
N
-acetyltransferases (NAT), and methyltransferases (MT).These conjugation reactions usually require the substrateto have oxygen (hydroxyl or epoxide groups), nitrogen,and sulfur atoms that serve as acceptor sites for a hydro-philic moiety, such as glutathione, glucuronic acid, sulfate,or an acetyl group, that is covalently conjugated to anacceptor site on the molecule. The example of phase 1 andphase 2 metabolism of phenytoin is shown in Figure 3–1.The oxidation by phase 1 enzymes either adds or exposes afunctional group, permitting the products of phase 1metabolism to serve as substrates for the phase 2 conjugat-ing or synthetic enzymes. In the case of the UGTs, glucu-ronic acid is delivered to the functional group, forming aglucuronide metabolite that is now more water solublewith a higher molecular weight that is targeted for excre-tion either in the urine or bile. When the substrate is adrug, these reactions usually convert the original drug to aform that is not able to bind to its target receptor, thusattenuating the biological response to the drug.
Sites of Drug Metabolism.
Xenobiotic metabolizingenzymes are found in most tissues in the body with thehighest levels located in the tissues of the gastrointestinaltract (liver, small and large intestines). Drugs that areorally administered, absorbed by the gut, and taken to the
Table 3–1
Xenobiotic Metabolizing Enzymes
ENZYMES REACTIONS
Phase 1 “oxygenases”
Cytochrome P450s (P450 or CYP) C and O oxidation, dealkylation, othersFlavin-containing monooxygenases (FMO) N, S, and P oxidationEpoxide hydrolases (mEH, sEH) Hydrolysis of epoxides
Phase 2 “transferases”
Sulfotransferases (SULT) Addition of sulfateUDP-glucuronosyltransferases (UGT) Addition of glucuronic acidGlutathione-S-transferases (GST) Addition of glutathione
N
-acetyltransferases (NAT) Addition of acetyl groupMethyltransferases (MT) Addition of methyl group
Other enzymes
Alcohol dehydrogenases Reduction of alcoholsAldehyde dehydrogenases Reduction of aldehydesNADPH-quinone oxidoreductase (NQO) Reduction of quinones
mEH and sEH are microsomal and soluble epoxide hydrolase. UDP, uridine diphosphate; NADPH, reduced nicotinamide adenine dinucleotidephosphate.
74
Section I / General Principles
liver, can be extensively metabolized. The liver is consid-ered the major “metabolic clearing house” for both endo-genous chemicals (
e.g.,
cholesterol, steroid hormones,fatty acids, and proteins), and xenobiotics. The smallintestine plays a crucial role in drug metabolism sincemost drugs that are orally administered are absorbed bythe gut and taken to the liver through the portal vein. Thehigh concentration of xenobiotic-metabolizing enzymeslocated in the epithelial cells of the GI tract is responsiblefor the initial metabolic processing of most oral medica-tions. This should be considered the initial site for first-pass metabolism of drugs. The absorbed drug then entersthe portal circulation for its first pass through the liver,where metabolism may be prominent, as it is for
β
adre-nergic receptor antagonists, for example. While a portionof active drug escapes this first-pass metabolism in the GItract and liver, subsequent passes through the liver resultin more metabolism of the parent drug until the agent iseliminated. Thus, drugs that are poorly metabolizedremain in the body for longer periods of time and theirpharmacokinetic profiles show much longer elimination
half-lives than drugs that are rapidly metabolized. Otherorgans that contain significant xenobiotic-metabolizingenzymes include the tissues of the nasal mucosa and lung,which play important roles in the first-pass metabolism ofdrugs that are administered through aerosol sprays. Thesetissues are also the first line of contact with hazardouschemicals that are airborne.
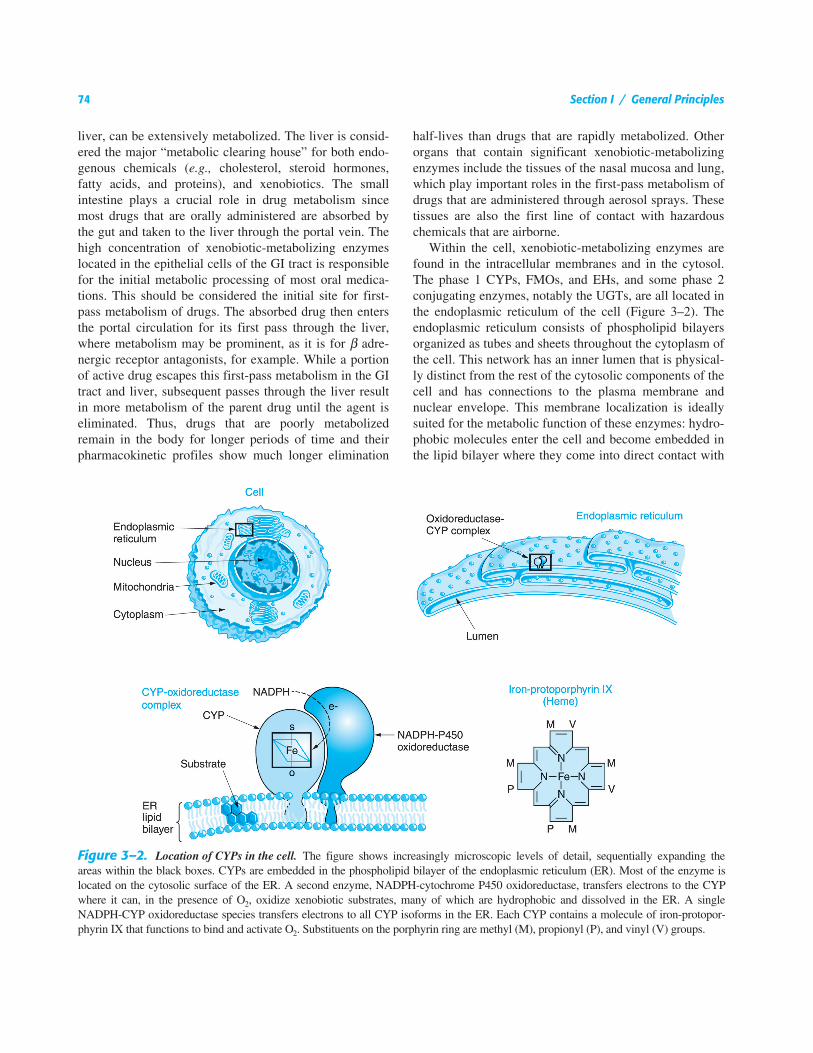
Within the cell, xenobiotic-metabolizing enzymes arefound in the intracellular membranes and in the cytosol.The phase 1 CYPs, FMOs, and EHs, and some phase 2conjugating enzymes, notably the UGTs, are all located inthe endoplasmic reticulum of the cell (Figure 3–2). Theendoplasmic reticulum consists of phospholipid bilayersorganized as tubes and sheets throughout the cytoplasm ofthe cell. This network has an inner lumen that is physical-ly distinct from the rest of the cytosolic components of thecell and has connections to the plasma membrane andnuclear envelope. This membrane localization is ideallysuited for the metabolic function of these enzymes: hydro-phobic molecules enter the cell and become embedded inthe lipid bilayer where they come into direct contact with
Figure 3–2.
Location of CYPs in the cell.
The figure shows increasingly microscopic levels of detail, sequentially expanding theareas within the black boxes. CYPs are embedded in the phospholipid bilayer of the endoplasmic reticulum (ER). Most of the enzyme islocated on the cytosolic surface of the ER. A second enzyme, NADPH-cytochrome P450 oxidoreductase, transfers electrons to the CYPwhere it can, in the presence of O
2
, oxidize xenobiotic substrates, many of which are hydrophobic and dissolved in the ER. A singleNADPH-CYP oxidoreductase species transfers electrons to all CYP isoforms in the ER. Each CYP contains a molecule of iron-protopor-phyrin IX that functions to bind and activate O
2
. Substituents on the porphyrin ring are methyl (M), propionyl (P), and vinyl (V) groups.

Chapter 3 / Drug Metabolism
75
the phase 1 enzymes. Once subjected to oxidation, drugscan be conjugated in the membrane by the UGTs or bythe cytosolic transferases such as GST and SULT. Themetabolites can then be transported out of the cell throughthe plasma membrane where they are deposited into thebloodstream. Hepatocytes, which constitute more than90% of the cells in the liver, carry out most drug metabo-lism and can produce conjugated substrates that can alsobe transported though the bile canalicular membrane intothe bile from which they are eliminated into the gut (
see
Chapter 2).
The CYPs.
The CYPs are a superfamily of enzymes, allof which contain a molecule of heme that is noncovalent-ly bound to the polypeptide chain (Figure 3–2). Manyother enzymes that use O
2
as a substrate for their reac-tions contain heme. Heme is the oxygen-binding moiety,also found in hemoglobin, where it functions in the bind-ing and transport of molecular oxygen from the lung toother tissues. Heme contains one atom of iron in a hydro-carbon cage that functions to bind oxygen in the CYPactive site as part of the catalytic cycle of these enzymes.CYPs use O
2
, plus H
+
derived from the cofactor-reducednicotinamide adenine dinucleotide phosphate (NADPH),to carry out the oxidation of substrates. The H
+
is sup-plied through the enzyme NADPH-cytochrome P450oxidoreductase. Metabolism of a substrate by a CYPconsumes one molecule of molecular oxygen and pro-duces an oxidized substrate and a molecule of water as aby-product. However, for most CYPs, depending on thenature of the substrate, the reaction is “uncoupled,” con-suming more O
2
than substrate metabolized and produc-ing what is called activated oxygen or O
2–
. The O
2–
isusually converted to water by the enzyme superoxidedismutase.
Among the diverse reactions carried out by mammali-an CYPs are
N
-dealkylation,
O
-dealkylation, aromatichydroxylation,
N
-oxidation,
S
-oxidation, deamination, anddehalogenation (Table 3–2). More than 50 individualCYPs have been identified in humans. As a family ofenzymes, CYPs are involved in the metabolism of dietaryand xenobiotic agents, as well as the synthesis of endoge-nous compounds such as steroids and the metabolism ofbile acids, which are degradation by-products of choles-terol. In contrast to the drug-metabolizing CYPs, theCYPs that catalyze steroid and bile acid synthesis havevery specific substrate preferences. For example, the CYPthat produces estrogen from testosterone, CYP19 or aro-matase, can metabolize only testosterone and does notmetabolize xenobiotics. Specific inhibitors for aromatase,such as
anastrozole,
have been developed for use in the
treatment of estrogen-dependent tumors (
see
Chapter 51).The synthesis of bile acids from cholesterol occurs in theliver, where, subsequent to CYP-catalyzed oxidation, thebile acids are conjugated and transported through the bileduct and gallbladder into the small intestine. CYPsinvolved in bile acid production have strict substraterequirements and do not participate in xenobiotic or drugmetabolism.
The CYPs that carry out xenobiotic metabolism have atremendous capacity to metabolize a large number ofstructurally diverse chemicals. This is due both to multi-ple forms of CYPs and to the capacity of a single CYP tometabolize many structurally distinct chemicals. A singlecompound can also be metabolized, albeit at differentrates, by different CYPs. In addition, CYPs can metabo-lize a single compound at different positions on the mole-cule. In contrast to enzymes in the body that carry outhighly specific reactions involved in the biosynthesis anddegradation of important cellular constituents in whichthere is a single substrate and one or more products, ortwo simultaneous substrates, the CYPs are consideredpromiscuous in their capacity to bind and metabolize mul-tiple substrates (Table 3–2). This property, which is dueto large and fluid substrate binding sites in the CYP, sac-rifices metabolic turnover rates; CYPs metabolize sub-strates at a fraction of the rate of more typical enzymesinvolved in intermediary metabolism and mitochondrialelectron transfer. As a result, drugs have, in general, half-lives of the order of 3 to 30 hours, while endogenouscompounds have half-lives of the order of seconds orminutes (
e.g.,
dopamine and insulin). Even though CYPshave slow catalytic rates, their activities are sufficient tometabolize drugs that are administered at high concentra-tions in the body. This unusual feature of extensive over-lapping substrate specificities by the CYPs is one of theunderlying reasons for the predominance of drug-druginteractions. When two coadministered drugs are bothmetabolized by a single CYP, they compete for binding tothe enzyme’s active site. This can result in the inhibitionof metabolism of one or both of the drugs, leading to ele-vated plasma levels. If there is a narrow therapeutic indexfor the drugs, the elevated serum levels may elicitunwanted toxicities. Drug-drug interactions are among theleading causes of adverse drug reactions.
The CYPs are the most actively studied of the xenobiotic metab-olizing enzymes since they are responsible for metabolizing the vastmajority of therapeutic drugs. CYPs are complex and diverse intheir regulation and catalytic activities. Cloning and sequencing ofCYP complementary DNAs, and more recently total genomesequencing, have revealed the existence of 102 putatively functionalgenes and 88 pseudogenes in the mouse, and 57 putatively functional

76
Table 3–2
Major Reactions Involved in Drug Metabolism
REACTION EXAMPLES
I
. Oxidative reactionsN
-Dealkylation RNHCH
3
→
RNH
2
+ CH
2
O Imipramine, diazepam, codeine, erythromycin, morphine, tamoxifen, theophylline, caffeine
O
-Dealkylation ROCH
3
→
ROH + CH
2
O Codeine, indomethacin, dextromethorphan
Aliphatic hydroxylation
RCH
2
CH
3
→
RCHOHCH
3
Tolbutamide, ibuprofen, phenobarbital, meprobamate, cyclosporine, midazolam
Aromatic hydroxylation
Phenytoin, phenobarbital, propanolol, ethinyl estradiol, amphetamine, war-farin
N
-Oxidation Chlorpheniramine, dapsone, meperidine
S
-Oxidation Cimetidine, chlorpromazine, thior-idazine, omeprazole
Deamination Diazepam, amphetamine
II
. Hydrolysis reactions
Carbamazepine
Procaine, aspirin, clofibrate, meperidine, enalapril, cocaine
Lidocaine, procainamide, indomethacin
III.
Conjugation reactions
Glucuronidation Acetaminophen, morphine, oxazepam, lorazepam
UDP-glucuronic acid
(Continued)
NHR1
R2
RNH2
N OHR1
R2
RNHOH
SH2
R1
R2
S OR1
R2
RCHCH3
NH2
R C CH3 + NH3
OH
NH2
R C CH3
O
R1COR2
O
R1CNR2
R1COOH
R1COOH
R2NH2
R2OH2
O
OHOH UDP
OH
COOH
O OHOH
+ UDR + OH
COOH
OO R

Chapter 3 / Drug Metabolism
77
genes and 58 pseudogenes in humans. These genes are grouped,based on amino acid sequence similarity, into a large number offamilies and subfamilies. CYPs are named with the root CYP fol-lowed by a number designating the family, a letter denoting the sub-family, and another number designating the CYP form. Thus,CYP3A4 is family 3, subfamily A, and gene number 4. While sever-al CYP families are involved in the synthesis of steroid hormonesand bile acids, and the metabolism of retinoic acid and fatty acids,including prostaglandins and eicosanoids, a limited number of CYPs(15 in humans) that fall into families 1 to 3 are primarily involved inxenobiotic metabolism (Table 3–1). Since a single CYP can metab-olize a large number of structurally diverse compounds, theseenzymes can collectively metabolize scores of chemicals found inthe diet, environment, and administered as drugs. In humans, 12CYPs (CYP1A1, 1A2, 1B1, 2A6, 2B6, 2C8, 2C9, 2C19, 2D6, 2E1,3A4, and 3A5) are known to be important for metabolism of xeno-biotics. The liver contains the greatest abundance of xenobiotic-metabolizing CYPs, thus ensuring efficient first-pass metabolism ofdrugs. CYPs are also expressed throughout the GI tract, and inlower amounts in lung, kidney, and even in the CNS. The expres-sion of the different CYPs can differ markedly as a result of dietaryand environmental exposure to inducers, or through interindividualchanges resulting from heritable polymorphic differences in genestructure, and tissue-specific expression patterns can impact onoverall drug metabolism and clearance. The most active CYPs fordrug metabolism are those in the CYP2C, CYP2D, and CYP3A sub-families. CYP3A4 is the most abundantly expressed and involved inthe metabolism of about 50% of clinically used drugs (Figure 3–3A). The CYP1A, CYP1B, CYP2A, CYP2B, and CYP2E subfami-lies are not significantly involved in the metabolism of therapeuticdrugs, but they do catalyze the metabolic activation of many protox-ins and procarcinogens to their ultimate reactive metabolites.
There are large differences in levels of expression of each CYPbetween individuals as assessed both by clinical pharmacologicstudies and by analysis of expression in human liver samples. Thislarge interindividual variability in CYP expression is due to thepresence of genetic polymorphisms and differences in gene regula-
tion (
see
below). Several human CYP genes exhibit polymorphisms,including
CYP2A6
,
CYP2C9
,
CYP2C19,
and
CYP2D6
. Allelic vari-ants have been found in the
CYP1B1
and
CYP3A4
genes, but theyare present at low frequencies in humans and appear not to have amajor role in interindividual levels of expression of these enzymes.However, homozygous mutations in the
CYP1B1
gene are associat-ed with primary congenital glaucoma.
Drug-Drug Interactions.
Differences in the rate ofmetabolism of a drug can be due to drug interactions.Most commonly, this occurs when two drugs (
e.g.,
a stat-in and a macrolide antibiotic or antifungal) are coadminis-tered and are metabolized by the same enzyme. Sincemost of these drug-drug interactions are due to CYPs, itbecomes important to determine the identity of the CYPthat metabolizes a particular drug and to avoid coadminis-tering drugs that are metabolized by the same enzyme.Some drugs can also inhibit CYPs independently of beingsubstrates for a CYP. For example, the common antifun-gal agent,
ketoconazole
(
NIZORAL
), is a potent inhibitorof CYP3A4 and other CYPs, and coadministration ofketoconazole with the anti-HIV viral protease inhibitorsreduces the clearance of the protease inhibitor and increasesits plasma concentration and the risk of toxicity. For mostdrugs, descriptive information found on the package insertlists the CYP that carries out its metabolism and thepotential for drug interactions. Some drugs are CYPinducers that can induce not only their own metabolism,but also induce metabolism of other coadministered drugs(
see
below and Figure 3–13). Steroid hormones and herb-al products such as St. John’s wort can increase hepaticlevels of CYP3A4, thereby increasing the metabolism of
Sulfation Acetaminophen, steroids, methyldopa
3'-phosphoadenosine-5' phosphosulfate
3'-phosphoadenosine-5'-phosphate
Acetylation Sulfonamides, isoniazid, dapsone, clonazepam (
see
Table 3–3)Methylation RO-, RS-, RN- + AdoMet
→
RO-CH
3
+ AdoHomCys
L
-Dopa, methyldopa, mercaptopurine, captopril
Glutathione conjugation
GSH + R
→
R-GSH Adriamycin, fosfomycin, busulfan
Table 3–2
Major Reactions Involved in Drug Metabolism (Continued)
REACTION EXAMPLES
SO2 OHORROH+ +PAPS PAP
CoAS CO CH3 CH3+ RNH2 + CoA-SHRNH CO

78
Section I / General Principles
many orally administered drugs. Drug metabolism canalso be influenced by diet. CYP inhibitors and inducersare commonly found in foods and in some cases these caninfluence the toxicity and efficacy of a drug. Componentsfound in grapefruit juice (
e.g.,
naringin, furanocoumarins)are potent inhibitors of CYP3A4, and thus some druginserts recommend not taking medication with grape-fruit juice because it could increase the bioavailabilityof a drug.
Terfenadine
, a once popular antihistamine, was removed fromthe market because its metabolism was blocked by CYP3A4 sub-
strates such as
erythromycin
and grapefruit juice. Terfenadine isactually a prodrug that requires oxidation by CYP3A4 to its activemetabolite, and at high doses the parent compound caused arrhyth-mias. Thus, elevated levels of parent drug in the plasma as a resultof CYP3A4 inhibition caused ventricular tachycardia in some indi-viduals, which ultimately led to its withdrawal from the market. Inaddition, interindividual differences in drug metabolism are signifi-cantly influenced by polymorphisms in CYPs. The CYP2D6 poly-morphism has led to the withdrawal of several clinically used drugs(
e.g.,
debrisoquine
and
perhexiline
) and the cautious use of othersthat are known CYP2D6 substrates (
e.g.
,
encainide
and
flecainide
[antiarrhythmics],
desipramine
and
nortriptyline
[antidepressants],and
codeine
).
Flavin-Containing Monooxygenases (FMOs).
The FMOsare another superfamily of phase 1 enzymes involvedin drug metabolism. Similar to CYPs, the FMOs areexpressed at high levels in the liver and are bound tothe endoplasmic reticulum, a site that favors interactionwith and metabolism of hydrophobic drug substrates.There are six families of FMOs, with FMO3 being themost abundant in liver. FMO3 is able to metabolizenicotine as well as H
2
-receptor antagonists (
cimetidine
and
ranitidine
), antipsychotics (
clozapine
), and anti-emetics (
itopride
). A genetic deficiency in this enzymecauses the fish-odor syndrome due to a lack of metabo-lism of trimethylamine
N
-oxide (TMAO) to trimethyl-amine (TMA); in the absence of this enzyme, TMAOaccumulates in the body and causes a socially offensivefish odor. TMAO is found at high concentrations, up to15% by weight, in marine animals where it acts as anosmotic regulator. FMOs are considered minor contrib-utors to drug metabolism and they almost always pro-duce benign metabolites. In addition, FMOs are notinduced by any of the xenobiotic receptors (
see
below)or easily inhibited; thus, in distinction to CYPs, FMOswould not be expected to be involved in drug-druginteractions. In fact, this has been demonstrated bycomparing the pathways of metabolism of two drugsused in the control of gastric motility, itopride and
cisapride
. Itopride is metabolized by FMO3 while cis-apride is metabolized by CYP3A4; thus, itopride is lesslikely to be involved in drug-drug interactions than iscisapride. CYP3A4 participates in drug-drug interac-tions through induction and inhibition of metabolism,whereas FMO3 is not induced or inhibited by any clini-cally used drugs. It remains a possibility that FMOsmay be of importance in the development of new drugs. Acandidate drug could be designed by introducing a sitefor FMO oxidation with the knowledge that selectedmetabolism and pharmacokinetic properties could beaccurately calculated for efficient drug-based biologi-cal efficacy.
Figure 3–3.
The fraction of clinically used drugs metabo-lized by the major phase 1 and phase 2 enzymes.
The relativesize of each pie section represents the estimated percentage ofdrugs metabolized by the major phase 1 (panel
A
) and phase 2(panel
B
) enzymes, based on studies in the literature. In somecases, more than a single enzyme is responsible for metabolismof a single drug. CYP, cytochrome P450; DPYD, dihydropyrim-idine dehydrogenase; GST, glutathione-S-transferase; NAT, N-acetyltransferase; SULT, sulfotransferase, TPMT, thiopurinemethyltransferase; UGT, UDP-glucuronosyltransferase.
Chapter 3 / Drug Metabolism 79
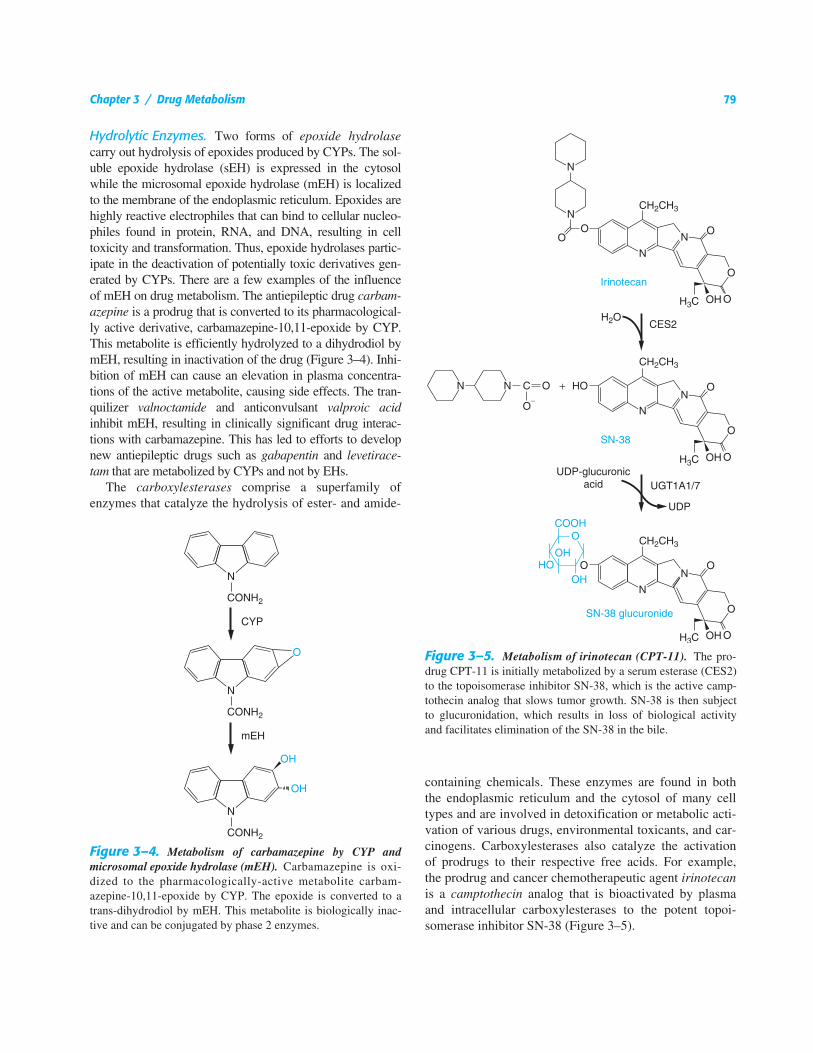
Hydrolytic Enzymes. Two forms of epoxide hydrolasecarry out hydrolysis of epoxides produced by CYPs. The sol-uble epoxide hydrolase (sEH) is expressed in the cytosolwhile the microsomal epoxide hydrolase (mEH) is localizedto the membrane of the endoplasmic reticulum. Epoxides arehighly reactive electrophiles that can bind to cellular nucleo-philes found in protein, RNA, and DNA, resulting in celltoxicity and transformation. Thus, epoxide hydrolases partic-ipate in the deactivation of potentially toxic derivatives gen-erated by CYPs. There are a few examples of the influenceof mEH on drug metabolism. The antiepileptic drug carbam-azepine is a prodrug that is converted to its pharmacological-ly active derivative, carbamazepine-10,11-epoxide by CYP.This metabolite is efficiently hydrolyzed to a dihydrodiol bymEH, resulting in inactivation of the drug (Figure 3–4). Inhi-bition of mEH can cause an elevation in plasma concentra-tions of the active metabolite, causing side effects. The tran-quilizer valnoctamide and anticonvulsant valproic acidinhibit mEH, resulting in clinically significant drug interac-tions with carbamazepine. This has led to efforts to developnew antiepileptic drugs such as gabapentin and levetirace-tam that are metabolized by CYPs and not by EHs.
The carboxylesterases comprise a superfamily ofenzymes that catalyze the hydrolysis of ester- and amide-
containing chemicals. These enzymes are found in boththe endoplasmic reticulum and the cytosol of many celltypes and are involved in detoxification or metabolic acti-vation of various drugs, environmental toxicants, and car-cinogens. Carboxylesterases also catalyze the activationof prodrugs to their respective free acids. For example,the prodrug and cancer chemotherapeutic agent irinotecanis a camptothecin analog that is bioactivated by plasmaand intracellular carboxylesterases to the potent topoi-somerase inhibitor SN-38 (Figure 3–5).
Figure 3–4. Metabolism of carbamazepine by CYP andmicrosomal epoxide hydrolase (mEH). Carbamazepine is oxi-dized to the pharmacologically-active metabolite carbam-azepine-10,11-epoxide by CYP. The epoxide is converted to atrans-dihydrodiol by mEH. This metabolite is biologically inac-tive and can be conjugated by phase 2 enzymes.
Figure 3–5. Metabolism of irinotecan (CPT-11). The pro-drug CPT-11 is initially metabolized by a serum esterase (CES2)to the topoisomerase inhibitor SN-38, which is the active camp-tothecin analog that slows tumor growth. SN-38 is then subjectto glucuronidation, which results in loss of biological activityand facilitates elimination of the SN-38 in the bile.

80 Section I / General Principles
Conjugating Enzymes. There are a large number ofphase 2 conjugating enzymes, all of which are consideredto be synthetic in nature since they result in the formationof a metabolite with an increased molecular mass. Phase 2reactions also terminate the biological activity of the drug.The contributions of different phase 2 reactions to drugmetabolism are shown in Figure 3–3B. Two of the phase 2reactions, glucuronidation and sulfation, result in the for-mation of metabolites with a significantly increased water-to-lipid partition coefficient, resulting in hydrophilicity andfacilitating their transport into the aqueous compartmentsof the cell and the body. Glucuronidation also markedlyincreases the molecular weight of the compound, a modifi-cation that favors biliary excretion. While sulfation andacetylation terminate the biological activity of drugs, thesolubility properties of these metabolites are alteredthrough minor changes in the overall charge of the mole-cule. Characteristic of the phase 2 reactions is the depen-dency on the catalytic reactions for cofactors such asUDP-glucuronic acid (UDP-GA) and 3′-phosphoadenos-ine-5′-phosphosulfate (PAPS), for UDP-glucuronosyltrans-ferases (UGT) and sulfotransferases (SULT), respectively,which react with available functional groups on the sub-strates. The reactive functional groups are often generatedby the phase 1 CYPs. All of the phase 2 reactions are car-ried out in the cytosol of the cell, with the exception of glu-curonidation, which is localized to the luminal side of theendoplasmic reticulum. The catalytic rates of phase 2 reac-tions are significantly faster than the rates of the CYPs.Thus, if a drug is targeted for phase 1 oxidation through theCYPs, followed by a phase 2 conjugation reaction, usuallythe rate of elimination will depend upon the initial (phase1) oxidation reaction. Since the rate of conjugation is fasterand the process leads to an increase in hydrophilicity of thedrug, phase 2 reactions are generally considered to assurethe efficient elimination and detoxification of most drugs.
Glucuronidation. Among the more important of thephase 2 reactions in the metabolism of drugs is that cata-lyzed by UDP-glucuronosyltransferases (UGTs) (Figure3–3B). These enzymes catalyze the transfer of glucuronicacid from the cofactor UDP-glucuronic acid to a substrateto form β-D-glucopyranosiduronic acids (glucuronides),metabolites that are sensitive to cleavage by β-glucu-ronidase. The generation of glucuronides can be formedthrough alcoholic and phenolic hydroxyl groups, carbox-yl, sulfuryl, and carbonyl moieties, as well as through pri-mary, secondary, and tertiary amine linkages. Examplesof glucuronidation reactions are shown in Table 3–2 andFigure 3–5. The structural diversity in the many differenttypes of drugs and xenobiotics that are processed through
glucuronidation assures that most clinically efficacioustherapeutic agents will be excreted as glucuronides.
There are 19 human genes that encode the UGT proteins. Nineare encoded by the UGT1 locus and 10 are encoded by the UGT2family of genes. Both families of proteins are involved in the metab-olism of drugs and xenobiotics, while the UGT2 family of proteinsappears to have greater specificity for the glucuronidation of endo-genous substances such as steroids. The UGT2 proteins are encodedby unique genes on chromosome 4 and the structure of each geneincludes six exons. The clustering of the UGT2 genes on the samechromosome with a comparable organization of the regions encod-ing the open reading frames is evidence that gene duplication hasoccurred, a process of natural selection that has resulted in the mul-tiplication and eventual diversification of the potential to detoxifythe plethora of compounds that are targeted for glucuronidation.
The nine functional UGT1 proteins are all encoded by the UGT1locus (Figure 3–6), which is located on chromosome 2. The UGT1locus spans nearly 200 kb, with over 150 kb encoding a tandemarray of cassette exonic regions that encode approximately 280amino acids of the amino terminal portion of the UGT1A proteins.Four exons are located at the 3′ end of the locus that encode the car-boxyl 245 amino acids that combine with one of the consecutivelynumbered array of first exons to form the individual UGT1 geneproducts. Since exons 2 to 5 encode the same sequence for eachUGT1A protein, the variability in substrate specificity for each ofthe UGT1A proteins results from the significant divergence insequence encoded by the exon 1 regions. The 5′ flanking region ofeach first-exon cassette contains a fully functional promoter capableof initiating transcription in an inducible and tissue-specific manner.
From a clinical perspective, the expression of UGT1A1 assumes animportant role in drug metabolism since the glucuronidation of biliru-bin by UGT1A1 is the rate-limiting step in assuring efficient bilirubinclearance, and this rate can be affected by both genetic variation andcompeting substrates (drugs). Bilirubin is the breakdown product ofheme, 80% of which originates from circulating hemoglobin and 20%from other heme-containing proteins such as the CYPs. Bilirubin is
Figure 3–6. Organization of the UGT1A Locus. Transcrip-tion of the UGT1A genes commences with the activation ofPolII, which is controlled through tissue-specific events. Con-served exons 2 to 5 are spliced to each respective exon 1sequence resulting in the production of unique UGT1A sequenc-es. The UGT1A locus encodes nine functional proteins.

Chapter 3 / Drug Metabolism 81
hydrophobic, associates with serum albumin, and must be metabolizedfurther by glucuronidation to assure its elimination. The failure to effi-ciently metabolize bilirubin by glucuronidation leads to elevated serumlevels and a clinical symptom called hyperbilirubinemia or jaundice.There are more than 50 genetic lesions in the UGT1A1 gene that canlead to inheritable unconjugated hyperbilirubinemia. Crigler-Najjarsyndrome type I is diagnosed as a complete lack of bilirubin glucu-ronidation, while Crigler-Najjar syndrome type II is differentiated bythe detection of low amounts of bilirubin glucuronides in duodenalsecretions. Types I and II Crigler-Najjar syndrome are rare, and resultfrom genetic polymorphisms in the open reading frames of theUGT1A1 gene, resulting in abolished or highly diminished levels offunctional protein.
Gilbert’s syndrome is a generally benign condition that is presentin up to 10% of the population; it is diagnosed clinically because cir-culating bilirubin levels are 60% to 70% higher than those seen innormal subjects. The most common genetic polymorphism associatedwith Gilbert’s syndrome is a mutation in the UGT1A1 gene promoter,which leads to reduced expression levels of UGT1A1. Subjects diag-nosed with Gilbert’s syndrome may be predisposed to adverse drugreactions resulting from a reduced capacity to metabolize drugs byUGT1A1. If a drug undergoes selective metabolism by UGT1A1,competition for drug metabolism with bilirubin glucuronidation willexist, resulting in pronounced hyperbilirubinemia as well as reducedclearance of metabolized drug. Tranilast [N-(3′4′-demethoxycin-namoyl)-anthranilic acid] is an investigational drug used for the pre-vention of restenosis in patients that have undergone transluminal cor-onary revascularization (intracoronary stents). Tranilast therapy in
patients with Gilbert’s syndrome has been shown to lead to hyperbili-rubinemia as well as potential hepatic complications resulting fromelevated levels of tranilast.
Gilbert’s syndrome also alters patient responses to irinotecan.Irinotecan, a prodrug used in chemotherapy of solid tumors (seeChapter 51), is metabolized to its active form, SN-38, by serum car-boxylesterases (Figure 3–5). SN-38, a potent topoisomerase inhibi-tor, is inactivated by UGT1A1 and excreted in the bile (Figures 3–7and 3–8). Once in the lumen of the intestine, the SN-38 glucuronideundergoes cleavage by bacterial β-glucuronidase and re-enters thecirculation through intestinal absorption. Elevated levels of SN-38in the blood lead to hematological toxicities characterized by leuko-penia and neutropenia, as well as damage to the intestinal epithelialcells (Figure 3–8), resulting in acute and life-threatening diarrhea.Patients with Gilbert’s syndrome who are receiving irinotecan thera-py are predisposed to the hematological and gastrointestinal toxici-ties resulting from elevated serum levels of SN-38, the net result ofinsufficient UGT1A activity and the consequent accumulation of atoxic drug in the GI epithelium.
The UGTs are expressed in a tissue-specific and often induc-ible fashion in most human tissues, with the highest concentrationof enzymes found in the GI tract and liver. Based upon their physi-cochemical properties, glucuronides are excreted by the kidneysinto the urine or through active transport processes through theapical surface of the liver hepatocytes into the bile ducts wherethey are transported to the duodenum for excretion with compo-nents of the bile. Most of the bile acids that are glucuronidated arereabsorbed back to the liver for reutilization by “enterohepatic
Figure 3–7. Routes of SN-38 transport and exposure to intestinal epithelial cells. SN-38 is transported into the bile followingglucuronidation by liver UGT1A1 and extrahepatic UGT1A7. Following cleavage of luminal SN-38 glucuronide (SN-38G) by bacteri-al β-glucuronidase, reabsorption into epithelial cells can occur by passive diffusion (indicated by the dashed arrows entering the cell)as well as by apical transporters. Movement into epithelial cells may also occur from the blood by basolateral transporters. IntestinalSN-38 can efflux into the lumen through P-glycoprotein (P-gp) and multidrug resistance protein 2 (MRP2) and into the blood viaMRP1. Excessive accumulation of the SN-38 in intestinal epithelial cells, resulting from reduced glucuronidation, can lead to cellulardamage and toxicity (Tukey et al., 2002).

82 Section I / General Principles
recirculation”; many drugs that are glucuronidated and excreted inthe bile can re-enter the circulation by this same process. The β-D-glucopyranosiduronic acids are targets for β-glucuronidase activi-ty found in resident strains of bacteria that are common in thelower GI tract, liberating the free drug into the intestinal lumen.As water is reabsorbed into the large intestine, free drug can thenbe transported by passive diffusion or through apical transportersback into the intestinal epithelial cells, from which the drug canthen re-enter the circulation. Through portal venous return fromthe large intestine to the liver, the reabsorption process allows forthe re-entry of drug into the systemic circulation (Figures 3–7 and3–8).
Sulfation. The sulfotransferases (SULTs) are located inthe cytosol and conjugate sulfate derived from 3′-phos-phoadenosine-5′-phosphosulfate (PAPS) to the hydroxylgroups of aromatic and aliphatic compounds. Like all ofthe xenobiotic metabolizing enzymes, the SULTsmetabolize a wide variety of endogenous and exoge-nous substrates. In humans, 11 SULT isoforms havebeen identified, and, based on evolutionary projections,have been classified into the SULT1 (SULT1A1, SULT1A2,SULT1A3, SULT1B1, SULT1C2, SYLT1C4, SULT1E1),SULT2 (SULT2A1, SULT2B1-v1, SULT2B1-v2), andSULT4 (SULT4A1) families. SULTs play an importantrole in normal human homeostasis. For example, SULT1B1is the predominant form expressed in skin and brain, car-rying out the catalysis of cholesterol and thyroid hor-mones. Cholesterol sulfate is an essential metabolite inregulating keratinocyte differentiation and skin develop-ment. SULT1A3 is highly selective for catecholamines,while estrogens are sulfated by SULT1E1 and dehydroepi-
androsterone (DHEA) is selectively sulfated by SULT2A1.In humans, significant fractions of circulating catechol-amines, estrogens, iodothyronines, and DHEA exist in thesulfated form.
The different human SULTs display a variety of unique sub-strate specificities. The SULT1 family isoforms are considered tobe the major forms involved in drug metabolism, with SULT1A1being the most important and displaying extensive diversity in itscapacity to catalyze the sulfation of a wide variety of structurallyheterogeneous xenobiotics. The isoforms in the SULT1 familyhave been recognized as phenol SULTs, since they have beencharacterized to catalyze the sulfation of phenolic molecules suchas acetaminophen, minoxidil, and 17α-ethinyl estradiol. Whiletwo SULT1C isoforms exist, little is known about their substratespecificity toward drugs, although SULT1C4 is capable of sulfat-ing the hepatic carcinogen N-OH-2-acetylaminofluorene. BothSULT1C2 and SULT1C4 are expressed abundantly in fetal tissuesand decline in abundance in adults, yet little is known about theirsubstrate specificities. SULT1E catalyzes the sulfation of endoge-nous and exogenous steroids, and has been found localized in liveras well as in hormone-responsive tissues such as the testis, breast,adrenal gland, and placenta.
The conjugation of drugs and xenobiotics is considered pri-marily a detoxification step, assuring that these agents are com-partmented into the water compartments of the body for targetedelimination. However, drug metabolism through sulfation oftenleads to the generation of chemically reactive metabolites, wherethe sulfate is electron withdrawing and may be heterolyticallycleaved, leading to the formation of an electrophilic cation. Mostexamples of the generation by sulfation of a carcinogenic or toxicresponse in animal or test mutagenicity assays have been docu-mented with chemicals derived from the environment or fromfood mutagens generated from well-cooked meat. Thus, it isimportant to understand whether genetic linkages can be made by
Figure 3–8. Cellular targets of SN-38 in the blood and intestinal tissues. Excessive accumulation of SN-38 can lead to bloodtoxicities such as leukopenia and neutropenia, as well as damage to the intestinal epithelium. These toxicities are pronounced in indi-viduals that have reduced capacity to form the SN-38 glucuronide, such as patients with Gilbert’s syndrome. Note the different bodycompartments and cell types involved (Tukey et al., 2002).
Blood Supply GI LumenGIEpithelium
UGT1A1
UGT1A7
Blood Cells
IntestinalEpithelium
Toxicity
ToxicityLeukopeniaNeutropenia
CarboxylesteraseSN-38 SN-38G
(bile)
SN-38G(excretion in feces)
SN-38
bacterialβ-glucuronidase
Diarrhea
IRINOTECAN(CPT-11)

Chapter 3 / Drug Metabolism 83
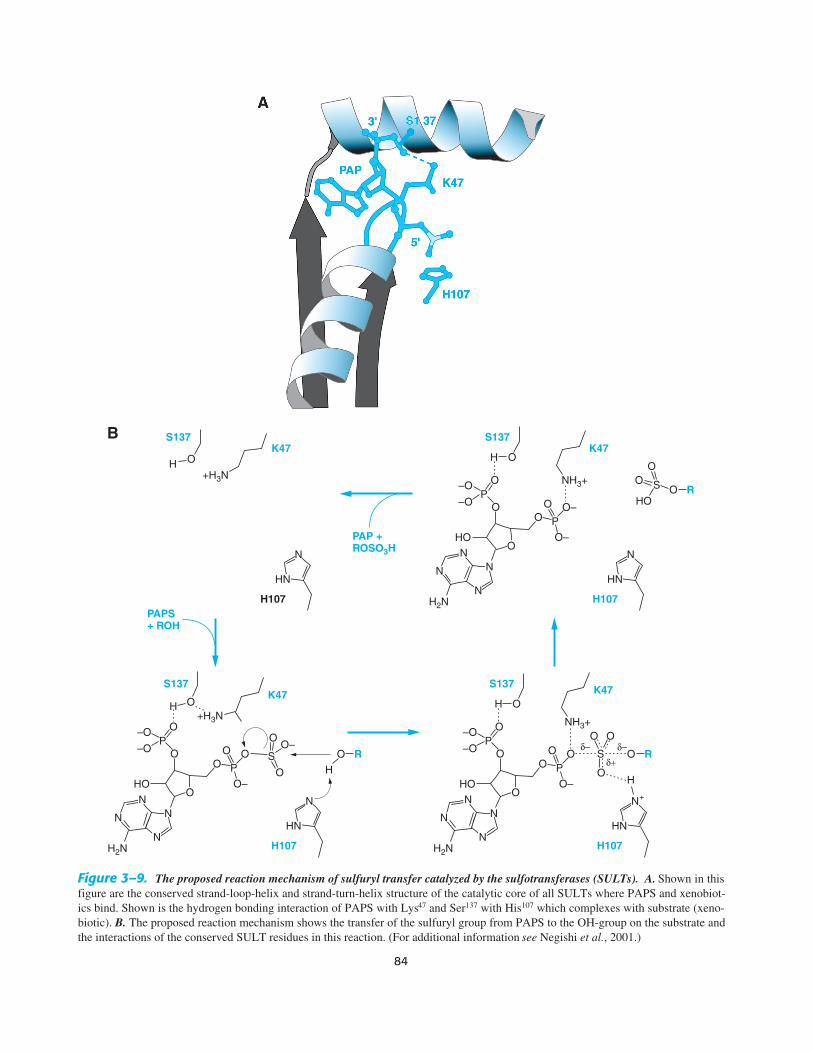
associating known human SULT polymorphisms to cancer epi-sodes that are felt to originate from environmental sources. SinceSULT1A1 is the most abundant in human tissues and displaysbroad substrate specificity, the polymorphic profiles associatedwith this gene and the onset of various human cancers is of con-siderable interest. An appreciation of the structure of the proteinsof the SULT family will aid in drug design and advance an under-standing of the linkages relating sulfation to cancer susceptibility,reproduction, and development. The SULTs from the SULT1 andSULT2 families were among the first xenobiotic-metabolizingenzymes to be crystallized and the data indicate a highly con-served catalytic core (Figure 3–9A). The structures reveal the roleof the co-substrate PAPS in catalysis, identifying the conservedamino acids that facilitate the 3′ phosphate’s role in sulfuryltransfer to the protein and in turn to the substrate (Figure 3–9B).Crystal structures of the different SULTs indicate that while con-servation in the PAPS binding region is maintained, the organiza-tion of the substrate binding region differs, helping to explain theobserved differences in catalytic potential displayed with the dif-ferent SULTs.
Glutathione Conjugation. The glutathione-S-trans-ferases (GSTs) catalyze the transfer of glutathione toreactive electrophiles, a function that serves to protectcellular macromolecules from interacting with electro-philes that contain electrophilic heteroatoms (-O, -N,and -S) and in turn protects the cellular environmentfrom damage. The co-substrate in the reaction is the trip-eptide glutathione, which is synthesized from γ-glutamicacid, cysteine, and glycine (Figure 3–10). Glutathioneexists in the cell as oxidized (GSSG) or reduced (GSH),and the ratio of GSH:GSSG is critical in maintaining acellular environment in the reduced state. In addition toaffecting xenobiotic conjugation with GSH, a severereduction in GSH content can predispose cells to oxida-tive damage, a state that has been linked to a number ofhuman health issues.
In the formation of glutathione conjugates, the reactiongenerates a thioether linkage with drug or xenobiotic tothe cysteine moiety of the tripeptide. Characteristically,all GST substrates contain an electrophilic atom and arehydrophobic, and by nature will associate with cellularproteins. Since the concentration of glutathione in cells isusually very high, typically ~7 µmol/g of liver, or in the10 mM range, many drugs and xenobiotics can react non-enzymatically with glutathione. However, the GSTs havebeen found to occupy up to 10% of the total cellular pro-tein concentration, a property that assures efficient conju-gation of glutathione to reactive electrophiles. The highconcentration of GSTs also provides the cells with a sinkof cytosolic protein, a property that facilitates noncovalentand sometimes covalent interactions with compounds thatare not substrates for glutathione conjugation. The cytoso-lic pool of GSTs, once identified as ligandin, has been
shown to bind steroids, bile acids, bilirubin, cellular hor-mones, and environmental toxicants, in addition to com-plexing with other cellular proteins.
Over 20 human GSTs have been identified and divided into twosubfamilies: the cytosolic and the microsomal forms. The major dif-ferences in function between the microsomal and cytosolic GSTsreside in the selection of substrates for conjugation; the cytosolicforms have more importance in the metabolism of drugs and xeno-biotics, whereas the microsomal GSTs are important in the endoge-nous metabolism of leukotrienes and prostaglandins. The cytosolicGSTs are divided into seven classes termed alpha (GSTA1 and 2),mu (GSTM1 through 5), omega (GSTO1), pi (GSTP1), sigma(GSTS1), theta (GSTT1 and 2), and zeta (GSTZ1). Those in thealpha and mu classes can form heterodimers, allowing for a largenumber of active transferases to form. The cytosolic forms of GSTcatalyze conjugation, reduction, and isomerization reactions.
The high concentrations of GSH in the cell, as well as the over-abundance of GSTs, means that few reactive molecules escapedetoxification. However, while there appears to be an overcapacityof enzyme and reducing equivalents, there is always concern thatsome reactive intermediates will escape detoxification, and bynature of their electrophilicity, will bind to cellular componentsand cause toxicity. The potential for such an occurrence is height-ened if GSH is depleted or if a specific form of GST is polymor-phic. While it is difficult to deplete cellular GSH levels, therapeu-tic agents that require large doses to be clinically efficacious havethe greatest potential to lower cellular GSH levels. Acetamino-phen, which is normally metabolized by glucuronidation and sul-fation, is also a substrate for oxidative metabolism by CYP2E1,which generates the toxic metabolite N-acetyl-p-benzoquinoneimine (NAPQI). An overdose of acetaminophen can lead to deple-tion of cellular GSH levels, thereby increasing the potential forNAPQI to interact with other cellular components. Acetaminophentoxicity is associated with increased levels of NAPQI and tissuenecrosis.
Like many of the enzymes involved in drug and xenobioticmetabolism, all of the GSTs have been shown to be polymorphic.The mu (GSTM1*0) and theta (GSTT1*0) genotypes express a nullphenotype; thus, individuals that are polymorphic at these loci arepredisposed to toxicities by agents that are selective substrates forthese GSTs. For example, the GSTM1*0 allele is observed in 50%of the Caucasian population and has been linked genetically tohuman malignancies of the lung, colon, and bladder. Null activity inthe GSTT1 gene has been associated with adverse side effects andtoxicity in cancer chemotherapy with cytostatic drugs; the toxicitiesresult from insufficient clearance of the drugs via GSH conjugation.Expression of the null genotype can be as high as 60% in Chineseand Korean populations. Therapies may alter efficacies, with anincrease in severity of adverse side effects.
While the GSTs play an important role in cellular detoxification,their activities in cancerous tissues have been linked to the develop-ment of drug resistance toward chemotherapeutic agents that areboth substrates and nonsubstrates for the GSTs. Many anticancerdrugs are effective because they initiate cell death or apoptosis,which is linked to the activation of mitogen-activated protein(MAP) kinases such as JNK and p38. Investigational studies havedemonstrated that overexpression of GSTs is associated with resis-tance to apoptosis and the inhibition of MAP kinase activity. In avariety of tumors, the levels of GSTs are overexpressed, which
84
Figure 3–9. The proposed reaction mechanism of sulfuryl transfer catalyzed by the sulfotransferases (SULTs). A. Shown in thisfigure are the conserved strand-loop-helix and strand-turn-helix structure of the catalytic core of all SULTs where PAPS and xenobiot-ics bind. Shown is the hydrogen bonding interaction of PAPS with Lys47 and Ser137 with His107 which complexes with substrate (xeno-biotic). B. The proposed reaction mechanism shows the transfer of the sulfuryl group from PAPS to the OH-group on the substrate andthe interactions of the conserved SULT residues in this reaction. (For additional information see Negishi et al., 2001.)
B

Chapter 3 / Drug Metabolism 85
leads to a reduction in MAP kinase activity and reduced efficacy ofchemotherapy. Taking advantage of the relatively high levels ofGST in tumor cells, inhibition of GST activity has been exploited asa therapeutic strategy to modulate drug resistance by sensitizingtumors to anticancer drugs. TLK199, a glutathione analog, serves asa prodrug that undergoes activation by plasma esterases to a GSTinhibitor, TLK117, which potentiates the toxicity of different anti-cancer agents (Figure 3–11). Alternatively, the elevated level ofGST activity in cancerous cells has been utilized to develop pro-drugs that can be activated by the GSTs to form electrophilic inter-mediates. TLK286 is a substrate for GST that undergoes a β-elimi-nation reaction, forming a glutathione conjugate and a nitrogenmustard (Figure 3–12) that is capable of alkylating cellular nucleo-philes, resulting in antitumor activity.
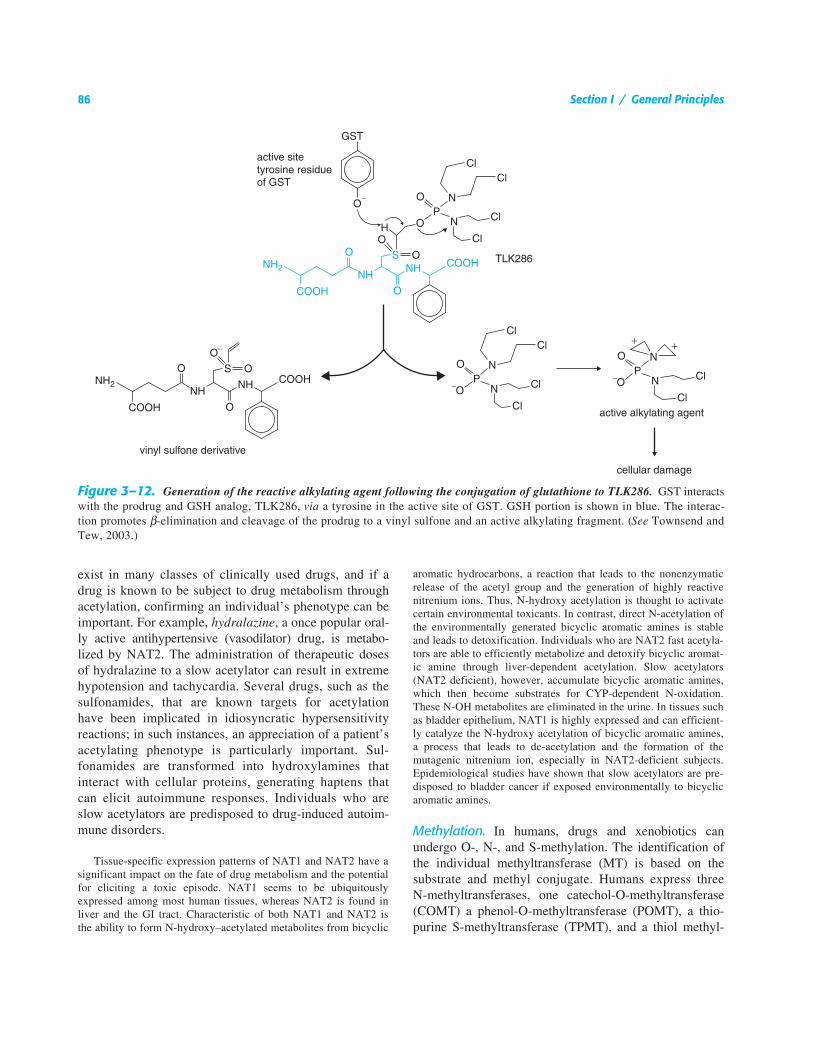
N-Acetylation. The cytosolic N-acetyltransferases (NATs)are responsible for the metabolism of drugs and environ-mental agents that contain an aromatic amine or hydrazinegroup. The addition of the acetyl group from the cofactoracetyl-coenzyme A often leads to a metabolite that is lesswater soluble because the potential ionizable amine isneutralized by the covalent addition of the acetyl group.NATs are among the most polymorphic of all the humanxenobiotic drug-metabolizing enzymes.
The characterization of an acetylator phenotype inhumans was one of the first hereditary traits identified,and was responsible for the development of the field ofpharmacogenetics (see Chapter 4). Following the discov-ery that isonicotinic acid hydrazide (isoniazid) could beused in the cure of tuberculosis, a significant proportionof the patients (5% to 15%) experienced toxicities thatranged from numbness and tingling in their fingers toCNS damage. After finding that isoniazid was metabo-lized by acetylation and excreted in the urine, researchersnoted that individuals suffering from the toxic effects ofthe drug excreted the largest amount of unchanged drugand the least amount of acetylated isoniazid. Pharmacoge-
netic studies led to the classification of “rapid” and“slow” acetylators, with the “slow” phenotype being pre-disposed to toxicity. Purification and characterization ofN-acetyltransferase and the eventual cloning of its RNAprovided sequence characterization of the gene for slowand fast acetylators, revealing polymorphisms that corre-spond to the “slow” acetylator phenotype. There are twofunctional NAT genes in humans, NAT1 and NAT2. Over25 allelic variants of NAT1 and NAT2 have been charac-terized, and in individuals in whom acetylation of drugs iscompromised, homozygous genotypes for at least twovariant alleles are required to predispose a patient to low-ered drug metabolism. Polymorphism in the NAT2 geneand its association with the slow acetylation of isoniazidwas one of the first completely characterized genotypesshown to impact drug metabolism, thereby linking pharma-cogenetic phenotype to a genetic polymorphism. Althoughnearly as many mutations have been identified in theNAT1 gene as the NAT2 gene, the frequency of the slowacetylation patterns are attributed mostly to polymor-phism in the NAT2 gene.
A list of drugs that are subject to acetylation and theirknown toxicities are listed in Table 3–3. The therapeuticrelevance of NAT polymorphisms is in avoiding drug-induced toxicities. The adverse drug response in a slowacetylator resembles a drug overdose; thus, reducing thedose or increasing the dosing interval is recommended.Drugs containing an aromatic amine or a hydrazine group
Figure 3–10. Glutathione as a co-substrate in the conjuga-tion of a drug or xenobiotic (X) by glutathione-S-transferase(GST).
Figure 3–11. Activation of TLK199 by cellular esterasesto the glutathione-S-transferase (GST) inhibitor TLK117.(For additional information, see Townsend and Tew, 2003.)
86 Section I / General Principles
exist in many classes of clinically used drugs, and if adrug is known to be subject to drug metabolism throughacetylation, confirming an individual’s phenotype can beimportant. For example, hydralazine, a once popular oral-ly active antihypertensive (vasodilator) drug, is metabo-lized by NAT2. The administration of therapeutic dosesof hydralazine to a slow acetylator can result in extremehypotension and tachycardia. Several drugs, such as thesulfonamides, that are known targets for acetylationhave been implicated in idiosyncratic hypersensitivityreactions; in such instances, an appreciation of a patient’sacetylating phenotype is particularly important. Sul-fonamides are transformed into hydroxylamines thatinteract with cellular proteins, generating haptens thatcan elicit autoimmune responses. Individuals who areslow acetylators are predisposed to drug-induced autoim-mune disorders.
Tissue-specific expression patterns of NAT1 and NAT2 have asignificant impact on the fate of drug metabolism and the potentialfor eliciting a toxic episode. NAT1 seems to be ubiquitouslyexpressed among most human tissues, whereas NAT2 is found inliver and the GI tract. Characteristic of both NAT1 and NAT2 isthe ability to form N-hydroxy–acetylated metabolites from bicyclic
aromatic hydrocarbons, a reaction that leads to the nonenzymaticrelease of the acetyl group and the generation of highly reactivenitrenium ions. Thus, N-hydroxy acetylation is thought to activatecertain environmental toxicants. In contrast, direct N-acetylation ofthe environmentally generated bicyclic aromatic amines is stableand leads to detoxification. Individuals who are NAT2 fast acetyla-tors are able to efficiently metabolize and detoxify bicyclic aromat-ic amine through liver-dependent acetylation. Slow acetylators(NAT2 deficient), however, accumulate bicyclic aromatic amines,which then become substrates for CYP-dependent N-oxidation.These N-OH metabolites are eliminated in the urine. In tissues suchas bladder epithelium, NAT1 is highly expressed and can efficient-ly catalyze the N-hydroxy acetylation of bicyclic aromatic amines,a process that leads to de-acetylation and the formation of themutagenic nitrenium ion, especially in NAT2-deficient subjects.Epidemiological studies have shown that slow acetylators are pre-disposed to bladder cancer if exposed environmentally to bicyclicaromatic amines.
Methylation. In humans, drugs and xenobiotics canundergo O-, N-, and S-methylation. The identification ofthe individual methyltransferase (MT) is based on thesubstrate and methyl conjugate. Humans express threeN-methyltransferases, one catechol-O-methyltransferase(COMT) a phenol-O-methyltransferase (POMT), a thio-purine S-methyltransferase (TPMT), and a thiol methyl-
Figure 3–12. Generation of the reactive alkylating agent following the conjugation of glutathione to TLK286. GST interactswith the prodrug and GSH analog, TLK286, via a tyrosine in the active site of GST. GSH portion is shown in blue. The interac-tion promotes β-elimination and cleavage of the prodrug to a vinyl sulfone and an active alkylating fragment. (See Townsend andTew, 2003.)

Chapter 3 / Drug Metabolism 87
transferase (TMT). All of the MTs exist as monomers anduse S-adenosyl-methionine (SAM; AdoMet) as the methyldonor. With the exception of a signature sequence that isconserved among the MTs, there is limited conservationin sequence, indicating that each MT has evolved to dis-play a unique catalytic function. Although the commontheme among the MTs is the generation of a methylatedproduct, substrate specificity is high and distinguishes theindividual enzymes.
Nicotinamide N-methyltransferase (NNMT) methylates seroto-nin and tryptophan, and pyridine-containing compounds such as nic-otinamide and nicotine. Phenylethanolamine N-methyltransferase(PNMT) is responsible for the methylation of the neurotransmitternorepinephrine, forming epinephrine; the histamine N-methyltrans-ferase (HNMT) metabolizes drugs containing an imidazole ringsuch as that found in histamine. COMT methylates neurotransmit-ters containing a catechol moiety such as dopamine and norepineph-rine, drugs such as methyldopa, and drugs of abuse such as ecstasy(MDMA; 3,4-methylenedioxymethamphetamine).
From a clinical perspective, the most important MT may beTPMT, which catalyzes the S-methylation of aromatic and hetero-
cyclic sulfhydryl compounds, including the thiopurine drugs aza-thioprine (AZA), 6-mercaptopurine (6-MP), and thioguanine.AZA and 6-MP are used for the management of inflammatorybowel disease (see Chapter 38) as well as autoimmune disorderssuch as systemic lupus erythematosus and rheumatoid arthritis.Thioguanine is used in acute myeloid leukemia, and 6-MP is usedworldwide for the treatment of childhood acute lymphoblasticleukemia (see Chapter 51). Because TPMT is responsible for thedetoxification of 6-MP, a genetic deficiency in TPMT can resultin severe toxicities in patients taking these drugs. When givenorally at clinically established doses, 6-MP serves as a prodrugthat is metabolized by hypoxanthine guanine phosphoribosyltransferase (HGPRT) to 6-thioguanine nucleotides (6-TGNs),which become incorporated into DNA and RNA, resulting inarrest of DNA replication and cytotoxicity. The toxic side effectsarise when a lack of 6-MP methylation by TPMT causes a build-up of 6-MP, resulting in the generation of toxic levels of 6-TGNs.The identification of the inactive TPMT alleles and the develop-ment of a genotyping test to identify homozygous carriers of thedefective allele have now made it possible to identify individualswho may be predisposed to the toxic side effects of 6-MP thera-py. Simple adjustments in the patient’s dosage regiment havebeen shown to be a life-saving intervention for those with TPMTdeficiencies.
Table 3–3Indications and Unwanted Side Effects of Drugs Metabolized by N-Acetyltransferases
DRUG INDICATION MAJOR SIDE EFFECTS
Acebutolol Arrhythmias, hypertension Drowsiness, weakness, insomniaAmantadine Influenza A, parkinsonism Appetite loss, dizziness, headache, nightmaresAminobenzoic acid Skin disorders, sunscreens Stomach upset, contact sensitizationAminoglutethimide Adrenal cortex carcinoma, breast cancer Clumsiness, nausea, dizziness, agranulocytosisAminosalicylic acid Ulcerative colitis Allergic fever, itching, leukopeniaAmonafide Prostate cancer MyelosuppressionAmrinone Advanced heart failure Thrombocytopenia, arrhythmiasBenzocaine Local anesthesia Dermatitis, itching, rash, methemoglobinemiaCaffeine Neonatal respiratory distress syndrome Dizziness, insomnia, tachycardiaClonazepam Epilepsy Ataxia, dizziness, slurred speechDapsone Dermatitis, leprosy, AIDS-related
complexNausea, vomiting, hyperexcitability, methemo-
globinemia, dermatitisDipyrone, metamizole Analgesic AgranulocytosisHydralazine Hypertension Hypotension, tachycardia, flush, headacheIsoniazid Tuberculosis Peripheral neuritis, hepatotoxicityNitrazepam Insomnia Dizziness, somnolencePhenelzine Depression CNS excitation, insomnia, orthostatic hypoten-
sion, hepatotoxicityProcainamide Ventricular tachyarrhythmia Hypotension, systemic lupus erythematosusSulfonamides Antibacterial agents Hypersensitivity, hemolytic anemia, fever,
lupuslike syndromes
For details, see Meisel, 2002.

88 Section I / General Principles
The Role of Xenobiotic Metabolism in the Safe andEffective Use of Drugs. Any compound entering thebody must be eliminated through metabolism and excre-tion via the urine or bile/feces. This mechanism keepsforeign compounds from accumulating in the body andpossibly causing toxicity. In the case of drugs, metabo-lism results in the inactivation of their therapeutic effec-tiveness and facilitates their elimination. The extent ofmetabolism can determine the efficacy and toxicity of adrug by controlling its biological half-life. Among themost serious considerations in the clinical use of drugsare adverse drug reactions. If a drug is metabolized tooquickly, it rapidly loses its therapeutic efficacy. Thiscan occur if specific enzymes involved in metabolismare overly active or are induced by dietary or environ-mental factors. If a drug is metabolized too slowly, thedrug can accumulate in the bloodstream; as a conse-quence, the pharmacokinetic parameter AUC (area underthe plasma concentration–time curve) is elevated andthe plasma clearance of the drug is decreased. Thisincrease in AUC can lead to overstimulation of sometarget receptors or undesired binding to other receptorsor cellular macromolecules. An increase in AUC oftenresults when specific xenobiotic-metabolizing enzymesare inhibited, which can occur when an individual istaking a combination of different therapeutic agents andone of those drugs targets the enzyme involved in drugmetabolism. For example, the consumption of grapefruitjuice with drugs taken orally can inhibit intestinalCYP3A4, blocking the metabolism of many of thesedrugs. The inhibition of specific CYPs in the gut bydietary consumption of grapefruit juice alters the oralbioavailability of many classes of drugs, such as certainantihypertensives, immunosuppressants, antidepressants,antihistamines, and the statins, to name a few. Amongthe components of grapefruit juice that inhibit CYP3A4are naringin and furanocoumarins.
While environmental factors can alter the steady-state levels of specific enzymes or inhibit their catalyticpotential, these phenotypic changes in drug metabo-lism are also observed clinically in groups of individu-als that are genetically predisposed to adverse drugreactions because of pharmacogenetic differences inthe expression of xenobiotic-metabolizing enzymes(see Chapter 4). Most of the xenobiotic-metabolizingenzymes display polymorphic differences in theirexpression, resulting from heritable changes in thestructure of the genes. For example, as discussedabove, a significant population was found to be hyper-bilirubinemic, resulting from a reduction in the abilityto glucuronidate circulating bilirubin due to a lowered
expression of the UGT1A1 gene (Gilbert’s syndrome).Drugs that are subject to glucuronidation by UGT1A1,such as the topoisomerase inhibitor SN-38 (Figures 3–5, 3–7, and 3–8), will display an increased AUCbecause individuals with Gilbert’s syndrome are unableto detoxify these drugs. Since most cancer chemothera-peutic agents have a very narrow therapeutic index,increases in the circulating levels of the active formcan result in significant toxicities. There are a numberof genetic differences in CYPs that can have a majorimpact on drug therapy.
Nearly every class of therapeutic agent has been reported toinitiate an adverse drug response (ADR). In the United States, thecost of such response has been estimated at $100 billion and to bethe cause of over 100,000 deaths annually. It has been estimatedthat 56% of drugs that are associated with adverse responses aresubjected to metabolism by the xenobiotic-metabolizing enzymes,notably the CYPs, which metabolize 86% of these compounds.Since many of the CYPs are subject to induction as well as inhibi-tion by drugs, dietary factors, and other environmental agents,these enzymes play an important role in most ADRs. Thus, it hasbecome mandatory that before a new drug application (NDA) isfiled with the Food and Drug Administration, the route of metabo-lism and the enzymes involved in the metabolism must be known.As a result, it has become routine practice in the pharmaceuticalindustry to establish which enzymes are involved in metabolism ofa drug candidate and to identify the metabolites and determinetheir potential toxicity.
Induction of Drug Metabolism. Xenobiotics can influ-ence the extent of drug metabolism by activating trans-cription and inducing the expression of genes encodingdrug-metabolizing enzymes. Thus, a foreign compoundmay induce its own metabolism, as may certain drugs.One potential consequence of this is a decrease in plas-ma drug concentration over the course of treatment,resulting in loss of efficacy, as the auto-induced metabo-lism of the drug exceeds the rate at which new drugenters the body. A list of ligands and the receptorsthrough which they induce drug metabolism is shown inTable 3–4. A particular receptor, when activated by aligand, can induce the transcription of a battery of targetgenes. Among these target genes are certain CYPs anddrug transporters. Thus, any drug that is a ligand for areceptor that induces CYPs and transporters could leadto drug interactions. Figure 3–13 shows the scheme bywhich a drug may interact with nuclear receptors toinduce its own metabolism.
The aryl hydrocarbon receptor (AHR) is a memberof a superfamily of transcription factors with diverseroles in mammals, such as a regulatory role in thedevelopment of the mammalian CNS and modulatingthe response to chemical and oxidative stress. This

Chapter 3 / Drug Metabolism 89
superfamily of transcription factors includes Per andSim, two transcription factors involved in developmentof the CNS, and the hypoxia-inducible factor 1α (HIF1α)that activates genes in response to low cellular O2
levels. The AHR induces expression of genes encoding
CYP1A1 and CYP1A2, two CYPs that are able to met-abolically activate chemical carcinogens, includingenvironmental contaminants and carcinogens derivedfrom food. Many of these substances are inert unlessmetabolized by CYPs. Thus, induction of these CYPsby a drug could potentially result in an increase in thetoxicity and carcinogenicity of procarcinogens. Forexample, omeprazole, a proton pump inhibitor used totreat gastric and duodenal ulcers (see Chapter 36), is aligand for the AHR and can induce CYP1A1 andCYP1A2, with the possible consequences of toxin/car-cinogen activation as well as drug-drug interactions inpatients receiving agents that are substrates for eitherof these CYPs.
Another important induction mechanism is due to type2 nuclear receptors that are in the same superfamily as thesteroid hormone receptors. Many of these receptors, iden-tified on the basis of their structural similarity to steroidhormone receptors, were originally termed “orphan recep-tors,” because no endogenous ligands were known tointeract with them. Subsequent studies revealed that someof these receptors are activated by xenobiotics, includingdrugs. The type 2 nuclear receptors of most importance to
Table 3–4Nuclear Receptors That Induce Drug Metabolism
RECEPTOR LIGANDS
Aryl hydrocarbon receptor (AHR) OmeprazoleConstitutive androstane receptor
(CAR)Phenobarbital
Pregnane X receptor (PXR) RifampinFarnesoid X receptor (FXR) Bile acidsVitamin D receptor Vitamin DPeroxisome proliferator activat-
ed receptor (PPAR)Fibrates
Retinoic acid receptor (RAR) all-trans-Retino-ic acid
Retinoid X receptor (RXR) 9-cis-Retinoic acid
Figure 3–13. Induction of drug metabolism by nuclear receptor–mediated signal transduction. When a drug such as atorva-statin (Ligand) enters the cell, it can bind to a nuclear receptor such as the pregnane X receptor (PXR). PXR then forms a complexwith the retinoid X receptor (RXR), binds to DNA upstream of target genes, recruits coactivator (which binds to the TATA box bind-ing protein, TBP), and activates transcription. Among PXR target genes are CYP3A4, which can metabolize the atorvastatin anddecrease its cellular concentration. Thus, atorvastatin induces its own metabolism. Atorvastatin undergoes both ortho and para hydrox-ylation. (See Handschin and Meyer, 2003.)
RXR
PXR
OH
PXR
Ligand Ligand
Ligand
CYP3A4
RXR TBP
RNAPIICo-activator
TATA

90 Section I / General Principles
drug metabolism and drug therapy include the pregnaneX receptor (PXR), constitutive androstane receptor(CAR), and the peroxisome proliferator activated receptor(PPAR). PXR, discovered based on its ability to be acti-vated by the synthetic steroid pregnane 16α-carbonitrile,is activated by a number of drugs including, antibiotics(rifampicin and troleandomycin), Ca2+ channel blockers(nifedipine), statins (mevastatin), antidiabetic drugs (tro-glitazone), HIV protease inhibitors (ritonavir), and anti-cancer drugs (paclitaxel). Hyperforin, a component of St.John’s wort, an over-the-counter herbal remedy used fordepression, also activates PXR. This activation is thoughtto be the basis for the increase in failure of oral contracep-tives in individuals taking St. John’s wort: activated PXRis an inducer of CYP3A4, which can metabolize steroidsfound in oral contraceptives. PXR also induces theexpression of genes encoding certain drug transportersand phase 2 enzymes including SULTs and UGTs. Thus,PXR facilitates the metabolism and elimination of xenobi-otics, including drugs, with notable consequences (Figure3–13).
The nuclear receptor CAR was discovered based on itsability to activate genes in the absence of ligand. Steroidssuch as androstanol, the antifungal agent clotrimazole,and the antiemetic meclizine are inverse agonists thatinhibit gene activation by CAR, while the pesticide 1,4-bis[2-(3,5-dichloropyridyloxy)]benzene, the steroid 5β-pregnane-3,20-dione, and probably other endogenouscompounds, are agonists that activate gene expressionwhen bound to CAR. Genes induced by CAR includethose encoding several CYPs (CYP2B6, CYP2C9, andCYP3A4), various phase 2 enzymes (including GSTs,UGTs, and SULTs), and drug and endobiotic transporters.CYP3A4 is induced by both PXR and CAR and thus itslevel is highly influenced by a number of drugs and otherxenobiotics. In addition to a potential role in inducing thedegradation of drugs including the over-the-counter anal-gesic acetaminophen, this receptor may function in thecontrol of bilirubin degradation, the process by which theliver decomposes heme.
Clearly, PXR and CAR have a capacity for binding agreat variety of ligands. As with the xenobiotic-metaboliz-ing enzymes, species differences also exist in the ligandspecificities of these receptors. For example, rifampicinactivates human PXR but not mouse or rat PXR. Meclizinepreferentially activates mouse CAR but inhibits geneinduction by human CAR. These findings further establishthat rodent model systems do not reflect the response ofhumans to drugs.
The peroxisome proliferator activated receptor (PPAR)family is composed of three members, α, β, and γ.
PPARα is the target for the fibrate class of hyperlipi-demic drugs, including the widely prescribed gemfi-brozil and fenofibrate. While activation of PPARαresults in induction of target genes encoding fatty acidmetabolizing enzymes that result in lowering of serumtriglycerides, it also induces CYP4 enzymes that carryout the oxidation of fatty acids and drugs with fattyacid–containing side chains, such as leukotriene andarachidonic acid analogs.
Role of Drug Metabolism in the Drug DevelopmentProcess. There are two key elements associated withsuccessful drug development: efficacy and safety. Bothdepend on drug metabolism. It is necessary to determinehow and by which enzymes a potential new drug ismetabolized. This knowledge allows prediction of wheth-er the compound may cause drug-drug interactions or besusceptible to marked interindividual variation in metabo-lism due to genetic polymorphisms.
Historically, drug candidates have been administered to rodentsat doses well above the human target dose in order to predict acutetoxicity. For drug candidates to be used chronically in humans, suchas for lowering serum triglycerides and cholesterol or for treatmentof type 2 diabetes, long-term carcinogenicity studies are carried outin rodent models. For determination of metabolism, the compoundis subjected to analysis by human liver cells or extracts from thesecells that contain the drug-metabolizing enzymes. Such studiesdetermine how humans will metabolize a particular drug, and to alimited extent, predict the rate of metabolism. If a CYP is involved,a panel of recombinant CYPs can be used to determine which CYPpredominates in the metabolism of the drug. If a single CYP, suchas CYP3A4, is found to be the sole CYP that metabolizes a drugcandidate, then a decision can be made about the likelihood of druginteractions. Interactions become a problem when multiple drugs aresimultaneously administered, for example in elderly patients, whoon a daily basis may take prescribed antiinflammatory drugs, cho-lesterol-lowering drugs, blood pressure medications, a gastric acidsuppressant, an anticoagulant, and a number of over-the-countermedications. Ideally, the best drug candidate would be metabolizedby several CYPs so that variability in expression levels of one CYPor drug-drug interactions would not significantly impact its metabo-lism and pharmacokinetics.
Similar studies can be carried out with phase 2 enzymes anddrug transporters in order to predict the metabolic fate of a drug. Inaddition to the use of recombinant human xenobiotic-metabolizingenzymes in predicting drug metabolism, human receptor–based(PXR and CAR) systems should also be used to determine whethera particular drug candidate could be a ligand for PXR, CAR, orPPARα.
Computer-based computational (in silico) prediction of drugmetabolism may also be a prospect for the future, since the struc-tures of several CYPs have been determined, including those ofCYPs 2A6, 2C9, and 3A4. These structures may be used to predictmetabolism of a drug candidate by fitting the compound to theenzyme’s active site and determining oxidation potentials of sites onthe molecule. However, the structures, determined by x-ray analysis

Chapter 3 / Drug Metabolism 91
of crystals of enzyme-substrate complexes, are static, whereasenzymes are flexible, and this vital distinction may be limiting. Thelarge size of the CYP active sites, which permits them to metabolizemany different compounds, also renders them difficult to model.The potential for modeling ligand or activator for the nuclear recep-tors also exists with limitations similar to those discussed for theCYPs. With refinement of structures and more powerful modelingsoftware, in silico drug metabolism may be a reality in the future.
BIBLIOGRAPHY
Blanchard, R.L., Freimuth, R. R., Buck, J., et al. A proposed nomencla-ture system for the cytosolic sulfotransferase (SULT) superfamily.Pharmacogenetics, 2004, 14:199–211.
Cashman, J.R. The implications of polymorphisms in mammalian fla-vin-containing monooxygenases in drug discovery and development.Drug Discov. Today, 2004, 9:574–581.
Coughtrie, M.W. Sulfation through the looking glass—recent advancesin sulfotransferase research for the curious. Pharmacogenomics J.,2002, 2:297–308.
Dutton, G.J. Glucuronidation of drugs and other compounds. CRCPress, Inc., Boca Raton, 1980.
Evans, W.E., and Relling, M.V. Pharmacogenomics: translating functionalgenomics into rational therapeutics. Science, 1999, 286:487–491.
Glatt, H. Sulfotransferases in the bioactivation of xenobiotics. Chem.Biol. Interact., 2000, 129:141–170.
Golka, K., Prior, V., Blaszkewicz, M. and Bolt, H.M. The enhancedbladder cancer susceptibility of NAT2 slow acetylators towards aro-matic amines: a review considering ethnic differences. Toxicol. Lett.,2002, 128:229–241.
Gong Q.H., Cho J.W., Huang T., et al. Thirteen UDP glucuronosyltrans-ferase genes are encoded at the human UGT1 gene complex locus.Pharmacogenetics, 2001, 11:357–368.
Hayes, J.D., Flanagan, J.U., and Jowsey, I.R. Glutathione transferases.Annu. Rev. Pharmacol. Toxicol., 2004.
Handschin, C., and Meyer, U.A. Induction of drug metabolism: the roleof nuclear receptors. Pharmacol. Rev., 2003, 55:649–673.
Kadakol, A., Ghosh, S.S., Sappal, B.S., et al. Genetic lesions of bilirubinuridine-diphosphoglucuronate glucuronosyltransferase (UGT1A1)causing Crigler-Najjar and Gilbert syndromes: correlation of geno-type to phenotype. Hum. Mutat., 2000, 16:297–306.
Kobayashi, K., Bouscarel, B., Matsuzaki, Y., et al. PH-dependent uptakeof irinotecan and its active metabolite, SN-38, by intestinal cells. Int.J. Cancer, 1999, 83:491–496.
Krynetski, E.Y., and Evans, W.E. Pharmacogenetics of cancer therapy:getting personal. Am. J. Hum. Genet., 1998, 63:11–16.
Landi, S. Mammalian class theta GST and differential susceptibility tocarcinogens: a review. Mut. Res., 2000, 463:247–283.
Lee, K.A., Fuda, H., Lee, Y.C., et al. Crystal structure of human choles-terol sulfotransferase (SULT2B1b) in the presence of pregnenoloneand 3'- phosphoadenosine 5'-phosphate. Rationale for specificity dif-ferences between prototypical SULT2A1 and the SULT2BG1 iso-forms. J. Biol. Chem., 2003, 278:44593–44599.
Meisel, P. Arylamine N-acetyltransferases and drug response. Pharma-cogenomics, 2002, 3:349–366.
Negishi, M., Pederson L. G., Petrotchenko, E., et al. Structure and func-tion of sulfotransferases. Arch. Biochem. Biophys., 2001, 390:149–157.
Nelson, D.R., Zeldin, D.C., Hoffman, S.M., et al. Comparison of cyto-chrome P450 (CYP) genes from the mouse and human genomes,including nomenclature recommendations for genes, pseudogenes andalternative-splice variants. Pharmacogenetics, 2004, 14:1–18.
Pedersen, L.C., Petrotchenko, E., Shevtsov, S., and Negishi, M. Crystalstructure of the human estrogen sulfotransferase-PAPS complex: evi-dence for catalytic role of Ser137 in the sulfuryl transfer reaction. J.Biol. Chem., 2002, 277:17928–17932.
Townsend, D.M., and Tew, K.D. The role of glutathione-S-transferase inanti-cancer drug resistance. Oncogene, 2003, 22:7369–7375.
Tukey, R.H., and Strassburg, C.P. Human UDP-glucuronosyltransferas-es: metabolism, expression, and disease. Annu. Rev. Pharmacol. Toxi-col., 2000, 40:581–616.
Tukey, R.H., Strassburg, C.P., and Mackenzie, P.I. Pharmacogenomicsof human UDP-glucuronosyltransferases and irinotecan toxicity. Mol.Pharmacol., 2002, 62:446–450.
Vatsis, K.P., Weber, W.W., Bell, D.A., et al. Nomenclature for N-acetyltransferases. Pharmacogenetics, 1995, 5:1–17.
Weinshilboum, R.M., Otterness, D.M., and Szumlanski, C.L. Methyla-tion pharmacogenetics: catechol O-methyltransferase, thiopurinemethyltransferase, and histamine N-methyltransferase. Annu. Rev.Pharmacol. Toxicol., 1999, 39:19–52.
Yamamoto, W., Verweij, J., de Bruijn, P., et al. Active transepithelialtransport of irinotecan (CPT-11) and its metabolites by human intesti-nal Caco-2 cells. Anticancer Drugs, 2001, 12:419–432.
Williams, P.A., Cosme, J., Vinkovic, D.M., et al. Crystal structures ofhuman cytochrome P450 3A4 bound to metyrapone and progesterone.Science, 2004, 305:683–686.

92—blank





























