Embed Size (px)
Citation preview

Online Submission ID: 0369
Generalizing Locomotion Style to New Animals With Joint Inverse Optimization
Abstract1
We present a technique for analyzing a set of animal gaits to predict2
the gait of a new animal from its shape alone. The core to our3
method is a novel algorithm called joint inverse optimization which4
learns coherent patterns in motion style from a database of example5
animal-gait pairs. This analysis gives rise to a generative model6
that can synthesize a realistic gait for a new animal by interpolating7
gaits of similarly shaped animals in the database. We quantify the8
predictive performance of our model by comparing its synthesized9
gaits to ground truth motions for a range of different animals. We10
also apply our method to the prediction of gaits for dinosaurs and11
other extinct creatures.12
CR Categories: I.3.7 [Computer Graphics]: Three-Dimensional13
Graphics and Realism—Animation;14
Keywords: character animation,optimization,locomotion15
1 Introduction16
The animals seen in nature come in a great variety of shapes and17
sizes, and move in a similarly diverse range of ways. Although the18
availability of video footage and motion capture data makes it easy19
to see how these animals locomote, natural curiosity compels us to20
wonder beyond this data to the motions of other creatures. How21
might animals which we don’t have data for move? Can the gaits22
of living animals be used to guess at the gait for a dinosaur?23
Our approach to gait synthesis takes a step toward answering these24
questions by using a database of real-world gaits to learn a model25
of how different animals move. We can then synthesize physically26
valid gaits for other animals according to the similarity of their27
shapes to the different animals in the database. This form of gen-28
eralization is made difficult by the fact that many of its most in-29
teresting applications are on to creatures (such as dinosaurs) with30
a shape substantially different than anything currently living. This31
necessitates a method which not only captures the salient aspects of32
the gaits for the animals for which we do have data, but which is33
also well-suited for generalizing these gaits to new creatures.34
Our approach to addressing these issues is based on a novel algo-35
rithm called joint inverse optimization. This method takes as input36
a database of animal gaits extracted from video footage, and pro-37
cesses it into a compact form describing the style of each gait. This38
style is captured with a set of biologically meaningful parameters39
describing, for instance, the relative preference for using different40
joints, the stiffness of these joints, and the preference for avoid-41
ing motions likely to cause the animal to trip. Unlike approaches42
such as [Liu et al. 2005] relying on traditional inverse optimization,43
our joint inverse optimization approach ensures that the learned pa-44
rameters are well-suited to generalization onto new animals. Thus,45
instead of learning specific values of these parameters from a single46
motion, we learn coherent patterns between the parameters for an47
entire set of different motions. These coherent patterns in style are48
then used to guess the manner in which a new input animal should49
move. This technique allows the synthesis of gaits for a wide range50
of extinct animals, and is validated to be more accurate then several51
alternative approaches at reproducing the gaits of a range of living52
animals for which we do have data.53
2 Related Work54
The synthesis of realistic legged locomotion is a well-studied but55
difficult problem in character animation. Although part of this dif-56
ficulty is inherent in the synthesis of locomotion (realistic or oth-57
erwise) for complex characters, synthesizing realistic locomotion58
presents the particular challenge in that it requires a precise defini-59
tion of what constitutes “realistic”. Although this problem can be60
addressed by relying on an artist or expert in the creation of the mo-61
tion [Raibert and Hodgins 1991; Yin et al. 2007; Coros et al. 2010;62
Kry et al. 2009; de Lasa et al. 2010], the most common method for63
realistic locomotion synthesis is probably to rely on motion capture64
or video data as a guide. This use of real-wold motion data is seen65
in methods which directly rearrange and replay existing motions66
[Bruderlin and Williams 1995; Witkin and Popovic 1995; Lee et al.67
2002; Arikan and Forsyth 2002; Kovar et al. 2002], as well as ap-68
proaches which transform an existing motion [Popovic and Witkin69
1999; Liu et al. 2005] or which take a set of related motions for a70
character and use these to create new motions for the same charac-71
ter [Safonova et al. 2004; Wei et al. 2011].72
The reliance of many methods on pre-existing motion data poses73
a special problem when it comes to the synthesis of realistic loco-74
motion for animals. Not only is it substantially harder to acquire75
motion data for animals than for humans, but there is also the more76
fundamental problem that acquiring motion data for extinct or fic-77
tional animals is impossible even in principle. Thus while some78
techniques have been developed to successfully synthesize legged79
animal locomotion, existing approaches either rely on motion data80
or artist interaction in specifying the style of the motions [Coros81
et al. 2011; Nunes et al. 2012], or do not focus on generating real-82
istic motions [Fang and Pollard 2003; Wampler and Popovic 2009;83
Mordatch et al. 2010; Mordatch et al. 2012].84
There are two extremes to the types of approaches which might hy-85
pothetically be taken to synthesize realistic animal motions. At one86
end of the spectrum one might attempt to increase the realism of87
the synthesized gaits by using a more faithful biomechanical model88
of an animal. This approach as been applied with some success to89
humanoid bipedal locomotion by the modeling of soft tissue defor-90
mations in the feet [Jain and Liu 2011] or by including a more accu-91
rate biomechanical and metabolic modeling of leg muscles [Wang92
et al. 2012]. Although these approaches have been successful in im-93
proving the realism of the synthesized motions, they have not yet94
been shown to generalize to a wide range of different animal shapes.95
One hurdle in creating a method of this sort which can be applied96
equally to any of a wide range of different animals lies in the burden97
of specifying the additional details about the biomechanical struc-98
ture of each animal. A second, perhaps more substantial hurdle lies99
in the fact that it is not yet clear that the biomechanical models used100
thus far are sufficient to capture the variations in motion observed in101
animals of diverse shapes, or even what the sufficient set of salient102
principles would be.103
On the other end of the spectrum one might attempt to synthesize104
an animal’s gait by forgoing any consideration of the principles un-105
derlying the motion and instead attempting to directly match the106
appearance of motions for which data does exist. This approach107
has the advantage that if a new animal happens to be very similar108
in shape to an animal for which there is data, then it would be ex-109
pected to give a very accurate prediction of the motion for the new110
animal. Unfortunately, it is not clear how well this sort of approach111
can work when applied to an animal that has a shape or mass sub-112
1

Online Submission ID: 0369
stantially different than anything data is available for. Indeed, the113
primary successful application of this approach to creatures with114
diverse shapes has thus far been limited to highly stylized motions115
[Hecker et al. 2008]. Moreover, these cases where the animal is116
relatively unlike anything one could obtain data for cover some of117
the most interesting applications of computational locomotion syn-118
thesis. A restriction to synthesizing gaits only for animals similar119
to those that exist would preclude the generation of gaits for most120
dinosaurs and many other extinct animals.121
Given the difficulties apparent in each of these two extremes on122
the spectrum of approaches to realistic locomotion synthesis, our123
method takes a middle ground and combines data-based interpo-124
lation with biologically and physically motivated factors. An in-125
stance of this type of ‘middle ground’ approach has been previously126
applied to human locomotion [Liu et al. 2005; Lee and Popovic127
2010] by learning the passive actuation characteristics at a charac-128
ter’s joints from a sequence of motion capture and then using this129
information to create new motions for the same character. Although130
these passive actuation characteristics are relatively simple biolog-131
ically motivated entities approximating the nature of tendons and132
ligaments, their particular values were derived from a sequence of133
motion capture data. Our method is in the same spirit of these ap-134
proaches, but applies to the analysis of a set of gaits for animals135
with widely varying shapes. The primary complication that arises136
in doing so is that in order to synthesize motions for new creatures,137
the parameters used to represent the style of each animal’s gait must138
be suited to interpolation on to animals for which there is no motion139
capture data available.140
3 Algorithm Overview141
Our approach to gait synthesis rests upon three primary compo-142
nents, a motion database, a generative model, and an algorithm for143
tuning the generative model to agree with the motion database we144
call joint inverse optimization. The motion database consists of a145
set of pairings (A1,M1), . . . , (An,Mn), each of which associates146
the shape of an animalAi with its ground-truth gaitMi as extracted147
from real-world video data. While this motion database covers a set148
of known animal motions, the synthesis of new motions is handled149
by a generative model, denoted f . This generative model takes150
as input an animal A and a vector of parameters φ describing the151
style of the motion to be synthesized, and outputs an associated gait152
f(A, φ). The goal of our gait synthesis technique can be summa-153
rized as follows: given a new animal Anew, find the parameters154
φnew of the generative model such that the style of the resulting155
motion f(Anew, φnew) matches what would be expected given the156
motion database.157
Intuitively this approach mimics that which an artist might take158
when animating an animal for which they do not have any video159
footage – try to guess the the motion for the new animal by appeal-160
ing to animals for which the artist does have video footage. Synthe-161
sizing locomotion in this manner requires that several sub-problems162
be addressed:163
Motion Database Creation The motion database contains164
ground-truth motions for a wide range of different animals.165
This database is created by tracking a set of points in a166
real-world video of the animal walking, then solving a167
spacetime constraints optimization to fit a physically realistic168
cyclic gait to the motion of these points.169
Generative Model The generative model f(A, φ) is used to syn-170
thesize new motions. In keeping with many existing locomo-171
tion synthesis techniques, we base the generative model on an172
optimization. That is, given an animal A, the motion f(A, φ)173
is chosen so as to minimize some objective function which is174
itself parameterized by φ. A key ingredient in the definition175
of f lies in choosing a set of parameters φ which is expressive176
enough to capture the variations in style between the different177
animals in the motion database.178
Joint Inverse Optimization To synthesize gaits for new animals,179
the motion database is preprocessed with a technique we term180
joint inverse optimization. This optimization estimates a vec-181
tor of parameters φi for each (Ai,Mi) in the motion database182
such that f(Ai, φi) ≈ Mi and such that the φi parameters183
are well-suited for interpolation to new animals by standard184
regression or sparse data interpolation techniques.185
With these three components in place, a motion for a new animal186
Anew can be synthesized by first interpolating φnew from the φi187
parameters associated with similar animals in the motion database,188
then solving for the final motion with f(Anew, φnew). The follow-189
ing three sections will cover each of these components in further190
detail.191
4 Motion Database Creation192
The motion database is used to define a mapping from the shape193
of an animal to the way that animal moves in the real world. Each194
entry associates a particular animal Ai with a cyclic gait Mi pre-195
scribing the animal’s ground truth walking motion. Although in196
principle our approach is applicable to non-walking gaits, all of the197
gaits in a motion database should be of the same type. We have198
therefore focused on walks due to the relative ease of obtaining199
video data of walks for a wide range of different animals. In syn-200
thesizing a gait for a new animal (described later in section 6) this201
motion database is used as a reference to estimate what parameters202
of the generative model are likely to result in a realistic motion for203
a new animal.204
An animal A is represented as a kinematic tree of limbs connected205
by joints. Each joint describes a parameterized rotation from its206
parent limb to its child limb, and each limb has an associated length207
and mass. In addition, each animal’s representation marks the limb208
corresponding to the head and the position of each foot. A pose for209
an animal is described by a vector giving the rotational parameters210
of each of its joints, the global translation and rotation of the animal,211
and the ground reaction forces and torques at each foot in contact212
with the ground. A sequence of such poses forms a motion M , and213
the pose associated with a particular frame at time t inM is denoted214
M(t).215
Although an ideal motion database would be constructed using 3D216
motion capture and force plates, this sort of data is currently diffi-217
cult to obtain for a wide range of different animals. Instead, each218
of the Mi motions in our motion database is created by fitting a219
cyclic 3D motion to standard 2D video footage of the animal walk-220
ing. We have obtained this data from online video sharing sites such221
as YouTube and Flickr Video, as they represent the most easily ac-222
cessible resource for such video footage. Each video consists of223
a side-on view of the animal walking with only rotational camera224
motion so as to avoid parallax.225
Our motion database consists of six bipeds and twelve quadrupeds226
spanning a wide range of animal shapes and sizes as shown in fig-227
ures 1 and 2. For each of these animals we determine the relative228
lengths and masses of the animal’s limbs by manually constructing229
a crude 3D model from a selected frame in the video, then uni-230
formly scaling each limb’s length and mass so that the animal’s231
height and total mass match the values in figures 1 and 2. For232
each video we also remove the effects of any camera motion by233
translationally stabilizing based on SURF features [Bay et al. 2008]234
tracked between the frames in the video.235
2

Online Submission ID: 0369
greater flamingo
1.1m 3kg
ostrich
2.3m 94.5kg
secretary bird
0.8m 3.3kg
greater roadrunner
0.22m 0.3kg
emu
1.5m 30.0kg
southern groundhornbill
0.9m 3.6kg
Figure 1: The six bipeds in the motion database along with theirassociated heights and masses.
Next, a number of points on the animal are manually tracked across236
the video. These points vary somewhat from animal to animal, but237
always include at least two points on the torso, at least one on the238
head, as well as points on the knees and feet. Each of these points239
gives a 2D trace of some point on the animal tracked through each240
frame in the video. A sub-segment of the frames in the video rep-241
resenting a single gait cycle is also selected. In order to match each242
2D point trace with the motion of a 3D point on the animal, a few243
frames are chosen on which a 3D model for the animal is superim-244
posed over the video. Using this superposition, each 2D point is245
attached to one of the animal’s limbs, allowing coordinates for each246
point to be determined in a coordinate system local to the point’s247
associated limb. This allows the accuracy with which a 3D gait248
matches the traces of the points in the video to be determined as249
described next.250
The final step is to solve for a cyclic gait M for each animal A251
such that the motion of this gait matches that of the 2D point traces252
extracted from the animal’s associated video. For a given frame253
at time ti in the video let dj(ti) denote the 2D position of the jth254
point in the extracted traces, and denote the 2D X-Y projection of255
the associated point on the animal in a motion M with pj(M(ti)).256
If M were a perfect fit to the data then it would hold that dj(ti) =257
pj(M(ti)) for each frame i and point j. Since in general a perfect258
fit to the data will not be possible, the degree to which a motion259
M matches the data is quantified independently at each frame with260
a sum of squared distances between each point in the traces ex-261
tracted from video and the associated point on the animal in the262
pose M(ti):263
fit(M(ti)) =∑j
wj‖dj(ti)− pj(M(ti))‖22 (1)
Here wj is a weight assigned to each point on the animal to give264
preference to a more accurate fit of the knee and foot positions265
(weight of 3) while allowing greater errors in the fit of points on266
the torso (weight of 0.5), all other points having weight 1.0.267
The motion M used to represent the ground truth gait for the an-imal in the motion database is then solved for with a spacetimeconstraints optimization. This generates a 3D gait closely matching
horse
1.55m 500kg
Thomson’sgazelle
0.55m 20kg
pronghornantelope
0.9m 50kg
bison
1.7m 700kg
house cat
0.24m 4.5kg
cheetah
0.85m 50kg
elephant
3.7m 5200kg
giraffe
5.5m 1000kgmoose
1.8m 320kg
rhinoceros
1.6m 1600kg
steenbok
0.5m 17kg
tiger
1.0m 180kg
Figure 2: The twelve quadrupeds in the motion database alongwith their associated heights and masses.
the data to as the solution of the nonlinear programming problem:
M = argmin∑i
[fit(M(ti)) + α · cost(M(ti))]
s.t. gkeq (M(ti)) = 0 ∀keq , ihkieq (M(ti)) ≤ 0 ∀kieq , i
(2)
where cost(M(ti)), gkeq (M(ti)), and hkieq (M(ti)) are all defined268
to match the objective, equality, and inequality constraints respec-269
tively described in [Wampler and Popovic 2009]. This definition of270
cost(M(ti)) penalizes high-energy motions, while gkeq (M(ti)),271
and hkieq (M(ti)) enforce the laws of Newtonian mechanics. Some272
contribution of cost(M(ti)) is included in the objective as a regu-273
larization term and helps to avoid overfitting to the data. The term274
α controls how strongly minimizing cost(M(ti)) is weighted ver-275
sus minimizing the data fit error. Larger values of α tend to lead to276
smoother motions at the cost of a poorer data fit. Although α is oc-277
casionally altered on a per-animal basis, a value of 10−5 works well278
for most of the animals in the database, with other values ranging279
from 10−3 to 10−8.280
5 Generative Model281
While the motion database represents the gaits of a set of animals as282
extracted from video footage, the synthesis of new gaits is handled283
by a generative model f , taking as input the shape of an animal284
A and a vector φ parameterizing the style of the gait to be gen-285
erated. This generative model is capable of synthesizing gaits for286
animals with greatly different shapes and sizes, as well as for ex-287
tinct or imaginary creatures. The remainder of this section will be288
concerned with the definition of f and φ, while section 6 will ad-289
dress which particular value of φ should be used for a given input290
animal.291
A good generative model should be flexible enough so that for any292
of a wide range of different input animals, it is possible to synthe-293
3

Online Submission ID: 0369
size a realistic gait of that animal by an appropriate choice of φ.294
More concretely, for each Ai and Mi in the motion database there295
should be some φi such that Mi ≈ f(Ai, φi). In addition, when296
synthesizing motions for new animals it will be useful for the val-297
ues of φ to be both relatively compact and coherently interpolable298
between different animals.299
Our generative model f is based on the spacetime constraints opti-mization described by [Wampler and Popovic 2009] because of itsability to automatically and relatively quickly synthesize motionsfor a wide range of different animals. In this approach, a motion issynthesized by finding the minimum of a large nonlinear program-ming problem:
f(A, φ) = argminM
∑i
cost(M(ti), φ)
s.t. gkeq (M(ti)) = 0 ∀keq , ihkieq (M(ti)) ≤ 0 ∀kieq , i (3)
Where gkeq and hkieq are the again the equality and inequality con-300
straints enforcing the physical validity of the resulting motion as301
described by [Wampler and Popovic 2009]. In or implementation,302
this optimization is initialized with the animal a default stand-still303
pose and is solved with the SNOPT nonlinear programming library304
[Gill et al. 2005].305
The objective function used in equation 3 dictates which sorts of306
gaits should be preferred over others, and thus altering this objec-307
tive function allows different gaits to be synthesized for the same308
animal. In previous work by [Liu et al. 2005] this idea has been309
used to synthesize different styles of human locomotion by choos-310
ing φ to represent to passive actuation characteristics at the charac-311
ter’s joints. For our task we extend this set to include parameters312
describing the strength and coactuation of different joints, the un-313
certainty of interaction with the ground, and the relative preference314
for low-torque, low-impact, and smooth motions.315
5.1 Generative parameters316
The elements of φ serving as parameters to our generative model arelisted in table 1 and are used to alter the per-frame objective func-tion cost(M(ti), φ) in equation 3. Adjusting the value of φ thusprovides a means by which the style of a synthesized gait can becontrolled. Our choice of generative parameters and the associatedobjective function were found by starting with an objective functionbased purely on torque-minimization, and iteratively adding termsuntil the gaits of the animals within the motion database could bereproduced. The result is a combination of six terms:
cost(M(ti), φ) = costtorque(M(ti), φ)
+ wf costforce(M(ti), φ)
+ wa costsmooth(M(ti), φ)
+ (ewh − 1) costhead(M(ti), φ)
+ costcoactuate(M(ti), φ)
+ (ewg − 1) costground(M(ti), φ) (4)
There are several sub-quantities which must be used to calculate317
these components, all of which are computed as a function of318
M(ti). For notational cleanness, however, we will leave this de-319
pendence on M(ti) implicit and write for instance τ instead of the320
more explicit τ(M(ti)). Accordingly, τ , f , and q will respectively321
represent a vector of the concatenated torques, forces, and joint ro-322
tation angles at each of the character’s joints at the frame M(ti).323
The quantities Rh, ph, and θh will refer to the rotation matrix, posi-324
tion, and angular acceleration for the limb representing the animal’s325
name biped descriptionhg X falloff of ground uncertainty distributionwg X weight for vertical ground penaltieswhg X weight for horizontal ground penaltieshw X maximum height of near-ground dragdw X drag coefficient for near-ground dragcl X weight on knee-ankle coactuationcrl X target ratio for knee vs. ankle velocitiesca weight on elbow-wrist coactuationcra target ratio for elbow vs. wrist velocitiesth X scaling for torques exerted at the hiptk X scaling for torques exerted at the kneeta X scaling for torques exerted at the anklets scaling for torques exerted at the shoulderte scaling for torques exerted at the elbowtw scaling for torques exerted at the wristwf X scaling for joint-force penaltieswa X scaling for joint-acceleration penaltieswh X scaling for head stability penaltiesksh X spring constant for hipqh X spring rest angle for hipkdh X damper coefficient for hipksk X spring constant for kneeqk X spring rest angle for kneekdk X damper coefficient for kneeksa X spring constant for ankleqa X spring rest angle for anklekda X damper coefficient for anklekss spring constant for shoulderqs spring rest angle for shoulderkds damper coefficient for shoulderkse spring constant for elbowqe spring rest angle for elbowkde damper coefficient for elbowksw spring constant for wristqw spring rest angle for wristkdw damper coefficient for wrist
Table 1: A table of the inverse parameters used to specify the styleof an animal’s gait. Quadrupedal animals make use of the wholeset of parameters, while bipeds make use only of those marked.
head as measured about its center of mass. Similarly, pf represents326
the position of a foot. Any temporal derivatives represented by a327
superscript dot are computed using finite differences.328
The first three components of the objective function in equation 4penalize torques, forces, and angular accelerations at the animal’sjoints:
costtorque(M(ti), φ) = wj · τ (5)costforce(M(ti), φ) = wj · f (6)
costsmooth(M(ti), φ) = ‖q‖2 (7)
where wj is a vector of weights with elements equal to eth − 1,329
etk−1, eta−1, ets−1, ete−1, or etw−1 at indices corresponding330
to the hip, knee, ankle, shoulder, elbow or wrist joints respectively.331
All other elements in wj are fixed equal to 1. Lower settings of332
an element in w approximates the effect of ‘stronger’ joints, which333
are actuated by stronger muscles and better able to withstand large334
forces.335
The term costhead(M(ti), φ) penalizes motion in the animal’s headand is computed as the sum of four sub-terms respectively penaliz-ing the rotation of the head away from forward, its velocity, linear
4

Online Submission ID: 0369
acceleration, and angular acceleration:
costhead(M(ti), φ) = (8)
100‖Rh − I‖22 + 2.5‖p2hyz‖22 + ‖ph‖22 + ‖θh‖22
The weights 100 and 2.5 scaling the first two sub-terms where cho-336
sen empirically. Although costhead(M(ti), φ) does not depend on337
φ, the strength with which it factors into equation 4 is scaled by338
(ewh − 1). The term phyz represents the motion of the head in the339
vertical and lateral directions. The function costhead is useful in340
modeling the fact that animals often take additional effort to stabi-341
lize their head motion to help with visual perception.342
The standard spacetime constraints formulation used to synthe-343
size motions treats each joint in the animal as being capable of344
moving entirely independently of all the other joints. In reality,345
however, some pairs of joints exhibit a tendency to be coactu-346
ated such that their motions occur in concert rather than indepen-347
dently. Rather than directly modeling the muscles responsible for348
this as in [Wang et al. 2012], we take a simplified approach where349
costcoactuate(M(ti), φ) penalizes deviations of the relative veloci-350
ties of the animal’s knee and ankle joints from crl :351
(ecl − 1)(crl qk − qa)2 (9)
Where qk and qa are the rotational velocities at the knee and ankle352
respectively. For a quadruped the parameters ca and cra are used to353
add an analogous additional penalty related to the relative velocities354
of the elbow and wrist joints.355
In the optimization defined by equation 3, the animal is implicitly356
assumed to have perfect knowledge of its environment. In real-357
ity this is of course not the case, and in order to avoid tripping an358
animal will often lift its feet higher than is strictly energetically op-359
timal. Since explicitly modeling this uncertainty as in [Wang et al.360
2010] is difficult within a spacetime constraints formulation, we in-361
stead use the term costground(M(ti), φ) to penalize motions where362
the animal’s foot moves quickly while too close to the ground as:363
costground(M(ti), φ) =∑f
pcx‖pfxz‖2 + max{
0,−pcy pfy
}(10)
where the sum is taken over each of the animal’s feet and pfxz andpfy represent the horizontal and vertical components of the foot’svelocity. The values pcxz and pcy approximate the probability ofan ‘unexpected’ contact between the foot and the ground due to thefoot’s horizontal and vertical motion respectively, calculated as:
pcx = whg (1− e−hgpfy ) (11)
pcy =e−hgp
[2]fy − e−hgpfy
1− e−hgpfy(12)
The value p[2]f indicates the position of the foot in the next frame in364
the motion, i.e. pf(M(ti+1)).365
In addition to the term costground(M(ti), φ) directly penalizing366
motions where the foot skirts too close to the ground, we also indi-367
rectly penalize such motions by approximating the resistive forces368
on the leg resulting from walking through shallow water or near-369
ground vegetation. For each foot f , we add a force f resisting the370
foot’s velocity based on the speed of the foot and it’s depth below371
the height defined by hw:372
f = −pf‖pf‖2 max{
0, hw − pfy
}edw (13)
In practice, we have found the effects of this near-ground resistance373
are useful in conjunction with those from costground(M(ti), φ) to374
shape the trajectory of the animal’s feet during their air phases.375
Finally, we include a number of parameters modeling the passive376
actuation characteristics of the animal’s leg joints. for each of377
the hip, knee, ankle, shoulder, elbow, and wrist joints we include378
three parameters specifying a spring rest length, spring constant,379
and damper coefficient for the joint. The use of these parameters is380
identical to that in [Liu et al. 2005], where they were successfully381
used to capture stylistic variations in human locomotion.382
6 Joint Inverse Optimization383
The motion database provides a reference for the real-world gaits of384
a number of different animals, but to create gaits for new animals it385
is necessary to generalize beyond those contained within the motion386
database. While the generative model f(A, φ) described in section387
5 is capable of synthesizing new motions given an animal A and a388
vector of parameters φ, it remains to be determined which value of389
φ is likely to lead to a realistic motion for a given animal.390
We achieve this by fitting a vector φi of parameters to each Mi in391
the motion database. Although we can fit each φi independently,392
we find that the resulting parameters do not allow us to predict393
the motion of new animals. Instead, we propose a new algorithm394
termed joint inverse optimization which jointly learns all of the φi395
together. The resulting parameters are constructed to allow φ to396
be determined for a new animal by simple interpolation. In the re-397
mainder of this section we will first look more closely at the naıve398
case of independently fitting each φi. We will then introduce our399
algorithm of joint inverse optimization, followed by details on its400
implementation.401
6.1 Naıve Inverse Optimization402
As its name implies, joint inverse optimization is based on the ap-403
proach of inverse optimization that has been previously employed404
in character animation [Liu et al. 2005; Lee and Popovic 2010].405
One possible formulation of an inverse optimization problem as ap-406
plied to animal gaits would determine the φi parameters for each407
(Ai,Mi) in the motion database by solving:408
φi = argminφ
D(f(Ai, φ),Mi) (14)
Where the function D(Ma,Mb) defines a distance metric repre-409
senting in the error in how closely the motion Ma matches Mb.410
See section 6.2.1 for the specifics of the distance function used in411
our implementation.412
The problem with using a standard inverse optimization formula-413
tion to determine the φi parameters for each entry in the motion414
database comes when trying to estimate the φnew which will lead415
to a realistic motion for a new animal. In particular, since equation416
14 solves for each φi independently, there is no guarantee of any417
consistency of these parameters between different animals. Indeed,418
as illustrated in figure 3 the φi parameters found using this method419
do not form any easily describable coherent pattern. This makes the420
use of sparse data interpolation to determine which φnew should be421
used to synthesize a gait for a new animal Anew problematic, and422
our attempts to synthesize plausible gaits for new animals using this423
approach were almost entirely unsuccessful.424
6.2 Joint Inverse Optimization425
Because the end goal of our approach is to use the examples in the426
motion database to determine how a new animal should move, we427
explicitly incorporate this requirement into our joint inverse opti-428
mization formulation. The result is an optimization which not only429
attempts to fit each φi with its associated motionMi, but which also430
5

Online Submission ID: 0369
Figure 3: Plots of the values of a parameter for each of thequadrupedal animals in the motion database. The y-axis of eachof the two plots is the value of the te parameter (see table 1) foreach animal while the x-axis represents the logarithm of each ani-mal’s mass. The top plot shows the parameter values resulting fromnaıve inverse optimization while the bottom plot shows the results ofa joint-inverse optimization. The parameters resulting from joint-inverse optimization are substantially better suited to regression.Note that distances along the x-axis are only an approximation tothe similarity of the animals as measured by equation 21, so a per-fectly smooth curve should not be expected in the lower plot.
minimizes a term ensuring that the result is well-suited to sparse431
data interpolation:432
argminθ,φ1,...,φn
∑i
[D(f(Ai, φi),Mi) + β
(‖φi −R(Ai, θ)‖22 + r(θ)
)](15)
This formulation differs from traditional inverse optimization433
(equation 14) by the addition of two functions: A regression func-434
tion and a regularization function. The regression functionR(A, θ)435
takes as input an animal A (not necessarily in the motion database)436
and a vector of regression parameters θ, and returns a vector of437
generative parameters φ suitable for using to synthesize a gait for438
A. The regularization function r(θ) is then used in preventing an439
overfitting of the regression parameters θ. It is the vector of regres-440
sion parameters θ which determines the motion of a new animal441
Anew, by first calculating φnew = R(Anew, θ), then solving for the442
final motion with f(Anew, φnew).443
Intuitively, the goal of a joint inverse optimization is to find a value444
of the regression parameters θ which simultaneously accurately re-445
produces each motion in the motion database via minimizing each446
D(f(Ai, R(Ai, θ)),Mi), and which avoids overfitting by mini-447
mizing r(θ). Unfortunately, the optimization resulting from at-448
tempting to directly minimize this quantity is extremely brittle due449
to the compounding factor that the generative function f is not guar-450
anteed to converge to a physically valid result, and thus for many451
values of θ there are some animals in the motion database for which452
D(f(Ai, R(Ai, θ)),Mi) is impossible to evaluate.453
The formulation for joint inverse optimization given in equation 15454
provides a more robust and efficient approach. In this approach,455
the φi parameters used to solve for an animal’s motion are allowed456
to differ from the parameters resulting from the regression function457
R(Ai, θ). In essence, although the φ parameters used to synthesize458
a gait for a new animal will be found via φnew = R(A, θ), this459
is treated as a soft constraint for the purposes for fitting θ to the460
motion database.461
When accounting for the possibility that f(Ai, φi) might fail to462
converge to a physically valid result, note that in equation 15 the463
only term which relies on the result of the generative model is464
D(f(Ai, φi),Mi). The fact that this term can be computed in-465
dependently for each animal makes it substantially easier to create466
an optimization which solves equation 15 by simply discarding any467
failed evaluations of f(Ai, φi). Our approach for achieving this468
(including the values used for the term β) is described in section469
6.2.3, but we will first provide the specific definitions of D, R, and470
r which we employ.471
6.2.1 Gait distance metric472
Solving the joint inverse optimization problem defined by equation15 requires a definition of D(Ma,Mb), describing a distance met-ric between different gaits. Although a simple sum-or-squared dif-ferences in the rotational degrees of freedom of the animal’s jointsover the course of the two gaits gives reasonable results, we havefound that more visually pleasing results can be obtained with aslightly more involved definition. Intuitively, this is because someaspects of a motion, such as the height by which the feet are raisedor the stability of the head, are more visually important than theprecise angles of rotation for the animal’s joints. We computeD(Ma,Mb) as a sum of per-frame costs, each computed as:
D(Ma(ti),Mb(ti)) =
‖0.25w · (q(Mb(ti))− q(Ma(ti))) ‖22+ (16)
‖0.6w · (q(Mb(ti))− q(Ma(ti))) ‖22+ (17)
‖0.5w · (q(Mb(ti))− q(Ma(ti))) ‖22+ (18)
2.25∑f
(pf (Mb(ti))y − pf (Ma(ti))y)2+ (19)
3.25∑
h∈{h1,h2}
‖ph(Mb(ti))− ph(Ma(ti))‖22+ (20)
Where q(M(ti)) is a vector of the rotational degrees of freedom473
inM at frame i, and ph1(M(ti)), ph2(M(ti)) give the position of474
the front and back of the head at frame i. Similarly, for each foot475
f , pf (M(ti)) gives the position of the foot at frame i. The vector476
w is used to scale the weight given to differences in the rotations at477
different joints. For bipeds w is 2 for the knee and 3 for ankle joints478
while for quadrupeds w is 3 for the knee/elbow joints and and 10479
for the ankle/wrist joints. All other elements in w are set to 1.480
6.2.2 Regression and Regularization Functions481
A joint inverse optimization additionally requires definitions for the482
regression and regularization functions R(Ai, θ) and r(θ). In our483
implementation we use a relatively simple formulation based on484
regression with radial basis functions regularized with quadratic485
smoothing [Boyd and Vandenberghe 2004]. This approach em-486
ploys a definition of θ which concatenates θ1, . . . , θn for each of487
the n animals in the motion database, where each θi has the same488
dimension as the generative parameters φi.489
The distance metric underlying the radial basis function interpola-tion measures the dissimilarity between two animals Aa and Abwith a weighted combination of the difference of the log-masses ofthe two animals and the difference in the lengths of their variouslimbs normalized by the total size of each animal:
d(Aa, Ab)2 =∑
i
(lai∑j laj
− lbi∑j lbj
)2
+ 0.002(ln(ma)− ln(mb))2
(21)
6

Online Submission ID: 0369
where laj and laj are the lengths of the jth limbs in Aa and Ab490
respectively, and ma and mb are the total respective masses of Aa491
andAb. The logarithmic scaling of the masses captures the fact that,492
for instance, a 10kg difference in mass is much more meaningful493
between a 10kg and a 20kg animal than between a 1000kg and a494
1010kg animal.495
Using this distance metric, the regression function is then defined496
in a manner similar to [Zhang et al. 2004] as:497
R(A, θ) =
∑i θie
−d2i∑i e−d2i
(22)
where di = d(A,Ai)dmin
with dmin = mini
d(A,Ai). Here θi are the498
regression parameters directly associated withAi (which in general499
need not be equal to φi due the decoupling of θ from φ1, . . . , φn in500
equation 15).501
To avoid overfitting, we employ a regularization function based on502
quadratic smoothing:503
r(θ) =∑i
‖R(Ai, θ′)− θi‖22 (23)
where θ′ are the regression parameters omitting the θi parameters504
associated with Ai, so this regularization function essentially com-505
putes a sum of leave-one-out errors.506
Finally, the period pi of an animal’s gait cycle and the speed vi atwhich it should move are determined with a separate method by:
pi = αpmiβp (24)
vi = αvllegiβv (25)
In these equations mi is the total mass of and llegi is the average507
length of the legs of Ai. Additionally, αp and βp are parameters508
used to model the relationship between an animal’s mass and the509
period of its gait cycle, while αv , and βv are parameters mod-510
eling relationship between the length of the animal’s legs and its511
speed. The parameters αp, βp, αv , and βv are found from the mo-512
tion database using a least-squares fit. Because the animals in our513
motion database do not exhibit significant differences in the timings514
for their foot contacts once normalized for period and speed, we fix515
the relative timings for the foot contacts in all synthesized gaits to516
match those of a default walk.517
6.2.3 Numerical Solution518
In order to solve the joint inverse optimization problem defined by519
equation 15, we first note that the optimization is in a form which is520
partially decoupled. In particular, only the ‖φi−R(Ai, θ)‖22+r(θ)521
regression error term relates the different animals to each other522
(via θ), and that remaining termD(f(Ai, φi),Mi) can be indepen-523
dently evaluated for each φi. This leads to an optimization tech-524
nique in which a series of otherwise independent optimizations for525
each φi are coupled together by the regression function R(Ai, φ)526
and the regularization function r(θ). This is achieved in a manner527
reminiscent of coordinate descent by alternating between minimiz-528
ing θ and minimizing φ1, . . . , φn.529
Our approach to minimizing equation 15 involves a set of coupled530
instances of the covariance matrix adaptation (CMA) algorithm531
[Hansen et al. 1996]. We maintain a separate mean and covariance532
matrix to solve for for each φi, denoted µi and Ci respectively. At533
the beginning of each iteration a fixed number of samples φi,1, φi,m534
(we use m = 64 in our tests) is drawn for each φi distributed ac-535
cording to µi and Ci. Treating these samples as the current esti-536
mates for each φi, equation 15 is minimized for θ while holding all537
method mean error median errordefault 0.495 0.435kinematic interpolation 0.451 0.387naıve inverse interpolation 0.473 0.360joint-inverse interpolation 0.327 0.245(base inverse fit) 0.142 0.127
Table 2: The mean and median errors over the combined biped andquadruped motion databases of leave-one-out tests in which oneanimal was excluded and its gait synthesized based on the otheranimals. The errors are measured using metric given by equations16-20. Note that the cheetah and emu were excluded from thesestatistics because the ‘naıve inverse interpolation’ spacetime con-straints optimization failed to converge for them.
φi,1, φi,m fixed. As only the ‖φi − R(Ai, θ)‖22 + r(θ) term de-538
pends on θ, solving for θ reduces to a straightforward regularized539
least-squares optimization and can be solved relatively efficiently540
either with an off-the-shelf unconstrained optimizer (or with a sin-541
gle linear least squares solve for appropriate choices of R and r).542
This yields a new estimate for θ, and allows the cost associated543
with each φi,j sample to be computed as defined by equation 15,544
after which the mean µi and covariance Ci associated with eachAi545
are independently updated using the standard CMA update [Hansen546
et al. 1996] omitting any samples for which f(Ai, φi,j) fails.547
Solving a joint inverse optimization problem also requires a setting548
for the parameter β used to weight the terms in the objective func-549
tion resulting from how closely each φi matchesR(Ai, θ). Because550
lower values of β tend to converge more quickly, a continuation is551
performed where β is started out at a low value and then gradually552
increased over the course of the optimization. For quadrupeds we553
set the value of β in iteration i to βi = 0.01 + 0.02 i over a total of554
50 iterations while for bipeds we use βi = 0.001 + 0.004 i over a555
total of 35 iterations.556
7 Results557
We demonstrate our approach of animal gait synthesis using joint558
inverse optimization on a motion database of walking gaits for six559
bipeds and twelve quadrupeds as illustrated in figures 1 and 2.560
Although in principle could use both the bipeds and quadrupeds561
simultaneously, for simplicity we automatically select whether to562
synthesize a motion using only the bipeds or only the quadrupeds563
depending of whether the input animal is a biped or a quadruped.564
Processing this motion database with the joint inverse optimization565
algorithm takes several days when run on a cluster of 96 computers,566
but need only be done once. Synthesizing a gait for a new animal567
once the motion database has been preprocessed takes only a few568
minutes.569
In order to quantitatively compare our approach with other alterna-570
tives, table 2 shows the result of a leave-one-out cross-validation571
for several potential synthesis techniques, including ours:572
default The gait is synthesized using a fixed default value of φi for573
all animals.574
kinematic interpolation The gaits in the motion database are di-575
rectly interpolated weighted according to equation 22. The576
resulting motion will generally not satisfy the laws of physics,577
so a final spacetime constraints optimization is performed578
which attempts to match the kinematic interpolation as closely579
as possible while satisfying the laws of physics.580
naıve inverse interpolation The gait is synthesized using param-581
eters interpolated using equation 22, but without any joint in-582
7

Online Submission ID: 0369
verse optimization (i.e. the generative parameters for each583
animal are optimized independently).584
joint-inverse interpolation Our proposed approach in which a585
gait is synthesized using parameters interpolated using equa-586
tion 22 with joint inverse optimization.587
(base inverse fit) For comparison, each gait is synthesized using588
the optimal independently-fit inverse parameters. In contrast589
to the other approaches listed here this one does make use of590
the ground-truth motion of the animal and is thus analogous591
to the approach of [Liu et al. 2005]. Although this approach592
gives the least error, it is fundamentally incapable of synthe-593
sizing motions for new animals.594
We also illustrate a comparison of the values of the foot height and595
several DOFs for the ground truth and leave-one-out synthesized596
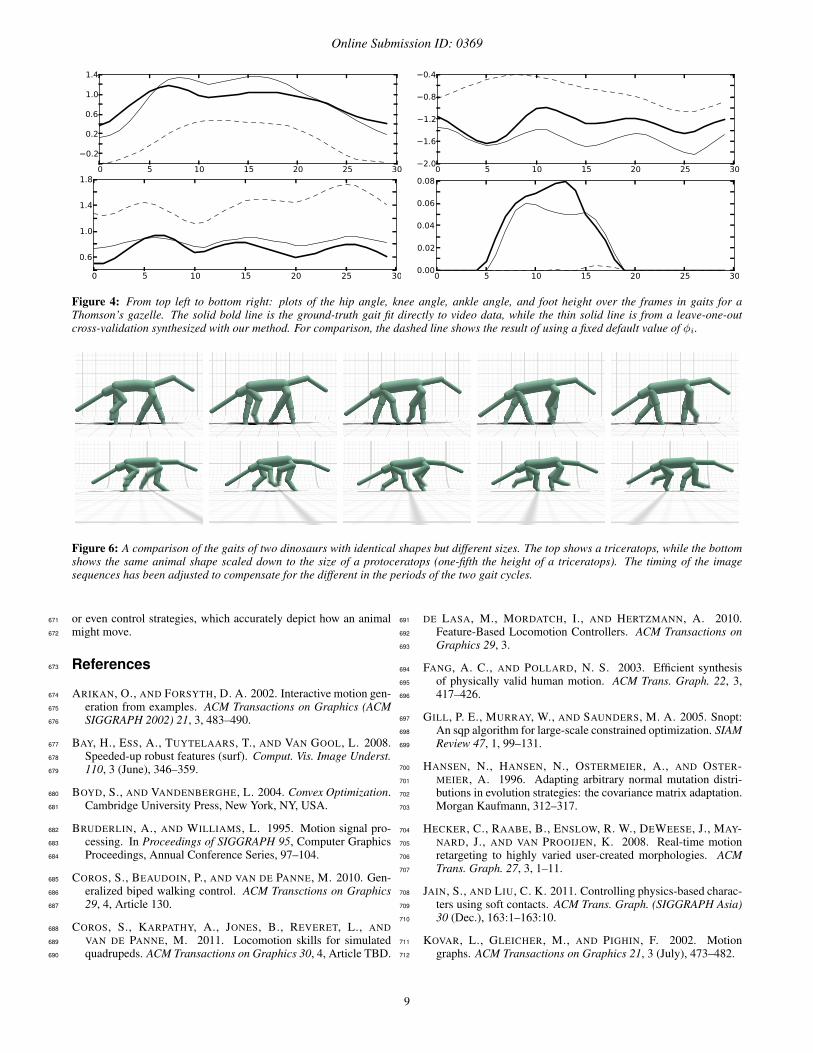
gaits for the Thomson’s gazelle in figure 4. As shown in table 2 our597
approach based on joint inverse optimization outperforms all of the598
other techniques we have tested. In addition, since our generative599
function only requires the shape of an animal and a vector of rela-600
tively general parameters, it is potentially better suited to creating601
a range of different motions for an animal as in [Liu et al. 2005]602
than an approach based on kinematic interpolation from the motion603
database.604
We have also tested our approach on synthesizing gaits for a number605
of extinct animals, including land-birds, mammals, and dinosaurs.606
Even when the shape of the animals differs significantly from any-607
thing in the motion database (or even from anything currently liv-608
ing) the gaits appear to be visually reasonable. A snapshot of these609
motions appears in figure 5. We also have found that the synthe-610
sized gaits adjust in a visually reasonable manner to the size of the611
animal. For example, figure 6 shows gaits for two dinosaurs with612
the same relative shape but different total sizes. The synthesized613
gaits are significantly different, and on a qualitative level capture614
the sort of differences which would be expected considering the615
triceratops’ substantially greater size and mass. Of course in these616
cases there is no possibility of a ‘ground truth’ motion to validate617
these synthesized gaits against. Nevertheless, the synthetic gaits618
still provide a compelling glimpse at how these creatures might619
have walked.620
There are a couple of situations where our proposed method does621
not perform as well. The first of these situations is when little data622
exists for a type of animal in the motion database. For instance, our623
motion database contains only three cats, and the the leave-one-out624
cross-validation for these motions shows an error greater than for625
the other animals in the database. The second situation in which this626
method can perform poorly is when large extrapolations are used.627
For instance the motions synthesized for a paraceratherium (four628
times the mass of an elephant) or for argentinosaurus huinculensis629
(4.5 times the mass of an elephant) exhibit some over-bending of630
the legs which is probably not realistic. Extrapolating further to631
the motion of a amphicoelias fragillimus (20 times the mass of an632
elephant) yields clearly unrealistic results. Nevertheless, when used633
on animals with a size near those in the motion database, the method634
given here performs well, even if the shape of the animal itself is635
relatively unlike anything currently living.636
8 Conclusion637
We have presented a technique for learning a model representing638
the style of walking gaits for a range of different animals, then us-639
ing this model to synthesize gaits for new animals. Our method is640
based on a novel algorithm called joint inverse optimization which641
learns coherent patterns underlying the gaits of different animals.642
This allows the synthesis of visually plausible gaits for a variety643
Figure 5: The top illustrates the gaits synthesized for a number ofextinct mammals and lan- birds, while the bottom shows the gaitssynthesized for a several dinosaurs. In the bottom image a giraffeappears in the background for size comparison.
of different extinct creatures, and has been verified to be superior to644
several other techniques at estimating the gaits of a range of animals645
for which we do have data.646
There are of course a great number of ways in which our work647
could be improved in the future. Perhaps the simplest of these is648
in the creation of a larger and more detailed motion database. Al-649
though we have achieved satisfactory results with a relatively small650
database extracted from 2D video footage, it is likely that the use651
of 3D motion capture and force plates could further improve the re-652
sults. Our approach also does not perform well when extrapolating653
gait style onto animals with a shape vastly different than anything654
we have data for. Although we expect that a more faithful biome-655
chanical model of each animal may help with this as in [Wang et al.656
2012], it remains an open question just what the most useful biome-657
chanical factors to include would be. In addition, our approach uses658
simple RBF-based interpolation to determine the style of a new an-659
imal, but it is likely that a more sophisticated regression technique660
could be better suited to large extrapolations.661
On a more theoretical level, our approach captures variations in662
style between different animals, but cannot account for the multi-663
tude of different motions often performed by even a single animal.664
This is because we treat the style of an animal’s gait as fixed by the665
shape of the animal. Instead, it would be fruitful for future tech-666
niques to be able to model the set of styles likely to be exhibited in667
the motions of an animal with a given shape. Even further in this di-668
rection, an ideal method would not be limited to just gait synthesis,669
but would be able to produce a diverse range of realistic motions,670
8
Online Submission ID: 0369
Figure 4: From top left to bottom right: plots of the hip angle, knee angle, ankle angle, and foot height over the frames in gaits for aThomson’s gazelle. The solid bold line is the ground-truth gait fit directly to video data, while the thin solid line is from a leave-one-outcross-validation synthesized with our method. For comparison, the dashed line shows the result of using a fixed default value of φi.
Figure 6: A comparison of the gaits of two dinosaurs with identical shapes but different sizes. The top shows a triceratops, while the bottomshows the same animal shape scaled down to the size of a protoceratops (one-fifth the height of a triceratops). The timing of the imagesequences has been adjusted to compensate for the different in the periods of the two gait cycles.
or even control strategies, which accurately depict how an animal671
might move.672
References673
ARIKAN, O., AND FORSYTH, D. A. 2002. Interactive motion gen-674
eration from examples. ACM Transactions on Graphics (ACM675
SIGGRAPH 2002) 21, 3, 483–490.676
BAY, H., ESS, A., TUYTELAARS, T., AND VAN GOOL, L. 2008.677
Speeded-up robust features (surf). Comput. Vis. Image Underst.678
110, 3 (June), 346–359.679
BOYD, S., AND VANDENBERGHE, L. 2004. Convex Optimization.680
Cambridge University Press, New York, NY, USA.681
BRUDERLIN, A., AND WILLIAMS, L. 1995. Motion signal pro-682
cessing. In Proceedings of SIGGRAPH 95, Computer Graphics683
Proceedings, Annual Conference Series, 97–104.684
COROS, S., BEAUDOIN, P., AND VAN DE PANNE, M. 2010. Gen-685
eralized biped walking control. ACM Transctions on Graphics686
29, 4, Article 130.687
COROS, S., KARPATHY, A., JONES, B., REVERET, L., AND688
VAN DE PANNE, M. 2011. Locomotion skills for simulated689
quadrupeds. ACM Transactions on Graphics 30, 4, Article TBD.690
DE LASA, M., MORDATCH, I., AND HERTZMANN, A. 2010.691
Feature-Based Locomotion Controllers. ACM Transactions on692
Graphics 29, 3.693
FANG, A. C., AND POLLARD, N. S. 2003. Efficient synthesis694
of physically valid human motion. ACM Trans. Graph. 22, 3,695
417–426.696
GILL, P. E., MURRAY, W., AND SAUNDERS, M. A. 2005. Snopt:697
An sqp algorithm for large-scale constrained optimization. SIAM698
Review 47, 1, 99–131.699
HANSEN, N., HANSEN, N., OSTERMEIER, A., AND OSTER-700
MEIER, A. 1996. Adapting arbitrary normal mutation distri-701
butions in evolution strategies: the covariance matrix adaptation.702
Morgan Kaufmann, 312–317.703
HECKER, C., RAABE, B., ENSLOW, R. W., DEWEESE, J., MAY-704
NARD, J., AND VAN PROOIJEN, K. 2008. Real-time motion705
retargeting to highly varied user-created morphologies. ACM706
Trans. Graph. 27, 3, 1–11.707
JAIN, S., AND LIU, C. K. 2011. Controlling physics-based charac-708
ters using soft contacts. ACM Trans. Graph. (SIGGRAPH Asia)709
30 (Dec.), 163:1–163:10.710
KOVAR, L., GLEICHER, M., AND PIGHIN, F. 2002. Motion711
graphs. ACM Transactions on Graphics 21, 3 (July), 473–482.712
9

Online Submission ID: 0369
KRY, P. G., REVERET, L., FAURE, F., AND CANI, M.-P. 2009.713
Modal locomotion: Animating virtual characters with natural vi-714
brations. Computer Graphics Forum.715
LEE, S. J., AND POPOVIC, Z. 2010. Learning behavior styles with716
inverse reinforcement learning. ACM Trans. Graph. 29, 4, 1–7.717
LEE, J., CHAI, J., REITSMA, P. S. A., HODGINS, J. K., AND718
POLLARD, N. S. 2002. Interactive control of avatars animated719
with human motion data. ACM Transactions on Graphics 21, 3720
(July), 491–500.721
LIU, C. K., HERTZMANN, A., AND POPOVIC, Z. 2005. Learning722
physics-based motion style with nonlinear inverse optimization.723
ACM Trans. Graph. 24, 3, 1071–1081.724
MORDATCH, I., DE LASA, M., AND HERTZMANN, A. 2010. Ro-725
bust physics-based locomotion using low-dimensional planning.726
ACM Transactions on Graphics 29, 4 (July), 71:1–71:8.727
MORDATCH, I., TODOROV, E., AND POPOVIC, Z. 2012. Dis-728
covery of complex behaviors through contact-invariant optimiza-729
tion. ACM Trans. Graph. 31, 4 (July), 43:1–43:8.730
NUNES, R. F., CAVALCANTE-NETO, J. B., VIDAL, C. A., KRY,731
P. G., AND ZORDAN, V. B. 2012. Using natural vibrations732
to guide control for locomotion. In Proceedings of the ACM733
SIGGRAPH Symposium on Interactive 3D Graphics and Games,734
ACM, New York, NY, USA, I3D ’12, 87–94.735
POPOVIC, Z., AND WITKIN, A. 1999. Physically based motion736
transformation. In SIGGRAPH ’99: Proceedings of the 26th737
annual conference on Computer graphics and interactive tech-738
niques, ACM Press/Addison-Wesley Publishing Co., New York,739
NY, USA, 11–20.740
RAIBERT, M. H., AND HODGINS, J. K. 1991. Animation of741
dynamic legged locomotion. SIGGRAPH Comput. Graph. 25742
(July), 349–358.743
SAFONOVA, A., HODGINS, J. K., AND POLLARD, N. S.744
2004. Synthesizing physically realistic human motion in low-745
dimensional, behavior-specific spaces. ACM Trans. Graph. 23,746
3, 514–521.747
WAMPLER, K., AND POPOVIC, Z. 2009. Optimal gait and form748
for animal locomotion. ACM Trans. Graph. 28, 3, 1–8.749
WANG, J. M., FLEET, D. J., AND HERTZMANN, A. 2010. Op-750
timizing walking controllers for uncertain inputs and environ-751
ments. ACM Trans. Graph. 29, 4 (July), 73:1–73:8.752
WANG, J. M., HAMNER, S. R., DELP, S. L., AND KOLTUN,753
V. 2012. Optimizing locomotion controllers using biologically-754
based actuators and objectives. ACM Trans. Graph. 31, 4, 25.755
WEI, X., MIN, J., AND CHAI, J. 2011. Physically valid statistical756
models for human motion generation. ACM Trans. Graph. 30757
(May), 19:1–19:10.758
WITKIN, A. P., AND POPOVIC, Z. 1995. Motion warping. In Pro-759
ceedings of SIGGRAPH 95, Computer Graphics Proceedings,760
Annual Conference Series, 105–108.761
YIN, K., LOKEN, K., AND VAN DE PANNE, M. 2007. Simbicon:762
Simple biped locomotion control. ACM Trans. Graph. 26, 3,763
Article 105.764
ZHANG, L., SNAVELY, N., CURLESS, B., AND SEITZ, S. M.765
2004. Spacetime faces: High-resolution capture for modeling766
and animation. In ACM Annual Conference on Computer Graph-767
ics, 548–558.768
10