Embed Size (px)
Citation preview

Curr Genet (1992)22:283-288 Current Genetics �9 Springer-Verlag 1992
Genetic mapping of 1,3-,fl-glucanase-encoding genes in Saccharomyces cerevisiae Jaime Correa, Carlos R. Vazquez de Aldana*, Pedro San Segundo, and Francisco del Rey
Instituto de Microbiologia-Bioquimica, Facultad de Biologia, CSIC-Universidad de Salamanca, E-37008 Salamanca, Spain
Received April 21, 1992
Summary. The map position of three 1,3-/%glucanase-en- coding genes in S. cerevisiae has been determined follow- ing conventional meiotic and mitotic mapping combined with recombinant DNA techniques. EXG1, EXG2 and SSG1 were localized to chromosomes XII, IV and XV, respectively, by hybridizing the cloned genes to Southern blots of chromosomes separated by pulsed-field gel elec- trophoresis, in conjunction with the rad52-1-dependent chromosome-loss mapping technique. Meiotic tetrad analyses further localized the EXG1 gene 6.1 centimor- gans centromere-proximal to CDC25 on the right arm of chromosome XII. EXG2 was positioned between LYS4 and GCN2 on the right arm of chromosome IV, at dis- tances of 6.2 centimorgans from LYS4 and 4.9 centimor- gans from GCN2. Finally, the SSG1 locus mapped on the right arm of chromosome XV, about 8.2 centimorgans to the centromere-proximal side of HIS3.
Key words: 1,3-fl-glucanase genes - Saccharomyces cere- visiae - Chromosomal mapping - Genetic mapping
Introduction
Yeast cells are surrounded by a complex structure in which 1,3-[3-glucan constitutes the main structural polysaccharide responsible for the shape and rigidity of the envelope. On this basis it has been suggested that the dynamics of this structure during morphogenetic events leading to a controlled hydrolysis of the cell wall, such as cell budding, expansion, conjugation and sporulation, demand the participation of 1,3-~-glucanases. These en- zymes have been detected in virtually all yeast species examined and this ubiquitous occurrence testifies to their importance at some stages during the cell life cycle.
* Present address: Laboratory of Molecular Microbiology, NIAID-NIH, Bethesda, MD 20205, USA Correspondence to." F. del Rey
To-date, the most complete study of these enzymes has been in Saccharomyces cerevisiae, revealing a complex system that encompasses several forms, some of which are present in very low amounts, differing not only in structural properties and substrate specificities but also in their fate and appearance through the cell cycle (Nombe- la et al. 1988). Thus, during vegetative growth several endo- and exo-l,3-[3-glucanases are synthesized, some of which are secreted only to remain trapped in the cell wall whereas others are released to the surrounding medium (Farkas et al. 1973; Sanchez et al. 1982; Cenamor et al. 1987). In turn, the meiotic cycle leads to the induction of a new 1,3-[3-glucanase not present in vegetatively growing cells and which is, therefore, specifically associated with the sporulation process (del Rey et al. 1979, 1980).
In recent years our work has focused on the character- ization of the system governing 1,3-[3-glucanase synthesis in yeast. Genetic approaches were initiated by the isola- tion of S. cerevisiae mutants with highly reduced exo-[3- glucanase activity by a screening method based on the capacity of these enzymes to hydrolyze synthetic deriva- tives, such as the fluorogenic substrate 4-methylumbel- liferyl-13-D-glucoside (MUG). By functional complemen- tation of the chromosomal mutation (exgl) harboured by these strains, we cloned and identified two different frag- ments that were able to restore the capacity to hydrolyze MUG (Nebreda et al. 1986). One of the fragments con- tains a structural gene (EXG1) coding for a protein whose differential glycosylation accounts for the two main ex- tracellular exo-l,3-13-glucanases (EXG I and EXG II) de- tected in culture supernatants of S. cerevisiae (Nebreda et al. 1986, 1987; Ramirez et al. 1989; Vazquez de Aldana et al. 1991). The second fragment identified produces on- ly partial complementation of the Exg- phenotype of exgl strains, and contains a gene (EXG2) that codes for a minor exo-l,3-[3-glucanase which we have designated EXG III. This enzymic form is a high molecular weight protein exhibiting a high carbohydrate content, but showing a significant degree of similarity in its protein fraction to that of the EXG I and EXG II exo-l,3-[3-glu- canases (del Rey et al. 1990). More recently, our work has

284
been directed towards the charac ter iza t ion of the gene responsible for the exo-l,3-13-glucanase produced specifi- cally dur ing the meiot ic cycle. The molecular c loning of the gene, S S G I (Sporulat ion-Specif ic Glucanase) , coding for this enzyme was achieved by screening a genomic l ibrary with a D N A probe ob ta ined by PCR-ampl i f ica- t ion using synthetic oligos designed according to the pre- dicted nucleot ide sequence of the amino - t e rmina l region of the purif ied pro te in (San Segundo et al., submit ted) .
The present work reports on the ch romosoma l assign- men t of these three genes to the S. cerevisiae genome.
Materials and methods
Strains and genetic methods. The haploid S. cerevisiae strains used in this study are listed in Table 1; crosses involving these strains are described in Tables 2 5. Standard yeast genetic media were used to grow cells, sporulate diploids and to score nutritional markers in auxotrophic strains (Sherman et al. 1982). Matings, sporulations, tetrad dissections and tetrad analyses were performed as described previously (Mortimer and Hawthorne 1969, 1975). Exo-l,3-[3-glu- canase activity was determined directly by overlaying colonies grown on YEPD-plates, with a solution containing 0.05% (w/v) 4-methylumbelliferyl-fI-D-glucoside (MUG) in 50 mM acetate buff- er (pH 5.5); after the overlay had solidified, the plates were incubat- ed at 37~ for 30min and examined under UV light where the exo-l,3-[3-glucanase-producing strains appeared fluorescent be- cause of the hydrolysis of the fluorogenic substrate. The tempera- ture sensitive (ts)-lethal cdc2, ede25, ede42 and cde64 markers were scored by replicating segregants on YEPD plates at the restrictive temperature (37 ~ In crosses involving more than one ts-lethal marker, each marker was scored by complementation; segregants were replicated onto lawns of MATa and MATs ede t~ strains on a YEPD plate and these replicas were then incubated at 37~ Link- age of markers to the RDN! locus was evaluated by scoring the segregation of the RDNI::LEU2 marker (the LEU2 gene inserted within the RDNI cluster, Petes 1980) in a leu2/leu2 background. Similarly, linkage of markers to the GCN2 locus was determined by scoring the segregation of the LEU2 gene inserted adjacent to it. Genetic map distances in centimorgans (cM) were calculated using the equation derived by Perkins (1949): Xp--100(T+6NPD)/ 2 (PD + NPD + T). Because of the underestimation of map distances over 35 eM using the Perkins equation, Xp was converted, when necessary, into the more accurate estimation given by X e (Ma and Mortimer 1983), according to the formula Xe=(80,7 Xp - 0.883 )(2)/(83.3 - X,).
Chromosome-loss mapping method. Chromosome loss in diploid strains homozygous for the rad52-1 mutation was induced by expos- ing the cells to methyl methanesulphonate (MMS) as described by Schild and Mortimer (1985). MMS plates were prepared according to the method of Prakash and Prakash (1977): MMS was added to autoclaved YEPD medium after it had cooled substantially and mixed in thoroughly before pouring. Plates were used within 2 days since MMS decomposes with time. A final concentration of 0.035 % MMS was used to score meiotic segregations for RAD + and rad52- I. To induce chromosome loss, cells grown to mid-exponential phase were directly plated on 0.005% MMS and allowed to grow into colonies on these plates.
Preparation of chromosomal DNA. Agarose plugs of chromosomal DNA were prepared using a modification of a previously described method (Schwartz and Cantor 1984). Yeast cells were grown to stationary phase in YEPD medium. The cells were harvested by eentrifugation, resuspended in 50 mM EDTA pH 8.0 (at a ratio of 6 vol EDTA solution to 4 vol of cells), and mixed with 100 p.1 (per 300 gl of cell suspension) of lyticase (2 mg/ml in 10 mM sodium
Table 1. List of strains
Strain Relevant genotype Source
YNN295 MATe, ura3, lys2, his7, trpl, ade- Y.G.S.C." YP148 MATe, ura3, lys2, his7, trpl, ade- Y.G.S.C."
(chromosome VII split at the RAD2 locus)
US80-10A MATe, ura3, leu2, inol Our collection JC2a MATe, ura3, leu2, his3 J.L. Revuelta JC2aT MATe, ura3, leu2, his3, EXG2::URA3 This study XS209-11C MATa, rad52-1, ade4, his3, leu2, R. Wickner
metlO, trpl, ura4 XS122-57D MATa, rad52-1, ura3 R. Wickner M1613 MATa, ura3, trpl, horn7, ilv5 J. Polaina US10-7A MATa, rad52-1, leu2 This study US5-7B MATe, rad52-1, trpl, ura4, exgl This study US50-13D MATe, rad52-1, ura3, his3, This study
EXG2:: URA3 MP2-14B MATa, rad52-1, ura3, ilv5, trpl This study DJMD2-7C MATe, cdc42, ura3, his4, leu2, gal2, J. Pringle
RDNI::LEU2 A5 MATa, adel, ade2, ade6, arg4, his7, A. Dominguez
trpl, ural, cde25 DMS122 MATa, leu2, his4, tyrl, exgl Our collection US21-7C MATe, leu2, his4, cdc42 This study US21-42C MATa, leu2, his4, cde42, exgl This study US23-16B MATe, ura3, his4, exgl This study US31-24B MATe, arg4, trpl, cde25 This study US32-7C MATa, arg4, ural, ura3, ede25, exgl This study US64-17C MATe, ura3, EXG2::URA3, horn2, This study
trp4, exgl, his7, arg9, adeI, ade2, ade8, pet8
US66-7C MATa, lys4, ura3 This study MLT44 MATa, ura3 Our collection US82-7B MATa, ura3, EXG2::URA3, lys4, This study
leu2, ade8, exgl US82-9C MATa, ura3, EXG2::URA3, Iys4, This study
leu2, exgl Hm753 MATe, ura3, leu2, GCN2::LEU2 M. Tamame US80-10AT MATe, ura3, SSGI::URA3, leu2, inol This study US90-8D MATa, ura3, cdc64, ade2 This study US91-30A MATa, ura3, SSG1.':URA3, ade2, ino! This study US92-5D MATe, ura3, his3, leu2 This study W3031A MATa, ura3, leu2, his3, trpl, ade2 S. Lindquist
Yeast Genetic Stock Center
phosphate containing 50% glycerol). After incubation at 37~ for 20 min, 0.3 ml of the cell enzyme suspension was mixed with 0.9 ml of warm melted agarose (1.0% low melting point agarose in 125 mM EDTA, pH 7.5), and the mixture was pipetted into mold chambers where it was allowed to solidify at 4~ for 20 min. Agarose samples were removed from the mold and placed in a Petri dish containing LET buffer (0.5 M EDTA pH 8.0, 10 mM Tris pH 7.5, 7.5% 2-mercaptoethanol). After 16-24 h at 37~ the LET buffer was replaced by an equal volume of NDS buffer (0.5 M EDTA pH 8.0, 10 mM Tris pH 7.5, 1% laurylsarcosine, 1 mg/ml proteinase K) and the samples were incubated at 50 ~ for 16-24 h. The agarose plugs were washed extensively with 50 mM EDTA pH 8.0 at room temperature and stored at 4~
Chromosome separation. Alternating field gel electrophoresis was performed with a Bio-Rad CHEF DR-II system in a 14 x 12.7 cm, 0.5 • buffer (Maniatis etal. 1982), 1% agarose gel, as de- scribed (Chu et al. 1986). Eleetrophoresis was carried out for ap- proximately 24 h in 0.5 x TBE buffer at 200 V with the switching intervals specified in the legend of Fig. 1.
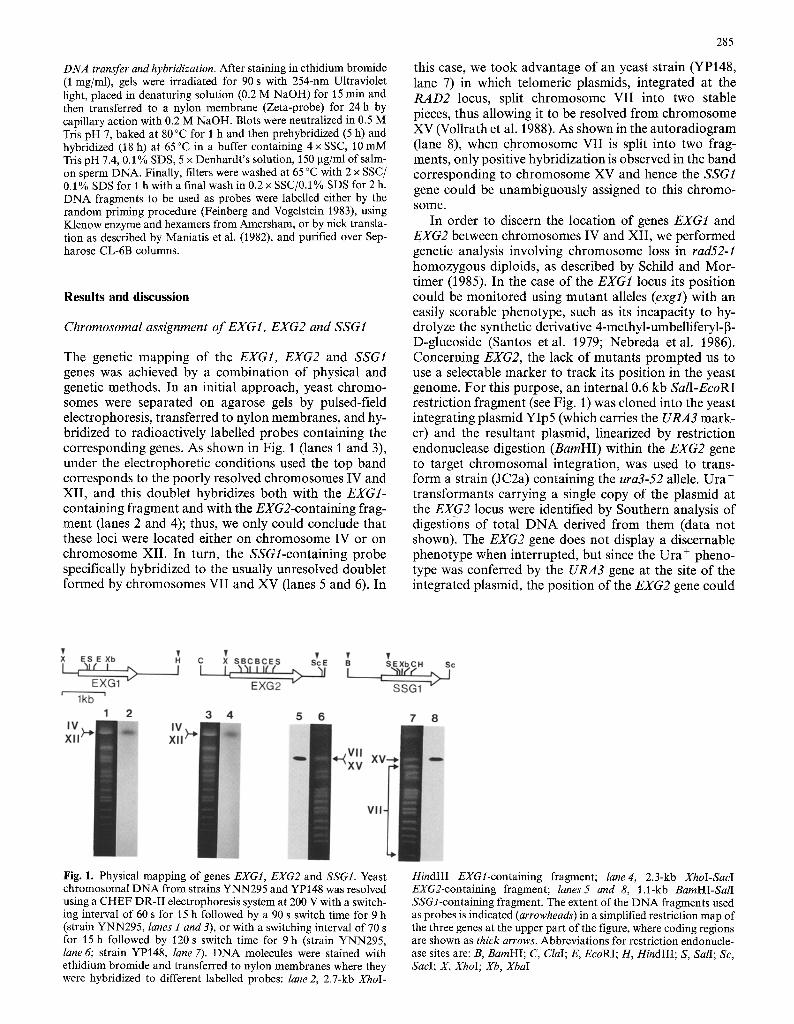
DNA transfer and hybridization. After staining in ethidium bromide (1 mg/ml), gels were irradiated for 90 s with 254-nm Ultraviolet light, placed in denaturing solution (0.2 M NaOH) for 15 rain and then transferred to a nylon membrane (Zeta-probe) for 24 h by capillary action with 0.2 M NaOH. Blots were neutralized in 0.5 M Tris pH 7, baked at 80 ~ for 1 h and then prehybridized (5 h) and hybridized (18 h) at 65~ in a buffer containing 4 • SSC, 10 mM Tris pH 7.4, 0.1% SDS, 5 x Denhardt's solution, 150 ~tg/ml of salm- on sperm DNA. Finally, filters were washed at 65 ~ with 2 x SSC/ 0.1% SDS for 1 h with a final wash in 0.2 • SSC/0.1% SDS for 2 h. DNA fragments to be used as probes were labelled either by the random priming procedure (Feinberg and Vogelstein 1983), using Klenow enzyme and hexamers from Amersham, or by nick transla- tion as described by Maniatis et al. (1982), and purified over Sep- harose CL-6B columns.
Results and discussion
Chromosomal assignment o f EXG1, EXG2 and SSG1
The genetic mapping of the EXG1, EXG2 and SSG1 genes was achieved by a combinat ion of physical and genetic methods. In an initial approach, yeast chromo- somes were separated on agarose gels by pulsed-field electrophoresis, transferred to nylon membranes, and hy- bridized to radioactively labelled probes containing the corresponding genes. As shown in Fig. 1 (lanes 1 and 3), under the electrophoretic conditions used the top band corresponds to the poorly resolved chromosomes IV and XII , and this doublet hybridizes both with the EXG1- containing fragment and with the EXG2-containing frag- ment (lanes 2 and 4); thus, we only could conclude that these loci were located either on chromosome IV or on chromosome XII. In turn, the SSGl-containing probe specifically hybridized to the usually unresolved doublet formed by chromosomes VII and XV (lanes 5 and 6). In
285
this case, we took advantage of an yeast strain (YP148, lane 7) in which telomeric plasmids, integrated at the R A D 2 locus, split chromosome VII into two stable pieces, thus allowing it to be resolved f rom chromosome XV (Vollrath et al. 1988). As shown in the autoradiogram ( lane 8), when chromosome VII is split into two frag- ments, only positive hybridization is observed in the band corresponding to chromosome XV and hence the SSG1 gene could be unambiguously assigned to this chromo- some.
In order to discern the location of genes EXG1 and EXG2 between chromosomes IV and XII , we performed genetic analysis involving chromosome loss in rad52-1 homozygous diploids, as described by Schild and Mor- timer (1985). In the case of the EXG1 locus its position could be moni tored using mutan t alleles (exgl) with an easily scorable phenotype, such as its incapacity to hy- drolyze the synthetic derivative 4-methyl-umbelliferyl-J3- D-glucoside (Santos etal . 1979; Nebreda et al. 1986). Concerning EXG2, the lack of mutants p rompted us to use a selectable marker to track its position in the yeast genome. For this purpose, an internal 0.6 kb SalI-EcoRI restriction fragment (see Fig. 1) was cloned into the yeast integrating plasmid YIp5 (which carries the URA3 mark- er) and the resultant plasmid, linearized by restriction endonuclease digestion (BamHI) within the EXG2 gene to target chromosomal integration, was used to trans- form a strain (JC2a) containing the ura3-52 allele. Ura § t ransformants carrying a single copy of the plasmid at the EXG2 locus were identified by Southern analysis of digestions of total D N A derived f rom them (data not shown). The EXG2 gene does not display a discernable phenotype when interrupted, but since the Ura + pheno- type was conferred by the URA3 gene at the site of the integrated plasmid, the position of the EXG2 gene could
Fig. 1. Physical mapping of genes EXG1, EXG2 and SSGI. Yeast chromosomal DNA from strains YNN295 and YP148 was resolved using a CHEF DR-II electrophoresis system at 200 V with a switch- ing interval of 60 s for 15 h followed by a 90 s switch time for 9 h (strain YNN295, lanes 1 and 3), or with a switching interval of 70 s for 15 h followed by 120 s switch time for 9 h (strain YNN295, lane 6; strain YP148, lane 7). DNA molecules were stained with ethidium bromide and transferred to nylon membranes where they were hybridized to different labelled probes: lane 2, 2.7-kb XhoI-
HindIII EXGl-containing fragment; lane4, 2.3-kb XhoI-SacI EXG2-containing fragment; lanes5 and 8, 1.1-kb BamHI-SalI SSGl-containing fragment. The extent of the DNA fragments used as probes is indicated (arrowheads) in a simplified restriction map of the three genes at the upper part of the figure, where coding regions are shown as thick arrows. Abbreviations for restriction endonucle- ase sites are: B, BamHI; C, ClaI; E, EcoRI; H, HindIII; S, SalI; Sc, SaeI; X, XhoI; Xb, XbaI
286
then be determined by following the segregation of the EXG2::URA3 marker on ura3/ura3 backgrounds.
A haploid rad52-1 strain was constructed that carried the exgl marker in addition to auxotrophic markers on chromosomes IV (trpl) and XII (ura4); this strain (US5- 7B) was mated to an opposite mating type rad52-1-bear- ing strain (USI0-7A) to create an homozygous rad52-1 diploid (US13) with the unmapped recessive muta t ion (exgl) in-coupling with known recessive chromosomal markers. In the same way, a haploid rad52-1 strain was constructed that carried representative auxotrophic markers on chromosomes IV (trpl) and XII (ilv5) and, in addition, was mutant at the URA3 locus on chromosome V. Crossing of this strain (MP2-14B) with an opposite mating type rad52-1 strain (US50-13D) harboring the integrated URA3 gene-containing plasmid at the EXG2 locus, resulted in a homozygous rad52-1 diploid (US55) with a ura3/ura3 background in which the Ura + pheno- type was conferred by the URA3 gene inserted at the unmapped EXG2 locus.
Chromosomal assignment by this mapping technique is based on the correlated expression of the gene in ques- tion with one of the known markers in the monosomic diploids generated by mitotic haploidization induced by X-ray or MMS treatment. The uncovering of recessive markers by chromosome loss in diploid US13, achieved by MMS treatment, demonstrated that exgl was only coordinately expressed with ura4 on chromosome XII: of the 178 colonies tested, all of them that became aux- ot roph for uracil (78) were also unable to hydrolyze M U G and, hence, of expressing EXG1 (Table 2). There- fore, we concluded that the EXG1 locus was located on chromosome XII. Regarding the EXG2 gene, the data after MMS treatment showed that whereas it was possi- ble to find auxotrophs for isoleucine-valine that were Ura + or U r a - , all the colonies requiring t ryptophan (45) were also unable to grow in the absence of uracil; the coordinate expression of the requirement for uracil (indi- cating loss of the chromosome copy containing the EXG2 locus interrupted by the URA3 gene) with the auxotrophy for t ryp tophan (trpl) clearly indicated that both loci, EXG2 and TRP1, were on the same chromosome, that is, chromosome IV.
Meiotic mapping of EXG1
Based on the above-mentioned results, EXG1 was subse- quently mapped more precisely on chromosome XII by conventional tetrad analysis. These studies included the reference loci RDN1, CDC42 and CDC25 which are lo- cated at known positions along the right arm of this chromosome. The results, summarized in Table 3, show
t h a t EXG1 is linked to CDC25: a majori ty of parental ditypes (199) and zero non-parental ditypes were found among 227 tetrads analysed f rom three different crosses. Significant linkage was also detected between EXG1 and CDC42, while EXG1 and RDN1 exhibited no detectable linkage. The genetic distances between EXG1, CDC25 and CDC42, as calculated f rom the segregation ratios, allow these loci to be arranged, relative to each other, in
Table 2. Expression of heterozygous recessive markers in diploids US13 and US55. Parent strains: USI3: US10-7A x US5-7B; US55: MP2-14B x US50-13D
Diploid Chromo- Second Total no. Markers some marker of colonies expressed"
examined + / - - / -
US13 IV trpl 178 31 33 XII ura4 178 0 78
US55 IV trpl 216 0 45 XII ilv5 216 38 12
" Diploid US13: Number of colonies wild-type for EXG1 and ex- pressing the second marker ( + / - ) , or expressing both exgl and the second marker ( - / - ) Diploid US55: Number of colonies Ura + (URA3 at the EXG2 locus) and expressing the second recessive marker ( + / - ) , or Ura (lacking URA3 at the EXG2 locus) and expressing the second reces- sive marker ( - / - )
Table 3. Meiotic linkage analyses of chromosome XII markers. Parent strains for each cross were as follows (see Table 1 for genotypes): US22: DJMD2-7C x DMS122; US31:A5 x US23-16B; US38: US31-24B x US21-42C; US39: US32-7C x US21-7C
Interval Cross Ascus type Total
PD NPD T
Map distance (cM)
RDN1-CDC42 US22 33 13 112 158 71.6" RDN1-EXG1 US22 27 20 111 158 Unlinked b CDC42-EXG1 US22 67 3 88
US38 27 2 38 US39 16 1 37
Pooled 110 6 163 279 36.8
CDC42-CDC25 US38 24 2 41 US39 15 2 37
Pooled 39 4 78 121 44.5
EXGI-CDC25 US31 91 0 15 US38 59 0 8 US39 49 0 5
Pooled 199 0 28 227 6.1
a Chi-square analysis indicates that the excess of parental ditypes over non-parental ditypes is statistically significant b Chi-square analysis indicates that the excess of parental ditypes over non-parental ditypes is not statistically significant
a map order of CDC42-EXG1-CDC25, separated at ap- proximately 36.8 and 6.1 cM, respectively (Fig. 2).
Compar ison of the size of intervals derived f rom this study with those summarized by Mort imer et al. (1989) reveals that the 44.5 cM distance estimated between CDC42 and CDC25 is in fairly good agreement with that previously proposed for this region (about 49 cM) al- though, according to our data, the distance between RDN1 and CDC42 (71.6 cM) is somewhat greater than the 58 cM estimated in previous studies (Johnson et al. 1987). As these authors stated, such variability might reflect variations in the severity of the suppression of meiotic recombinat ion that occurs in the RDN1 region (Petes 1979) or in the extent of the region affected by this phenomenon.

287
An interesting conclusion derived from the genetic mapping of EXG1 is that our results could help to clarify the discrepancy concerning the chromosomal location of CDC25, since it has been assigned to two different posi- tions, chromosome II (Portillo and Maz6n 1986) and chromosome XII (Johnson et al. 1987). We used a cdc25 t~ allele non-complementing to that employed by Portillo and Maz6n, and from our data CDC25 is clearly located on chromosome XII. Thus, it seems that it is not a prob- lem of the same name being used to describe two different loci, as stated in the 10th edition of the genetic map of S. cerevisiae (Mortimer et al. 1989), but rather, for some reason that we cannot account for, an incorrect assign- ment of CDC25 to chromosome II.
Meiotic mapping o f EXG2
To determine which arm of chromosome IV contains the EXG2 locus, we made a three-point cross with the left- arm marker cdc2, the right-arm marker hom2 and EXG2.'." URA3. This cross (data not shown) indicated that CDC2 and EXG2 are unlinked and that H O M 2 and EXG2 are linked. To confirm this chromosomal assign- ment and to identify a more precise location for the EXG2 gene, we meiotically mapped EXG2:: URA3 rela- tive to four markers (hom2, lys2, gen2 and trp4) on the right arm of chromosome IV (Table 4). Analysis of te- trads from four different diploids revealed that the inte- grated URA3 gene mapped between LYS4 and GCN2. On comparing LYS4 to the plasmid marker URA3 in 186 asci from three crosses, 163 parental ditypes and 23 te- tratypes were found, suggesting a map distance of ap- proximately 6.2 cM. The linkage between GCN2 and EXG2:." URA3 was analysed in 121 tetrads from two inde- pendent crosses which yielded 109 parental ditypes and 12 tetratypes, corresponding to a genetic distance of about 4.9 cM. It should be noted that data compiled from Mortimer et al. (1989) give a map distance of about 28 cM between L Y S 4 and GCN2, significantly greater than the 10.3 cM estimated for the same interval from our results. The only reported gene within this region is SUF24, which maps half-way between both loci, slightly closer to GCN2 (15 cM, LYS4-SUF24; 13 cM, SUF24- GCN2). Although we have not checked for genetic link- age between SUF24 and EXG2, taking into account the position of these loci relative to the flanking genes de- duced from our data and from that compiled by Mor- timer et al. (1989), it can be surmised that both loci would be tightly linked in chromosome IV (Fig. 2).
Meiotic mapping o f SSG1
As in the case of EXG2, mutants affecting the sporula- tion-specific 1,3-13-glucanase-encoding gene were not available, so that to determine its position on chromo- some XV we had to insert a selectable marker at the SSG1 locus. To do this, an internal 0.3 kb SalI-HindIII restric- tion fragment (see Fig. 1) was cloned into the yeast inte- grating vector YIp5, and the resultant plasmid was used
- - - - C D C 4 2 4'-
%
3 6 . 8
- E X G 1 -.,--
6 .1 - C D C 2 5 -.,-- - C D C 3
,v+E - - - - H O M 2
gl ( ~ 4 5 . 7
---- - L Y S 4 - -
6 . 2
S U F 2 4 : ' E X G 2 " -
o~ 4 . 9
~ - - G C N 2 ~ -
. . ~ - - A D E 2 -,,-
( , )
"q 4 4 . 2
- - - - - A D E 9 - - S S G 1 ~.-
. .k
" 8 . 2
- - - . - H I S 3 4 -
Fig. 2. A partial map of the right arms of chromosomes IV, XII and XV derived from the 10th edition of the genetic map of S. cerevisiae (Mortimer et al. 1989), illustrating the position of EXG1, EXG2 and SSG1 in relationship to other markers on the same chromo- some arm. Numbers at left indicate the approximate genetic dis- tances (cM) proposed for a given interval (delimited by arrows), and those at right express the values inferred in the present study
Table 4. Meiotic linkage analyses of chromosome IV markers. Par- ent strains for each cross were as follows (see Table 1 for genotypes): US67: US64-17C xUS66-7C; US68: US64-17CxMLT44; US83: US82-7B x Hm753; US84: US82-9C x Hm753
Interval Cross Ascus type Total
PD NPD T
Map distance (cM)
HOM2-LYS4 US67 19 2 44 65 45.7 HOM2-EXG2" US67 15 3 47
US68 15 4 25
Pooled 30 7 72 109 58.2
LYS4-EJIG2 a US67 56 0 9 US83 58 0 8 US84 49 0 6
Pooled 163 0 23 186 6.2
EXG2 "-GCN2 US83 60 0 6 US84 49 0 6
Pooled 109 0 12 121 4.9
Lu US83 53 0 13 US84 43 0 12
Pooled 96 0 25 121 10.3
HOM2-TRP4 US67 8 11 46 65 Unlinked LYS4-TRP4 US67 10 5 50 65 Unlinked b EXG2 a. TRP4 US67 10 6 49 65 Unlinked b
" Plasmid marker integrated at the EXG2 gene b Chi-square analysis indicates that the excess of parental ditypes over non-parental ditypes is not statistically significant

288
Table 5. Meiotic linkage analyses of chromosome XV markers. Parent strains for each cross were as follows (see Table 1 for genotypes): US91: US90-8DxUS80-10AT; US96: US91-30Ax US92-5D; US99:W303 1A x US80-10AT
Interval Cross Ascus type Total
PD NPD T
Map distance (cM)
ADE2-SSGI a US91 7 0 26 US96 14 2 39 US99 16 1 31
Pooled 37 3 96 136 44.2
SSG1 "-HIS3 US96 48 0 7 US99 38 0 10
Pooled 86 0 17 103 8.2
ADE2-HIS3 US96 11 5 39 US99 12 2 34
Pooled 23 7 73 103 63.8
ADE2-CDC64 US91 6 5 22 33 Unlinked SSG1 a-CDC64 US91 4 3 26 33 Unlinked
" Plasmid marker integrated at the SSG1 gene
to t r ans fo rm a s t ra in con ta in ing the ura3-52 allele (US80- 1 0A). L inea r i za t i on by res t r ic t ion endonuc lease d iges t ion (XbaI) within the SSG1 gene f r agmen t he lped to ensure tha t c h r o m o s o m a l in teg ra t ion w o u l d occur wi th in the SSG! gene and no t wi th in ura3. Genet ic tests es tab l i shed tha t the m a r k e r p l a smid (wi ld- type copy o f URA3) h a d in teg ra ted at only one pos i t ion , and D N A blots con- f i rmed tha t inser t ion occur red at the a p p r o p r i a t e SSG1 locus (da t a no t shown). C o m p a r a b l e to EXG2 gene dis- rup t ion , the SSGl-interrupted s t ra ins do no t d i sp lay a d iscernible p h e n o t y p e but , on ura3/ura3 b a c k g r o u n d s , U r a + segrega t ion cou ld be m o n i t o r e d as SSG1 segrega- t ion.
S t a n d a r d t e t rad analyses were then p e r f o r m e d us ing the SSGI:.URA3 i n t eg ran t to search for genet ic l inkage wi th k n o w n genes on c h r o m o s o m e XV (Table 5). F r o m a first cross (US91), in which the reference m a r k e r s ade2 and cdc64 were used, the segrega t ion ra t ios revealed a s ignif icant l inkage be tween SSG1 a n d ADE2 indica t ive o f a c o m m o n pos i t i on ing o f b o t h loci on the r ight a r m o f this c h r o m o s o m e . Two add i t i ona l crosses (US91 a n d US96) were then tes ted which, besides the ade2 ma rke r , inc luded the m o r e cen t romere -d i s t a l his3 marke r . The resul t ing t e t r a d s sugges ted a pu ta t ive m a p o rde r o f ADE2-SSGI-HIS3, with a close l inkage be tween SSGI and HIS3: in 103 asci ana lyzed , 86 p a r e n t a l d i types and 17 t e t ra types were found , c o r r e s p o n d i n g to a d i s tance o f a b o u t 8.2 c M (Fig. 2). D a t a f rom M o r t i m e r et al. (1989) ind ica te a m a p d is tance be tween HIS3 and the m o r e cen- t r o m e r e - p r o x i m a l ADE9 locus o f a p p r o x i m a t e l y 11 cM. Since the overa l l length f rom ADE2 to HIS3, es t ima ted f rom the sum o f the ADE2/SSGI and SSGI/HIS3 seg- ments , is fa i r ly cons is ten t wi th tha t r epo r t ed for the sum o f the ADE2/ADE9 and ADE9/HIS3 in tervals , it can be inferred tha t SSG1 is loca ted be tween ADE9 and HIS3, p r e s u m a b l y more closely l inked to the former .
Acknowledgements. We are grateful to J.R. Pringle, R.B. Wickner, A.G. Hinnebusch, A. Dominguez, M. Tamame, J.L. Revuelta, Y. Sanchez and J. Polaina for providing yeast strains, C. Belinch6n for photographic work, and N. Skinner for correcting the English manuscript. This research was supported by grants from the Comi- si6n Interministerial de Ciencia y Tecnologia (BIO89-O261) and from the European Economic Community (BIOT-CT90-0165). J. Correa, C.R. Vazquez de Aldana and P. San Segundo were recipi- ents of fellowships from Ministerio de Educaei6n y Ciencia.
References
Cenamor R, Molina M, Galdona J, Sanchez M, Nombela C (1987) J Gen Microbiol 133:619-628
Chu G, Vollrath D, Davis RW (1986) Science 234:1582-1585 Farkas VP, Biely P, Bauer S (1973) Biochim Biophys Acta 321:246-
255 Feinberg AP, Vogelstein B (1983) Anal Biochem 132:6-13 Johnson DI, Jacobs CW, Pringle JR, Robinson LC, Carle GF, Olson
MV (1987) Yeast 3:243-253 Ma C, Mortimer RK (1983) Mol Cell Biol 3:1886-1887 Maniatis T, Fritsch EF, Sambrook J (1982) Molecular cloning: a
laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York
Mortimer RK, Hawthorne DC (1969) Yeast Genetics. In: Rose AH, Harrison JS (eds) The yeasts I. Academic Press, New York, pp 385-460
Mortimer RK, Hawthorne DC (1975) Genetic mapping in yeast. In: Prescott DM (ed) Methods in cell biology, vol. 11. Academic Press, New York, pp 221-233
Mortimer RK, Schild D, Contopoulou CR, Karts JA (1989) Yeast 5:321-403
Nebreda AR, Vazquez de Aldana CR, Villa TG, Villanueva JR, Rey F del (1987) FEBS Lett 220:27-30
Nebreda AR, Villa TG, Villanueva JR, Rey F del (1986) Gene 47:245-259
Nombela C, Molina M, Cenamor R, Sanchez M (1988) Microbiol Sci 5:328-332
Perkins D (1949) Genetics 38:187-197 Petes T (1979) J Bacteriot 138:185-192 Petes Y (1980) Cell 19:765-774 Portillo F, Maz6n MJ (1986) J Bacteriol 168:1254-1257 Prakash L, Prakash S (1977) Genetics 86:33-55 Ramirez M, Hernandez LM, Larriba G (1989) Arch Microbiol
151:391-398 Rey F de1, Santos T, Garcia-Acha I, Nombela C (1979) J Bacteriol
139:924-931 Rey F del, Santos T, Garcia-Acha I, Nombela C (1980) J Bacteriol
143:621-627 Rey F del, Vazquez de Aldana CR, Correa J, San Segundo P (1990)
Yeast 6, S 504 Sanchez A, Villanueva JR, Villa TG (1982) FEBS Lett 138:209-212 Santos T, Rey F del, Conde J, Villanueva JR, Nombela C (1979) J
Bacteriol 139:333-338 Schild D, Mortimer RK (1985) Genetics 110: 569- 589 Schwartz DC, Cantor CR (1984) Cell 37:67-75 Sherman F, Fink GR, Hicks JB (1982) Methods in yeast genetics.
Laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York
Vazquez de Aldana CR, Correa J, San Segundo P, Bueno A, Ne- breda AR, Mendez E, Rey F del (1991) Gene 97:173-182
Vollrath D, Davis RW, Connelly C, Hieter P (1988) Proc Natl Acad Sci USA 85:6027-6031
C o m m u n i c a t e d by A. H innen





























