Embed Size (px)
Citation preview

252 GENETICS: MUKHERJEE AND SINHA PROC. N. A. S.
4Hemmingsen, E., private communication.6 Roughton, F. J. W., Progr. Biophys. Biophys. Chem., 9, 55 (1959).6 Unpublished data.7Collins, R. E., Science, 133, 1593 (1961).8 Wang, J. H., J. Theoret. Biol., 4, 175 (1963).9 La Force, R. C., and I. Fatt, Trans. Faraday Soc., 58, 1451 (1962).
SINGLE-ACTIVE-X HYPOTHESIS: CYTOLOGICAL EVIDENCE FORRANDOM INACTIVATION OF X-CHROMOSOMIES IN A
FEMALE M1ULE COMPLEMENT*
BY BARID B. M\IUKHERJEE AND ANIL K. SINHA
DEPARTMENT OF GENETICS, MCGILL UNIVERSITY, MONTREAL, CANADA
Communicated by Bentley Glass, December 24, 1963
According to the dosage compensation hypothesis, postulated by Lyon," 2 oneof the two X-chromosomes in a normal mammalian female complement becomesgenetically inactivated at an early stage of embryonic development, and this in-active X, which is usually condensed or heteropycnotic at prophase, could be eitherpaternal or maternal in origin in different cells of the same individual. This hy-pothesis is supported by evidence from a biochemical study of glucose-6-phosphatedehydrogenase activity in the human female' and has been confirmed by a recentstudy of Davidson, Nitowsky, and Childs.4 In an attempt to test this hypothesiscytologically, Ohno and Cattanach5 studied the prophase skin cells of a stock ofmice whose wild-type alleles for the coat color genes of linkage group I had beentranslocated to the X. The male mice with XtXnY and females with XtXn chromo-somes had a variegated coat color with light and dark patches, and a distinct hetero-pycnotic element was observed in their prophase skin cells. These investigatorsinterpreted the heteropycnotic elements as inactive X-chromosomes and claimed thatthey could distinguish the heteropycnotic chromosome from the albino patches asthe translocated-X (Xt) and its counterpart in the wild patches as normal-X (Xn)by comparing their total lengths. They thus concluded that random inactivationof X-chromosomes does occur in mammalian females as postulated by Lyon." 2But considering the extent of condensation and other morphologic changes thatchromosomes undergo during early to late prophase, one could not be absolutelysure of distinguishing a heteropycnotic X-chromosome from a heteropycnoticXt in prophase figures, particularly since the only criterion of identification is basedmerely on the comparative lengths of these two chromosomes, when X' was de-rived from one prophase figure and Xt from another.
Autoradiographic studies on chromosome duplications have revealed that incultured cells from normal mammalian females, or in diploid polysomic-X cells,only one X completes its duplication along with the autosomes, while replicationcontinues in the remaining X-chromosome or chromosomes after it is completein the rest of the complement. Although the exact morphologic identification ofthe late-replicating chromosome in the human complement is difficult, autoradio-
Dow
nloa
ded
by g
uest
on
Aug
ust 2
8, 2
021

VOL. 51, 1964 GENETICS: MUKHERJEE AND SINHA 253
graphic studies on chromosome duplication in cultured leucocytes from a domesticcow confirm unequivocally that the chromosome which replicates its DNA out ofphase with the rest of the complement is indeed an X-chromosome. 10 Consideringall the available information, it now appears that the late-replicating X-chromosomeis in fact the one which becomes genetically inactivated during embryogenesis.If this is so, the hypothesis of random inactivation of the maternal or paternalX-chromosome could be tested in a mammalian species whose two X-chromosomesare morphologically distinguishable in a female complement. We report here ourpreliminary observations on chromosome duplication in cultured leucocytes from afemale mule, which is unique in that its paternal and maternal X-chromosomes canbe clearly distinguished morphologically.
Materials and Methods.-Leucocytes from peripheral blood were cultured by thestandard technique" with minor modifications. The autoradiographic techniqueused in this study was similar to the one previously used.'2 After 72-85 hr of in-cubation, thymidine-H3 (specific activity 0.36 c/mM, Schwarz BioResearch Inc.,Orangeburg, New York) was added to each culture in a final concentration of 1/Ic/ml. Colchicine was added one hr later. Five to six hr after the addition ofthymidine-H3 the cells were harvested directly from the medium in which isotopewas still present. The cells were then washed, treated with hypotonic solution,fixed, and air-dried on slides. Well-spread metaphase figures were photographed,and their locations on the slides were recorded. Kodak NTB3 emulsion was thenapplied to each slide and exposed for four days before being developed. Eachmapped metaphase figure was rephotographed at the focal plane of the overlyingsilver grains. The photographs of each metaphase figure taken before applicationof the film were used to prepare a karyotype which served as a guide in karyotypingthe autoradiogram of the same metaphase figure. Since thymidine-H3 remainedavailable from its addition until termination of each culture, any unlabeled chromo-some or chromosome segments must have completed DNA synthesis before thymi-dine-H3 was added to the medium, and any labeled chromosome must have com-pleted DNA replication after the addition of the isotope.
Results and Discussion.-The mule is a hybrid between a female horse (Equuscaballus) and a male donkey (Equus asinus). Although all male and most femalemules are sterile, females are occasionally fertile. Except for sterility, mules ofboth sexes are perfectly normal and healthy. The somatic chromosome numberof this species is 63, which is the exact total of two haploid sets, n = 32 chromosomesfrom E. caballus and n = 31 from E. asinus. In the mule karyotype, the paternaland maternal chromosomes can be rather clearly distinguished. In the female,the haploid set from E. caballus consists of 14 metacentric or submetacentric chro-mosomes and 18 that are acrocentric. The X-chromosome is the second biggestin this set, with a submedian centromere. The haploid set from E. asinus consistsof 20 chromosomes with median, submedian, or subterminal centromeres, and 11that are acrocentric. The X-chromosome is the fourth largest chromosome of thiscomplement and has a subterminal centromere. The two X's of a female mulecomplement can therefore be easily distinguished morphologically.'3, 14 Thelargest autosome of the E. asinus complement has a subterminal centromere, and in90 per cent of the cells that we examined this chromosome showed a marked con-densed region in the upper part of its long arm, just below the centromere.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
8, 2
021

254 GENETICS: MUKHERJEE AND SINHA PROC. N. A. S.
Grain density among metaphase figures varied greatly, from completely unlabeledcells to very heavily labeled ones. In many labeled metaphase figures a varyingnumber of chromosomes had no overlying silver grains and were presumed to haveterminated DNA replication before isotope became available to the culture. Itwas thus possible to determine the presumptive order in which specific chromosomesin the complement terminated DNA synthesis. Out of 149 metaphase figures thatwere first photographed for this study, only 42 cells were labeled in a way that wasinformative. In 33 of these 42 metaphase figures one chromosome was found tobe comparatively heavily labeled in comparison with the rest of the chromosomesin the complement. In 17 of these 33 figures the late-replicating chromosomethadthe size and morphology of the X-chromosome of E. asinus (Fig. 1), and in the re-
,*"'x'S g. * q
m~~~~~
V~~~~~~~~~~~~~
someof th :eE Xcaas(i.2.In temjryofthemeapas figre th ealy
replicatnXse r of i trm~~~~~~~~~~~~.A A--041 3°-;it . A.S.
replicti eachrlethnhe fromat (Figs. 3 an 4), bat both typestof the harn w her
xisit̂4 A. ': i: chr omoomin^ th com lem nt
s.o.e ;:.f th j;. :aalu (i. 2) In the maort of the mea s fiue the.early' I'
repIca1.Ting X-chromosomeseithromE. asinusorofEasbllsterminateddulctoDltrtanAte
replication earlier than the late-X (Figs. 3 and 4), but both types of the early-X's werestill continuing their replication when approximately half of the chromosomes of theentire complement had completed duplication. In many of the metaphase figures, theshort arm and the part of the long arm close to the centromere of the X-chromosomeof E. asinus appeared to terminate replication earlier than the telomeric region ofthe long arm, but no such- pattern of DNA replication was seen with the X-chromo-some of E. caballus.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
8, 2
021

VOL. 51, 1964 GENETICS: MUKHERJEE AND SINHA 255~~~~~~~~~~~* ,%@*
Ire F .
*~~~~~~~~~~~~~ ~~~~~~ So'as
::.~~~~~~ ~~~X A..
9 * #
* L4 o
IR *'^," 00%1 E | * X 5 *
* . *0 * ,.* 0X ..
IG. 2 h -o o f E.s atle
A~~ ~ ~ ~ ~ ~ ~
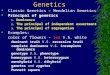
FIG. 2.-The X-chromosome from E. caballus is seen as the late replicator.
Six of the 42 metaphase figures did not show a particular late-replicating chro-mosome, but several chromosomes, including one or both X-chromosomes, showedapproximately equal numbers of silver grains on them. For example, in a partic-ular metaphase figure which must have come in contact with the isotope very latein the DNA synthetic period, only 13 chromosomes were labeled; others werecompletely free of grains. Of these 13 only 2 chromosomes, including the X-chro-mosome of E. caballus, contained three grains, whereas the remaining 11 chromo-somes showed only one or two grains. The X-chromosome from E. asinus in thiscomplement was completely free of any silver grains.The remaining 3 of the 42 metaphase figures did not conform to any of the afore-
said patterns of chromosome duplication. In one of these 3 metaphase figures inwhich all the chromosomes were labeled, the condensed portion of the largest auto-some of the E. asinus complement and a small acrocentric chromosome were com-paratively heavily labeled. In another figure, the long arm of one of the threelargest submetacentric autosomes of the mule complement was the most heavilylabeled chromosome segment; the last figure showed both X-chromosomes as com-paratively late-replicators.
In general, one medium-sized acrocentric and one of the three largest submeta-centric autosomes of the mule complement completed duplication only slightlyearlier than the late-replicating X, and this was preceded by another medium-sized submetacentric chromosome. Two of the smallest acrocentric chromosomes
Dow
nloa
ded
by g
uest
on
Aug
ust 2
8, 2
021

256 GENETICS: MUKHERJEE AND SINHA PROC. N. A. S.
g 4 A g a 4F:: fiAk V:
~~~~S*5
A:m.
*~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.'.~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~'.
FIG. 3.-The comparative grain densities of two X-chromosomes of a female mule complementfrom 18 different metaphase figures. In nine of them on the left, the X-chromosomes from E.asinus are most heavily labeled, whereas in the nine on the right, the X-chromosomes from E.caballus are the late-replicators.
were the first to complete duplication, followed by two or three additional medianor small acrocentric autosomes and one or two smallest metacentric chromosomes.Although some of the smaller chromosomes were the first to complete duplication,there seemed to be no correlation between the size of a particular chromosome and itstime of termination of DNA synthesis. Many of the larger chromosomes in thiscomplement completed duplication earlier than some of the smaller chromosomes.
It is interesting to note also that in most of the metaphase figures that came incontact with the isotope comparatively early in their DNA synthetic period, thecondensed portion of the long arm of the largest autosome of the E. asinus comple-ment was the second most heavily labeled chromosome segment after the late-X,but in metaphase figures that came in contact with thymidine-H3 very late in theirDNA synthetic period this chromosome consistently showed fewer grains than thelate-replicating X-chromosome and the chromosomes which completed duplicationslightly before the late-X. Furthermore, in a number of metaphase figures, whichwere also very lightly labeled, the short arm of this autosome and the tip of the longarm away from the centromere showed one or more grains, but the condensed re-gion which was very heavily labeled during earlier stages was completely free of anysilver grains. Thus, this appears to be a very clear example of the variation in therate of DNA synthesis among different chromosomes of the complement or evenamong different segments of the same chromosome.
This autoradiographic study on chromosome duplication in cultured leucocytesfrom a female mule unquestionably demonstrates that either the paternal or thematernal X-chromosome becomes the late-replicator in different cells of the sameanimal, and if the late-replicating X-chromosome is the one which becomes genet-ically inactivated during the early stages of embryonic development, then this
Dow
nloa
ded
by g
uest
on
Aug
ust 2
8, 2
021

VOL. 51, 1964 GENETICS: MUKHERJEE AND SINHA 257
FIG. 4.--One distinct sex-chromatin mass observed in interphase nuclei from vaginal mucosacells of the mule, stained by the Feulgen reaction.
study provides one of the clearest cytological pieces of evidence in support of therandom inactivation hypothesis postulated by Lyon."1 2
The question of whether or not the entire portion -of the late-replicating-X in amammalian female complement becomes inactivated is difficult to answer. Therandom inactivation of one of the X-chromosomes in each cell along its entire lengthdoes not explain either the variation in the amount of variegation Russell'" observedin mice with different rearrangements of the same two chromosomes (X-chromo-some and the linkage group I), or the absence of two red cell populations, Xg(a+)and Xg(a-), in women heterozygous for the X-linked Xga blood group demon-strated by Reed et al.'6 Observations on the human sex-chromosome anomaliesalso suggest that inactivation does not involve the entire X.
Autoradiographic studies on chromosome duplication in cultured cells fromseveral mammalian species, including the mule, show that it is not the entire por-tion of the late-replicating X-chromosome which terminates duplication later thanthe rest of the complement; some segments of it complete replication along withsome of the other chromosomes. In the female Chinese hamster, for example, theshort arm of one of the X-chromosomes replicates early, while the long arm and theentire length of the other X replicate late.'7 In the human female, the short arm
Dow
nloa
ded
by g
uest
on
Aug
ust 2
8, 2
021

258 GENETICS: MUKHERJEE AND SINHA PROC. N. A. S.
of the late-X appears to complete duplication earlier than the long arm, whereasin cultured leucocytes from the domestic cow the short arm of the late-X completesduplication later than the long arm.'0 In the mule, the telomeric region of thelong arm of the X-chromosome of E. asinus terminates duplication later than itsother parts. Whether or not the portion of the late-X which completes duplica-tion earlier, with other chromosomes of the complement, represents the activesegment of the inactive-X is difficult to decide, but considering the facts which havebeen mentioned here and Russell's15 elegant demonstration that the amount ofvariegation in X-autosome translocated mice depends on the point of rearrange-ment of the translocated-X, it is reasonable to suppose that inactivity does notinvolve the entire X-chromosome.Although the majority of the 42 suitably labeled cells that were studied showed
either the paternal or the maternal X-chromosomes as late-replicators, a number ofcells did not conform to this general pattern. Considering the degree of dis-similarity between the parental chromosomes, different duplication patterns insome cells might be expected. On the other hand, the different duplication patternobserved in these cells might be caused simply by the technical factors involvedin this study.Very little is known about the genetic mechanism which controls the inactivation
of X-chromosomes in a mammalian female complement, but evidence from studiesof sex-chromatin in diploid and polyploid human cells18 and heteropycnotic pre-sumptive X-chromosomes from tetraploid prophase nuclei from female rat livercells19 supports the concept that a diploid set of autosomes suppresses the activityof all but one X-chromosome in each diploid complement.20 In the light of thisconcept, what has been observed in the female mule complement is rather inter-esting. In the mule the morphologic dissimilarities between parental chromosomesare considerable, and in histological studies of mule testes, Makinol4 observed thatdegeneration of germ cells took place during the early stages of meiotic prophase,so that the meiotic division was entirely absent. Such divisions are a very rarephenomenon in female germ cells also, and this is probably due to the fact that noneof the pairs of chromosomes in this complement are truly homologous. The chro-mosome complement of the somatic cells of the mule is composed of two mor-phologically dissimilar haploid sets rather than a true diploid one. But in spite ofthis, only one X-chromosome, either of paternal or maternal origin, becomes thelate-replicator in the majority of the cells that were studied. This observationsuggests that probably the genomes of E. asinus and E. caballus are not as dis-similar as the morphological appearances of their chromosomes suggest.Although this study very clearly demonstrates the random inactivation of either
the maternal or the paternal X-chromosome in different cells of a mammalian fe-male, in one presumptive XO/X iso-X mosaic21 and another X iso-X22 humanfemale, the iso-X was always found to be the late-replicator. This may indicatethat the abnormal X always becomes inactivated in preference to the structurallynormal X-chromosome, but on the other hand the cells with one active X deficientfor the short arm and the other inactivated normal X may become eliminated dur-ing early embryonic life, as suggested by Gartler and Sparkes23 and Miller et al.2'Summary.-Patterns of chromosome duplication in cultured leucocytes from a
female mule have been studied by the method of autoradiography. Cultured
Dow
nloa
ded
by g
uest
on
Aug
ust 2
8, 2
021

NIOL. 51, 1964 GENETICS: MUKHERJEE AND SINHA 259
leucocytes were exposed to thymidine-H3 continuously for 5-6 hr and were harvesteddirectly from the isotope-containing medium. Out of 42 metaphase figures thatwere labeled in a way that was informative, 33 showed one chromosome that washeavily labeled in comparison with the rest of the chromosomes in the complement.In 17 of these 33 figures the heavily labeled chromosome was identified as theX-chromosome from E. asinus, and in the remaining 16 this chromosome had thesize and morphology of the X-chromosome of E. caballus. Six other metaphasefigures did not show a particular late-replicating chromosome, and the remaining3 did not conform to any of the aforesaid patterns. If the late-replicating X-chro-mosome in the mammalian female complement is the one which becomes geneticallyinactivated during early embryogenesis, then this study provides one of the clearestpieces of cytological evidence in support of the random inactivation of the X-chro-mosome hypothesis postulated by Lyon.
The authors wish to thank Dr. F. Clarke Fraser for his helpful criticism of the manuscript, andDr. C. B. Baker for his kind cooperation in collecting mule blood. Technical assistance of Mr.Nikolaos Plevritis is gratefully acknowledged.
* This work has been supported by grant MA-1026 from the Medical Research Council ofCanada.
1 Lyon, M. E., Nature, 190, 372 (1961).2Lyon, M. E., Am. J. Hum. Genet., 14, 135 (1962).Beutler, E., M. Yeh, and V. F. Fairbanks, these PROCEEDINGS, 48, 9 (1962).
4 Davidson, R. G., H. M. Nitowsky, and C. Childs, these PROCEEDINGS, 50, 481 (1963).5 Ohno, S., and B. M. Cattanach, Cytogenetics, 1, 129 (1962).6 Morishima, A., M. M. Grumbach, and J. H. Taylor, these PROCEEDINGS, 48, 756 (1962).7 German, J. L., Trans. N. Y. Acad. Sci., 24, 395 (1962).8 Rowley, J., S. Muldal, C. W. Gilbert, L. G. Lajtha, J. Lindsten, M. Fraccaro, and K. Kaijser,
Nature, 197, 251 (1963).9 Mukherjee, B. B., 0. J. Miller, W. R. Breg, and S. Bader, Exptl. Cell Res., in press.10 Mukherjee, B. B., and A. K. Sinha, Can. J. Genet. Cytol., 5, 490 (1963).11 Moorhead, P. S., P. C. Nowell, W. J. Mellman, D. M. Batipps, and D. A. Hungerford, Exptl.
Cell Res., 20, 613 (1960).12Bader, S., 0. J. Miller, and B. B. Mukherjee, Exptl. Cell Res., 31, 100 (1963).13Trujillo, J. M., C. Stenins, L. C. Christian, and S. Ohno, Chromosoma, 13, 243 (1962).14 Makino, S., T. Sofuni, and M. S. Sasaki, Proc. Japan Acad., 39, 176 (1963).16 Russell, L. B., Science, 140, 976 (1963).16 Reed, T. E., N. E. Simpson, and B. Chown, Lancet, II, 467 (1963).17 Taylor, J. H., J. Biophys. Biochem. Cytol., 7, 455 (1960).18 Klinger, H. P., and N. A. Schwarzacher, J. Biophys. Biochem. Cytol., 8, 345 (1961).19 Ohno, S., W. D. Kaplan, and R. Kinosita, Exptl. Cell Res., 18, 415 (1959).2 Editorial, Lancet, II, 191 (1960).21 Miller, 0. J., B. B. Mukherjee, S. Bader, and A. C. Christakos, Nature, 200, 918 (1963).21 Muldal, S., C. W. Gilbert, L. G. Lajtha, J. Lindsten, J. Rowley, and M. Fraccaro, Lancet, I,
861 (1963).21 Gartler, S. M., and R. S. Sparkes, Lancet, II, 411 (1963).
Dow
nloa
ded
by g
uest
on
Aug
ust 2
8, 2
021