Embed Size (px)
Citation preview

2012. október 3.
Mészáros Klára
Genetikai térképezés,
gén azonosítás
Mészáros Klára, Karsai Ildikó

Növénynemesítés feladatai
2012. október 3.
Mészáros Klára
Növénytermesztés hatékonyságának és a termésbiztonság növelése
speciális termesztési rendszerek biztosítása (herbicid tolerancia)
biotikus stressz tolerancia növelése
környezeti adaptáció és abiotikus stressz tolerancia javítása. Víz (WUE) és nitrogén
hasznosítás (NUE) javítása, fagyállóság és szárazságtűrés javítása
Funkcionális élelmiszer alapanyag előállítására alkalmas növényfajta
Bioenergetikai célra alkalmas növények nemesítése
Technológiai rendszerekre adaptált és/vagy nemesített fajták (gyógyszer alapanyag, oltóanyag)
sejt fermentorokban
szántóföldi növénytermesztésben

2012. október 3.
Mészáros Klára
diagnosztikai markerek, módszerek kidolgozása
természetes variabilitás tesztelése vad és termesztett fajtákon belül, valamint a rokon
fajok csoportjában
hasznos gének, gén allélek beépítése a nemesítési alapanyagokba
marker szelekcióval egybekötött hagyományos keresztezések során
gén transzformációs technikák alkalmazásával
Gén azonosítás
Adott tulajdonság fenotípusos megnyilvánulásában meghatározó szerepet játszó genetikai
komponensek, szabályozó mechanizmusok, biokémiai folyamatok megértése.

Minőségi és mennyiségi tulajdonságok
Azok a tulajdonságokat amelyek mértékegységgel
nem, vagy csak nehezen mérhetők, kialakulásuk
kevéssé függ a környezet hatásától, minőségi
tulajdonságoknak nevezzük. Megnyilvánulásukat
egy vagy néhány gén határozza meg. Pl: betegség
ellenállóság
A mértékegységgel mérhető tulajdonságok,
amelyek kialakulásában a környezet hatása
jelentős szerepet játszik: mennyiségi
tulajdonságok. Kifejeződésük általában sok gén
kölcsönhatásának eredménye pl: termőképesség
2012. október 3.
Mészáros Klára

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térképek
Két szülős térképező populációk
Széles genetikai bázist képviselő fajtakör
Jelölt gén megközelítése
Genom pozíció függő stratégiák: pozicionális klónozás, deléciós vonalak
Összehasonlító genomikai stratégiák: modell növények
Mesterséges genetikai variáció (indukált mutációs populációk)
Gén csapdázás: T-DNS, transzpozon
Gén expressziós mintázatok elemzése:
Differential Display
DNS microarray, DNS chip
Gén azonosítás főbb módszerei

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térképek
Két szülős térképező populációk
Széles genetikai bázist képviselő fajtakör
Jelölt gén megközelítése
Genom pozíció függő stratégiák: pozicionális klónozás, deléciós vonalak
Összehasonlító genomikai stratégiák: modell növények
Mesterséges genetikai variáció (indukált mutációs populációk)
Gén csapdázás: T-DNS, transzpozon
Gén expressziós mintázatok elemzése:
Differential Display
DNS microarray, DNS chip
Gén azonosítás főbb módszerei

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térképek
Térképező populáció
Molekuláris marker technikák
Számítógépes térképező programok

Térképező populáció
Két homozigóta szülő keresztezéséből származó utódok
a térképező populáció, amelynek minden egyedéről
meghatározható, hogy a szülőktől milyen kombinációt
örökölt két vizsgált allélpárra, P vagy R típust.
Ennek ismeretében a rekombináció gyakorisága
meghatározható akár közvetlenül (r=R/P+R), akár
LOD számítással.
Ezt az r (rekombinaciós gyakoriság) értéket genetikai
térképezésre használhatjuk.
2012. október 3.
Mészáros Klára

2012. október 3.
Mészáros Klára
Térképező populáció
Keresztezési partnerek kiválasztása: távoli vagy speciális
Populáció típusa:
hasadó F2 populáció
homozigóta populációk (DH, RIL, SSD)
egyéb speciális populációk (NIL, egy kromoszómára rekombináns stb.)
Populáció mérete
Marker kapcsoltsági térképek
DNS izolálás markeres vizsgálatok
Genom szintű különbségek

Öntermékenyülő növények térképezési populációinak
főbb típusai
2012. október 3.
Mészáros Klára

2012. október 3.
Mészáros Klára 2012. október 3. Mészáros Klára
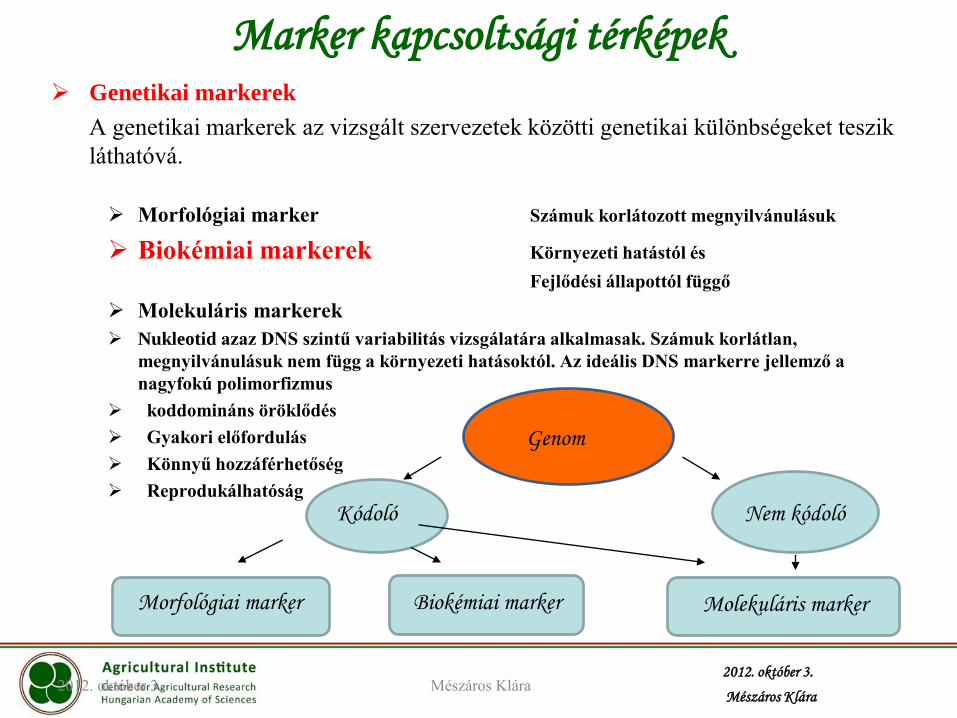
Genetikai markerek
A genetikai markerek az vizsgált szervezetek közötti genetikai különbségeket teszik
láthatóvá.
Morfológiai marker Számuk korlátozott megnyilvánulásuk
Biokémiai markerek Környezeti hatástól és
Fejlődési állapottól függő
Molekuláris markerek
Nukleotid azaz DNS szintű variabilitás vizsgálatára alkalmasak. Számuk korlátlan,
megnyilvánulásuk nem függ a környezeti hatásoktól. Az ideális DNS markerre jellemző a
nagyfokú polimorfizmus
koddomináns öröklődés
Gyakori előfordulás
Könnyű hozzáférhetőség
Reprodukálhatóság
Marker kapcsoltsági térképek
Genom
Kódoló Nem kódoló
Morfológiai marker Biokémiai marker Molekuláris marker

2012. október 3.
Mészáros Klára
Oregon Wolf Barley Morfológiai markerek
2012. október 3.
Mészáros Klára 2012. október 3. Mészáros Klára
Genetikai markerek
A genetikai markerek az vizsgált szervezetek közötti genetikai különbségeket teszik
láthatóvá.
Morfológiai marker Számuk korlátozott megnyilvánulásuk
Biokémiai markerek Környezeti hatástól és
Fejlődési állapottól függő
Molekuláris markerek
Nukleotid azaz DNS szintű variabilitás vizsgálatára alkalmasak. Számuk korlátlan,
megnyilvánulásuk nem függ a környezeti hatásoktól. Az ideális DNS markerre jellemző a
nagyfokú polimorfizmus
koddomináns öröklődés
Gyakori előfordulás
Könnyű hozzáférhetőség
Reprodukálhatóság
Marker kapcsoltsági térképek
Genom
Kódoló Nem kódoló
Morfológiai marker Biokémiai marker Molekuláris marker
2012. október 3.
Mészáros Klára
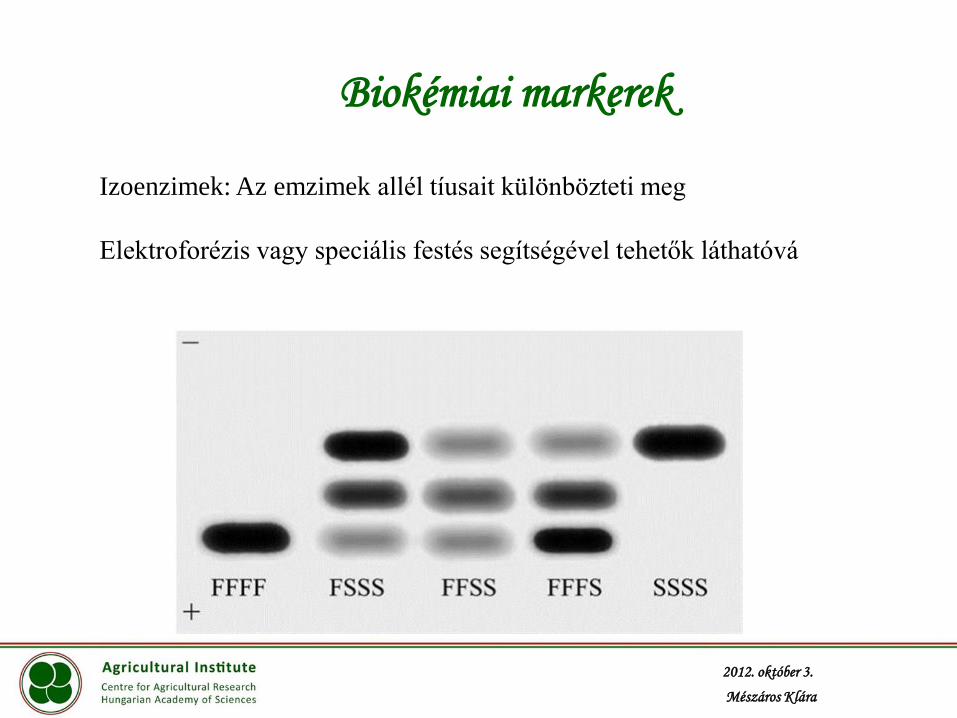
Biokémiai markerek
Izoenzimek: Az emzimek allél tíusait különbözteti meg
Elektroforézis vagy speciális festés segítségével tehetők láthatóvá

2012. október 3.
Mészáros Klára 2012. október 3. Mészáros Klára
Genetikai markerek
A genetikai markerek az vizsgált szervezetek közötti genetikai különbségeket
teszik láthatóvá.
Morfológiai marker Számuk korlátozott megnyilvánulásuk
Biokémiai markerek környezeti hatástól és
fejlődési állapottól függő
Molekuláris markerek Nukleotid azaz DNS szintű variabilitás vizsgálatára alkalmasak. Számuk korlátlan,
megnyilvánulásuk nem függ a környezeti hatásoktól. Az ideális DNS markerre jellemző a
nagyfokú polimorfizmus
koddomináns öröklődés
Gyakori előfordulás
Könnyű hozzáférhetőség
Reprodukálhatóság
Marker kapcsoltsági térképek
Genom
Kódoló Nem kódoló
Morfológiai marker Biokémiai marker Molekuláris marker

Monomorf és polimorf markerek
2012. október 3.
Mészáros Klára

2012. október 3.
Mészáros Klára
Molekuláris marker technikák
Hibridizáción alapuló
RFLP
PCR alapú markerek
ismeretlen kromoszómális elhelyezkedésű: RAPD, AFLP
ismert kromoszómális elhelyezkedésű: SSR, STS
funkcionális markerek: EST, SNP, Tillling
HTP marker technikák (DArT)
Szekvencia alapú
Új generációs szekvenálás (Illumina platform)
Marker kapcsoltsági térképek

2012. október 3.
Mészáros Klára 2012. október 3. Mészáros Klára
RFLP (Restriction Fragment Lenght Polimorphism)
A DNS molekula restrikciós endonukleázzal történő emésztésén és Southern-hibridizáción alapuló módszer.
A hasító helyeken történt pont mutáció (SNP) vagy inszerció és deléció (INDEL) okozta különbségek mutathatók ki.
Restrikciós endonukleázok:
– bakteriális eredetű enzimek
– 4-10 nukleotid hosszúságú palindróma szekcvenciát ismernek fel
– nagyon specifikusak az adott DNS-szekvenciára; (ha akár csak egy bázisnyi különbség van a hasítási helyen, már nem történik meg a hasítás)
– a hasítással ragadós (EcoRI) vagy tompa (SmaI; HaeIII) vég jön létre
Marker kapcsoltsági térképek

2012. október 3.
Mészáros Klára
RFLP
Lépések: a vad típusú és az ismeretlen DNS hasítása restrikciós endonukleázokkal (ha a
mutáció a restrikciós hasítóhelyen volt, akkor ott az enzim nem képes hasítani)
a mintát gélre vinni, elektromos erőtérben megfuttatni
blottolás membránra
hibridizáció – általában radioaktív próbával
Autoradiográfia
Előnye: Nem szükséges a templát DNS szekvenciájának ismerete
A heterozigóták megkülönböztetését biztosító kodomináns módszer
Megbízható, jól ismételhető
Hátránya: Egyes fajokban a restrikciós hasítóhelyek megfelelő szintű
polimorfizmusának hiánya
Nagy mennyiségű DNS-t igényel
Southern hibridizációhoz jelölt próba szükséges
Hosszadalmas és költséges

2012. október 3.
Mészáros Klára
Molekuláris marker technikák
RFLP
PCR alapú markerek
ismeretlen kromoszómális elhelyezkedésű: RAPD, AFLP
ismert kromoszómális elhelyezkedésű: SSR, STS
funkcionális markerek: EST, SNP, Tillling
HTP marker technikák (DArT)
Új generációs szekvenálás (Illumina platform)
Marker kapcsoltsági térképek

2012. október 3.
Mészáros Klára
PCR (polymerase chain reaction)
Akár 1 sejtből származó, egyetlen nukleinsav molekula megsokszorozására enzimatikus úton, élő
szervezet (baktérium, élesztő) igénybevétele nélkül
……..95C………
……………………
….
DNS szálak
kitekerednek
…..denaturáció…
……..
Hűtés primerek
olvadáspontja alá
1-2 fokkal
Primerek
bekötődése
hibridizáció
72 C
DNS polimeráz a
primerektől építi a
DNS láncot
polimerizáció
DNS
Nukleotidok
Primerek
DNS polimeráz
puffer

2012. október 3.
Mészáros Klára
PCR (polymerase chain reaction)
Agaróz gélelektroforézis
EtBr festés
Horizontális gél elektroforézis
PCR készülék PCR alapú markerek
Előnye: Gyors, kis mennyiségű DNS-t igényel, nem kell jelölt
próbát alkalmazni
Hátránya: Megbizhatósága függ az alkalmazott markertípustól

2012. október 3.
Mészáros Klára 2012. október 3. Mészáros Klára
Molekuláris marker technikák
ismeretlen kromoszómális elhelyezkedésű:
RAPD (Randomly Amplyfied Polymorphic DNA),
Nagyfokú polimorfizmus
Reprodukálhatósága függ a kisérleti körülményektől
A termék szekvenálása után specifikus primer tervezhető (SCAR) Kodominánsá
tehető
CAPS (Cleaved Amplified Polymorphic Secuence)
A random amplifikáció során kapott monomorf mintázat (azonos méret, de
különböző szekvencia) restrikciós hasitással polimorffá tehető. Kodomináns
AFLP (Amplified Fragment Lenght Polimorphism)
A genomi DNS restrikciós emésztésén és a fragmantumok szelektív PCR
amplifikációján alapul. Sok fragmentumot eredményez, domináns markereket
eredményez, lokusz-specifikus markerek fejlesztése nemhéz.
Marker kapcsoltsági térképek

2012. október 3.
Mészáros Klára
Molekuláris marker technikák
ismert kromoszómális elhelyezkedésű:
SSR (Simple Sequence Repeat)
Fajtól függően a genom 10-90% monoton ismétlődő nukleotid motívumokat tartalmaz (AC, AG, CG, TA, AG/TC, AC/TG)
A határszekvenciákra tervezett primerekkel hossz polimorfizmus mutatható ki.
Jól használható genotipizálásra, ujjlenyomat meghatározásra.
STS (Sequence-Tagged-Site)
Egyedi szekvenciájú rövid DNS szakasz
Kromoszómális elhelyezkedése ismert
Igen gyors és megbízható eszközzé vált a genetikai vizsgálatokban, mely térképezésre és markerszerlekcióra egyaránt használható
Marker kapcsoltsági térképek

2012. október 3.
Mészáros Klára
DNS markerek, mint ujjlenyomatok
„A”
szülő
„B”
szülő
„A” x „B” keresztezés
utódpopulációjának növényegyedei
(N1………)
N1 N2 N5 N10 N15
Mar
ker
ek (
M1…
……
)
M1
M2
M3

2012. október 3.
Mészáros Klára
Molekuláris marker technikák
funkcionális markerek: EST (Expressed sequence Tagg)
cDNS-ek részei
RNS izolálás > cDNS klónozás > szekvenálás > EST adatbázis > primer tervezés
Összehasonlító térképezés
Marker kapcsoltsági térképek
SNP (Single nucleotide polymorphysms) DNS szekvencia variáció, mely akkor jön létre, ha egy nukleotid a genomban megváltozik (pl.: AAGCCTA AAGCTTA ) – tulajdonképpen pontmutáció
Csak akkor tekinthetjük a variációt SNP-nek, ha a populáció legalább 1%-ában megjelenik
Allélvariációk kimutatása Szekvencia ismeret szükséges
Tillling (Targeting Induced Local Lesions IN Genomes) Mutáns populáció előállítás HTTP mutáns azonosítás

2012. október 3.
Mészáros Klára
eSNP detektálás
Árpában több, mint 420,000 EST áll rendelkezésre 8 különböző genotípusból
EST jelentős többségében kimutatható SNP a fajták között
10,985 eSNP azonosítottak 3,334 génben
Elsődleges szempont, az eredmények egységesíthetősége az ÁRPA1 Gén-Chip (kapcsolat a gén expressziós mintázatokkal), valamint a BAC- könyvtár alapú fizikai térkép információkkal

2012. október 3.
Mészáros Klára
Molekuláris marker technikák
HTP marker technikák
Speciális műszerezettséget igényel
Multiplex módszerek
Rövid idő alatt több száz vagy ezer marker
Szolgáltatás jellegűek DArT (Diversity Arrays Technology)
hybridizáción alapuló marker technológia
Nem igényel szekvencia információkat
Nagy mennyiségű adat létrehozása
Új generációs szekvenálás Illumina platform,
Marker kapcsoltsági térképek
2012. október 3.
Mészáros Klára
http://www.illumina.com
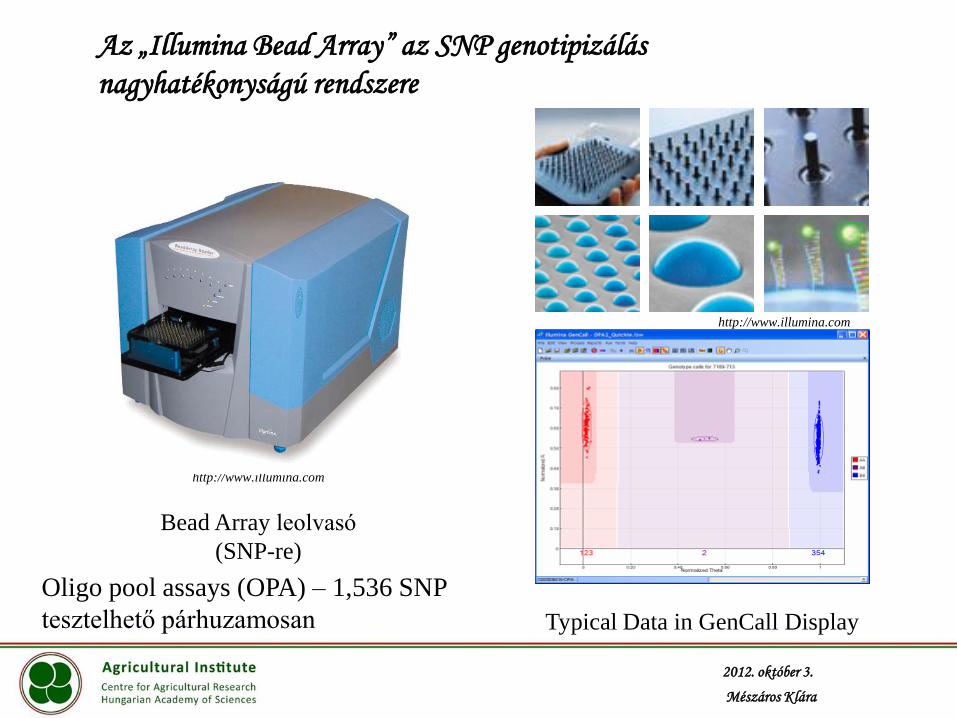
Az „Illumina Bead Array” az SNP genotipizálás
nagyhatékonyságú rendszere
http://www.illumina.com
Bead Array leolvasó
(SNP-re)
Typical Data in GenCall Display
Oligo pool assays (OPA) – 1,536 SNP
tesztelhető párhuzamosan

2012. október 3.
Mészáros Klára
A növénynemesítésben és a genetikában az egyik alapvető cél, hogy a
tulajdonságokat meghatározó gének kromoszómális lokalizációját, és
egymáshoz viszonyított sorrendjüket meg tudjuk határozni.
A géntérképezés során a gének egymástól való távolságát, a genomban
való viszonylagos elhelyezkedésüket határozzuk meg anélkül, hogy
valóban megszekvenálnánk a DNS-t.
Ezt oly módon érik el, hogy az együtt öröklődő tulajdonságok
(koszegregáció) gyakoriságát figyelik, tehát a Mendeli független
öröklődéstől való eltérést, és ebből következtetnek a gének egymástól
való távolságára a kromoszómán.
Amint készen áll a térkép, képesek vagyunk egy ismert gén vagy marker
alapján megmondani egy másik gén helyét a genomban.
Marker kapcsoltsági térképek

Rekombináció
Molekuláris rekombináció: DNS törése és újraegyesülése
(nem feltétlenül homológ szakaszoké)
2012. október 3.
Mészáros Klára
Genetikai rekombináció: új (nem-szülői) allélkombinációk
létrejötte, mely a meiózis profázisában történő nem testvér
kromatidák közötti molekuláris rekombináció eredménye
(crossing over)

Két tulajdonság együttes öröklődésmenete
P1 (AABB) x P2 (aabb)
F1 AaBb (AB|ab –haplotípus: kapcsolt allélek sora)
nem rekombináns nem rekombináns
rekombináns
F1 gamétái: AB Ab aB ab
ha A és B független 25 25 25 25
ha A és B kapcsolt 35 15 15 35
ha szorosan kapcsoltak 48 2 2 48
ha teljesen kapcsoltak 50 0 0 50
2012. október 3.
Mészáros Klára

Testcross rekombináció gyakorisága
A testcross megmutatja az F1 szülőben keletkező 4-féle
gaméta arányait, mert a tester szülő mindig ab-t ad.
F1 x tester: AaBbx aabb
utódok: AaBb Aabb aaBb aabb(4 kül fenotípus)
2012. október 3.
Mészáros Klára
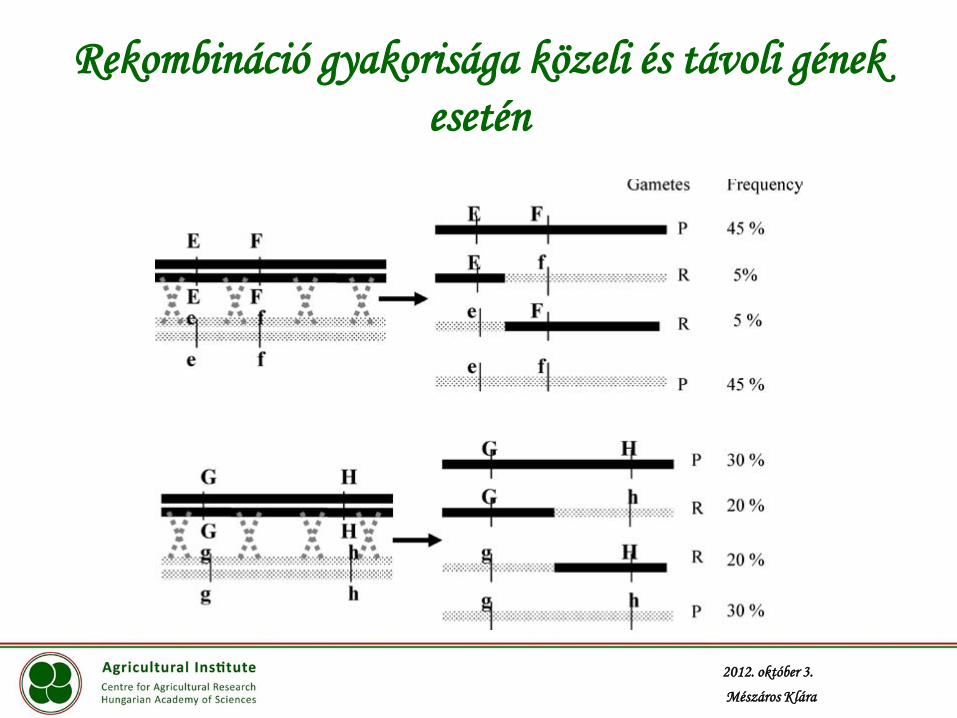
Rekombináció gyakorisága közeli és távoli gének
esetén
2012. október 3.
Mészáros Klára

Térképtávolság
r = rekombináns utódok gyakorisága
rekombináns gaméták száma/ összes gaméta
d = térkép távolság
Kis távolságok (10 cM) esetén d=r
2012. október 3.
Mészáros Klára
A gének közötti, rekombinációs gyakoriság
felhasználásával számolható a térképtávolság.
Mértékegysége a centimorgan (cM).
Egy cM távolság van két gén között, ahol a gaméták
1%-a rekombináns, vagyis a szülőitől eltérő kombinációt
hordoz.

2012. október 3.
Mészáros Klára

Használatos függvények
Morgan d = r (r ≤ 0,5)
Haldane-függvény C=1 (tfh. nincs interferencia)
d = -½ ln(1-2r)
r = ½ (1-e-2d)
Kosambi-függvény C=2r (ha r=0,5 => nincs interferencia,
ha r=0 => teljes interferencia)
d = ¼ ln[(1+2r)/(1-2r)]
r= ½ e4d-1/e4d+1
2012. október 3.
Mészáros Klára

2012. október 3.
Mészáros Klára
Számítógépes térképező programok
marker térkép készítés (MAPMAKER, GMENDEL, JOINMAP)
A LOD - dal kapott r (rekombinációs) gyakoriságokat (megfelelő
lötyögéssel) a térképező programok a legvalószínűbb géntérképekké rakják
össze.
Az r értékekből levezetett d értékeket (Haldane függvény) a kétfaktoros
térképezési logika (additivitások) alapján rendezik.
Marker kapcsoltsági térképek

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térkép elkészítése 8> make chromosome chrom2h chrom4h chrom5h chromosomes defined: chrom2h chrom4h chrom5h 9> seq hvm36 bmag125 sequence #1= hvm36 bmag125 10> anchor chrom2h HvM36 - anchor locus on chrom2h Bmag125 - anchor locus on chrom2h chromosome chrom2h anchor(s): HvM36 Bmag125

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térkép elkészítése 8> make chromosome chrom2h chrom4h chrom5h chromosomes defined: chrom2h chrom4h chrom5h 9> seq hvm36 bmag125 sequence #1= hvm36 bmag125 10> anchor chrom2h HvM36 - anchor locus on chrom2h Bmag125 - anchor locus on chrom2h chromosome chrom2h anchor(s): HvM36 Bmag125
16> assign HvM36 - anchor locus on chrom2h...cannot re-assign Bmag125 - anchor locus on chrom2h...cannot re-assign Bmac30 - anchor locus on chrom4h...cannot re-assign HvM67 - anchor locus on chrom4h...cannot re-assign Bmag337 - anchor locus on chrom5h...cannot re-assign ABG702 - anchor locus on chrom5h...cannot re-assign ABG8 - assigned to chrom2h at LOD 20.1 ABC311 - assigned to chrom2h at LOD 24.4 OPH121 - assigned to chrom2h at LOD 28.6 OPB162 - assigned to chrom2h at LOD 23.8 ABG318 - assigned to chrom2h at LOD 22.9 ABG358 - assigned to chrom2h at LOD 13.8 ABG459 - assigned to chrom2h at LOD 24.4

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térkép elkészítése 8> make chromosome chrom2h chrom4h chrom5h chromosomes defined: chrom2h chrom4h chrom5h 9> seq hvm36 bmag125 sequence #1= hvm36 bmag125 10> anchor chrom2h HvM36 - anchor locus on chrom2h Bmag125 - anchor locus on chrom2h chromosome chrom2h anchor(s): HvM36 Bmag125
16> assign HvM36 - anchor locus on chrom2h...cannot re-assign Bmag125 - anchor locus on chrom2h...cannot re-assign Bmac30 - anchor locus on chrom4h...cannot re-assign HvM67 - anchor locus on chrom4h...cannot re-assign Bmag337 - anchor locus on chrom5h...cannot re-assign ABG702 - anchor locus on chrom5h...cannot re-assign ABG8 - assigned to chrom2h at LOD 20.1 ABC311 - assigned to chrom2h at LOD 24.4 OPH121 - assigned to chrom2h at LOD 28.6 OPB162 - assigned to chrom2h at LOD 23.8 ABG318 - assigned to chrom2h at LOD 22.9 ABG358 - assigned to chrom2h at LOD 13.8 ABG459 - assigned to chrom2h at LOD 24.4
18> three point Linkage Groups at min LOD 3.00, max Distance 37.2 Triplet criteria: LOD 3.00, Max-Dist 37.2, #Linkages 2 'triple error detection' is on. counting...246 linked triplets in 2 linkage groups log-likelihood differences count markers a-b-c b-a-c a-c-b 1: ABG8 ABC311 OPH121 0.00 -0.20 0.00 2: ABG8 ABC311 HvM36 0.00 -0.20 -0.61 3: ABG8 ABC311 OPB162 0.00 -0.19 -1.92 4: ABG8 ABC311 ABG318 0.00 -0.18 -2.95 5: ABG8 ABC311 ABG358 0.00 -0.09 -19.21 6: ABG8 ABC311 ABG459 0.00 -0.08 -19.77 7: ABG8 ABC311 Bmc222a 0.00 -0.09 -18.78 8: ABG8 ABC311 OPP102b 0.00 0.00 -5.44 9: ABG8 ABC311 OPA192 -0.16 0.00 -6.41 10: ABG8 ABC311 OPQ10 -0.11 0.00 -16.98

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térkép elkészítése 8> make chromosome chrom2h chrom4h chrom5h chromosomes defined: chrom2h chrom4h chrom5h 9> seq hvm36 bmag125 sequence #1= hvm36 bmag125 10> anchor chrom2h HvM36 - anchor locus on chrom2h Bmag125 - anchor locus on chrom2h chromosome chrom2h anchor(s): HvM36 Bmag125
16> assign HvM36 - anchor locus on chrom2h...cannot re-assign Bmag125 - anchor locus on chrom2h...cannot re-assign Bmac30 - anchor locus on chrom4h...cannot re-assign HvM67 - anchor locus on chrom4h...cannot re-assign Bmag337 - anchor locus on chrom5h...cannot re-assign ABG702 - anchor locus on chrom5h...cannot re-assign ABG8 - assigned to chrom2h at LOD 20.1 ABC311 - assigned to chrom2h at LOD 24.4 OPH121 - assigned to chrom2h at LOD 28.6 OPB162 - assigned to chrom2h at LOD 23.8 ABG318 - assigned to chrom2h at LOD 22.9 ABG358 - assigned to chrom2h at LOD 13.8 ABG459 - assigned to chrom2h at LOD 24.4
18> three point Linkage Groups at min LOD 3.00, max Distance 37.2 Triplet criteria: LOD 3.00, Max-Dist 37.2, #Linkages 2 'triple error detection' is on. counting...246 linked triplets in 2 linkage groups log-likelihood differences count markers a-b-c b-a-c a-c-b 1: ABG8 ABC311 OPH121 0.00 -0.20 0.00 2: ABG8 ABC311 HvM36 0.00 -0.20 -0.61 3: ABG8 ABC311 OPB162 0.00 -0.19 -1.92 4: ABG8 ABC311 ABG318 0.00 -0.18 -2.95 5: ABG8 ABC311 ABG358 0.00 -0.09 -19.21 6: ABG8 ABC311 ABG459 0.00 -0.08 -19.77 7: ABG8 ABC311 Bmc222a 0.00 -0.09 -18.78 8: ABG8 ABC311 OPP102b 0.00 0.00 -5.44 9: ABG8 ABC311 OPA192 -0.16 0.00 -6.41 10: ABG8 ABC311 OPQ10 -0.11 0.00 -16.98
Placing at log-likelihood threshold 3.00...
Start: 9 5 1 11 12
Npt-2: 9 5 1 (10) 11 12
No unique placements for 6 remaining markers
Map:
Markers Distance
9 Bmc222a 21.5 cM
5 OPB162 8.2 cM
1 ABG8 10.6 cM
10 OPP102b 6.3 cM
11 OPA192 10.6 cM
12 OPQ10 ----------
57.1 cM 6 markers log-likelihood= -98.26
Markers placed relative to above map:
9 5 1 10 11 12
:-21-:--8-:-11-:--6-:-11-:
8 2 .*.:.**.:....:....:....:....:...
7 2 .*.:.**.:....:....:....:....:...
6 2 ...:.**.:..*.:....:....:....:...
4 2 ...:..*.:.**.:....:....:....:...
3 2 ...:....:.**.:..*.:....:....:...
2 2 ...:....:.**.:..*.:....:....:...
------------------------------------------------------------------
Placing at log-likelihood threshold 2.00...
Start: 9 5 1 10 11 12
Npt-4: 9 5 (2) 1 10 11 12
Npt-4: 9 5 (4) 2 1 10 11 12
Npt-4: 9 5 4 (3) 2 1 10 11 12

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térkép elkészítése 8> make chromosome chrom2h chrom4h chrom5h chromosomes defined: chrom2h chrom4h chrom5h 9> seq hvm36 bmag125 sequence #1= hvm36 bmag125 10> anchor chrom2h HvM36 - anchor locus on chrom2h Bmag125 - anchor locus on chrom2h chromosome chrom2h anchor(s): HvM36 Bmag125
16> assign HvM36 - anchor locus on chrom2h...cannot re-assign Bmag125 - anchor locus on chrom2h...cannot re-assign Bmac30 - anchor locus on chrom4h...cannot re-assign HvM67 - anchor locus on chrom4h...cannot re-assign Bmag337 - anchor locus on chrom5h...cannot re-assign ABG702 - anchor locus on chrom5h...cannot re-assign ABG8 - assigned to chrom2h at LOD 20.1 ABC311 - assigned to chrom2h at LOD 24.4 OPH121 - assigned to chrom2h at LOD 28.6 OPB162 - assigned to chrom2h at LOD 23.8 ABG318 - assigned to chrom2h at LOD 22.9 ABG358 - assigned to chrom2h at LOD 13.8 ABG459 - assigned to chrom2h at LOD 24.4
18> three point Linkage Groups at min LOD 3.00, max Distance 37.2 Triplet criteria: LOD 3.00, Max-Dist 37.2, #Linkages 2 'triple error detection' is on. counting...246 linked triplets in 2 linkage groups log-likelihood differences count markers a-b-c b-a-c a-c-b 1: ABG8 ABC311 OPH121 0.00 -0.20 0.00 2: ABG8 ABC311 HvM36 0.00 -0.20 -0.61 3: ABG8 ABC311 OPB162 0.00 -0.19 -1.92 4: ABG8 ABC311 ABG318 0.00 -0.18 -2.95 5: ABG8 ABC311 ABG358 0.00 -0.09 -19.21 6: ABG8 ABC311 ABG459 0.00 -0.08 -19.77 7: ABG8 ABC311 Bmc222a 0.00 -0.09 -18.78 8: ABG8 ABC311 OPP102b 0.00 0.00 -5.44 9: ABG8 ABC311 OPA192 -0.16 0.00 -6.41 10: ABG8 ABC311 OPQ10 -0.11 0.00 -16.98
Placing at log-likelihood threshold 3.00...
Start: 9 5 1 11 12
Npt-2: 9 5 1 (10) 11 12
No unique placements for 6 remaining markers
Map:
Markers Distance
9 Bmc222a 21.5 cM
5 OPB162 8.2 cM
1 ABG8 10.6 cM
10 OPP102b 6.3 cM
11 OPA192 10.6 cM
12 OPQ10 ----------
57.1 cM 6 markers log-likelihood= -98.26
Markers placed relative to above map:
9 5 1 10 11 12
:-21-:--8-:-11-:--6-:-11-:
8 2 .*.:.**.:....:....:....:....:...
7 2 .*.:.**.:....:....:....:....:...
6 2 ...:.**.:..*.:....:....:....:...
4 2 ...:..*.:.**.:....:....:....:...
3 2 ...:....:.**.:..*.:....:....:...
2 2 ...:....:.**.:..*.:....:....:...
------------------------------------------------------------------
Placing at log-likelihood threshold 2.00...
Start: 9 5 1 10 11 12
Npt-4: 9 5 (2) 1 10 11 12
Npt-4: 9 5 (4) 2 1 10 11 12
Npt-4: 9 5 4 (3) 2 1 10 11 12
chrom2h framework:
Markers Distance
7 ABG358 20.6 cM
5 OPB162 2.4 cM
4 HvM36 3.1 cM
3 OPH121 0.0 cM
2 ABC311 2.1 cM
1 ABG8 10.6 cM
10 OPP102b 6.3 cM
11 OPA192 10.6 cM
12 OPQ10 ----------
55.6 cM 9 markers
log-likelihood= -101.39

2012. október 3.
Mészáros Klára
DK1DK2DK3DK4DK7DK8DK9DK10DK11DK12DK13DK14DK15DK16DK17DK18DK19DK20DK21DK22DK24DK25DK26DK27DK28DK29DK30DK31DK32DK35DK36DK37DK38DK39
*mst102 A B A A A B B B A A B B A B B B A B B B A B B A B B A A B A A B A B
*Bmac144d A B A A A B B B A B B A B B B B B B B B A B B A B B A A B A A B A B
TCGA96 A B A A A B B B A B B A B B B A B B B B A B B A B B A A B A A B A B
*MWG938 A B A A B B B B A B B A B B B A B B B B A B B A B B A A B A A B A B
AGAT166/162 A B A A B B B B A B B A B B B A B B B B A B B A B B A A B A A B A B
GCTC58 A B A A B B B B A B B A B B B A B A B B A B B A B B A A B A B B B B
OPK16 A B A A B B B B A B B A B B B A B A B B A B B A B B A A B A B B B B
*abc156.1 A B A A B B B B A B B A B B B A B A B B A B B A B B A A B A B B B B
*BMAC213 A B A A B B B B A B B A B B B A B A B B A B B A B B A A B A B B B B
GAAT76 A B A A B B B B A B B A B B B A B A B B A B B A B B A A B A B A B B
*OPE171 A B A A B B B B A B B - B B B A B A B B A B B A B B B A B A B A B B
GAAT414 A B A B B B B B A B B A B B B A B A B B A B B A B B B A B A B A B A
*OPO18 A B A B B B B B A B B A B B B A B A B B A B B A B B B A B A B A B A
*OPB16-1 A B A B B B B B A A B A B B B A B A A A A B B A B B B A B A B A B A
*Bmag211 A B A B B B B B B A B A B B B A B A A A A B B A B B B A B A B A B A
*OPS16 A B A B B B B B B A B A B B B A B A A A A B B A B B A A B A B A B A
GCGA428 A B A B B B B B B A B A B B B A B A A A A B B A B B A A B A B A B A
*Ebmac560a A B A B B B B B B A B A B B B A B A A A A B B A B B A A B A B A B A
*OPB41 A B A B B B B B B A B A B B B A B A A A A B B A B B A A B A B A B A
GAAT550 A B A B B B B B B A B A B B B A B A A A A B B A B B A A B A B A B A
AGGT290 A B A B B B B B B A B A B B B B B A A A A B B A B B A A B A B A B A
TCGA119 A B A B B B B B B A B A B B B B B A A A A B B A B B A A B A B A B A
*OPT152 A B A B B B B B B A B A B B B B B A A A A B B A B B A A B A B A B A
GAAT510 A B A B B B B B B A B A B B B B B A A B A B A A A A A B A A B B A A
AGAT51 A B A B A A A B B A B A B B B B B B A B B B A A A A A B A A A B A B
*Bmac144a A B A B A A A A B A B A B B B B B B A B B B A A A A A B A A A B A B
GCGA269/270 A B A B A A A A B A B A B B B B B B A B B B A A A A A B A A A B A B
*ABC152b A B A B A A A A B A B A B B B B B B A B B B A A A A A B A A A B A B
*HvHVA1 A B B B A A A A B A B A B B B B B B A B B B B A A A A B A A A B B B
*BCD265c A B B B A A A A B A B A B B B B B B A B B B B A A A A B A A A B B B
GCGT132 A B B B A A A A B A B A B B B B B B B B B B B A A A A B A A A B B B
*OPP142 A B B B A A A A B A B A B B B B B B B B B B B A A A A B A A A B B A

2012. október 3.
Mészáros Klára
mst102 0 Bmac144d 4 TCGA96 6 MWG938 8 AGAT166 11 GCTC58 15 OPK16 17 abc156.1 18 Bmac213 22 GAAT76 24 OPE171 28 GAAT414 30 OPO18 31 OPB161 41 Bmag211 48 OPS16 50 GCGA428 53 Ebmac560a 54 OPB041 56 GAAT550 57 AGGT290 58 TCGA119 59 OPT152 61 GAAT510 86 AGAT51 99 Bmac144a 100 GCGA269 104 ABC152b 109 HvHVA1 125 BCD265c 126 GCGT132 130 OPP142 136 OPX031 163
1H
Bmac222a 0 ABG459 1 ABG318 18 HvM36 25 ABC311 28 GAGT494 29 ABG8 31 GCGA52 37 GAGA72 39 TCTC212 40 GAAT338 41 OPA192 42 AGAT347 51 AGAT248 56 GAGA573 57 GAGT62 58 GCGT250 88 abg459a 101 Bmac216 102 Bmac144b 108 bcd175b 109 Bmag125 110 GAGT670 113 TCGT280 115 TCAT572 116 TCTC631 126 Bmag003c 127 GCAT142 128 OPG07 142
2H
OPD07 0 OPS141 6 OPO072 11 GATC355 14 GCGA288 16 TCAT132 17 GAGA642 19 TCTC626 22 AGTC201 50 GAAT308 55 GCAT191 59 OPI041 60 AGAT498 72 OPS202 73 Bmag384b 75 TCTC548 76 GCGT270 87 OPK111 92 HvM60 99 AGAT175 102 AGAT598 103 GAAT182 104 AGGT472 106 GCGT164 108 OPH15 132 ABG4 152 TCAT477 155 Bmag13 159 GCAT104 172 TCTC216 175 OPS192 179 Hvm62 186 GAGA650 189 OPH122 194 Hvm70 195 GCGT68 197 TCAT228 204 GCAT227 205 GCGA205 209 TCGA205 210 ABC174 211 AGTC335 212 GAAT147 220
3H
HvPhyA 0 GCAT76 8 TCAT300 TCAT77 9
AGAT142 16 HvPhyB 21 CAB 22 Bmac30 24 AGAT312 29 Bmag353 32 Bmag173a 33 Bmac310 39 OPJ15 40 TCAT452 41 OPU162 42 AGAT46 59 ABG366 62 abg54 64 AGAT148 65 GCGT161 72 GCAT312 79 AGAT406 84 HvM67 85 OPS031 86
VRN-H2 HvSnf2 92
AGGT650 93 Hdamyb 100
4H
OPT151 0 GCAT50 7 Bmag136b 8 GAGA556 9 Bmag337 13 GATC565 18 AGTC122 21 AGGA575 24 GAGA220 27 AGAT42 32 GAGA58 33 Bmag113 35 abc156.5 37 AGAT676 38 TCTC502 40 Bmag223 mR 41
GATC106 47 abg459b 51 OPH123 54 GCGT406 58 GCGA434 71 TCAT332 88 abc306d 90 OPK112 92 TCTC510 95 ABG702 96 AGTC50 101 Bmag381b 112 gms061 114 HvPhyC
VRN-H1 117
abc155 137 TCGT372 142 GCAT406 156 ABC310 161
5H
BMAC316 0 AGGT256 2 TCTC386 9 AGAT658 13 ABC152c 23 Bmag500 26 Bmag21a 51 GAGA626 62 GCGT262 64 Bmag219 65 OPH081 68 Bmag009 69 HvCry1a HvCry2 GCGA48
71
AGGA62 72 OPI042 75 OPA203 78 abc306b 80 abc306c 81 abc303 84 AGGT336 87 Bmag003b 89 ebmac806 90 TCTC576 93 OPT14 95 GCTC258 97 GAGA170 99 AGGT484 102 TCAT216 107 GCAT212 108 AGAT298 111 wg464a GAGA622 136
GAGA596 143 TCTC528 149 TCAT232 150 GCGT382 151
6H
Hvm4 0 Bmac222b 9 GCAT188 11 TCAT516 12 TCTC250 13 abg460 14 TCTC623 15 abc152a
16
GCGT69 25 AGTC400 28 TCAT123 30 Bmag21c 32 GCAT299 33 Ebmac603 36 TCGT144 42 TCGA273 48
VRN-H3 50 KSuA1a 52 abc306a 59 TCGT153 60 TCGT64 62 AGGT64 65 OPE172 73 Bmac187 75 TCGT143 78 Bmag120 81 OPR152 86 OPS032 87 mst101b 88 Bmag369 90 AGAT87 107 AGAT223 118 GCGT206 123 OPQ17 127 TCTC71 143 OPB031 145 AGGT86 147 GCTC158 157 GAAT418 172
7H
B1
B3
B7
B9
B10 B12 B13
B5 B3
B3
B7
B7
B8
B15
B14
B16
B18
B19
B4
B5
B6
B1
0
B1
2
B1
4
B4
B5
B7
B11
B13
B18
B1
B3 B4
B8
B1
0
B1
2
B3
B6
B5
B9
B12
B10
Dicktoo × Kompolti korai marker térkép
(246 lókusz)

Oregon Wolfe Barley Map
Centromere
Morphological marker
ABG7040.0
Bmag000714.2
ABG38033.9
HVCMA44.3Bmac018749.6X1.162.4Bmac047B65.9DAK64268.2MWG80870.1nud81.6
lks294.2
Bmag0120104.0WG380B107.1
Ris44117.3
ABC253132.5
ABG461A142.6WG380A146.9X1.2153.6
HVM5172.0ThA1175.6
1 (7H)
ABG0580.0DAK6058.8ABG00814.8X2.139.2Pox42.4MWG949B55.8Hot160.6EBmac068461.8ABG35666.3X2.268.5Bmag0113E77.9Bmac0144F84.1vrs191.3Bmag012597.0MWG503104.2MWG844E104.9X2.3105.7KFP203106.7MWG882A110.0ABG072124.9EBmac0415137.8cnx1139.5zeo148.9X2.4161.1MWG720164.4X2.5170.2wst172.7X2.6178.8MWG949A180.3
2 (2H)
BCD9070.0
ABC171A26.4X3.131.5
ABG46041.3
alm62.2Bmac020967.9ABC32570.7BCD114574.6Bmac006783.0
Bmag022599.8HVM60106.2
X3.2124.6ABG499125.8X3.3129.3
Pub149.7
MWG883169.4
HVM62183.7
ABC805191.8
ABC172202.0
3 (3H)
MWG6340.0MWG07721.6HVM4024.4CDO54231.2CDO12232.3hvknox337.3Dhn640.9ABC30342.9HVM353.1X4.164.2Bmag035365.2X4.266.2Bmac018669.6ABG47284.6X4.388.2KFP22197.5MWG652B100.0EBmac0701102.7X4.4109.3Hsh119.6HVM67120.6X4.5132.4ABG601134.4
4 (4H)
Rh0.0Act8A8.8kAzmil15.6MWG837B21.2MWG837A24.5Bmac039929.0X5.130.1BCD09836.3X5.248.6Bmag021158.3ABG49459.2X5.368.9ABC16075.4Bmac0144A85.0Bmag0113A88.3MWG706A98.2Blp109.8MWG2028119.2ABC261120.8X5.4127.1WMC1E8130.3MWG912133.8ABG387A137.1
5 (1H)
Bmac03160.0MWG6204.0
MWG652A28.8MWG602B33.0
X6.149.4ABG45854.5X6.262.0HVM3168.7rob70.5Bmag000971.6X6.381.1Bmac0218C88.1ABG38893.8MWG820103.3
X6.4121.6MWG934124.6Tef1132.0X6.5137.2Bmac0040145.5
X6.6165.0
6 (6H)
MWG6180.0ABC4835.5ABG6106.6
ABG39529.0
Bmac0273B38.2Bmac009645.3KFP00249.8
ABC30268.3
srh88.3
Bmag0113D102.0
ABG003B126.8
MWG877149.8X7.1151.9
ABG496163.6
ABG391182.8
ABC622191.4
Bmag0113C201.3
MWG602A210.5
7 (5H)

2012. október 3.
Mészáros Klára
Összehasonlító térképezés
fajon belül különböző populációk között,
rokon fajok között
Genom szerveződés
makro és mikro kolinearitás
kromoszóma átrendeződések
intergénikus és génklaszter szakaszok azonosítása
Marker kapcsoltsági térképek alkalmazási
területei

2012. október 3.
Mészáros Klára
Egyedi térképek – fajra jellemző konszenzus
térkép
Stein et al. TAG(2007)114:823

2012. október 3.
Mészáros Klára
A búza kromoszómák BIN térképe
Qi et al. Genetics 2004

2012. október 3.
Mészáros Klára
2BS-1
2BS-3
2BL-6
wPt-1494 wPt-3188 wPt-3983 wPt-4701 wPt-4737 wPt-4997
wPt-7408 wPt-7757 wPt-8070
wPt-5249 wPt-5556 wPt-5597 wPt-7348 wPt-7610 wPt-8072 wPt-8294 wPt-8460 wPt-8720 wPt-9402 wPt-6144 wPt-6053 wPt-6627
wPt-1650 wPt-8074 wPt-0408 wPt-0615 wPt-1489
wPt-1549 wPt-2249 wPt-2397 wPt-3807 wPt-5287
wPt-6970 wPt-7123 wPt-9288 wPt-6311 wPt-8737
wPt-4916 wPt-9423 wPt-5587 wPt-5934 wPt-6575
wPt-9859 wPt-9220 wPt-3459
wPt-7995
wPt-3188
wPt-1549
wPt-5128
P92201D5-2/P91193D1-10 Ajana/WAWHT2074 Cadoux/Reeves EGA Blanco/Millewa
wPt-5195* wPt-0643wPt-6575 wPt-5587wPt-3459 wPt-5934wPt-6627*wPt-4916*wPt-0100*wPt-8235*gwm614b*wPt-8404*wPt-1041*wmc25*gwm319*wPt-3569*wPt-1650*wPt-1068*wPt-7937* wPt-0775*wPt-8294* wPt-0395*wPt-1494* wPt-2249*wPt-2907* wPt-0434*wPt-9131*barc183*wPt-0615*wPt-1127*wPt-5440*wPt-6144*wPt-0079*gwm630*wPt-9350*cfd56*AF112966*wPt-7625*gwm47*wmc477*gwm120*wPt-3109*wPt-9336* wPt-7350*wPt-4701wPt-2266*gwm526c*barc1147*barc92
0.30.7
16.42.9
10.58.0
34.60.7
14.122.40.52.24.13.80.21.41.20.50.30.20.32.63.37.67.34.02.03.12.11.3
12.11.9
10.45.4
11.215.0
2B
gwm614wPt-1663wPt-5567wPt-4527cfd238wmc25abarc318wPt-9423 wPt-9402wPt-1489 wPt-8072wPt-4301 wPt-0462wPt-3561 wPt-6932wPt-3983 wPt-5707wPt-2314wPt-6199wPt-8583wPt-6477 wPt-4125wPt-9644 wPt-7757wPt-0408wPt-0615 wPt-5672wPt-8492barc183b*gwm46barc7barc55†wmc474cfa2278wPt-0335wPt-0709wPt-7750gwm374gwm319 gwm630bgwm271gdm14agwm120agwm191a*wPt-1140*wPt-7200wPt-3132 wPt-0950wPt-7404wPt-5128*gwm120b†wPt-4199gdm61*
14.619.69.64.32.96.25.60.68.90.70.60.70.60.6
21.126.612.22.57.7
12.010.10.60.64.60.66.18.2
11.07.57.22.31.27.6
22.028.320.9
2B
wPt-4527 cfd238wPt-8004 wPt-8326wPt-9423 wPt-9402wPt-1489 wPt-8072wPt-5707wPt-3983 wPt-4997gwm429barc13†wPt-6477wPt-0615wPt-7750wPt-2430*wPt-8569*
9.210.85.02.28.17.60.70.72.9
37.8
1.4
2B
wPt-6575* wPt-5587*wPt-3459* wPt-5934*wPt-6970* wPt-6627*wPt-5195* wPt-0643*wPt-6805*wPt-0100*wPt-8760*wPt-2410*wPt-3565*gwm614*wPt-6223*barc35*barc297a*wPt-2106*wPt-8004* wPt-8326*wmc154wPt-5374wPt-3561wPt-5707wPt-7757 wPt-5672wPt-5556 wPt-4125barc55wPt-6278barc1114cfa2043bgwm120wPt-4199*wPt-2430wPt-8569wPt-0094wPt-7200wPt-3132wPt-8460wPt-5680wPt-5242wPt-0694stm509acagwPt-4701wPt-7004wPt-4210wPt-0047barc1147wPt-2135 wPt-3378barc159wPt-4559
0.93.21.00.40.53.83.40.41.52.36.0
21.79.9
12.510.32.3
12.910.48.25.0
11.23.33.22.41.00.52.40.40.54.17.71.91.8
14.92.12.8
19.11.48.01.4
2B
Chromosome 2B
Marker kapcsoltsági térkép – kromoszóma fizikai térképe

Térképegység (cM) és megabázis (Mb)
1 cM 1Mb
1 Mb = 106bázis
A rekombinációs gyakoriság változó genomon belül is, ezt
jellemzi a rekombinációs ráta, amely 1 körül változik.
Vannak rekombinációs hot-spotok, míg máshol, pl. a
centroméra környékén szinte nincs rekombináció.
2012. október 3.
Mészáros Klára

2012. október 3.
Mészáros Klára
Mennyiségi tulajdonságok lókuszainak (QTL) elemzése
lókuszok számának és helyének azonosítása
Adott lókusz szerepének elemzése; QTL dinamika, tulajdonság
komponensek vizsgálata
Marker kapcsoltsági térképek alkalmazási
területei
Gének azonosítása, fenotípusos hatásuk vizsgálata
finom térképezés, pozicionális klónozás
modell növény ismert génszekvenciája alapján
Allél gyakoriságok vizsgálata szélesebb genetikai bázison
Marker szelekció

2012. október 3.
Mészáros Klára
Single intervall mapping (SIM): Két szomszédos marker közötti
intervallumot vizsgál az egész kromoszóma mentén.
Kiküszöböli a rekombinációból származó torzítást.
QTL elemzés fő módszerei
Composit interval mappiung (CIM): egyesíti az intervallum
térképezést a lineáris regresszió analízissel és a szomszédos
markerpárokat további markerekkel egészíti ki.
Sigle-marker analízis: nincs szükség kapcsoltsági térképre.
Egy marker és az adott tulajdonság kapcsolatát vizsgálja
A linearis regresszió determinációs koefficiense (R2) megadja, hogy
az adott marker a fenotípusos variancia hány százalékét
magyarázza.
Hátránya: a marker és a QTL közti rekombináció esetén
alulbecsli a kapcsoltságot
Nagy populáció méret csökkenti ezt a hatást

Adott tulajdonsággal kapcsolt marker
2012. október 3.
Mészáros Klára

2012. október 3.
Mészáros Klára
QTL azonosítás a teljes marker kapcsoltsági térképre kiterjedve
Markerek közötti intervallumok vizsgálata
Az adott QTL hatásának nagyságát a LOD (likelihood ratio) értékkel jellemezzük
LOD= lg(QTL csúcs jelenlétének valószínűsége/annak a valószínűsége, hogy nincs QTL hatás)
LOD küszöbérték: genom méret
marker típus és sűrűség
populáció típusa és méret
QTL elemzés marker kapcsoltsági térkép
alkalmazásával
Fenotípusos vizsgálatok
ismételhetőség, pontosság
Fenomika

2012. október 3.
Mészáros Klára
Hasadó populáció Genotípusos jellemzés
szülők
A B AAABBAB ABBABABB BBBAA
Fenotípusos jellemzés
*mst1011 ABAAABBBABBBABBBBBABBABBA
*mst1012 ABBAABAABBAABBBBBBABBBBBB
*mst102 AAABBBAABBABBBABBBBABBABB
*wg4642 ABBAABAABBAABBBBBBABBBABB
*bcd1752 ABBAAABBBA---ABBABBBABABAA
*wg541 AA-BBBA---ABBABBABBABABAABA
*v8h49 164.5 157.5 149.5 146.5 154 171 250
*uv16h49 43 128.5 138 159 163.5 143.5 49 51 38
*v16h49 40 57.5 60.5 60.5 69.5 38 44 63 38 62.5
*uv24h49 27 97.5 111.5 123.5 125 103.5 29.5 31.5
Adat mátrix
QTL elemzés marker kapcsoltsági térkép alkalmazásával

2012. október 3.
Mészáros Klára
=============================================================
QTL-Map for peak 3:
Confidence Interval: Left Boundary= OPS31-Snf2 + 2.0
Right Boundary= Snf2-Hdamyb (off end)
INTERVAL LENGTH QTL-POS GENETICS WEIGHT
Snf2-Hdamyb 8.0 0.0 free 78.700
chi^2= 105.665 (1 D.F.) log-likelihood= 22.94
mean= 16.300 sigma^2= 102.058 variance-explained= 93.8%
=============================================================
QTL elemzés marker kapcsoltsági térkép alkalmazásával

2012. október 3.
Mészáros Klára
=============================================================
QTL-Map for peak 3:
Confidence Interval: Left Boundary= OPS31-Snf2 + 2.0
Right Boundary= Snf2-Hdamyb (off end)
INTERVAL LENGTH QTL-POS GENETICS WEIGHT
Snf2-Hdamyb 8.0 0.0 free 78.700
chi^2= 105.665 (1 D.F.) log-likelihood= 22.94
mean= 16.300 sigma^2= 102.058 variance-explained= 93.8%
=============================================================
-------------------------------| OPJ15-ABG366 28.0 cM
0.0 35.24 17.7% 1.585 |
2.0 38.40 21.2% 1.758 |
4.0 41.51 25.0% 1.941 |
6.0 44.33 28.8% 2.126 | *
8.0 46.56 32.0% 2.303 | **
10.0 48.44 34.9% 2.464 | **
12.0 49.63 36.7% 2.599 | ***
14.0 50.31 37.8% 2.704 | ***
16.0 50.48 38.2% 2.776 | ****
18.0 50.03 37.5% 2.815 | ****
20.0 49.35 36.5% 2.822 | ****
22.0 48.24 34.8% 2.800 | ****
24.0 46.68 32.5% 2.754 | ****
26.0 44.82 29.9% 2.688 | ***
28.0 42.79 27.1% 2.609 | ***
-------------------------------| ABG366-abg54 5.5 cM
0.0 42.79 27.1% 2.608 | ***
2.0 45.76 30.5% 2.667 | ***
4.0 44.87 29.1% 2.372 | **
-------------------------------| abg54-OPR195 7.9 cM
0.0 36.31 19.1% 1.753 |
2.0 37.80 20.9% 1.885 |
4.0 38.66 22.0% 1.992 |
cM Weight R2 LOD
QTL elemzés marker kapcsoltsági térkép alkalmazásával

2012. október 3.
Mészáros Klára
=============================================================
QTL-Map for peak 3:
Confidence Interval: Left Boundary= OPS31-Snf2 + 2.0
Right Boundary= Snf2-Hdamyb (off end)
INTERVAL LENGTH QTL-POS GENETICS WEIGHT
Snf2-Hdamyb 8.0 0.0 free 78.700
chi^2= 105.665 (1 D.F.) log-likelihood= 22.94
mean= 16.300 sigma^2= 102.058 variance-explained= 93.8%
=============================================================
-------------------------------| OPJ15-ABG366 28.0 cM
0.0 35.24 17.7% 1.585 |
2.0 38.40 21.2% 1.758 |
4.0 41.51 25.0% 1.941 |
6.0 44.33 28.8% 2.126 | *
8.0 46.56 32.0% 2.303 | **
10.0 48.44 34.9% 2.464 | **
12.0 49.63 36.7% 2.599 | ***
14.0 50.31 37.8% 2.704 | ***
16.0 50.48 38.2% 2.776 | ****
18.0 50.03 37.5% 2.815 | ****
20.0 49.35 36.5% 2.822 | ****
22.0 48.24 34.8% 2.800 | ****
24.0 46.68 32.5% 2.754 | ****
26.0 44.82 29.9% 2.688 | ***
28.0 42.79 27.1% 2.609 | ***
-------------------------------| ABG366-abg54 5.5 cM
0.0 42.79 27.1% 2.608 | ***
2.0 45.76 30.5% 2.667 | ***
4.0 44.87 29.1% 2.372 | **
-------------------------------| abg54-OPR195 7.9 cM
0.0 36.31 19.1% 1.753 |
2.0 37.80 20.9% 1.885 |
4.0 38.66 22.0% 1.992 |
cM Weight R2 LOD
0
10
20
30
40
50
60
70
80
HD16h uv HD16h vern HD24h uv HD24h vern
recombination distance (cM)
LO
D v
alue
s
2H 4H 5H
QTL elemzés marker kapcsoltsági térkép alkalmazásával

2012. október 3.
Mészáros Klára
Kalászolási idő, mint vizsgált
tulajdonság
16h + hő ciklus
0
10
20
30
40
20 30 40 50 60 70 80 90 100 110 120 130 140 150
Xátl=77 nap
Kompolti Dicktoo
Kalás zolás i idő
DH
vo
nala
k s
zám
a
Populáció egyedei közti változékonyság

2012. október 3.
Mészáros Klára
ZCCTH
HvSnf2
Dicktoo x Kompolti korai DH
vonalak HvPhyA 0
GCAT76 8
TCAT300
TCAT77 9
AGAT142 16
HvPhyB 21
CAB 22
Bmac30 24
AGAT312 29
Bmag353 32
Bmag173a 33
Bmac310 39
OPJ15 40
TCAT452 41
OPU162 42
AGAT46 59
ABG366 62
abg54 64
AGAT148 65
GCGT161 72
GCAT312 79
AGAT406 84
HvM67 85
OPS031 86
VRN-H2 HvSnf2
92
AGGT650 93
Hdamyb 100
4H árpa kromoszóma vern
aliz
ála
tlan
vern
aliz
ált k
ezelé
s
QTL elemzés

2012. október 3.
Mészáros Klára
Pozicionális klónozás
Alapja: a QTL hatás 1 cM-nál kisebb marker intervallumhoz köthető
Finomtérképezés, nagy populáció méret bevonásával
A génhez közeli rekombinációt hordozó egyedek azonosítása,
fenotípusos jellemzése
A kromoszóma régióhoz köthető átfedő BAC klónok azonosítása,
térképezése
A keresett gént tartalmazó BAC klón szekvenálása
A BAC klónon talált gén(ek) funkciójának tisztázása
Gén expresszió
transzformáció

2012. október 3.
Mészáros Klára
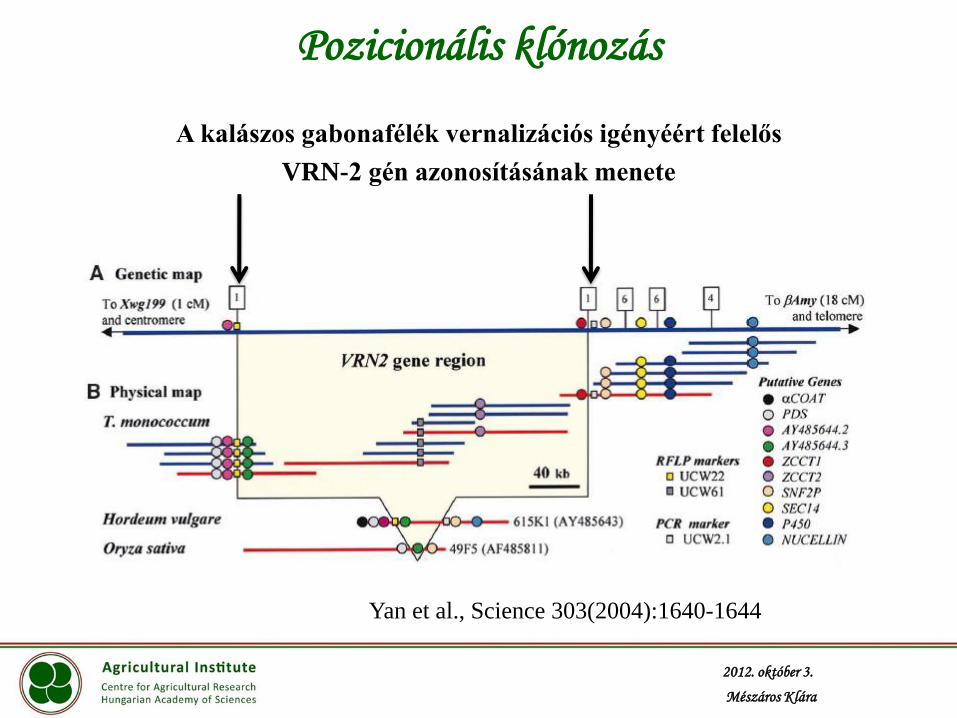
Pozicionális klónozás
A kalászos gabonafélék vernalizációs igényéért felelős
VRN-2 gén azonosításának menete
Yan et al., Science 303(2004):1640-1644
2012. október 3.
Mészáros Klára
Pozicionális klónozás
A kalászos gabonafélék vernalizációs igényéért felelős
VRN-2 gén azonosításának menete
Yan et al., Science 303(2004):1640-1644

2012. október 3.
Mészáros Klára
Pozicionális klónozás
A kalászos gabonafélék vernalizációs igényéért felelős VRN-2 gén azonosításának
menete
Yan et al., Science 303(2004):1640-1644

2012. október 3.
Mészáros Klára
Deléciós vonalak alkalmazása búza
fagytűrésének vizsgálatában

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térképek
Két szülős térképező populációk
Széles genetikai bázist képviselő fajtakör
Jelölt gén megközelítése
Genom pozíció függő stratégiák: pozicionális klónozás, deléciós vonalak
Összehasonlító genomikai stratégiák: modell növények
Mesterséges genetikai variáció (indukált mutációs populációk)
Gén csapdázás: T-DNS, transzpozon
Gén expressziós mintázatok elemzése:
Differential Display
DNS microarray, DNS chip
Gén azonosítás főbb módszerei

2012. október 3.
Mészáros Klára
Széles genetikai bázisú fajtakör – asszociációs
genetika Hagyományos két-szülős térképező populációk hátrányai:
Előállításuk idő és munka igényes
Lókuszonként csak két allél hatásának vizsgálatára van mód
A két szülő közti hasonlóság korlátozza az egyes gének azonosítási lehetőségeit
Adott populációban azonosított QTL csúcshoz szorosan kapcsolt marker nem polimorf a nemesítési anyagban
Gén (allél) kölcsönhatások vizsgálata limitált

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térképek
Két szülős térképező populációk
Széles genetikai bázist képviselő fajtakör
Jelölt gén megközelítése
Genom pozíció függő stratégiák: pozicionális klónozás, deléciós vonalak
Összehasonlító genomikai stratégiák: modell növények
Mesterséges genetikai variáció (indukált mutációs populációk)
Gén csapdázás: T-DNS, transzpozon
Gén expressziós mintázatok elemzése:
Differential Display
DNS microarray, DNS chip
Gén azonosítás főbb módszerei

2012. október 3.
Mészáros Klára
Összehasonlító genomikai stratégiák
Ortológ lokusz: Különböző fajokban ugyanabból az ősi lokuszból
származó hasonló szekvenciájú és funkciójú géneket hordozó lokusz.
Paralóg lokusz: Ősi lokusz duplikációjából származó lokusz.
Szinténia: Két különböző fajban a gének és markerek hasonló sorrendje.
Mikroszinténia: Szekvencia szintű szinténia. A homológ
szekvenciájú gének hasonló sorrendje, szekvenciája és orientációja
a genomban.

2012. október 3.
Mészáros Klára
Gu et al. 2004 Plant Phys

2012. október 3.
Mészáros Klára
Stein et al. TAG(2007)114:823

2012. október 3.
Mészáros Klára
Cockram et al. JEXB(2007)58:1231

2012. október 3.
Mészáros Klára
Marker kapcsoltsági térképek
Két szülős térképező populációk
Széles genetikai bázist képviselő fajtakör
Jelölt gén megközelítése
Genom pozíció függő stratégiák: pozicionális klónozás, deléciós vonalak
Összehasonlító genomikai stratégiák: modell növények
Mesterséges genetikai variáció (indukált mutációs populációk)
Gén csapdázás: T-DNS, transzpozon
Gén expressziós mintázatok elemzése:
Differential Display
DNS microarray, DNS chip
Gén azonosítás főbb módszerei

2012. október 3.
Mészáros Klára
A mutáció az örökítő anyag spontán, maradandó
megváltozása, amelynek során új tulajdonság keletkezik.
A Hugo De Vries által a múlt század elején bevezetett fogalmat
ma szűkebb értelemben a génen belüli bázissorrend-változásra
vonatkoztatjuk.
Tágabb értelemben mutációnak tekinthető a kromoszóma-
szerkezetben, ill. a kromoszómák számában bekövetkezett
minden változás, ideértve a poliploidiát is.
Mutáció

2012. október 3.
Mészáros Klára
A génmutációk (pontmutációk) egy gén bázissorendjének változását
jelentik. Ennek hatása ritkán jelenik meg azonnal a fenotípusban, csak ha
domináns a mutáció.
Bázis csere (tranzicio, transzverzió)
Keret eltolódás (Frameshift)
Hatásuk: same sence azonos aminósav
missense aminósav csere
nonsense stop kodon kialakulása
Kromoszómamutáció:
A kromoszóma egyes részei változnak meg. Ez lehet szerkezeti változás, például a
kromoszómák törlődése (deléció), megfordulása (inverzió) kicserélődése
(transzlokáció), megduplázódása (duplikáció).
Másrészt lehet számbeli, melyen belül megkülönböztetünk poliploidiát
(kromoszómagarnitúra a normális egész számú többszöröse) és aneuploidiát (csak
egy kromoszómánál van eltérés).

Mutagenezis
kémiai (EMS, ENU, DEB, HNO2, …)
besugárzás (R, a-g, gyors neutron, UV)
inszerció [T-DNS és/v. (retro)transzpozon]
kimérikus (RNS-DNS) oligonukleotidok
géncsendesítés (antiszensz, RNSi)
véletlen,
azonosítás??
letalitás

mutáns populáció előállítása
HTP mutáns azonosítás
értékelés
Lépései
• (EMS) mutagenezis + genomika = HTP azonosítás
Szelekció a genotípus alapján (‘reverz’ genetika)
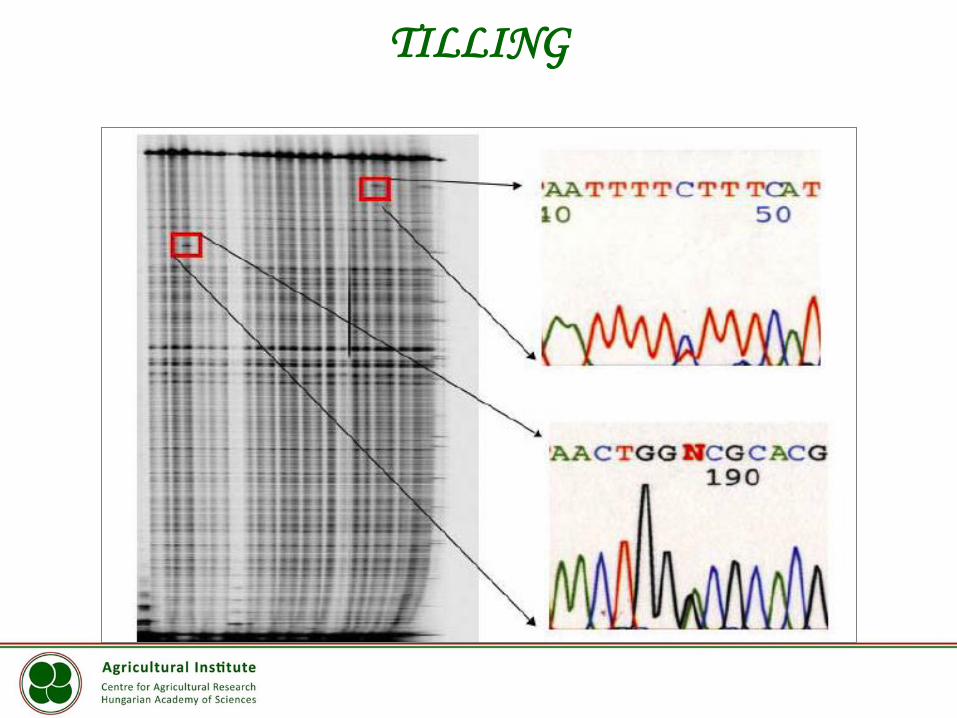
TILLING
(Targeting Induced Local Lesions IN Genomes)

TILLING – mutáns populáció előállítása

TILLING – mutáns populáció előállítása

TILLING Mutáció kimutatása

Heteroduplex formációk
TILLING – a módszer

IR 700 IR 800
A mutációk kimutatása
IR 700 IR 800
TILLING – a módszer

LI-COR 4300 DNA Analyzer
TILLING – a módszer

TILLING
IR 700 IR 800
Elválasztás
TILLING

ECOTILLING
TILLING a természetben létező ökotípusokkal vagy
populációkkal
és
TILLING mesterségesen fenntartott genetikai forrásokkal,
pl. fajta- és mutánsgyűjtemények, izogén vonalak (NIL),
hasadó és térképezési populációk (BSA), fajtajelöltek, ...

ecoTILLING

allél(sorozatok) azonosítása (funkc. genomika, SNP)
többszörös mutánsok azonosítása
HTP szelekció (MAS, GAS, SAS …)
pedigrévizsgálat
rokonsági körök igazolása (kukorica)
(fejlődés)rendszertan
genetikai térképezés (génizolálás)
TILLING – alkalmazások

Ajánlott irodalom
2012. október 3.
Mészáros Klára
Balázs E. és Duduts D.: Molekuláris növénybiológia
Dudits D., Heszky L.: Növényi biotechnológia és géntechnológia
Heszky L., Fésüs L., Hornok L.: Mezőgazdasági biotechnológia
B.C.Y. Collard1,4,∗, M.Z.Z. Jahufer2, J.B. Brouwer3 & E.C.K.
Pang (2005) An introduction to markers, quantitative trait
loci (QTL) mapping and
marker-assisted selection for crop improvement: The basic
concepts.
Euphytica 142: 169–196 DOI: 10.1007/s10681-005-1681-5

2012. október 3.
Mészáros Klára
Köszönöm a figyelmet!






![A biometrikus[1] azonosítás új irányaP/2010/1/Nagy.pdf · A biOMeTRikus AzOnOsíTás új iRányA 73 Napjainkban a személyes adatok védelméhez fűződő jognak egyre nagyobb](https://img.pdfslide.net/doc/110x75/5f7e21d5c1dc9658a0425ea2/a-biometrikus1-azonosts-j-irnya-p20101nagypdf-a-biometrikus-azonosts.jpg)






















