Embed Size (px)
Citation preview

The FASEB Journal • Research Communication
Germ-free C57BL/6J mice are resistant tohigh-fat-diet-induced insulin resistance and havealtered cholesterol metabolism
Sylvie Rabot,*,1 Mathieu Membrez,†,1 Aurelia Bruneau,* Philippe Gerard,*Taoufiq Harach,† Mireille Moser,‡ Frederic Raymond,‡ Robert Mansourian,‡
and Chieh J. Chou†,2
*Institut National de la Recherche Agronomique, UMR1319 Micalis, Jouy-en-Josas, France; and†Department of Nutrition and Health, ‡Department of Bioanalytical Science, Nestle Research Center,Lausanne, Switzerland
ABSTRACT Recent studies showed that germ-free(GF) mice are resistant to obesity when consuming ahigh-fat, high-carbohydrate Western diet. However, itremains unclear what mechanisms are involved in theantiobesity phenotype and whether GF mice developinsulin resistance and dyslipidemia with high-fat (HF)feeding. In the present study, we compared the meta-bolic consequences of HF feeding on GF and conven-tional (conv) C57BL/6J mice. GF mice consumed fewercalories, excreted more fecal lipids, and weighed sig-nificantly less than conv mice. GF/HF animals alsoshowed enhanced insulin sensitivity with improved glu-cose tolerance, reduced fasting and nonfasting insuline-mia, and increased phospho-Akt(Ser-473) in adipose tis-sue. In association with enhanced insulin sensitivity,GF/HF mice had reduced plasma TNF-� and totalserum amyloid A concentrations. Reduced hypercholes-terolemia, a moderate accretion of hepatic cholesterol,and an increase in fecal cholesterol excretion suggestan altered cholesterol metabolism in GF/HF mice.Pronounced nucleus SREBP2 proteins and up-regula-tion of cholesterol biosynthesis genes indicate thatenhanced cholesterol biosynthesis contributed to thecholesterol homeostasis in GF/HF mice. Our resultsdemonstrate that fewer calorie consumption and in-creased lipid excretion contributed to the obesity-resistant phenotype of GF/HF mice and reveal thatinsulin sensitivity and cholesterol metabolism are met-abolic targets influenced by the gut microbiota.—Rabot, S., Membrez, M., Bruneau, A., Gerard, P.,Harach, T., Moser, M., Raymond, F., Mansourian, R.,Chou. C. J. Germ-free C57BL/6J mice are resistant tohigh-fat-diet-induced insulin resistance and have al-tered cholesterol metabolism. FASEB J. 24, 4948–4959(2010). www.fasebj.org
Key Words: insulin sensitivity � obesity � inflammation
Since the reproduction of germ-free (GF) rats in anisolator was established, the ability to maintain andreproduce GF animals has allowed scientists to address
questions related to the gut microbiota. Recently, GFmice have been used to study the effects of the intesti-nal microbiota on obesity and metabolic energy bal-ance. Backhed et al. (1, 2) showed that GF C57BL/6Jmice gained less weight than conventional (conv) micewhen consuming a chow or a sugar and fat-rich Westerndiet, but the mechanisms explaining the obesity resis-tant phenotype of GF mice are not clear. Existing datashow that food intake and total fecal calorie weresimilar in GF and conv mice on the Western diet (2),but these results were observed in a small number of GFanimals. In addition, fecal caloric values determined bybomb calorimetry are not suitable for comparing en-ergy excretion of GF and conv mice because the energycontribution from bacteria mass and mucins to totalfecal calories is largely different between the 2 groups.Therefore, further studies are needed to examinewhether food intake and energy excretion in fecescontribute to obesity resistance of GF mice.
In association with an excessive weight gain, West-ern diet feeding affected the cecal microbiota byincreasing Firmicutes and reducing Bacteroidetes inconv C57BL/6J mice (3). In particular, propagationof Erysipelotrichi, a class of Firmicutes, was signifi-cantly pronounced in cecal microbiota of Westerndiet-associated animals (3, 4). Functionally, the West-ern diet-associated intestinal metagenome is charac-terized by enrichment of bacterial genes involved inthe transport and fermentation of simple sugars andhost glycans (3). It has been hypothesized that theability of intestinal bacteria to ferment dietary fiberscan lead to an increase in the production of shortchain fatty acids, which contributes to weight gainsobserved in obese mice (5). In addition to inducing
1 These authors contributed equally to this work.2 Correspondence: Nestle Research Center, Rte. du Jorat
57, Vers-chez-les-Blanc, CH-1000 Lausanne 26, Switzerland.E-mail: [email protected]
doi: 10.1096/fj.10-164921This article includes supplemental data. Please visit http://
www.fasebj.org to obtain this information.
4948 0892-6638/10/0024-4948 © FASEB

obesity, high-fat (HF) feeding also leads to insulinresistance in mice, but the role of the gut microbiotain diet-induced insulin resistance has not been fullyexplored. Recently, Larsen et al. (6) reported that thefecal microbiota composition differs between type 2diabetic subjects and healthy volunteers. In addition,a number of publications (7, 8) have demonstratedthat antibiotic administration improves oral glucosetolerance in ob/ob and HF diet-induced insulin resis-tant C57BL/6J mice. These data suggest that not onlya gut microbiota profile is associated with insulinresistance but that modulation or reduction of thegut microbiota can be a successful strategy in man-aging insulin resistance. Another consequence of HFfeeding is the development of dyslipidemia. In convC57BL/6J mice, chronic HF feeding induces theexpression of hepatic HMG-CoA reductase(HMGCR) mRNA (9, 10), increases liver HMGCRactivity, and enlarges cholesterol storage (9). How-ever, it is not clear whether GF mice would respondto long-term HF feeding in a similar way to conv miceby increasing their cholesterol biosynthesis.
In the present study, we examined the effects ofthe GF condition on the development of obesity,insulin resistance, and dyslipidemia in mice fed asterilized HF diet. Transcriptional profiling of he-patic genes was compared to reveal different meta-bolic adaptations of GF and conv mice to a HF diet.Results from our study suggest that lack of gutmicrobiota prevents diet-induced obesity by reducingfood intake and increasing fecal lipid outputs. Lackof commensal gut microbiota also affected the devel-opment of diet-induced insulin resistance and intro-duced multiple changes in cholesterol metabolism inC57BL/6J mice.
MATERIALS AND METHODS
Animal experimentation
Fourteen GF male C57BL/6J mice were reared from GFbreeding pairs in GF isolators at Institut National de laRecherche Agronomique in Micalis Germfree Rodent Fa-cilities (ANAXEM, Jouy-en-Josas, France). In parallel, 16conventional male C57BL/6J mice were ordered fromCharles River Laboratories (L’Arbresle, France). All micewere housed individually and received a sterile rodentchow diet (R03; SAFE, Augy, France) and water ad libitum.At 6 wk of age, a sterilized HF diet [D12492 with 60%energy from fat and 0.03% w/w cholesterol (ResearchDiets, New Brunswick, NJ, USA) sterilized with �-irradia-tion 25 kGy] was introduced for an additional 11 wk. BothGF and conv mice were kept in the same room to ensurecomparable conditions. Body weight and diet consumptionof mice were measured weekly. Spillage of food pellets onthe bottom of the cage was carefully collected to ensure thequality of food intake measurements. Mice were deprivedof food for 6 h before an oral glucose tolerance test(OGTT). At 2 h after completing the OGTT, all mice werekilled with an overdose of isoflurane (Aerrane; Baxter,Maurepas, France). White (epididymal, mesenteric, andretroperitoneal) adipose tissues, liver, total gut, and cecum
content were collected immediately after exsanguinationand flash-frozen in liquid nitrogen. Procedures were car-ried out in accordance with the European Guidelines forthe Care and Use of Laboratory Animals and with permis-sion 78-63 of the French Veterinary Services.
OGTT
The OGTT was performed after 6 h food deprivation at theend of HF diet feeding. Blood glucose concentrations wereanalyzed from tail vein samples using an Ascensia Elite XLglucometer (Bayer AG, Zurich, Switzerland), before (time 0min) and at 15, 30, 60, and 120 min after the animals weregiven a glucose solution at 2 g/kg body weight (BW) by oralgavage. Samples collected at 0, 15, and 120 min were put intochilled EDTA-coated tubes for insulin determination.
Measurements of plasma parameters
Blood taken from the vena cava at death was collected intochilled EDTA-coated tubes and centrifuged at 2000 rpm for10 min. Plasma was aliquoted and frozen at �80°C until it wasanalyzed. Plasma samples were analyzed for triglycerides(Roche Diagnostics, Basel, Switzerland), free fatty acids(Wako, Neuss, Germany), insulin (IBL, Hamburg, Germany),total cholesterol (Roche Diagnostics), HDL-cholesterol (Bio-vision Research, Mountain View, CA, USA), serum amyloid A(SAA; Invitrogen Corp., San Diego, CA, USA), and leptin(Linco Research, St. Charles, MO, USA) concentrations withcommercial kits. The plasma proinflammatory cytokinesIFN�, IL-10, IL-1�, IL-6, and TNF-� were measured by immu-noassay with electrochemiluminescence detection (MesoScale Discovery, Gaithersburg, MD, USA) according to themanufacturer’s instructions.
Measurement of fecal and liver lipids
Fecal pellets from individual GF/HF mice were pooled tohave sufficient material for analysis. For conv/HF and conv/chow mice, each pool consisted of fecal pellets from 12individuals. Lyophilized feces (150 mg) was used for lipidextraction according to Hara and Radin (11). Total choles-terol (Roche Diagnostics), triglyceride (Roche Diagnostics),and free fatty acid (Wako) concentrations were quantifiedusing commercial kits. For triglyceride measurements, lipidswere first hydrolyzed in a basic solution (0.5 N KOH inethanol). For the determination of liver triglyceride andcholesterol concentrations, 200 mg frozen liver was used toextract lipids according to Folch et al. (12). Total cholesteroland triglycerides were quantified as above. Results were thennormalized to the weight of the sample.
Measurement of liver glycogen
Frozen liver (50–100 mg) was incubated in 30% KOH solu-tion for 15–30 min in a boiling water bath. After homogeni-zation, glycogen was precipitated with 96% ethanol, followedby centrifugation at 5000 rpm for 10 min. Pellets wereresuspended in 1 ml distilled water for the amyloglucosidasedigestion. Samples (200 �l) were incubated in 1.8 ml oflyophilized amyloglucosidase dissolved in acetate buffer (0.2M, pH 4.8 at a final concentration of 10 U/ml) for 2 h at40°C. The resulting glucose solution was then measured witha Quantichrom glucose assay kit (BioAssay Systems, Hayward,CA, USA). Results were then normalized to the weight of thesample.
4949EFFECTS OF HF FEEDING ON GF MICE

Western blot
Frozen epididymal adipose tissues were homogenized intubes containing Lysing matrix D (MP Biomedicals,Illkirch, France) and an ice-cold buffer A (10 mM KHEPES,pH 7.9; 10 mM KCl; 1.5 mM MgCl2; 0.5 mM DTT; 1 mMPMSF; and protease inhibitor cocktail tablets; Roche). Thehomogenates were centrifuged at 14,000 rpm for 10 min at4°C before the protein concentration was measured. Thepurified protein fractions were stored at �80°C beforeWestern blotting. Frozen livers were pulverized in liquidnitrogen and incubated in buffer A for 20 min. Thesamples were homogenized with a Polytron PT1300D (Ki-nematica AG, Lucerne, Switzerland) for 10 s at mediumspeed and centrifuged at 4000 rpm for 3 min at 4°C. Thesupernatant containing the cytosolic fraction was kept in aprechilled tube. The pellet was resuspended in ice-coldbuffer B (20 mM KHEPES, pH 7.9; 400 mM NaCl; 1.5 mMMgCl2; 0.5 mM DTT; 0.2 mM EDTA; 1 mM PMSF; andprotease inhibitor cocktail tablets) and was gently rotatedfor 20 min at 4°C. The samples were then centrifuged at14,000 rpm for 10 min at 4°C. Total protein concentrationof each sample was determined using a BCA protein assaykit (Pierce, Rockford, IL, USA). For Western blot analysis,50 �g of protein was loaded into a 12-well InvitrogenNuPAGE Novex 10% Bis-Tris gel for separation by electro-phoresis and then transferred to a nitrocellulose mem-brane as indicated in the manufacturer’s instructions(Invitrogen). Primary antibodies, rabbit polyclonal anti-SREBP2 (NB100-74543; Novus Biological, Littleton, CO,USA), rabbit polyclonal anti-�-actin (Invitrogen), and rab-bit polyclonal anti-Akt and anti-phospho-Akt (Ser-473; CellSignaling Technology, Inc., Danvers, MA, USA) were incu-bated at 4°C overnight, followed by secondary horseradishperioxidase-anti-rabbit antibody (Invitrogen) incubation.The blots were visualized with BM chemiluminescenceWestern blot kit (Roche).
Gene expression analysis by microarray
Liver samples (10 mg) were homogenized in lysis buffer,using a FastPrep instrument, in lysing tubes containing ce-ramic beads (MP Biomedicals, Irvine, CA, USA). Total RNAwas extracted and purified with the RNAdvance tissue kit(Agencourt, Beverly, MA, USA) using an automated proce-dure on a robotic station. The quality of RNA samples waschecked by using the Agilent 2100 Bioanalyser (RNA integritynumbers �8 for high quality; Agilent Technologies, SantaClara, CA, USA). All cRNA targets were synthesized, labeled,and purified according to the Illumina TotalPrep RNA am-plification protocol (Applied Biosystems/Ambion, Austin,TX, USA), using an automated procedure that has beenadapted to the Illumina procedure. Then, 15 �l of eachhybridization mix was dispensed on the arrays. After over-night hybridization (16 h, 58°C), the arrays were washed toremove nonhybridized material and stained with Streptavi-din-Cy3, which binds with biotin. All samples were analyzedwith MouseRef-8 v1.1 Expression BeadChips (Illumina, SanDiego, CA, USA). Scanning was performed using a BeadArrayReader (Illumina). Absolute signal intensities were extractedand summarized in the BeadStudio software (Illumina). Datawere quantile normalized, and log2 transformations weremade using Partek software (St. Louis, MO, USA). Microarraydata files have been deposited in GEO Omnibus (http://www.ncbi.nlm.nih.gov/geo/) with the accession numberGSE19038. Quality control of the data was performed on allsamples with a Pearson correlation matrix and a principalcomponent analysis on all probes. To assess which probes
were differentially expressed between the GF/HF and theconv/HF groups, 1-way ANOVA was performed, followed by aglobal error assessment (GEA; ref. 13). The GEA results in arobust MSE that replaces the current MSE from the classicalANOVA; a new F statistic is recalculated, and a robust P valueis derived.
A list of probes was then selected, based on their P value(cutoff P�0.001) and their fold change, taken to be �1.3 or ��1.3between the 2 groups. Selected probes, along with theirdifferential expression value, were loaded into the IngenuityPathways Analysis software (IPA; Ingenuity Systems; http://www.ingenuity.com) for annotation, redundancy checks, andnetwork and pathway analysis, with the following criteria:reference set, MouseRef-8 v1.1; direct and indirect relation-ships included. Semantic relationship analysis with IPA soft-ware generated significant networks of genes that are well-characterized metabolic and cell-signaling pathways asdocumented in published articles and books and the KEGGLigand database, with the satisfaction of the right-tailedFisher’s exact test.
Gene expression analysis by quantitative PCR
Reverse transcription was performed on 1.5 �g total RNA usingthe first-strand cDNA synthesis kit for real-time PCR (AMV;Roche Biomedical) with oligo d(T)15 as primer. Real-time PCRanalyses were performed in a fluorescent temperature cycler(GeneAmp PCR 5700 Sequence Detection System; AppliedBiosystems). Values were normalized to GAPDH expression.The effects of treatments on gene expression were evaluated bycalculating the relative expression level of each gene of interest)(GOI) as follows: 2�(Ct GOI � Ct GAPDH), using the mean rawcycle-threshold (Ct) values.
Statistical analyses of biological data
All biological parameters were tested by Wilcoxon tests dueto the presence of outliers or abnormally distributed data.Results are presented as medians � se. The se was com-puted using a robust sd based on Sn of Rousseeuw (14). Alltests were performed 2-sided. Analysis was performed withR 2.6.1 (15).
RESULTS
GF mice are resistant to diet-induced obesity
All male GF C57BL/6J mice were reared from GFbreeding pairs residing in isolators. At 6 wk old, a sterileHF diet was provided ad libitum to GF (GF/HF) andconv (conv/HF) mice for 11 wk. The GF status of micewas monitored weekly by culturing fresh fecal samplesin aerobic and anaerobic conditions; none of the fecalsamples from GF mice contained replicating bacteria(Supplemental Table S1). BW of GF and conv mice onthe HF diet are shown in Fig. 1A and Table 1. Incontrast to the age-matched conv/HF male mice whogained 15.0 � 1.1 g BW in 11 wk, GF/HF mice onlygained 5.3 � 0.9 g (P�0.001). Epididymal (3.5-fold),mesenteric (3.6-fold), and retroperitoneal (4.7-fold) fatpads also weighed less in GF/HF than in conv/HF mice(Table 1). As expected, plasma leptin concentrationswere lower in GF/HF mice (Table 2). Cecum weight
4950 Vol. 24 December 2010 RABOT ET AL.The FASEB Journal � www.fasebj.org

was 11-fold greater in GF/HF than in conv/HF mice(Table 1), which is one of the hallmark features of GFrodents.
GF mice have reduced food intake and increasedfecal lipid excretion
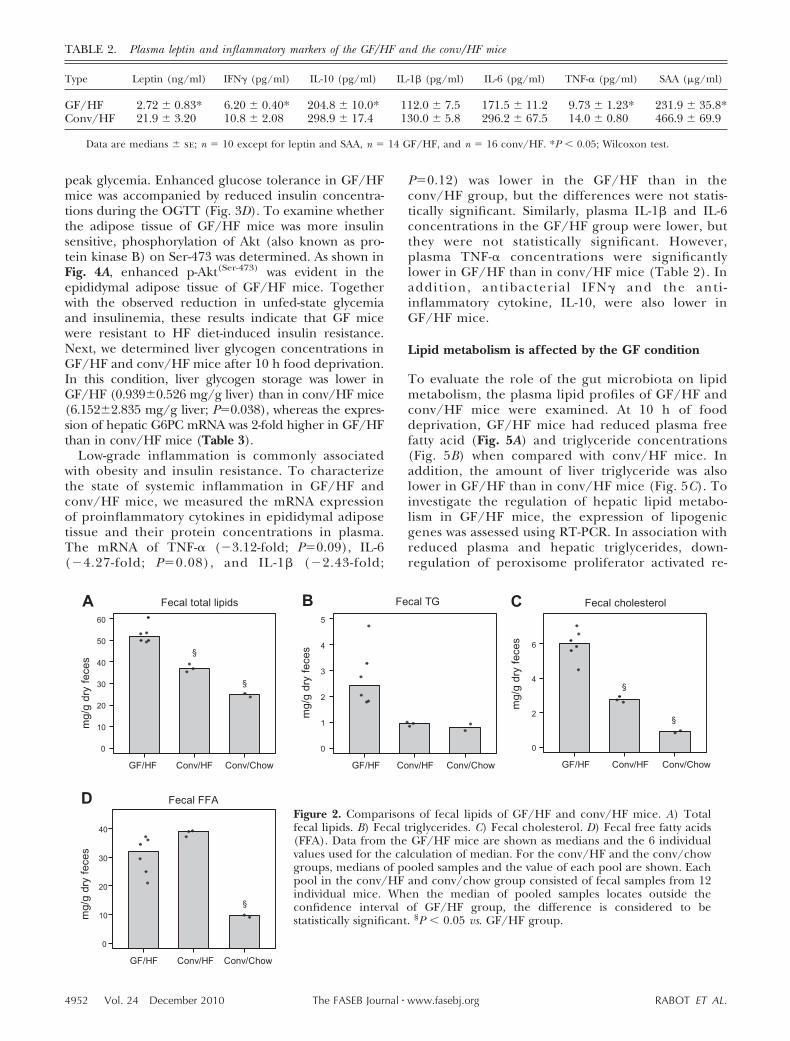
Food intake of individual mice was measured weekly.The cumulated food intakes of GF/HF and conv/HFmice are illustrated in Fig. 1B. GF mice ate 16% lessfood than conv controls in 11 wk, and the differencewas statistically significant (168.0�4.2 vs. 200.5�3.7 g;P�0.001). When food intake of each mouse was nor-malized to its BW or metabolic mass (BW0.75), GF micestill consumed less of the HF diet than their convcounterparts (Fig. 1C). In addition, food efficiency wasreduced by 54% in GF/HF mice (Fig. 1D), indicatingthat these mice are inefficient in converting dietaryenergy into body mass. To examine the possible rea-sons for the reduction in food efficiency, fecal lipidswere measured in GF/HF and conv/HF mice. Fecalsamples collected from conv C57BL/6J mice fed astandard chow diet were used as a reference. Feces ofGF/HF mice contained 40% more total lipids than thatof conv/HF mice (Fig. 2A). When the lipid composi-tion of the feces was compared, a trend for hightriglycerides (Fig. 2B) and significantly more choles-
terol (Fig. 2C) but not fecal free fatty acids (Fig. 2D) wasobserved in GF/HF group. Our data indicate that GFmice are resistant to HF-diet-induced obesity and thatreduced food intake together with increased fecal lipidoutput contributes to the lean phenotype of GF/HFmice.
GF mice are resistant to diet-induced insulinresistance
C57BL/6J mice develop obesity and insulin resistancewhen fed an HF diet, and insulin resistance of DIOmice can be improved by reducing the gut microbiotausing antibiotics (8). To study whether the gut micro-biota is required for diet-induced insulin resistance,parameters indicating insulin sensitivity were measuredin GF/HF and conv/HF mice. When compared withGF/HF mice, elevation of blood glucose in conv/HFmice was apparent after 6 h of food deprivation but notin a fed state (Fig. 3A). Plasma insulin concentrationswere lower in GF/HF mice than in conv/HF controls inboth the 6 h unfed and fed states (Fig. 3B). To furtherexamine the insulin sensitivity of mice, OGTT wereconducted at the end of the study. As illustrated in Fig.3C, conv/HF mice developed glucose intolerance asexpected, whereas GF/HF mice showed improvedblood glucose clearance with reduced baseline and
TABLE 1. Body weight and tissue weights of the GF/HF and the conv/HF mice
Type Body weight Cecum Epi Mes Rp Liver Pancreas
GF/HF 26.75 � 1.25* 1.52 � 0.08* 0.48 � 0.10* 0.13 � 0.03* 0.11 � 0.03* 0.79 � 0.03* 0.21 � 0.01*Conv/HF 32.75 � 0.66 0.14 � 0.01 1.68 � 0.21 0.47 � 0.07 0.52 � 0.06 1.02 � 0.06 0.28 � 0.01
Data are median � se weight (g); n 14 GF/HF and n 16 conv/HF. Epi, epididymal fat pad; Mes, mesenteric fat pad; Rp, retroperitonealfat pad. *P � 0.05; Wilcoxon test.
Figure 1. Comparisons between GF/HF and conv/HF mice. A) BW. B) Cumulativefood intake. C) Normalized food intake (FI; based on BW or BW0.75). D) Foodefficiency. Data are medians � se; n 14 for GF/HF, n 16 for conv/HF mice.*P � 0.05; Wilcoxon test
4951EFFECTS OF HF FEEDING ON GF MICE
peak glycemia. Enhanced glucose tolerance in GF/HFmice was accompanied by reduced insulin concentra-tions during the OGTT (Fig. 3D). To examine whetherthe adipose tissue of GF/HF mice was more insulinsensitive, phosphorylation of Akt (also known as pro-tein kinase B) on Ser-473 was determined. As shown inFig. 4A, enhanced p-Akt(Ser-473) was evident in theepididymal adipose tissue of GF/HF mice. Togetherwith the observed reduction in unfed-state glycemiaand insulinemia, these results indicate that GF micewere resistant to HF diet-induced insulin resistance.Next, we determined liver glycogen concentrations inGF/HF and conv/HF mice after 10 h food deprivation.In this condition, liver glycogen storage was lower inGF/HF (0.939�0.526 mg/g liver) than in conv/HF mice(6.152�2.835 mg/g liver; P0.038), whereas the expres-sion of hepatic G6PC mRNA was 2-fold higher in GF/HFthan in conv/HF mice (Table 3).
Low-grade inflammation is commonly associatedwith obesity and insulin resistance. To characterizethe state of systemic inflammation in GF/HF andconv/HF mice, we measured the mRNA expressionof proinflammatory cytokines in epididymal adiposetissue and their protein concentrations in plasma.The mRNA of TNF-� (�3.12-fold; P0.09), IL-6(�4.27-fold; P0.08), and IL-1� (�2.43-fold;
P0.12) was lower in the GF/HF than in theconv/HF group, but the differences were not statis-tically significant. Similarly, plasma IL-1� and IL-6concentrations in the GF/HF group were lower, butthey were not statistically significant. However,plasma TNF-� concentrations were significantlylower in GF/HF than in conv/HF mice (Table 2). Inaddit ion, antibacterial IFN� and the anti -inflammatory cytokine, IL-10, were also lower inGF/HF mice.
Lipid metabolism is affected by the GF condition
To evaluate the role of the gut microbiota on lipidmetabolism, the plasma lipid profiles of GF/HF andconv/HF mice were examined. At 10 h of fooddeprivation, GF/HF mice had reduced plasma freefatty acid (Fig. 5A) and triglyceride concentrations(Fig. 5B) when compared with conv/HF mice. Inaddition, the amount of liver triglyceride was alsolower in GF/HF than in conv/HF mice (Fig. 5C). Toinvestigate the regulation of hepatic lipid metabo-lism in GF/HF mice, the expression of lipogenicgenes was assessed using RT-PCR. In association withreduced plasma and hepatic triglycerides, down-regulation of peroxisome proliferator activated re-
Figure 2. Comparisons of fecal lipids of GF/HF and conv/HF mice. A) Totalfecal lipids. B) Fecal triglycerides. C) Fecal cholesterol. D) Fecal free fatty acids(FFA). Data from the GF/HF mice are shown as medians and the 6 individualvalues used for the calculation of median. For the conv/HF and the conv/chowgroups, medians of pooled samples and the value of each pool are shown. Eachpool in the conv/HF and conv/chow group consisted of fecal samples from 12individual mice. When the median of pooled samples locates outside theconfidence interval of GF/HF group, the difference is considered to bestatistically significant. §P � 0.05 vs. GF/HF group.
TABLE 2. Plasma leptin and inflammatory markers of the GF/HF and the conv/HF mice
Type Leptin (ng/ml) IFN� (pg/ml) IL-10 (pg/ml) IL-1� (pg/ml) IL-6 (pg/ml) TNF-� (pg/ml) SAA (�g/ml)
GF/HF 2.72 � 0.83* 6.20 � 0.40* 204.8 � 10.0* 112.0 � 7.5 171.5 � 11.2 9.73 � 1.23* 231.9 � 35.8*Conv/HF 21.9 � 3.20 10.8 � 2.08 298.9 � 17.4 130.0 � 5.8 296.2 � 67.5 14.0 � 0.80 466.9 � 69.9
Data are medians � se; n 10 except for leptin and SAA, n 14 GF/HF, and n 16 conv/HF. *P � 0.05; Wilcoxon test.
4952 Vol. 24 December 2010 RABOT ET AL.The FASEB Journal � www.fasebj.org

ceptor-� (PPAR�) and stearoyl-CoA desaturase 1(SCD1) mRNA was found in the liver of GF/HF mice(Table 3).
As illustrated in Fig. 5D, E, GF/HF mice had reducedplasma total cholesterol and HDL cholesterol concen-trations when compared with the conv/HF controls.However, a moderate elevation in liver cholesterol (Fig.5F) and a 2.8-fold increase in hepatic HMGCR mRNAexpression (Table 3) were found in GF/HF mice. Tostudy whether SREBP2 was involved in the transcrip-tional control of the HMGCR gene, we measured theprotein quantity of SREBP2 in different subcellular
fractions of the liver by Western blots. As illustrated inFig. 4B, nuclear SREBP2 was higher in liver of GF/HFthan of conv/HF mice, whereas cytosolic SREBP2 pro-teins were similar in both groups. Our data indicatethat the accumulation of liver cholesterol in GF/HFmice is associated with an increased ability to synthesizecholesterol.
Comparison of global gene expression in the liver ofGF/HF and conv/HF mice
To reveal the effect of the GF condition on livermetabolism, we performed a transcriptomic analysiswith the livers of GF/HF (n14) and conv/HF(n16) mice. A total of 236 differentially regulatedunique transcripts were identified based on a cutoffof P � 0.001 and �1.3-fold change; 121 transcriptswere up-regulated and 115 transcripts were down-regulated in the liver of GF/HF mice. The mostoverrepresented canonical pathways and the propor-tions of differentially regulated genes in these path-ways are shown in Fig. 6A. Many of the most overrep-resented canonical pathways are defensivemechanisms that respond to environmental stimula-tions. For example, pathways related to the metabo-lism of xenobiotics, such as PXR/RXR activation andcytochrome P450 pathways, as well as pathways thatreact against bacterial components, such as LPS-1/IL-1-mediated inhibition of RXR function, were over-represented in the liver of GF/HF mice. Comple-ment factor D (CFD, �3.33-fold), hepcidinantimicrobial peptide 1 (HAMP, �2.32-fold), serumamyloid A1 (SAA1, �2.06-fold) and lipocalin 2(LCN2, �2.06-fold) were among the most down-regulated molecules in GF/HF mice (SupplementalTable S2), further indicating that many componentsof the innate immune system in mice were reduced in
Figure 3. Insulin sensitivity of GF/HF and conv/HF mice. A, B) Bloodglucose (A) and plasma insulin (B) at 6 h food deprivation (fasting) andfood availability (nonfasting) in GF/HF and conv/HF mice were compared.C, D) Results of blood glucose (C) and the glucose area-under-the-curve(AUC; inset) and plasma insulin (D) and the insulin AUC (inset) duringOGTTs in GF/HF and conv/HF mice are shown. All mice were deprived offood for 6 h before the OGTT. Data are medians � se; n 14 for GF/HF,n 16 for conv/HF mice. For nonfasting blood glucose and plasma insulinlevels, n 6. *P � 0.05; Wilcoxon test.
Figure 4. A) Total Akt and phosphorylated Akt on Ser-473 inthe epididymal adipose tissue from 3 GF/HF and 3 conv/HFindividual mice were measured by Western blot. B) SREBP2protein in nuclei, cytosol, and total extract (TE) of the frozenliver from 2 individual animals in each group were measuredwith Western blot, and expression of �-actin protein served asa loading control.
4953EFFECTS OF HF FEEDING ON GF MICE

a GF condition. Expression of HAMP, SAA1, andLCN2 is a part of acute-phase responses. Recent datasuggested that a high amount of SAA proteins causesinsulin resistance in 3T3-L1 adipocytes (16). In thepresent study, plasma total SAA concentrations werelower in GF/HF than in conv/HF mice (Table 2) and
the data is in accordance with improved glucosetolerance in GF/HF mice (Fig. 3C).
Biosynthesis of steroids is another overrepresentedcanonical pathway. Unlike other pathways, all differen-tially regulated genes, such as HMG-CoA synthase 1(HMGCS1, 2.15-fold), HMGCR (1.47-fold), farnesyl
TABLE 3. Analysis of hepatic gene expression by RT-PCR
Gene GF/HF Conv/HF Primer sequences
G6PC 0.486 � 0.059* 0.234 � 0.035 For 5-GAGGAAAGAAAAAGCCAACGTATG-3Rev 5-CCCAGAATCCCAACCACAAG-3
HMGCR 0.090 � 0.014* 0.032 � 0.003 For 5-TGTGGCCAGGAGTTTGGTGACTGA-3Rev 5-TAAGATTCAACAACTCTGCTGACC-3
PPAR� 0.002 � 0.000* 0.004 � 0.001 For 5-CATGACCAGGGAGTTCCTCAA-3Rev 5-CTTAGGCTCCATAAAGTCACCAAAG-3
SCD1 0.015 � 0.002* 0.031 � 0.002 For 5-GACCAGTCAATGTGCAAGACTACCT-3Rev 5-ATCTCATCCAGGGAAGGGAAA-3
ABCG5 0.175 � 0.017* 0.100 � 0.012 For 5-CGTGGCGGACCAAATGA-3Rev 5-CGCTCGCCACTGGAAATT-3
ABCG8 0.298 � 0.037* 0.173 � 0.022 For 5-GGAGCACTGTGCCTACGTCAT-3Rev 5-CGCAGGTTTGTCAGCCAGTA-3
GAPDH For 5-ATGTGTCCGTCGTGGATCTGA-3Rev 5-CCTGCTTCACCACCTTCTTGA-3
Data are median � se expression (2���Ct); n 6 for G6Pc, PPAR�, and HMGCR or n 12 for SCD1, ABCG5, and ABCG8. Samples werecollected after 10 h food deprivation. Expression of target genes was normalized to the level of GAPDH. For, forward; Rev. reverse. *P � 0.05;Wilcoxon test.
Figure 5. Comparisons of plasma and liver lipids in GF/HF and conv/HF mice. All samples were collected after 10 h fooddeprivation. A) Plasma free fatty acids. B) Plasma triglycerides. C) Liver TG. D) Plasma total cholesterol. E) Plasma HDLcholesterol. F) Liver cholesterol. Data are medians � se. For plasma samples, n 10 or 11 for the GF/HF and n 9 or 11 forthe conv/HF mice. For liver samples, n 8. *P � 0.05; Wilcoxon test.
4954 Vol. 24 December 2010 RABOT ET AL.The FASEB Journal � www.fasebj.org

diphosphate synthase (FDPS, 1.94-fold), squalene ep-oxydase (SQLE, 2.77-fold), and 7-dehydrocholesterolreductase (DHCR7, 1.47-fold), were up-regulated inGF/HF mice (Supplemental Table S2), and the in-
crease in HMGCR mRNA expression was confirmedwith RT-PCR (Table 3). Figure 6B illustrates the coregu-lation of SREBP2 and its target genes based on knownsemantic relationships. Synergistic up-regulation of
Figure 6. Global gene expression analysis from GF/HF and conv/HF livers. A) Compared with data from the conv/HFgroup, the 10 most overrepresented canonical pathways in liver from GF/HF mice are illustrated. Total of number of genesin each respective pathway is indicated in parentheses. Up-regulation and down-regulation of genes in each pathway areindicated by red and green bars, respectively. Length of the bar in each direction indicates the proportion of thedifferentially regulated genes in each specific pathway. Numbers located inside the red or green areas show the numberof differentially regulated genes in each respective direction. B) Differentially regulated hepatic genes shown in the GF/HFto conv/HF comparison and their semantic associations to nuclear SREBP2. The darker the red color, the greater the foldchange of gene expression. Data were analyzed and the networks were generated through the use of IPA (IngenuitySystems; www.ingenuity.com).
4955EFFECTS OF HF FEEDING ON GF MICE

SREBP2 mRNA, SREBP2 protein in the nuclei, and theSREBP2 target genes suggests that SREBP2 is an impor-tant transcriptional factor that controls cholesterolmetabolism in a GF condition. Down-regulation oflipogenic genes, including PPAR� (�1.71-fold) andSCD1 (�1.73-fold), in the liver of GF/HF mice was alsoobserved in the transcriptomic analysis, and theresults were confirmed by RT-PCR (Table 3). Unex-pectedly, ATP-binding cassette transporter G5 andG8, the 2 genes involved in reverse cholesteroltransport, were up-regulated in GH/HF mice, 1.223-fold and 1.338-fold, respectively. Since the differenceis near the level of detection limited by the microar-ray analysis, we then examined the expression ofboth genes by RT-PCR. As shown in Table 3, up-regulation of hepatic ABCG5 and ABCG8 in liver ofGF/HF mice was confirmed.
DISCUSSION
In the present study, we examined the role of the gutmicrobiota in diet-induced obesity and insulin resis-tance. Our results show that the absence of the gutmicrobiota has profound effects on the development ofobesity, glucose regulation, and cholesterol metabolismhomeostasis.
Regulation of BW in GR mice fed an HF diet
Previously, Backhed et al. (2) reported that GF micegained less weight on a Western diet than conv mice.Our results agree with their findings and demonstratethat GF C57BL/6J mice are resistant to the develop-ment of obesity when consuming an HF diet. In con-trast, Fleissner et al. (17) showed that GF C3H micegained more weight than conv mice when mice con-sumed an HF diet. The discrepancy of the weight gainbetween the studies is probably due to the strain ofmice used in each study. Future studies comparing theobesity development of different strains of mice areneeded to confirm this hypothesis. In the present study,we observed a significant reduction in cumulative foodintake by GF mice during the 11-wk feeding period.After normalization to BW or metabolic mass, foodconsumption was still lower in GF/HF than conv/HFmice. In addition, increased energy excretion in GFmice can explain much of the reduced food efficiency.Although previous data demonstrate that total fecalcalories, measured by a bomb calorimeter, are similarin GF and conv mice (2), we observed that total fecallipids were higher in GF/HF than in conv/HF mice.The reason fecal lipids instead of total fecal calorieswere measured is because the fecal composition is verydifferent between GF and conv mice. In GF animals,feces contain undigested plant polysaccharides andendogenously produced mucins, which are normallydigested and fermented by the bacteria residing in thedistal intestine of conv mice, while bacteria mass, anabsent component in GF feces, represents a large
portion of the calorie content of conv feces. Because ofthe intrinsic difference in fecal composition betweenGF and conv mice, we decided to examine the fecallipid composition to estimate the contribution of en-ergy excretion to energy balance in GF and conv mice.In our experiment, reduced energy intake and en-hanced lipid excretion are the two contributing factorsthat explain the lack of weight gain in GF/HF mice.
Mechanisms of improved insulin sensitivity in GF/HFmice
Here we demonstrate that GF mice are resistant toHF-diet-induced insulin resistance. While reduction inBW and body fat can significantly contribute to enhanc-ing insulin sensitivity in GF/HF mice, absence of theintestinal microbiota may be an independent contrib-utor to whole-body insulin sensitivity. This argument issupported by a recent publication (8) demonstratingthat antibiotic treatment reduces the number of cecalbacteria and normalizes glucose tolerance in ob/ob andDIO. However, the molecular mechanisms behind theimproved oral glucose tolerance, as well as reducedglycemia and insulinemia, are unknown. Backhed et al.(2) have shown that phosphorylation of AMPK is ele-vated in skeletal muscle of GF mice. Since activation ofAMPK has been shown to increase insulin-stimulatedglucose transport in ex vivo epitrochlearis muscles (18),enhanced AMPK activity in muscle can positively influ-ence glucose clearance in GF/HF mice during anOGTT.
Alternatively, many groups, including our own,have suggested that lipopolysaccharides (LPS) ab-sorbed from gram-negative bacteria residing in theintestine may induce insulin resistance in the host (7,8, 19). Abrogation of intestinal bacteria, as in the GFcondition, should minimize the exposure of GF miceto LPS and avoid LPS-induced insulin resistance.Although the LPS concentration was not measured inthe present study, the confirmation of GF status andlow plasma TNF-� concentrations let us assume thatthere was little or no LPS in GF/HF mice. Whenconv/HF mice are considered, infiltration of macro-phages into the adipose tissue results in inflamma-tory responses, with excessive production of proin-flammatory cytokines such as TNF-� (20). TNF-� hasbeen shown to induce insulin resistance in obeseZucker rats (21), and its concentration in plasmapositively correlates with obesity and insulin resis-tance in animal models (22). More important, thefact that insulin sensitivity can be normalized byneutralization of TNF-� receptors in obese Zuckerrats (22) and by deletion of TNF-� in mice (23, 24)clearly demonstrates the causal role of TNF-� ininsulin resistance. Plasma TNF-� concentrations weresignificantly less in GF/HF than conv/HF mice.Therefore, it is plausible that the ability of GF mice toresist diet-induced insulin resistance is partially dueto low plasma TNF-� concentrations.
Another hallmark feature of insulin resistance is
4956 Vol. 24 December 2010 RABOT ET AL.The FASEB Journal � www.fasebj.org

the inability of insulin to suppress lipolysis in adipo-cytes. Consequentially, elevated plasma free fatty acidconcentrations lead to lipotoxicity and insulin resis-tance in the liver, skeletal muscle, and pancreas (25).Recently, Berggreen et al. (26) showed that insulin-mediated suppression of lipolysis depends on func-tional Akt. Inhibition of Akt with a pharmacologicalinhibitor, Akti, decreased insulin-stimulated phos-phorylation of Akt on Thr-308 and Ser-473 andattenuated insulin-mediated antilipolytic effects inisolated rat adipocytes and 3T3-L1 cells (26). In thepresent study, decreased phosphorylation of Akt onSer-473 in adipose tissue and elevated plasma freefatty acid concentrations were found in conv/HFmice, suggesting that their adipose tissue is resistantto elevated basal insulin concentrations after 10 hfood deprivation when compared with GF/HF micewho are sensitive to lower concentrations of insulinunder the same condition.
Changes of cholesterol metabolism in GF/HF mice
In the present study, moderate elevations in livercholesterol and marginally reduced plasma total andHDL cholesterols were observed in GF/HF mice, whichis in agreement with previous data by Wostmann andWiech (27). The moderate increase of liver cholesterolin GF/HF mice can be partially explained by increasedcholesterol biosynthesis, as supported by several keyobservations: overexpression of SREBP2 protein in thenuclei of GF/HF livers, and up-regulation of HMGCS1,HMGCR, FDPS, SQLE, and DHCR7 mRNAs in GF/HFlivers. Because the expression of HMGCR mRNA ispositively correlated with its enzymatic activity in theliver of C57BL/6J mice (9, 28), it is highly possible thatcholesterol biosynthesis is increased in GF/HF mice. Inaddition to increased endogenous cholesterol produc-tion, increased dietary cholesterol absorption and en-hanced cholesterol uptake by hepatocytes can contrib-ute to elevated hepatic cholesterol storage. However,the influence of these 2 mechanisms is unlikely, be-cause the cholesterol content of the HF diet was verylow (0.03% w/w), and GF/HF mice consumed less foodin the present study. Also, mRNA expression of theLDL receptor was similar in GF/HF and conv/HF mice,suggesting that mechanisms other than cholesteroluptake are involved in the homeostasis of cholesterolmetabolism in GF/HF mice.
We unexpectedly observed a 2-fold increase infecal cholesterol in GF/HF mice. This observationcan be explained by a lack of cholesterol absorptionin the intestine or an increase in cholesterol excre-tion by the liver. The first possibility is unlikelybecause it contradicts published data demonstratingthat GF animals absorb dietary cholesterol moreefficiently than conv controls (29). In support of thesecond possibility, we found that hepatic ABCG5 andABCG8 expression was elevated in the GF/HF liver.ABCG5 and ABCG8 are membrane transporters thatare expressed primarily in hepatocytes and entero-
cytes in the proximal small intestine of humans andmice (30). Functionally, these transporters are im-portant for the excretion of cholesterol in feces viareverse cholesterol transport. Up-regulation ofABCG5/G8 via LXR-mediated activation is requiredfor transporting cholesterols from lipid laden macro-phages to HDL particles and then from hepatocytesto the gallbladder in the liver (31). Overexpressionof human ABCG5/G8 in mice results in a 3-foldincrease in fecal cholesterols in males when com-pared with gender matched wild-type littermates(32). In this transgenic mouse model, an increase inhepatic cholesterol biosynthesis and biliary choles-terol secretion has also been reported (33). Interest-ingly, changes of cholesterol metabolism in GF/HFmice are remarkably similar to those of ABCG5/G8transgenic mice. In both models, a slight increase inliver cholesterol, a mild reduction in plasma choles-terol, and an up-regulation of cholesterol biosynthe-sis were observed (33). Yu et al. (33) suggested that inABCG5/G8 transgenic mice the accelerated loss ofcholesterol from the body is compensated for by anincrease in hepatic cholesterol biosynthesis and thatthe balance of synthesis and secretion of cholesterolresults in mild differences in the plasma and livercholesterols when compared with wild-type controls.
The reason for the elevated ABCG5/G8 expressionin GF/HF mice is not clear. Diminished exposure tointestinal bacteria and bacterial compounds couldhave positively affected ABCG5/G8 expression. Re-cently, endotoxemia has been shown to suppress theexpression of ABCG5/G8 in the liver and to decreasefecal cholesterol excretion (33–35). Under conven-tional conditions, plasma LPS concentrations areincreased in mice fed an HF diet (36), which sup-ports our observation that ABCG5/G8 mRNA levelswere suppressed in the liver of conv/HF mice. In GFconditions, mice are exposed to minimal amounts ofbacterial LPS; hence, the mechanism by which LPSinhibits the expression ABCG5/G8 genes is notstrongly induced. As a consequence, high hepaticABCG5/G8 expression was observed, which couldexplain the very large fecal cholesterol excretionfound in the feces of GF/HF mice.
Another possible explanation for elevated fecal cho-lesterol is the lack of bacterial activity in GF conditions.Under conventional conditions, cholesterol is con-verted to coprostanol by the gut microbiota (37).Species in the genus Eubacterium isolated from thececum of rats (38) and Bacteroides sp strain D8 that isclosely related to Bacteroides dorei of human origin (39)have been shown to contain cholesterol reducing activ-ity. In the absence of intestinal bacteria and bacterialactivity in GF mice, the conversion of cholesterol tocoprostanol would not occur and the composition offecal steroids would be different. To better understandthe role of gut microbiota in the homeostasis of cho-lesterol metabolism, future studies should examine thephysiological effects of intestinal cholesterol conversion
4957EFFECTS OF HF FEEDING ON GF MICE

by intestinal microbiota and its reabsorption by thehost.
Previously, we demonstrated that reduction of intes-tinal bacteria with an antibiotic treatment improvedoral glucose tolerance of ob/ob and DIO mice (8). In thepresent study, we confirmed that C57BL/6J mice bornand raised in a GF environment are resistant to diet-induced obesity. Reduced food intake and increasefecal lipid excretion contributed to the limited weightgain and decreased food efficiency. In addition, we alsodemonstrated that GF C57BL/6J mice are resistant toHF feeding associated insulin resistance, hepatic steato-sis, dyslipidemia, and elevated systemic proinflamma-tory markers. More interestingly, GF/HF mice displayunexpected changes in cholesterol metabolism withsynergistic elevation of hepatic SREBP2 mRNA andprotein levels in the nuclei as well as the expression ofSREBP2 target genes in the cholesterol biosynthesispathway. In summary, the results of this study revealmany fundamental differences in the physiology of GFmice and demonstrate the importance of intestinalbacteria in the regulation of lipid and carbohydratemetabolism of the host.
The authors thank Drs. Kevin Acheson and KatherineMace for valuable scientific comments, Alain Joffre (InstitutNational de la Recherche Agronomique, Jouy-en-Josas,France) for breeding and husbandry of GF mice, SylvianeMetairon for microarray analysis, Sylvie Pridmore-Merten(Nestlé Research Center, Lausanne, Switzerland) for provid-ing mouse fecal samples, and Irina Monnard for fecal sampleanalysis. Nestec S.A. provided funding to S.R., A.B., and P.G.for the present work. M.M., T.H., M.M., F.R., S.M., R.M., andC.J.C. are employees of Nestec S.A.
REFERENCES
1. Backhed, F., Ding, H., Wang, T., Hooper, L. V., Koh, G. Y.,Nagy, A., Semenkovich, C. F., and Gordon, J. I. (2004) The gutmicrobiota as an environmental factor that regulates fat storage.Proc. Natl. Acad. Sci. U. S. A. 101, 15718–15723
2. Backhed, F., Manchester, J. K., Semenkovich, C. F., andGordon, J. I. (2007) Mechanisms underlying the resistance todiet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci.U. S. A. 104, 979–984
3. Turnbaugh, P. J., Backhed, F., Fulton, L., and Gordon, J. I.(2008) Diet-induced obesity is linked to marked but reversiblealterations in the mouse distal gut microbiome. Cell Host Microbe3, 213–223
4. Turnbaugh, P. J., Ridaura, V. K., Faith, J. J., Rey, F. E., Knight,R., and Gordon, J. I. (2009) The effct of diet on the human gutmicrobiome: a metagenomic analysis in humanized gnotobioticmice. Sci. Translat. Med. 1, 1–9
5. Turnbaugh, P. J., Ley, R. E., Mahowald, M. A., Magrini, V.,Mardis, E. R., and Gordon, J. I. (2006) An obesity-associated gutmicrobiome with increased capacity for energy harvest. Nature444, 1027–1031
6. Larsen, N., Vogensen, F. K., van den Berg, F. W., Nielsen, D. S.,Andreasen, A. S., Pedersen, B. K., Al-Soud, W. A., Sorensen, S. J.,Hansen, L. H., and Jakobsen, M. (2010) Gut microbiota inhuman adults with type 2 diabetes differs from non-diabeticadults. PLoS One 5, e9085
7. Cani, P. D., Amar, J., Iglesias, M. A., Poggi, M., Knauf, C.,Bastelica, D., Neyrinck, A. M., Fava, F., Tuohy, K. M., Chabo, C.,Waget, A., Delmee, E., Cousin, B., Sulpice, T., Chamontin, B.,Ferrieres, J., Tanti, J. F., Gibson, G. R., Casteilla, L., Delzenne,
N. M., Alessi, M. C., and Burcelin, R. (2007) Metabolic endo-toxemia initiates obesity and insulin resistance. Diabetes 56,1761–1772
8. Membrez, M., Blancher, F., Jaquet, M., Bibiloni, R., Cani,P. D., Burcelin, R. G., Corthesy, I., Mace, K., and Chou, C. J.(2008) Gut microbiota modulation with norfloxacin andampicillin enhances glucose tolerance in mice. FASEB J. 22,2416 –2426
9. Lin, C. C., and Yin, M. C. (2008) Effects of cysteine-containingcompounds on biosynthesis of triacylglycerol and cholesteroland anti-oxidative protection in liver from mice consuming ahigh-fat diet. Br. J. Nutr. 99, 37–43
10. Chan, M. Y., Zhao, Y., and Heng, C. K. (2008) Sequentialresponses to high-fat and high-calorie feeding in an obesemouse model. Obesity 16, 972–978
11. Hara, A., and Radin, N. S. (1978) Lipid extraction of tissues witha low-toxicity solvent. Anal. Biochem. 90, 420–426
12. Folch, J., Lees, M., and Sloane Stanley, G. H. (1957) A simplemethod for the isolation and purification of total lipides fromanimal tissues. J. Biol. Chem. 226, 497–509
13. Mansourian, R., Mutch, D. M., Antille, N., Aubert, J., Fogel,P., Le Goff, J. M., Moulin, J., Petrov, A., Rytz, A., Voegel, J. J.,and Roberts, M. A. (2004) The Global Error Assessment(GEA) model for the selection of differentially expres-sed genes in microarray data. Bioinformatics 20, 2726 –2737
14. Rousseeuw, P. J., and Croux, C. (1993) Alternatives to themedian absolute deviation. J. Am. Statist. Assoc. 88, 1273–1283
15. R Development Core Team (2007) R 2.6.1, R Foundation forStatistical Computing, Vienna, Austria; ISBN 3-900051-07-0;http://www.R-project.org
16. Ye, X. Y., Xue, Y. M., Sha, J. P., Li, C. Z., and Zhen, Z. J. (2009)Serum amyloid A attenuates cellular insulin sensitivity by in-creasing JNK activity in 3T3–L1 adipocytes. J. Endocrinol. Invest.32, 568–575
17. Fleissner, C. K., Huebel, N., Abd El-Bary, M. M., Loh, G.,Klaus, S., and Blaut, M. (2010) Absence of intestinal micro-biota does not protect mice from diet-induced obesity. Br. J.Nutr. 5, 1–11
18. Hayashi, T., Hirshman, M. F., Kurth, E. J., Winder, W. W., andGoodyear, L. J. (1998) Evidence for 5 AMP-activated proteinkinase mediation of the effect of muscle contraction on glucosetransport. Diabetes 47, 1369–1373
19. Cani, P. D., Neyrinck, A. M., Fava, F., Knauf, C., Burcelin,R. G., Tuohy, K. M., Gibson, G. R., and Delzenne, N. M.(2007) Selective increases of bifidobacteria in gut microfloraimprove high-fat-diet-induced diabetes in mice through amechanism associated with endotoxaemia. Diabetologia 50,2374 –2383
20. Xu, H., Barnes, G. T., Yang, Q., Tan, G., Yang, D., Chou, C. J.,Sole, J., Nichols, A., Ross, J. S., Tartaglia, L. A., and Chen, H.(2003) Chronic inflammation in fat plays a crucial role in thedevelopment of obesity-related insulin resistance. J. Clin. Invest.112, 1821–1830
21. Hotamisligil, G. S., Budavari, A., Murray, D., and Spiegelman,B. M. (1994) Reduced tyrosin kinase activity of the insulinreceptor in obesity-diabetes. central role of tumor necrosisfactor-alpha. J. Clin. Invest. 94, 1543–1549
22. Hotamisligil, G. S., Shargill, N. S., and Spiegelman, B. M.(1993) Adipose expression of tumor necrosis factor-alpha:direct role in obesity-linked insulin resistance. Science 259,87–91
23. Uysal, K. T., Wiesbrock, S. M., Marino, M. W., and Hotamisligil,G. S. (1997) Protection from obesity-induced insulin resis-tance in mice lacking TNF-alpha function. Nature 389, 610 –614
24. Ventre, J., Doebber, T., Wu, M., MacNaul, K., Stevens, K.,Pasparakis, M., Kollias, G., and Moller, D. E. (1997) Targeteddisruption of the tumor necrosis factor-alpha gene: metabolicconsequences in obese and nonobese mice. Diabetes 46,1526 –1531
25. Unger, R. H. (2003) Minireview: weapons of lean body massdestruction: the role of ectopic lipids in the metabolic syn-drome. Endocrinology 144, 5159–5165
4958 Vol. 24 December 2010 RABOT ET AL.The FASEB Journal � www.fasebj.org

26. Berggreen, C., Gormand, A., Omar, B., Degerman, E., andGoransson, O. (2009) Protein kinase B activity is required forthe effects of insulin on lipid metabolism in adipocytes. Am. J.Physiol. Endocrinol. Metab. 296, E635–E646
27. Wostmann, B. S., and Wiech, N. L. (1961) Total serum and livercholesterol in germfree and conventional male rats. Am. J.Physiol. 201, 1027–1029
28. Hwa, J. J., Zollman, S., Warden, C. H., Taylor, B. A., Edwards,P. A., Fogelman, A. M., and Lusis, A. J. (1992) Genetic anddietary interactions in the regulation of HMG-CoA reductasegene expression. J. Lipid Res. 33, 711–725
29. Wostmann, B. S. (1973) Intestinal bile acids and cholesterolabsorption in the germfree rat. J. Nutr. 103, 982–990
30. Repa, J. J., Berge, K. E., Pomajzl, C., Richardson, J. A., Hobbs,H., and Mangelsdorf, D. J. (2002) Regulation of ATP-bindingcassette sterol transporters ABCG5 and ABCG8 by the liver Xreceptors alpha and beta. J. Biol. Chem. 277, 18793–18800
31. Calpe-Berdiel, L., Rotllan, N., Fievet, C., Roig, R., Blanco-Vaca, F., and Escola-Gil, J. C. (2008) Liver X receptor-mediated activation of reverse cholesterol transport frommacrophages to feces in vivo requires ABCG5/G8. J. LipidRes. 49, 1904 –1911
32. Yu, L., Li-Hawkins, J., Hammer, R. E., Berge, K. E., Horton,J. D., Cohen, J. C., and Hobbs, H. H. (2002) Overexpressionof ABCG5 and ABCG8 promotes biliary cholesterol secretionand reduces fractional absorption of dietary cholesterol.J. Clin. Invest. 110, 671– 680
33. McGillicuddy, F. C., de la Llera, M. M., Hinkle, C. C., Joshi,M. R., Chiquoine, E. H., Billheimer, J. T., Rothblat, G. H., andReilly, M. P. (2009) Inflammation impairs reverse cholesteroltransport in vivo. Circulation 119, 1135–1145
34. Khovidhunkit, W., Moser, A. H., Shigenaga, J. K., Grunfeld, C.,and Feingold, K. R. (2003) Endotoxin down-regulates ABCG5and ABCG8 in mouse liver and ABCA1 and ABCG1 in J774murine macrophages: differential role of LXR. J. Lipid Res. 44,1728–1736
35. Jahnel, J., Fickert, P., Langner, C., Hogenauer, C., Silbert, D.,Gumhold, J., Fuchsbichler, A., and Trauner, M. (2009)Impact of experimental colitis on hepatobiliary transporterexpression and bile duct injury in mice. Liver Int. 29, 1316 –1325
36. Cani, P. D., Bibiloni, R., Knauf, C., Waget, A., Neyrinck, A. M.,Delzenne, N. M., and Burcelin, R. (2008) Changes in gutmicrobiota control metabolic endotoxemia-induced inflamma-tion in high-fat diet-induced obesity and diabetes in mice.Diabetes 57, 1470–1481
37. Dam, H. (1934) The formation of coprosterol in the intestine:the action of intestinal bacteria on cholesterol. Biochem. J. 28,820–825
38. Eyssen, H. J., Parmentier, G. G., Compernolle, F. C., De, P. G.,and Piessens-Denef, M. (1973) Biohydrogenation of sterols byEubacterium ATCC 21,408–Nova species. Eur. J. Biochem. 36,411–421
39. Gerard, P., Lepercq, P., Leclerc, M., Gavini, F., Raibaud, P., andJuste, C. (2007) Bacteroides sp. strain D8, the first cholesterol-reducing bacterium isolated from human feces. Appl. Environ.Microbiol. 73, 5742–5749
Received for publication May 20, 2010.Accepted for publication August 5, 2010.
4959EFFECTS OF HF FEEDING ON GF MICE





























