Embed Size (px)
Citation preview

1
Table of Contents
German Programme on Biodiversity and Global Change
(Phase I, 2000 - 2004) Funded by BMBF
Status Report 2001
INTRODUCTION TO THE PROGRAMME BIOLOG.................................... 10
BIOLOG EUROPE.........................................................................................................................12BIOLOG AFRICA..........................................................................................................................14
BIODIVERSITY INFORMATICS IN THE BIOLOG PROGRAMME.........................................16
BIOLOG EUROPE
Project ID: 01LC0002.....................................................................................................................20GLOBAL CHANGE-DEPENDENT SPREAD OF NEOPHYTES: RELATIONSHIP TO ECOSYSTEMCHANGE (CASE STUDY: FLOODPLAINS OF THE RIVER RHINE)
Project ID: 01LC0004.....................................................................................................................22RECONSTRUCTING HISTORICAL BIODIVERSITY FROM ARCHIVAL SOURCES
Project ID: 01LC0005.....................................................................................................................24BIODIVERSITY CHANGE DURING 20 YEARS OF SUCCESSION: IMPLICATIONS FOR MANAGEMENTOF RESTORATION
Project ID: 01LC0006.....................................................................................................................26ECOLOGY AND BIODIVERSITY OF HIGHER FUNGI IN CENTRAL EUROPE (GERMANY)
Project ID: 01LC0010.....................................................................................................................28CONDITIONS OF A RATIONAL BIODIVERSITY POLICY FOR GERMANY, AS A PART OF THEEUROPEAN UNION
Project ID: 01LC0012 (Subproject 1)..............................................................................................30EVOLUTIONARY CONSEQUENCES OF BIOLOGICAL INVASIONS – INTROGRESSIVEHYBRIDIZATION BETWEEN INVASIVE AND NATIVE RORIPPA SPECIES (BRASSICACEAE).
Project ID: 01LC0012 (Subproject 2)..............................................................................................32EVOLUTIONARY CONSEQUENCES OF BIOLOGICAL INVASIONS – ONGOING HYBRIDIZATIONPROCESSES BETWEEN CARDAMINE TAXA AT THE URNERBODEN, CENTRAL SWITZERLAND

2
Project ID: 01LC0013.....................................................................................................................34BIODIVERSITY AND ECOSYSTEM FUNCTION IN GRASSLANDS - THE EFFECTS OF INSECTHERBIVORY AND DROUGHT STRESS
Project ID: 01LC0013 (Subproject STOFF)....................................................................................36THE CARBON AND NITROGEN CYCLES IN GRASSLAND ECOSYSTEMS OF DIFFERENTDIVERSITIES
Project ID: 01LC0013 (Subproject RAUM)....................................................................................38A SPATIAL MODEL FOR INDICATING ECOSYSTEM FUNCTIONS
Project ID: 01LC0013 (Subproject ENTO).....................................................................................40THE RELATIONSHIP BETWEEN INSECT HERBIVORIE, PLANT DIVERSITY, INSECT DIVERSITY ANDECOSYSTEM PROCESSES
Project ID: 01LC0013 (Subproject GENMOD) ..............................................................................42GENETIC AND PHENOTYPIC DIVERSITY AND RESPONSE TO DISTURBANCES IN GRASSLANDSYSTEMS: MODELLING AND EMPIRICAL RESEARCH
Projekt ID: 01LC0014.....................................................................................................................44BIODIVERSITY AND ECOSYSTEM FUNCTIONS IN GRASSLAND
Projekt ID: 01LC0014 (Subproject Vegetation) ..............................................................................46TAXONOMIC, FUNCTIONAL AND STRUCTURAL CHARACTERISTICS OF ABOVE-GROUND PLANTFEATURES AND THEIR INFLUENCE ON ECOSYSTEM FUNCTIONS
Projekt ID: 01LC0014 (Subproject Roots) ......................................................................................48THE ROLE OF BIODIVERSITY FOR STORAGE OF CARBON AND MINERALS IN ROOTS, ANDACQUISITION OF SOIL RESSOURCES IN GRASSLAND
Projekt ID: 01LC0014 (Subproject Soil) .........................................................................................50THE IMPACT OF PLANT DIVERSITY OF GRASSLANDS ON WATER- AND NUTRIENT-FLUXES INSOILS AND SOIL ORGANIC MATTER
Project ID: 01LC0015 (Subproject VEGETATION ECOLOGY) ...................................................52DEVELOPMENT OF BIOLOGICAL DIVERSITY IN SALT MEADOWS OF THE 'VORPOMMERSCHEBODDENLANDSCHAFT' – SPATIO-TEMPORAL DYNAMICS AND ECOSYSTEM FUNCTION OFBIODIVERSITY IN BRACKISH GRASSLANDS
PROJECT ID: 01LC0015 (Subproject LANDSCAPE ECONOMY)...............................................54DEVELOPMENT OF BIOLOGICAL DIVERSITY IN SALT MEADOWS OF THE 'VORPOMMERSCHEBODDENLANDSCHAFT' – ECONOMICAL VALUATION OF BIODIVERSITY
Project ID: 01LC0015 (Subproject ANIMAL ECOLOGY).............................................................56DEVELOPMENT OF BIOLOGICAL DIVERSITY IN SALT MEADOWS OF THE 'VORPOMMERSCHEBODDENLANDSCHAFT' - DYNAMICS IN DIVERSITY OF ANIMAL SPECIES AND COMMUNITIES IN ACHANGING ENVIRONMENT WITH SPECIAL REGARD TO DISPERSAL PROCESSES
Project ID: 01LC0015 (Subproject SOIL SCIENCE) .....................................................................58DEVELOPMENT OF BIOLOGICAL DIVERSITY IN SALT MEADOWS OF THE 'VORPOMMERSCHEBODDENLANDSCHAFT' - WATER AND SALT DYNAMICS OF SALTY MEADOW AND MARSH SOILS
Project ID: 01LC0018 (SUBICON Subproject 1)............................................................................60SUBICON - SUCCESSIONAL CHANGE AND BIODIVERSITY CONSERVATION IN A RED OAKCHRONOSEQUENCE – SUCCESSION OF DIVERSITY AND FUNCTION OF ECTOMYCORRHIZAL TYPES IN RED OAK FORESTECOSYSTEMS
Project ID: 01LC0018 (SUBICON Subproject 2)............................................................................62BIODIVERSITY OF PLANT COMMUNITIES AND SELECTED PHYTOPHAGES IN RED OAKDOMINATED ECOSYSTEMS

3
Project ID: 01LC0018 (SUBICON Subproject 3)............................................................................64GENETIC DIVERSITY OF SELECTED PLANTS, PHYTOPHAGES AND ZOOPHAGES IN SUCCESSIONALRED OAK ECOSYSTEMS
Project ID: 01LC0018 (SUBICON Subproject 4)............................................................................66STRUCTURE OF THE PREDATOR COMMUNITY IN A RED OAK CHRONOSEQUENCE
Project ID: 01LC0020 (BIOPLEX Subproject 1) ............................................................................68BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURAL LANDSCAPES UNDER GLOBALCHANGE (BIOPLEX) - MODELLING AND GEOSTATISTICS
Project ID: 01LC0020 (BIOPLEX Subproject 2) ............................................................................70BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURAL LANDSCAPES UNDER GLOBALCHANGE (BIOPLEX) - AGROECOLOGY
Project ID: 01LC0020 (BIOPLEX Subproject 3) ............................................................................72BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURAL LANDSCAPES UNDER GLOBALCHANGE (BIOPLEX) - PHYTOPATHOLOGY
Project ID: 01LC0020 (BIOPLEX Subproject 4) ............................................................................74BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURAL LANDSCAPES UNDER GLOBALCHANGE (BIOPLEX) - Soil BIOTA
Project ID: 01LC0020 (BIOPLEX Subproject 5) ............................................................................76BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURAL LANDSCAPES UNDER GLOBALCHANGE (BIOPLEX) - SOCIAL-ECONOMIC EVALUATION AND APPRAISAL WITH RESPECT TOBIODIVERSITY IN AGRARIAN LANDSCAPES
Project ID: 01LC0021 (Subproject 1)..............................................................................................78BIODIVERSITY OF NITROGEN-FIXING MICROORGANISMS
Project ID: 01LC0021 (Subproject 2)..............................................................................................80BIODIVERSITY OF NITRIFYING AND SULFATE-REDUCING PROKARYOTES
Project ID: 01LC0021 (Subproject 3)..............................................................................................82BIODIVERSITY OF METHANOTROPHIC AND NITRATE-REDUCING BACTERIA
Project ID: 01LC0021 (Subproject 4)..............................................................................................84BIODIVERSITY AND SUCCESSIONS IN TEMPORARY TERRESTRIAL-AQUATIC INTERFACES
Project ID: 01LC0021 (Subproject 5)..............................................................................................86INVESTIGATION OF PROKARYOTIC DIVERSITY BY OPTIMIZED CULTIVATION METHODS
Project ID: 01LC0021 (Subproject 6)..............................................................................................88THE ROLE OF N-ACYL-L-HOMOSERINE LACTONE (AHL) MEDIATED COMMUNICATION FOR THESTRUCTURE AND PERSISTENCE OF MICROBIAL RHIZOSPHERE COMMUNITIES
Project ID: 01LC0021 (Subproject 7)..............................................................................................90BIODIVERSITY OF PROTOZOA IN THE RHIZOSPHERE OF PLANTS: EFFECTS ON ACTIVITY,PERFORMANCE AND DIVERSITY OF MICROORGANISMS AND ON PLANT GROWTH.
Project ID: 01LC0101 (Subproject 1)..............................................................................................92AN ANALYSIS OF PREVAILING ECONOMIC AND POLITICAL STRATEGIES FOR THE PROTECTIONOF BIODIVERSITY - THE TRADE IN GENETIC RESSOURCES: A VIABLE CONTRIBUTION TO THEPROTECTION OF BIODIVERSITY?
Project ID: 01LC0101 (Subproject 2)..............................................................................................94AN ANALYSIS OF PREVAILING ECONOMIC AND POLITICAL STRATEGIES FOR THE PROTECTIONOF BIODIVERSITY - THE PROTECTION OF BIODIVERSITY: NEW APPROACHES AND INSTRUMENTSIN INTERNATIONAL LAW

4
BIOLOG AFRICA
Project ID: 01LC0024 (BIOTA Southern Africa) ............................................................................98A RESEARCH PROJECT MONITORING AND ANALYSING BIODIVERSITY AND ITS CHANGE ALONGA TRANSECT IN NAMIBIA AND SOUTH AFRICA
Project ID: 01LC0024 (BIOTA Southern Africa S01)................................................................... 100REMOTE SENSING AND GIS BASED SURVEY OF SPATIAL AND TEMPORAL BIODIVERSITYDYNAMICS AND ANALYSIS OF BIODIVERSITY AND GEODIVERSITY INTERRELATIONSHIPS
Project ID: 01LC0024 (BIOTA Southern Africa S02)................................................................... 102EDAPHICAL DIVERSITY AND BIODIVERSITY IN MUTUAL DEPENDENCY
Project ID: 01LC0024 (BIOTA Southern Africa S03)................................................................... 104BIODIVERSITY OF SOUTHWESTERN AFRICAN FUNGI: INTERACTIONS WITH HIGHER PLANTS,FUNCTIONAL DIVERSITY AND DNA-TAXONOMY
Project ID: 01LC0024 (BIOTA Southern Africa S04)................................................................... 106DELVELOPMENT OF AN INTEGRATED SYSTEM FOR THE INPUT, STORAGE AND ANALYSIS OFBIODIVERSITY DATA IN SOUTHERN AFRICA
Project ID: 01LC0024 (BIOTA Southern Africa S05)................................................................... 108BIOLOGICAL SOIL CRUSTS OF TERRESTRIAL ECOSYSTEMS: MODES OF PATTERN FORMATIONAND THEIR MODELLING CONCERNING DIVERSITY, STRUCTURE AND FUNCTION INCRYPTOGAMIC COMMUNITIES
Project ID: 01LC0024 (BIOTA Southern Africa S06)................................................................... 110CHANGES IN BOTANICAL BIODIVERSITY WITH REGARD TO CHANGES IN LAND USE PRACTICESAND CLIMATE: STANDARDISED MONITORING AND TRANSECT ANALYSIS
Project ID: 01LC0024 (BIOTA Southern Africa S07)................................................................... 112FUNCTIONAL ZOODIVERSITY IN SOUTHERN AFRICA UNDER CHANGING ENVIRONMENTS ANDHUMAN USE
Project ID: 01LC0024 (BIOTA Southern Africa S08)................................................................... 114DIVERSITY OF NAMIBIAN DRAGONFLIES: EFFECTS OF ANTHROPOGENIC CHANGES ANDMODELLING ON DIFFERENT SCALES
Project ID: 01LC0024 (BIOTA Southern Africa S09)................................................................... 116STRUCTURAL AND SPECIES DIVERSITY IN SEMI-ARID SAVANNAHS OF SOUTHERN AFRICA: THEIMPACT OF LAND USE AND CLIMATIC CHANGE
Project ID: 01LC0024 (BIOTA Southern Africa S10)................................................................... 118WHICH PROCESSES AND MECHANISMS DETERMINE THE BIODIVERSITY IN (SEMI-) ARID PLANTCOMMUNITIES?
Project ID: 01LC0024 (BIOTA Southern Africa S11)................................................................... 120SOCIO-ECONOMIC ASPECTS OF CHANGES IN BIODIVERSITY IN SOUTHERN AFRICA
Project ID: 01LC0007................................................................................................................... 122BIOLOGICAL STUDIES ON MYCO-HETEROTROPHIC PLANTS WITH SPECIAL EMPHASIS OF THEIRMYCORRHIZAE
Project ID: 01LC0017 (BIOTA West Africa) ................................................................................ 124BIOTA WEST AFRICA – GENERAL SUMMARY
Project ID: 01LC0017 (BIOTA West Africa W01)........................................................................ 128REMOTE SENSING AND GIS BASED SURVEY OF SPATIAL AND TEMPORAL BIODIVERSITYDYNAMICS AND ANALYSIS OF BIODIVERSITY AND GEODIVERSITY INTERRELATIONSHIPS

5
Project ID: 01LC0017 (BIOTA West Africa W02)........................................................................ 130BIOPHYSICAL AND HYDROMETEOROLOGICAL PARAMETERS WITHIN THE FRAMEWORK OFTERRESTRIAL BIODIVERSITY RESEARCH
Project ID: 01LC0017 (BIOTA West Africa W03)........................................................................ 132BIODIVERSITY OF AFRICA IN THE GLOBAL CONTEXT: SPATIAL PATTERNS OF VASCULAR PLANTDIVERSITY IN A CHANGING ENVIRONMENT (BIOMAPS PROJECT)
Project ID: 01LC0017 (BIOTA West Africa W04)........................................................................ 134PHYTODIVERSITY AND DYNAMICS OF HABITAT FRAGMENTS IN THE IVORY COAST: SPATIALAND TEMPORAL PATTERNS OF BIODIVERSITY CHANGES, EDGE EFFECTS, AND THEIRFUNCTIONAL CONSEQUENCES IN NATURAL AND DISTURBED ECOSYSTEM MOSAICS
Project ID: 01LC0017 (BIOTA West Africa W06)........................................................................ 136DETERMINANTS OF SMALL-SCALE MOSAICS OF ARTHROPOD COMMUNITIES IN NATURAL ANDANTHROPOGENICALLY DISTURBED HABITATS
Project ID: 01LC0017 (BIOTA West Africa W07)........................................................................ 140COMPARATIVE STUDIES ON EPHEMERAL SAVANNA WATERS AND THEIR SIGNIFICANCE FORDIVERSITY AND PRODUCTIVITY OF FISH COMMUNITIES IN NATURAL ANDANTHROPOGENICALLY DISTURBED RIVERS
Project ID: 01LC0017 (BIOTA West Africa W08)........................................................................ 142AMPHIBIAN DIVERSITY AND DIVERSITY CHANGES IN WEST AFRICA: GLOBAL, REGIONAL ORLOCAL CAUSES?
Project ID: 01LC0017 (BIOTA West Africa W09)........................................................................ 144THE INFLUENCE OF LOCAL AND REGIONAL FACTORS ON THE DIVERSITY, STRUCTURE, ANDFUNCTION OF WEST AFRICAN BAT COMMUNITIES (CHIROPTERA)
Project ID: 01LC0017 (BIOTA West Africa W11)........................................................................ 146PHYTODIVERSITY IN THE WEST AFRICAN SAHEL AND SUDAN ZONE – DEVELOPMENT ANDEVALUATION
Project ID: 01LC0022................................................................................................................... 148THE CUVETTE CENTRALE AS RESERVOIR OF MEDICINAL PLANTS
Project ID: 01LC0025 (BIOTA East Africa E01) .......................................................................... 150BIOTA EAST AFRICA: BIODIVERSITY IN CONVERSION – THE INFLUENCE OF FRAGMENTATIONAND MAN-MADE EXPLOITATION ON THE BIODIVERSITY OF TROPICAL MONTANE FORESTS
Project ID: 01LC0025 (BIOTA East Africa E02) .......................................................................... 152IMPACT OF BIODIVERSITY CHANGE ON THE EXCHANGE POTENTIAL OF TRACE GASES BETWEENECOSYSTEMS AND ATMOSPHERE
Project ID: 01LC0025 (BIOTA East Africa E03) .......................................................................... 154REGENERATION OF TROPICAL MONTANE TREE SPECIES – SPATIAL AND TEMPORAL DYNAMICSOF FEED BACK PROCESSES
Project ID: 01LC0025 (BIOTA East Africa E04) .......................................................................... 156INFLUENCE OF NATURAL AND ANTHROPOGENIC FRAGMENTATION ON THE VEGETATION INMONTANE RAINFORESTS OF KAKAMEGA AND MT. KENYA
Project ID: 01LC0025 (BIOTA East Africa E06) .......................................................................... 158LEPIDOPTERA AS INDICATORS OF HUMAN IMPACT ON TROPICAL RAINFOREST ECOSYSTEMS INEASTERN AFRICA
Project ID: 01LC0025 (BIOTA East Africa E07) .......................................................................... 160DIVERSITY AND SPECIES COMPOSITION OF ODONATA AS INDICATORS OF BIOTOPE QUALITY OFEAST AFRICAN RAIN FORESTS AND THEIR REPLACEMENT COMMUNITIES

6
Project ID: 01LC0025 (BIOTA East Africa E08) .......................................................................... 162MEASURING AND MONITORING AMPHIBIAN DIVERSITY IN EAST AFRICA: SEPARATING GLOBALFROM REGIONAL AND LOCAL CAUSES
Project ID: 01LC0025 (BIOTA East Africa E09) .......................................................................... 164THE INFLUENCE OF LAND USE MODES ON DIVERSITY, ABUNDANCE AND GUILD STRUCTURE OFCOPROPHAGOUS BEETLES IN THE AFRICAN FOREST-SAVANNA-MOSAIC
Project ID: 01LC0025 (BIOTA East Africa E10) .......................................................................... 166EFFECTS OF HABITAT FRAGMENTATION ON PLANT-POLLINATOR INTERACTIONS IN EASTAFRICAN RAINFORESTS
Project ID: 01LC0025 (BIOTA East Africa E11) .......................................................................... 168EFFECT OF FRAGMENTATION ON SEED DISPERSAL, REGENERATION AND DYNAMICS OFKAKAMEGA FOREST, KENYA
Project ID: 01LC0025 (BIOTA East Africa E12) .......................................................................... 170MONTANE FOREST ECOSYSTEMS IN KENYA - DIVERSITY AND POTENTIAL FOR USE
Project ID: 01LC0025 (BIOTA East Africa, Yemen E13-E16)...................................................... 172BIOTA-EAST YEMEN-PROJECT-GROUP: BIODIVERSITY AND ECOLOGY OF THE PALAEO-AFRICANREFUGE AREAS IN SOUTHERN ARABIA AND ON SOCOTRA (REPUBLIC OF YEMEN)
Project ID: 01LC0025 (BIOTA East Africa, Yemen E13) ............................................................. 174PHYTODIVERSITY AND VEGETATION TYPES OF REPRESENTATIVE PALEO-AFRICAN REFUGIA INTHE COASTAL MOUNTAINS OF SOUTHERN YEMEN AND ON SOCOTRA (REPUBLIC OF YEMEN)
Project ID: 01LC0025 (BIOTA East Africa, Yemen E14) ............................................................. 176ECOLOGICAL CAUSES OF BIODIVERSITY IN THE COASTAL MOUNTAINS OF SOUTHERN YEMENAND ON SOCOTRA (REPUBLIC OF YEMEN)
Project ID: 01LC0025 (BIOTA East Africa, Yemen E15) ............................................................. 178PHYLOGEOGRAPHIC AND POPULATION GENETIC ASPECTS OF THE PHYTODIVERSITY INPALAEO-AFRICAN REFUGIA OF THE COASTAL MOUNTAINS OF SOUTHERN YEMEN ANDSOCOTRA (REPUBLIC OF YEMEN)
Project ID: 01LC0025 (BIOTA East Africa, Yemen E16) ............................................................. 180EVOLUTIONARY HISTORY AND PHYLOGEOGRAPHY OF SOUTH ARABIAN FAUNAL ELEMENTS
Project ID: 01LC0001................................................................................................................... 182BIODIVERSITY OF SELECTED MODEL GROUPS OF THE PHYTOPLANKTON IN STANDING INLANDWATERS OF KENYA
BIODIVERSITY INFORMATICS
Project ID: 01LC9901 (SysTax) .................................................................................................... 186SYSTAX - ELECTRONIC DATA PROCESSING FOR RECORDING AND ANALYSING BIODIVERSITYDATA WITH THE SYSTEMATIC AND TAXONOMIC DATABASE SYSTEM SYSTAX
Project ID: 01LC9902 (ARB)........................................................................................................ 188AUTOMATED MAINTAINANCE OF INTEGRATED DATABASES FOR RRNA AND OTHER RELEVANTGENE SEQUENCES; SOFTWARE DEVELOPMENT, OPTIMIZATION, AND PARALLELIZATION FORAUTOMATED SEARCHING OF SEQUENCE IDENTIFIERS FOR ANALYZING DIVERSITY
Project ID: 01LC9903 (ZEFOD)................................................................................................... 190REGISTER FOR BIOLOGICAL RESEARCH COLLECTIONS IN GERMANY
Project ID: 01LC9904 (EDIS) ....................................................................................................... 192EDIS – ENTOMOLOGICAL DATA INFORMATION SYSTEM

7
Project ID: 01LC9904 (EDIS, Subproject ABIS) .......................................................................... 194ABIS - AUTOMATED IDENTIFICATION OF BEE SPECIES
Project ID: 01LC9904 (EDIS, Subproject DNATAX 1)................................................................. 196DNATAX – DNA TAXONOMY AS A TOOL IN BIODIVERSITY RESEARCH: RECORDING,ASSESSMENT, MONITORING AND UTILIZATION OF THE GENETIC DIVERSITY OF EUKARYONTSYSTEMS
Project ID: 01LC9904 (EDIS, Subproject DNATAX 2 - MOLART)............................................. 198MOLART (DNATAX 2): MOLECULAR IDENTIFICATION OF COLLEMBOLA: METHOD OPTIMISATIONVERIFICATION - DOCUMENTATION
Project ID: 01LC9904 (EDIS, Subproject DORSA) ...................................................................... 200DORSA - GERMAN ORTHOPTERA COLLECTIONS
Project ID: 01LC9904 (EDIS, Subproject GART) ........................................................................ 202GART – GLOBAL SPECIES REGISTER BUTTERFLIES
Project ID: 01LC9904 (EDIS, Subproject INGE).......................................................................... 204INGE – INVENTORY OF GEOMETRID MOTHS OF EUROPE (INSECTA, LEPIDOPTERA): PROVIDINGMULTIMEDIAL DATA ON SYSTEMATICS, ZOOGEOGRAPHY AND ECOLOGY
Project ID: 01LC9904 (EDIS, Subproject OBIF) .......................................................................... 206OBIF - OPTIMIZATION OF BIODIVERSITY INFORMATION FACILITIES ON APPLICATION-ORIENTEDRESEARCH
Project ID: 01LC9905 (GLOPP)................................................................................................... 208GLOPP - GLOBAL INFORMATION SYSTEM FOR THE BIODIVERSITY OF PLANT PATHOGENICFUNGI
Project ID: 01LC9905 (GLOPP, Subproject IT) ........................................................................... 210GLOPP-IT - BIOINFORMATICS AND DEVELOPMENT OF THE INFORMATION SYSTEM
Project ID: 01LC9905 (GLOPP, Subproject LIT)......................................................................... 212GLOPP-LIT - DIGITIZATION OF IMPORTANT EUROPEAN HOST-PATHOGEN INDICES FOR FUNGI
Project ID: 01LC9905 (GLOPP, Subproject Oomycetes).............................................................. 214GLOPP-OOMYCETES
Project ID: 01LC9905 (GLOPP, Subproject Erysiphales)............................................................. 216GLOPP-ERYSIPHALES - A GLOBAL INFORMATION SYSTEM FOR THE BIODIVERSITY OF POWDERYMILDEWS (ERYSIPHALES, ASCOMYCOTA)
Project ID: 01LC9905 (GLOPP, Subproject Smut Fungi) ............................................................ 218GLOPP – SMUT FUNGI
Project ID: 01LC9905 (GLOPP, Subproject Uredinales) .............................................................. 220GLOPP-UREDINALES - REGISTRATION OF TROPICAL AND SUBTROPICAL RUST FUNGI
Project ID: 01LC9906 (INFOBOT)............................................................................................... 222INFOBOT - DEVELOPMENT AND TESTING OF INFORMATION SYSTEMS COVERING DIFFERENTASPECTS OF BOTANICAL COLLECTIONS
Project ID: 01LC9906 (INFOBOT, Subproject INFOCOMP/TYPES).......................................... 224COMPOSITAE TYPES IN GERMAN HERBARIA
THE BOHLMANN-FILES – A DATABASE OF NATURAL SUBSTANCES IN THE COMPOSITAE
Project ID: 01LC9906 (INFOBOT, Subproject BoGART) ............................................................ 228BOGART - THE INFORMATION SYSTEM OF THE BERLIN BOTANIC GARDEN

8
Project ID: 01LC0026 (AlgaTerra) ............................................................................................... 230ALGATERRA – AN INFORMATION SYSTEM FOR TERRESTRIAL ALGAL BIODIVERSITY: ASYNTHESIS OF TAXONOMIC, MOLECULAR AND ECOLOGICAL INFORMATION
Project ID: 01LC0026 (AlgaTerra Subproject 1) .......................................................................... 232THE ALGATERRA INFORMATION SYSTEM INCLUDING THE IDEAL-MODULE
Project ID: 01LC0026 (AlgaTerra Subproject 2) .......................................................................... 234ACCESSIBILITY OF THE EHRENBERG COLLECTION
Project ID: 01LC0026 (AlgaTerra Subproject 3) .......................................................................... 236MOLECULAR AND ECOLOGICAL DATA OF TERRESTRIAL DIATOMS
Project ID: 01LC0026 (AlgaTerra Subproject 4) .......................................................................... 238DOCUMENTATION OF CHARACTERS OF TERRESTRIAL ALGAE IN CULTURE: R-DNA SEQUENCEANALYSES, MORPHOLOGY AND CRYOPRESERVATION OF REFERENCE STRAINS
Project ID: 01LC0026 (AlgaTerra Subproject 5) .......................................................................... 240CHARACTERISTIC DATA OF GREEN AND BLUEGREEN AERO-TERRESTRIAL ALGAE(CHLOROPHYTINA AND CYANOBACTERIA)
APPENDIX ...................................................................................................... 242
Index of Names ............................................................................................................................. 242BIOLOG-Coordinators ................................................................................................................. 246

9

10
Introduction to the Programme BIOLOG
Erwin Beck, University of Bayreuth, Dept. of Plant PhysiologyUniversitätsstr.30, D-95440 Bayreuth
BIOLOG: The history BIOLOG is Germany’s most outstanding scientific and also financial responseto the UN-Convention on Biological Diversity, the so-called 1992 RIO-Convention. It is part of theFederal Government’s programme for research into aspects of global change. In 1997 the GermanNational Committee for Global Change recommended the launching of a research programme on“biodiversity in the scope of global change”. In April 1999 the guidelines of the “BIOLOG”programme and a call for projects were published. From 251 proposals 93 projects were finally selectedwhich started their work during the course of the last year. Affiliation with other programmes onbiodiversity research, e.g. the ICSU and IUBS Diversitas programme is expected.
Biodiversity is severely threatened by man There is general agreement among scientists and politiciansthat biodiversity is severely threatened by the activities of the tremendously increasing humanpopulation. Estimates of the worldwide loss of biodiversity range between 2 and 130 species per day.Even if a number on the lower side of this range is assumed, the extinction rate by far exceeds that atthe end of the Cretaceous when the Saurians died out.Research into biodiversity is therefore not a luxury but a necessity. It is a major challenge, not onlybecause of the great variety of related scientific aspects and the multitude of disciplines involved, butalso with respect to the high expectations of the public as to practically applicable results and solutionsof urgent environmental problems arising in the context of global change.
Only a small part of the kingdom of organisms is known at present A primary problem inbiodiversity research is the lack of basic knowledge. Estimates of the total number of species on earthrange between 5 and 100 million, of which less than 2 million are known at present. Therefore furtherinventories are indispensable, but it is clear that the progress in knowledge can hardly keep up with therate of disappearance of species. This holds in particular for the realm of microorganisms where we arejust beginning to develop methods for identifying organisms which can not be cultivated in thelaboratory.
The value of biodiversity is inestimable, and proper use of it has not yet been accomplished If only aminor part of the estimated wealth of organisms is known, can we validate biodiversity and especiallythe losses of biodiversity? In an economic sense this is not possible and from the biological viewpointwe must admit that the value of biodiversity is in fact inestimable. With each extinct species afunctioning genetic system is lost and with it genes and gene products which could have been of greatimportance for the needs of the rapidly growing human population. This holds for all kinds oforganisms, but will be illustrated only by the example of crop plants: Of 270,000 species of higherplants about 3000 serve as food plants, and of these 3000 only 30 species are more or less domesticatedcrop plants of which only the 4 species maize, rice, wheat and potato produce more than 70 % of thefood supply for 6.4 billion people. There is no doubt that even in the food sector man is making not yetproper use of the resource biodiversity.
Biodiversity and the stability of ecosystems Biodiversity is understood as a major factor involved in thestability of ecosystems. However, the investigation of the functional roles of all species of an ecosystemis not feasible because of our incomplete knowledge of the respective biological inventory. Thereforeso-called key species must be identified which may be essential for the ecosystem under consideration,and their specific functions in that ecosystem must be elucidated. This kind of research is not onlynecessary for the understanding of functionality in an ecosystem, but also for the recognition ofenvironmental changes which may destabilize or even cardinally change the ecosystem.

11
Biodiversity and sustainable use of an ecosystem Stability of an ecosystem is the primary prerequisitefor its sustainable use by man. Sustainable use means an almost indefinite number of possibilities ofexploitation, which depend on the ecosystem itself and the people living in it. Important examples withspecial regard to global change are the worldwide endangered tropical mountain forests which gatherthe rain for the benefit of the lowlands and the semiarid ecosystems which are threatened bydesertification. Even in these ecosystems knowledge of functionality of individual organisms, such aspresumptive key species is still missing.
The Pilot Phase: Research into biodiversity requires capacity building It is conceivable that researchinto the already mentioned and additional topics of biodiversity, especially with respect to globalchange, will require special scientific and logistic capacity and in particular sufficient time.The scientific advisory committee which has been in charge of the arrangements of BIOLOG hastherefore given priority to R&D projects which are based on a fair degree of international integrationand capacity building. Especially with respect to these requirements and to a strong and effectivecooperation of the disciplines involved, the committee’s policy was - and still is - to concentrate theprojects to regions expressing different levels of biodiversity, regions of different levels of relevantknowledge and expertise, and of differently endangered biodiversity.
BIOLOG Africa and BIOLOG Europe Projects operating in several regions of subsaharian Africa (theso-called BIOTA-projects: 3 individual projects and 3 clusters of projects with a financial volume of30.1 million DM) and, as an antipole, projects in Central Europe (“BIOLOG EUROPE”, comprising 13projects, awarded grants of 14.2 million DM) were selected to start BIOLOG. A pilot phase of 3 yearsshould be used to develop scientific as well as logistic capacity, and to examine the methodologies. Ageographical information system (GIS) will be established in order to model scenarios when sufficientdata become available.In the BIOTA projects special emphasis is given to biodiversity inventories at different scales,combined with long-term monitoring in so-called “observatories” which are standardized plots ofusually 1 km2 left untreated or subjected to selected kinds and varying intensities of utilization. In theBIOLOG EUROPE projects, the effects of fragmentation of the landscape, of intentional and accidentalinvasion of exotic species and other impacts on biodiversity are being studied, as well as the diversity,dynamics and function of microorganisms in terrestrial ecosystems. In both BIOLOG EUROPE andBIOLOG AFRICA special attention is given to history and tradition, policy implications and socio-economic dimensions of the loss and preservation of biodiversity.
BIOLOG: Biodiversity Informatics In addition to the projects investigating aspects of terrestrialbiodiversity, a subprogramme “Biodiversity Informatics” was advertised. An immense amount oforganismic data are deposited at the big Natural History Museums, e.g. at Frankfurt ,Berlin, Stuttgart,Braunschweig and Munich and also at smaller specialized collections of biological materials. Asubstantial part of these data is not yet readily accessible or is incomplete; in particular moleculargenetic data is still missing for the majority of the collected specimens. The aims of “BiodiversityInformatics” are to improve the methods of biodiversity data capture and maintenance, to digitalize andnetwork the wealth of the already available data collections, and to interlink biodiversity-relatedinfrastructures nationally and worldwide. Out of 56 drafted proposals 31 were selected and have beenorganised within 7 projects with a financial volume of 14.6 million DM. Three of these projectsrepresent major clusters comprising e.g. insects, pathogenic fungi and special families of higher plants.
The BIOLOG Status Symposia Status Symposia will report on the achieved scientific progress of theprojects, on their difficulties, their future plans and on their collaborative integration. The firstsymposium of this series in December 2001 will be of special interest with respect to the announcementof the main phase of BIOLOG.

12
BIOLOG EUROPEA MULTIDISCIPLINARY APPROACH TO EUROPEAN BIODIVERSITY
Volkmar Wolters & Barbara Reinhold-Hurek
As a continent, Europe is unique in the extent to which its natural and cultural landscape is fragmentedinto a patchwork pattern. Humans have deliberately changed Europe’s environment for hundreds ofyears (1). Today there is no place in Europe below 2000 m that has not been affected by anthropogenicimpacts in one-way or the other. The resulting mosaic of natural, semi-natural and cultural landscapefragments is a major factor supporting European biodiversity by creating a complex spatio-temporalpattern of ecosystems and habitats (2). There is growing concern, however, about potentiallydetrimental consequences of large-scale changes on biodiversity, because recent alterations induced byhuman beings occur at unprecedented rates and scales (3, 4).
Biodiversity is the variety of life at all levels of biological organization (5). The 13 projects ofBIOLOG-Europe that are currently funded study the impact of environmental change on Europe’sbiodiversity by focusing on a diversity of ecosystems such as floodplains, grasslands, coastal areas,forests and arable fields (Tab. 1). The following topics are covered:
• Evolution, maintenance and alterations of biodiversity as a result of natural andanthropogenic changes in global environmental conditions;
• Links between biodiversity and both ecosystem stability and functioning;
• Development of indicator systems for rapid survey and evaluation of biodiversity change;
• Policy dimensions of biodiversity.
Ecological processes observed and quantified in experimental approaches include secondarysuccession, invasion of non-native species, changes in landscape complexity, modifications in wateravailability, manipulation of diversity, and management. The various taxa considered include woodyand herbaceous plants, grasses, different guilds of insects, soil biota, basidiomycetes, and terrestrialmicroorganisms. In addition, past biodiversity is reconstructed from historical sources, policydimensions of biodiversity are studied, socio-economic aspects of preserving biodiversity are explored,evolutionary consequences of hybridisation are evaluated, and strategies for the conservation of geneticresources are developed.
A particular focus is on the development and adaptation of methods for analysing diversity,dynamics and function of prokaryotic microorganisms in terrestrial ecosystems. Prokaryotesexclusively or predominantly catalyse key transformations within the global N, S and C cycles, andsignificantly contribute to the mineralization of xenobiotics. According to recent estimates more thanhalf of the earth’s biomass is stored in prokaryotes. Conservative estimates suggest that only 0.4 % ofthe naturally occurring species of the Archaea and Bacteria have yet been described, makingprokaryotes by far the most unexplored group of organisms. This apparent lack of knowledge reflectsthe limitations of traditional techniques for studying complex microbial communities. Only a minorityof the naturally occurring prokaryotes can be obtained by classical enrichment and cultivation methods.Recently developed DNA- or RNA-based techniques allow studying the diversity, in situ populationstructure and activity of complex microbial consortia. Special attention will be given to those terrestrialprokaryotes, which are essential to global nitrogen and sulphur cycling and/or to the production ofgreenhouse gases.

13
Table 1: Overview over the 13 BIOLOG-Europe projects that are currently funded.
Monitoring and Complexity Analysis •Biodiversity and spatial complexity in agricultural landscapes under global change. •Biodiversity change in salt meadows. •Ecology and biodiversity of fungi in Central Europe (with emphasis on Homobasidiomycetidae).Experimental Approaches and Process Analysis •Biodiversity and ecosystem function in grasslands – the effects of insect herbivory and draught
stress. •Biodiversity and Ecosystem Function: Grassland as a Model System. •Biodiversity of terrestrial microbiota. •Evolutionary consequences of biological invasions – Hybridisation between invasive and native
Brassicaceae taxa. •Global change-dependent spread of neophytes: relationship to ecosystems change (case
study: floodplains of the river Rhine).Temporal Dimensions and Socio-economic Perspectives •Mechanisms of biodiversity changes of selected functional groups in Red oak ecosystems. •Management concepts based on biodiversity change during a 20-years succession. •Reconstruction of historic biodiversity from archival sources. •Conditions of a biodiversity-oriented policy for Germany as a part of the EU. •Use and conservation of genetic resources: A strategy for preserving biological diversity?
References
(1) Wolters V (1999) Terrestrische Ökosysteme. Spektrum der Wissenschaft 2/1999 (A), 35-39.(2) Troumbis A, Beier C, Wolters V, Peter D (2001) Terrestrial ecosystem research in Europe: Lessons from the European
Union’s research policy 1996-2001. Environmental Science & Policy 4, 51 – 58.(3) Brown JH, Morgan Ernest SK, Parody JM, Haskell JP (2001) Regulation of diversity: maintenance of species richness in
changing environments. Oecologia 126, 321-332.(4) Wolters V et al. (2000) Effects of global changes on above- and belowground biodiversity in terrestrial ecosystems:
Implications for ecosystem functioning. BioSience 50, 1089-1098.(5) Sala OE et al. (2000) Global biodiversity scenarios for the year 2100. Science 287, 1770-1774.

14
BIOLOG AFRICA
BIODIVERSITY MONITORING TRANSECT ANALYSIS IN AFRICA
BIOTA AFRICA (Biodiversity Monitoring Transect Analysis in Africa) is the joint frame for thethree BIOLOG projects, BIOTA West Africa, BIOTA East Africa and BIOTA Southern Africa, andaltogether includes 34 sub-projects.
BIOTA AFRICA is the synonym for the corporate identity of these BIOLOG projects and sub-projects, aiming at coherent research concepts and monitoring activities in various regions ofAfrica. During the planning phase, a number of basic paradigms were defined which finallyresulted in a large number of research proposals:
• The programme puts a major focus on the assessment, understanding and prediction of thechange of biodiversity on the African continent, in the context of global environmental changeand impacts of human land use. In the main phase, approaches towards a scientifically basedmanagement and monitoring of biodiversity will gain higher importance. The programme willnot be restricted to one in-depth case study, but will aim at comparable results derived fromvarious scales and different regions.
• In order to record the driving factors controlling biodiversity, research sites were selected alongthe predominant environmental gradients, allowing better comparability by standardised scalesand measurement techniques.
• The programme will strengthen the impulse towards the initiation of a global network ofstandardised biodiversity observation sites.
• Biodiversity in various land use systems is being analysed, monitored and compared withnatural systems.
• Continental scale and ecosystem scale will be linked by suitable research structures that allowan up- and down-scaling.
• The major biomes of the continent will be represented.
• Special emphasis is placed on research close to the boundaries or transition zones betweenmajor biomes, as these regions might show more rapid changes of biodiversity.
• The possibility of the extrapolation of results will be optimised by putting a clear focus on zonalecosystems of a larger spatial representation.
• The strong interdisciplinary structure of the project aims at the development of integrative,applied management and monitoring concepts.
Based on these goals and paradigms, as well as on the already existing disciplinary and regionalcompetence and co-operation, a regional concentration along two large continental transects inSouthern and Western Africa was combined with regional investigations into altitudinal gradients,aridity as well as land use intensities in Eastern Africa and Southern Arabia.
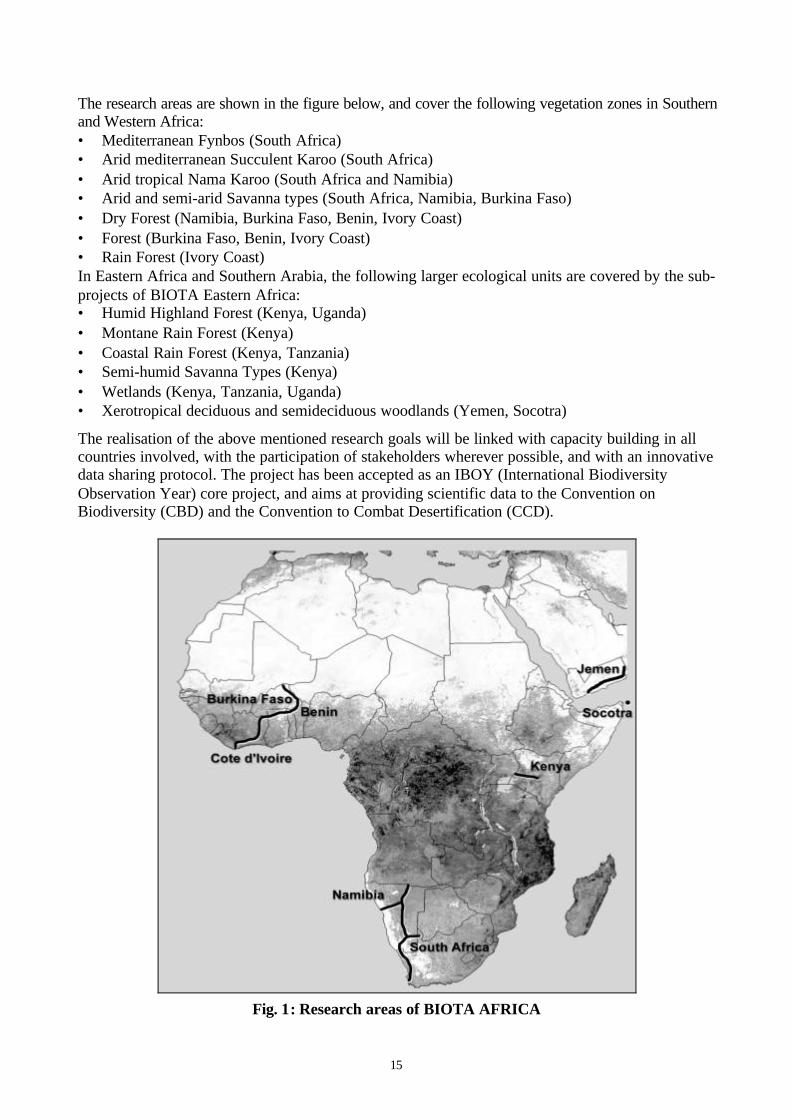
15
The research areas are shown in the figure below, and cover the following vegetation zones in Southernand Western Africa:• Mediterranean Fynbos (South Africa)• Arid mediterranean Succulent Karoo (South Africa)• Arid tropical Nama Karoo (South Africa and Namibia)• Arid and semi-arid Savanna types (South Africa, Namibia, Burkina Faso)• Dry Forest (Namibia, Burkina Faso, Benin, Ivory Coast)• Forest (Burkina Faso, Benin, Ivory Coast)• Rain Forest (Ivory Coast)In Eastern Africa and Southern Arabia, the following larger ecological units are covered by the sub-projects of BIOTA Eastern Africa:• Humid Highland Forest (Kenya, Uganda)• Montane Rain Forest (Kenya)• Coastal Rain Forest (Kenya, Tanzania)• Semi-humid Savanna Types (Kenya)• Wetlands (Kenya, Tanzania, Uganda)• Xerotropical deciduous and semideciduous woodlands (Yemen, Socotra)
The realisation of the above mentioned research goals will be linked with capacity building in allcountries involved, with the participation of stakeholders wherever possible, and with an innovativedata sharing protocol. The project has been accepted as an IBOY (International BiodiversityObservation Year) core project, and aims at providing scientific data to the Convention onBiodiversity (CBD) and the Convention to Combat Desertification (CCD).
Fig. 1: Research areas of BIOTA AFRICA

16
BIODIVERSITY INFORMATICS IN THE BIOLOG PROGRAMME
Walter G. Berendsohn
Department of Biodiversity Informatics and Laboratories, Botanic Garden and Botanical MuseumBerlin-Dahlem, Freie Universität Berlin, Königin-Luise-Str. 6-8, D-14191 [email protected]; http://www.bgbm.org/BioDivInf/
Key words: biodiversity informatics, BIOLOG programme
Abstract:This article briefly introduces the field of Biodiversity Informatics in general and details the role of thisdiscipline within the BIOLOG programme.
Introduction “Biodiversity Informatics” circumscribes the application of IT tools and technology to biodiversityinformation, principally at the organismic level. It thus deals with information capture, storage,provision, retrieval, and analysis, focusing on individual organisms, populations, and taxa, and theirinteraction. It covers the information generated by the fields of systematics (including molecularsystematics), evolutionary biology, population biology, behavioural sciences, and synecological fieldssuch as pollination biology, parasitism, and phytosociology. Biodiversity Informatics is considered apart of biological informatics sandwiched between - and strongly overlapping with - environmentalinformatics and molecular bioinformatics. [1]
BIOLOG and Biodiversity InformaticsThe choice to include a section titled Biodiversity Informatics in the BIOLOG Programme [2] wasundoubtedly influenced by the German government’s decision to become a founding member of theGlobal Biodiversity Information Facility (GBIF). The rationale for the GBIF [3] also applies toBIOLOG: vast information resources have to be accessed, and are produced, by a biodiversity researchprogramme of BIOLOG’s scale. An effective electronic information infrastructure is the prerequisitefor an efficient dissemination of results and a sustained effect of the invested research funds. Effectiveaccess to past research results and to data resources like collections is essential because it will providethe necessary background for knowledge gain from the new results. The information provided has to beof high quality, it must be accessible, relevant, accurate, complete, referenced, and affordable. [4]BIOLOG’s sub-programme on biodiversity informatics is to explore and develop the very substanceof information processing present in the German biodiversity research infrastructure. It is a showcasefor the biodiversity research community’s ability to co-operate, both within the community and withrelated fields such as geography and climatology. With its explicit funding focus on biodiversityinformatics, BIOLOG treads new ground. The US, Australia, Mexico and the UK, among others,have started funding such research much earlier than the German government, but on a project-by-project basis. However, the results are so convincing that, for example, the Australian government isnow providing Au$ 10 million to digitise all Australian herbarium specimens over the next 5 years inthe Australian Virtual Herbarium [5].The novelty of the topic in Germany is perhaps also the reason for the explicit limitation of the sub-programme to 3 years, while the rest of BIOLOG is seen as a potential a long-term programme. Thestated intention is to integrate biodiversity informatics research into the general terrestrial biodiversityresearch programme. In some cases this has already happened: there are several projects within BIOTAand BIOLOG-Europe with a distinct focus on biodiversity informatics. Further build-up of theinfrastructure and the provision of base data is to be covered by other programmes such as GBIF-

17
Germany, and eventually the biodiversity informatics infrastructure is to be institutionalised to ensurelong term sustainability.For the planned creation of a system of German GBIF nodes [6], we can already draw on the results ofseveral projects in the sub-programme. The following EU-funded projects are also directly supportedby collection-oriented activities in the BIOLOG programme: ENHSIN (The European Natural HistorySpecimen Information Network [7]), BioCASE (A Biological Collection Access Service for Europe[8]), and the forthcoming ENBI (European Network of Biodiversity Information) [9].
Presentation of projectsThe sub-programme on biodiversity informatics consists of 7 main projects, 4 of which are more or lessclosely-knit assemblages of sub-projects; in total 24 individual projects are funded. Most of theinvestigators participating in the ongoing projects were brought together in 3 workshops to discussoptions for collaboration and for the presentation of the sub-programme in the Statusseminar. For the 3time slots available within the conference’s programme, workshop participants generally preferred acoordinated thematic focus to a sequence of individual project presentations. The three subject areasselected cover (i) the discovery and identification of organisms with various computer-aided methods,(ii) the documentation and data mining of the enormous information resources represented by biologicalcollections in Germany, and (iii) a summary of efforts directed at the interoperability and networking ofthe project results as well as their inclusion and interrelation with international projects such as theaforementioned European initiatives and GBIF. This selection of activities is to be supported by a rangeof horizontal subjects, such as software development, georeferencing, IPR, and data quality aspects.After little more than a decade of development, the field of Biodiversity Informatics is still in itsinfancy. Presently the overriding objective – also within BIOLOG - is to provide a sound andeffective information management infrastructure for biodiversity and Global Change research. TheBIOLOG sub-programme is already making an impact in that respect. Although manymethodological, operational, and research questions remain to be solved, BIOLOG represents animportant step towards the realization of a vision: mobilizing existing biodiversity informationresources and the application of new methods of analysis to that information will stimulate newinsight into the evolution, functioning and role of ecosystems and their components.
References:1. Berendsohn, W.G. 2001: Biodiversity informatics. Pp. 89-94 in: Contributions to Global ChangeResearch: A Report by the German National Committee on Global Change Research. Bonn.2. BMBF 2000: Federal Ministry of Education and Research. Announcement of the Funding RegulationsGoverning "Biodiversity and Global Change (BIOLOG)" under the Federal Government's Programme on"Research for the Environment" of 7 April 1999.. – Bonn.3. GBIF 2000: Business Plan for the Global Biodiversity Information Facility. Discussion Draft 5. –Canberra. [ http://www.gbif.org ]4. Berendsohn, W. G. 2001 ["1998"]: Problems and perspectives in biological databasing. In: Feral, J. P.(ed.): Concepts and methods for studying marine biodiversity. Oceanis 24: 319-334.5. Fagg, E. 2001 [Oct. 26]: Australia’s Virtual Herbarium. Canberra. [http://www.anbg.gov.au/avh.html]6. Berendsohn W. G. (ed.) 2001 [Oct. 12]: GBIF-D. Berlin. [www.gbif.de]7. NHM 2001: ENHSIN. London. [http://www.nhm.ac.uk/science/rco/enhsin/]8. BioCASE 2001: BioCASE work programme on-line. [www.biocase.org]9. ENBI 2001: European Network for Biodiversity Information, Information Site. Amsterdam.[http://www.faunaeur.org/enbi/info.html]

18

19
BIOLOG EUROPE

20
Project ID: 01LC00021.3.2001 - 31.12.2003
GLOBAL CHANGE-DEPENDENT SPREAD OF NEOPHYTES: RELATIONSHIP TOECOSYSTEM CHANGE (CASE STUDY: FLOODPLAINS OF THE RIVER RHINE)
Rainer Lösch, Ulf Schmitz, Guido Dericks
Abt. Geobotanik, H. Heine-Universität Düsseldorf, Universitätsstr. 1, D-40 225 Dü[email protected]; http://www.geobotanik.uni-duesseldorf.de
Key words : neophytes, floodplains, Lower Rhine, C4 plants, increase of vegetation period,Amaranthus, Solanum, Impatiens, plant productivity, competition, river flood regimes
Abstract:Biodiversity changes due to increased neophyte colonization of river banks and floodplains in the LowerRhine area are described and their impact upon ecosystem peculiarities assessed.Generally, these habitatsshow high short-term changes of a-diversity from disturbances by the river activity and diaspore inputs viathe waterway. Under the global change effects of extended vegetation periods due to increased averagetemperatures and of changed flood regimes, the spread of neophytes in these habitats is intensified. Theimpact of these processes on the floodplain vegetation can be indicative for future trends and for per-spectives of stability of this ecosystem complex. .Since few decennia the percentage of C4 centrospermic taxa and of other neophytes of subtropical origin onthe river banks has considerably increased. In other floodplain vegetation units high growing neophytes arespreading. This is mapped, and the competition dynamics of autochthonous taxa and of already establishedresp. of recently spreading neophytes is analyzed both by detailed biomass harvests in the floodplain vege-tation units and by pairwise cultivation of relevant species pairs under controlled conditions (oblique-window rhizotrons, moisture gradient beds). Nutrient relations both of the soils and the plants are quanti-fied, the impact of submergence, waterlogging or drought upon the neophytic species is measured by eco-physiological methods, and the environmental requirements for growth of these taxa are determined (ger-mination studies, productivity measurements, biometric measures). First results after half a year durationof the project are presented as examples for the approaches of this study.
Results:Mapping of the Rhine river bank vegetation of the Lower Rhine area with emphasis on neophytecolonization is underway. A locality-point map of Amaranthaceae and Chenopodiaceae on the
Fig. 1: Centrospermic taxa on Lower Rhine river banks
Rhine river banks between Dormagen andDuisburg is given in Fig. 1. If relevées from thesecond half of the 20th century are com-pared(Tab. 1), the increase of neophytes of thesefamilies is obvious. Only Amaranthusretroflexus from temperate North America isnaturalized in the area since the 19th century.The other taxa, with meridional/subtropicalorigins started spreading only two decenniaago. Another nine Amaranthus spp. of tropi-calorigin have not yet left the bridgeheads ofharbors. A similar situation is given withvarious Solanum species. The floodplain forestunderstory is, among other neophyte facies,dominated by four Impatiens species. Only I.noli-tangere is indigeneous, and in one location(KNEBEL 1998) it meets since
21
few years I. capensis, the vicariant taxon fromtemperate North America: It would be a uniquecase of documentation if we could follow in thenext years a hybridogenic introgression amongthis trans-atlantic species pair from the verybeginning onwards.
Tab. 1: Number of neophytic Amaranthaceae andChenopodiaceae in Polygono-Chenopodietumrelevées alongside the Middle and Lower Rhineriver between 1964 and 2001.Year Source Number of
species1964 LOHMEYER & SUKOPP 1992 2 A.blitum
A.retroflex
1983 MAY 1988 2 A.blitumA.retroflex.
1986 - 89
WISSKIRCHEN 1995 6 A. albusA.blitoidesA.blit:2 sspA.buchoniiA.powellii
1989 LOHMEYER & SUKOPP 1992 6 A. albusA.blitoidesA.blitumA.buchoniiA.powelliiA. retrofl.
1998-
2001SCHMITZ, unpubl. 10 A. albus
A.blitoidesA.blitumA.buchoniiA.powelliiA.retroflex.A.tubercul.Ch. botrysCh. pumilioCorisp. lept.
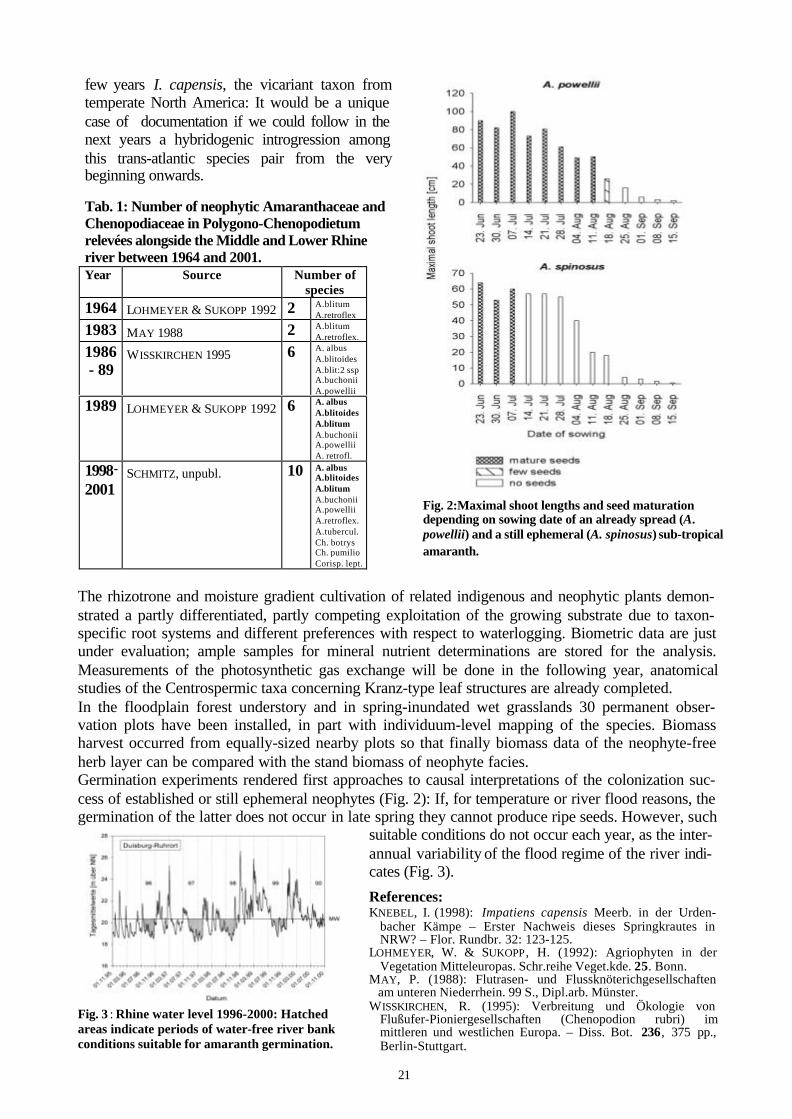
Fig. 2:Maximal shoot lengths and seed maturationdepending on sowing date of an already spread (A.powellii) and a still ephemeral (A. spinosus) sub-tropicalamaranth.
The rhizotrone and moisture gradient cultivation of related indigenous and neophytic plants demon-strated a partly differentiated, partly competing exploitation of the growing substrate due to taxon-specific root systems and different preferences with respect to waterlogging. Biometric data are justunder evaluation; ample samples for mineral nutrient determinations are stored for the analysis.Measurements of the photosynthetic gas exchange will be done in the following year, anatomicalstudies of the Centrospermic taxa concerning Kranz-type leaf structures are already completed.In the floodplain forest understory and in spring-inundated wet grasslands 30 permanent obser-vation plots have been installed, in part with individuum-level mapping of the species. Biomassharvest occurred from equally-sized nearby plots so that finally biomass data of the neophyte-freeherb layer can be compared with the stand biomass of neophyte facies.Germination experiments rendered first approaches to causal interpretations of the colonization suc-cess of established or still ephemeral neophytes (Fig. 2): If, for temperature or river flood reasons, thegermination of the latter does not occur in late spring they cannot produce ripe seeds. However, such
Fig. 3 : Rhine water level 1996-2000: Hatchedareas indicate periods of water-free river bankconditions suitable for amaranth germination.
suitable conditions do not occur each year, as the inter-annual variability of the flood regime of the river indi-cates (Fig. 3).
References:KNEBEL, I. (1998): Impatiens capensis Meerb. in der Urden-
bacher Kämpe – Erster Nachweis dieses Springkrautes inNRW? – Flor. Rundbr. 32: 123-125.
LOHMEYER, W. & SUKOPP , H. (1992): Agriophyten in derVegetation Mitteleuropas. Schr.reihe Veget.kde. 25. Bonn.
MAY, P. (1988): Flutrasen- und Flussknöterichgesellschaften am unteren Niederrhein. 99 S., Dipl.arb. Münster.WISSKIRCHEN, R. (1995): Verbreitung und Ökologie von
Flußufer-Pioniergesellschaften (Chenopodion rubri) immittleren und westlichen Europa. – Diss. Bot. 236, 375 pp.,Berlin-Stuttgart.

22
Project ID: 01LC000401.11.2000–31.10.2003
RECONSTRUCTING HISTORICAL BIODIVERSITY FROM ARCHIVALSOURCES
Dr. Peter-M. Steinsiek, Dipl.-Biol. Antje Jakupi, Prof. Dr. Bernd Herrmann
Institut für Zoologie und Anthropologie, Abteilung Historische Anthropologie undHumanökologie, Georg-August-Universität Göttingen, Bürgerstraße 50, D-37073 Gö[email protected]; http://www.anthro.uni-goettingen.de
Key words: biodiversity, nature conservation, environmental history, Brandenburg (Germany)
Abstract:The research project aims at developing a tool for nature restoration to make more reliablejudgements on the diversity of ecosystems by considering historical biological information.For that, proxy-data are made available out of archival sources and, standards are setaccording to which historical species stocks can be compiled from archival sources, bechecked and be used as references for changes of biodiversity in the past. By taking alandscape drained in the 18th century (Oderbruch, Brandenburg) as an example, we aredeveloping a method which guarantees a systematic approach to the relevant archivalsources and which allows their efficient analysis.
Results:So far, the stocks of archives have not been consulted for measuring historical biodiversity. Incomparison with exclusively temporary data collections on biodiversity, they represent, however,an irreplaceable source of information for reconstructing the conditions of nature in the past. Theproject’s aim is to trace back the functional changes certain elements of biodiversity fulfilled forhuman needs by looking at historical examples at different stages in the past. Apart from that, it isthe project’s aim to determine parameters for assessing shifts of biodiversity. By following bothaims the project addresses a deficiency of relevant research. At the same time, we will gain someinformation on how reliable historical data are for the reconstruction of the fauna and flora of thepast.
In our research, the land reclamation measures in 18th century Prussia and their impacts on theexistence of plants and animals serve as an example. Land reclamation mainly served the goal toraise agricultural production and to create new land for foreign colonists to settle on. The processof land transformation lasted many years and transformed our sample region, the Oder marshlands– situated east of Berlin and in the state of Brandenburg – which were formerly mainly used byfisheries, into fertile grass and arable land. This resulted in considerable alterations of therespective ecosystems. After more than two hundred years, we still do not know which elementsof fauna and flora actually disappeared.
Our investigations focus mainly on the stocks of the former Prussian State Archives in Potsdamand Berlin-Dahlem. The significance of archival records relating to certain questions on matters ofhistorical species diversity varies considerably and thus, the records need to be classified toguarantee that they can be used in an efficient way. For the adequate processing of archivalsources we are developing a catalogue of features which should allow to distinguish betweentypes of sources which contain a lot of information relevant to us and types of sources which areirrelevant.
23
In general, there are no proper historical lists of species which will give us an immediate idea ofthe fauna and flora of the “Oderbruch” at the time of its melioration. Therefore, it is necessary toput the main emphasis on Indicator Species which point to the existence of other species withsimilar necessaries of life. The concept of Keystone Species allows to concentrate the searchspecifically on those species the existence of which determines significantly the structures andfunctions of an ecosystem.
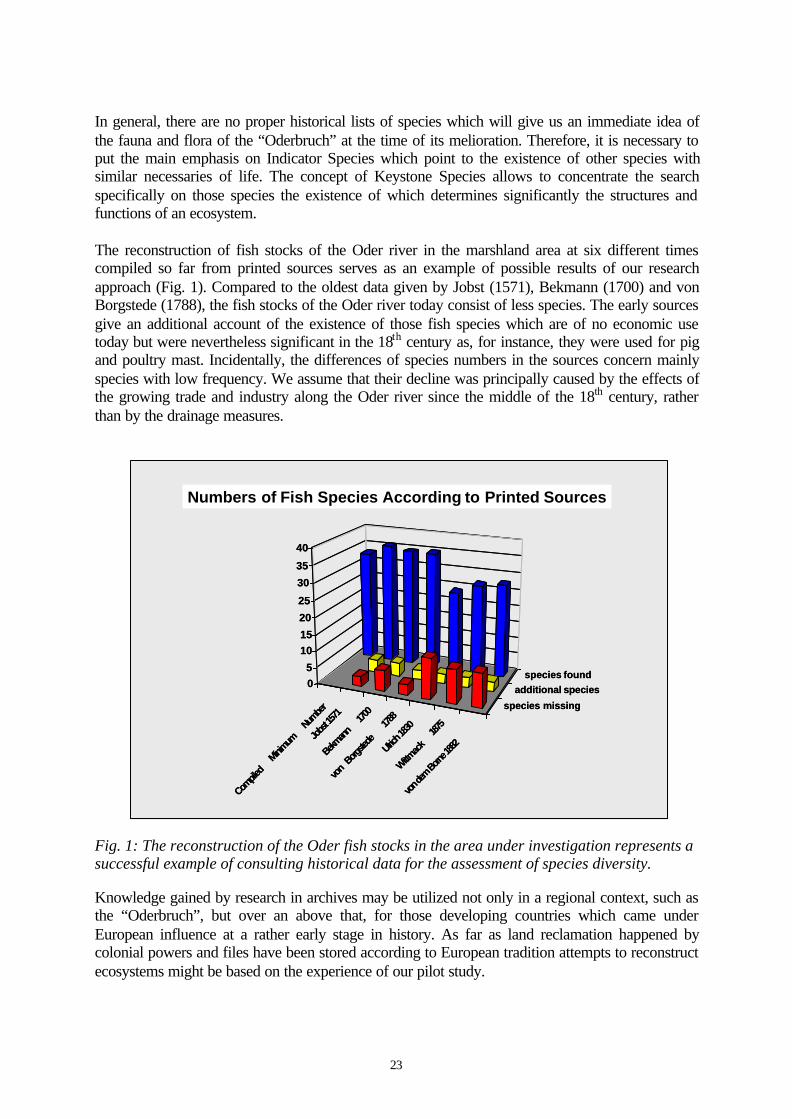
The reconstruction of fish stocks of the Oder river in the marshland area at six different timescompiled so far from printed sources serves as an example of possible results of our researchapproach (Fig. 1). Compared to the oldest data given by Jobst (1571), Bekmann (1700) and vonBorgstede (1788), the fish stocks of the Oder river today consist of less species. The early sourcesgive an additional account of the existence of those fish species which are of no economic usetoday but were nevertheless significant in the 18th century as, for instance, they were used for pigand poultry mast. Incidentally, the differences of species numbers in the sources concern mainlyspecies with low frequency. We assume that their decline was principally caused by the effects ofthe growing trade and industry along the Oder river since the middle of the 18th century, ratherthan by the drainage measures.
Fig. 1: The reconstruction of the Oder fish stocks in the area under investigation represents asuccessful example of consulting historical data for the assessment of species diversity.
Knowledge gained by research in archives may be utilized not only in a regional context, such asthe “Oderbruch”, but over an above that, for those developing countries which came underEuropean influence at a rather early stage in history. As far as land reclamation happened bycolonial powers and files have been stored according to European tradition attempts to reconstructecosystems might be based on the experience of our pilot study.
Compile
dMinim
umNum
ber
Jobs
t 1571
Bekm
ann
1700
von
Borgs
tede
1788
Ulrich
1830
Wittmac
k187
5
von de
m Borne
1882
species missing
additional speciesspecies found
05
1015
20
25
30
35
40
Compile
dMinim
umNum
ber
Jobs
t 1571
Bekm
ann
1700
von
Borgs
tede
1788
Ulrich
1830
Wittmac
k187
5
von de
m Borne
1882
species missing
additional speciesspecies found
05
1015
20
25
30
35
40
Numbers of Fish Species According to Printed Sources
24
Project ID: 01LC000501.10.2000 – 30.09.2002
BIODIVERSITY CHANGE DURING 20 YEARS OF SUCCESSION: IMPLICATIONSFOR MANAGEMENT OF RESTORATION
Hartmut Koehler, Josef Müller
Universität Bremen, Zentrum für Umweltforschung und Umwelttechnologie (UFT)Leobener Str., 28359 [email protected]; http://www.uft.uni-bremen.de/oekologie/hartmutkoehler.2.htm
Key words: succession, long-term monitoring, biodiversity, vegetation, seed bank, soil fauna, Gamasina,restoration, management
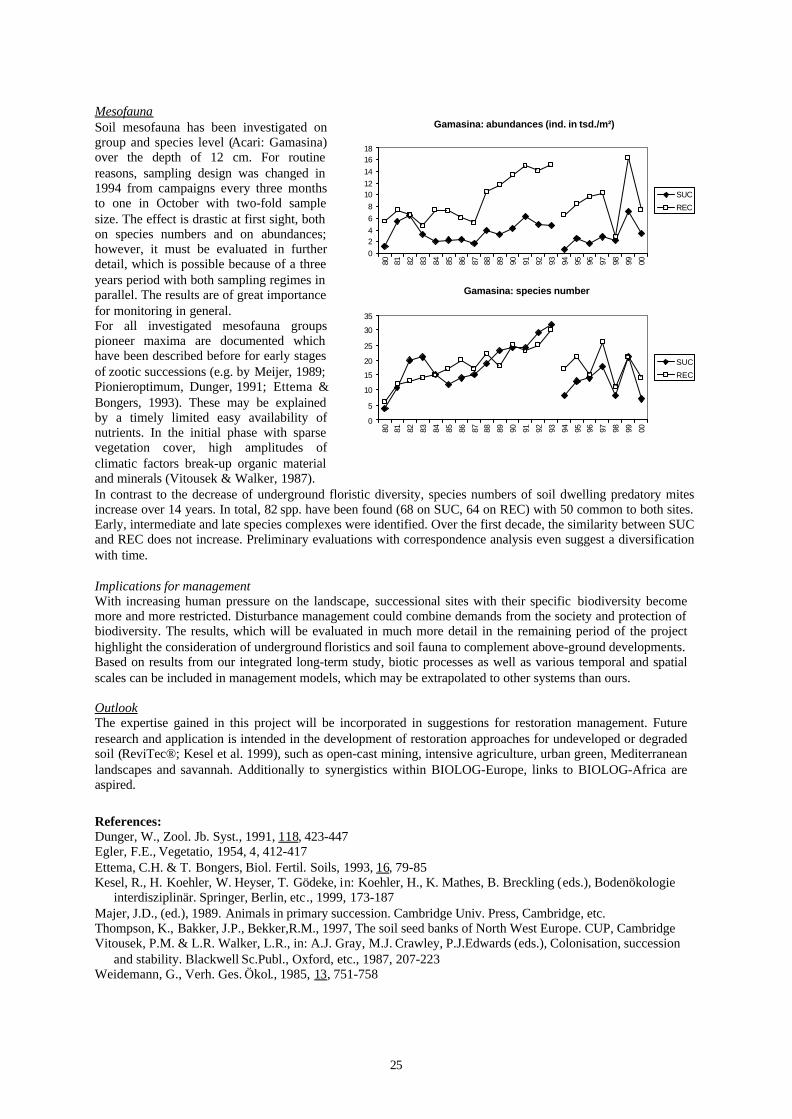
Abstract:The succession of soil mesofauna and vegetation has been studied since 1980 on two experimental plots, one leftfor undisturbed succession, the other managed as a lawn until 1987 (thereafter ruderalization). The resultsdocument very different patterns for soil mesofauna and vegetation, both at group and species levels. Within thefirst three years, recultivation leads to a transitory pioneer-maximum, followed by a steady increase over morethan 10 years. The development of vegetation depends on historical conditioning of the seed bank, whereassurvivors of fauna are of minor importance. Although above- and below-ground plant structure influence thedevelopment of soil mesofauna, their dynamics are not synchronised. Diversity of predatory mites (Acari,Mesostigmata: Gamasina) is increasing over long periods of time, whereas the seed bank is impoverished.Restoration management with focus on biodiversity should consider processes in the soil, and allow or introducepatchiness diversified in space and time.
Results:The former rubble-and-debris dump in the vicinity of Bremen (Northern Germany) was covered in the late1970ies with loamy sand (C= 1.6 – 3-5; N= 0.1 – 0.3). In 1980, the plateau was graded and two plots wereprepared, one with standard recultivation measures (REC), the other was left for natural succession (SUC)(Weidemann 1985).
VegetationThe development of the vegetation was studied on permanent plots and with germination experimentssynchronously to succession, in the greenhouse and also in the field.
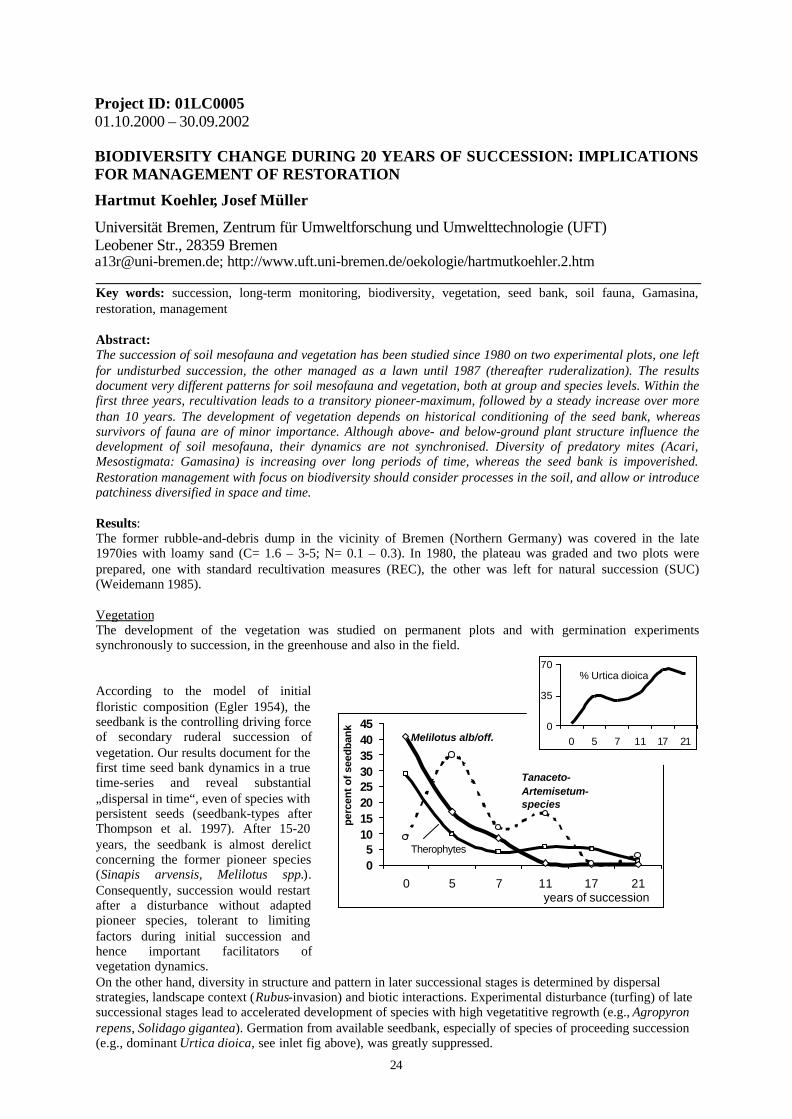
According to the model of initialfloristic composition (Egler 1954), theseedbank is the controlling driving forceof secondary ruderal succession ofvegetation. Our results document for thefirst time seed bank dynamics in a truetime-series and reveal substantial„dispersal in time“, even of species withpersistent seeds (seedbank-types afterThompson et al. 1997). After 15-20years, the seedbank is almost derelictconcerning the former pioneer species(Sinapis arvensis, Melilotus spp.).Consequently, succession would restartafter a disturbance without adaptedpioneer species, tolerant to limitingfactors during initial succession andhence important facilitators ofvegetation dynamics.On the other hand, diversity in structure and pattern in later successional stages is determined by dispersalstrategies, landscape context (Rubus-invasion) and biotic interactions. Experimental disturbance (turfing) of latesuccessional stages lead to accelerated development of species with high vegetatitive regrowth (e.g., Agropyronrepens, Solidago gigantea). Germation from available seedbank, especially of species of proceeding succession(e.g., dominant Urtica dioica, see inlet fig above), was greatly suppressed.
05
1015202530354045
0 5 7 11 17 21
Melilotus alb/off.
Tanaceto-Artemisetum-species
per
cent
of
seed
bank
years of succession
Therophytes
% Urtica dioica
0
35
70
0 5 7 11 17 21
25
MesofaunaSoil mesofauna has been investigated ongroup and species level (Acari: Gamasina)over the depth of 12 cm. For routinereasons, sampling design was changed in1994 from campaigns every three monthsto one in October with two-fold samplesize. The effect is drastic at first sight, bothon species numbers and on abundances;however, it must be evaluated in furtherdetail, which is possible because of a threeyears period with both sampling regimes inparallel. The results are of great importancefor monitoring in general.For all investigated mesofauna groupspioneer maxima are documented whichhave been described before for early stagesof zootic successions (e.g. by Meijer, 1989;Pionieroptimum, Dunger, 1991; Ettema &Bongers, 1993). These may be explainedby a timely limited easy availability ofnutrients. In the initial phase with sparsevegetation cover, high amplitudes ofclimatic factors break-up organic materialand minerals (Vitousek & Walker, 1987).In contrast to the decrease of underground floristic diversity, species numbers of soil dwelling predatory mitesincrease over 14 years. In total, 82 spp. have been found (68 on SUC, 64 on REC) with 50 common to both sites.Early, intermediate and late species complexes were identified. Over the first decade, the similarity between SUCand REC does not increase. Preliminary evaluations with correspondence analysis even suggest a diversificationwith time.
Implications for managementWith increasing human pressure on the landscape, successional sites with their specific biodiversity becomemore and more restricted. Disturbance management could combine demands from the society and protection ofbiodiversity. The results, which will be evaluated in much more detail in the remaining period of the projecthighlight the consideration of underground floristics and soil fauna to complement above-ground developments.Based on results from our integrated long-term study, biotic processes as well as various temporal and spatialscales can be included in management models, which may be extrapolated to other systems than ours.
OutlookThe expertise gained in this project will be incorporated in suggestions for restoration management. Futureresearch and application is intended in the development of restoration approaches for undeveloped or degradedsoil (ReviTec®; Kesel et al. 1999), such as open-cast mining, intensive agriculture, urban green, Mediterraneanlandscapes and savannah. Additionally to synergistics within BIOLOG-Europe, links to BIOLOG-Africa areaspired.
References:Dunger, W., Zool. Jb. Syst., 1991, 118, 423-447Egler, F.E., Vegetatio, 1954, 4, 412-417Ettema, C.H. & T. Bongers, Biol. Fertil. Soils, 1993, 16, 79-85Kesel, R., H. Koehler, W. Heyser, T. Gödeke, in: Koehler, H., K. Mathes, B. Breckling (eds.), Bodenökologie
interdisziplinär. Springer, Berlin, etc., 1999, 173-187Majer, J.D., (ed.), 1989. Animals in primary succession. Cambridge Univ. Press, Cambridge, etc.Thompson, K., Bakker, J.P., Bekker,R.M., 1997, The soil seed banks of North West Europe. CUP, CambridgeVitousek, P.M. & L.R. Walker, L.R., in: A.J. Gray, M.J. Crawley, P.J.Edwards (eds.), Colonisation, succession
and stability. Blackwell Sc.Publ., Oxford, etc., 1987, 207-223Weidemann, G., Verh. Ges. Ökol., 1985, 13, 751-758
Gamasina: species number
0
5
10
15
20
25
30
35
80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 00
SUC
REC
Gamasina: abundances (ind. in tsd./m²)
02468
1012141618
80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 00
SUC
REC

26
Project ID: 01LC00061.4.2001 – 31.12.2003
ECOLOGY AND BIODIVERSITY OF HIGHER FUNGI IN CENTRAL EUROPE(GERMANY)
A. Bresinsky, L. Krieglsteiner, C. Düring
Institut für Botanik, Universität Regensburg, 93040 [email protected] (Prof. Dr. A. Bresinsky, Am Katzenbichl 22, D-93161 Sinzing)_______________________________________________________________________________
Key words: database, PILZOEK, fungi, Basidiomycetes, Hymenomycetes, Germany, Rhön,biosphere reservation, biodiversity, ecological parameters, chorology, mycoflora
Abstract: Studies on fungal diversity in regard to ecological and chorological parametersare carried out in the biosphere reservation Rhön and by the aid of the database PILZOEKin general.
Results:The database PILZOEK has been developed in order to collect and to compute data connectedwith fungi. It is possible to establish lists of fungi in association with various other data likedifferent nutritional behaviour, substrates, habitats, soil characters, geology, microclimate,chorological features, grid square frequency, red list status, occurrence in different areasdefined by their geological and geomorphological background (“Naturraum”; landscapenatural unit) and presence or absence in the different federal states of Germany. Informationfrom the database can be drawn in whatever combination. The database PILZOEK has beenchecked for its capability to store and to recall data from literature and the field as well.Nearly 1000 different sets of data have been established so far. The reference list of thedatabase is available viawww.biologie.uni-regensburg.de/Botanik/Bresinsky/Pilzoek_homepage.html
According to present calculation the 4185 species within Basidiomycetes occurring inGermany contribute to the different trophic groups as follows: parasites (19%), saprotrophes(48%) and symbionts (33%). Among the latter most of the species are involved inectomycorrhizal interrelationships between various trees and fungi. In case of theAscomycetes - the other main group within the Mycobionta - lack of knowledge about thepresence of species in Central Europe does not allow to present even tentative calculationsabout species diversity in different trophic groups. Even in our country which is thought to beexplored intensively, there is a considerable lack of knowledge regarding species diversity.This is an obstacle since fungi are important members of the different ecosystems for instanceas decomposers of wood and litter, as parasites on trees, herbals, mosses, other fungi andanimals and last but not least as symbionts interacting with higher plants, microorganisms andanimals.
The project being here in focus aims to contribute data to a “Mycoflora Germaniae Program(MGP)” with special emphasis on ecological and chorological data linked withBasidiomycetes (Hymenomycetes s. l.). The genus Hygrocybe will be treated in a pilot studyin the context of MGP and of the here discussed project. The expected approach in case ofHygrocybe and other fungi is based on the evaluation of widespread publications in variousjournals (partly also in books) and on a field work project in the biosphere reservation Rhönas well. Amazingly enough, the Rhön has been neglected by mycologists to a considerableextent so far.

27
Regarding field work in the Rhön, carried out by L. Krieglsteiner, 69 daily excursions havebeen undertaken with a total crop of 1881 observed fungal species, being collected,determined and in case of interest dried as herbarium specimens: Basidiomycetes (1128species; i.e. Boletales 49, Agaricales 662, Russulales 101, “Aphyllophorales” 284,Heterobasidiomycetes 109, “Gasteromycetes” 23), Ascomycetes (336; i.e. Pezizales 79,Leotiales and other groups 257), mitosporic and lower fungi (53), Myxomycetes (81). Quite anumber of those species are to be regarded as the first records for the Rhön, others for one ofthe three federal states (Bayern, Hessen, Thüringen) covered partially by the Rhön. Each ofthe fungal species has been recorded together with ecological data at the place offructification, i.e. substrate (including host species), vegetation (if possible identified as plantassociations) and other habitat characters, soil type, including acidity and nitrification.Fungi could be observed in about 70 different types of vegetation, 45 of them being identifiedas plant associations. Some of the latter apparently never have been examined for thepresence of fungi so far, like for example in case of Betuletum carpaticae, Caricetumappropinquatae, Caricetum paniculatae, Caricetum rostratae, Chaerophylletum temuli,Epilobietum angustifolii, Geranio-Trisetetum and others. The fungal components of thedifferent habitats will be evaluated in respect to various parameters, for example share ofendangered species in different trophic groups etc.
The observations in the Rhön will also be evaluated comparatively to the adjacent landscapenatural unit “Mainfränkische Platten”, treated in the doctoral thesis of L. Krieglsteiner.More then 16% of the species (i.e. rare species, species with concentration in montaneousregions, on acid soils, or sensitive to fungicides) observed in the Rhön have not been found inMainfränkische Platten with a total of 2600 observed fungal species. An example for fungibeing sensitive to fungicides, occurring in the Rhön, however missing in MainfränkischePlatten, are some of the smut fungi ( 6 of a total number of 13 species) like Microbotryummarginale and M. pustulatum, both on Polygonum bistorta as their host.
References:
Bresinsky, A., Düring, C. (2001): PILZOEK, ein Erfassungsprogramm für Daten zur Ökologie undChorologie von Pilzen in Mitteleuropa. Z. Mykol. 67: 157-168
Krieglsteiner, L. (1999): Pilze im Naturraum Mainfränkische Platten und ihre Einbindung in dieVegetation. Regensb. Mykologische Schr. 9: 1-901

28
Project ID: 01LC00101.1.2001-31.12.2003
CONDITIONS OF A RATIONAL BIODIVERSITY POLICY FOR GERMANY, AS APART OF THE EUROPEAN UNION
Andreas Hoffmann, Marko Ehlebe , Sönke Hoffmann, Stefanie Richter and JoachimWeimann
Lehrstuhl für Wirtschaftspolitik, Otto-von Guericke Universität Magdeburg. Postfach 4120, D-39016 [email protected] , http://www.uni-magdeburg.de/vwl3
Key words: biodiversity policy, cost-benefit analysis, conservation, opportunity costs
Abstract:The aim of the project is to determine the requirements of a rational biodiversity policy forGermany and its institutional framework within the European Union (EU). Methods for theevaluation of cost and benefits associated with biodiversity conservation activities will beanalysed. Furthermore, instruments for an implementation of a biodiversity policy based onrationality and efficiency will be developed.
Results:The global biodiversity loss is mainly an outcome of the human use of the biosphere. Exploitationof natural resources results in social welfare benefits. However, costs associated with biodiversityloss are widely ignored in cost-benefit analysis evaluating human activities.From an economic perspective, biodiversity is characterized by attributes (lacking property rights,public good) constituting a market failure. Therefore, decentralised decisions by autonomousagents, which are typical for markets, will not provide a solution to the biodiversity decline,instead governmental intervention is required. The aim of our project is to determine therequirements of such a rational biodiversity policy and its institutional framework. It is crucial toidentify the aims of such policies in order to use rationality as a criterion for evaluating alternativepolicies. Therefore, the determination of biodiversity functions influencing human well-being is aprerequisite.In a first step, we are trying to identify biodiversity measures, which could be used as a basis forthe evaluation of cost and benefits associated with biodiversity conservation activities.Therefore a preceding task is to explore latest theoretical approaches on measuring diversity ingeneral (1,2,3). After having outlined aspects about its essential theoretical characters we will beable to examine critically if some properties might be helpful in delimiting appropriate measuresof biodiversity. In this context an important and interesting objective is to find out how farbiodiversity can be a matter of calculus and where subjective valuations become inevitable. To acertain point biodiversity policy is always based on normative decisions. The question is to definethe limits of such a normative approach in order to establish the starting point for rationaldecisions in the context of real word policies. Another topic of interest is the spatial distribution ofbiodiversity (regional, national) and its potential for specialization and allocation of conservationefforts between regions and nations within the borders of the European Union. After theidentification of relationships between social welfare and biodiversity and establishing the

29
relevant policy goals, the appropriate instruments to achieve these goals in an economicallyefficient way have to be chosen. Standard methods in economic theory will be used to assess theapplicability of the chosen instruments.Based on the characterisation of a rational national biodiversity policy, also international aspectshave to be considered. The European Union assumed already many tasks related to biodiversityconservation. However, we have to examine if this centralisation is in accordance with ourproposed rational biodiversity policy. Another area of research will be the eastern extension of theEU. This extension provides a variety of perspectives and opportunities for a pan-Europeanbiodiversity strategy. The efficient use of these opportunities and the required institutionalcondition are of major importance. In addition, there is still the necessity to negotiate biodiversityconservation issues with non-EU countries. Analysing the available instruments to create self-enforcing treaties will be another point of interest.Preliminary analyses of strategies for the identification of priority areas for the conservation ofbiodiversity revealed the “complementarity approach” (4,5) as the most promising. In this processareas are ranked according their complementarity value, that is the degree to which a single areaor subset of areas represents the total number off biodiversity attributes found in the evaluatedsystem, or adds unrepresented attributes to already selected areas. Comparisons between theimplementation of the Habitat Directive of the EU and the complementarity approach inidentifying the most efficient network of areas for protection and specific management are inprogress. The analysis will focus on social-economic factors and their incorporation asopportunity costs in conservation planning methods.
References:1. Weitzman, M. L. 1992. On diversity. Quarterly Journal of Economics 58: 157-183.2. Faith, D.P. (1994): Phylogenetic pattern and the quantification of organismal biodiversity.–
Phil. Trans. Roy. Soc. Lond. B 345: 45-58.3. Nehring, K. & Puppe, C. (in press): A theory of diversity. – Econometrica.4. Vane-Wright, R. I., Humphries, C. J. & Williams, P. H. (1991) What to protect? - Systematics and
the agony of choice. - Biological Conservation 55: 235-254.5. Pressey, R. L., Humphries, C. J., Margules, C. R., Vane-Wright, R. I. & Williams, P. H. (1993)
Beyond opportunism: key principles for systematic reserve selection. - Trends in Ecology and Evolution 8:124-128.

30
Project ID: 01LC0012 (Subproject 1)01.10.2000 – 30.09.2003
EVOLUTIONARY CONSEQUENCES OF BIOLOGICAL INVASIONS –INTROGRESSIVE HYBRIDIZATION BETWEEN INVASIVE AND NATIVERORIPPA SPECIES (BRASSICACEAE).
Walter Bleeker and Herbert Hurka
Department of Systematic Botany, University of Osnabrück, Barbarastr. 11, 49076 Osnabrü[email protected]
Key words: AFLP, chloroplast DNA, evolution, introgression, invasive species
Abstract:The aim of the project is to assess the evolutionary consequences of interspecific gene flowbetween invasive and native plant species using the Brassicaceae as a model system. Firstresults of molecular analyses (cp DNA, AFLP) provide evidence for the spread of molecularmarkers of an invasive species (Rorippa austriaca) via native species (R. sylvestris) and theevolution of a new invasive taxon (R. ×armoracioides) via hybridization. Such ongoingprocesses may even be reinforced in the future when global warming and further habitatdestruction favour the invasion of R. austriaca and other invasive species into northern partsof Central Europe.
Results:Since several decades Rorippa austriaca is extending its range from southeastern Europe intoCentral Europe. Clonal growth provides the capacity to compete with indigenous Rorippaspecies. Hybridization is common in Rorippa (Bleeker & Hurka 2001) and can be expectedalong the invasive and native species contact zones since several hybrids between R.austriaca, R. sylvestris, and R. amphibia have been reported. One of these hybrids, R.×armoracioides (putative hybrid R. austriaca × R. sylvestris) is itself invasive in man-madehabitats in northern Germany and adjacent regions (Bleeker, in press). The aims of the projectare to study invasion dynamics of R. austriaca, to localize contact zones between R.austriaca and native Rorippa species, to analyse interspecific gene flow using molecularmarkers, and to assess the evolutionary consequencesSeveral native R. austriaca populations in the Tchech Republic, Hungary, and eastern Austriaand invasive R. austriaca populations in Germany have been visited in 2001. In southeasternEurope R. austriaca occurs in flood-plain meadows and at ruderal sites. Invasive R. austriacapopulations in Germany were predominantly found along disturbed river banks and in urbanand industrial sites. Contact zones between the invasive R. austriaca and the native R.sylvestris were detected along disturbed river banks of the rivers Ruhr and Main. Themorphologically intermediate R. ×armoracioides was found at several locations either with orwithout its putative parent species, R. austriaca and R. sylvestris.Maternally inherited chloroplast DNA (trnL intron) and biparentally inhereted AFLP markers(amplified fragment length polymorphism) were used to analyse interspecific gene flowbetween the invasive R. austriaca and native Rorippa species. First results provide substantialevidence for hybridization and introgression between R. austriaca and R. sylvestris. Analysesof noncoding chloroplast DNA (trnL intron, trnL/F spacer) showed that the trnL intron ofR.austriaca is characterized by a deletion of 169 bp in length. (Bleeker et al., accepted).R.austriaca shows a trnL intron length variant of approximately 420 bp, the native species R.amphibia, R. sylvestris, and R. palustris are characterized by a trnL intron length variant of

31
Fig. 1: Distribution of two trnL intron length variants in the invasive R. austriaca, R. ×armoracioides, and R. sylvestris from contact zones with R. austriaca (from Hurka et al, in press).
590 bp. The 420 bp length variant typical for R. austriaca was found in ten indivíduals of R.sylvestris which were collected in contact zones between both species indicating hybridizationand introgression of the R. austriaca chloroplast into R. sylvestris (Fig. 1). Themorphologically intermediate R. ×armoracioides showed either the chloroplast character of R.austriaca (26 individuals) or the character of the native species (16 individuals, Fig. 1).AFLP analysis provided further evidence for introgressive hybridization between the invasiveR. austriaca and the native R. sylvestris and the spread of R. austriaca characters via thehybrid R. ×armoracioides. Hybrid zones were localized at the river Ruhr near Mülheim and atthe river Main near Würzburg. 102 individuals were analysed so far. We were able to identifyseveral species specific AFLP markers of R. austriaca and R. sylvestris. In an invasive R.austriaca population at the river Ruhr, three hybrids could be detected showing an additivebanding pattern of diagnostic R. austriaca and R. sylvestris markers (Tab. 1).
ReferencesBleeker, W.: Entstehung und Etablierung hybridogener Sippen: Drei Fallbeispiele aus Nordwest-
deutschland. Schriftenreihe NEOBIOTA, in press.Bleeker, W. and Hurka, H. (2001): Introgressive hybridization in Rorippa (Brassicaceae): gene flow
and its conequences in natural and anthropogenic habitats. Molecular Ecology 10: 2013-2022.Bleeker, W., Weber-Sparenberg, C., and Hurka, H.: Chloroplast DNA variation and biogeography in
Rorippa (Brassicaceae). Plant Biology, accepted.Hurka, H., Bleeker, W., and Neuffer, B.: Evolutionary processes associated with biological invasions
in the Brassicaceae. Biological Invasions, in press.
Tab. 1: Distribution of selected diagnostic R. austriaca and R. sylvestris AFLP markers in three hybridscollected in an invasive R. austriaca population near Mülheim.
Primer combination /Fragment length
R. austriacainvasive
hybrid1
hybrid2
hybrid3
R. sylvestrisnative
Mse I-CTA / Eco RI-AAC52 bp - + + + +54 bp - + + + +56 bp + + + + -99 bp - + + + +103 bp + + + + -110 bp - + + + +112 bp + - - - -117 bp - + + + +137 bp - + + + +194 bp - + + + +236 bp - + + + +269 bp + + + + -
R. austriaca R. sylvestrisR. xarmoracioides

32
Project ID: 01LC0012 (Subproject 2)01.10.2000 – 30.09.2003
EVOLUTIONARY CONSEQUENCES OF BIOLOGICAL INVASIONS –ONGOING HYBRIDIZATION PROCESSES BETWEEN CARDAMINE TAXA ATTHE URNERBODEN, CENTRAL SWITZERLAND
Barbara Neuffer, Herbert Hurka, Ulrich Mönninghoff
Department of Systematic Botany, University of Osnabrück, Barbarastr. 11, 49076 Osnabrü[email protected]
Key words: AFLP fingerprinting, evolution, introgression, invasive species, vegetational records
Abstract:The aim of the project is to assess the evolutionary consequences of interspecific geneflowbetween invasive and native species using the Brassicaceae as a model system.Natural hybrids between Cardamine rivularis auct. non Schur (2n = 2x = 16; genome RR) andC. amara L. (2n = 2x = 16; genome AA) were discovered in 1971 at the Urnerboden inSwitzerland (Urbanska et al. 1997). The triploid hybrid C.x insueta Urbanska forms vigorouspopulations in man-made, fertilised hay meadows which were created early in the 20th
century. This hybrid species by polyploidisation gave rise to the the hexaploid C. schulziiUrbanska (genome constitution RRRRAA) which also occupies only man-made habitats(drainage ditches), and its origin may not be older than 50 - 70 years when farmers began todrain land. Molecular studies carried out in our group (cp- and nrDNA, isozymes, RAPD)confirmed the hybridisation event and established the precise parentage of C.x insueta(Neuffer & Jahncke 1997, Urbanska et al. 1997, Franzke & Mummenhoff 1999). We haveevidence for multiple origin, introgressive hybridisation and recurrent formation. In 1994, thelowland species C. pratensis L. s .str. (2n = 4x = 30) was first observed at the Urnerbodenand since then began to spread. The invasion of C. pratensis into the Urnerboden has alreadyled to the formation of contact zones with the native species. This might have direct orindirect evolutionary consequences. A new natural experiment in evolution has started at theUrnerboden and offers the rare opportunity for long term monitoring of the evolutionaryimpact of an invading species along altitudinal belts.
Results:Distribution of Cardamine pratensis and contact zones between C. pratensis and theindigenous Cardamine taxa have been carefully mapped, and permanent plots for long termmonitoring have been established. The plots enclose different species compositons withregard to the Cardamine species (Tab. 1). In each plot, the actual vegetation has been
Tab. 1: Field observations and sample numbers at Urnerboden, Central Switzerland, during the vegetation period2001. Ca = Cardamine amara, Cr = C. rivularis, Cp = C. pratensis, Cs = C. schulzii, Ci = C. x insueta; n =number of individuals; veg. survey = vegetational survey following Braun-Blanquet.
permanent plots 1a 1b 1c 1d 1e 2a 2b 2c 3a
species Ca/Cp Ca/Cp Ca/Cp Ca/Cr Ca/Cr/Cs Ci/Cr Cr/Ci/Cs Ca/Ci Cp
n (silica gel) 15/15 17/17 16/17 17/16 39/2/28 17/17 11/11/11 16/16 10
n (cytology) 5/5 5/5 5/5 5/5 5/5/5 5/5 5/5/5 5/5 5
n (seeds) 3/1 0/0 1/11 2/2 1/2/5 8/8 1/5/2 5/8 3
n (living plants) 2/2 2/0 0/0 0/0 2/0/2 3/0/0 0/3/3 0/0 2
veg. survey yes yes yes yes yes yes yes yes yes

33
analysed by the method of Braun-Blanquet.Samples from individual Cardamine plants within -and in addition also outside - the plotshave been collected for several purposes: for cytological studies, for AFLP fingerprinting andDNA sequencing, and seeds for fitness studies.
AFLP technique has been optimised for Cardamine. In a first experiment, we compared theAFLP banding patterns of living material, silica dried material, and herbarium material. Itturned out that silica dried leaves revealed high quality banding patterns comparable with theresults of fresh material. This is an important result which makes sampling a lot easier.In another set of experiments we searched for intra- and interspecific variation in the AFLPpatterns. Knowledge of the genetic variation is a prerequisite for analysing the evolutionaryprocesses at the Urnerboden. Summarising the results, we now know (1) that all speciesinvolved in the evolutionary scenario at the Urnerboden display species-specific AFLP bands,(2) that hybridisation and introgression can be detected, (3) that geographic variation withinspecies is obvious allowing to analyse colonisation events. Hybrid nature of C.x insueta andC. schulzii was verified by AFLP as with other molecular markers before, and analyses of C.pratensis contact zones are in progress.
ReferencesFranzke, A. & Mummenhoff, K (1999): Recent hybrid speciation in Cardamine
(Brassicaceae). Conversion of nuclear ribosomal ITS sequences in statu nascendi.Theor.Appl. Genet. 98, 831-834.
Hurka, H., Bleeker, W., Neuffer, B. (in press) Evolutionary processes associated withbiological invasions in the Brassicaceae. Biological Invasions.
Neuffer, B. & Jahncke, P. (1997): RAPD analyses of recent hybrid speciation events inCardamine (Brassicaceae). Folia Geobot. Phytotax. 32, 57-67.
Urbanska, K.M., Hurka, H., Landolt, E., Neuffer, B. & Mummenhoff, K. (1997):Hybridization and evolution in Cardamine L. (Brassicaceae) at Urnerboden (Switzerland):biosystematic and molecular evidence. Plant Syst. Evol. 204, 233-256.
Fig. 1: Distribution of the invasive Cardamine pratensis and the „indigenous“Cardamine species at Urnerboden in 2001.

34
Project ID: 01LC00131.1.2000 – 31.12.2003
BIODIVERSITY AND ECOSYSTEM FUNCTION IN GRASSLANDS - THEEFFECTS OF INSECT HERBIVORY AND DROUGHT STRESS
Prof. Dr. Wolfgang W. Weisser
Institut für Ökologie, Friedrich-Schiller-Universität Jena, Dornburger Str. 159, 07743 [email protected]; http://www.uni-jena.de/biologie/ecology/
Key words: Biodiversity, Ecosystem function, Nutrient cycling, Grassland, Experiment
Abstract:This project is a collaborative research effort of the Friedrich-Schiller-University and theMax-Planck-Institute for Biogeochemistry, both in Jena, and the Bureau for EcologicalStudies in Bayreuth/Erfurt. Using grasslands in Thuringia, Germany, as model ecosystems,the project is designed to investigate the relationship between genetic/phenotypic diversityand ecosystem processes such as carbon and nitrogen fluxes. Seven research groups in theareas of plant physiology, biogeochemistry, entomology, mathematical modelling, planttaxonomy, mycorrhizal biology, and landscape modelling will use empirical and theoreticalapproaches to investigate the importance of biodiversity for the stability and functioning ofecosystems. Characteristic for this research network will be a close cooperation between thevarious research groups for example by using the same sites for empirical investigations.
Results:In the past decades, it has been increasingly realised that species are disappering from ourplanet at an unprecedented rate. This extinction crisis has raised the question how the loss inbiological diversity affects the functioning of ecosystems. First experimental studies in thepast years have shown that a loss in biodiversity will have negative consequences forecosystem function. For example, a number of studies have shown that decreasing the numberof plant species in a grassland will decrease biomass production and will adversely affectother processes in the ecosystem. These studies have also made clear, however, that a generalunderstanding of the relationship between biodiversity and ecosystem processes can only begained if research programs include both various groups of organisms (e.g. animals, plants,microbes) and different levels of organisation (e.g. leaf, ecosystem, landscape) (Fig. 1, left).The present research network with its six subprojects therefore uses an interdisciplinaryapproach to address current scientific questions. Grasslands in Thuringia, Germany, will serveas model ecosystems and all research groups will focus their activities on the same set ofnatural and manipulated grasslands.
In an experimental approach (simulation of a disturbance) the project aims to unravelcausal relationships between the genetic and phenotypic diversity and ecological processes ingrasslands (Fig 1, middle). The project will be the first to investigate the importance of insectherbivory for ecosystem processes in stressed vs. unstressed ecosystems. In an exemplarymanner, various biological disciplines will work together to quantify both various aspects ofbiological diversity (subprojects MYKO, ENTO, GENMOD, STOFF) and importantecological processes (subprojects PHOTO, GENMOD, STOFF), using modern technology.In addition, the results obtained will be implemented in a spatial model that will findapplication in landscape planning procedures (subproject RAUM, Fig. 1, right).

35
Levels of organisation Research approach Subprojects in network
Fig. 1: Schematic view of research approach and links between subprojects in network
In the first year (2001) of the study, ca. 80 different grasslands in Thuringia at a range of plantspecies diversity will be visited in a collaborative effort of subprojects ENTO, GENMOD,STOFF and RAUM. The biodiversity of these grasslands will be charakterised, in particularplant species richness and abundance, vegetation structure and insect diversity. In addition, anumber of parameters that characterise ecosystem processes both above- and below ground(C- and N-cycling, temperature, humidity), the ecophysiology of dominant plant species (e.g.N- and C-use, leaf water potential) and the field sites (exposition, land use etc.) will bemeasured. The results of the study will reveal first insights into the relationship betweenbiodiversity and ecosystem function. The results will be implemented in a spatial model(RAUM). Further activities in the first year are the development of suitable methodology toinvestigate the genetic diversity of selected plant species in grasslands, and the developmentof a first mathematical model on the relationship between biodiversity and ecosystem function(GENMOD).
In the second and third year (2002 and 2003), results from the first year will be used to selectgrasslands at different levels of diversity in which experimental manipulations will be carriedout. In each of 20 blocks, the following manipulation will be carried out:
T- Water regime unchangedT+ Simulated droughtIns+ Insect herbivory unchangedIns- Insect herbivory reduced
In plots with where drought stress is applied, mobile roofs will be placed over the grasslandfor a short while in spring. In plots with reduced insect herbivory, insecticides will be appliedrepeatedly. All subprojects will collaborate in measuring ecosystem processes in allexperimental plots.
T -Ins +
T +Ins +
T +Ins -
T -Ins -
5 m
5 m
Soil
Landscape
Ecosystem
Insects ? Plants ?
Microorganisms
Indication
Diversity Processes
= Disturbance
RAUM
MYKOENTOGENMODSTOFF
PHOTOGENMODSTOFF

36
Project ID: 01LC0013 (Subproject STOFF)1.1.2001 – 31.12.2003
THE CARBON AND NITROGEN CYCLES IN GRASSLAND ECOSYSTEMS OFDIFFERENT DIVERSITIES
Ansgar Kahmen, PD Dr. Nina Buchmann
Max-Planck-Institut für Biogeochemie, P.O. Box 10 01 64, 07701, [email protected], [email protected]; http://www.bgc-jena.mpg.de
Key Words: biodiversity, ecosystem function, pools, fluxes, C, N, nutrients
Abstract:The subproject STOFF investigates the influence of biodiversity on bio-geochemical
ecosystem functions in well-established, managed grasslands, with particular emphasis on theeffect of plant and insect diversity on carbon and nitrogen pools and fluxes. In the first part ofthe project, relevant site and ecosystem parameters have been measured at 80 sites in theThuringian Schiefergebirge to evaluate the relationship between plant diversity and productivityat the regional scale. These data will also serve as criteria for the selection of experimental sitesin the second part of the project, when the influence of biodiversity on ecosystem resistance andresiliance will be tested at the local scale. First results show that plant diversity is lowest ingrasslands with high and low productivities, whereas grasslands with highest levels of plantdiversity show only average productivity.
Results:Research within the subproject STOFF is conducted at two different spatial scales. During
the 1st year, investigations are covering the entire region of the Thuringian Schiefergebirge(regional scale). During the 2nd and 3rd year, app. 20 sites will be selected for an experimentalapproach (local scale). During the first part of the project (2001), a well defined dataset has beencollected, to determine the relationship of biodiversity and ecosystem functioning at the regionalscale. Thus, data collection in summer 2001 at all 80 sites focused on the determination oftaxonomic and structural plant diversity (e.g., species composition, life forms, LAI, height, etc.)and ecosystem parameters (e.g., carbon and nitrogen pools in the soil and the vegetation, soil pH,soil moisture, nutrient availability in the soil as well as nutrient concentrations of plant tissues ofdifferent species). In addition, this large dataset will be used to select the 20 experimental sitesfor the second part of the project (2002-2003).
In the 2nd and 3rd year, causal relationships between biodiversity and ecosystem functionwill be determined at our app. 20 sites, focussing on resistance and resiliance of grasslandecosystems with respect to experimental disturbances. By manipulating the water regime and theabundance of insects and invertebrates, we will determine if well-established, managedgrasslands with low levels of biodiversity are less resistant and resiliant to disturbances thancomparable systems with higher diversity levels.
Since several decades, ecologists have tried to reveal the relationship of biodiversity andecosystem productivity in order to understand fundamental determinants of biodiversity(Mittelbach, et al., 2001). Species richness is often hypothesized to produce a hump-shaped oruni-modal relationship with productivity (Grime, 1973). Our results from the first part of thissub-project support this hypothesis for managed grasslands. Grasslands with low as well as veryhigh productivities showed low plant diversities, whereas grasslands with intermediateproductivity support the highest levels of plant diversity (Fig. 1).
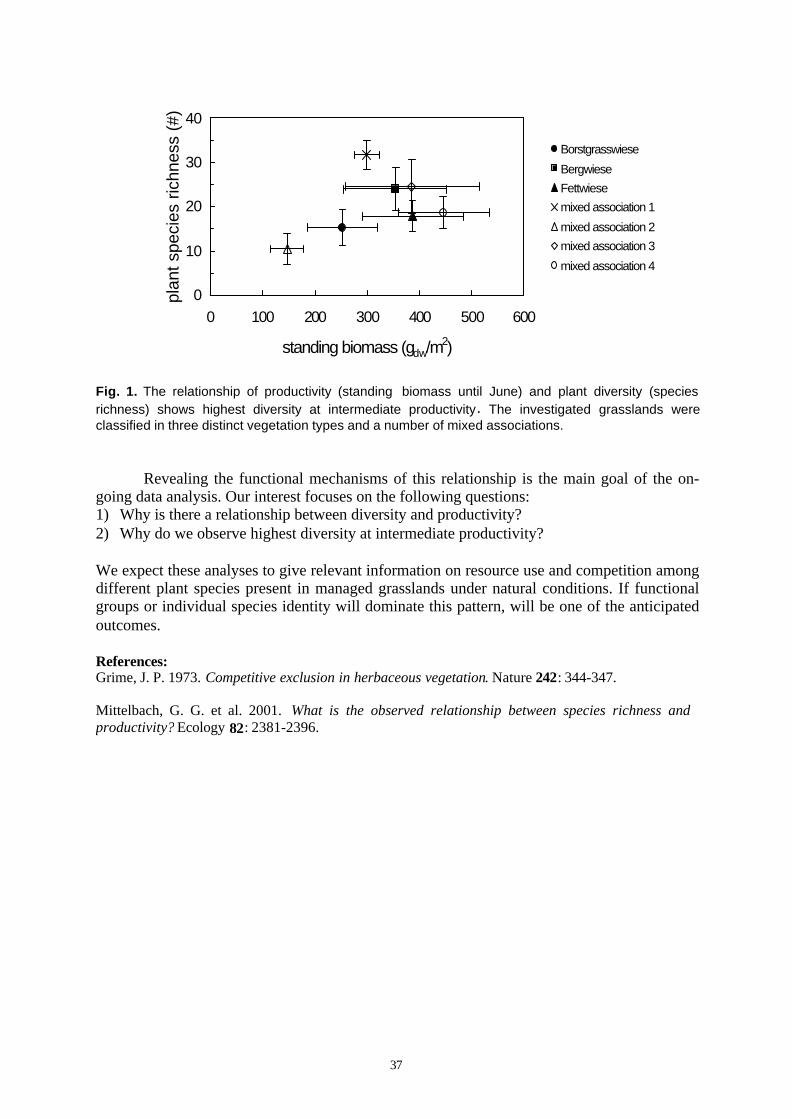
37
0
10
20
30
40
0 100 200 300 400 500 600
standing biomass (gdw/m2)
plan
t spe
cies
ric
hnes
s (#
)Borstgrasswiese
Bergwiese
Fettwiese
mixed association 1
mixed association 2
mixed association 3
mixed association 4
Fig. 1. The relationship of productivity (standing biomass until June) and plant diversity (speciesrichness) shows highest diversity at intermediate productivity. The investigated grasslands wereclassified in three distinct vegetation types and a number of mixed associations.
Revealing the functional mechanisms of this relationship is the main goal of the on-going data analysis. Our interest focuses on the following questions:1) Why is there a relationship between diversity and productivity?2) Why do we observe highest diversity at intermediate productivity?
We expect these analyses to give relevant information on resource use and competition amongdifferent plant species present in managed grasslands under natural conditions. If functionalgroups or individual species identity will dominate this pattern, will be one of the anticipatedoutcomes.
References:Grime, J. P. 1973. Competitive exclusion in herbaceous vegetation. Nature 242: 344-347.
Mittelbach, G. G. et al. 2001. What is the observed relationship between species richness andproductivity? Ecology 82: 2381-2396.

38
Project ID: 01LC0013 (Subproject RAUM)1.1.2000 – 31.12.2003
A SPATIAL MODEL FOR INDICATING ECOSYSTEM FUNCTIONS
Volker Audorff
Büro für ökologische Studien, Alexanderstr. 5, 95444 [email protected]; http://www.bfoes.de/zebene2/leistg/bsp/biodiv.htm
Key words: Biodiversity, grasslands, indicator system, modelling, geographic informationsystem (GIS)
Abstract:Based on measurements of the relationship between biotic and abiotic variables and
ecosystem function, the subproject RAUM aims at developing a predictive indicator systemduring 2001 and 2002. By using few and easily measurable variables, the indicator systemwill allow to predict certain ecosystem processes within grasslands with the help of aGeographical Information Systems (GIS). This model will be validated in the field incooperation with the other subprojects in 2003. In addition, the subproject RAUM willspatially visualisize the results obtained in the entire research network using (GIS) andanalyze vegetation data obtained in 2001. In the first year, 80 sites were studied in closecooperation with the subprojects STOFF, ENTO and GENMOD. First results show that theinvestigated grasslands can be differenciated into three distinct plant associations. In order toobtain additional information on managemetn history, a questionnaire for the landowners iscurrently developed.
Results:Subproject RAUM will utilize the dataset that was collected in first project phase in
2001 for the development of an indicator system (2002), in which taxonomic and structuralparameters of plant diversity are used to draw conclusions on ecosystem function in well-established managed grasslands in montane regions of central Germany. This indicator systemwill allow to predict certain ecosystem processes within these grasslands based on themeasurement of easily measurable variables. This model will be validated in the field with thehelp of GIS by testing its spatial prediction in the second phase of the project (2003). Afterfurther development, the model will be made available for applications in landscapemodelling and landscape planning. Thus, the subproject RAUM explicitly aims at aknowledge-transfer from science to the applied sector.
In cooperation with the other subprojects a broad dataset was collected in the year2001 as a basis for the described model. The data collected include taxonomic and structuralplant diversity such as the Ellenberg Indicator Values, life forms and plant functional groups,species cover, LAI, height and species composition as well as core ecosystem parameters suchas carbon and nitrogen stocks in the soil and vegetation, soil moisture, available nutrients (N,P, Ca, K, Mg, S) and soil acidity. Also, the plant associations of the investigated grasslandswere characterized by using species composition and abundance.First results of the first project phase show that three distinct grassland associations(Borstgraswiese, Bergwiese und Fettwiese) with different ecosystem properties such asstanding biomass and species diversity (see subproject STOFF) can be distinguished. UsingGIS, we spacially visualized these results in order to obtain an overview of the research area(Fig. 1).

39
An additional goal of the first project phase is to inquire detailed information on themanagement history of the 80 investigated sites. For this reason, we have developed an explicitequestionnaire for the responsible management agencies and farmers. From this questionnaire, wewill obtain information on harvest method and intensity, fertilizer applications, age of theecosystem and participation in special management and conservation programms. These data willnot only be essential for the indicator model, but are also of fundamental interest for thecooperating subprojects and the selection of the 20 core sites for the further experimentalmanipulations.
%[
%[
%[
%[
%[
%[
#S
#S
#S
#S
#S
#S
#S
#S
#S
#S#S
#S
#S
#S
#S
#S#S
#S
#S
#S #S
#S#S
#S
#S#S
#S#S
#S #S
#S
#S
#S
#S
#S
#S#S
#S
#S
#S
#S
#S
#S
#S#S
#S
#S
#S
#S#S
#S
#S
#S
#S
#S#S
#S
#S
#S
#S
#S#S
#S
#S
#S#S
#S
#S#S#S#S
#S
#S#S
#S
#S
#S
#S
Steinach
Sonneberg
Lobenstein
Leutenberg
Neuhaus am Rennweg
Ludwigsstadt
Itz
Roda
ch
Stei nach
Haß la
ch
K re m
nitz
Tett a
u
Loquit z
Sormitz
Lic h
te
Schwarza
Effel
der
Schlage
E inf luss der B io divers it ät aufÖkosystemprozesse in Grünlandbeständen
Artzahl
0 2 4 6 8 10
Kilometer
#S 8 - 1 1#S 12 - 15
#S 16 - 19
#S 20 - 23
#S 24 - 27
#S 28 - 31
#S 32 - 33
Fig. 1: Location and species richness in the grassland communities visited in 2001(2 x 2 m-Plots).

40
Project ID: 01LC0013 (Subproject ENTO)1.1.2000 – 31.12.2003
THE RELATIONSHIP BETWEEN INSECT HERBIVORIE, PLANT DIVERSITY,INSECT DIVERSITY AND ECOSYSTEM PROCESSES
Carla Wytrykush, Dr. Jörg Perner, Prof. Dr. Wolfgang W. Weisser
Institut für Ökologie, Friedrich-Schiller-Universität Jena, Dornburger Str. 159, 07743 [email protected]; http://www.uni-jena.de/biologie/ecology/
Key words: Herbivory, Insects, Ecosystem Function, Insecticide, Experiment, Grassland
Abstract:Only recently has the significance of insect herbivores for the structure and dynamics of plantcommunities been recognized. There is some evidence that manipulating plant communities hasan effect on insect herbivores, but little research has been performed to demonstrate the effects ofinsect herbivory on ecosystem processes. The purpose of the ENTO subproject is elucidate therole of above-ground insect herbivory for the structure and dynamics of the plant community.We will investigate the relationships between insect herbivory, plant diversity and ecosystemfunctioning and stability. Insect herbivore pressure will be removed using insecticides and theeffects on natural grassland ecosystems determined. The manipulation of insect herbivory willalso help us to determine the role of insects in ecosystem responses to stress (drought). Inaddition, the dependence of insect diversity on plant diversity will be determined for grasslandsof different diversities.
Results:It is generally accepted that large, plant-eating (herbivorous) vertebrates exert a great influenceon vegetation dynamics. The importance of herbivorous insects for the structure and dynamics ofnatural plant communities has, however, only recently been recognized and investigated.Manipulative studies where insects were removed using biocides have shown that herbivory maynot only change the diversity and composition of plant communities but also affect the directionof succession. In addition to trophic effects that arise directly or indirectly from removal ofbiomass by herbivores, herbivorous insects may also affect ecosystem processes through anumber of non-trophic effects. Surprisingly, there have been very few studies on the effects ofinsect herbivores on ecosystem processes. The most convincing evidence that herbivores affectecosystem processes comes from direct manipulations of herbivore populations. For terrestrialsystems, such experiments have provided some clear demonstrations of significant effects ofherbivores on ecosystem processes such as primary productivity and nutrient cycling, but so farthey have only been performed with a focus on large mammalian herbivores. In almost all of therecent biodiversity-ecosystem function field experiments, the emphasis was on the manipulationof plant diversity only. Because insect herbivory will depend on plant species diversity, it can behypothesized that insect herbivory will modify the relationship between plant diversity andecosystem processes.
While insects affect plant communities, the plant community itself exerts a strong influence onthe diversity of the insects associated with the plant community. Virtually all ecological modelspredict that an increase in resource diversity supports a more diverse array of consumers.Similarly, increased herbivore density is predicted to result in an increased diversity of parasitesand predators but there may also be direct effects of plant diversity on parasite and predatordiversity as well as changes in the interactions between herbivores and their natural enemies.Thus, it is likely that increasing plant diversity affects the diversity of the whole insect complex,but the relationship can be complicated.

41
The subproject ENTO has the following goals:
1. To manipulate the presence of herbivorous insects in grassland plots to study therelationships between plant diversity, insect herbivory and ecosystem processes. Themanipulation of insect herbivory will also help us to determine the role of insects inecosystem responses to stress (drought).
2. To assess insect diversity in the experimental plots to investigate the relationship betweenthe compositional structure of the insect complex, plant diversity and plant productivity.
In the first year (2001) of the study, ca. 80 different grasslands in Thuringia at a range of plantspecies diversity were visited in a collaborative effort of subprojects ENTO, GENMOD,STOFF and RAUM. The task of this subproject is to quantify insect diversity in these plots.Suction samplers in connection with 'biocenometers' were used to obtain standardized samplesof the insect fauna. The specimens in the samples will be separated into taxonomic groups.For certain taxa, all individuals will be determined to species level (Heteroptera,Auchenorrhyncha, certain groups of Diptera and Coleoptera etc.). The data will allow us todetermine the relationship between plant diversity, plant productivity and insect diversity inthese grasslands.
In the second and third year (2002 and 2003), results from the first year will be used to selectgrasslands at different levels of diversity in which experimental manipulations will be carriedout. In the herbivore removal plots (see abstract of main project), insecticides will be appliedrepeatedly. Biomass production and other ecosystem processes will be measured inmanipulated and control plots to determine the importance of insect herbivores for ecosystemfunction. The manipulation of insect herbivory in drought-stressed grasslands will also allowus to determine the role of insect herbivores in the stress response of grasslands. In addition,the insect diversity in the (unsprayed) experimental plots will be assessed to investigate theimportance of plant diversity, plant productivity and stress for the insect complex.

42
Project ID: 01LC0013 (Subproject GENMOD)1.1.2000 – 31.12.2003
GENETIC AND PHENOTYPIC DIVERSITY AND RESPONSE TO DISTURBANCESIN GRASSLAND SYSTEMS: MODELLING AND EMPIRICAL RESEARCH
Gottfried Jetschke, Frank Hellwig, Nidal Odat
Institut für Ökologie, Friedrich-Schiller-Universität, Dornburger Straße 159, 07743 JenaInstitut für Spezielle Botanik, Friedrich-Schiller-Universität, Philosophenweg 16, 07743 [email protected]; http://www.uni-jena.de/biologie/ecologyhellwigotto.biologie.uni-jena.de, [email protected]
Key words: genetic diversity, phenotypic diversity, grasslands, ecosystem functions, modelling,RAPD-PCR, reproductive success,
Abstract:The aim of the project is a two-fold one: the development of a mathematical model to cover therelationship between both phenotypic and genetic biodiversity and ecosystem functions and thequantification of genetic diversity of selected species in the field. The model will include multipletrophic levels and different functional types and shall give a conceptual answer how productivityand nutrient cycling are related to the diversity of species. Representative species of ourgrasslands will be selected to quantify genetic diversity within and between populations byRAPD-PCR.
The modelling part covers the development of an appropriate mathematical model which shallespecially include the genetic diversity of the community and the response of the ecosystem toexternal disturbances, like drought stress or removal of herbivores. The results of the modelinvestigations will give a more general answer how the response of the system depends on itsdiversity (with respect to functional types) and on the number of trophic levels. A specialemphasis is given to the question how the degree of genetic diversity will influence thefunctioning of the community under disturbances and environmental changes.
The empirical part of the project is focussed on the quantification of the genetic diversity in thefield. Typical species, like annual or perennial herbs and grasses of different growth form, will beselected to quantify the genetic diversity by means of a RAPD-PCR. These investigations will beperformed over two years to detect possible changes after an experimental manipulation.Moreover, the genetic diversity will also be measured by using quantitative traits of reproductivesuccess, as pollen and seed production and vitality and differences in germination success.
Results:A basic model structure has been developed to include several functional types of primaryproducers plus an additional trophic level of herbivores with different food preferences. Themodel is explicitly based on the uptake of a resource (e.g. nitrogen), different ways of utilisationby the plants and cycling through local and regional pools. It can be shown that the indirectinteractions through local resource use can support the coexistence of a certain number of speciesand, therefore, maintain diversity. Some other results obtained until now show that herbivoresmay essentially influence the performance of the community (net primary productivity) by theirfeeding behaviour and spatial mobility.

43
First attempts have been made to include the genetic structure and its change betweengenerations. Growth throughout the year is modelled in continuous time, but reproduction(and genetic mixing) is a discrete event based on random mating, seed production andcreation of vegetation gaps by mortality. Also a spatially explicit model structure has beentested to serve as a starting point for an individual-based modelling approach.About 14 species have been sampled from the potential sites to check which of them are mostsuitable to be used for the genetic investigations. The genetic analysis by PCR is just beingperformed and first results will be available by the end of the year. Based on these preliminaryscreening and on the species list of the sites selected for our experiments, about fivecharacteristic species will be selected for the detailed genetic analysis in the next two years.

44
Projekt ID: 01LC00141.4.2001 – 31.3.2004
BIODIVERSITY AND ECOSYSTEM FUNCTIONS IN GRASSLAND
G. Aas1, C. Beierkuhnlein2, G. Kossmann3, E. Matzner3, C. Neßhöver2, A. Reuter4 andC. Engels4
1 Ökologisch-Botanischer Garten, Universität Bayreuth, 95440 Bayreuth;2 Landschaftsökologie, Institut für Landschaftsplanung und Landschaftsökologie, UniversitätRostock, 18059 Rostock; 3 Bayreuther Institut für Terrestrische Ökosystemforschung,Lehrstuhl für Bodenökologie, Universität Bayreuth, 95440 Bayreuth; 4 AbteilungAgrarökologie, Universität Bayreuth, 95440 [email protected]; http://www.bitoek.uni-bayreuth.de/biolog
Grassland is an important type of agroecosystem covering about 60 million ha of land in middleEurope. The number of plant species in grassland communities may strongly vary depending onclimate and soil conditions. In agricultural grasslands, plant diversity is strongly affected byagricultural measures such as frequency and date of harvest, and fertilization. Recent studies haveshown that reduction of plant diversity may decrease the productivity of grassland (Hector et al.,1999). However, up to now there is strong debate on the general validity of diversity effects, andthe mechanisms involved in the positive diversity effects found in some experimental studies(Grime, 1997; Kaiser, 2000).It is the aim of this project to assess biodiversity effects on ecosystem functions in grassland.Biodiversity may be defined as taxonomical diversity, i.e. number of plant species, or functionaldiversity, i.e. number of functional types. Ecosystem functions include net primary production,accumulation of minerals and carbon in plants and soil, and filtering of water for ground waterformation. The importance of taxonomical and functional diversity on these ecosystem functionsis investigated in experimental grassland communities established in the field, in large lysimeters,or in small root boxes.In the field, biodiversity effects are measured in grassland communities that were established in1996 on 2 x 2 m plots („BIODEPTH“ project). On 64 Plots, communities with 1, 2, 4, 8 and 16species were established. Species were selected from 30 grassland species typical for the area. Theplots were weeded from 1996 to 1998 to sustain composition and species number. In 1999, theweeding was stopped to allow succession to occur within the plots. The plots were underharvested twice a year (June and September) and were not fertilized. Emphasis of theinvestigation within the BIOLOG-Bayreuth project is put on the plots with communities showingsimilar taxonomical and functional diversity as that in the lysimeters (see below).In the lysimeters with a surface area of 1.3 x 1.3 m and a depth of 1 m and in the root boxes (0.2m diameter, 1 m depth) biodiversity effects are measured in grassland communities that wereestablished in 2001. The plant communities are composed of either 2, 4, or 8 different plantspecies that may be attributed to 4 functional groups: grasses with low growth habitus, grasseswith high growth habitus, herbs with low growth habitus and rosettes, herbs with high growthhabitus. Legumes were not included. All communities are grown in 5 replicates. In the root boxes,in addition to the plant communities in the lysimeters, also monocultures of all species grown inthe lysimeters are cultured.The effects of biodiversity on ecosystem functions are studied in a research group including thedisciplines botany, ecology, and soil science. The project is divided into three subprojects thatinvestigate biodiversity effects on (1) above-ground and (2) below-ground plant characteristicsand (3) soil characteristics and water and nutrient fluxes.

45
References:1. Grime, J. P., Science, 1997, 277, 1260-1261.2. Hector, A. and 33 coauthors, Science, 1999, 286, 1123-1127.3. Kaiser, J., Science, 2000, 289, 1282-1283.

46
Projekt ID: 01LC0014 (Subproject Vegetation)1.4.2001 – 31.3.2004
TAXONOMIC, FUNCTIONAL AND STRUCTURAL CHARACTERISTICS OFABOVE-GROUND PLANT FEATURES AND THEIR INFLUENCE ONECOSYSTEM FUNCTIONS
C. Neßhöver1*, G. Aas2 and C. Beierkuhnlein1
1 Landscape Ecology, Institute for Landscape Planning and Landscape Ecology, University ofRostock, 18059 Rostock2 Ecological-Botanical Garden, University of Bayreuth, 95440 Bayreuth* current address: Bayreuth Institute on Ecosystem Research (BITÖK), University ofBayreuth, 95440 [email protected]
Key words : biodiversity, grassland, productivity, functional diversity, structural diversity,functional attributes
Abstract:Biodiversity can be examined from different viewpoints. Besides the taxonomical view, structuraland functional aspects of biodiversity get more in focus, especially when regarding therelationship between diversity and ecosystem functions.These different aspects of diversity are the main objective of the subproject. Earlier studies inexperimental grasslands have shown, that there is a relationship between the existence of specificfunctional groups of plants for example for productivity and nitrate leaching in grasslandcommunities. These functional relationships shall be further investigated by dividing plant speciesinto funtional groups and observing their performance within the communities.
Experimental questions and approaches:How are productivity and biodiversity in grasslands related to each other? Several studies haveshown a positive correlation (e.g. Hector et al. 1999). But the debate about this relationshipremains controversial (e.g. Huston et al. 2000). For example, there is evidence that the diversity offunctional types of plants is more important than taxonomic diversity (Spehn et al. 2001). Moststudies have focused on this functional viewpoint only marginally, for functional classificationsentail several problems (Beierkuhnlein & Schulte 2000). Within the vegetation-subproject ofBIOLOG-Bayreuth, the functional classification of species and individuals and their role shall beexamined in depth, using existing data on the species and biometric measurements of individualsin the experiments. The species composition of the lysimeter-experiment was designed accordingto this aim.Besides the taxonomic and functional aspect of diversity, also a structural aspect will beaddressed. Continuous LAI-measurements in different heights are carried out to look at thestructural development of different plant mixtures on the lysimeters and the experimental fieldplots. Relating these data to biomass production, the role of a higher light use efficiency due tomore diverse temporal and spatial development of above-ground plant-organs for productivitywill be examined.Adressing these different aspects of biodiversity it is aimed to evaluate the varying informationcontents implied with premise to a prediction on ecosystem functions. As perspective, the resultsof BIOLOG-Bayreuth shall be applied to the landscape scale by identifying the main traits ofgrassland diversity in central Europe.

47
References:1. Hector, A. and 33 coauthors, Science, 1999, 286, 1123-1127.2. Huston, M.A. et al., Science, 2000, 289, 1255a.3. Spehn, E.M., Scherer-Lorenzen, M., Schmid, B., Körner, Ch. and BIODEPTH-consortium,Verh. Ges. Ökol., 2001, 31, 178.4. Beierkuhnlein, C. and Schulte, A., pp. 45-64. In: Jax, K. (ed.): Theorie in der Ökologie:Funktionbegriff und Unsicherheit in der Ökologie, 2000. Peter Lang

48
Projekt ID: 01LC0014 (Subproject Roots)1.4.2001 – 31.3.2004
THE ROLE OF BIODIVERSITY FOR STORAGE OF CARBON AND MINERALS INROOTS, AND ACQUISITION OF SOIL RESSOURCES IN GRASSLAND
A. Reuter and C. Engels
Abteilung Agrarökologie, Universität Bayreuth, 95440 [email protected]
Key words : nutrient uptake ability, root diameter, rooting depth, root growth, rhizodeposition
Abstract:In this subproject the role of roots for biodiversity effects on productivity and cycling of carbonand mineral nutrients in grassland is investigated. The effects of plant diversity on rootcharacteristics that are related to (a) net primary production and mineral accumulation of roots,(b) acquisition of soil resources by living roots, and (c) release of organic carbon and nutrientsinto the soil by decaying roots are assessed in grassland communities that were experimentallyestablished in the field, or in lysimeters, or in root boxes. It is expected that the results willimprove our knowledge about the processes involved in diversity effects on ecosystem functions,and finally may help to develop recommendations of agricultural measures for sustainablegrassland management.
Experimental questions and methods:
Is below-ground productivity and mineral nutrient accumulation in roots affected by plantdiversity ?Below-ground productivity is assessed by measuring root growth in a well defined period of timeusing the ingrowth-core method (Oliveira et al., 2000). The soil cores are sampled twice per yearto a depth of 1 m for measurement of biomass and nutrient content of roots.Is acquisition of soil ressources affected by plant diversity ?To examine this question morphological root characteristics related to nutrient and wateracquisition such as root length and surface area are measured in the roots sampled from ingrowth-cores (see above). Physiological root characteristics related to nutrient and water acquisition aremeasured in these root samples using 15N (Buljovcic and Engels, 2001), or are indirectly assessedfrom accumulation of tracers in the shoot. For the assessment of root characteristics related tonutrient acquisition, special emphasis is put on „critical“ time periods where root growth andactivity are expected to be low (e.g. after cutting, in spring and autumn), and soil layers whereroot length density is expected to be suboptimal for complete capture of soil resources (i.e. belowthe main rooting zone). In the communities growing in root boxes, root growth along a transparentinterface with the soil can continuously be measured, allowing assessment of root growth with thehigh resolution in time that is necessary for the critical periods.Is carbon and nutrient release from roots to soil affected by plant diversity ?It is to be expected that in agriculturally used grassland roots are the most important source forcarbon and mineral input into the soil. The rate of rhizodeposition is dependent on root longevitythat is affected in a species-specific manner by shoot pruning. Root longevity and root turnoverare estimated by repeated observations of the same section of the root boxes allowing to estimatethe time interval between appearance and disappearance of specific roots from the interface (Smitet al., 2000).

49
References:1. Buljovcic, Z., and Engels, C., Plant Soil, 2001, 229, 125-135.2. Oliveira, M. R.G., van Noordwijk, M., Gaze, S. R., Brouwer, G., Bona, S., Mosca, G., andHairiah, K., pp. 175-210. In: A.L. Smit et al. (eds.) Root Research: a Handbook of Methods,2000. Springer-Verlag.3. Smit, A. L., George, E., and Groenwald, J, pp. 235-272. In: A.L. Smit et al. (eds.) RootResearch: a Handbook of Methods, 2000. Springer-Verlag.

50
Projekt ID: 01LC0014 (Subproject Soil)1.4.2001 – 31.3.2004
THE IMPACT OF PLANT DIVERSITY OF GRASSLANDS ON WATER- ANDNUTRIENT-FLUXES IN SOILS AND SOIL ORGANIC MATTER
G. Kossmann and E. Matzner
Bayreuther Institut für Terrestrische Ökosystemforschung (BITÖK), Lehrstuhl fürBodenökologie, 95440 [email protected]
Key words: soil water- and nutrient-fluxes, leaching of NO3 and DOM, characteristics ofphysical fractions of SOM
Abstract:Main aims of this subproject are to investigate changes in soil parameters and to measurewater- and nutrient-fluxes in BIODEPTH-plots and lysimeters. The soil inventories coverCEC, Nmin, PCAL and physical soil fractionation with subsequent analysis of the fractions. Soilsolution and leachates are collected to evaluate changes in the solution phase and tocalculate seepage fluxes of mineral elements and dissolved organic carbon.
Approach:Does plant diversity affect water- and nutrient-fluxes in soils and soil organic mattercharacteristics?It is postulated that increased diversity causes a higher use of soil-born resources which leadsto detectable lower concentrations of nutrients in soil solution. Higher use of nutrients resultsin a decrease of NO3-leaching to the groundwater (also HECTOR et al, 1999). Furthermore,the higher diversity is supposed to result in an increasing carbon-sequestration of the soil.Soil parameters such as CEC, Nmin, PCAL, etc. will be investigated in an initial (autumn 2001)and a final soil inventory in autumn 2003 on BIODEPTH-plots and lysimeters. Specialemphasis is given to physical fractionation of SOM (CHRISTENSEN, 2001) and subsequentanalysis of the fractions.The experimental grassland communities (BIODEPTH-plots) will be used to investigate theimpacts of plant diversity on soil moisture, potential leaching of NO3 and dissolved organicmatter (DOM). Soil matrix potentials are continuously measured by tensiometers (30 and 50cm soil depth) and soil solution is collected periodically by ceramic suction cups at 40 cm soildepth.The main aim of the lysimeter studies is to balance water- and nutrient-fluxes in response toplant diversity. This task is achieved by acquisition of soil solution from 15 cm and 30 cmdepth (using suction-cups) and leachates from 100 cm depth.Solutions are analysed on a monthly basis for mineral elements (K, Mg, Ca, Mn, Fe, Al, Na,NO3, NH4, Cl, SO4, HPO4, HCO3) and DOM. The study started in May 2001, the installationswere recently finished, thus, results are not available yet.
References:1. Christensen, B.T., 2001, European Journal of Soil Science, 52, 345-353.2. Hector, A. and 33 co-authors, Science, 1999, 286, 1123-1127.

51

52
Project ID: 01LC0015 (Subproject VEGETATION ECOLOGY)1.3.2001 – 29.02.2004
DEVELOPMENT OF BIOLOGICAL DIVERSITY IN SALT MEADOWS OF THE'VORPOMMERSCHE BODDENLANDSCHAFT' – SPATIO-TEMPORALDYNAMICS AND ECOSYSTEM FUNCTION OF BIODIVERSITY IN BRACKISHGRASSLANDS
Dipl. Geogr. E. Held, Dipl. Biol. S. Seiberling, Dipl. Biol. R. Zimmermann, and Prof. Dr.U. Schickhoff
Stiftungsprofessur Biodiversität, Department of Botany, University of Greifswald, Grimmer Str.88, 17489 [email protected]; http://www.uni-greifswald.de/~botanik/botinst.htm
Key words: salt meadows, biodiversity dynamics, ecosystem function, multi-temporal analysis,ecological modelling
Abstract:The project is part of the interdisciplinary research program “Development of biodiversity ofsalt meadows in the Vorpommersche Boddenlandschaft“, which is carried out by severalresearch groups of the University of Greifswald, the University of Bremen and the TechnicalUniversity of Berlin. Biodiversity of salt meadows on the German coast of the Baltic Sea isinvestigated with regard to temporal and spatial dynamics as well as ecosystem function inorder to explore quantitative and qualitative dimensions of biodiversity dynamics.By means of vegetation and phytomass analyses and soil samplings on different study sitesabiotic and biotic site factors and biodiversity will be correlated. Local and regionaldistribution patterns of phytodiversity are assessed and human impact is evaluated. Multi-temporal analyses are applied to detect previous biodiversity dynamics. GIS-integratedmodelling is developed to enable predictions of future changes of biodiversity in the saltmeadows. Studies of ecosystem function with regard to stability and productivity are based onexperiments imitating changes of environmental conditions. Experiments include simulationsof sea level change and nutrient decrease. 30 permanent plots comprising the whole variety ofsalt meadows were established to assess future biodiversity changes.
Results:
1. Spatial Aspects
In the beginning of this project vegetation ecologists and animal ecologists jointly selectednineteen study sites in order to analyse relationships between phytodiversity, zoodiversity andenvironmental conditions.In summer 2001, vegetation was analysed at each of the 360 pitfall traps, which were set by theanimal ecologists involved. Shannon-index and the Evenness-index were calculated for thevegetation data to characterize α-diversity. Concurrently with the vegetation records, phytomasswas harvested within an area of 25 x 25 cm nearby the traps. The dry weight of the phytomasswill be used as a measure of productivity of the different study sites. Furthermore, total C, N, P,and degree of salinity will be analysed in order to assess nutrient conditions.Soil samples were taken at the end of the vegetation period. Examined soil samples will includesoil moisture, mineral nitrogen, available phosphorus, C/N-ratio, organic matter, and soil-pH.According to first results there is a negative correlation between productivity and salinity. Furtherevaluations are carried out to find relations between site conditions and phytodiversity.

53
2. Temporal aspects
Temporal changes in species and ecosystem diversity were investigated with multi-temporalanalyses using a geographic information system. Research data were acquired from maps of the17th, 19th and 20th century. The evaluation of a floristic database, which is available at theDepartment of Botany, shows a distinct decline for the most of the typical salt meadow speciesafter the intensification of land use since 1964. Further data source materials (e.g. aerial andsatellite images) were collected for the ongoing analyses.The design of a model, which enables the development of different kinds of scenarios regardingfuture biodiversity dynamics (e.g. assuming a sea-level rise), is under development stage.To ensure and improve data exchange between the different sub-projects of the research program,a database was designed and implementation has started. Furthermore this database serves as thebasis for the projected database “Biodiversity Vorpommersche Boddenlandschaft”, which isgoing to put results of the research program at public’s disposal.To assess diversity changes over a longer period, 30 permanent plots were established. Study sitescomprise a salinity range from 2 0/00 to 10 0/00 and include natural types as well as embanked sites.Vegetation cover and above-ground phytomass were recorded and soil samples were taken toassess the abiotic / biotic site conditions as a reference for a regular monitoring program.
3. Functional Aspects
In 2001, two of three experiments on the ecosystem function of phytodiversity with regard tostability and productivity have been launched. Both of them focus on the functional role diversityunder changing environmental conditions (i.e. sea level change and nutrient decrease).For the experiments, three different types of species composition growing under comparable siteconditions were selected in a 1 ha field-exclosure of a salt meadow 10 km north of Greifswald.Type 1: poor (4 grasses and herbs), type 2: rich (7 grasses and herbs), and type 3: rich withlegumes (7 grasses, herbs, and legumes) serve for a comparison of different richness in species aswell as in plant functional types.To simulate rising sea level conditions 20 sods of each type were transplanted 15 cm beneathfrom their original level, 10 remained as a control and 10 were dug out and re-implanted to assessthe mechanical impact of transplantation. Previously, species cover and above-ground phytomasswere recorded for each sod.Moreover, phytomass was measured for 20 sods and 10 controls of each type to detect theinfluence of nutrient decrease due to reduced N and P import by inundation. A N/P/K-solutionwill be added in next year´s early summer as a period of three years is to short to achieve asufficient reduction in nitrogen as the limiting factor of salt meadows by mowing. Thus, theopposite trend will be simulated in order to draw respective conclusions for a nutrient reduction.By the end of the vegetation period in 2001 the following site parameters are to be recorded forboth experiments: soil moisture and bulk density, mineral nitrogen, available phosphorus, soil pH,cation exchange capacity (CEC), base saturation, C/N-ratio, and organic matter. Phytomass willbe analysed with regard to the content of water, total N, P, C, and Cl.The data base of 2001 will serve as a reference for the experimental effects to be recorded overthe following two years.

54
PROJECT ID: 01LC0015 (Subproject LANDSCAPE ECONOMY)1.3.2001 – 29.2.2004
DEVELOPMENT OF BIOLOGICAL DIVERSITY IN SALT MEADOWS OF THE'VORPOMMERSCHE BODDENLANDSCHAFT' – ECONOMICAL VALUATION OFBIODIVERSITY
Prof. Dr. U. Hampicke , Dr. R. Döring, A. Kowatsch
Fachgebiet Landschaftsökonomie, Botanisches Institut, Universität Greifswald, Grimmer Str. 88,17487 [email protected]; http://www.uni-greifswald.de
Key words: Contingent Valuation, Use of salt meadowland, Costs and Benefits of Dyke building
Abstract:The decline in biodiversity is also a great problem from an economic viewpoint. Thereforeeconomists search for possibilities to preserve biological diversity with economic arguments.Within in the project there will be a calculation of a cost-benefit-analysis for a dyke-removingproject at the German Baltic Sea coast. This includes a contingent valuation for the existencevalue of rare species and the costs of different use systems of salt meadowland were developed.
Results:The decline in biodiversity is also a great problem from an economic viewpoint. It is clear that inthe future we will depend even more on stable ecosystems with a high biodiversity if todaysprocesses of biodiversity loss continue. Therefore economists search for possibilities to preservebiological diversity also with economic arguments. Economic valuation of costs and benefits ofdyke building as an example must introduce not only benefits from use values of an ecosystemchanged by a different water regime, also existence values and option values are important. Inaddition the costs of dyke building and the repair work must be introduced. This is the reason whyfor most cost-benefit analysis only estimations of the total economic value are possible. Over thelast three decades economists developed a lot of different methods of economic valuation ofindividual preferences for rare species, ecosystems or historic landscapes (for an overview abouteconomic valuation Gronemann und Hampicke 1997).Within the project as part of the research on salt meadowland we calculate costs and benefits ofdyke-removing and combine the results in a cost-benefit-analysis. To compare different situationsthree study areas were chosen. The Sundische Wiese, as an area where the dyke-removing isplanned, the Karrendorfer Wiesen where the removing has been realized a few years ago and theKooser Wiesen as a typical salt meadowland. The main use system on the meadowland isextensive pasturing with cattle and we estimate that after a dyke-removing 50% of the SundischeWiese is not usable that way anymore. Also the provender quality will change after the ecosystemwill come under the water regime of the Baltic Sea again (Kowatsch et al. 1997). What this meansfor the user was part of the work in the first year. In addition an overview about the literature withrespect to the history of the use systems should give us informations about the possible usealternatives in the future and what might happen after the dyke-removing. All these parts wereintroduced in a scenario for the Sundische Wiese and will be part of the cost-benefit-analysis atthe end.During the next year we’ll prepare the questionnaires for two examinations: On one side we wantto know what people around the area think about the dyke-removing (acceptance of this) and onthe other side we want to ask tourists about their preferences for rare species after the naturalhabitats will be restored. Not many contingend valuations deal with existence values and we hope

55
that we can get some useful results here. Most of the species on natural salt meadowlands are notvery common and not well known by the people (like orchids). In addition costs of dyke-removing, dyke-building and repair will be analysed over the next two years. After the firstproject phase (bejond 2004) the economic part will be oriented close to the research program ofthe other working groups within the project.
ReferencesGronemann, S. und U. Hampicke (1997): Die Monetarisierung der Natur – Möglichkeiten, Grenzen
und Methoden in: Jahrbuch Ökonomie und Gesellschaft Nr. 14 – Nachhaltigkeit in derökonomischen Theorie, Frankfurt (Main) (Campus), S. 164 – 207
Kowatsch, A., T. Fock, M. Köhler & J. Walter (1997): Landnutzung, Naturschutz und Küstenschutzim Biosphärenreservat Südost-Rügen, Neubrandenburg (Schriftenreihe der FHNeubrandenburg Reihe A), 137 S.

56
Project ID: 01LC0015 (Subproject ANIMAL ECOLOGY)1.3.2001 – 29.2.2004
DEVELOPMENT OF BIOLOGICAL DIVERSITY IN SALT MEADOWS OF THE'VORPOMMERSCHE BODDENLANDSCHAFT' - DYNAMICS IN DIVERSITY OFANIMAL SPECIES AND COMMUNITIES IN A CHANGING ENVIRONMENTWITH SPECIAL REGARD TO DISPERSAL PROCESSES
Prof. Dr. D. Mossakowski, Prof. Dr. G. Müller-Motzfeld, A. Broll, W. Dormann, T.Görlich, A. Hochkirch, N. Kamer, G. Mathiak, R. Mißkampf, R. Vierk, A. Ziebarth,M. Zimmermann
Institut für Ökologie und Evolutionsbiologie, Abteilung Evolutionsbiologie, FB 2, UniversitätBremen, PF 33 04 40, 28334 [email protected]; [email protected]://www.biologie.uni-bremen.de/biologie/ifoe/evolutionLehrstuhl für Spezielle Zoologie, Ernst-Moritz-Arndt Universität, Bachstraße 11/12, [email protected]; [email protected]; http://uni-greifswald.de/~zoologie
Key words: salt meadows, salt marshes, Vorpommersche Boddenlandschaft, Baltic Sea, pitfalltrap, lifestock taking, Carabidae, Saldidae, distribution patterns, area value rating, dispersal,population genetics
Abstract:Ecological analysis is used to show the significance of the distinctiveness and dynamics ofenvironmental factors which essentially determinate the diversity of animal species, communitiesand landscape sectors. The focus will be on the impact of human activities, with special regard toclimatic changes and the dispersal abilities of animal species.In close co-operation with the other project partners, recent developments and their influencingfactors will be analyzed by molecular techniques, field ecological (and, if necessary, laboratory)methods, as well as a Geographical Information System (GIS).Based on these results, prognoses (e.g. scenarios) to be used for the recommendation of measuresfor the maintainance and development of biodiversity with respect to economical relations can bestatet. The following questions will be investigated:Ø Which regional dispersion patterns characterize the diversity of species, coenoses and
landscape sectors? Are there areas with a high concentration of species that show a highspatial significance?
Ø Are there small-scaled and regional differences in the genetical equipment of populationsthat might give a hint for the genetical pauperisation due to isolation, or that reflect therange and intensity of dispersal processes?
Ø Which are the abiotic/biotic factors to determine diversity (distinctiveness, dynamics)?How do extreme events, such as inundations due to heavy storms, influence diversity?How important are area size, age and isolation?
Ø Which are the historical and recent impacts of land-use type and intensity (especiallyagriculture, coastal protection and tourism)?
Ø How does the dispersal ability of animals influence the distinction of biodiversity? Do thehydrodynamics of the Baltic Sea cause a ‘hydrochoric network’ of populations orcoenoses? How far do human activities influence such a network (increasing isolation)?
Ø Which impacts can be expected of the predicted climatic changes? (direct: temperature,precipitation, wind; indirect impacts: sea level rise, rising frequency of inundations dueto heavy storms, change of landuse type and intensity).

57
Ø Are there areas for the emigration of arthropods, which live exclusively in salt- andcoastal habitats? Can biodiversity be regenerated/re-established by a retraction of thedike position towards inland (expansion of foreshore areas)?
Ø How can the results be used effectively within an Integrated Coastal Zone Management(databases, GIS, expert or indicator systems, monitoring concepts)?
In cooperation with the project partners, along the “Vorpommersche Boddenküste” (investigationregion) 20 investigation areas were selected with special regard to geographical location,geological development, hemerobiotic degree, soil type and recent agricultural use.In the first year of study (2001), epigeic arthropods (especially Carabidae and Saldidae) werecaught at 65 investigation sites (1 to 9 investigation sites per area) with pitfall traps from the endof March until the beginning of November. The pitfall traps were changed 16 times in 14-days-intervalls and remained outside for at most 224 days. The time consuming sorting of the animalsis still in progress. Additionally in 2001, vegetation, biomass, inundation frequency (based on anivellement; measuring in 14-days-intervalls) as well as several soil parameters are investigatedin cooperation with the working group of Prof. Schickhoff.Parallel, lifestock taking of Carabids in the entire investigation region was started: Nearly 25%of the 800 km coastline have been under survey so far. Much effort had to be done to get freshmaterial for DNA extraction because of the complex spatial situation. In order to make use ofmaterial already fixed, we tested specimens collected in ethylene glycol, the medium used inpitfall traps in order to get species which are rare or not easily to collect.In Carabid beetles living north of the Alps, species are known without variability in theirdistribution area, others display variability from population to population. Therefore, wescreened core and mitochondrial genes of those species which were collected first from mostsites: Carabus clatratus, a species flightless and living on peat ground like coastal salt marshesand meadows with a high content of organic matter.
Results:First results show partly considerable quantitative and qualitative differences in the Carabidcommunities between the investigation sites of the saltmarshes. Intensity and frequency ofinundation (often regulated by coastal defense management), water salinity and landuse intensity(e.g. vegetation structure and soil compaction) seem to be the key factors for the development ofdiversity in Carabid coenoses.In general, isolated populations of Carabus clatratus show sequence differences; the highest rateof substitutions was found in ND1 and ND5 (0.2 – 0.5 %), the lowest in the control region, ITS1and ITS2 (about 0.1 %). Additional data exist for 12S rRNA, CO1, CO2, CO3. The mostinteresting result is a relatively high genetical difference in two populations (islands GrosserWerder and Hiddensee (Gellen)) that are about 7 km apart.Although at this stage we are unable to interpret the pattern of variability, this flightless speciesdisplay a high degree of within species diversity.
References:VAGTS, I., CORDES, H., WEIDEMANN, G., & MOSSAKOWSKI, D. (2000): Auswirkungen von
Klimaänderungen auf die biologischen Systeme der Küsten (Salzwiesen & Dünen), Abschlußberichtdes BMBF-Verbundvorhabens (Teil A u. B )
MÜLLER-MOTZFELD, G.; SCHMIDT, J, & BERG, C. (1997): Zur Raumbedeutsamkeit der Vorkommengefährdeter Tier- und Pflanzenarten in Mecklenburg-Vorpommern. – Natur und Naturschutz inMecklenburg-Vorpommern 33, 42-70.

58
Project ID: 01LC0015 (Subproject SOIL SCIENCE)1.3.2001-29.2.2004
DEVELOPMENT OF BIOLOGICAL DIVERSITY IN SALT MEADOWS OF THE'VORPOMMERSCHE BODDENLANDSCHAFT' - WATER AND SALT DYNAMICSOF SALTY MEADOW AND MARSH SOILS
L. Ikels, K. Schwärzel and G. WessolekWessolek
Ecology Institute, Soil Science Dept., TU Berlin, Salzufer 11-12, D-10587 [email protected]; [email protected];http://www.tu-berlin.de/fb7/ioeb/bodenkunde
Key words: biodiversity, global climate change, salt marshes, salt meadows, physical andchemical soil parameters, modelling
Abstract:Climate scenarios predict a global temperature increase within the next decades. Aim of theproject is to gain insight in the future development of the salt meadows of the VorpommerscheBoddenlandschaft, a region of outstanding ecological relevance. Analyses of new collecteddata on chemical, physical and hydrological soil parameters in this region will supply thebasis for a prediction of the soil lifetime dynamics associated with climate, groundwater andland use changes in this region.The specific study plan of the project includes:
- Exemplary soil ecological mapping of typical salt meadow areas.- Determination of relevant soil parameters for typical soil profiles.- Characterisation of the water and salt dynamics for two selected sites by means of soil
hydrological measurements and numerical model calculations.- Derivation of parameters concerning to soil water and soil salt balance as determined
by temperature, humidity, rainfall patterns, sea level height and land use due topredicted climatic shifts.
Project description and first results:Targets of the investigations in the first project phase have been the selection of theappropriate study sites and of the suitable measuring probes.For the purpose of a prolonged surveillance during the project, a transect with 10experimental plots was set up at the “Kooser Wiesen”. Measurements of soil water contentsand soil water quality are performed every 14 days in two soil depths. The soil pore water issubsequently analysed for pH-values, electric conductivity, concentrations of anionic andcationic salts and nutrients. The set-up of an intensive measurement field on the “KooserWiesen” is planned for next year. It is designed for measurements with higher analysisresolution in time and space (automatic probes measurements over soil profiles).The selection of the appropriate water content measuring devices is up to now challenging,since most of the commercially available TDR-probes (time domain reflectometry) do notdeliver reliable values at pore water salt contents above 20 mS/cm.A first attempt to gauge probes coated with epoxy resin seemed promising up to watercontents around 40% v/v, but since the peaty soils typical for the investigational sites mayreach values of 70% v/v, probably the use of the coated probes will not always be possible.
Next to the set-up of the measuring plots for the continuous data collection, soils of twotransects on the “Kooser” and “Karrendorfer” salt meadows have been investigated for

59
important chemical and physical soil parameters. The results of the analyses will supply,along with water tension curves, the bulk densities, water contents, C/N ratio, organic mattercontents, salt concentrations, and pH-values of the investigated soils.Future investigations will focus additionally on soil mapping of typical vegetation units of thesalt meadows and a subsequently soil ecological state assessment.Modelling of the dynamic development of the soil water parameters under changing siteconditions (salt contents, climate constraints, groundwater supply and land use pressure) shallhelp to answer the following questions:
1) Which consequences will arise from the current climate change scenarios in respect tothe site parameters of the ecologically relevant salt meadows?
2) How will the so-called ecological soil moisture parameter develop and how will thelength of the current dry and moist periods change in future, taking the different sitecharacteristics into account?
3) How will the salt meadows develop, considering particularly the scenarios predictingrecurrent situations of climate extremes (e.g., frequencies of flooding, dry periods)?

60
Project ID: 01LC0018 (SUBICON Subproject 1)1.3.2001 – 29.02.2004
SUBICON - SUCCESSIONAL CHANGE AND BIODIVERSITY CONSERVATION INA RED OAK CHRONOSEQUENCE – SUCCESSION OF DIVERSITY AND FUNCTION OF ECTOMYCORRHIZALTYPES IN RED OAK FOREST ECOSYSTEMS
J. Wöllecke, S. Gebhardt, B. Münzenberger and R.F. Hüttl
Chair of Soil Protection and Recultivation, Brandenburg University of Technology Cottbus,P.O. Box 101344, D-03013 [email protected]; http://www.tu-cottbus.de/roteichen/teilprojekte/index.htm
Key words: Dispersion of ectomycorrhizal types, Primary succession, Competition, Neophytes,Quercus rubra, Mining, Reclamation
Abstract:The region of Lower Lusatia (Eastern Germany) is characterized by many active and reclaimedlignite mining sites. Of these, 15 % have been reafforested with red oak (Quercus rubra L.). Thus,this introduced tree species is nowadays widespread on the former mining sites. As a pioneerspecies, red oak is well adapted to extrem soil conditions. Like other oak species, red oak formsectomycorrhizae with higher fungi (Ascomycetes and Basidiomycetes). In forest stands thesemycorrhizae hold a key position for the water and nutrient cycle. The mycobionts are able tomobilize, accumulate and transport water as well as nutrients from different soil substrates andtranslocate them to the phytobiont.The mycorrhizae and the extramatrical mycelium are part of the detritus food chain. The litter isintensively colonised by saprophytic and mycorrhizal fungi, bacteria and soil animals likeearthworms, enchytraeid worms, nematodes, collemboles, mites and others which deminute anddecompose organic components of the soil. Thus, the nutrients are being fixed in organiccomponents which can be taken up very fast by the growing mycelium. The huge diversity offungal species is based on the fungi`s different functions in mycorrhizal symbiosis. In consequenceof the high variety of functional groups the determination of fungal species will be the first part ofthe investigation. After characterization of the autecological importance of single species, theirinteractions with other heterotrophic organisms will be estimated.
Questions:The project will highlight some topics related to function, temporal and spatial dispersion ofectomycorrhizal types. The investigations will answer aspects of the following questions:- How does primary succession of ectomycorrhizal types work?- Which changes in successional stages can be observed along the chronosequence?- Are there differences in the ectomycorrhizal species composition associated to native and
neophytic oak species?- Are there differences in the ectomycorrhizal species composition of mining substrate and
undisturbed soils?- Which ectomycorrhizal fungi are key species for the development of forest ecosystems on
mining substrate, and how do these fungi disperse on different scales?- How important is the role of ectomycorrhizal fungi within the detritus food chain?
Methods:Our plots are located in former lignite mining sites reclaimed for forestry and planted with red

61
oak. The chosen Quercus rubra stands are forming a chronosequence. To show the differences inmycorrhization between the meliorized mining substrate and naturally grown soils we preparedalso a control plot in the neighbourhood of the mining area. To interprete the role of the treespecies in community stucture of mycorrhizae, we compare our results from the Quercus rubrastands with stands of native Quercus and Pinus species formerly investigated in the same region.For an analysis of the ectomycorrhizal community we identify the mycorrhizal fungi on threeways. First, we conduct a morphological and anatomical characterization of the mycorrhizae. Thismakes possible a quantitative analysis of the community composition. In a second step, we usemolecular genetic techniques like PCR, RFLP and sequencing of ITS regions.In using a capillar electrophoresis we try to analyse the content of different nutrient elements inthe mycorrhizae. These results, together with the morphological and anatomical features and theautecological data, will give us hints to the function of a specific fungal species in this ecosystem.
First results:After a preliminary sampling of our investigation sites we could differentiate 14 ectomycorrhizaltypes by means of anatomical and morphological characterization. Till now, we were able todetermine four of these types to species level. Cenococcum geophilum is a well known fungusestablishing mycorrhizae with a very broad range of phytobiots. In comparison to othermycorrhizal fungi, this species shows a wider ecological valency. Piloderma croceum andScleroderma citrinum are also known as euryoecious species. Former investigations in reclaimedmining sites in the same region detected mycorrhizae of these three species as dominant in Scotspine stands. A fourth species was determined as Paxillus involutus which is the dominant speciesin our samples. All of these four fungi are typical early stage fungi. Thus, they were expected tooccur in this sites of primary succession.Other mycorrhizal fungi which we identified by their fruitbodies were Amanita muscaria, A.citrina, Boletus edulis, B. luridus and several species out of the Russulales.In comparing natural forst stands with reclaimed mining sites our first samples indicate that rootmorphology can differ between these different habitats. Nevertheless, these findings have to beconfirmed by further samples.
The first place we presented our project was the congress of the German Society of Mycology andLichenology (GML 20. – 23. 09.01) in Halle, Germany.
Outlook:First we want to create a sampling design which allows an up/down scaling of the dispersion ofectomycorrhizal types to different scales.In experiments we want to investigate the influence of mycophagous insects on the establishmentof mycorrhizas and the development of mycorrhizal communities under grazing pressure.
Information on ongoing studies is available at www.tu-cottbus.de/roteichen/teilprojekte/tp1/index.htm.

62
Project ID: 01LC0018 (SUBICON Subproject 2)1.3.2001 – 29.02.2004
BIODIVERSITY OF PLANT COMMUNITIES AND SELECTED PHYTOPHAGES INRED OAK DOMINATED ECOSYSTEMS
P. Denkinger and U. Bröring
Dr. Udo Bröring, Lehrstuhl Allgemeine Ökologie, BTU Cottbus, Postfach 101344, [email protected], www.subicon.de
Key words: Biodiversity, Brown coal mining areas, Disturbance, Community Function, Heteroptera,Succession, Quercus rubra
Abstract:The temporal and spatial variation of biodiversity of plant communities and selected phytophagous arthropodcommunities in red oak dominated ecosystems is described. Species diversity of producers (higher plants andmosses) and first order consumers (phytophagous heteropteran species) are analysed in relation to influences ofhabitat diversity, disturbance regime, biotic interactions, and genetic diversity. Aim of the project is to giveexplanations for the generation, establishment, maintenance, and development of biodiversity, which should beconfirmed by experimental analyses.
Questions:In the restoration management of post mining landscapes widespread afforestation with non native red oak(Quercus rubra) took place. The main questions to be answered in the framework of the SUBICON researchproject are:What factors determinate the species diversity of red oak dominated ecosystems and adjacent open landscapes?Which of the various explications of biodiversity (e.g. number of species, Shannon-Wiener-Index, Preston´sgamma in species-abundance relations) are useful to describe biodiversity adequately in order to generate modelsto explain the maintenance and development of biodiversity?What about the relations among the biodiversity of different compartments of the communities (producers, firstand second order consumers, different sets of species with special ecological functions)?What about the temporal variation of biodiversity at different spatial scales and which of the general theories onthe development of biodiversity in ecosystems can be confirmed as to explanatory and predictive power?
Methods:The investigations are carried out in four former brown coal mining areas of Lower Lusatia (Eastern Germany)as well as in an undisturbed reference.The following investigations have already been realized:- Analyses of vegetation at 112 SUBICON plots in red oak forests as well as in the adjacent open landscape,- Starting of the experimental approach to recolonization after disturbance.- Quantitative sweepnet sampling of Heteroptera (30 sites in total, 50 sweeps per 100 qm on each of the
sampling sites).- Heteropteran species sampled by pitfall trapping will be included to analyse biodiversity of different vertical
strata.
The influence of soil disturbance on the diversity development of the vegetation in Lusatian post mining areas isanalysed by an experimental approach (see figure 1). It is widely accepted that disturbance has a strong influenceon biotic communities and their functioning on various spatial scales. In the post-mining landscape of Lusatia anobvious disturbance factor is the wild boar (Sus scrofa) that causes soil disturbances in all vegetation structuresby churning up the soil. This kind of disturbance is imitated. The experimental approach is to analyse themechanisms of colonization of plants and the development of diverse communities.Three open-cast mines of different ages and three vegetation types (red oak forests, Calamagrostis epigejosstands, psammophytic grassland) are included into the studies, six disturbance and six reference plots per standare analysed (sum of study plots: N = 108). The following procedure is applied:

63
- Soil disturbance: Removal of the above-ground vegetation. Digging up of the soil up to a depth of about12 centimetres;
- Recording of the colonizing individuals;- Soil seed bank analysis;- Exposition of seed traps.
(S = disturbance plot, 4 m², R = reference plot, 4 m², Distance between neighboured plots: 2 m)
Fig. 1: Experimental design
The results of the vegetation surveys and the second order consumers studies (TP 4) will be linked in order togenerate a general model comprising different functional groups of species.
First Results:Previous research on open landscape vegetation show that the change of vegetation type is slow. Directordination revealed that environmental parameters such as pH, the level of available phosphate and organiccarbon as well as the age of the study sites do not significantly account for the variance (Wiegleb & Felinks2001). For succession in red oak forests no likewise studies are known.The investigations started in summer 2001. Preliminary results can be presented at this state.The mean number of plant species in red oak forests of the former coal mining area is 13 (4 - 38 species), inundisturbed red oak forests 17 species (14 – 19 species), respectively, while in Calamagrostis stands 11 species(4 – 17) have been found. In psammophytic grasslands a mean of 9 species (2 – 17) occured.The number of heteropteran species found on oak trees is low, a very high amount of common species regularlyoccurring on native oak trees in Brandenburg are missing. This is in contradiction to the sampling results of openlandscapes, where species number is high: including results of former studies roughly calculated 30 % of allopenland species occuring in Brandenburg established populations at the post mining landscape.
Outlook:The next steps of the investigations will be:- Continuing with the data collection,- Determination of Heteroptera including material collected by pitfall trapping (Tp 4),- Statistical analyses, modelling,- Application of the experimental approach,- Data link between sampling results for different groups of species.
References:1. Wiegleb, G. and Felinks, B., Applied Vegetation Science, 2001, 4, 5-18.
RSRS
SRSR
RSRS

64
Project ID: 01LC0018 (SUBICON Subproject 3)1.3.2001 – 29.02.2004
GENETIC DIVERSITY OF SELECTED PLANTS, PHYTOPHAGES ANDZOOPHAGES IN SUCCESSIONAL RED OAK ECOSYSTEMS
W. Durka, K. Heyde and M. Güth
Dr. Walter Durka, Sektion Biozönoseforschung, Theodor-Lieser-Str. 15, D-06110 Halle,[email protected], www.subicon.de
Key words: Colonization, Genetic diversity, Molecular markers, Population genetics,Quantitative genetics, Succession
Abstract:The project will analyse genetic diversity of different functional groups (plants, phytophagousbugs, spiders and zoophagous beetles) within a Red Oak chronosequence. Molecular markers willbe used to elucidate genetic diversity and population structure as a basis for the understanding ofcolonization and microevolutionary processes. Genetic diversity within a population and betweenvarious populations as well as between different species are analysed in detail and compared toeach other.In plants quantitative genetics of morphological traits will be analysed in order to understandadaptive evolutionary changes due to varying selection regimes (light, competition, nutrients)during the chronosequence.The results will be used together with those of other subprojects in a comparative analysis of thedevelopment of species diversity and genetic diversity through time in a highly disturbedecosystem. The current state of the project including species selection, site selection, samplingand laboratory progress is reported.
Questions:The project addresses the following questions:- How do different functional groups of plants and animals differ with respect to spatial and
temporal population structure?- Does colonization take place through continuous immigration or through a single founder
population?- Does genetic diversity change through the course of succession?- Do molecular markers and quantitative traits differ in their spatial pattern and temporal
development?- Is there microevolutionary change during succession in adaptive traits connected with
competition or light regime?
Methods:The main method of analysis is the Amplified Fragment Length Polymorphisms (AFLP). TheAFLP technique is based on the selective PCR amplification of resriction fragments from atotal digest of genomic DNA (Vos et al. 1995).

65
Fig. 1: AFLP (Amplified Fragment Length Polymorphisms ) gel image of Rumex acetosellasampled in the Schlabendorf-Süd area.
First results:Species selection - plantsFrom a number of potential plant species two species with contrasting ecology and habitatwhere selected. Whereas a number of species were present either at the beginning or at theend of the chronosequence, only these two were present both early and late in thechronosequence.(1) Rumex acetosella (Sheep´s Sorrel) a herbaceous pioneer species with low competitive
ability, present in 13 sites.(2) Calamagrostis epigejos (Wood Small-reed) a generalistic stress-tolerant grass, present in
18 sites.
Species selection – animalsArthropods of different functional groups were selected, namely the phytophagous true bugStenodema laevigata, the spider Alopecosa fabrilis and the zoophagous ground beetleCalathus ambiguus.
The sampling both for the molecular and the quantitative genetics analyses was undertakenfrom July to September 2001. For molecular analyses of plants a total of 444 samples from 16(R. acetosella) and 21 (C. epigejos) sites has been taken. For quantitative genetics 1398 plantsfrom 16 (R. acetosella) and 21 (C. epigejos) sites are being cultivated and cloned vegetativelyfor experiments to be performed in 2002 and 2003.Of the target animal species 97 spiders, 173 true bugs and 366 ground beetles in 5 sites werecollected for molecular analyses. Next captures will be carried out in spring 2002.
For molecular analyses, AFLP (Amplified Fragment Length Polymorphisms) are currentlybeing developed. Preliminary results are shown in figure 1.
Outlook:The molecular analyses and laboratory work will be going on in 2002. Focuses on data analysisand modelling of the data will start in autumn 2002.
References:1. Vos, P., Hogers, R., Bleeker, M., Reijans, M., Vandelee, T., Hornes, M., Frijters, A., Pot, J.,
Peleman, J., Kuiper, M. and Zabeau, M., Nucleic Acids Research, 1995, 23 (21), 4407-4414.

66
Project ID: 01LC0018 (SUBICON Subproject 4)1.3.2001 – 29.02.2004
STRUCTURE OF THE PREDATOR COMMUNITY IN A RED OAKCHRONOSEQUENCE
A. Balkenhol, K. Voigtländer and W. Xylander
Staatliches Museum für Naturkunde Görlitz, Postfach 300154, 02806 Gö[email protected], http://www.subicon.de
Key words: Araneae, Carabidae, Chilopoda, Coal mining areas, Habitat structure, Red Oakchronosequence, Staphilinidae, Succession
Abstract:Araneae, Carabidae, Staphilinidae and Chilopoda are studied based on a comparative analysis ofdifferent aged red oak stands and grassland of post mining areas. The hypothesis of habitatstructure being the most determinating factor of biodiversity of a predator community is tested.
Questions:In the end of the 19th century, the red oak (Quercus rubra) was imported from North America toEurope as a fast growing tree being able to achieve a high productivity on poor stands. Until nowthere is only little knowledge about the importance of these forests for indigenous arthropodpredators. Several investigations have shown the importance of habitat characteristics such ashabitat structure for arthropod colonisation.In this context, we investigate both the development of red oak forest habitat diversity and thedevelopment of predator diversity in a chronosequence of former coal mining areas. Recultivatedareas are most suitable for investigations of biodiversity development because any establishedarthropod populations affecting new colonisators are absent.- What is the influence of tree species on the predator community?- What are the differences between forests with indigenous species (Quercus robur, Quercus
petraea) and unforested areas on reclaimed soil?- How much time does it take for establishment of biodiversity in disturbed landscapes?
Methods:The colonisation of the study areas by predator groups of different mobility and dispersal strategy(Araneae, Staphylinidae, Carabidae, and Chilopoda) is studied. The epigaeic activity of thesegroups is investigated by pitfall traps and tree trunk eclectors throughout the vegetation period.Additionally, soil cores and sweep-net catches are analysed for arthropod abundance in spring andautumn. Expected results refer to species richness, dominance structure, habitat preference ofspecies, life form diversity, age structure of populations, and also species and individual mobility.To investigate the influence of habitat structure and vegetation architecture of the forest stands onpredators, a method according to Curtis Bignal and Meißner, which analyses habitat structure anddiversity of different vertical quadrats, is applied.The investigations are performed in cooperation with the working groups of the BrandenburgischeTechnische Universität Cottbus and the Umweltforschungszentrum Leipzig-Halle. The synopticconsideration of the diversity of mycorrhiza (in both their symbiotic and degrading aspect), ofprimery producers, of selected phytophages, and of zoophages enables a comprehensive picture ofthe functional relations between spatial structures and biocenotic diversity.

67
First Results:In previous studies (Mrzljak & Wiegleb 2000) of open country vegetation of post mining areas itwas shown, that all epigaeic arthropod groups being present in the undisturbed surroundings canbe found after 20 years in considerable number and species rich community. No specific trend inspecies number development was detected. Species number of a site is obviously limited bystratification and height of the colonized vegetation type. Differentation among e.g. Calamagrostisand other tall grass stand is not strongly developed. In Europe no comparable data for Red Oakforests exists.
Overview:
Fig. 1: Investigated arthropod groups and environmental parameters
Outlook:Outdoor sampling and determination of material was started in July this year and will becontinued until end of 2003. First statistical analysis were carried out. Modelling of the data willstart after the end of collection.
References:1. Mrzljak, J. and Wiegleb, G., Landscape and Urban Planning, 2000, 51, 131-146.
Quercus rubra forestS U CC E S S I ON Indigenous
Quercus speciesGrassland
Small scalehabitat structure
Large scale habitat structure
Araneae
Staphylinidae
Carabidae
Chilopoda
• Ecological diversity
• Habitat preferences
• Form of life
• Age structure
• Mobility of species and individuals
• Influence of vegetation type
Quercus rubra forestS U CC E S S I ON Indigenous
Quercus speciesGrassland
Small scalehabitat structure
Large scale habitat structure
Araneae
Staphylinidae
Carabidae
Chilopoda
• Ecological diversity
• Habitat preferences
• Form of life
• Age structure
• Mobility of species and individuals
• Influence of vegetation type

68
Project ID: 01LC0020 (BIOPLEX Subproject 1)1.10.2000 – 30.9.2003
BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURALLANDSCAPES UNDER GLOBAL CHANGE (BIOPLEX) - MODELLING ANDGEOSTATISTICS
Kerstin Wiegand, Thorsten Behrens , Ute Visser, and Wolfgang Köhler
Biometry and Population Genetics, IFZ, U of Giessen, Heinrich-Buff-Ring 26, 35392 [email protected]; http://www.uni-giessen.de/biometrie/
Key words: landscape ecology, Geographical Information System (GIS), study site selection,spatially-explicit simulation models, ecological profiles
Abstract:Among several fields of activity we highlight two examples. First, we introduce PaDS, a tool forobjective study site selection for landscape ecology. PaDS showed that the landscape investigatedin BIOPLEX can be characterized by mean slope and mean perimeter-to-area ratio of the landparcels. BIOPLEX study sites were selected spanning the entire dimension of landscapecomplexity. Second, a spatially explicit, multi-species metapopulation model has been developedwhich will be used to investigate the functional influence of landscape structure on biodiversity.
Results:BIOPLEX investigates the connections among biodiversity and spatial complexity in agriculturallandscapes. This subproject combines the theoretical aspects of BIOPLEX. The fields of activityinclude (1) the development and application of a standardized procedure for study site selection,(2) the identification of meaningful landscape indices describing spatial landscape complexity asit influences biodiversity, (3) linking geostatistical information of the other subprojects in a GIS,(4) the development of spatially explicit simulation models facilitating a scaling up of findingsobtained at small spatial scales to the landscape scale, and (5) the development of hypotheses onthe functional influence of landscape complexity on biodiversity using spatially explicitsimulation models. Here, we present (1) and (5) in more detail.
Study site selectionIn large complex landscapes the spatial representativity of sample sites becomes important asfinancial and human resources of research projects limit the practible sample size. Therefore,we developed PaDS as a tool for standardised landscape analyses to guarantee reproducibleand representative study site selection. The main objective of PaDS is to detect patches that fituser specified criteria based on patch metrics. We explain PaDS using the selection of studysites situated in wheat fields for BIOPLEX subprojects 2 – 4 as an example.
First, PaDS was used to examine 714 Patches of 7 different sizes ranging from (250m)2 upto (3000m)2. For each of these patches a number of landscape metrics and terrain attributeswere derived from the DEM (Digital Elevation Model) and the official German land usedatabase called ATKIS; both converted to grids with 20m mesh size. The parameters used inthis project were: proportion of different types of land use, acreage/pasture ratio, (total)number of parcels, perimeter-to-area ratio (PAR), Shannon Index, (derived from ATKIS), aswell as mean slope, mean global radiation, mean topographical roughness, mean altitude,mean height above drainage channel, proportion of stretched hill slopes (derived from theDEM) and mean precipitation. Using principal component analysis (PCA) this plethora of
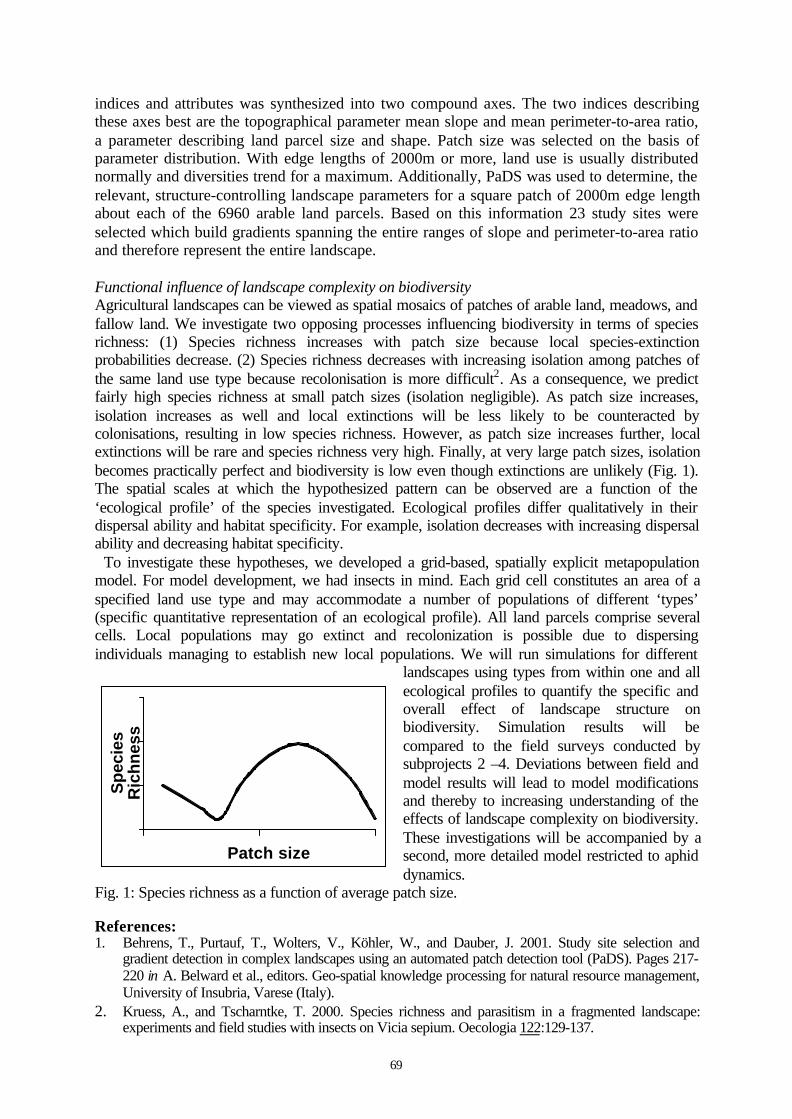
69
indices and attributes was synthesized into two compound axes. The two indices describingthese axes best are the topographical parameter mean slope and mean perimeter-to-area ratio,a parameter describing land parcel size and shape. Patch size was selected on the basis ofparameter distribution. With edge lengths of 2000m or more, land use is usually distributednormally and diversities trend for a maximum. Additionally, PaDS was used to determine, therelevant, structure-controlling landscape parameters for a square patch of 2000m edge lengthabout each of the 6960 arable land parcels. Based on this information 23 study sites wereselected which build gradients spanning the entire ranges of slope and perimeter-to-area ratioand therefore represent the entire landscape.
Functional influence of landscape complexity on biodiversityAgricultural landscapes can be viewed as spatial mosaics of patches of arable land, meadows, andfallow land. We investigate two opposing processes influencing biodiversity in terms of speciesrichness: (1) Species richness increases with patch size because local species-extinctionprobabilities decrease. (2) Species richness decreases with increasing isolation among patches ofthe same land use type because recolonisation is more difficult2. As a consequence, we predictfairly high species richness at small patch sizes (isolation negligible). As patch size increases,isolation increases as well and local extinctions will be less likely to be counteracted bycolonisations, resulting in low species richness. However, as patch size increases further, localextinctions will be rare and species richness very high. Finally, at very large patch sizes, isolationbecomes practically perfect and biodiversity is low even though extinctions are unlikely (Fig. 1).The spatial scales at which the hypothesized pattern can be observed are a function of the‘ecological profile’ of the species investigated. Ecological profiles differ qualitatively in theirdispersal ability and habitat specificity. For example, isolation decreases with increasing dispersalability and decreasing habitat specificity.
To investigate these hypotheses, we developed a grid-based, spatially explicit metapopulationmodel. For model development, we had insects in mind. Each grid cell constitutes an area of aspecified land use type and may accommodate a number of populations of different ‘types’(specific quantitative representation of an ecological profile). All land parcels comprise severalcells. Local populations may go extinct and recolonization is possible due to dispersingindividuals managing to establish new local populations. We will run simulations for different
landscapes using types from within one and allecological profiles to quantify the specific andoverall effect of landscape structure onbiodiversity. Simulation results will becompared to the field surveys conducted bysubprojects 2 –4. Deviations between field andmodel results will lead to model modificationsand thereby to increasing understanding of theeffects of landscape complexity on biodiversity.These investigations will be accompanied by asecond, more detailed model restricted to aphiddynamics.
Fig. 1: Species richness as a function of average patch size.
References:1. Behrens, T., Purtauf, T., Wolters, V., Köhler, W., and Dauber, J. 2001. Study site selection and
gradient detection in complex landscapes using an automated patch detection tool (PaDS). Pages 217-220 in A. Belward et al., editors. Geo-spatial knowledge processing for natural resource management,University of Insubria, Varese (Italy).
2. Kruess, A., and Tscharntke, T. 2000. Species richness and parasitism in a fragmented landscape:experiments and field studies with insects on Vicia sepium. Oecologia 122:129-137.
Patch size
Spe
cies
R
ich
nes
s

70
Project ID: 01LC0020 (BIOPLEX Subproject 2)01.10.2000-30.09.2003
BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURALLANDSCAPES UNDER GLOBAL CHANGE (BIOPLEX) - AGROECOLOGY
Carsten Thies, Judith Schmid and Teja Tscharntke
Georg-August-University, Agroecology, Waldweg 26, 37073 Goettingen, GermanyContact: [email protected]; [email protected]; http://www.gwdg.de/~uaoe
Key words:biodiversity, ecological functions, landscape structure, landscape complexity, biologicalcontrol, trophic interactions, top down, bottom up, scale dependence, wheat, cereal aphids,parasitoids.
Abstract:The Agroecology group investigates the influence of landscape structure and farmingmanagement on biodiversity and functioning of ecological processes in agriculturallandscapes of northern and middle Germany. The analyses focus on species diversity andspecies interactions in wheatcrop fields and grasslands. In wheat fields, we address factorsthat affect the presence, abundance and species richness of natural enemies of cereal aphids(and weeds) as well as the role of pathogens (with project group 3: Phytopathology) andbelow- and above-ground processes (with project group 4: Soil Ecology) in relation tolandscape complexity. We evaluate the ability of ”natural enemies” of pests (and weeds) toact as ”biocontrol agents”, i.e. species which suppress the damage caused to crops by pestsbelow an economic threshold. In grasslands we focus on the combined effects of pollinatorsand seed predators on plant’s seed set. The analysis of the spatio-temporal variability of thesecounter-rotating processes will give a deeper insight to multitrophic interactions of importantfunctional groups in terrestrial ecosystems, and to modeling biodiversity (with project group1: Modeling and Geostatistics) at a range of spatial scales upscaling from microsites, via thefield and farm scale to landscapes and the global context.
Results:Since there is an increasing recognitation that community structure, species abundance andbiotic interactions may depend on scales much larger than a single habitat, a landscapeperspective considering spatial arrangement, connectivity and environment of habitats may beundispensable in determining local ecological pattern and processes (Kareiva 1990, Kruess &Tscharntke 1995, Roland und Taylor 1997, Thies & Tscharntke 1999). However, landscapecontext does not influence all species equally, and interacting communities are made up ofspecies with very different spatial strategies. Species show specific responses to landscapestructure facilitating or impeding movement among habitat patches, and with a specificperception of spatial scale of landscape complexity (With et al. 1999). Here, we present initialresults from herbivore-parasitoid interaction in wheat fields (Triticum aestivum) located in 46landscapes differing in structural complexity. We analysed the most important crop pest onwheat in Germany, the cereal aphid Sitobion avenae (Homoptera: Aphididae), and itsmortality due to parasitoids (Hymenoptera parasitica). The abundance of the aphid S. avenaeas well as its mortality due to parasitism was correlated with the perimeter/area ratio (P/A).The P/A ratio serves as a useful index of shape complexity in a landscape. For a given area, ahigh P/A ratio indicates a complex or elongated boundary shape, and a low P/A ratio indicatesa more compact and simple shape.
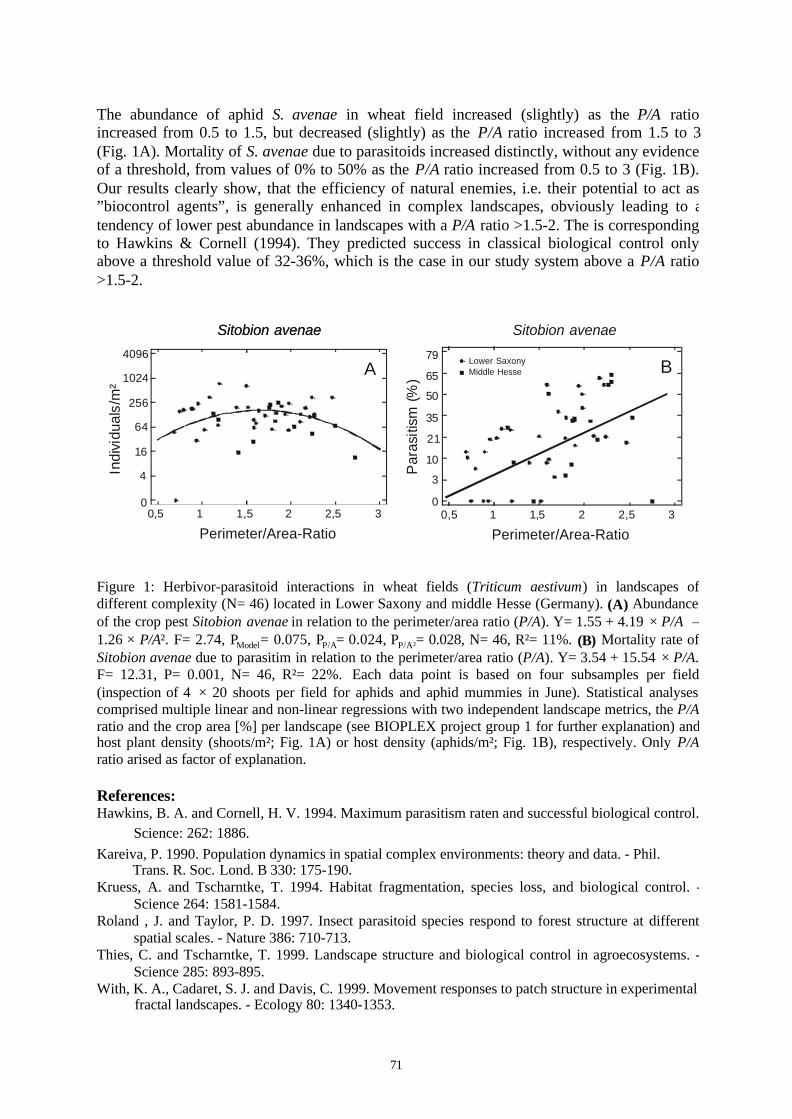
71
The abundance of aphid S. avenae in wheat field increased (slightly) as the P/A ratioincreased from 0.5 to 1.5, but decreased (slightly) as the P/A ratio increased from 1.5 to 3(Fig. 1A). Mortality of S. avenae due to parasitoids increased distinctly, without any evidenceof a threshold, from values of 0% to 50% as the P/A ratio increased from 0.5 to 3 (Fig. 1B).Our results clearly show, that the efficiency of natural enemies, i.e. their potential to act as”biocontrol agents”, is generally enhanced in complex landscapes, obviously leading to atendency of lower pest abundance in landscapes with a P/A ratio >1.5-2. The is correspondingto Hawkins & Cornell (1994). They predicted success in classical biological control onlyabove a threshold value of 32-36%, which is the case in our study system above a P/A ratio>1.5-2.
Figure 1: Herbivor-parasitoid interactions in wheat fields (Triticum aestivum) in landscapes ofdifferent complexity (N= 46) located in Lower Saxony and middle Hesse (Germany). (A) Abundanceof the crop pest Sitobion avenae in relation to the perimeter/area ratio (P/A). Y= 1.55 + 4.19 × P/A –1.26 × P/A². F= 2.74, PModel= 0.075, PP/A= 0.024, PP/A²= 0.028, N= 46, R²= 11%. (B) Mortality rate ofSitobion avenae due to parasitim in relation to the perimeter/area ratio (P/A). Y= 3.54 + 15.54 × P/A.F= 12.31, P= 0.001, N= 46, R²= 22%. Each data point is based on four subsamples per field(inspection of 4 × 20 shoots per field for aphids and aphid mummies in June). Statistical analysescomprised multiple linear and non-linear regressions with two independent landscape metrics, the P/Aratio and the crop area [%] per landscape (see BIOPLEX project group 1 for further explanation) andhost plant density (shoots/m²; Fig. 1A) or host density (aphids/m²; Fig. 1B), respectively. Only P/Aratio arised as factor of explanation.
References:Hawkins, B. A. and Cornell, H. V. 1994. Maximum parasitism raten and successful biological control.
Science: 262: 1886.Kareiva, P. 1990. Population dynamics in spatial complex environments: theory and data. - Phil.
Trans. R. Soc. Lond. B 330: 175-190.Kruess, A. and Tscharntke, T. 1994. Habitat fragmentation, species loss, and biological control. -
Science 264: 1581-1584.Roland , J. and Taylor, P. D. 1997. Insect parasitoid species respond to forest structure at different
spatial scales. - Nature 386: 710-713.Thies, C. and Tscharntke, T. 1999. Landscape structure and biological control in agroecosystems. -
Science 285: 893-895.With, K. A., Cadaret, S. J. and Davis, C. 1999. Movement responses to patch structure in experimental
fractal landscapes. - Ecology 80: 1340-1353.
Sitobion avenaeSitobion avenae Sitobion avenae
Indi
vidu
als/
m²
Par
asiti
sm (%
)
0
4
16
64
256
1024
4096
Perimeter/Area-RatioPerimeter/Area-Ratio
BA
0,50,5 11 1,51,5 22 2,52,5 330
3
10
21
35
50
65
79Middle HesseLower Saxony

72
Project ID: 01LC0020 (BIOPLEX Subproject 3)01.10.2000-30.09.2003
BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURALLANDSCAPES UNDER GLOBAL CHANGE (BIOPLEX) - PHYTOPATHOLOGY
Lars Reimer, Julia Morzfeld, Stefan Vidal
Institute of Plant Pathology and Plant Protection, Goettingen University, Grisebachstr 6, [email protected]; http://www.uni-goettingen.de
Key words: Sitobion avenae, DNA-Fingerprinting, Septoria tritici, genetic variability,landscape-structure, agricultural utilization
Abstract:The BIOPLEX subproject Phytopathology aims at evaluating the influence of landscapestructure on population processes at the genetic level. Using cereal aphid species and plantpathogens on winter wheat as targets we investigate the genetic diversity of these specieswithin and between fields at a local and a regional scale to evaluate the influence of thestructural and agronomical diversity of the surrounding landscape at different spatial scales.The project started with the year 2001. Sampling methods were determined in cooperation withother subprojects. Samples were taken two or three times, respectively (according to specificdevelopmental stages of the wheat) in up to 40 fields in the environment of Goettingen and in theLahn-Dill-Bergland. The wheat fields were selected by means of GIS-Analysis. During samplingdates additional plants were collected to determine biomass as a covariate. On each field in theenvironment of Goettingen small areas were excluded from insecticide application for aphidsampling. All farmers were questioned on information concerning agricultural cultivationtreatments (tillage strategies, application of fungicides, fertilization, cultivars and crop rotation).
Insects (L. Reimer)About 30 to 100 cereal aphids, predominantly Sitobion avenae, were collected during twosampling dates from 36 field sites in the region around Giessen and Goettingen. Sweepnetcollections were performed along a transect, the length of which was adapted to field size andthe number of aphids available. Aphid captures were located at least one meter apart fromeach other to collect as many clones within one field as possible. >From each sweepnetcapture only one nonparasitised individual, identified to species level, was taken for furtherDNA analysis. Indentifications were verified by Dr. T. Thieme (BTL GmbH Sagerheide).Samples were stored either deep frozen at –20 °C or in 99 % Ethanol. For the analysis ofmicrosatellite length polymorphism (Sunnucks et al. (1)) six primers are actually available.Between 91 and 127 individuals from five field sites of the Goettingen region were screenedfor five primers and 5 to 25 different alleles were found per locus. The number of alleles sofar found is as high or even higher than reported in papers that used a higher number ofindividuals. The distribution of distinct alleles within a site appears clumped with regard tocertain alleles. An example on the allele frequencies at one locus is given in Tab. 1. There arepredominant alleles and also rare ones. The allelic distribution seems to be similar at differentfield sites, but some shifts in allele frequencies do occur (field site 2/1, Tab. 1). Thegenotypes, defined as individuals sharing the same alleles at all loci, show a high diversity.Only two genotypes were found more than once in the different field sites. These genotypesare homozygous at three or four loci and consist mainly of common alleles.
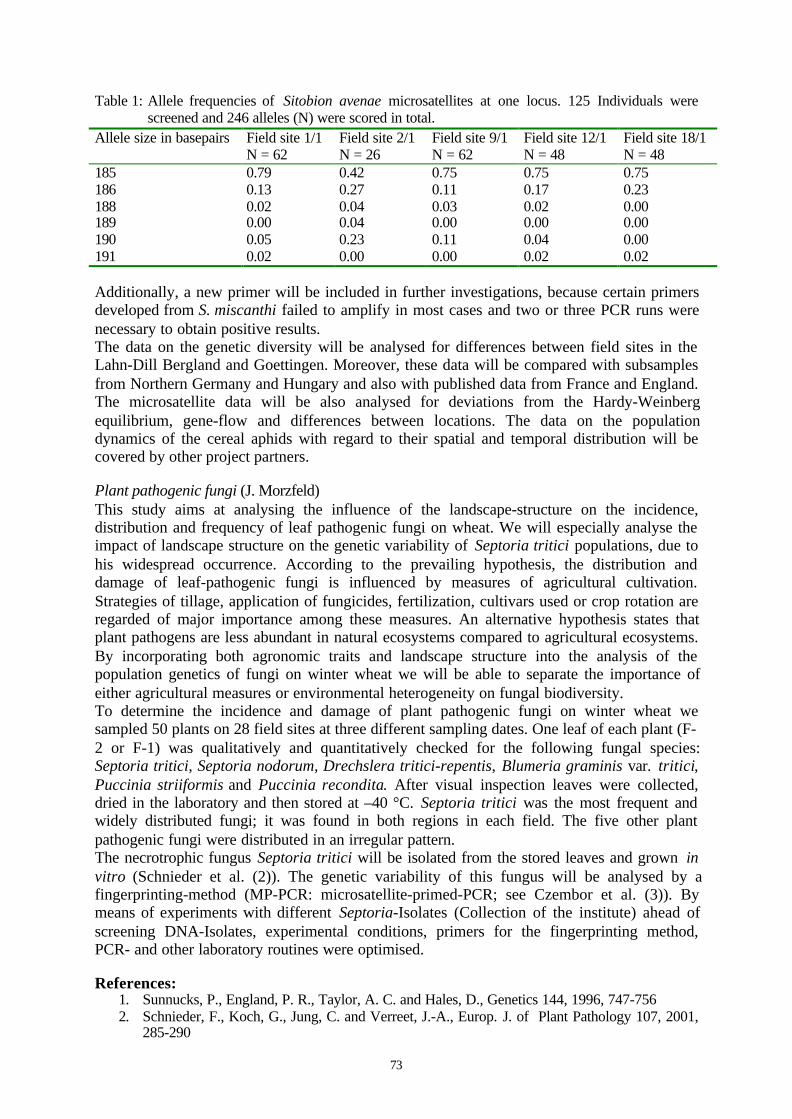
73
Table 1: Allele frequencies of Sitobion avenae microsatellites at one locus. 125 Individuals werescreened and 246 alleles (N) were scored in total.
Allele size in basepairs Field site 1/1N = 62
Field site 2/1N = 26
Field site 9/1N = 62
Field site 12/1N = 48
Field site 18/1N = 48
185 0.79 0.42 0.75 0.75 0.75186 0.13 0.27 0.11 0.17 0.23188 0.02 0.04 0.03 0.02 0.00189 0.00 0.04 0.00 0.00 0.00190 0.05 0.23 0.11 0.04 0.00191 0.02 0.00 0.00 0.02 0.02
Additionally, a new primer will be included in further investigations, because certain primersdeveloped from S. miscanthi failed to amplify in most cases and two or three PCR runs werenecessary to obtain positive results.The data on the genetic diversity will be analysed for differences between field sites in theLahn-Dill Bergland and Goettingen. Moreover, these data will be compared with subsamplesfrom Northern Germany and Hungary and also with published data from France and England.The microsatellite data will be also analysed for deviations from the Hardy-Weinbergequilibrium, gene-flow and differences between locations. The data on the populationdynamics of the cereal aphids with regard to their spatial and temporal distribution will becovered by other project partners.
Plant pathogenic fungi (J. Morzfeld)This study aims at analysing the influence of the landscape-structure on the incidence,distribution and frequency of leaf pathogenic fungi on wheat. We will especially analyse theimpact of landscape structure on the genetic variability of Septoria tritici populations, due tohis widespread occurrence. According to the prevailing hypothesis, the distribution anddamage of leaf-pathogenic fungi is influenced by measures of agricultural cultivation.Strategies of tillage, application of fungicides, fertilization, cultivars used or crop rotation areregarded of major importance among these measures. An alternative hypothesis states thatplant pathogens are less abundant in natural ecosystems compared to agricultural ecosystems.By incorporating both agronomic traits and landscape structure into the analysis of thepopulation genetics of fungi on winter wheat we will be able to separate the importance ofeither agricultural measures or environmental heterogeneity on fungal biodiversity.To determine the incidence and damage of plant pathogenic fungi on winter wheat wesampled 50 plants on 28 field sites at three different sampling dates. One leaf of each plant (F-2 or F-1) was qualitatively and quantitatively checked for the following fungal species:Septoria tritici, Septoria nodorum, Drechslera tritici-repentis, Blumeria graminis var. tritici,Puccinia striiformis and Puccinia recondita. After visual inspection leaves were collected,dried in the laboratory and then stored at –40 °C. Septoria tritici was the most frequent andwidely distributed fungi; it was found in both regions in each field. The five other plantpathogenic fungi were distributed in an irregular pattern.The necrotrophic fungus Septoria tritici will be isolated from the stored leaves and grown invitro (Schnieder et al. (2)). The genetic variability of this fungus will be analysed by afingerprinting-method (MP-PCR: microsatellite-primed-PCR; see Czembor et al. (3)). Bymeans of experiments with different Septoria-Isolates (Collection of the institute) ahead ofscreening DNA-Isolates, experimental conditions, primers for the fingerprinting method,PCR- and other laboratory routines were optimised.
References:1. Sunnucks, P., England, P. R., Taylor, A. C. and Hales, D., Genetics 144, 1996, 747-7562. Schnieder, F., Koch, G., Jung, C. and Verreet, J.-A., Europ. J. of Plant Pathology 107, 2001,
285-290

74
3. Czembor, P. C. and Arseniuk, E., J. Phytopathology 147, 1999, 539-546Project ID: 01LC0020 (BIOPLEX Subproject 4)1.10.2000 – 30.09.2003
BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURALLANDSCAPES UNDER GLOBAL CHANGE (BIOPLEX) - Soil BIOTA
J. Dauber, T. Purtauf and V. Wolters
Department of Animal Ecology, Justus-Liebig-University, Heinrich-Buff-Ring 26-32 (IFZ), D-35392 [email protected], http://www.uni-giessen.de/bioplex
Key words: landscape complexity, soil fauna, ground beetles, ants, wheat, grassland
Abstract:The character of Central European landscapes is shaped by human land-use, especiallyagriculture. The continuous expansion of agriculture over the last centuries created openlandscapes with new habitats and led to an increase of biodiversity. Therefore biodiversity ofCentral Europe depends on agriculture. However, industrialisation of agriculturalproduction and intensification of land-use over the last decades led to enormous habitatdestruction and thereby to a decrease in biodiversity. The question arising from thisdevelopment is: What kind of agricultural landscape do we need to sustain high levels ofbiodiversity? The multidisciplinary project BIOPLEX approaches this question by analysingthe relationship between biodiversity and landscape complexity at different spatial scales.The studies focus on wheat fields and managed grasslands in two agricultural regions ofGermany (Lahn-Dill-Bergland, Hesse and Southern Lower Saxony). The subproject SoilEcology aims at: (i) recording the species richness of soil living macroinvertebrates (groundbeetles, rove beetles, woodlice, diplopods) and soil engineers (ants), (ii) analysing the effectof epigaeic predatory arthropods on cereal aphids in winter wheat, and (iii) quantifying thefunctional role of decomposers depending on landscape complexity. A comparison betweenthe data derived from these analyses with data on phytophagous insects, plant pathogens andvegetation will reveal some of the relationships between above- and belowgroundbiodiversity.
Results:Sustainable management of biodiversity at the landscape level urgently needs simpleindicators of biodiversity. Landscape structure affects ecological processes and speciesrichness (Wiens et al. 1993). Landscape pattern descriptors, which correspond to spatialcomplexity and to varying amounts of land-use types might serve as useful tools to explaindifferences in species richness across landscapes (Miller et al. 1997). Field experiments andsurveys on this topic rely on representative selection of study sites by landscape analyses,taking into calculation the landscape features of interest for the respective survey. Ideally,there is a small set of metrics, which span the important dimensions of landscape structure(Riitters el al. 1995). The selection of study sites in a gradient of landscape complexity isshown in Behrens et al. 2001. Base maps used for the study site selection were a DEM(Digital Elevation Model) and the official German land-use database called ATKIS(Amtliches topographisch-kartographisches Informationssystem). The decision to use thesebase maps was taken on base of the results of a preliminary study out of which some of theresults are presented here:
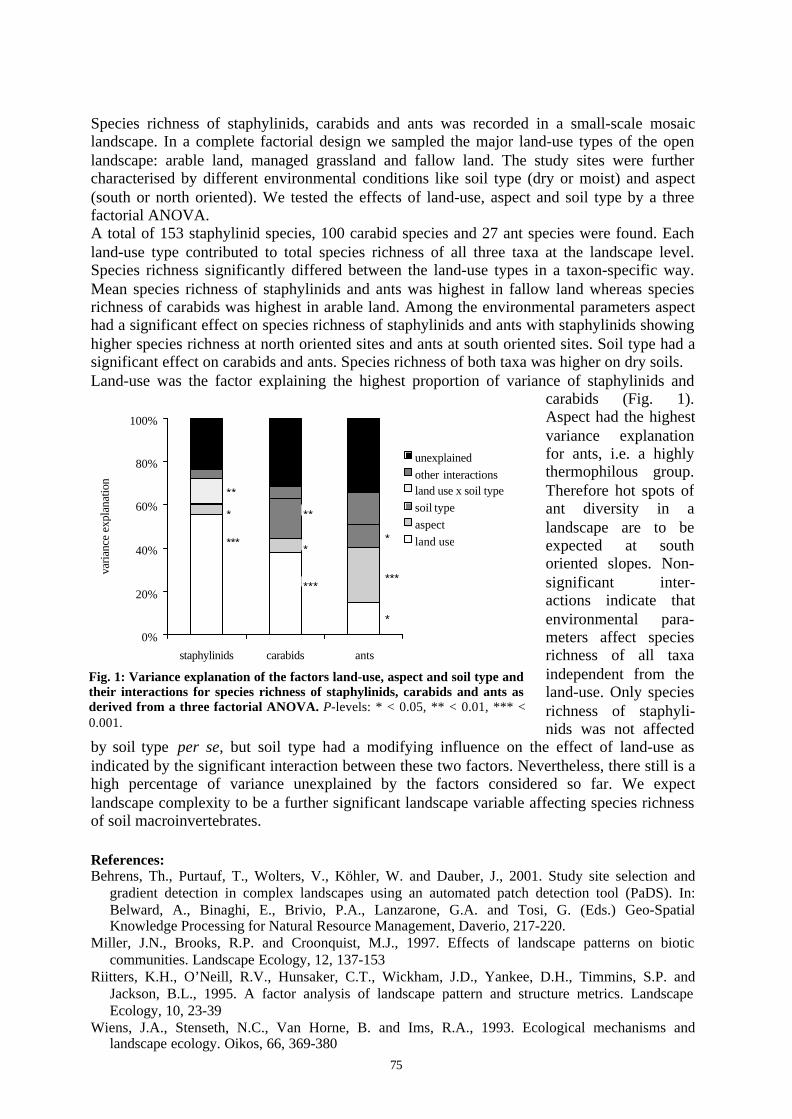
75
0%
20%
40%
60%
80%
100%
staphylinids carabids ants
varia
nce
expl
anat
ion
unexplainedother interactionsland use x soil typesoil typeaspectland use
*
***
*
***
*
**
***
*
**
Species richness of staphylinids, carabids and ants was recorded in a small-scale mosaiclandscape. In a complete factorial design we sampled the major land-use types of the openlandscape: arable land, managed grassland and fallow land. The study sites were furthercharacterised by different environmental conditions like soil type (dry or moist) and aspect(south or north oriented). We tested the effects of land-use, aspect and soil type by a threefactorial ANOVA.A total of 153 staphylinid species, 100 carabid species and 27 ant species were found. Eachland-use type contributed to total species richness of all three taxa at the landscape level.Species richness significantly differed between the land-use types in a taxon-specific way.Mean species richness of staphylinids and ants was highest in fallow land whereas speciesrichness of carabids was highest in arable land. Among the environmental parameters aspecthad a significant effect on species richness of staphylinids and ants with staphylinids showinghigher species richness at north oriented sites and ants at south oriented sites. Soil type had asignificant effect on carabids and ants. Species richness of both taxa was higher on dry soils.Land-use was the factor explaining the highest proportion of variance of staphylinids and
carabids (Fig. 1).Aspect had the highestvariance explanationfor ants, i.e. a highlythermophilous group.Therefore hot spots ofant diversity in alandscape are to beexpected at southoriented slopes. Non-significant inter-actions indicate thatenvironmental para-meters affect speciesrichness of all taxaindependent from theland-use. Only speciesrichness of staphyli-nids was not affected
by soil type per se, but soil type had a modifying influence on the effect of land-use asindicated by the significant interaction between these two factors. Nevertheless, there still is ahigh percentage of variance unexplained by the factors considered so far. We expectlandscape complexity to be a further significant landscape variable affecting species richnessof soil macroinvertebrates.
References:Behrens, Th., Purtauf, T., Wolters, V., Köhler, W. and Dauber, J., 2001. Study site selection and
gradient detection in complex landscapes using an automated patch detection tool (PaDS). In:Belward, A., Binaghi, E., Brivio, P.A., Lanzarone, G.A. and Tosi, G. (Eds.) Geo-SpatialKnowledge Processing for Natural Resource Management, Daverio, 217-220.
Miller, J.N., Brooks, R.P. and Croonquist, M.J., 1997. Effects of landscape patterns on bioticcommunities. Landscape Ecology, 12, 137-153
Riitters, K.H., O’Neill, R.V., Hunsaker, C.T., Wickham, J.D., Yankee, D.H., Timmins, S.P. andJackson, B.L., 1995. A factor analysis of landscape pattern and structure metrics. LandscapeEcology, 10, 23-39
Wiens, J.A., Stenseth, N.C., Van Horne, B. and Ims, R.A., 1993. Ecological mechanisms andlandscape ecology. Oikos, 66, 369-380
Fig. 1: Variance explanation of the factors land-use, aspect and soil type andtheir interactions for species richness of staphylinids, carabids and ants asderived from a three factorial ANOVA. P-levels: * < 0.05, ** < 0.01, *** <0.001.

76
Project ID: 01LC0020 (BIOPLEX Subproject 5)01.11.2000 – 31.10.2003
BIODIVERSITY AND SPATIAL COMPLEXITY IN AGRICULTURALLANDSCAPES UNDER GLOBAL CHANGE (BIOPLEX) -SOCIAL-ECONOMIC EVALUATION AND APPRAISAL WITH RESPECT TOBIODIVERSITY IN AGRARIAN LANDSCAPES
Elke Bertke, Katja Diedrich, Anke Fischer, Sonja Katerina Hespelt, Christoph Tute,Dr. Bärbel Gerowitt, Prof. Dr. Johannes Isselstein, Prof. Dr. Rainer Marggraf
Research Centre for Agriculture and the EnvironmentFaculty of Agriculture, Georg-August-University of Göttingen,Am Vogelsang 6, 37075 D-Gö[email protected]; http://www.uni-goettingen.de/zlu (BIOPLEX - Social-economic concepts)
Key words: system of payment, ecological services, information, contingent valuation method,biodiversity, production value, agriculture
Abstract:The subproject V is structured into three parts. The part "Result-Oriented System of Payment forEcological Services“ deals with the development of an economic system of payment for ecologicalservices provided by farmers. These services will be measured by ecological goods arising fromplant biodiversity. The part of the project that deals with “The Influence of Information on theEconomic Valuation of Natural Resources” has two objectives. On the one hand, it aims at aneconomic evaluation of ecological goods defined in the first part of the subproject (s. a.). On theother hand, different influences of information that occur during the process of evaluation in acontingent valuation, are investigated in an interdisciplinary approach. Another part of theproject concerns „The Impact of Certain Components of Biodiversity on the Production Value inAgriculture“. This aims at assessing the utility certain components of biodiversity and,simultaneously, ecological goods have got for agriculture.
Results:“Result-Oriented System of Payment for Ecological Services“Ecological services performed by the farming community are positive external effects. At presentsome of these ecological services are financed under agri-environmental programs or agri-environmental contracting. In these programs the farmers get paid for acting according to specialcropping instructions, e.g. dispensing pesticide use.In theory, such services could be increased if the financial incentives on offer are improved. Ourresearch project focuses on developing a concept of a regional market and result-oriented systemof payment for ecological services provided by farmers. The results of ecological services areecological goods. Goods arising from plant biodiversity can be considered to be those mostsuitable for a result-oriented system of payment. These ecological goods should be supplied bythe farmers voluntarily and without set regulations. Furthermore, the public demand for ecologicalgoods should be represented by a regional advisory board in the sense of the Europeansubsidiarity principle. This advisory board has to award public contracts for the production ofecological goods.In this way, environmental programs could be made more flexible and the farmers would receivefinancial incentives to find innovative solutions for an environmentally compatible management.

77
The concept further takes into account the preferences of the local population for ecological goodsand for the allocation of public funds.In our transdisciplinary research work we develop this result-oriented system of payment for arepresentative region in the south of Lower Saxony (County Northeim) and check the possibilitiesof its application in practice with the relevant institutions of that county. The members of theregional advisory board represent the administrations of agriculture and environment protection,the local affairs, and the associations of agriculture, conservation, and land owners. Up to now wedefined regional specific ecological goods associated with grassland use, we worked outmanagement proposals to obtain these ecological goods, and calculated the costs. The awardprocedures for public orders, complying with the law, were examined with regard to theirtransferability on a market for ecological goods.
“The Influence of Information on the Economic Valuation of Natural Resources“Non-market common goods, such as ecological commodities, cannot be evaluated by their marketprice. To estimate their monetary value, ecological economists have developed several methods ofindirect and direct evaluation, for example, the contingent valuation method (CVM), which elicitswillingness to pay-values by means of an interview.In this study, CVM will be applied to estimate the willingness to pay of the citizens living in therepresentative region, county Northeim (s. a.), for certain ecological goods (s. a.), such ashedgerows in agrarian landscapes. At the same time, the impacts of information imparting andinformation processing on the valuation of the ecological good in question during an interviewsituation are investigated. For this purpose, in a first step the findings of other disciplines havebeen summarised and analysed, based on a common theoretical framework, the Theory ofPlanned Behaviour. In a second step, hypotheses about information processing and decisionmaking of respondents in CV situations have been derived from this analysis. They will be testedin a study orientated by findings and methods of social and consumer psychology.
“The Impact of Certain Components of Biodiversity on the Production Value in Agriculture“Local explorations of natural sciences in this field have shown that certain components ofbiodiversity may be economically important for agricultural production. With the aid of an expertquestionnaire, these sporadic results shall be tested by their relevance in different agriculturallandscapes, and the capability of generalising them be assessed, respectively.So far, the recent results presented in literature have been explored and summarised in causes andeffects. As the analysis will be conducted according to the Delphi method, the summaries willnow be used for developing a standardised questionnaire. With the aid of the Delphi-Questioning,the opinion of a group of experts will be ascertained and structured. As the experts stayanonymous in this kind of questionnaire, biases in the results which may result from the opinionsof too strong characters can be avoided. This is, however, often the case in group discussions.Apart from that, various data bases have been checked to find persons with expertise in this fieldof research. This is crucial as the quality of the results depends substantially on a good choice ofexperts.

78
Project ID: 01LC0021 (Subproject 1)1.1.2000 - 31.12.2003
BIODIVERSITY OF NITROGEN-FIXING MICROORGANISMS
Lei Zhang,1 Torsten Thünen,1 Thomas Hurek,2 and Barbara Reinhold-Hurek1
1Laboratorium für Allgemeine Mikrobiologie, 2 Kooperationslabor des Max-Planck-Institutsfür marine Mikrobiologie Bremen, Universität Bremen FB 2, Leobener Strasse NW2, D-28359 [email protected], [email protected]
Key words: Uncultured bacteria, nitrogenase genes, rice roots, DNA-microarrays, mRNAdetection
Abstract:The project aims to analyze the biodiversity of nitrogen-fixing prokaryotes by molecular andclassical techniques. During this project, the molecular methods to address questions ofterrestrial biodiversity are developed and established: Development of monitoring systems forfast, reliable assessment of the biodiversity of diazotrophic bacteria in association with plantroots, and for analysis of the dynamics of their population structure and their key activities(expression of nitrogenase genes). Protocols were developed to quantify bacterial nifH(nitrogenase gene) mRNA extracted from roots, and clone libraries of bacterial nifH genescompleted, demonstrating that the majority of rice root-associated diazotrophs has not yetbeen cultured. One of the major achievements is the development of a DNA-microarray whichcan specifically detect and identify a large variety of specific nitrogenase gene fragmentsamplified by PCR, without the necessity for clone libraries.
Results:The diversity of prokaryotic organisms is classically estimated by attempts to cultivate them andby characterization of the isolates. However, during the last decade DNA- or RNA- basedtechniques led to the conclusion that the organismal diversity is much higher than estimated bythese classical techniques: the majority of bacteria present in a given habitat has in most cases notbeen cultured as yet, and often these bacteria are phylogenetically very distant to known,cultivated bacterial strains [1]. This is also the case for root-associated bacteria [2, 3]. In order toassess the biodiversity and the population structure in a given habitat, their changes in response toenvironmental changes, and also in order to understand functions of microorganisms in thehabitat, culture-independent methods are indispensable.The most widely used target molecules for molecular ecological studies are ribosomal rRNAgenes [1, 4]. This allows an estimation of the biodiversity, but does –in most cases- not permit tolink the results to physiological functions of the bacteria. Functions however may be analyzedwhen key genes for certain metabolic processes are used as target genes. In the global nitrogencycle, biological fixation of atmospheric nitrogen is an important microbial activity. Theavailability of combined nitrogen affects plant productivity and the structure of the plantcommunity [5]. Increasing anthropogenic activities result in an increased contamination of theatmosphere by nitrogenous compounds, and in deposition of nitrogen from the atmosphere onsoils, even without application of fertilizer [6]. N-deposits are also likely to affect the populationof diazotrophs. One of the structural genes of nitrogenase, nifH, is particularly suitable forphylogenetic analysis [3, 7].We established protocols which allow to retrieve bacterial mRNA from soil-grown rice roots. Thedegradation of RNA was sufficiently inhibited for the detection of nifH mRNA by reversetranscriptase (RT) analysis [8]. Template dilution allowed a quantitative analysis of transcript

79
abundance, the application to different rice varieties revealing significant differences. In clonelibraries constructed from these amplification products, most sequences analyzed were onlydistantly related to nitrogenase genes of known, cultured bacteria. Thus, up to now uncultureddiazotrophs appear to be predominantly active in rice roots.A new method for the assessment of biodiversity is being developed which may allow the specificdetection and quantification of a large numbers of different nifH mRNA fragments withoutconstruction and analysis of clone libraries. DNA microarrays or DNA-chips are widely used forhighly parallel and high througput analyses of gene expression in genome research.Oligonucleotides or DNA-fragments which are complementary to target genes are linked tocarriers such as membranes or glass slides, and used as baits for hybridization with labeled DNAor RNA under study. This methodology is currently adopted by us to construct a „phylo-chip“, aglass microarray for specific detection of the diversity of nif genes. Our database of nifH genesfrom cultured diazotophs and fragments retrieved from environmental samples was used to designshort oligonucleotides which are specific to single genes or groups of genes with at least 1-2mismatches to each other. The oligonucleotides were attached to activated glass slides via amino-links at specific postions, hybridized to fluorescence (Cy-5)-labeled amplification products of anifH-directed PCR, and the hybridizations quantified with a microarray scanner. Theoligonucleotide design was optimized with respect to the position of hybridization within thefragment and other factors. Using genes of the Azoarcus group as examples, a hybridizationprotocol was developed in which one mismatch was sufficient to result in a specific hybridizationsignal. Thus from mixtures of DNA of different bacteria, their nitrogenases were specificallyidentified. The next steps of development will be the detection of root-associated nifH fragmentsusing the phylo-chip, and the enlargement of the number of oligonucleotide baits per chip to covermost of the nitrogenase gene diversity.
References:1. Pace, N.R. (1997) A molecular view of the microbial diversity and the biosphere. Science 276: 734-740.2. Engelhard, M., Hurek, T. and Reinhold-Hurek, B. (2000) Preferential occurrence of diazotrophic
endophytes, Azoarcus spp., in wild rice species and land races of Oryza sativa in comparison withmodern races. Environ. Microbiol. 2: 131-141.
3. Ueda, T., Suga, Y., Yahiro, N. and Matsuguchi, T. (1995b) Remarkable N2-fixing bacterial diversitydetected in rice roots by molecular evolutionary analysis of nifH gene sequences. J. Bacteriol. 177:1414-1417.
4. Amann, R.I., Ludwig, W. and Schleifer, K.H. (1995) Phylogenetic identification and in situ detection ofindividual microbial cells without cultivation. Microbiol. Rev. 59: 143-169.
5. Bakker, J.P. and Berendse, F. (1999) Constraints in the restoration of ecological diversity in grasslandand heathland communities. TREE 14: 63-68.
6. Vitousek, P.M., Aber, J.D., Howarth, R.W., Likens, G.E., Matson, P.A., Schindler, D.W., Schlesinger,W.H. and Tilman, D.G. (1997) Human alteration of the global nitrogen cycle: sources andconsequences. Ecol. Appl. 7:737-750.
7. Hurek, T., Egener, T. and Reinhold-Hurek, B. (1997) Divergence in nitrogenases of Azoarcus spp.,Proteobacteria of the ß-subclass. J. Bacteriol. 179: 4172-4178.
8. Hurek, T., Handley, L., Reinhold-Hurek , B. and Piché, Y. (2001) Azoarcus grass endophytes function innitrogen fixation by contributing fixed nitrogen to the plant in an unculturable state. Mol. Plant-Microb. Interact.: in press.

80
Project ID: 01LC0021 (Subproject 2)1.1.2001 – 31.12.2001
BIODIVERSITY OF NITRIFYING AND SULFATE-REDUCING PROKARYOTES
A. Loy, J. Adamczyk, A. Lehner, and M. Wagner
Lehrstuhl für Mikrobiologie, Technische Universität München, Am Hochanger 4, [email protected]; www.microbial-ecology.de
Key words: Nitrification, sulfate-reduction, 16S rDNA, DNA-microarray, ammonia-monooxygenase gene amoA, dissimilatory sulfite reductase genes dsrA/B
Abstract:Nitrifying and sulfate-reducing prokaryotes catalyze key steps in the nitrogen and carbon-cycle,respectively, and are thus of primary importance for the ecology of most ecosystems. The projectaims to develop and adapt methods suitable for cultivation-independent investigations of thediversity and function of nitrifying (ammonia-oxidizing and nitrite-oxidizing) and sulfate-reducingprokaryotes living in the rhizosphere. For both groups of microorganisms 16S rDNA as well assuitable functional genes are used as marker molecules (1, 2 ,4 ,6). Based on encompassing datasets of these marker molecules, oligonucleotide microarrays for highly parallel, high throughputscreening of these prokaryotes in environmental samples were developed and initially applied toinvestigate samples from a Sphagnum bog. These techniques will be supplemented by optimizingthe combination of FISH and microautoradiography (3) for its use in rhizosphere and soil inorder to determine the links between the availability of specific substrates (for example by plantroot exudation) and the microbial biodiversity. These investigations will provide novel insightsinto diversity and activity patterns of soil and rhizosphere microorganisms and into interactionsbetween plants and microorganisms.
Results:During the first project phase we have established encompassing 16S rDNA data sets fornitrifying and sulfate-reducing prokaryotes. Based on this information, microarrays for nitrifyingand sulfate-reducing bacteria were designed. These arrays, which consist of several hundredoligonucleotide probes, were thoroughly evaluated under different hybridization conditions usingreference sets of suitable pure cultures and are now ready for application in environmentaldiversity surveys. Furthermore, we started to design and evaluate DNA microarrays for ammonia-oxidizing and sulfate-reducing prokaryotes by exploiting the genes encoding the ammonia-monooxygenase (amoA) and the dissimilatory sulfite-reductase (dsrA/B), respectively, as targetmolecules.
The developed microarrays are now used to investigate the diversity of nitrifiers and sulfate-reducers in a variety of different rhizosphere samples. In collaboration with Prof. Drake and Dr.Küsel (Lehrstuhl für ökologische Mikrobiologie, Bayreuth) we also began to investigate thecommunity composition of sulfate-reducing prokaryotes in different depths of a Sphagnum-bog, asystem for which sulfate-reduction rates comparable to those of marine systems have beenreported (for example 5).

81
References:1. Daims, H., Nielsen, J.L., Nielsen, P.H., Schleifer, K.H., and Wagner, M., Appl. Environ. Microbiol.,
2001, 67,5273-5284.2. Klein, M., Friedrich, M., Roger, A.J., Hugenholtz, P., Fishbain, S., Abicht, H., Blackall, L.L., Stahl,
D.A., and Wagner, M., J. Bacteriol., 2001, 183,6028-6035.3. Lee, N., Nielsen, P. H., Andreasen, K. H., Juretschko, S., Nielsen, J. L., Schleifer, K. H., and Wagner,
M., Appl. Environ. Microbiol, 1999, 65,1289-1297.4. Purkhold, U., Pommerening-Röser, A., Juretschko, S., Schmid, M.C., Koops H.P., and Wagner, M.
Appl. Environ. Microbiol., 2000, 66, 5368–5382.5. Urban, N.R., Eisenreich, S.J., and Grigal, D.F., Biogeochem., 1989 7,81-90.6. Wagner, M, Flax, J. L., Brusseau, G. A., and Stahl, D. A., J. Bacteriol., 1998, 180, 2975-2982.

82
Project ID: 01LC0021 (Subproject 3)1.1.2000 – 31.12.2003
BIODIVERSITY OF METHANOTROPHIC AND NITRATE-REDUCING BACTERIA
G. Braker, P. Dunfield, H.P. Horz and R. Conrad
Max-Planck-Institut für terrestrische Mikrobiologie, Karl-von-Frisch-Str., 35043 [email protected]; http://www.uni-marburg.de/mpi
Key words: methane oxidation, nitrate reduction, denitrification, nitric oxide, nitrous oxide
Abstract:We investigate microbial processes in soils. The research focuses on the question which processeson a microscopic scale are catalyzed by which microorganisms. Besides the functional diversityin soil with respect to processes, there is an even higher structural diversity with respect to themicroorganisms that are responsible for the functions. The extent of the soil microbial diversity iscompletely unknown. In particular, it is unclear how much microbial diversity is required tomaintain soil functions. Within BIOLOG we study the biodiversity of methanotrophic bacteria andnitrate-reducing bacteria in relation to their functioning in soil environments. Biodiversity isstudied by targeting both the 16S rRNA genes and the structural genes of key enzymes. Here, wetargeted the pmoA genes of methanotrophs coding for the particulate methane monooxygenase,and the nirK/nirS and norB genes of denitrifiers coding for nitrite reductase and nitric oxidereductase, respectively.
Results:We have studied the diversity of the methanotrophic microbial community (i) in rice field soilsfrom different geographic regions1; and (ii) in a meadow soil with seasonally changing watertable2. Furthermore, (iii) the genetic diversity of denitrifiers was explored by PCR amplification ofnirK/nirS3 and norB4 genes from pure cultures and environmental samples.
(i) Methanotrophic bacteria play a crucial role in regulating the emission of CH4 from rice fieldsinto the atmosphere. We investigated the CH4 oxidation activity together with the diversity ofmethanotrophic bacteria in ten rice field soils from different geographic locations. Uponincubation of aerated soil slurries under 7% CH4, rates of CH4 oxidation increased after a lagphase and reached values of 3-10 µmol d-1g-dw-1 soil. The methanotrophic community wasassayed by retrieval of the pmoA gene which encodes the a subunit of the particulate methanemonooxygenase. After extraction of DNA from actively CH4-oxidizing soil samples and PCR-amplification of the pmoA, the community was analyzed by Denaturant Gradient GelElectrophoresis (DGGE) and Terminal Restriction Fragment Length Polymorphism (T-RFLP).DGGE bands were excised, the pmoA re-amplified, sequenced and the encoded amino acidsequence comparatively analyzed by phylogenetic treeing. The analyses allowed the detection ofpmoA sequences related to the following methanotrophic genera: the type-I methanotrophsMethylobacter, Methylomicrobium, Methylococcus and Methylocaldum, and the type-IImethanotrophs Methylocystis and Methylosinus. T-RFLP analysis detected a similar diversity, buttype-II pmoA more frequently than DGGE. All soils but one contained type-II in addition to type-Imethanotrophs. Type-I Methylomonas was not detected at all. Different combinations ofmethanotrophic genera were detected in the different soils. However, there was no obviousgeographic pattern of the distribution of methanotrophs.

83
(ii) Methane oxidation activity and the structure of the methane-oxidising microbial community ina wet meadow soil in Germany were investigated using biogeochemical, culturing, and molecularfingerprinting techniques. Most-probable-number counting of methane-oxidising bacteriafollowed by isolation and characterisation of strains from the highest positive dilution stepssuggested that the numerically dominant member of the methane-oxidising community was aMethylocystis strain (≅106 cells g-1 d.w. soil). Calculations based on kinetic data support theconclusion that this organism could account for most of the observed methane oxidation activityin the soil. DNA extraction directly from soil and PCR amplification of the pmoA gene alsorecovered Methylocystis-like sequences. However, molecular community fingerprinting analysesalso revealed a more diverse and dynamic picture of the methane-oxidising community, and therewere large variations across samples. Retrieved pmoA sequences included, besides Methylocystissequences, others related to the genera Methylomicrobium and Methylocapsa, and a novel lineage.A combination of classical microbiological methods and molecular fingerprinting therefore gave abetter picture of the microbial community in this meadow soil than either method alone, i.e. themolecular methods revealed a higher species richness and uncovered the presence ofMethylocapsa, but failed to demonstrate the consistently high abundance of Methylocystis, andoccasionally even failed to detect this organism despite its abundance.
(iii) The genes nirK and nirS encode two structurally different but functionally equivalentenzymes occurring within the same genus but not within the same strain of a denitrifier. Twoclasses of norB genes exist with low levels of identity: one type with an adjacent norC gene and asecond type with a N-terminal extension instead. Amplification of nirK, nirS, and norB wassuccessful from pure cultures within the Proteobacteria. Cloning of nir genes from environmentalsamples revealed gene clusters distinct from those of known denitrifiers suggesting novel lineagesof yet uncultivated denitrifiers, probably also within the Proteobacteria. Gene clusters werespecific for the habitat from which these clones were obtained. Phylogenetic analysis of norBgrouped genes according to two classes; within the norCB class, norB genes from isolatesgrouped together with clones suggesting that most of these environmental sequences might bederived from Proteobacteria. Within the second class, genes from all known isolates except onewere grouped in one cluster, clones were unrelated and scattered. Interestingly, in contrast to nirgenes, which clustered strongly according to the environment from which they were derived, thiswas not observed for norB. With our current knowledge, the origin of novel sequences and theirdistribution patterns in the environment remain obscure. Future research will focus on exploringdenitrification genes from denitrifiers from culture collections and isolates from environmentalsamples to extend the databases of functional genes by also considering the phylogeny of theorganisms.
References:1. Hoffmann T, Horz HP, Conrad R, submitted.2. Horz HP, Raghubanshi AS, Heyer J, Kammann C, Conrad R, Dunfield P, submitted.3. Braker G, Fesefeldt A, Witzel K.P. 1998. Appl. Environ. Microbiol. 64:3769-3775.4. Braker G, Tiedje, JM. 2001. Poster VAAM Jahrestagung 2001.

84
Project ID: 01LC0021 (Subproject 4)1.1.2001 – 31.12.2003
BIODIVERSITY AND SUCCESSIONS IN TEMPORARY TERRESTRIAL-AQUATICINTERFACES
J. Pernthaler, A. Pernthaler, F. Warnecke and R. Amann
Max-Planck-Institut für Marine Mikrobiologie, Celsiusstrasse 1, D-28359 [email protected]
Key words : Actinobacteria, automated microscopy, substrate gradients
Abstract:The diversity of microorganisms found in freshwater aquatic habitats is probably a small subsetof the total microbial biodiversity of terrestrial habitats. One aspect of the current project attempsto establish the diversity and ecology of freshwater Actinobacteria. This group of gram positivemicrobes is of considerable biotechnological interest, and many actinobacterial lineages ofterrestrial origin are known (5). Some Actinobacteria have recently been discovered to play aprominent role in freshwater microbial communities, too (1), and moreover provide an interestingmodel for food-web controlled microbial survival in habitats exposed to high and selectivemortality (4). Other facets of the project deal with the establishment of high-throughput methodsfor the quantification of different microbial populations in water samples, and with laboratorymodels that investigate the competitive performance of individual microbial populations exposedto different substrate gradients (3).
Results:During the first year, one focus of the project was the development and refinement of appropriatemethods for the culture-independent characterization of microbial diversity within one definedphylogenetic group. A PCR-based protocol for the rapid screening of 16S rDNA clone librariesfrom freshwater systems for comparatively rare actinobacterial sequences (typically 1-10% of allobtained sequences) was developed. So far, 20 almost complete sequences from this lineage couldbe obtained from several hundred screened clones, which more than doubles the availabledatabase for freshwater Actinobacteria. After the successful establishment of the screening assaywe now progress to produce at least 50 different actinobacterial sequences from 4-5 aquatichabitats that greatly differ in their temporal stability, and to search for eventual biogeographicalpatterns. In addition, this collection will serve as a base for the design of specific oligonucleotideprobe for fluorescence in situ hybridization (FISH), so that the population sizes of differentactinobacterial lineages can be followed in selected habitats.
FISH probes are an increasingly popular tool to visualize different bacteria in variousenvironments. So far, the quantification of FISH-stained cells is tedious and time-consuming, andthe relative number of different bacteria in a sample have to be established by epifluorescencemicroscopy and manual counting. This limits the application of the approach to relatively simpleecological questions, and e.g., discourages more comprehensive studies of microbialbiogeographical distribution and population dynamics. We are, therefore developing techniquesfor the automated counting of double-stained bacteria in environmental samples by computer-assisted image analysis. In the first year of the project we have established an image acquisitionand evaluation system for the automated microscopic evaluation of comparatively simplesamples, i.e. the counting of bacterial cells in suspension that are filtered and immobilized onmembrane filters. For this we have developed a robust computerized focussing system that

85
combines a propriatory autofocus with Haralick texture parameters (2), feedback loops for imagecontrast maximation, and decisions on image content during acquisition. The system is thuscapable of excluding inappropriate or badly focussed microscopic fields prior to evaluationwithout operator interaction with high reliability, and to evaluate >50 FISH stained samples perday at a total of 2000-3000 counted cells per sample. Future improvements are aimed at betterdiscrimination of bacteria and non-bacterial objects in complex (e.g., sediment) samples by meansof advanced shape recognition and/or colour.
A third line of research, although originally not explicitely focussed on freshwatersystems, is conceptionally nevertheless closely related to one of the basic ecological questions tobe adressed in the project. We tested the hypothesis that aquatic bacteria found in different nichesin the natural habitat (particle-attached and planktonic, respectively), might in principle also differin their adaptations to substrate gradients. Two bacterial isolates from a previously establishedculture collection were exposed to different modes of substrate addition in batch cocultures.During cocultivation on a batch of substrates the final cell densities of the free-living bacterialstrain were reduced three times as much as of the attached ecotype, as compared to growth yieldsin pure cultures. In contrast, the gradual addition of substrates to stationary phase cocultures wasclearly disadvantageous for the population of the typically particle-attached bacteria (3). Adifferent growth response to substrate gradients could thus be another facet affecting thecompetition between bacteria at the aquatic-terrestrial interface.
References:1. Glöckner, F.-O., E. Zaichikov, N. Belkova, L. Denissova, J. Pernthaler, A. Pernthaler, and R.
Amann, Comparative 16S rRNA analysis of lake bacterioplankton reveals globally distributedphylogenetic clusters including an abundant group of Actinobacteria , Appl. Environ. Microbiol.,2000, 66, 5053-5065.
2. Haralick, R. M., Shanmuga.K, and I. Dinstein, Textural features for image classification, IEEETrans. Syst. Man Cybern., 1973, 3, 610-621.
3. Pernthaler, A., J. Pernthaler, H. Eilers, and R. Amann, Growth patterns of two marine isolates:Adaptations to substrate patchiness?, Appl. Environ. Microbiol., 2001, 67, 4077-4083.
4. Pernthaler, J., T. Posch, K. Simek, J. Vrba, A. Pernthaler, F. O. Glockner, U. Nübel, R. Psenner,and R. Amann, Predator-specific enrichment of actinobacteria from a cosmopolitan freshwaterclade in mixed continuous culture, Appl. Environ. Microbiol., 2001, 67, 2145-2155.
5. Rheims, H., A. Felske, S. Seufert, and E. Stackebrandt, Molecular monitoring of an unculturedgroup of the class Actinobacteria in two terrestrial environments, J. Microbiol. Methods, 1999, 36,65-75.

86
Project ID: 01LC0021 (Subproject 5)1.1.2000 - 31.12.2003
INVESTIGATION OF PROKARYOTIC DIVERSITY BY OPTIMIZEDCULTIVATION METHODS
Frederic Gich, Herbert Hoffelner and Jörg Overmann
Bereich Genetik und Mikrobiologie, Department I, Fakultät für Biologie, Ludwig-Maximilians-Universität München, Maria-Ward-Str. 1a, D-80638 Mü[email protected]
Key words: Not-yet-cultured bacteria, TGGE, dialysis culture, molecular fingerprints,cultivation, dialysis culture, chemotaxis
Abstract:The biodiversity of selected groups of prokaryotes is investigated by a combination ofcultivation methods with culture-independent molecular techniques. The target groups arechosen based on their numerical dominance in the natural environment. The present projectfocuses on those prokaryotes which are not-yet-cultured, but have been shown to occur inlarge populations in the environment, especially in soil. For a first inventory of the importantgroups of prokaryotes, group-specific PCR protocols are established and combined withtemperature gradient gel electrophoresis (TGGE) to yield typical molecular fingerprints. Asuite of novel cultivation methods is employed for the enrichment, isolation and subcultivationof prokaryotes. These methods comprise the formulation of novel mineral media, chemotacticenrichments, growth on inert solid substrates, treatment with specific bacterial pheromones,and dialysis cultures. Cultivation success is monitored by most probable numbers and bymolecular fingerprints to establish the most promising experimental strategies for theisolation of certain dominant prokaryotes. By its combination of molecular biologicalmethods with cultivation techniques, the present project offers the perspectivetof ultimatelyresult in the isolation and characterization of ecologically significant prokaryotic isolates andto understand their impact on the performance of the soil ecosystem. We expect that some ofthe isolates obtained can also be employed as indicators for certain types of ecosystems.
Results:Culture independent methods have repeatedly demonstrated that the diversity of prokaryotes inthe natural environment is unexpectedly high. This is especially true for soil, where the number ofbacterial species is estimated to range between 13,000 and 26,000 species [1,2]. By comparison,only 4500 species of prokaryotes so far have been validly described [3]. The diversity in aquatichabitats appears to be lower (~550 species, [4]). So far, only a minor fraction of the diverse andecologically significant assemblages of prokaryotes can be isolated in pure culture (i.e. 0.3% ofthe total cell counts [5]) and is thus available for physiological studies. Traditional cultivationtechniques have been shown to select for bacteria which appear to occur in minor numbers only[6]. The isolation and characterization of ecologically significant bacteria is thus a prerequisite foran understanding of microbial diversity and the function of microbial assemblages in naturalenvironments.In the present project eight taxonomic groups of prokaryotes were chosen based on theiroccurrence in the natural environment: the Acidobacteria, Actinobacteria, Low GC gram-positivebacteria, α-Proteobacteria, β-Proteobacteria, Cytophaga-Flexibacter-Bacteroides Group,nonthermophilic Crenarchaeota, and the Green Nonsulfur Bacteria. Selective PCR protocols forthe amplification of 600 bp-long 16S rRNA gene fragments were established. In combinationwith Temperature Gradient Gel Electrophoresis (TGGE), our technique permits a culture-

87
independent and rapid monitoring of the composition of natural communities. Since the DNAbands generated are available for sequencing, the dominant prokaryotes of each group can also beidentified taxonomically in a cultivation-independent manner.
Fig. 1. 16S rDNA fingerprints of the group Green Nonsulfur Bacteria in various environments. Different numberscorrespond to different bacteria. All 16S rDNA sequences obtained were previously unknown (after [7]).
Several novel cultivation methods are employed for the enrichment, isolation and subcultivationof prokaryotes especially from soil. Three different soils (fir forest, beech forest, alpine soil at1400 altitude) located south of Munich were chosen as sampling sites. Our methods comprise theformulation of novel mineral media, chemotactic enrichments, growth on inert solid substrates,treatment with specific bacterial pheromones, and dialysis cultures. Cultivation success ismonitored by the most probable number (MPN). We divised a new MICRODROP technique forthe automated inoculation of defined numbers of single bacterial cells in liquid media in microtiterplates. To date, a total of 200 bacterial strains could be isolated in liquid cultures with thisMICRODROP technique. Employing the group-specific fingerprinting methods described above,the strains obtained are identified and compared to the original composition of the complexprokaryotic community. By the combination of cultivation attempts with culture-independentmethods, the most promising experimental strategies for the isolation of certain dominantprokaryotes will finally be established.
References:1. Atlas RM, Horowitz A, Krichevsky M, Bej AK (1991) Microb Ecol 22, 249-2562. Torsvik V, Sørheim R, Goksør J (1996) J Industrial Microbiol 17, 170-1783. DSMZ, Deutsche Sammlung von Mikroorganismen und Zellkulturen (2000)
http://www.dsmz.de/bactnom/bactname.htm4. Jaspers E, Nauhaus K, Cypionka H, Overmann J (2001) FEMS Microbiol Ecol 36,153-1645. Amann RI, Ludwig W, Schleifer K-H (1995) Microbiol Rev 59, 143-1696. Dunbar J, Takala S, Barns SM, Davis JA, Kuske CR (1999) Appl Environ Microbiol 65, 1662-16697. Gich F, Garcia-Gil J, Overmann J (2001) Arch Microbiol (in press)
1
2
345
6 6 6 6
7
8 8 8 8 8 8 8
910
11
1213
14
15 1617
187
10
19

88
Project ID: 01LC0021 (Subproject 6)1.1.2001 – 31.12.2001
THE ROLE OF N-ACYL-L-HOMOSERINE LACTONE (AHL) MEDIATEDCOMMUNICATION FOR THE STRUCTURE AND PERSISTENCE OFMICROBIAL RHIZOSPHERE COMMUNITIES
A. Steidle, K. Sigl, K. Riedel, and L. Eberl
Lehrstuhl für Mikrobiologie, Technische Universität München, Am Hochanger 4, 85350 [email protected]; http//www.quorum-sensing.de
Key words: Quorum-sensing, rhizosphere, cell-cell communication, Pseudomonas putida,Serratia sp.
Abstract:The plant rhizosphere represents a highly complex ecosystem that is influenced by a numberof abiotic (soil chemistry, atmos- and hydrosphere, climate, etc.) and biotic factors (plantspecies, number and diversity of rhizosphere organisms). The bacterial communities of therhizosphere have strong influence on the growth and health of the plant as well as on theirability to adopt to changed environmental conditions. In recent years it has become evidentthat a number of bacteria associated with plants produce N-acyl-L-homoserine lactone (AHL)signal molecules that act as chemical communication signals between bacterial cells as wellas between bacteria and plants. These interactions appear to be of great importance for theecology of the rhizosphere. The aim of the proposed project is to investigate the role of AHL-mediated intergeneric communication for the diversity and the stability/dynamic of therhizosphere ecosystem. These investigations should also allow to monitor changes of theenvironment by assessing the fluctuations in the communication and population structure ofthe rhizosphere consortium.
Results:We have developed and characterized novel Gfp-based monitor strains that allow in situvisualization of AHL-mediated communication between individual cells in the plantrhizosphere. For this purpose three Gfp-based AHL sensor plasmids (1) were transferred intoAHL-negative derivatives of Pseudomonas putida IsoF and Serratia liquefaciens MG1, twostrains that are good colonizers of tomato roots. Each of these sensor plasmids is capable ofdetecting a different range of AHL molecules, depending on the components used for itsconstruction. The resulting strains were rigorously characterized with respect to theirsensitivities for various AHL molecules and then employed to visualize AHL-basedcommunication between defined bacterial populations in the rhizosphere of axenically growntomato plants.We screened over 300 bacterial strains isolated from the rhizosphere of tomato plants for theirability to synthesize AHL signal molecules. Approximately 12% provoked a positive signalwith at least one of the AHL monitor strains used. We selected four isolates for further studiesand these were, on the basis of their 16S rDNA, affiliated to the genera Pseudomonas andRahnella. The AHL profiles (i.e. the amounts and structure of the AHL molecules) of theseisolates were determined by thin-layer chromatography in combination with the AHLbiosensor pSB403 (Fig. 1) (4, 2).

89
To investigate whether these strains do produce AHL molecules when colonizing the tomatorhizosphere we inoculated axenically cultured tomato plants with an AHL monitor straintogether with an AHL-producing rhizosphere isolate. When the tomato roots were inspectedunder a confocal laser scanning microscope, green fluorescent cells could be observedindicating the presence of AHL molecules in the plant rhizosphere. Expectedly, no signalswere detected when the sensor strains were inoculated alone. In conclusion, these experimentsprovide strong evidence that AHLs are produced by bacteria colonizing the rhizosphere ofaxenically grown tomato plants and that these signals can be perceived by other bacteria (3).
Furthermore, we integrated an AHL sensor cassette, that responds to the presence of longchain AHLs with the expression of Gfp, into the chromosome of the DsRed-tagged P. putidastrain F117. This monitor strain was used to demonstrate that the indigenous bacterialcommunity colonizing the roots of tomato plants growing in non-sterile soil produces AHLmolecules. In a first step this monitor strain was used to inoculate tomato plants grown in non-sterile soil together with the untagged wild type strain IsoF. As for the axenically grownplants we observed that most red fluorescent cells were also green fluorescent indicating thatIsoF also produces AHL in the natural root habitat. However, in contrast to the gnotobioticsystem, we also detected green fluorescent cells when the monitor strain was inoculated alone.These results clearly demonstrate that the indigenous rhizosphere community produces AHLsat concentrations high enough to activate the monitor strain and thus provides strong evidencethat these signal molecules are utilized by the consortium for communication betweendifferent populations.
References:1. Andersen, J. B., A. Heydorn, M. Hentzer, L. Eberl, O. Geisenberger, B. B. Christensen, S. Molin, and
M. Givskov. 2001. Appl. Environ. Microbiol. 67:575-585.2. Geisenberger, O., M. Givskov, K. Riedel, N. Høiby, B. Tümmler, and L. Eberl. 2000. FEMS
Microbiol. Lett. 184:273-278.3. Steidle, A., K. Sigl, R. Schuhegger, A. Ihring, M. Schmid, S. Gantner, M. Stoffels, K. Riedel, M.
Givskov, A. Hartmann, C. Langebartels, and L. Eberl. 2001. Appl. Environ. Microbiol. 67. In press.4. Winson, M. K., S. Swift, L. Fish, J. P. Throup, F. Jørgensen, S. R. Chhabra, B. W. Bycroft, P.
Williams, and G. S. A. B. Stewart. 1998. FEMS Microbiol. Lett. 163:185-192.
1 2 3 4 5 6 7
Fig. 1. Thin layer chromatographic analysisof AHLs produced by bacterial strainsisolated from the tomato rhizosphere. AHLsextracted from cell-free culture supernatantsof Pseudomonas putida IsoF (4), P. putidaZ2D (5), Rahnella aquatilis TAA (6), and R.aquatilis T13 (7) were separated by TLC, andspots were detected with the aid of the AHLbiosensor E. coli MT102(pSB403). SyntheticAHLs were included as reference substancesas follows: 3-oxo-C6-HSL (1), 3-oxo-C12-HSL (2), and 3-oxo-C10-HSL (3).

90
Project ID: 01LC0021 (Subproject 7)1.1.2000-31.12.2003
BIODIVERSITY OF PROTOZOA IN THE RHIZOSPHERE OF PLANTS:EFFECTS ON ACTIVITY, PERFORMANCE AND DIVERSITY OFMICROORGANISMS AND ON PLANT GROWTH.
M. Bonkowski, K. Kreuzer and S. Scheu
Institut für Zoologie, Darmstadt University of Technology, Schnittspahnstrasse 3, D-64287Darmstadt, [email protected]; http://neuro.bio.tu-darmstadt.de/newww/zoology/scheu
Key words: manipulation of soil diversity, rhizosphere interactions, plant competition
Abstract:Competition between plants is high, because plant growth in natural environments frequentlytakes place in dense stands of established vegetation. Thus both, beneficial and deleteriousassociations of plants with microorganisms and the regulation of these associations by soilfaunal activity may be of fundamental importance for individual plant species, but also for thespecies composition of the whole community. We investigate the mechanisms and ecologicalsignificance of interactions between plants and microorganisms (bacteria, fungi), and theirfaunal regulation. We focus on three general aspects: (1) the regulation of plant growth bymicroorganisms and their consumers (protozoa, nematodes, collembola), (2) the competitionfor nutrients between microorganisms and plants, (3) the regulation of plant-plantcompetition in the rhizosphere.
Results:Hypotheses 1.3.1. Protozoa affect size, turnover, diversity and function of microbialcommunities in the rhizosphere and 1.3.4. Protozoa increase the resilience of the microbialcommunity.
We isolated species of different protozoan taxonomic (amoebae, flagellates, ciliates) andfunctional groups (bacterial, fungal feeders) from soil. An abundant soil amoebae(Acanthamoeba sp.) was selected as model-organism and in cooperation with Prof. M.Wagner, TU-München, TP2) could be genetically related to the A. castelanii group. Wetransferred this naked amoebae with specific soil bacteria in monoxenic cultures. The effectsof amoebal grazing on the diversity of rhizobacteria and root growth of experimental plantshave to be tested in subsequent experiments.
A first protoype of a new microbial respirometer has been constructed in our department inorder to measure microbial activity, biomass, turnover and functional attributes on amicroscale.
Investigations of microbial diversity with molecular approaches are currently in progress.
Hypothesis 1.3.2. Protozoa increase plant growth by changing the microbial community inthe rhizosphere, by increasing nutrient cycling and hormonal effects.
In cooperation with Prof. I.T. Baldwin (MPI for Chemical Ecology, Jena) and Dr. B.S.Griffiths (Scottish Crop Research Institute, U.K.) we are currently investigating theinteractions of protozoa and root-feeding nematodes in the rhizosphere of tobacco plants(Nicotiana attenuata). Root herbivores such as nematodes are known as crop pests and maystrongly affect plant diversity in the field. Current studies, focussing on effects of visible

91
above-ground herbivory, demonstrated that tobacco plants can discriminate attacks fromdifferent above-ground herbivore species. About 500 plant genes are involved in the'recognition' and specific defense strategies against different herbivores. However, themechanism of plant response to below-ground herbivores is not yet known. In addition, theinteractions in the rhizosphere are complicated by species interactions. Protozoa are known toincrease root biomass due to hormonal effects from grazing on rhizosphere bacteria. Root-feeding nematodes commonly have the opposite effect, but leaking of root cell contents due tonematode feeding might fuel growth of rhizobacteria and stimulate subsequent effects ofprotozoan grazing. We aim to get deeper insight in the molecular regulation of root growthand plant defense to attack from root herbivores.
Hypothesis 1.3.3. Protozoa increase plant growth via interactions with mycorrhiza.An experiment in cooperation with Prof. G. Bécard (C.N.R.S., Toulouse, France), testing
effects of protozoa and AM-fungi on growth of Plantago lanceolata is currently inpreparation.
Hypothesis 1.3.5. Protozoa affect the competition between plants via species specific effectson microbial communities and root morphology.
In cooperation with Dr. J. Roy (C.N.R.S., Montpellier, France) we investigated theinfluence of microbial diversity on growth and competition among grasses in experimentalmicrocosms.
The performance of the six different grass species was highly dependent on soil diversity(F = 13.0, p = 0.0001 for the interaction plant species x soil diversity). A subsequentregression of the average shoot biomass per plant species in a sequence of increasing soildiversity (sterile < microorganisms < microfauna < mesofauna) was taken as a measure ofplant performance in relation to soil diversity.
The homogeneity of slopes (ANCOVA), gave significant differences for three out of thesix investigated plant species. A significant negative slope (F = 4.4, p = 0.04) for Bromushordeaceus indicated a decrease in shoot growth with increasing soil diversity inmonocultures. Bromus species turned out to be weak competitors in mixtures compared tomonocultures. Particularly B. hordeaceus gained the lowest shoot biomass in competitionwith other plant species in the mixture treatment. However, with increasing soil diversityshoot biomass of B. hordeaceus increased in plant mixtures (F = 4.8, p = 0.04). This was dueto a release of Bromus from competition with Aegilops geniculata (F = 5.3, p = 0.03) and B.madritensis (F = 15.8, p < 0.001) which both were negatively affected by increasingbiodiversity. The two strongest competitors in plant mixtures, Hordeum murinum and Loliumrigidum did not respond to any soil diversity treatment.
In conclusion, soil biodiversity had strong effects on the performance of certain grasseswhen different plant species were in competition. Apparently, slight modifications in thefitness of a grass by soil organisms multiplied to strong effects when the plant had to competewith other grass species. Since competition among plants is the natural situation in the field,our experiment indicates that effects of soil biodiversity on grassland community structurehave so far been underestimated.
Hypothesis 1.3.6. Protozoan effects on plant growth depend on the diversity of the faunalcommunity in the rhizosphere.
This hypothesis is tested by including other faunal groups in experiments with soil protozoa(see examples in hypotheses 1.3.2. and 1.3.5.). Our preliminary results indicate that speciesinteractions in the soil food-web are of major importance for understanding faunal effects onrhizosphere microorganisms and plant growth.

92
Project ID: 01LC0101 (Subproject 1)01.04.2001-30.09.2003
AN ANALYSIS OF PREVAILING ECONOMIC AND POLITICAL STRATEGIESFOR THE PROTECTION OF BIODIVERSITY - THE TRADE IN GENETICRESSOURCES: A VIABLE CONTRIBUTION TO THE PROTECTION OFBIODIVERSITY?
S.L. Franck-Oberaspach1, O. Deke2, G. Klepper2* und P.T. Stoll1,3**
1Max-Planck-Institute (MPI) for Foreign Public Law and International Law, D-69120 Heidelberg,Im Neuenheimer Feld 535, e-mail: [email protected];2Institute for World Economics (IfW), D-24100 Kiel, Düsternbrooker Weg 120; e-mail:[email protected], [email protected];3New adress: Institut für Völkerrecht der Universität Göttingen, D-37073, Platz der GöttingerSieben 5, e-mail: [email protected]; *Project leader IfW Kiel; **Project leader MPI Heidelberg
Keywords: biological diversity; genetic resources; public good; property rights; access andbenefit sharing, value, trade
Abstract:It is the purpose of this sub-project to assess the potential to contribute to the conservation ofbiodiversity of the international trade in genetic resources (GR). Initial results show that thecontribution of the trade in genetic resources to the conservation of biological diversityprobably will be limited even without taking into account the concerns unsually cited here,e.g. the incomplete implementation of relevant international legislation or the possiblyinefficient use of monetary returns by providers of genetic resources.
Results:Genetic resources are often considered as one of the major contributions of biodiversity(Swanson 1996). In addition, it is often assumed that genetic resources are the majorprivatizeable good in the vector of outputs produced by biodiversity. We define, for thepurpose of our study, as genetic resources (GR) only biological material the geneticinformation of which is directly used by a potential recipient. The biological material itself(e.g. wood or a medical plant that is used as raw material, i.e. a pharmaceutical is directlyextracted) is not regarded as a genetic resource here since it has different economic properties:biological material is a rival, exclusive good. Genetic information, to the contrary, is a publicgood as soon as the biological material it is embedded in is publicly accessible. Exclusivepossession of a biological material implies the exclusive possession of the genetic informationcontained. Access to a genetic resource in this case can be provided via sale of the material, orvia a contract that entitles the providing party to a percentage of the income derived from thisgenetic information. Accordingly, legal experts discussed as a major problem theimplementation of legislation to regulate access and benefit sharing (e.g. Glowka 1997).Economists, on the other hand, tried to assess supply of and demand for genetic resources,including the problem of of transboundary habitats, that may cause ‚races to the bottom‘ ofmarket prices of genetic resources1. Recent economic research also focuses on the survey ofbilateral contracts in order to examine their design and economic efficiency (e.g. Rausser andSmall 2000).
1A related problem is the fact that biodiversity- rich areas amy be substituted, in part, by ex situ collections in a
partial consideration, i.e. if only the production/conservation of genetic resources is looked at.

93
We follow a more specific, theoretical approach in the first part of our analysis. We considerappropriabilities in the GR-related industries and markets, i.e. in all phases of GR-relatedresearch and development (R&D) processes and marketing of respective R&D results. Weexamine different institutional frameworks (legal rights of exclusion) and their potentialeffects on property rights as defined by Barzel (1989). Initial results gained in the area ofplant genetic resources for food and agriculture (PGRFA) show that weak property rightshave a major impact on the market value of genetic resources. These property rights, however,do not simply depend on an institutional framework (i.e. legal rights of exclusion) that can becreated and implemented at the discretion of the national or international lawmaker, since thephysical properties of the materials and R&D processes involved largely determine thepossibilities of effective legislation, mainly by influencing the enforcability of contractsand/or intellectual property rights. Inherent public good aspects, however, are likely to bemuch smaller in other industries.Further study will focus on the assessment of the gap between the social and private value ofgenetic resources, and on the potential of (international) legislation and implementation ofexisting legislation to close this gap.
References:
Barzel, Y. (1989): Economic analysis of property rights. Cambridge University Press.Glowka, L. (1997): Emerging legislative approaches to implement Article 15 of the Convention on
Biological Diversity. Review of European Community & International Environmental Law 6 (3): 249ff.
Rausser, G.C. And Small, A.A. (2000): Valuing research leads: bioprospecting and the conservation of genetic resources. Journal of Political Economy 108 (1), 173-206.
Swanson, T. (1996): The reliance of northern economies on southern biodiversity: biodiversity as information. Ecological Economics 17, 1-8.

94
Project ID: 01LC0101 (Subproject 2)01.04.2001-30.09.2003
AN ANALYSIS OF PREVAILING ECONOMIC AND POLITICAL STRATEGIESFOR THE PROTECTION OF BIODIVERSITY - THE PROTECTION OFBIODIVERSITY: NEW APPROACHES AND INSTRUMENTS IN INTERNATIONALLAW
S.L. Franck-Oberaspach1, O. Deke2, G. Klepper2* und P.T. Stoll1,3**
1Max-Planck-Institute (MPI) for Foreign Public Law and International Law, D-69120Heidelberg, Im Neuenheimer Feld 535, e-mail: [email protected];2Institute for World Economics (IfW), D-24100 Kiel, Düsternbrooker Weg 120; e-mail:[email protected], [email protected];3New adress: Institut für Völkerrecht der Universität Göttingen, D-37073, Platz der GöttingerSieben 5, e-mail: [email protected]; *Project leader IfW Kiel; **Project leader MPI Heidelberg
Keywords: biological diversity; international regimes; international law, economicincentives, transfer mechanisms; institutional coordination
Abstract:The protection of biodiversity is a relatively new objective for the international community. In1995, the prevailing international treaty in the field, the Convention on Biological Diversity(CBD), entered into force. Its efficient implementation, however, has yet to be achieved.Another unresolved task is the harmonization of this new objective with related goals (e.g.,the protection of species, landscapes, or cultural achievements of indigenous peoples) and theeffective adaptation of the existing system of international law towards this objective. The taskincludes, inter alia, innovative mechanisms with respect to financial transfers. Her, theexisting international regime aiming at the protection of biodiversity is analysed. Economicanalysis focuses on incentives provided by available instruments. Legal analysis focuses onthe identification and examination of existing treaties and regulations that often do notmention ‚biodiversity‘ but focus on elements thereof. The systematic connection of theseinternational treaties is analysed. Our final goal for this first part of the BIOLOG project isdirected towards the description of possibilities of efficient implementation and institutionalcoordination at the international level.
Results:The term ‚biodiversity‘ requires and promotes the joining together of regimes and instrumentsthat have originally been intended to promote the protection either of ‚parts‘ of biodiversity(e.g., single species) or of areas (e.g., certain landscapes) with existing regimes especiallydesigned to protect biodiversity (maily the Convention on Biological Diversity (CBD, enteredinto force in 1995), with bodies of the United Nations in charge of environmental problems(e.g., the United Nations Environmental Programme (UNEP) and the Commission onSustainable Development (CSD)), and with to-be-constructed new instruments.In the actual phase of the project, we concentrate on an analysis of existing internationalregimes and instruments. We identified a considerable amount of international regimes andinstruments the objective of which is the protection of biodiversity-related elements. Theorigin of most of these treaties is pre-CBD; they do not usually refer to the ‚biodiversity‘directly. Most of these regimes do not have mechanisms for international financial transfer.Reseach with respect to the legal aspects of these instruments has largely focused on problemsrelated to the collision of norms between different regimes (e.g. Raustiala, 1997). Aspects of

95
institutional coordination, however, only rarly have been examined in-depth. Economicresearch, on the other hand, has analysed incentives provided by these regimes, and incentivesof differents regimes have been compared to each other (e.g. Moyle 1998). Results of theseeconomic approaches led to the proposal of potentially more effective new instruments.Aspects of compatibility of these instruments with the existing system of internationalregimes, however, have been largely neglected by most of these approaches.We identified as one major goal, at the theoretical/analytical level as well as with respect tothe actual implementation of biodiversity-related regimes, the systematic joining together ofexisting treaties, instruments, and regulations that have potential to contribute to theprotection of biodiversity-rich areas with available mechanisms of financial transfer(s).Another major objective, again at the theoretical/analytical level and with respect to the actualimplementation, is the analysis and development of possibilities to impose sanctions oncountries that do not comply with the objective of the conservation of biodiversity. Bothaspects involve very complex problems already at the level of international law andgovernance. Our preliminary conclusion is that there are serious problems to be solved at theinternational level, and that the constant degradation and the progressive loss of biodiversityis not a problem of implementation at the national level alone.
References:
Rosell, M. (1997): The sovereignty challenge. In: Review of European Community & International Environmental Law, 6 (3), 274-283.
Moyle, B. (1998): Species Conservation and the principal agent problem. Ecological Economics 26: 313-320.

96

97
BIOLOG AFRICA

98
Project ID: 01LC0024 (BIOTA Southern Africa)01.10.2000 – 30.09.2003
A RESEARCH PROJECT MONITORING AND ANALYSING BIODIVERSITY ANDITS CHANGE ALONG A TRANSECT IN NAMIBIA AND SOUTH AFRICA
BIOTA Southern Africa is located in Namibia and the Republic of South Africa. BIOTASouthern Africa has an integrated interdisciplinary research approach on biodiversity inAfrica that includes the following disciplines: animal production, botany, climatology,modelling, mycology, remote sensing, socio-economics, soil science, and zoology.
The project is active along an approximately 2000 km long transect that extends from theMediterranean Fynbos Biome at the Cape, the semi-arid Mediterranean Succulent KarooBiome, the arid Desert Biome, the semi-arid tropical Nama Karoo Biome up to the tropicalsemi-arid Savanna Biome. Based on this transect approach, the target area of BIOTASouthern Africa covers the entire area parallel to the main rainfall gradient, from thetemperate Cape with its winter rainfall climate up to the semi-humid Kavango region ofnorth-eastern Namibia.
Within the three BIOTA AFRICA regional projects, BIOTA Southern Africa examines themost arid regions. Hereby, the project activities are located at either margin of the aridsubtropical high pressure cell (wider Karoo-Namib region).
Scientific goals of BIOTA Southern Africa
The need to develop new strategies to deal with the dramatic environmental changes in Africa is afundamental challenge to the scientific community. The protection and management ofbiodiversity plays a key role in such strategies. The overall goal of BIOTA Southern Africa is togain knowledge for decision-makers for a feasible and sustainable management of biodiversity.Within the framework of long-term interdisciplinary fieldwork, applicable land use systems areanalysed in combination with scientifically based conservation concepts for the resourcebiodiversity. Socio-economic information is fundamental for the comprehension and themanagement of the systems.
In order to understand the bio-physical and socio-economic drivers affecting biodiversity, somerelevant questions are:
• To what extent are environmental factors responsible for biodiversity?• How important are abiotic and biotic soil compartments and mechanisms?• How important are slow processes and preceding environmental and evolutionary history?• What are the mechanisms by which land use practices control biodiversity and vice versa?• How important are spatial movements of taxa and environmental factors?• Are future changes of biodiversity predictable?• What is the social, economic, and political importance of biodiversity and how can it be
valued?
In contrast to dense rainforests, relatively open savanna, shrubland, and semi-desertvegetation allow a more convenient long-term monitoring. In these semi-arid vegetationtypes, BIOTA Southern Africa has established a standardised monitoring approach thatconsiders the following aspects:
• Based on remote sensing and GIS, a large-scale approach monitors changes to vegetationstructures, to soil characteristics, and to biodiversity within a specified transect band.
• More than 30 Biodiversity Observatories have been established along the transect band.These Observatories represent a large number of major ecosystems and vegetation units.

99
• Often, these Biodiversity Observatories are arranged in pairs, in identical ecosystems, butexposed to different land use practices. This allows the identification of managementpractices and land use intensities and their impact on natural resources.
Biodiversity Observatories ensure the comparability of research sites within the BIOTASouthern Africa project. The standardisation of scales and applied methods, as well as theintegration of different disciplines in a defined research area are the major advantages of thisresearch approach.
Priorities of the first months
Directly after the start in October 2000, and after the formation of the working groups,BIOTA Southern Africa principally focused on the following important issues:
• Strengthening links with the partners and research institutions in Namibia and theRepublic of South Africa (MoUs, contracts)
• Strengthening links with landowners and local communities (MoUs, contracts)• Preparation and carrying out of a first Kick-Off-Trip with all project heads• Organisation / establishment of the project´s infrastructure (field equipment, cars, weather
stations, logistic and research stations, surveyors, permits)
Practical steps of the first months
As a result of these activities, all practical preparations for the field work had been completedby April 2001, and can be summarised as follows:
• Several workshops were organised by our Head office in Hamburg/Germany. Threeworkshops took place during the Kick-Off-Trip in March and April 2001 in the hostcountries (two in Cape Town, one in Windhoek). The detailed and multidisciplinarypresentation of BIOTA Southern Africa´s intentions and methodology were discussedwith the scientific community of Namibia and the Republic of South Africa. The projectinitiative was welcomed, and in some cases an even stronger integration into the nationalscientific communities was requested. Based on the motivating spirit of co-operation,clear agreements on the future co-operation were defined. The formation of the “SouthAfrica BIOTA Liaison Committee” (SABLC) will guarantee transparent discussions andsupport well-founded decisions on the future and joint development of the BIOTAAFRICA project in South Africa.
• A number of more technical matters had to be settled: six cars, technical field equipment,and weather stations were bought. The issues of registration, insurance, and safe storagewere solved.
• 30 Biodiversity Observatories and a very large number of additional sampling sites havealready been established in co-operation with national and local institutions andlandowners. For an easy orientation in the field, the Observatories have been subdividedinto 100 hectares, and numbered. Weather stations were installed and have started withdata recording. For all sites, monitoring protocols and general rules were defined, underwhich field work in the Biodiversity Observatories is to be carried out.
• For a large proportion of Observatories, first inventories of organisms have beendocumented. This baseline information can be used by all scientists who work inaccordance with the scientific needs as defined by the Namibian and South Africanscientific community and responsible institutions.
• At several conferences, preliminary field results have been presented to a broad scientificcommunity, and discussion papers are in the making.

100
Project ID: 01LC0024 (BIOTA Southern Africa S01)01.10.2000 – 30.09.2003
REMOTE SENSING AND GIS BASED SURVEY OF SPATIAL AND TEMPORALBIODIVERSITY DYNAMICS AND ANALYSIS OF BIODIVERSITY ANDGEODIVERSITY INTERRELATIONSHIPS
B. Hörsch, M. Vogel, C. Schultz and G. Braun
Deutsches Zentrum für Luft- und Raumfahrt DLR, DFD-UG, Linder Höhe, 51147 Kö[email protected] - http://www.dfd.dlr.de
Key words: Remote Sensing, GIS, Biodiversity-Geodiversity, Monitoring, VegetationMapping, Capacity Building
Abstract:This project shall create the basis for a system in which both, remote sensing and GISapplications, form an analysis and monitoring system for the assessment of human impacts onbiological diversity. It is our aim to make the existent biodiversity more understandable andpredictable, taking into account the UN-Convention on Biological Diversity (UNCBD).It is planned to establish a coherent GIS along the entire BIOTA transect in order to allow theanalysis of spatial patterns of biodiversity, its temporal changes and the mutual dependencyof geodiversity and biological diversity. Focus will lie on the comparison of naturalecosystems with those where socio-economic activity has led to various degradation levels ofthe former natural ecosystems. Furthermore, the detection of indications of spatial patterns ofdiversity by remote sensing techniques will contribute to future monitoring and conservationconcepts.Further tasks are the “capacity building” for co-operating partners within the fields ofRemote Sensing and GIS.
Results:Within the last 14 months the data base of a Remote Sensing based GIS for the monitoring ofpresent-state biodiversity within the Southern African transect was built up (1). In closeinteraction with the project partners the different localities of field work were identified during afield trip in March and respective observatories were geographically referenced.Tasks performed included first of all the investigation of appropriate multitemporal data sets fromcommercial multispectral remote sensing systems (LANDSAT-7-ETM, IKONOS, CORONAetc.). A set of LANDSAT-7-ETM scenes was acquired, covering the identified biodiversityobservatories along the transect. All satellite scenes ordered were georectified to the commonUTM system and were integrated as image base into the GIS. Additionally, aerial photogrphs ofthe Namibian observatories were georectified and added to the data base.Besides Digital Chart of the World (DCW)-data basic GIS data (topography, infrastructure) wereintegrated into the spatial database at a scale of 1: 50.000. For Namibia, the integration of farmboundaries, settlements, river and road networks has been completed, whereas Southern Africandata based on topographic maps at 1: 50.000 are partly still being processed. These primaryinformations were further used to produce geometrically referenced image maps which areessential as an auxiliary means for the orientation of the fieldworkers. They also act as a base formapping training samples which form the basis in the supervised classification of vegetationpatterns from the multispectral images (2, 3). The maps were enhanced by topologicalinformation (roads, settlements, farm boundaries) and were superimposed by a fine resolutionlatitude/longitude grid in order to facilitate orientation in the field and to enable the directcomparison of GPS recordings with mapped features. The maps were distributed to the projectpartners in paper printed form. On request, digital maps were also supplied.

101
An important feature of a spatial database supporting biodiversity analysis in a more indirect wayare topographical data (4, 5). They form not only the basis for the absolute geographicalorientation of features in space, but are essential to derive topoclimatological information likepotential water and solar irradiance distributions in the terrain (6, 7). Due to the lack of fine scaletopographical maps, interferometric analysis of European Remote Sensing Satellite (ERS)-tandemdata has been performed to derive Digital Terrain Models from the research areas. Respecive datafor Namibia have almost entirely been processed. For Southern Africa, topographic information toproduce Digital Terrain Models is being purchased at the moment, as appropriate ERS-data arelacking. Due to the importance of spatial climatic data for biodiversity modelling, research hadbeen carried out on downscaling spatially coarse climatic datasets in order to derive climaticinformation on a spatial scale which is compatible to the scale of the research approach (8, 9).. Upto now, long term (1960-1990) precipitation and temperature data have been downscaled to aspatial resolution of 5km and 2 km, respectively, as long-term monthly means. In the near future itis intended to add additional climatic parameters and to cover the area with non-aggregatedmonthly mean data.Resulting from various field campaigns by the project partners of subprojects S05 and S06vegetation training samples have been integrated into the spatial database for study areas at theRichtersveld/Fish River Canyon, the Knersvlakte and the Wlotzka´s Baken area. Based on thesesamples vegetation maps have been derived from respective LANDSAT-7-ETM scenes.Validation of these maps is in progress.To support the use of spatial data and information systems by the project partners two capacitybuilding activities took place at the DLR in Cologne and at the Institute of Botany in Hamburg.Topics of the first course were primarily remote sensing basics with a special emphasis on sensorsensitivity, reflectance of different surfaces, and generation and interpretation of false colourcomposites. The second course held in Cologne focussed on Remote Sensing basics incombination with GIS architecture for biodiversity research and monitoring (11). Topics likegeneral data structure, the integration of referenced geospatial together with Remote Sensing dataand the main steps for building up of an information system were dealt with. Integrationpossibilities of raster and vector based spatial data with ecological/biological attributes stored indatabases were addressed partially.
References:1. Dale, V.H., Offerman, H. et al. (1995): Landscape characterization and biodiversity research. In:
Boyle, T.J.B. & Boontawee, B.: Measuring and Monitoring Biodiversity in Tropical andTemperate Forests; Proceedings of a IUFRO Symposium held at Chiang Mai, Thailand August27. - September 2. 1994, Centre for International Forestry Research: Bogor, Indonesia.
2. Allner, T., Backhaus, R. et al. (1992): Biodiversity Indication by Earth Observation Data. In:Proc. Of the Central Symp. of the Int. Space Year Conference, Munich, Germany. 30 March- 4April 1992, 863-867.
3. Podolsky, R. H. (1990): Monitoring Biodiversity and Landscape Richness with digital EarthImagery. In: Archives of Photogrammetry and Remote Sensing Vol. 28, Part 7-1, Proceedings ofthe Symp. On Global and Encironmetal Monitoring.
4. Burnett, M. R., August, P. V. et al. (1998): The influence of Geomorphological Heterogeneity onBiodiversity. A Patch-Scale Perspective. In: Conservation Biology, 12, 363-370.
5. Nichols, W. F., Killingbeck, K. T. et al. (1998): The Influence of GeomorphologicalHeterogeneity on Biodiversity. A Landscape Perspective. In: Conservation Biology, 12, 371-377.
6. Antonic, O., Marki, A. et al. (2000): A Global Model for Monthly Mean Hourly Direct SolarRadiation. In: Ecological Modelling, 129, 113-118.
7. Florinsky, I. V., Kulagina, T. B. et al. (1994): Influence of Topography on Landscape RadiationTemperature Distribution. In: Int. Journal of Remote Sensing, 15, 3147-3153.
8. Franklin, J. (1998): Predicting the Distribution of Shrub Species in Southern California fromClimate and Terrain-derived Variables. Journal of Vegetation Science, 9, 733-748.
9. Aspinall, R. (1992). Spatial analysis of wildlife distribution and habitat in a GIS. In: Proc. 5thInternational Symposium on Spatial Data Handling.

102
Project ID: 01LC0024 (BIOTA Southern Africa S02)01.10.2000 – 30.09.2003
EDAPHICAL DIVERSITY AND BIODIVERSITY IN MUTUAL DEPENDENCY
A. Gröngröft, A. Petersen and G. Miehlich
Institut für Bodenkunde, Universität Hamburg, Allende-Platz 2, 20146 [email protected]
Key words: soil variability, pH value, Namibia
Abstract:Soil as one major compartment of terrestrial ecosystems has a significant impact on thecomposition of terrestrial flora and fauna and vice versa. The soil science BIOTA projectaims to analyse the mutual dependency of the soil as a controlling factor of biodiversity alongthe entire BIOTA transect following a continental climatic gradient of high geodiversity. Thedocumentation of the soil diversity, especially with respect to the interdependency of soil,flora and fauna, requires a standardised and applicable methodology which proofs to bepowerful also in the analysis and interpretation of land use practises and managementstrategies. This includes the selection of useful indicators to document the influence of soilparameters on the biodiversity.
General Aims:The main focus of the Soil Science BIOTA project lies on the inter-disciplinary approach toanalyse the structures, reasons, processes, and impacts of biodiversity along a gradient-orientated transect. The hypothesis that a higher variability in abiotic parameters in a givenarea will necessarily lead to a higher biodiversity is to be proofed. Considering that abioticparameters are of varying importance to flora and fauna depending on the biotic group to beanalysed, this project does not only focus on a standardised documentation of soil diversity,but also considers the problem of scale. Different methodological scales have to be appliedwhen analysing (a) changes in soil parameters according to changes in vegetation units and/oranalysing the effects that single bushes or trees have (b) on animals (shading effect) or (c) onplants (safe sites for germination). Important questions to be answered include whether or nota systematic nutrient or salt accumulation takes place, which effects small burrowingmammals create on soil properties and wether soil crusts by cyanobacterias or lichens have animpact on the nutrient cycle of the soil and thus again on the biotic diversity. Furthermore,the impact of different land use practises on the geodiversity will be analysed. As one majorabiotic part of terrestrial ecosystems the soil and thus this project forms an importantexplanatory and functional link between the various biotic aspects (fauna, vegetation, lichens,crusts, fungi) within the BIOTA project.
Results:In this first phase main focus lies on the testing of soil spatial variability in the Biotaobservatories. For the selection of the sampling sites within the observatory a stratifiedrandom procedure was developed, in which the observatories are stratified in different habitattypes, according to topography, vegetation patterns etc.. Each hectare square is assigned to ahabitat type.
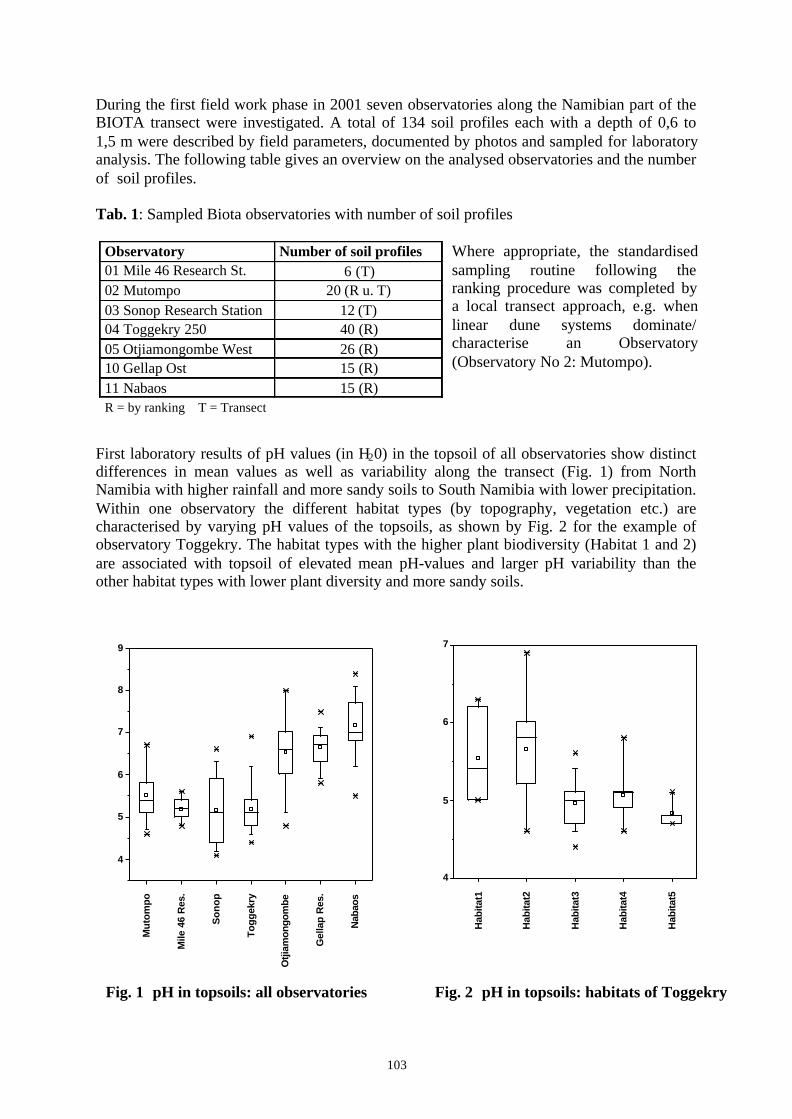
103
During the first field work phase in 2001 seven observatories along the Namibian part of theBIOTA transect were investigated. A total of 134 soil profiles each with a depth of 0,6 to1,5 m were described by field parameters, documented by photos and sampled for laboratoryanalysis. The following table gives an overview on the analysed observatories and the numberof soil profiles.
Tab. 1: Sampled Biota observatories with number of soil profiles
Where appropriate, the standardisedsampling routine following theranking procedure was completed bya local transect approach, e.g. whenlinear dune systems dominate/characterise an Observatory(Observatory No 2: Mutompo).
First laboratory results of pH values (in H20) in the topsoil of all observatories show distinctdifferences in mean values as well as variability along the transect (Fig. 1) from NorthNamibia with higher rainfall and more sandy soils to South Namibia with lower precipitation.Within one observatory the different habitat types (by topography, vegetation etc.) arecharacterised by varying pH values of the topsoils, as shown by Fig. 2 for the example ofobservatory Toggekry. The habitat types with the higher plant biodiversity (Habitat 1 and 2)are associated with topsoil of elevated mean pH-values and larger pH variability than theother habitat types with lower plant diversity and more sandy soils.
Hab
itat1
Hab
itat2
Hab
itat3
Hab
itat4
Hab
itat5
4
5
6
7
Observatory Number of soil profiles01 Mile 46 Research St. 6 (T)02 Mutompo 20 (R u. T)03 Sonop Research Station 12 (T)04 Toggekry 250 40 (R)05 Otjiamongombe West 26 (R)10 Gellap Ost 15 (R)11 Nabaos 15 (R)R = by ranking T = Transect
Mut
ompo
Mile
46
Res
.
So
no
p
Togg
ekry
Otji
amon
gom
be
Gel
lap
Res
.
Nab
aos
4
5
6
7
8
9
Fig. 1 pH in topsoils: all observatories Fig. 2 pH in topsoils: habitats of Toggekry

104
Project ID: 01LC0024 (BIOTA Southern Africa S03)01.10.2000 – 30.09.2003
BIODIVERSITY OF SOUTHWESTERN AFRICAN FUNGI: INTERACTIONS WITHHIGHER PLANTS, FUNCTIONAL DIVERSITY AND DNA-TAXONOMY
R. Berndt1, C. Görke1, E. Langer2, M. Mennicken1, E. Uhlmann1 and F. Oberwinkler1
1 Spezielle Botanik/Mykologie, Botanisches Institut, University of Tübingen, Auf derMorgenstelle 1, D-72076 Tübingen;
2 Vertretungsprofessur für Ökologie, FB 19 Biologie/Chemie, University of Kassel, Heinrich-Plett-Str. 40, D-34123 [email protected] - http://www.systbot.uni-tuebingen.de
Key words: Arbuscular mycorrhizae, Namibia, rust fungi, soil fungi
Abstract:Our project aims to ascertain data on the biodiversity and ecology of soil fungi, arbuscularmycorrhizae, and plant parasitic rust fungi in important biomes of southwestern Africa.During a first trip to Namibia in March 2001, samples were taken on the observatories Mile46, Sovo, Sonop 903, Otjiamongombe, Toggekry 250, Gellap Ost 3 and Nabaos 7. Firstobservations and results from this trip are presented here.
Results:Soil samples examined for the presence of arbuscular mycorrhizal spores revealed differencesin the number of spores and – although species identification is still pending – speciescomposition. The dominating spore type in the north (Mile 46 and Sovo) is a small, darkbrown Glomus-spore, in the thornbush savanna (Otjiamongombe, Toggekry 250) a lightbrown Glomus-spore and in the south (Gellap Ost 3 and Nabaos 7) a small, yellowish-brownGlomus-spore. The dominance of Glomus-spores is in accordance with the reports by Stutz etal. (1). The number of spores is within the range reported in previous studies from comparableecosystems (2, 3).Root samples stained for the presence of arbuscular mycorrhizal fungi were mainly found tobe mycorrhizal with the exception of Poaceae in Nabaos 7. As spores were present in soilsamples near the plants from which the non-mycorrhizal roots were taken, this warrantsfurther attention. Molecular studies (especially on the samples from Nabaos 7) are under wayto check for the presence of non-staining mycorrhizal fungi and to verify the results obtainedby microscopic techniques.The soil samples that were examined for arbuscular mycorrhiza were used for the study of soilfungi as well. Following the procedure of Gams & Domsch (4) the samples were washed with6 l of water and transferred to isolation media. Over thousand cultures of soil fungi wereisolated. A part of the species is already identified, but identification of most isolates is stillpending. So far, the isolates that have been identified belong to 20 genera; eight genera seemto be new to Namibia.The preliminary results confirm the impression gained during the study of arbuscularmycorrhiza, that Gellap Ost 3 and Nabaos 7 are different from the other areas. From the soilin Gellap Ost 3 and Nabaos 7, fewer fungi were isolated. The number of Zygomycetes isespecially low, possibly due to the lack of organic litter. The dominating isolate was aCoelomycete. However, in this area samples were taken before the onset of the rainingseason, while the raining season in the other areas was well under way. Therefore, furthersampling is necessary to compare the areas under the same water conditions.
105
Our work on rust fungi in Namibia aims to contribute to the rust mycota of selectedbiodiversity observatories and the whole country.The knowledge about Namibian rust fungi is extremely scanty and mostly derived from theworks of Ethel Doidge in southern Africa (e.g. 5). Although the Namibian flora comprises ca.4.000 species of higher plants (6) our extensive literature search revealed only 13 species ofrust fungi (comp. Tab. 1). This species number is extremely low compared with figuresavailable for other African countries [e.g. South Africa ca. 400 (7), Uganda 294 species (8)].A first species inventory of the BIOTA observatories in 2001 indicates that a considerablyhigher number of rusts can be expected to occur in Namibia.
Tab. 1: Rust species hitherto reported for Namibia (compiled from literature)
Rust fungi Host/Family Host/Species
Aecidium acanthopsidis H. & P. Sydow Acanthaceae Acanthopsis sp. Harv.
Aecidium clarum Sydow Iridaceae Ferraria viscaria Schinz
Aecidium dinteri Doidge Mimosaceae Acacia uncinata Engl.
Aecidium lebeckiae P. Hennings Fabaceae Lebeckia simsiani E. & Z.
Aecidium talinophilum P. & H. Sydow Portulacaceae Talinum caffrum (Thunb.) Eckl. & Zeyh.
Puccinia desertorum Sydow Convolvulaceae Evolvulus alsinoides (L.) L.
Puccinia chaseana Arth. & Fromme Poaceae Anthephora schinzii Hack.
Puccinia galeniae Dietel Aizoaceae Galenia africana L.Galenia sarcophylla Fenzl.
Puccinia sorghi Schwein.Syn.: Puccinia maydis Bereng.Syn.: Aecidium oxalidis Thuem.
Poaceae
Oxalidaceae
Zea mays
Oxalis obtusa Jacq.
Uredo augeae H. & P. Sydow Zygophyllaceae Augea capensis Thunb.
Uromyces comptus H. & P. Sydow Convolvulaceae Merremia bipinnatipartita (Engl.) Hallier f.
Uromyces saginatus Sydow Hyacinthaceae Drimia altissima (L.f) Ker Gawl.
Uromyces trollipi Kalchbr. & MacOwan Zygophyllaceae Zygophyllum sp. L.
References:1. Stutz, J. C., Copeman, R., Martin, C. A., Morton, J. B., Can. J. Bot., 2000, 78, 237-245: Patterns of
species composition and distribution of arbuscular mycorrhizal fungi in arid regions ofsouthwestern North America and Namibia, Africa
2. Gaur, A., van Greuning, J. V., Sinclair, R. C., Eicker, A., South African Journal of Botany, 1999,65, 434-436: Arbuscular mycorrhizas of Vangueria infausta Burch. subsp. infausta (Rubiaceae)from South Africa
3. Michel-Rosales, A., Valdés, M., Mycorrhiza, 1996, 6, 105-109: Arbuscular mycorrhizalcolonization of lime in different agroecosystems of the dry tropics
4. Gams, W. & Domsch, K. H., Archiv für Mikrobiologie, 1967, 58, 134-144: Beiträge zurAnwendung der Bodenwaschtechnik für die Isolierung von Bodenpilzen
5. Doidge, E. M., Bothalia, 1950, 5, 1-1094: The South African fungi and lichens to the end of 19456. Craven, P. (ed.), Southern African Botanical Diversity Network Report, 1999, 7, 1-206: A checklist
of Namibian plants7. Crous, P. W., Phillips, A. J. L., Baxter, A. P., Phytopathogenic fungi from South Africa, 2000, 1-
3588. Kåre, A. L., Namaganda, M., Gjærum, H. B., Lidia, 2000, 5, 9-12: Ugandan rust fungi 1. Key to
genera

106
Project ID: 01LC0024 (BIOTA Southern Africa S04)01.10.2000 – 30.09.2003
DELVELOPMENT OF AN INTEGRATED SYSTEM FOR THE INPUT, STORAGEAND ANALYSIS OF BIODIVERSITY DATA IN SOUTHERN AFRICA
A: G. Rambold1, R. Siber2, B. Weber2; B: B. Marbach1, R. Siber2, B. Weber2 and C: B.Büdel2, G. Rambold1
1 Universität Bayreuth, Lehrstuhl für Pflanzensystematik, Abt. Mykologie, Universitätsstrasse 30– NW I, 95440 Bayreuth
2 Universität Kaiserslautern, Fachbereich Biologie, Abt. Allgem. Botanik, Postfach 3049,67653 [email protected]://www.uni-bayreuth.de/departments/planta2/wgr/fmycology.html
Key words: biological soil crusts, checklists, database, ecological and descriptive data, GIS, GUI,interoperability, functional diversity, LIAS, modelling, Mycology.Net, spatial analysis
Abstract:A central task of BIOTA subproject S04 was the development of a uniform standardizedmethod for data collection in the field. According to the observatory transect method, all datacollected in the field were categorized into four different hierarchical levels. The developmentof the database module “DiversityEcology”, which reflects this hierarchical structure,represents the second major achievement of the project. A third major task is the developmentof a graphic user interface (“DiversityNavigator“) for remote editing and analysis of databeing stored in Postgres implementations of database modules, based on the “Diversity” datamodels, which have been developed within the BIOLOG-project GLOPP.
Results:Three main issues had to be addressed during the first year of the project:
A) Composition of a common method for the data collection in the fieldB) Development of a software structure for the recording, storage and analysis of project dataC) Establishment of a Java-based application („DiversityNavigator“) for the request and
processing of data from the Diversity Database Modules via internet
A) Composition of a common method for the data collection in the field
One main task of the project is the development of a uniform standardized method for datacollection in the field to enable interdisciplinary analysis and modeling. According to theobservatory transect method, which forms a common standard of the three BIOTA groups, alldata collected in the field could be categorized into four different hierarchical levels. The selectionand classification of the parameters, which are collected on the different levels, was realized inaccordance to the Database-project SOTER (Global and National Soils and Terrain DigitalDatabases) of the FAO (FAO, 1995). The uppermost level contains data on the observatories asunits, like their geographic coordinates, the vegetation type and the local climate. One levelbelow, data on the habitats delimited against each other within the observatories is combined. Onthe third level, all data on the sites which are investigated by the researchers are combined. Thedetailed characterization of the investigated sites is a fundamental requirement for spatial analysisand modeling of the distribution patterns of species. On the undermost level, data on allspecimens which are investigated or collected in the field and on the applied sampling method arestored. In concordance with all subprojects of BIOTA South the parameters being collected on the

107
two upper levels of the database were determined and their classification adopted to the localconditions. The parameters of the two lower levels were elaborated together with subproject S05.Only such a standardized interdisciplinary approach facilitates detailed spatial and statisticalanalyses and the yield of new knowledge.
B) Development of a software structure for the recording, storage and analysis of project data
The implementation of a modern networked software architecture with continuous databaseaccess and fully developed GIS analysis functionality was another major task of the first projectperiod. The database SQLServer was chosen as DBMS, since this software also allows to processgeographic coordinates. A direct database connect of the GIS-software was realized via anArcSDE gateway. This software architecture facilitates the storage of all data within ageodatabase, which allows an object oriented data structure. ArcInfo was selected as GIS-software, because of the comprehensive analysis and visualization techniques and the newfunctionalities in the area of geostatistical analysis. For the analysis of remote sensing and otherraster data Erdas Imagine is applied. In the near future, the software ArcIMS will be applied topresent the results via internet in the form of interactively generated maps.
During the last months, the database module “DiversityEcology” was created, in whichthe data collected in the field by BIOTA South (data on the observatories and habitats) and alldata collected by subproject S05 are stored. The database module was implemented underconsideration of the Relational Model by Codd (CODD, 1990) and the normalization rules. Thedatabase “DiversityEcology” is an independent database module, that has an interface to the otherexisting Diversity Database modules. The data input in the database module is realized by agraphical user interface (GUI), that facilitates the input and editing of the acquired data viainternet.
C) Establishment of a Java-based application („DiversityNavigator“) for the request andprocessing of data from the Diversity Database Modules via internet
The database modules being developed in the BIOLOG-projects GLOPP (01LC9905) and nowbeing provided to public on the Internet (www.glopp.net) offer suitable tools for themaintenance of taxonomic data (DiversityTaxonomy), literature (DiversityReferences),descriptive data (DiversityDescription) and collection data (DiversityCollection). Their imple-mentation under MS Access will be completed in the near future. The establishment of theJava-based graphic user interface “DiversityNavigator”, established in Biota S04, will allowan Internet-based, platform-independent usage of these databases. The first period of thisproject was dedicated to the establishment of various program technical basics like thedefinition of data exchange formats, the establishment of data import routines and an experi-mental prototype network, as well as the establishment of Diversity-compatible Postgresdatabases. A recently completed draft version of the DiversityNavigator enables the user toimport and edit the taxonomic and collection data, stored in Postgres databases. It further-more allows parallel data queries of databases, which may be located under several domains.The interoperability of the modules could be successfully tested by a configuration with thetwo database modules DiversityTaxonomy and DiversityReferences which enable users tomaintain and query species checklist data. In the forthcoming period of the project theestablished technique will be used to store and maintain all taxonomic and descriptive datagathered in subproject S05.
References:1. Codd, 1990: The Relational Model for Database Management, Version 2. Menlo Park.2. Fao, 1995: Global and National Soils and Terrain Digital Databases (SOTER). World Soil
Resources Reports 74, Rev. 1, 1-125.

108
Project ID: 01LC0024 (BIOTA Southern Africa S05)01.10.2000 – 30.09.2003
BIOLOGICAL SOIL CRUSTS OF TERRESTRIAL ECOSYSTEMS: MODES OFPATTERN FORMATION AND THEIR MODELLING CONCERNING DIVERSITY,STRUCTURE AND FUNCTION IN CRYPTOGAMIC COMMUNITIES
A: B. Büdel1, K. Loris3, V. Wirth8, L. Zedda6 – B: S. Dojani1, T. Feuerer2, M. Küppers 3,R. Lücking4, S. Ott5, W. Reisser7, M. Salisch7, V. Wirth8 – C: B. Büdel1, G. Rambold6
1Universität Kaiserslautern, Fachbereich Biologie, Allgemeine Botanik, Postfach 3049, D-67653 Kaiserslautern2Universität Hamburg, Botanisches Institut und Botanischer Garten, Ohnhorststraße 18, D-22609 Hamburg3Universität Hohenheim, Botanisches Institut, Biologie II, Garbenstraße 30, D-70599 Stuttgart4The Field Museum, 1400 S. Lake Shore Dr., Chicago, Il 60605-2496, U.S.A.5Universität Düsseldorf, Botanisches Institut, Universitätsstraße 1, D-40225 Düsseldorf6Universität Bayreuth, Lehrstuhl für Pflanzensystematik, Universitätsstraße 30-NW 1, D-95447 Bayreuth7Universität Leipzig, Botanisches Institut, Talstraße 33, D-04103 Leipzig8Staatliches Museum für Naturkunde, Erbprinzenstraße 13, D-76133 Karlsruhe
Contact person: [email protected], http://www.uni-kl.de/FB-Biologie/AG-Buedel/
Key words: biological soil crusts, biodiversity, lichens, algae, cyanobacteria, vegetationecology, microclimate, dew fall, fog
AbstractAn overview of the field and laboratory work performed so far is given and first results arepresented. In almost all observatories along the transect, biological soil crusts, composed ofeither cyanobacteria, algae or lichens, or a combination of them, have been discovered.Biomass, expressed as milligrams chlorophyll a per m², ranged from 1 to 80 mg/m². So far, 14genera of cyanobacteria, 21 genera of green algae and 32 lichen species have beendetermined. A new species of the lichens genus Peltula was discovered growing underneathtranslucent quartz pebbles with an inverse internal thallus morphology.Special care was given to the establishment of measuring devices for fog and dew fall along a40 km long gradient from the coast to the inland at Wlotzkasbaken, Namib Desert, in order tocorrelate this data with the vegetation pattern of lichens.
ResultsIt is the intention of our project to study the species (cyanobacteria, algae and lichens) diver-sity, pattern formation and biomass of biological soils crusts and their role in the ecosystem.We want to find out their dynamics, their establishment after disturbance and their influenceon the diversity of the phanerogamous vegetation (e.g. N input via cyanobacterial N-fixation).
Three of us took part in the kick-off trip and actively participated in the selection ofthe observatories. A key on the priority of interest was developed and assigned to the newlyestablished observatories. High priority was granted to observatories with well-developedbiological crusts and high lichen diversity (12 observatories). Intermediate priority wasassigned to observatories without biological soil crusts, but with a rich epiphytic and epilithiclichen flora (8 observatories), where only biodiversity studies are carried out. Observatorieswithout biological crusts or lichens will not be investigated by S05 at this stage.
During this trip, first collections were made and a very rough estimation of thepresence or absence of soil crusts of each observatory was achieved. At each investigationsite, 4 samples were collected for the following purposes: determination of a) cyanobacterial

109
diversity, b) green algal diversity, c) N- and C-content as well as the isotope fractionation, d)chlorophyll a/b content. Two main classes of crusts can be distinguished 1) pioneer crusts(succession stage after disturbance) and 2) climax crusts (part of permanent vegetation).Further differentiation can be made according to the dominating taxonomical group (e.g.cyanobacterial, algal, lichen or moss crust). Determinations of biomass, expressed aschlorophyll a content per m², resulted in values between 1 and 80 mg/m², with an average of33 mg/m² for the first 16 samples along the transect. However, this can only be seen as verypreliminary results. This is also the case for the first diversity analysis of cyanobacteria andalgae. In total, we found cyanobacterial species (free living and in culture) belonging to 14different genera and green algal species (in culture only), belonging to 21 different genera.
In the Nama Karoo, a new species of the lichen genus Peltula was discovered (fig. 1),growing underneath quartz pebbles, exposing its cyanobiont towards the quartz and thushaving an inverse internal morphology. The new description of this species is in preparationand will be published in 2002.
An additional transect from the coast 40 km to the inland was established in the lichenfield of Wlotzkasbaken, meeting the observatory in the center. Along this transect, measuringdevices for fog and dew fall were installed in order to gather data on the events and amount.Those results will be correlated with the very peculiar vegetation pattern of lichens in thisarea. The biodiversity of the lichen vegetation along this transect was investigated on 21 sta-tions and to a distance of 35 km from the coast. Close to the coast, lichen coverage rises dis-tinctly for a short distance, then diminishes constantly inland along the transect. Most speciesoccur from the coast to the end of the investigated transect section, frequency of some specieshowever changes dramatically. At least one species occurs only at inland stations, severaloccur only near the coast.
Lichens were collected in the observatories 18 (Koeroegap), 20 (Numees), 21 (GrootDerm 10), 22 (Quaggafontein 478), 26 (Faminkvlakte 111), 27 (Luiperskop 211), 28 (Moed-verloren 208) and 29 (Rocherpan) in order to investigate the general biodiversity. In theobservatory of Numees vegetation relevés on soil were also carried out by means of a newmethod. It consists of taking digital pictures of cryptogamic communities on a soil surface of60 x 60 cm using an aluminum square frame construction. Since May 2001, lichens have beenidentified in the laboratory. TLC was applied to many samples. Problematic specimens ofvarious genera were sent to specialists. A first list of lichens forming biological soil crustsincludes 32 species. Information on lichen characters are entered into a Diversity databasemodule, and those on lichen specimens in the collection database module (http://www.-diversitycam-pus.net). For the analysis of digital pictures, the program PanoramaFactory(freeware) is used to compose the entire relevé of 60 x 60 cm from 36 single digital pictures.

110
Project ID: 01LC0024 (BIOTA Southern Africa S06)01.10.2000 – 30.09.2003
CHANGES IN BOTANICAL BIODIVERSITY WITH REGARD TO CHANGES INLAND USE PRACTICES AND CLIMATE: STANDARDISED MONITORING ANDTRANSECT ANALYSIS
N. Jürgens , B. Strohbach; M. Akhtar-Schuster, R. Austermühle, T. Becker, B. Hachfeld,U. Schmiedel, M. Strohbach
Coordinators:Prof. Dr. Norbert Jürgens, Botanical Institute, University of Hamburg, Ohnhorststr. 18, D-22609Hamburg, Germany; e-mail: [email protected] Strohbach, National Botanical Research Institute, Private Bag 13184, Windhoek, Namibia, e-mail: [email protected]
Key words: Phytodiversity, vegetation mapping, community ecology, population ecology,population dynamics, monitoring, land use impacts, desertification
Abstract: Vegetation plays an important and multidirectional role within the mechanismsthat control biodiversity. Firstly, vegetation forms an important resource for humankind andis strongly influenced by land use practices. Secondly, vegetation is controlled byenvironmental factors and, therefore, is an indispensable indicator for documentingecological changes. Thirdly, due to its structural, biological, and ecological properties,vegetation itself is an important driver of biodiversity. The botanical subproject S06 aims atlong-term analyses and monitoring of the vegetation in over 30 Biodiversity Observatories.
Results:Earth's biodiversity includes “all organisms and species, their immense genetic variation, aswell as their complex assemblages of communities and ecosystems...“ DIVERSITASPROGRAM Based on this broad definition, S06 integrates the assessment and analyses ofstructures and mechanisms at different temporal and spatial scales. The development of ageneral working protocol will supply all surveyors with a standardised methodology for allBiodiversity Observatories.Structural diversity: The mapping of vegetation along the BIOTA transect band, from theKavango to the Cape, will allow large and medium scale analyses of the vegetation structure.This can only be achieved by the application of remote sensing techniques (S01). Thevegetation map is in the validation process. It will serve for an in-depth understanding of theimpact of changing natural gradients, e.g. rainfall gradient, as well as the influence ofdifferent land use systems [1,2,3].Community diversity:The species composition of plant communities reflects environmental conditions. Based on amulti-scale standardised working protocol, a large number of vegetation data were sampledand classified into different vegetation units. Within the 1 km² large BIOTA Observatories,the assessment of the spatial patterns of different communities was carried out on a regulargrid of 100 hectare.Species diversity:In 2001, for a large proportion of BIOTA Observatories, first inventories of flora andvegetation were documented at four different spatial scales, ranging from 100m², 1,000m²,10,000m² up to 1 square kilometre (Table 1). In some cases, these are linked to demographicanalyses of populations and species, focusing on migratory dynamics. Additionally, themonitoring of the BIOTA Observatories also considers the individual and population level(e.g. location, size, number of fruits) [4,5,6]. The long-term monitoring of the permanent plotsin the western Richtersveld (South Africa) forms the basis for the simulation models insubproject S10.
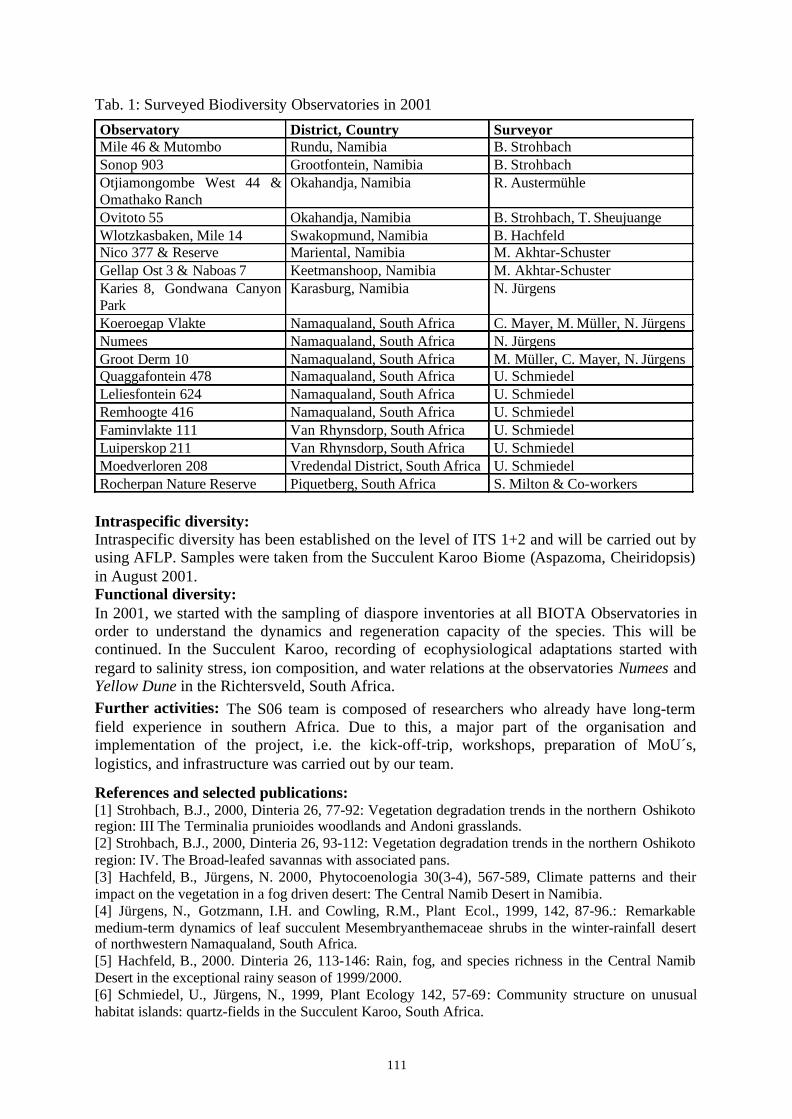
111
Tab. 1: Surveyed Biodiversity Observatories in 2001
Observatory District, Country SurveyorMile 46 & Mutombo Rundu, Namibia B. StrohbachSonop 903 Grootfontein, Namibia B. StrohbachOtjiamongombe West 44 &Omathako Ranch
Okahandja, Namibia R. Austermühle
Ovitoto 55 Okahandja, Namibia B. Strohbach, T. SheujuangeWlotzkasbaken, Mile 14 Swakopmund, Namibia B. HachfeldNico 377 & Reserve Mariental, Namibia M. Akhtar-SchusterGellap Ost 3 & Naboas 7 Keetmanshoop, Namibia M. Akhtar-SchusterKaries 8, Gondwana CanyonPark
Karasburg, Namibia N. Jürgens
Koeroegap Vlakte Namaqualand, South Africa C. Mayer, M. Müller, N. JürgensNumees Namaqualand, South Africa N. JürgensGroot Derm 10 Namaqualand, South Africa M. Müller, C. Mayer, N. JürgensQuaggafontein 478 Namaqualand, South Africa U. SchmiedelLeliesfontein 624 Namaqualand, South Africa U. SchmiedelRemhoogte 416 Namaqualand, South Africa U. SchmiedelFaminvlakte 111 Van Rhynsdorp, South Africa U. SchmiedelLuiperskop 211 Van Rhynsdorp, South Africa U. SchmiedelMoedverloren 208 Vredendal District, South Africa U. SchmiedelRocherpan Nature Reserve Piquetberg, South Africa S. Milton & Co-workers
Intraspecific diversity:Intraspecific diversity has been established on the level of ITS 1+2 and will be carried out byusing AFLP. Samples were taken from the Succulent Karoo Biome (Aspazoma, Cheiridopsis)in August 2001.Functional diversity:In 2001, we started with the sampling of diaspore inventories at all BIOTA Observatories inorder to understand the dynamics and regeneration capacity of the species. This will becontinued. In the Succulent Karoo, recording of ecophysiological adaptations started withregard to salinity stress, ion composition, and water relations at the observatories Numees andYellow Dune in the Richtersveld, South Africa.Further activities: The S06 team is composed of researchers who already have long-termfield experience in southern Africa. Due to this, a major part of the organisation andimplementation of the project, i.e. the kick-off-trip, workshops, preparation of MoU´s,logistics, and infrastructure was carried out by our team.
References and selected publications:[1] Strohbach, B.J., 2000, Dinteria 26, 77-92: Vegetation degradation trends in the northern Oshikotoregion: III The Terminalia prunioides woodlands and Andoni grasslands.[2] Strohbach, B.J., 2000, Dinteria 26, 93-112: Vegetation degradation trends in the northern Oshikotoregion: IV. The Broad-leafed savannas with associated pans.[3] Hachfeld, B., Jürgens, N. 2000, Phytocoenologia 30(3-4), 567-589, Climate patterns and theirimpact on the vegetation in a fog driven desert: The Central Namib Desert in Namibia.[4] Jürgens, N., Gotzmann, I.H. and Cowling, R.M., Plant Ecol., 1999, 142, 87-96.: Remarkablemedium-term dynamics of leaf succulent Mesembryanthemaceae shrubs in the winter-rainfall desertof northwestern Namaqualand, South Africa.[5] Hachfeld, B., 2000. Dinteria 26, 113-146: Rain, fog, and species richness in the Central NamibDesert in the exceptional rainy season of 1999/2000.[6] Schmiedel, U., Jürgens, N., 1999, Plant Ecology 142, 57-69: Community structure on unusualhabitat islands: quartz-fields in the Succulent Karoo, South Africa.

112
Project ID: 01LC0024 (BIOTA Southern Africa S07)01.10.2000-30.09.2003
FUNCTIONAL ZOODIVERSITY IN SOUTHERN AFRICA UNDER CHANGINGENVIRONMENTS AND HUMAN USE
U. Zeller1, M. Ade 1, J. Deckert1, S. Eiseb2, P. Giere1, A. Hoffmann1, F. Koch1, E.Marais2, W. Mey1, J. Plötner1, M. Uhlig1, K. Vohland1, H. Wendt1
1) Museum für Naturkunde, Humboldt Universität, Invalidenstr. 43; 10099 Berlin, Germany;contact person email: [email protected]) National Museum of Namibia, Robert Mugabe Avenue 59; Windhoek, Namibia
Key words: Functional zoodiversity, food webs, reproduction, population ecology
Abstract:
The main objective of this study is to learn about biodiversity, its composition (taxonomy),how it is functioning and how it can be maintained and managed as seen from a zoologistsperspective. In the pilot phase research is focused on small ground living mammals, their re-production and their interactions with arthropods and plants, especially trophic interrelation-ships between these groups.
Hypotheses: We hypothesize that
1) specific small mammal communities differ in their direct influence on vegetation and soilcharacteristics by their digging habits seed dissemination and bioturbation in general;
2) specific small mammal communities directly influence arthropod species richness andabundance mainly due to predation, and thus indirectly affect vegetation communities (con-sumption of herbivores, pollinators, detritivores...);
3) the differential resource utilisation due to changing nutritional demands of the small mam-mals in the course of reproduction contributes significantly to vegetation and arthropod com-munity composition;
4) large mammals (in different land use practices) and climate (esp. water availability) influ-ence performance of soil, vegetation, arthropods, and small mammals. Accordingly the func-tional relationships and ecosystem performance differ between differently managed farms andclimatic zones;
5) the above mentioned indicators of biodiversity can be used for the formulation of manage-ment tools with respect to global change.
Results:
Investigations took place on iodiversity observatories in Karies 8 (Fish River Canyon), GellapOst - Nabaos (Berseba) and Otjiamongombe (Erichsfelde) - Toggekry (Omatako) in Namibia,April - Mai 2001. In South Afrika the biodiversity observatories Yellow dunes, Numees andthe Koerogapvlakte at the Richtersveld were sampled in October 2001.
One main goal of the first phase of the project is to develop and optimize adequate methods tolong term monitor biodiversity. Plots within the biodiversity observatories were selected ac-cording to the ranking method developed by BIOTA-South. A first species inventory was be-gun. Arthropods were collected with Malaise traps, different light traps, pitfall traps, yellowplates, sweeping nets, pyrethrum fogging and various methods of hand collecting.A first analysis of the well-known bio-indicator group Carabidae (ground beetles), based onpitfall trap and light trap sampling as well as singling, showed the following results:

113
Whereas the differences inspecies inventory, diversity andabundance of the ground beetlescommunities between Karies andGellap Ost are obviously due tometeorological, geological, geo-botanical, zoogeographical andgeographical factors on theBIOTA transect, the greatdifference between the Karakulsheep research farm Gellap Ostand the over-grazed observatoryNabaos results from the influenceof different land use.
Trapping for small mammals consisted of a 150 m transectholding 30 Sherman traps and three bucket traps with someadditional traps selectively placed in bushes and trees. Thecomposition of trapped species reflected the assumptionaccording to literature research; except for Mus minutoides,for which the most nothern record could be supplied for theKoerogapvlakte in the Richtersveld.Comparable to the arthropods, small mammals differ in theirspecies composition and abundance according to the positionon the BIOTA transect and the intensitiy of land use. AgainNabaos is impoverished in biodiversity, only the widely distributed G. vallinus was trapped.To analyse trophic relationships, the stomach content of selected small mammals was investi-gated. For this, the stomach was removed from the dead animal and emptied completely. Thevolume was measured and the content was first analysed according to volumen relations. Adetailled analysis on a more elaborated taxonomic basis followed.Anatomy of the reproductive tract and embryology of most trapped small mammal specieswas only poorly or not at all known. First results show that the anatomical structure and re-productive status of the animals deviated from other well documented animals. These dataprovide a necessary basis for the newly developed method of ultrasonic investigation for thein-vivo determination of the reproductive status in the field.
Discussion and outlook
Monitoring zoological biodiversity is a compromise between effort and results. We can eithercover the transect of observatories completely, focus on selected biodiversity observatories toobtain a more comprehensive data set, or record annual cycles.The last point will be approached for the adjacent biodiversity observatories in Gellap Ost andNabaos, as here studies on population dynamics on small mammals have started to gather in-formation on population processes and time slots for reproduction. For the main phase, moreemphasis will be put on population processes, and functional hypothesis approximated by firstmodels will be tested in the field.Another point concerns sustainability, which can only be achieved if the project is acceptedboth in Namibia and South Africa. For this, we are working on the implementation of a MSc-course in "Functional Biodiversity" in cooperation with the University of Namibia. Further-more, native students are introduced into our project and employed in.

114
Project ID: 01LC0024 (BIOTA Southern Africa S08)01.10.2000 – 30.09.2003
DIVERSITY OF NAMIBIAN DRAGONFLIES: EFFECTS OF ANTHROPOGENICCHANGES AND MODELLING ON DIFFERENT SCALES
Frank Suhling2, Erik Braune 1, Heike Hadrys3, Andreas Martens 2, Dagmar Söndgerath1
and Otto Richter1
1 Institut für Geographie und Geoökologie, Technische Universität Braunschweig, LangerKamp 19c, D-38106 Braunschweig, [email protected], [email protected],[email protected].
2 Zoologisches Institut, Technische Universität Braunschweig, Fasanenstraße 3, D-38106Braunschweig, [email protected], [email protected].
3 Institut für Tierökologie, Tierärztliche Hochschule Hannover, Bünteweg 17, D-30559Hannover, [email protected].
Key words: Odonata, ephemeral rivers, species and community diversity, populationdiversity, isolation, habitat suitability, population development, distribution maps.
Description of the project and first resultsThe aim of our project is the development and application of a system for the modelling ofOdonata diversity — on population, species and community level — in arid and semiaridregions of Africa. The model-system will help to understand the dynamics of Odonatacommunities and to evaluate different scenarios of global change and anthropogenic effectsfor ecosystem-management. Odonate diversity can be used as an indicator in this context.
Faunistic survey, monitoring and experimental studiesThe Odonata fauna of Namibia consists of 100 species (Martens et al. in prep.). The highestspecies diversity – reflecting the presence of permanent rivers – is in the NE (Jödicke &Martens in prep.). To study diversity in arid regions on the species and community level wesurvey the Odonata of about 250 aquatic habitats in the western catchments, namely theSwakop, Kuiseb, Tschauchab, Ugab, Huab and Hoanib, and, additionally, in the Fish river.We recorded 52 species in these systems. We characterized the environment and thegeographical situation of all sites. Based on these data habitat suitability models (see below)will help to evaluate the relevance of factors like habitat duration, vegetation, salinity, altitudeand connectivity for the structuring of Odonata assemblages and the distribution of species. Anumber of sites are selected for long term monitoring to study phenology and variations inassemblage structures. Using artificial pond systems we studied habitat selection, life cycles,phenology, and priority. Our studies revealed clear patterns of habitat choice via oviposition,which effects larval density in different structured ponds (Suhling et al. 2001). All species ofthese ponds were able to develop within less than two months; Pantala flavescens requiresonly 31 days from oviposition to emergence. The short development allows species to useeven short existing habitats and may be a major trait of desert Odonata (Suhling et al. inpress). The field studies are accompanied by laboratory experiments, which revealed cleareffects of priority, growth rate and intraguild predation in a two species assemblage,combining the widespread species Trithemis kirbyi and Sympetrum fonscolombii.
Molecular geneticsThe molecular genetic analyses of several ncDNA and mtDNA marker reveal first data on thegenetic diversity and genealogical relationships of the selected study species. The dataprovide first insights into spatial and temporal dynamics of selected key-species in correlationto different ecological settings (see below). The latter will help us to identify pathways used

115
by Odonata to colonize the ephemeral desert habitats, which is not possible with traditionalmethods. The study includes different geographical scales: (a) between habitats in onecatchment, (b) between catchments, (c) between the western catchments and other regions ofNamibia (such as the savanna regions in the northeast), and (d) between Namibia and otherregions like Kenya, in co-operation with BIOTA E07. Based on monitoring data we selected aset of key species, e.g. Paragomphus genei, which exclusively colonizes habitats within thefloodplains. Others such as Crocothemis erythraea and Anax imperator are not limited, sincethey are excellent flyers. We compare the population structures/dynamics of these species inconnection with differences in their mobility. Using additional information on environmentalstress related genes (e.g. variation in heat shock proteins) provide insights into the adaptivepotential of the selected species. Our study also includes genetic information on rare andextremely isolated species, which is a major task when setting up a conservation managementplan. For analyses at the genus level we have chosen Pseudagrion, Orthetrum and Trithemis,which seem to be promising model systems to obtain insights into different stages ofspeciation and differences in species diversity patterns in combination with recent andhistorical habitat shifts.
Ecological modellingModelling is done on different scales. On the habitat scale suitability and population dynamicmodels will be developed and applied for selected species. We use logistic regressionmethods to model habitat suitability. These models are based on presence / absence datatogether with habitat parameters recorded during the survey (see above). As a result we getprobabilities of occurrence in a given habitat for each species. Up to now data of the firstsurvey were explored to find species suitable for modelling and to run first suitability models.For population dynamics we used a model based on “delay-differential-equation”, evolved byCrowley et al (1987) for Odonata of the temperate region. This model was fitted to thespecific conditions of tropical desert Odonata using literature data as well as data of our ownstudies (Schroeder 2001). However, since we found that this model will cause some problemswhile combining it with habitat suitability models we decided to use the Leslie-matrix modelinstead, which allows the incorporation of more complex data. On the catchment scaledifferent habitat models are coupled by a geographical information system (GIS) to form amulti-habitat model. To do so we take into account the spatial spread of the individuals andthe gene flow between different populations. With this model system we will describe thediversity of Odonata under different scenarios, e.g increase of drought, increase of water use,building of dams. We produce distribution maps (basing on GIS), which allows theapplication of countrywide models the diversity in the second phase.
Identification keysAs a general preparation we produced keys to the adults and larvae of the Namibian Odonata,which will be published with the National Museum of Namibia, Windhoek. The larval keyalready allows identification of all genera, but not all species, since a number of larvae areundescribed. The adult key already allows identification of most of the species.
ReferencesCrowley, P.H., R.M. Nisbet, W.S.C. Gurney & J.H. Lawton 1987. Population regulation in animals with
complex life-histories: formulation of a damselfly model. Adv. Ecol. Res. 17: 1-59.Schroeder, E. 2001. Analysis of population dynamic models for dragonflies based on delay-differential
eqautions. Diploma-thesis, Technische Universität Braunschweig, 89+XIV pp.Jödicke, R. & A. Martens 2002. The Odonata of Caprivi. Int. J. Odonatol. in press.Martens, A., R. Jödicke & F. Suhling 2002. Odonata of Namibia. a priliminary checklist. CimbebasiaSuhling, F., R. Jödicke & W. Schneider 2002. The Odonata of African arid regions: are there desert species?
Proceedings of the 1st Symposium on African Odonata. CimbebasiaSuhling, F., K. Schenk & T. Padeffke (2001): Dragons of desert ponds: Colonization patterns, life cycles and
structural diversity. International Symposium of Odonatology, Schweden.

116
Project ID: 01LC0024 (BIOTA Southern Africa S09)01.10.2000 – 30.09.2003
STRUCTURAL AND SPECIES DIVERSITY IN SEMI-ARID SAVANNAHS OFSOUTHERN AFRICA: THE IMPACT OF LAND USE AND CLIMATIC CHANGE
F. Jeltsch, M. Schwager and J. Tews
Coordinator: Prof. Dr. Florian Jeltsch, Plant Ecology and Nature Conservation, Institute forBiochemistry and Biology, University of Potsdam, Maulbeerallee 2, D-14469 Potsdam, Germany;e-mail: [email protected] German Partners: Prof. Dr. P. Poschlod (Nature Conservation, University ofMarburg/Regensburg); Prof. Dr. A. Rasa (Zoology, University of Bonn); Dr. K.-H. Müller(Geography, University of Marburg); Prof. Dr. R. Brandl (Ecology, University of Marburg/ UFZHalle); PD Dr. M. Bollig (Ethnology, University of Cologne) South African Partners: Prof. Dr. S.J. Milton (Nature Conservation, University of Stellenbosch);Dr. W.R.J. Dean (FitzPatrick Institute for Ornithology, University of Capetown); Prof. Dr. N. vanRooyen (Botany, University of Pretoria)
Key words: population ecology, extinction risk, spatial-explicit simulation models, remotesensing, landscape structures
Abstract:In arid and semiarid savannahs species diversity is strongly influenced by the structuraldiversity (landscape structures at different scales), which in these systems is characteristicallyformed by woody vegetation. The southern Kalahari based subproject S09 uses aninterdisciplinary approach to investigate these structural-dependent relationships, combiningfield studies (zoological, botanical, genetic and socio-economic), remote sensing, and spatial-explicit simulation models. The aim of the project is to evaluate the impact of current andrealistic future land use intensities and climatic change scenarios on the survival of selectedanimal and plant species and on species diversity in general.
Results:According to a basic definition of ‘biodiversity’ (UN Environmental Program 1992)biological diversity means the variability among living organisms and the ecologicalcomplexes of which they are part. This not only includes species diversity but also thediversity of ecological structures or ‘structural diversity’. Structural diversity encompassesboth functional structures such as food webs and spatial structures, i.e. landscape structures atdifferent scales. Both elements of biodiversity – species and structural diversity – are closelyinterlinked. Any changes in habitat structure inherit the risk of species loss. The aim of thisinterdisciplinary project is an improved understanding of the complexity of impacts oflandscape dynamics (structural diversity) on species diversity under the major driving forcesin the southern Kalahari, namely land use and climatic change.
Structural diversity: - In the case of the southern Kalahari structural diversity ismainly formed by woody vegetation. Major impacts on structural diversity in this system canbe addressed to land use and climate change. Land use, e.g. in the form of cattle farming orfire wood cutting have evolved major changes of landscape structure. Socio-economicsurveys show, among other results, that the majority of farmers are utilizing camelthorn trees,both for domestic and commercial purposes. Roughly about 20 % of the farmerscommercially utilize the wood and remove very large quantities. Long- to mid term changesof tree cover ratios have also been quantified by remote sensing techniques using satelliteimages and aerial photographs covering the last 60 years. An automatic tree detection

117
software has been developed writing a vector database into GIS. These spatio-temporal dataof landscape structure can be compared with existing spatial vegetation models (e.g. Jeltsch etal. 1999) and linked to spatially explicit population models under varying land use andclimate change scenarios. In this context a new model focuses on the population dynamics ofGrewia flava, a common shrub in the southern Kalahari. Cattle-induced shrub encroachment –an ecologically and economically important form of land degradation in savannas - is usuallyexplained in terms of livestock grazing altering shrub-grass competition in favour of shrubs.However, empirical studies and modelling results show that under grazing of domesticlivestock shrub seed dispersal in cattle dung has a significantly higher influence on Grewiaencroachment. This has important implications for land use and management since, onceyoung shrubs have established, encroachment is almost irreversible.
Species diversity: - To understand the impact of climate change and land use onspecies diversity both directly and via structural diversity, several target species were selectedthat depend on different elements of vegetation structure (trees, shrubs) on different spatialscales. The target species include animal species (tawny eagle Aquila rapax, small carnivorese.g. yellow mongoose Cynictis penicillata, tree rat Thallomys nigricauda, sociable weaverPhiletairus socius and others) and plant species (Grewia flava, Lycium sp. and others). Forthese species field studies on habitat use, population dynamics and population genetics will belinked with population simulation models. For the tawny eagle first model results show thatthe viability of the Kalahari-population is directly dependent (i) on the mean of annual rainfalland (ii) on the variability and chronological order of rainfall. It is therefore suggested thatpossible climatic changes will have a major effect on the survival of this population. Futuremodel investigations will show the effect of changes in vegetation structure (mainly largenesting trees), on the survival of the population. For sociable weavers a metapopulation modelhas been built to investigate (i) the effect of the spatial distribution of rainfall and (ii) theeffect of changes in size and distribution of trees on this population. Similarly to the tawnyeagle, preliminary results suggest a strong dependence on rainfall patterns. First results of thefield investigations on tree rats show a dependence of the population distribution as well as ofthe individual behaviour of animals on the spatial structure of trees and shrubs. Furtherinvestigations on population dynamics and on population genetics were started and data willbe used in a simulation model to show the effect of tree- and shrub structure on thepopulation. In a new study on the effects of land use on the biodiversity of small carnivores inthree different grazing regimes (cattle vs. sheep, vs. game) first results show, that yellowmongooses strongly depend on vegetation structure: they dig their sleep and reproductionburrows preferably under shrubs of Acacia hebeclada and Rhigozum trichotomum (>10 mdiameter), whereas shrubs of Acacia mellifera were rarely used. Density and distribution ofshrubs with an equivalent size differ between the three grazing regimes. The aim of ourfurther study is to investigate the consequences of these observed differences on thecomposition of the small carnivore guild. Data on vegetation patterns show the importance ofAcacia trees on the overall plant species diversity and abundance: the number of plant speciesand total vegetation cover were always higher underneath Acacia trees than in treeinterspaces. In a comparison between three sites of different land use intensities the effect ofAcacia trees were strongest at the unmanaged site. Increasing land use intensity wascorrelated with a decline in species/cover of several physiognomic categories. Experimentalapproaches to test which factor - shading, nutrient enrichment or dispersal - is limiting theoccurrence were started with a shrub, Lycium sp. First results on shading and nutrientenrichment have shown that germination rate depends on nutrients. Linking of individualpopulation models with models of large scale vegetation dynamics is in progress.
Reference:Jeltsch, F., Moloney, K.A. and Milton, S.J., 1999. Detecting process from snap-shot pattern: lessonsfrom tree spacing in the southern Kalahari.- Oikos 85, 451-467

118
Project ID: 01LC0024 (BIOTA Southern Africa S10)01.10.2000 – 30.09.2003
WHICH PROCESSES AND MECHANISMS DETERMINE THE BIODIVERSITY IN(SEMI-) ARID PLANT COMMUNITIES?
B. Reineking and C. Wissel
Department of Ecological Modelling, UFZ-Centre for Environmental Research Leipzig-Halle,P.O. Box 2, D-04301 [email protected]
Key words: allocation, allometric scaling, coexistence, competition, generic plant model,individual-based simulation model, plant functional types, plant species diversity, spatial andtemporal pattern, stochastic environment, trade-off
Abstract:The arid winter-rainfall region of the western Richtersveld (South Africa) harbours anunusually high plant species richness, with maximum values of more than 40 perennials per100 m2 [1]. Species coexistence in this system has been attributed to several factors, includingthe plants’ relatively short life span, the high spatial and temporal heterogeneity inenvironmental conditions, notably water availability, and the compartmentalisation of theshallow soil by rocks, which reduces the spatial scale of competitive interactions betweenplants.The aim of the project is to assess the relative importance of these and other factors, using anindividual-based, generic plant model. Simulated spatial and temporal dynamics of the plantcommunity under different scenarios are compared with those observed in long-term fieldstudies, and with predictions from general theories of biodiversity.
RESULTS:In generic plant models, different species are distinguished by only a few characteristics (2-4).In particular, these are the biomass allocation patterns to different compartments such asroots, stem, leaves, seeds and storage tissue. Other characteristics, such as seed size ordispersal distance, may be included. The distinguishing characteristics form a low-dimensional ‘design space’. ‘Species’ are points within this design space and may be bestinterpreted as representations of plant functional types.The costs and benefits of biomass allocation to different compartments, in terms of water andphotosynthate gain or loss, are based on allometric scaling relationships (5). A sub-model isdeveloped of the dependence of occupied soil volume and water uptake on root biomass.Model results include characteristic size and abundance distributions of different plant typesgiven specific environmental conditions (cf. 6).The analysis of the simulation model has two main points of reference: general theories ofbiodiversity on a more abstract level, and long-term field studies on a more concrete level.Several ‘unified’ or ‘general’ theories of biodiversity and competitive coexistence haverecently been proposed (7, 9), each emphasising different possible mechanisms. Thesimulation model’s dynamics will be interpreted in the light of these theories to assess thedegree to which they bear on the specific system under study.The project’s empirical basis are long-term observations carried out by Norbert Jürgens andco-workers over the last 20 years, in which location, size and number of fruits of individualplants have been recorded on permanent plots in the western Richtersveld (South Africa).

119
By mapping real species’ allocation patterns into the model’s plant design space, the modelresults can be directly compared to the specific fine scale patterns observed in the westernRichtersveld region. However, because of the model’s generic structure, key findings of thestudy are likely to generalise to other ecosystems of similar environmental conditions.
REFERENCES:1. Jürgens, N., Gotzmann, I.H. and Cowling, R.M., Plant Ecol., 1999, 142, 87-96.2. Tilman, D., Plant Strategies and the Dynamics and Structure of Plant Communities, 1988,
Princeton University Press.3. Kleidon, A. and Mooney, H.A., Glob. Change Biol., 2000, 6, 507-523.4. Schwinning, S. and Ehleringer, J.R., J. Ecol., 2001, 89, 464-480.5. West, G.B., Brown, J.H. and Enquist, B.J., Nature, 1999, 400, 664-667.6. Enquist, B.J. and Niklas, K.J., Nature, 2001, 410, 655-660.7. Chesson, P., Theor. Popul. Biol., 2000, 58, 211-237.8. Hubbell, S.P., The Unified Neutral Theory of Biodiversity and Biogeography, 2001, Princeton
University Press.9. Pachepsky, E., Crawford, J.W., Bown, J.L. and Squire, G., N

120
Project ID: 01LC0024 (BIOTA Southern Africa S11)01.10.2000 – 30.09.2003
SOCIO-ECONOMIC ASPECTS OF CHANGES IN BIODIVERSITY IN SOUTHERNAFRICA
Buss, H.-J., Falk, T., Heimpel, B., Hoffmann, I., Isaacs, G., Kirk, M., Knight, R., Krug,M., Nuppenau, E.-A., Schneiderat, U., Werner, D.
Contact: Dept. of Livestock Ecology, Justus-Liebig-University Giessen, Ludwigstr. 21, 35390Giessen: [email protected]; http://www.uni-giessen.de/ghi5
Key words: household, communal and commercial farming, livestock, game, rangelandmanagement, natural resources, agro-biodiversity, property rights, new institutional economics,resource economics
Abstract:The study aims at giving an overview over past and present livestock and rangeland managementstrategies in communities and on commercial farms along the BIOTA transect. It also focuses oninstitutions ruling the management. About 60 households in three rural communities and 20 farmowners in two locations in South Africa and Namibia have been interviewed by structured andsemi-structured interviews. RRA tools were used in the communities. Preliminary results indicatethat- livestock production systems differ along the transect, depending on the environmental
conditions, technologies applied, as well as on cultural background and traditions of thedifferent groups,
- the production systems have flexibly adapted to changing environmental, economic andpolitical conditions,
- water access, water quantity and quality are identified as a key problem for people living inrural areas (esp. communal),
- rangeland quality with respect to bush encroachment is an issue for commercial farmers, whotake countermeasures, whereas livestock owners in communal areas do not actively react onchanges in vegetation composition.
Results:Activities of BIOTA S11 started from July 2001 in the four sub-projects (SP)1. identifying and analysing the actual land use practices and animal husbandry systems,2. identifying suitable management practices for bio-economic models and rangeland policy,3. assessing property rights over resources, organisational structure, and national tenure
policies,4. assessing the economic value of savannahs and biodiversity with respect to human use.Research methods applied included structured and semi-structured interviews, and interviewswith key persons.SP1 and SP3 did their research jointly in three rural communities (Mutompo; Okamboro,Soebatsfontein). About 40 households were interviewed in a full census in the first two villages,and 50% of all 52 households in the last village. RRA tools such as, resource mapping, and groupdiscussion were applied.Population density is high in all three villages. Household composition showed a high proportionof old people and children below school age, particularly in Okamboro. Livestock holding is a

121
secondary enterprise, since most of the household income is derived from government transfers orwaged labour. Livestock holding differs, since the Herero and Kavango keep mainly cattle,whereas the Nama/Damara keep small stock. Extensive livestock keeping in the communities isless productivity- or market- than security- and savings-oriented. Data on the production of localsmall ruminants were obtained in progeny histories. The body weight and body measurements ofsheep and goats of different ages were taken in more than 20 herds. Most animals were found tobe crossbreeds.A long-term rangeland assessment tool is being developed. It aims at assessing the quality ofcommunal rangeland and at recording the changes from a scientific and farmers' point of view.Local institutions that regulate access to and use of natural resources characterise all communities,however, the importance of traditional vs. modern institutions and organisations variesconsiderably. Local committees as resource allocating and sanctioning authorities have usuallybeen implemented by modern organisations or the state. In all communities, former commonproperty resources are partially privately appropriated and fenced in. The extent of this is mappedby GPS and will be integrated in a GIS analysis in future. Migration and absentee livestockownership is of increasing importance.SP 2 interviewed farm owners in three locations (18 around Erichsfelde, 3 in the Kalahari, 2around Fish River Canyon) using a questionnaire on farm practices. Cattle production is the mostimportant enterprise around Erichsfelde, but tourism contributes to up to 50 % of the income.Farmers notified overgrazing and bush encroachment as problems and take countermeasures.Small ruminant production is most prevalent in the two locations in Southern Namibia. Allfarmers try to make their production more extensive, to lower production costs, and to simplifymanagement.Data on hunting and tourism operations were also collected on farms and integrated in a LinearProgramming model (first LP). It covers range management of wildlife and domestic herbivores.It is temporal, i.e. covers overlapping resource use of several periods, is able to depict the impactsfrom current grazing on future range quality, and integrates sound practices.SP 4 is conducted in co-operation with the Dept. of Botany, University of the Western Cape. Theproject focuses on 4 protected areas (Richtersveld, Namaqualand (Soebatsfontein community),Rocherpan and Riverlands). All stakeholders, such as park management, adjacent commercialfarms and communities, will be interviewed to develop an inventory for the consumptive and non-consumptive use of the natural resources in these areas. An online questionnaire will be set up forinternet users (especially park managers); tourists and locals will be interviewed by structuredquestionnaires. A trial questionnaire has been forwarded to a flower tour operator for distributionto tourists (clients) and the data collected. Specific economic valuations methods will be applied.Aspects of the landscape will be modelled within a GIS and future scenarios generated.Besides the research agenda, contacts to local organisations, extension agents, and researchpartners were made, covering in addition the Richtersveld and Paulshoek area. A vivid exchangewith DRFN, NEPRU, the UNAM (Law Dept.) in Namibia, with the Dept. of Plant Conservationof UCT and with Dept. of Agricultural Economics and Range Ecology, Orange FreestateUniversity Bloemfontein, already exists and is further deepened.
References:1. Hoffmann, I.: The BIOTA project. Paper read at the Namibian Rangeland Forum, Okahandja,
28.6.012. Hoffmann, I., E.-A. Nuppenau, M. Kirk, R. Knight, U. Schneiderat, H.-J. Buss, T. Falk, G. Isaacs:
Socio-economic aspects of changes in biodiversity in South Africa. Poster, Arid Zone EcologyForum, Calitzdorp, 4.-7.9.2001
3. Isaacs, G., R. Knight, I. Hoffmann, T. Leymann: Protected areas. Evaluation techniques forassessing socio-economic impacts for local communities. Poster, Arid Zone Ecology Forum,Calitzdorp, 4.-7.9.2001

122
Project ID: 01LC00071.10.2000-30.09.2003
BIOLOGICAL STUDIES ON MYCO-HETEROTROPHIC PLANTS WITH SPECIALEMPHASIS OF THEIR MYCORRHIZAE
Thassilo Franke1) and Reinhard Agerer2)
Ludwig-Maximilians-University, Faculty of Biology, Department Biology I, Biodiversity –Section Mycology, Menzinger Straße 67, 80638 München1) [email protected] , 2) [email protected]
Key words: myco-heterotrophic plants, arbuscular mycorrhiza, anatomy, Cameroon
Abstract:The project is carried out in the Southwest Province of Cameroon. 12 species (7 genera; 4families) of myco-heterotrophic plants could be recorded. A new species of Gymnosiphon(Burmanniaceae) is described. All species were analysed morphologically and anatomically.Mycorrhizal structures were observed. “Trap cultures” were established, using Hieraciumpilosella L. (Asteraceae) as a mycorrhizal host plant. “In situ” germination tests were carriedout. Observation plots were established near Limbe. Soil sampling, as well as plant monitoringstarted in October 2001.
Introduction:The aim of the project is the investigation of the poorly known biology of myco-heterotrophicplants. Myco-hetrotrophic plants receive all essential nutrients (including carbohydrates) fromendomycorrhizal fungi, which are colonizing their roots and/or rhizomes [1]. The lack ofleaves and chlorophyll, as well as their subterraneous habit lead to the assumption that theylive as parasites on their associated fungi [2]. In tropical Africa comparatively few species ofmyco-heterotrophic plants occur. An exception are glacial refuges like e.g. the rain forestsaround Mt Cameroon in Cameroon’s Southwest Province, which are housing one third of allAfrican species of myco-heterotrophic plants [3].Data concerning the ecology of myco-heterotrophic plants are very scarce and in the case ofmost species - especially those from Africa - not existent. The main goal of this project is theregistration of the species composition of myco-heterotrophic plants in South-west Cameroon,as well as the investigation of the mycorrhiza and the determination of host fungi.
Results:Since the beginning of the project in October 2000, three trips to Cameroon were made. Twotrips were undertaken during the declining rainy season, that comprises the flowering seasonof all myco-heterotrophic plants (orchids excluded). The main mission was the registration ofmyco-heterotrophic plants in the Southwest Province. For this reason the type localities ofseveral species were visited. The rainforests that formerly covered the hills around the villageof Moliwe turned out to be completely replaced by oil palm plantations. This area is the typelocality of Afrothismia pachyantha Schltr. (Burmanniaceae) and Oxygyne triandra Schltr.(Burmanniaceae). As the screening of all nearby patches of remaining forest was without anysuccess, it is to be feared that Oxygyne triandra, which is only known from a singlecollection, became extinct. The type locality of Kupea martinetugei Cheek ined.(Triuridaceae), that also comprises the only presently known population of A. pachyantha(discovered by S. Williams in 1993 [3]) is highly threatened to be transformed into farmland.

123
The presumably endemic K. martinetugei could still be found in abundance. In the EkunduKundu community forest near Mundemba a new species of Gymnosiphon (Burmanniaceae)was discovered and will be described soon. The taxonomical situation of the African speciesof Gymnosiphon turned out to be very confusing. The careful study of type material mightreveal, that another two species (both collected near Mundemba) are new to science.Altogether 12 species (7 genera; 4 families) of myco-heterotrophic plants could be recorded.All detected species were analyzed morphologically and anatomically using light- as well aselectron microscopical (SEM, TEM) techniques. With regard to root anatomy, attention wasmainly paid to the endomycorrhizal structures of the involved host fungi. The mycorrhizalpatterns of all plants, except Auxopus macranthus Summerh. (Orchidaceae), showed the typicalcharacters of an arbuscular mycorrhiza. In order to determine the associated fungus, ‘trap cultures’were established, using Hieracium pilosella L. (Asteraceae) as a mycorrhizal host plant. Rootfragments of Afrothismia winkleri (Engl.) Schltr. (Burmanniaceae), Burmannia congesta (Wright)Jonk. (Burmanniaceae) and Sciaphila ledermannii Engl. (Triuridaceae) were placed onto theliving roots of H. pilosella. As arbuscular mycorrhizal (glomalean) fungi have no host specificity,it is expected that the hyphae will grow over to the roots of H. pilosella. Later on the fungi will bedetermined by molecular-biological comparison of isolated spores (extracted from ‘trap-cultures’)and the fungal hyphae within the root tissues. Further on, it is planned to trace the host.-fungus inthe roots of adjacent photo-autotrophic plants, with which they perform mutualisticendomycorrhizal relations. This also demands a careful screening of the surrounding photo-autotrophic plant species. For this reason observation plots were established in Bimbia-Bonadikombo community forest near Limbe. Staff members of the Limbe Botanical andZoological Garden (LBZG) are carrying out a one-year plant monitoring that started in October2001. Besides this, the abiotic environmental conditions are going to be investigated, especiallythose concerning the physical and chemical soil composition of the rhizosphere. Soil samples thathad been taken at the plots were analysed in the soils research laboratory / Ekona, a facility of the‘Institut de la Recherche Agricule pour le Development’ (IRAD). As the reproduction biology,ontogeny and phenology of African myco-heterotrophic plants is completely unknown, severalfield experiments are performed. “In situ” germination tests are carried out to investigate theontogeny. For this reason cell sieves (Falcon®), were provided with seeds of e.g. B. congesta andburied in the soil. After six months some of the sieves were removed from the soil and examined.The seeds were still vital but no germination process was observed. After 12 months only oneseedling of B. congesta could be detected within the sieves.
References:1. LEAKE J (1994) The biology of myco-heterotrophic (saprophytic) plants. In Tansley Review
No.69, New Phytol. 127: 171-216.2. RAYNAL-ROQUES A. & PARE, J. (1998) Biodiversité des Phanérogames parasites: leur place dans
la classification systématique. Adansonia, sér. 3, 20 (2) : 313-322.3. CHEEK M. & CABLE S. (1997) Plant inventory for conservation management: the Kew-Earthwatch
programme in Western Cameroon, 1993 – 96. Proceedings of the Limbe Conference LimbeBotanic Garden, Cameroon 17-24 January 1997: 29 – 38.

124
Project ID: 01LC0017 (BIOTA West Africa)01.01.2001 – 30.12.2003
BIOTA WEST AFRICA – GENERAL SUMMARY
a) Overall goals
BIOTA West Africa consists of ten subprojects that are part of the joint project BIOTA AFRICA.Each of the three main BIOTA projects aims in its complimentary subprojects at assessing
• dimensions,• spatial and temporal patterns and• functional roles
of biodiversity in some of the most important African ecosystems.The special focus in the upcoming main phase of the BMBF’s BIOLOG programme will be onthe anthropogenic change of biodiversity in recent times, both in its continent-wide impacts aswell as its profound regional peculiarities. The dramatic change is correlated with
• large scale habitat conversion and habitat destruction,• overuse and mismanagement of natural resources,• and is also affected by the summed spectrum of small scale human interferences with
the environment.To design advanced management tools, it is necessary to know the status quo ante, compare itwith the current situation and comprehend the already detectable direct and indirect, immediateand long-term consequences of the change on the ecological and socio-economic stage from thelocal to the sub-continental scale in West Africa. Such tools will be better suited than any of thepresently available management measures for conserving the local and regional biodiversity andfor preserving the ecosystems’ functional integrity. These scientifically designed tools shouldallow us to develop wiser and more sustainable forms of use of both the remaining natural as wellas converted ecosystems. These goals shall be reached by
• concentrating research on a limited geographical area (see Fig. 1, page 15),• posing convergent questions and• applying a harmonised methodology.
The closely aligned, co-operative and interdisciplinary research programme of BIOTA-Westintegrates the following research areas: remote sensing, meteorology, biogeography, botany(cross-cutting over different fields among those ethnobotany and archaeobotany), zoology (withstrong emphasis on entomology, herpetology, chiropterology, and limnology), differentsubdisciplines of ecology, ethnology, archaeology, socio-economics.
b) Research localities and infrastructure
The studies of the Frankfurt-Group (W11) are being executed in Burkina Faso and in Benin withtheir counterparts at the Université de Ouagadougou and the Université National du Bénin. Afurther study (of Barbara Fruth), included under the umbrella of BIOTA-West, will conduct itsfield work in the Republic of Congo. The main geographic focus within BIOTA-West is,however, on Côte d’Ivoire, with the Centre de Recherche en Ecologie (CRE, Université de AboboAdjamé) and the Department of Botany at the Université de Cocody as the main counterpartinstitutions in the country. In Côte d’Ivoire, six of the eight subprojects working in the field arecarrying out their empirical studies. During the first year the permanent research camp of theUniversity of Würzburg (originally financed by the Volkswagen Foundation) has served as thealready fully functioning infrastructural and spatial pivot, allowing some of the projects a flyingstart. In due course the newly constructed and now nearly completed permanent Ivoro-German

125
Field Station in the Comoé National Park will replace the University of Würzburg Field Camp asone of our main infrastructural centres. This new research station will provide excellent workingconditions for both, demanding field studies as well as ambitious laboratory research. The stationis predominantly financed by (1) a single grant for the construction and most of the generaltechnical and laboratory equipment by the Fritz Thyssen Foundation, by (2) valuablecontributions to the infrastructure by the BMBF in the framework of BIOTA West, and (3) by along-term commitment by the Free State of Bavaria and the University of Würzburg to coversome of the basic running costs. In addition to BIOTA-West, some of the BIOTA-East projectsare using this Research Station for comparative investigations. The rights and obligations of theco-operating parties have been stipulated in an exchange of diplomatic notes between thegovernments of the Federal Republic of Germany and the Republic of Côte d’Ivoire, dated March3,1997.
The Comoé Research Station allows the lodging and boarding of up to twenty researchers at agiven time. There is enough space and all the basic equipment in the laboratory building to allowall accommodated users to perform their research efficiently. Apart from the proper ResearchStation inside the Park an Information/Education Centre is being built at one of the entrances ofthe Park. It will serve public relations and educational purposes in a very broad sense and on verydifferent levels (from primary school level to advanced students and scientists) for both, visitorsof the Park and residents living in the Park’s wider vicinity. Through mediating education-adequate knowledge a widely lacking comprehension for the value of natural resources, for thevulnerability of the fragile web of life, the necessity of nature conservation and of a wise andsustainable use of natural resources shall be roused in the local population and far beyond. To thisend much valuable scientific contributions are expected from the BIOTA West projects.
c) The basic natural setting for BIOTA-West’s research
There is a steep climatic gradient prevalent along the north-south transect in the investigated partof West Africa (see map, Fig. x), between the arid southern fringes of the Sahara and the humidrain forests on the Atlantic Coast. This offers especially interesting research opportunitiesregarding a number of aspects of biodiversity alterations within the framework of anthropogenicglobal change. Most of our subprojects utilise this gradient, at least partly, by focussing on localand/or regional patterns of diversity in different taxa. Comparative studies along this gradient willallow the retrospective understanding of local and regional changes as a consequence of previousnatural climate variability within the passed millennia as well as predicting some probablechanges in the future under anthropogenicly altered conditions.
d) Understanding the natural background – a highly important objective
All the human-driven ecological changes can only be clearly identified against• the background of the original patterns of biodiversity, and against• the dynamic processes in the respective ecosystem not yet under strong pressure of
modern man.Since the necessary previous knowledge that is needed here is not available at present for any ofthe investigated biomes, basic research requires most of the room in the preparatory phase of thefirst three years of BIOTA Africa in general and BIOTA West in particular. Such knowledge thatcan nowadays only be acquired by conducting research in least disturbed systems within protectedareas is absolutely unrenouncable. Only with a sound scientific understanding of natural structuresand dynamics will we be able to achieve the already mentioned pre-eminent goal of mitigatingand eventually bringing to a halt further losses of biodiversity. Also for the additional goal ofgiving recommendation for enhancing and broadening the wise and sustainable use of organismal

126
diversity, as claimed by the Convention on Biological Diversity (UNCBD), much basic researchis required.
The efficacy of all management measurements, be it for conservation or the ameliorated use ofbiodiversity, will be scrutinised via long-term monitoring. This monitoring will also serve todetect further changes of biodiversity and to recognise their consequences at the earliest possiblestage and help to clarify their causes. The latter, however, requires to distinguish naturalenvironmental variability from anthropogenicly caused alterations. Also this task cannot beachieved without a sound knowledge of the natural systems’ structural and functional properties.Given the high temporal variability in all natural systems, only long-term research will allow us togain the necessary insights. Time is running out for effectively curing the problems humans havealready inflicted on the worlds ecosystems in general and those of the African continent inparticular. We cannot continue to rely further on a pure trial and error strategy when searching forthe best means to effectively conserve and wisely use biodiversity.
e) Recognising and understanding biodiversity patterns on different spatial scales
BIOTA West is tackling its scientific problems on very different spatial scales –fromsubcontinental to regional, landscape and to a more or less fine-grained local level. During thefirst three years (the preparatory phase that will also serve for capacity building of ourcounterparts and of our own working groups) we strive among others
• to find patterns of biodiversity at different spatial scales,• to correlate them with other variables (e.g., climatic, edaphic, orographic, structural,
temporal), and• to observe short-term processes that influence biodiversity patterns and dynamic
processes.At the basis of such correlational data some modelling and up-scaling on larger areas that can onlypunctually be checked by ground truthing is already possible. This holds especially true whenconsidering what in some instances older data (e.g., former floristic inventories – see the Biomapsproject –W03) and remote sensing are offering us as additional information. However, toeventually understand the true cause-effect relationship, which is our final goal, experimentalanalyses are required wherever possible. Such analyses have usually to be confined to small,exemplary spatial and temporal sectors of the systems and yet are very time-consuming. Suchexperimental studies have been commenced in a number of subprojects, but they will take theirtime (and often will need to be extended into BIOLOG’s main phase) until one reaches firstconclusive results.
f) Achievements up to date
The project BIOTA West started in January 2001. Most subprojects needed some time forpractical preparations, for forming or modifying their institutional links, establishing theirinfrastructure, etc. In the meanwhile this phase has been completed in all ten projects. Severalsubprojects already started with their scientific work. The hitherto achieved results within all thesesubprojects are mentioned in the following short abstracts, due to the very limited space, however,only in sketchy form.

127

128
Project ID: 01LC0017 (BIOTA West Africa W01)01.01.2001 – 31.12.2003
REMOTE SENSING AND GIS BASED SURVEY OF SPATIAL AND TEMPORALBIODIVERSITY DYNAMICS AND ANALYSIS OF BIODIVERSITY ANDGEODIVERSITY INTERRELATIONSHIPS
B. Hörsch, M. Vogel, C. Schultz and G. Braun
Deutsches Zentrum für Luft- und Raumfahrt DLR, DFD-UG, Linder Höhe, 51147 Kö[email protected]; www.dfd.dlr.de
Key words: Remote Sensing, GIS, Biodiversity-Geodiversity, Monitoring, VegetationMapping, Capacity Building
Abstract:This project shall create the basis for a system in which both, remote sensing and GISapplications, form an analysis and monitoring system for the assessment of human impacts onbiological diversity. It is our aim to make the existent biodiversity more understandable andpredictable, taking into account the UN-Convention on Biological Diversity (UNCBD).It is planned to establish a coherent GIS along the entire BIOTA transect in order to allow theanalysis of spatial patterns of biodiversity, its temporal changes and the mutual dependencyof geodiversity and biological diversity. Focus will lie on the comparison of naturalecosystems with those where socio-economic activity has led to various degradation levels ofthe former natural ecosystems. Furthermore, the detection of indications of spatial patterns ofdiversity by remote sensing techniques will contribute to future monitoring and conservationconcepts.Further tasks are the “capacity building” for co-operating partners within the fields ofRemote Sensing and GIS.
Results:Within the last 10 months the data base of a Remote Sensing based GIS for the monitoring ofpresent-state biodiversity within the Western African study sites was built up [1]. In closeinteraction with the project partners the different localities of field work were identified andgeographically referenced.Tasks performed included first of all an investigation of appropriate multitemporal data sets fromcommercial multispectral remote sensing systems (LANDSAT-7-ETM, IKONOS, CORONAetc.). A set of LANDSAT-7-ETM scenes were acquired, covering the identified areas. All scenesordered were georectified to the common UTM system and were integrated as image base into theGIS. Due to the lack of reliable topographical maps, Digital Chart of the World (DCW)-data wereused to integrate features like road networks, populated places and river networks into the spatialdatabase. These primary informations were further used to produce geometrically referencedimage maps which are essential as an auxiliary means for the orientation of the fieldworkers. Theyalso act as a base for mapping training samples which form the basis in the supervisedclassification of vegetation patterns from the multispectral images [2, 3]. The maps wereenhanced by topological information (roads, settlements as far as available) and weresuperimposed by a fine resolution latitude/longitude grid in order to facilitate orientation in thefield and to enable the direct comparison of GPS recordings with mapped features. The mapswere distributed to the project partners in paper printed form. On request, digital maps were alsosupplied.

129
An important feature of a spatial database supporting biodiversity analysis in a more indirect wayare topographical data [4, 5, 6]. They form not only the basis for the absolute geographicalorientation of features in space, but are essential to derive topoclimatological information likepotential water and solar irradiance distributions in the terrain [7, 8]. Due to the lack of mid to finescale topographical maps, interferometric analysis of European Remote Sensing Satellite (ERS)-tandem data has been started to derive Digital Terrain Models from the research areas in the IvoryCoast and Benin.Due to the importance of spatial climatic data for biodiversity modelling, research had beencarried out on downscaling spatially coarse climatic datasets in order to derive climaticinformation on a spatial scale which is compatible to the scale of the research approach [9, 10].Up to now, long term (1960-1990) precipitation and temperature data have been downscaled to aspatial resolution of 5km and 2 km, respectively, as long-term monthly means. In the near future itis intended to add additional climatic parameters and to cover the area with non-aggregatedmonthly mean data.To support the use of spatial data and information systems by the project partners two capacitybuilding activities took place at the DLR in Cologne and at the Institute of Botany in Hamburg.Topics of the first course were primarily remote sensing basics with a special emphasis on sensorsensitivity, reflectance of different surfaces, and generation and interpretation of false colourcomposites. The second course held in Cologne focussed on Remote Sensing basics incombination with GIS architecture for biodiversity research and monitoring [11]. Topics likegeneral data structure, the integration of referenced geospatial together with Remote Sensing dataand the main steps for building up of an information system were dealt with. Integrationpossibilities of raster and vector based spatial data with ecological/biological attributes stored indatabases were addressed partially.
References:1. Dale, V.H.; Offerman, H.; Frohn, R. & R. H. Gardner. (1995). Landscape characterization andbiodiversity research. In: Boyle, T.J.B. & Boontawee, B.: Measuring and Monitoring Biodiversity inTropical and Temperate Forests; Proceedings of a IUFRO Symposium held at Chiang Mai, ThailandAugust 27. - September 2. 1994, Centre for International Forestry Research: Bogor, Indonesia.2. Allner, T., Backhaus, R., Gaito, S. & J. McGlade: 1992: Biodiversity Indication by EarthObservation Data. In: Proc. Of the Central Symp. of the Int. Space Year Coference, Munich, Germany.30 March- 4 April 1992, 863-867.3. Podolsky, R. H. 1990: Monitoring Biodiversity and Landscape Richness with digital Earth Imagery.In: Archives of Photogrammetry and Remote Sensing Vol. 28, Part 7-1, Proceedings of the Symp. OnGlobal and Encironmetal Monitoring.4. Burnett, M. R., August, P. V., Brown, J. H., & Killingbeck, K. T. (1998). The influence ofGeomorphological Heterogeneity on Biodiversity. A Patch-Scale Perspective. Conservation Biology,12, 363-370.5. Nichols, W. F., Killingbeck, K. T., & August, P. V. (1998). The Influence of GeomorphologicalHeterogeneity on Biodiversity. A Landscape Perspective. Conservation Biology, 12, 371-377.6. Wohlgemuth, T. (1998). Modelling floristic species richness on a regional scale: a case study inSwitzerland. Biodiversity and Conservation, 7, 159-177.7. Antonic, O., Marki, A., & Krizan, J. (2000). A Global Model for Monthly Mean Hourly DirectSolar Radiation. Ecological Modelling, 129, 113-118.8. Wang, J., White, K., & Robinson, G. J. (2000). Estimating surface net solar radiation by use ofLandsat-5 TM and digital elevation models. International Journal of Remote Sensing, 21, 31-44.9. Florinsky, I. V., Kulagina, T. B., & Meshalkina, J. L. (1994). Influence of Topography onLandscape Radiation Temperature Distribution. International Journal of Remote Sensing, 15, 3147-3153.10. Franklin, J. (1998). Predicting the Distribution of Shrub Species in Southern California fromClimate and Terrain-derived Variables. Journal of Vegetation Science, 9, 733-748.11. ASPINALL, R. (1992). SPATIAL ANALYSIS OF WILDLIFE DISTRIBUTION AND HABITAT IN A GIS. IN:PROC. 5TH INTERNATIONAL SYMPOSIUM ON SPATIAL DATA HANDLING.

130
Project ID: 01LC0017 (BIOTA West Africa W02)1.1.2001 – 31.12.2003
BIOPHYSICAL AND HYDROMETEOROLOGICAL PARAMETERS WITHIN THEFRAMEWORK OF TERRESTRIAL BIODIVERSITY RESEARCH
Dr. Jörg Szarzynski 1, Dipl. Geogr. Thomas Motzer 1 and Prof. Dr. Dieter Anhuf 2
1 Institut für Physische Geographie und Länderkunde, Universität Mannheim, L 9, 1-2,D-68131 Mannheim2 Instituto de Estudos Avançados, Av. Prof. Luciano Gualberto, Travessa J, 374, térreo, CidadeUniversitária, 05508-900, São Paulo, [email protected]
Key words: abiotic environmental conditions, microclimate, energy budget, water balance
Abstract:
Since many ecological aspects of plants and animals are strongly related to local abioticconditions, measurements of meteorological and geophysical data provide comprehensive basicinformation within the discussion of biodiversity stearing mechanisms. The scientific contributionintends to link spatial and temporal variations of abiotic components within dry island forests andsavannas in Comoé National Park (Ivory Coast) to actual questions of biodiversity research.
Results:
Two representative experimental sites have been established within savanna and island forestlocations in order to determine relevant data concerning the energy- and water-budget, as well asvertical profiles of micrometeorological parameters (Fig. 1). Furthermore, the analysis of digitalhemispherical photographs enables us to describe the structure of vegetation within the observedformations (Fig. 2).
In addition, comparative studies will be conducted in the primary tropical rain forest insouthwestern Taï National Park. Comprising an area of 457.000 ha this location representsone of the last large humid forest habitats in West Africa. Known as a centre of highbiodiversity the Taï Park was declared as a World Heritage Region by the UNESCO in 1982.
In the further course of the project the bioclimatological investigations will be carried outsimultaneously within dry forests/savannas in Comoé Park and evergreen forests of Taï Park inorder to analyse the largescale interannual precipitation variability and its impact on the differentvegetation formations.
Appendix:

131
Fig: 1: Installation of technical equipment at the savanna station
Fig. 2: Partly analyzed hemispherical photography with annual sun-tracks

132
Project ID: 01LC0017 (BIOTA West Africa W03)1.1.2001 – 31.12.2003
BIODIVERSITY OF AFRICA IN THE GLOBAL CONTEXT: SPATIAL PATTERNSOF VASCULAR PLANT DIVERSITY IN A CHANGING ENVIRONMENT(BIOMAPS PROJECT)
J. Mutke, W. Küper, G. Kier and W. Barthlott
Botanisches Institut, Abt. Systematik und Biodiversität, Rheinische Friedrich-Wilhelms-Universität, Meckenheimer Allee 170, 53115 [email protected]; www.botanik.uni-bonn.de/system
Key words: biodiversity mapping, biogeography, vascular plants, geodiversity, spatial analysis,databases, GIS, information system
Abstract:BIOTA BIOMAPS aims at the analysis of spatial patterns of African phytodiversity at regional tocontinental scale. Using an inventory-based biodiversity mapping approach, a computer basedmap of total vascular plant species richness for continental Africa has been generated (usingdiversity figures for some 470 geographical units). For the first time, detailed statistical analysesof the correlation of total plant species richness with climate, geodiversity and other abioticparameters are now available on the continental scale for Africa. Additionally, we computed thefirst continental map of endemism richness. For more detailed analyses using a higher spatial andthematic resolution, we are establishing a database on the distributions of selected plant taxa.Until now, distribution data for about 1600 plant taxa are analysed. The established databasesprovide a standardised data structure to make the scat tered data on floristics, biogeography, andbiodiversity of African vascular plants available.
Results:As a baseline against which the results of more detailed analyses can be tested, we mapped andstatistically analysed spatial patterns of overall species richness of African plants on a continentalscale [1]. For these analyses, the complete datasets used by Lebrun [2] and Barthlott et al. [3, 4]were revised, significantly expanded and reanalysed using an inventory-based biodiversitymapping approach. The dataset currently includes 470 records of summary data taken fromliterature on the phytodiversity of geographical units such as mountain ranges, national parks,provinces, and others. Using a new methodological approach, we mapped endemism richnessvalues (an index combining endemism and species richness) for the seed plant flora of the 20African biogeographical regions [5].More detailed analyses were started using a taxon-based biodiversity mapping approach. Therespective dataset with distribution data for single taxa of African vascular plants allows not onlyanalyses with higher spatial resolution. It is as well the basis for more detailed studies of differentaspects of phytodiversity patterns such as endemism, phylogenetic diversity and complementarityanalyses. During the first months of the project the basic infrastructure for this approach wasestablished. Using the BioCise data model [6] a database for the management of plant distributiondata was developed and linked to GIS. Details of the database and possible co-operation are co-ordinated with W. Berendsohn (BIOLOG Biodiversity informatics). A continent-wide GIS onhigh-resolution geodata (e.g., DEM, protected areas, climate, geodiversity and vegetation) wasestablished using internet-based information as well as data provided by BIOTA W01. These dataare used for the spatial analysis of plant distributions in relation to the abiotic environment toallow predictive modelling of the probable range of a species. In addition, a Gazetteer with more

133
than 1 Mio geographical features on the African continent was compiled using different datasources (NIMA, CIAT, etc.). This reference system facilitates the assignment of exact spatial co-ordinates to plant record localities. Using different online-databases (mainly the Kew Record oftaxonomic literature) a dataset on more than 5000 monographs and journal articles on taxonomy,floristics, biogeography and phytosociology of African plants was established and revised for ourpurposes. Extracts from the dataset were provided for other BIOTA subprojects, as well. Based onour information about available plant distribution data, we are establishing a list of selected spe-cies, which are of interest considering our analytical purposes and the work of our co-operatingpartners. For this work, we specifically co-operate with BIOTA sub-projects W04, W09, W11,E04 and further taxonomic experts.Until now, two main types of taxon-based data are analysed. The larger dataset at the moment aredot maps of the distribution of some 1240 vascular plant taxa belonging to 124 genera digitisedfrom the Distributiones Plantarum Africanarum. The second dataset is based on the localitydescriptions of plant specimens cited in taxonomic revisions. Until now, data on some 420 taxafrom 19 plant families have been entered in the database. Furthermore, we are in contact withspecialists for various plant groups with the aim of joint analyses of their distribution data. Firstplant groups to be analysed are Scrophulariaceae, Aristolochiaceae, Melinideae. In addition, weare in contact with working groups (e.g., H.P. Linder, J. Lovett, P. Williams) doing comparableanalyses. Altogether, our taxon-based datasets currently include distribution data of some 1600plant taxa. First analyses of diversity patterns for various plant functional types illustrate thevariety of analytical applications of this datasets [7].
References:1. Mutke, J., Kier, G., Braun, G., Schulz, C., and Barthlott W. (in press), Patterns of African vascular
plant diversity - a GIS based analysis. Syst. Geogr. Pl.2. Lebrun, J. (1960), Sur la richesse de la flore de divers territoires africains. Bull. Séances Acad. Roy.
Sci. Outre-Mer 6, 669-690.3. Barthlott, W., Lauer, W. and Placke, A. (1996), Global Distribution of Species Diversity in vascular
Plants: Towards a World Map of Phytodiversity. Erdkunde 50, 317-327.4. Barthlott, W., Biedinger, N., Braun, G., Feig, F., Kier, G. and Mutke, J. (1999), Terminological and
methodological aspects of the mapping and analysis of global biodiversity. Act. Bot. Fennica 162, 103-110.
5. Kier, G. and Barthlott, W. (2001), Measuring and Mapping endemism and species richness: a newmethodological approach and its application on the flora of Africa. Biodiversity and Conservation 10,1513-1529.
6. Berendsohn, W. G., Anagnostopoulos, A., Hagedorn, G., Jakupovic, J., Nimis, P. L., Valdés, B.,Güntsch, A., Pankhurst, R. J. and White, R. J. (1999), A comprehensive reference model for biologicalcollections and surveys. Taxon 48, 511-562.
7. Mutke, J., Küper, W., Braun, G. and Barthlott, W. (2001), Analysing diversity patterns of differentplant functional types in Tropical Africa. Verhandlungen der Gesellschaft für Ökologie 31, 63.

134
Project ID: 01LC0017 (BIOTA West Africa W04)1.1.2001 – 31.12.2003
PHYTODIVERSITY AND DYNAMICS OF HABITAT FRAGMENTS IN THE IVORYCOAST: SPATIAL AND TEMPORAL PATTERNS OF BIODIVERSITY CHANGES,EDGE EFFECTS, AND THEIR FUNCTIONAL CONSEQUENCES IN NATURALAND DISTURBED ECOSYSTEM MOSAICS
Dethardt Goetze 1, Annick Koulibaly2, Klaus Hennenberg1, Rudolf Pivarci1, PhilippeKersting3, Dossahoua Traoré 2, Stefan Porembski1, Jörg Grunert3
1 Universität Rostock, Institut für Biodiversitätsforschung, Allgemeine und Spezielle Botanik,Wismarsche Strasse 8, D-18051 Rostock, Germany2 Laboratoire de Botanique, UFR Biosciences, Université de Cocody, 22 B.P. 582, Abidjan22, Côte d’Ivoire3 Universität Mainz, Geographisches Institut, D-55099 Mainz, [email protected];http://www.biologie.uni-rostock.de/abt/botanik/ botanik.html
Keywords : forest island, forest regeneration, forest-savanna ecotone, global change,landscape structure, fire, soil properties, sustainable land management, socio-economicalconditions, modelling, vegetation map, remote sensing, GIS, Comoé National Park
Abstract:In the forest-savanna mosaic of West Africa, habitat fragmentation caused by man leads to adecline of natural forests, their increasing isolation, a growing influence of edge effects fromthe surrounding savanna [1], and an alteration of species composition. As a result,biodiversity, as the most important natural resource of the country and basic foundation forthe livelihood of the inhabitants, is declining. In a period of intensification of land use andglobal change, the consequences of habitat fragmentation on biodiversity are being studiedwith a focus on temporal and spatial patterns of biodiversity changes, as well as theincreasing significance of edge effects. The core questions deal with the consequences for theecological functioning of natural and cultivated ecosystems [2,3], the quantification oflandscape diversity, and the establishment of recommendations for a sustainable land useaiming at a long-term preservation of biodiversity. For this purpose, field data are beingassessed in the Comoé National Park and neighbouring utilized areas by use of varioustransect methods and the newly installed biodiversity observatories. They are being mergedwith remote sensing data and processed to models using advanced statistical procedures andGIS.
Results:The analysis is subdivided into five thematic sections:1. Spatio-temporal analysis of the development of habitat patterns and extrapolation ofthe results to larger areas. Based on the analysis of new remote sensing data and intensivefield work, a map of the important vegetation types of the National Park is being generated. Itserves as a basis for the subsequent analysis of landscape structure and diversity. Whententatively related to former data [4], spatio-temporal dynamics of the forest islands becomeobvious; however, a consistent trend has not been detected yet. The predominance of certainvegetation types in the respective forest islands shows that under natural conditions we mayreckon with high beta-diversity among particular forests, whereas the diversity of vegetationtypes within a forest island is limited. Currently, the data of training areas are being

135
extrapolated onto larger areas within the satellite image in order to generate a GIS vegetationmap of the entire region through multiple ground truthing. (Cooperation with W01, W03,W09, W11)2. Analysis and modelling of edge effects between forest and savanna. Along selectedmonitoring transects in forest islands of the National Park, botanical and structural relevés arebeing collected, and the long-term influences of the factors grazing, soil disturbance, and fireare being investigated by means of experimental interference. Furthermore, naturalregeneration of woody plants and herbaceous standing crop are being recorded, andexperiments on the establishment of transplanted seedlings and in-situ germination of selectedwoody species are being carried out. Germination characteristics are being compared undercontrolled laboratory conditions. The biotic parameters are being merged with thedetermining parameters (microclimate, soil, structure, fire). Though a clear structural andmicroclimatic gradient was shown to exist along the forest edges during the rainy season, thetransplanted seedlings of characteristic tree species showed the same survival rates andrelative growth during the first 2 months. Thus, the dynamics of forest edges are presumablydetermined by the microclimatic conditions of the dry season as well as timing and intensityof the fire events. (Cooperation with W02, W03, W06, W09, W11, S09)3. Study of the land utilization regime in neighbouring areas of the National Park,comparison of the consequences of habitat fragmentation in the utilized and protectedareas. In the cultivated areas, monitoring plots were established in close agreement with thelocal inhabitants, and the botanical inventory was started. Currently, the vegetation structure[5] and the occurrence of species groups indicating certain types of land use are beinganalyzed in monitoring transects [6]. It becomes evident that due to utilization, forest edgeecotones are becoming larger at the expense of the forest area. Hence, the effects of abioticparameters (microclimate, structure, soil, fire) are being investigated. On the basis ofextensive polls at local administrative offices and questionnaires of the population in the areasunder study, data are being obtained and analyzed on past and present land-use practices aswell as the socio-economic conditions of the inhabitants. This in turn will aid interpretation ofpresent and future ethnobotanical developments. Intensive capacity building is beingundertaken by an Ivorian PhD candidate working on this topic. (Cooperation with W01, W03,W09, W11)4. Study of the soil properties. Currently, catenas and chemistry of the soils of the area arebeing analyzed in order to systematically quantify the impact of soil properties on theformation of forest-savanna mosaics and on the regeneration of forests. (Cooperation withW02, W06)5. Comparative long-term monitoring of biodiversity changes at monitoring plots inrelation to anthropogenic factors. An observatory was installed each inside and outside thePark by means of a DGPS. Its vegetation was assessed and classified in all 1 ha subplots [7].The subplots to be analyzed by all working teams were determined by means of a stratifiedranking procedure. A detailed inventory of the vegetation of the subplots and of additionalpermanent transects has been commenced. Another pair of observatories will shortly beinstalled in an adjacent region. (Cooperation with W01, W02, W03, W06, W07, W09, W11)
References:1. Forman, R. T. T., 1995: Land mosaics: the ecology of landscapes and regions. Cambridge.2. Schulze, E. D. & Mooney, H. A., 1993: Biodiversity and ecosystem function. Berlin a. o.3. Walker, B. & Steffen, W. (eds.), 1996: Global change and terrestrial ecosystems. Cambridge.4. Hovestadt, T., Yao, P. & Linsenmair, K. E., 1999: Plant Ecol. 144: 1–25.5. Gautier, L., Chatelain, C. & Spichiger, R., 1994: XIIIth AETFAT Congress, Malawi: 1339–1350.6. Koulibaly, A., 2000: Charactéristiques de la végétation et de la flore de la savane soudanienne de
Côte d’Ivoire: Cas des alentours de la ville de Korhogo. DEA. Univ. Cocody.7. Trochain, J-L., 1957: Bull. Inst. Centrafrique N. S., Brazzaville, 13–14: 55–93.

136
Project ID: 01LC0017 (BIOTA West Africa W06)1.1.2001 – 31.12.2003
DETERMINANTS OF SMALL-SCALE MOSAICS OF ARTHROPODCOMMUNITIES IN NATURAL AND ANTHROPOGENICALLY DISTURBEDHABITATS
Karsten Mody, Christian Kost, and K. Eduard Linsenmair
Lehrstuhl für Tierökologie und Tropenbiologie, Universität Würzburg, Biozentrum, Am Hubland,97074 Wü[email protected]; http://www.biozentrum.uni-wuerzburg.de/zoo3/
Key words: animal-plant-interactions, arthropod communities, community structure, ComoéNational Park, Dorylus, local processes
Abstract:The diversity and structure of ecological communities is determined by many parameters. Inorder to understand the influence local processes exert on the maintenance of biodiversity innatural and anthropogenically altered environments, two arthropod systems are studied in theWest African Comoé National Park. In arthropod communities living on individual savannashrubs and trees, the role of deterministic small scale variation in plant and habitatcharacteristics for community composition is investigated. Repeatedly sampled, the arthropodcommunities of individual shrubs and trees were distinctive even for conspecific plants growing inclose neighborhood. Thus, each plant was inhabited by a unique, characteristic set of arthropods.This striking pattern suggested a community organization governed rather by deterministic, smallscale environmental variables than by stochastic events. Focal observations on several arthropodspecies belonging to different guilds supported this interpretation. Driver ants (Dorylusnigricans) are studied as a stochastic disturbance of arthropod communities. Driver ants areconsidered to be an important, community-structuring source of disturbance especially forground living arthropod communities. In a first analysis indirect evidence is presented that driverant raids reduce the abundance of some potential prey taxa as well as that of potentiallycompeting species.
Fig.1: A: Apparatus for standardized measurement of herbivory: area of leaf samples (ls) is quantified before and afterfeeding trials using a digital camera (dc); B: Caterpillar of Chrysopsyche imparilis; C: Dorylus nigricans driver ants.
dc
lsA B C

137
Results:Repeatedly sampling complete arthropod faunas of individual plants revealed that savannashrubs and trees are inhabited by distinctive sets of arthropods which turned out to be stablefor – at least - more than a year. Such arthropod communities, unique for individual plants,could be found for trees and shrubs exceeding a certain minimum size (Fig.2). This findingcould be confirmed even for conspecific plants growing in close neighborhood. Since thespecies pool for colonization is identical for these plants, it has to be concluded thatdeterministic habitat characteristics, acting on a restricted local scale, are influential oncommunity composition. Such deterministic habitat parameters could be variable plantcharacteristics like plant morphology and chemistry or environmental variables like soilheterogeneity (determined in coordination with W02 and W04), generating variation inarthropod-relevant parameters on a local scale [1]. Thus, small scale variability in habitatcharacteristics seems to be a crucial factor for diversity in arthropod communities. It may playa key role in the interpretation of similarities and differences found for regions or sitesdifferentially influenced by humans and may have profound implications for managementmeasures.
The distribution of herbivorous insects seems to reflect the intraspecific variability ofplant characteristics mentioned above. This is, as an example, shown in the distribution of themobile scarabaeid beetle Apogonia fatidica and is probably causing food plant changingbehavior of less mobile moth caterpillars. A. fatidica occurred highly aggregated on certainindividuals of the savanna tree Combretum nigricans while it was completely absent on someneighboring C. nigricans plants (Fig.3A). Feeding experiments (cf. Fig.1A) demonstrated thatthis characteristic distribution pattern was tightly coupled with leaf attractiveness whichsignificantly varied between individual C. nigricans plants for the beetles (Fig.3B).
Caterpillars are limited in choosing between different plants and in reaching the mostattractive and best suited plants. However, some caterpillars feeding on C. nigricans changefood plants regularly. This change of food plants reduces – in view of highly variable plants -the probability of using exclusively unfavorable plants and results in a diversified diet.Feeding on different plant individuals can be positively correlated with fitness which wasexperimentally shown for Chrysopsyche imparilis (Lepidoptera, Lasiocampidae)(Figs.1B&3C).
Fig.2 shows similarities in species composition of arthropod communities (excluding ants) eight times sampled from thesame and different Combretum nigricans plants respectively. It becomes clear that communities obtained from the sameplants are significantly more similar than communities from different plants when shrubs of Size Class 1 (SC1: plants with175 to 1090 leaves) are considered. No difference in species similarity of communities was detectable for small plants ofSize Class 2 (SC2: plants with 23 to 94 leaves). Paired t-test; N per size class: 14 plants; JI = Jaccard Index.
mea
n si
mila
rity
[mea
n JI
]
-0,04
0,00
0,04
0,08
0,12
0,16
0,20
0,24
same plant other plants same plant other plants
±Std.Dev.
±Std.Err.
Mean
SC1 SC2
P < 0.0001
P = 0.68
138
Fig.3 A: Aggregated distribution of Apogonia fatidica beetles on 14 Combretum nigricans plants (P1-P14) which areexemplarily presented for their regularly assessed beetle abundance. Nine consecutive collection events (Coll.) are shown.Differing beetle densities on individual plants reflect variable plant characteristics (attractiveness). B: Comparison of leafattractiveness of paired C. nigricans plants (C) to A. fatidica beetles. Each plant pair tested in the lab contains a plantindividual which attracts high beetle densities in the field (attractive Combretum = aC) and a plant individual which attractsonly small numbers of beetles in the field (unattractive Combretum = uC). Significant differences in consumed leaf mass(dw = dry weight) are indicated (paired t-test): ns = not significant; ** = P<0.01; *** = P<0.001; Box-Whisker-Plots:Central line = Mean, Box = Std. Err., Whisker = Std. Dev.; N (per plant pair): 12 leaf pairs, 120 beetles. C: Comparison ofegg numbers laid by Chrysopsyche imparilis females raised either on leaves of a single Combretum nigricans plant (single-plant food) or on leaves of a mix of six C. nigricans plants (mixed-plant food). Females raised on the mixed diet laidsignificantly more eggs. This indicates that changing food plants can be advantageous in view of variable plants and mayevidently enhance reproductive output
Plant ID
cons
umed
leaf
mas
s [m
g dw
]
-5
5
15
25
35
45
55
aC
1
uC
1
aC
2
uC 2
aC 3
uC 3
aC 4
uC 4
aC 5
uC 5
aC 6
uC 6
aC 6
uC 6
aC 3
uC 3
*** *** *** ns ** *** *** ***
0
10
20
30
40
50
60
70
P1
P2
P3
P4
P5
P6
P7
P8
P9
P1
0
P1
1
P1
2
P1
3
P1
4
Plant ID
num
ber
of b
eetle
s
9. Coll.
8. Coll.
7. Coll.
6. Coll.
5. Coll.
4. Coll.
3. Coll.
2. Coll.
1. Coll.
A
B
egg
num
ber
0
40
80
120
160
200
single-plant food mixed-plant food
n = 20
n = 23
P = 0,03 ±Std.Dev.
±Std.Err.
Mean
C
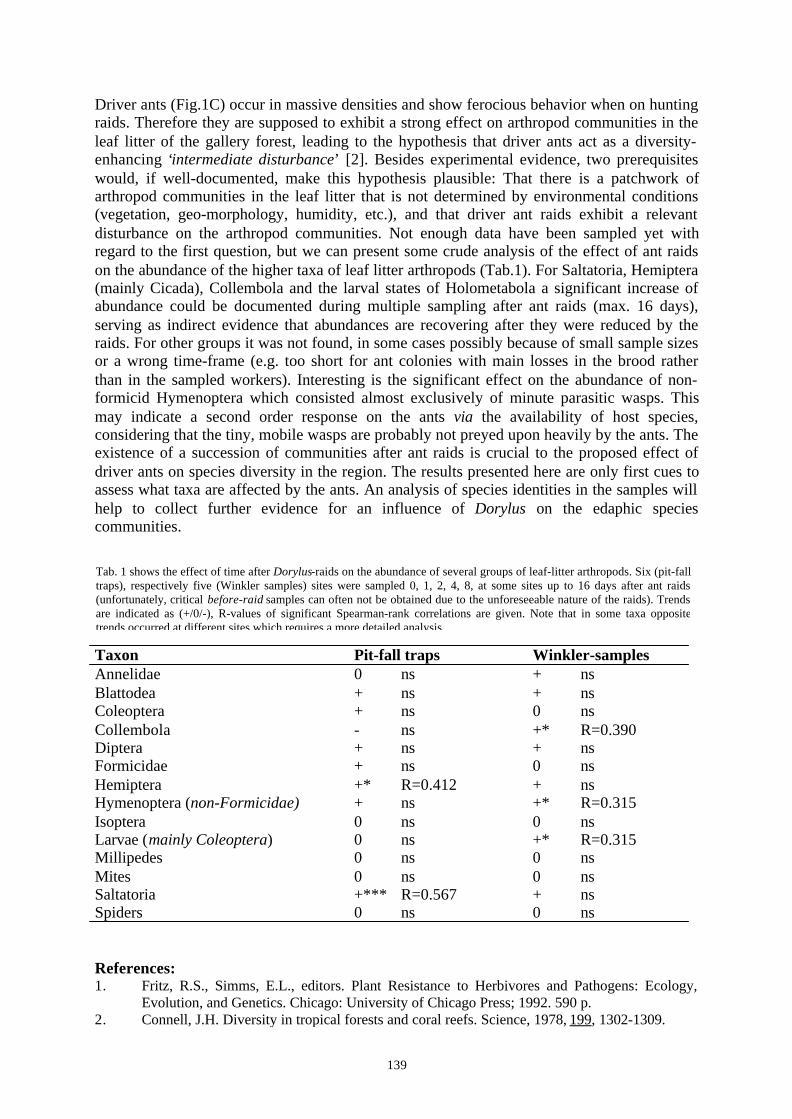
139
Driver ants (Fig.1C) occur in massive densities and show ferocious behavior when on huntingraids. Therefore they are supposed to exhibit a strong effect on arthropod communities in theleaf litter of the gallery forest, leading to the hypothesis that driver ants act as a diversity-enhancing ‘intermediate disturbance’ [2]. Besides experimental evidence, two prerequisiteswould, if well-documented, make this hypothesis plausible: That there is a patchwork ofarthropod communities in the leaf litter that is not determined by environmental conditions(vegetation, geo-morphology, humidity, etc.), and that driver ant raids exhibit a relevantdisturbance on the arthropod communities. Not enough data have been sampled yet withregard to the first question, but we can present some crude analysis of the effect of ant raidson the abundance of the higher taxa of leaf litter arthropods (Tab.1). For Saltatoria, Hemiptera(mainly Cicada), Collembola and the larval states of Holometabola a significant increase ofabundance could be documented during multiple sampling after ant raids (max. 16 days),serving as indirect evidence that abundances are recovering after they were reduced by theraids. For other groups it was not found, in some cases possibly because of small sample sizesor a wrong time-frame (e.g. too short for ant colonies with main losses in the brood ratherthan in the sampled workers). Interesting is the significant effect on the abundance of non-formicid Hymenoptera which consisted almost exclusively of minute parasitic wasps. Thismay indicate a second order response on the ants via the availability of host species,considering that the tiny, mobile wasps are probably not preyed upon heavily by the ants. Theexistence of a succession of communities after ant raids is crucial to the proposed effect ofdriver ants on species diversity in the region. The results presented here are only first cues toassess what taxa are affected by the ants. An analysis of species identities in the samples willhelp to collect further evidence for an influence of Dorylus on the edaphic speciescommunities.
Taxon Pit-fall traps Winkler-samplesAnnelidae 0 ns + nsBlattodea + ns + nsColeoptera + ns 0 nsCollembola - ns +* R=0.390Diptera + ns + nsFormicidae + ns 0 nsHemiptera +* R=0.412 + nsHymenoptera (non-Formicidae) + ns +* R=0.315Isoptera 0 ns 0 nsLarvae (mainly Coleoptera) 0 ns +* R=0.315Millipedes 0 ns 0 nsMites 0 ns 0 nsSaltatoria +*** R=0.567 + nsSpiders 0 ns 0 ns
References:1. Fritz, R.S., Simms, E.L., editors. Plant Resistance to Herbivores and Pathogens: Ecology,
Evolution, and Genetics. Chicago: University of Chicago Press; 1992. 590 p.2. Connell, J.H. Diversity in tropical forests and coral reefs. Science, 1978, 199, 1302-1309.
Tab. 1 shows the effect of time after Dorylus-raids on the abundance of several groups of leaf-litter arthropods. Six (pit-falltraps), respectively five (Winkler samples) sites were sampled 0, 1, 2, 4, 8, at some sites up to 16 days after ant raids(unfortunately, critical before-raid samples can often not be obtained due to the unforeseeable nature of the raids). Trendsare indicated as (+/0/-), R-values of significant Spearman-rank correlations are given. Note that in some taxa oppositetrends occurred at different sites which requires a more detailed analysis.

140
Project ID: 01LC0017 (BIOTA West Africa W07)01.01.2001-31.12.2003
COMPARATIVE STUDIES ON EPHEMERAL SAVANNA WATERS AND THEIRSIGNIFICANCE FOR DIVERSITY AND PRODUCTIVITY OF FISHCOMMUNITIES IN NATURAL AND ANTHROPOGENICALLY DISTURBEDRIVERS
A. Kahlheber, M. Boutros and K. E. Linsenmair
Lehrstuhl für Tierökologie und Tropenbiologie, Zoologie III, Universität Würzburg,Biozentrum, Am Hubland, 97074 Wü[email protected]; http://www.biozentrum.uni-wuerzburg.de/zoo3/
Key Words: Comoé National Park, ephemeral waters, fish communities, lateral migration, localprocesses, spatial-temporal utilization patterns
Abstract:
Freshwaters in tropical savanna biomes are currently subject to massive changes.Deterioration of water quality, elimination of whole spectra of habitats and profoundalterations in the basic hydrological conditions are especially serious consequences of humanactivities. In order to understand the natural organization of local fish assemblages and toassess the effects of anthropogenic influences on the composition and functionalcharacteristics of fish communities, the fish fauna of undisturbed savanna waters in ComoéNational Park, Ivory Coast, and in disturbed adjacent areas is investigated. The aim is todescribe the spatial and temporal use patterns of savanna waters by age classes, species andassemblages of fish and to understand the factors that are causing these use patterns. In thefirst year of the project, the main emphasis is put on surveying the composition of fishcommunities that use ephemeral savanna waters temporarily connected to Comoé river, theonly permanent water in the region. 65 species of fish have been caught in these waters so far,among them some new to the fish community hitherto reported for Comoé river. The generalcomposition of fish assemblages of ephemeral waters is comparable to the fish community ofComoé river. However, analyzing parameters of distribution more precisely, a speciesspecific, fine grained use pattern becomes evident. It points to the important role ephemeralwaters can play for many species of fish as feeding, growing or spawning sites.
Results:Our first results indicate that ephemeral savanna waters are an important fish habitat although theyare useable only for a limited time of the year. A comparison of the fish fauna of the great riversystems of Ivory Coast (Sassandra, Bandama, Comoé) and the fish assemblages of our study areashows that most fish locally occurring in permanent waters can also be found in ephemeralwaters. While 91 species of fish are reported for the whole Comoé river system (length: 1,160 km;drainage area: 72,000 km²) [1,2], 65 species of fish could already be recorded in the extremelylimited savanna waters studied so far (three 200 m sections of two ephemeral savanna rivers andfifteen ponds with a total surface area of less than 1 km²).An analysis of assemblage composition indicates that use of ephemeral habitats is notrestricted to certain groups but is to be found in a wide range of taxonomic groups of fish(Fig.1). Simply the Cyprinodontidae seem to be relatively rare in ephemeral savanna waters incomparison to the total fish fauna of Comoé river [3]. This might be caused by the existenceof many species in estuarine or rainforest waters which contribute to the total Comoé fishfauna. On the other hand, we have recorded one annual cyprinodontid as a characteristicinhabitant of ephemeral savanna ponds which previously has not been reported for Comoériver. Another group of fish, the Cyprinidae, appear, however, especially species-rich in

141
ephemeral savanna ponds and streams. With the documentation of three species, new to theComoé fish fauna, more species of cyprinids are known from the ephemeral as compared tothe river community. This finding stresses the great importance of ephemeral savanna watersto these fishes. It also points to gaps in the current knowledge of the distribution of many fishspecies in the region. As all fish - with the exception of annual cyprinodontids andProtopterus-lungfish which can survive dry seasons without permanent water as eggs orburied in mud holes - have to newly re-colonize ephemeral savanna waters from permanentwaters, i.e. from the Comoé river in our case, they have to occur also in the river itself.
Analyzing parameters of distribution and habitat characteristics more precisely (incooperation with W01, W02, W04), a species specific, fine grained use pattern of ephemeralwaters is becoming evident. In dependence of water availability, the composition of fishassemblages in savanna waters changed. On the one hand, species of Barbus, Aplocheilichthysand Hemichromis re-colonized ephemeral waters directly after first rains. They may either tryto migrate in savanna ponds far away from Comoé river or to take advantage of low predationpressure by predatory fish in newly developing waters. On the other hand, fish like Brycinusnurse, Labeo parvus or Schilbe spec. started lateral migration much later, entering savannawaters at a time when water level remained high and rather predictable and stable foodsources had been established. The different patterns in utilization of ephemeral savannawaters by a species-rich fish community point to the important role these waters can play forfishes as feeding, growing or spawning sites.
References:1. Teugels, G.G., Lévêque, C., Paugy, D. and Traoré, K. État des conaissances sur la faune
ichtyologique des bassins côtiers de Côte d`Ivoire et de l`ouest du Ghana. Revue d`Hydrobiologie Tropicale, 1988, 21, 221-237.
2. Livingstone, D.A., Rowland, M. and Bailey, P.E. On the size of African riverine fish faunas.American Zoologist, 1982, 22, 361-369.
3. Lévêque, C., Paugy, D., Teugels, G.G. Faune des poissons d'eaux douces et saumâtres del'Afrique de l'Ouest, Tome 2. Tervuren and Paris: MRAC-ORSTOM; 1992. 902 p.
Fig.1: Taxonomic composition of fish assemblages of ephemeral savanna waters (our study) and a great WestAfrican river, the Comoé (published data). Total species number for each group mentioned is given in brackets.“Others” comprise: Anabantidae, Centropomidae, Channidae, Mastacembelidae, Polypteridae, Protopteridae(for ephemeral waters) and Anabantidae, Centropomidae, Channidae, Clupeidae, Eleotridae, Mastacembelidae,Notopteridae, Polypteridae, Protopteridae (for Comoé river).
Ephemeral waters Comoé
Cyprinodontidae6% (4)
Cichlidae8% (5)
Siluriformes18% (12)
Cyprinidae26% (17)
Characiformes18% (12)
Mormyridae14% (9)
Others9% (6)
Cyprinodontidae16% (15)
Cichlidae9% (8)
Siluriformes18% (16)
Characiformes18% (16)
Cyprinidae18% (16)
Mormyridae11% (10)
Others11% (10)

142
Project ID: 01LC0017 (BIOTA West Africa W08)01.01.2001-31.12.2003
AMPHIBIAN DIVERSITY AND DIVERSITY CHANGES IN WEST AFRICA:GLOBAL, REGIONAL OR LOCAL CAUSES?
M.-O. Rödel1, 2, 3, J. Kosuch2, R. Ernst1, V. Rudolf1, G. Schorr1, C. Harbig1,S. Frohschammer1, G. Leistner1, M. Wegmann1, P.J. Adeba3, G.N. Kouamé 3, A.A.Cudjoe4, K.E. Linsenmair1 and M. Veith2
1 Lehrstuhl für Tierökologie und Tropenbiologie, Zoologie III, Universität Würzburg,Biozentrum, Am Hubland, D-97074 Würzburg, Germany; 2 Institut für Zoologie, Saarstrasse21, D-55099 Mainz, Germany; 3 Université Abobo Adjamé et Université Cocody, ProjetBiodiversité, BP 20 Taï, Côte d’Ivoire; 4 Ghana Wildlife Division, PB M 239, Accra, [email protected]; http://www.gadag.org; http://www.biozentrum.uni-wuerzburg.de/zoo3/; http://perdix.biologie.uni-mainz.de/___________________________________________________________________________
Key Words: Amphibia, bio-monitoring, community ecology, diversity, forest, Ghana, genetics,Ivory Coast, primary habitats, savanna, secondary habitats, West Africa
Abstract:Amphibian diversity along environmental and disturbance gradients in West Africa has beeninvestigated. Diversity was higher than expected and at least equals that of neotropicalregions. Habitat specific anuran communities could be characterized and basic data on lifehistory traits of possible target species for a long term monitoring program have beencompiled.
Results:As a base line for our future research amphibian diversity throughout different West Africanecotones has to be documented. So far we compiled species inventories of five national parksin Ivory Coast and the Ghanian Volta region (Fig. 1). By means of standardized samplingtechniques we obtained basic data on the amphibian community composition in differenthabitat types (forest, savanna, primary, secondary). This enables us to detect diversity changesover time, and/or along environmental and disturbance gradients.In total 96 amphibian species were recorded from Ivory Coast. From 61 species DNA sampleshave been collected and partly sequenced. For most of the species at least basic biological andecological data are now available. We were able to document several reproductive strategiesso far unknown for African anurans [4, 7], as well as unknown interactions of other groups oforganisms with amphibians [5]. By means of morphological, behavioral, acoustical andgenetic comparisons we detected five species new to science [6].Analyses of the leaf litter anuran assemblage of Taï National Park by means of niche overlapanalysis, non-metric-multidimensional-scaling, and Mantel-test comparison revealed that theobserved multivariate structure is only weakly influenced by biotic interactions and therefore mostlikely not the sole result of past or present species interactions. Results indicate that different lifehistories are likely to explain the observed pattern. In primary forests, distinct species assemblagesexisted but could be explained by geographic proximity and therefore the existence of commonlocal species pools. Assemblages in secondary forests were predictable based on environmentalparameters. This can be inferred to be the result of physiologically more restrictive conditionswithin secondary forest habitats especially due to an altered microclimate. Only highly adapted ortolerant species may be able to cope with these conditions.

143
These results lead to important implications for bio-monitoring and conservation managementand provide useful information, which is urgently needed to assist conservation efforts.Surveys of the populations of selected species may yield information about the state ofdisturbance of an area. We currently selected several anuran species for further investigations,that proved to be specialized on certain habitat types and/or show peculiar life history traitsand therewith represent different ecotypes. These comprise three species with directdevelopment that are restricted to disturbed forests (Phrynobatrachus alticola) or occur invarying abundances in primary and secondary forests (Arthroleptis variabilis, A. crusculum),as well as three species that all maintain tadpole stages but reproduce in different habitats[Phrynobatrachus guineensis: breeds in tree holes in drier parts of primary forest; Kassinalamottei: breeds in large ponds in swampy parts of primary forest (Fig. 2); Cardioglossaleucomystax: reproduces in slow running forest creeks].
Fig. 1: Similarity of West African amphibiancommunities; presence-absence data, speciesnumbers in parentheses, CI = Côte d’Ivoire,GH = Ghana, LI = Liberia, ² = [3], ³ = [1, 2, 8].
Fig. 2: Kassina lamottei an endemic frog of theUpper Guinea forest zone, confined to primaryrainforest, drawing: M.-O. Rödel.
References:1. Guibé, J. and Lamotte, M. La réserve naturelle intégrale du Mont Nimba. XII. Batraciens (sauf
Arthroleptis, Phrynobatrachus et Hyperolius). Mem. Inst. fond. Afr. noire, 1958, 53, 241–273.2. Guibé, J. and Lamotte, M. La réserve naturelle intégrale du Mont Nimba. XXVIII. Batraciens du
genre Phrynobatrachus. Mem. Inst. fond. Afr. noire, 1963, 66, 601–627.3. Lamotte, M. Les batraciens de la région de Gpakobo (Côte d’Ivoire). Bull. Inst. fond. Afr. noire,
1967, 29, 218–294.4. Rödel, M.-O. and Ernst, R. A new reproductive mode for the genus Phrynobatrachus Günther,
1862: Phrynobatrachus alticola Guibé and Lamotte, 1961 has non-feeding, non-hatching tadpoles.J. Herpetol., in press.
5. Rödel, M.-O., Range, F., Seppänen, J.-T. and Noë, R. Caviar in the rain forest: monkeys as frogspawn predators in Taï National Park, Ivory Coast. J. Trop. Ecol., in press.
6. Rödel, M.-O. and Ernst, R. A new Phrynobatrachus species from the Upper Guinean rain forest,West Africa, including the description of a new reproductive mode for the genus. J. Herpetol., inpress.
7. Rödel, M.-O., Krätz, D. and Ernst, R. The tadpole of Ptychadena aequiplicata (Werner, 1898) withthe description of a new reproductive mode for the genus (Amphibia: Anura: Ranidae). Alytes, inpress.
8. Schiøtz, A. The treefrogs (Rhacophoridae) of West Africa. Spolia zool. Mus. haun., 1967, 25, 1–346.
euclidian distance
Mt. Nimba, LI (57)³
Tai, CI (56)
Mt. Péko, CI (35)
Volta Re., GH (32)
Mt. Sangbé, CI (42)
Lamto , CI (39)²
Marahoué, CI (31)
Comoé, CI (35)
3,0 3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0

144
Project ID: 01LC0017 (BIOTA West Africa W09)1.1.2001-31.12.2003
THE INFLUENCE OF LOCAL AND REGIONAL FACTORS ON THE DIVERSITY,STRUCTURE, AND FUNCTION OF WEST AFRICAN BAT COMMUNITIES(CHIROPTERA)
Jakob Fahr, Njikoha M. Ebigbo and Elisabeth K.V. Kalko
Abt. Experimentelle Ökologie (Bio 3), Universität Ulm, Albert-Einstein Allee 11, 89069 [email protected]; http://www.biologie.uni-ulm.de/bio3
Key words : local assemblages, community ecology, guild structure, habitat heterogeneity, scaledependency, distribution patterns, regional species pools, GIS, modeling, potential distribution,frugivory, seed dispersal, plant-animal interactions, vegetation dynamics
Abstract:Using a comprehensive approach, we examine the functional significance of bats for themaintenance of biological diversity in Africa. The project goals are to determine the roles of batsin fundamental ecological processes such as seed dispersal and to assess factors that influencethe structure and diversity of chiropteran assemblages on local and regional scales. In a firstapproach we determine the composition and guild structure of selected bat communities on alocal scale (Ivory Coast) by conducting comparative field studies with standardised methodologyat two localities contrasting in vegetation composition (savanna and rain forest). In a modellingapproach, we analyse the large-scale, regional diversity patterns of West African bats based ondata of species distribution as well as biotic and abiotic habitat variables. We study the spatialpatterns of bat diversity with the ultimate goal to test underlying factors such as habitatheterogeneity and productivity that might covary with these patterns. In a third approach weexamine the role of flying foxes (Megachiroptera) as keystone species in plant-animal interactionsand their influence on the spatial and seasonal vegetation patterns within the highly dynamicforest-savanna mosaics of the Guinea zone.
Results:In our first working package we have established a standardized protocol to sample local batassemblages. We ask how individual species and entire communities are associated with differenthabitat types. Until now we have obtained long-term data for two local assemblages, one in therain forest zone (Taï-N.P.), the other in the Guinea zone (Comoé-N.P.), both of which are fairlyundisturbed. The study areas represent important examples of West African biomes that areincreasingly threatened by degradation. At present the data set for Taï-N.P. covers 2 years(1999/2000) while the one for Comoé-N.P. was initiated in 1993/95 and now embraces 3 years(1999-2001) of intense sampling, including the Biodiversity Observatory. The preliminary resultssuggest that the habitat preferences of bats are generally very species-specific, thus making themwell-suited candidates to evaluate and monitor the effects of habitat alteration through landconversion and global change. Furthermore, the structure of the assemblages (species and guildcomposition, abundance patterns) reflect the different habitat types that have been sampled. Thesedata will serve as a base-line against which the effect and magnitude of land alterations can betested in the main phase of BIOLOG. So far, a total of 53 species which represent 84% of theestimated 63±4 species (calculated with EstimateS, © R. Colwell) have been found in Comoé-N.P. In Taï-N.P. we have recorded 37 species, representing 80% of the expected 46±2 species.These preliminary data support our initial assumption (see BIOTA application) that the diversitypeak of bats in West Africa is not found in the rain forest zone but in the adjacent forest-savanna

145
mosaic of the Guinea zone. This diversity peak is caused by the exceptional habitat richnesswhich fosters an influx of species from habitats adjacent to the ecotone. The species richness ofComoé-N.P. by far surpasses all other African bat communities that have been studied in the past.It underlines our general hypothesis [1] that African bat communities are not depauperate asproposed by FINDLEY [2] but attain levels comparable to the Neotropics. We conclude that habitatdegradation – especially in highly dynamic mosaic systems – is likely to negatively affect speciesrichness, composition and community pattern, potentially altering the function of dependentecosystem levels. This latter question will be addressed in the main phase of BIOLOG.
While our first working package focuses mainly on the local factors determining bat assemblages,we investigate in our second working package the spatial patterns of bat diversity on a regional /subcontinental scale and its interdependency with the local scale. For this approach we model thehabitat preferences and potential distribution patterns of all bat species occurring in West Africa.To build species-specific habitat models, the known occurrences of each species (verified pointlocalities only) are superimposed onto biotic and abiotic variables (vegetation, topography,climate) already implemented in the GIS of BIOTA-subproject W01. By superimposing themodelled distribution patterns we identify the regional diversity patterns and regions withexceptionally high species richness and / or endemism (hot spots). We have already established adata base which comprises all known point localities of West African bat species (121) that havebeen published (ca. 470 publications) and where specimens (about 20.000) are available inmuseum collections in Europe and North America. It is the most comprehensive data set for anygroup of African vertebrates. The modeling approach will allow us to overcome many of thesampling biases by building species-specific habitat models and extrapolating them into space.For this working package we are collaborating closely with BIOTA-subprojects W01 and W03.
Finally, in our third working package we study the role of flying foxes on the vegetation dynamicsof the Guinea savanna by determining their influence as seed dispersers. We investigate theirimportance in detail by focusing on the tree species Cola cordifolia (Sterculiaceae) and Miliciaexcelsa (Moraceae). First results indicate that both species are dominant elements of island forestsbut with varying densities and/or patchy distribution within these forests. This pattern might becaused by the activities of highly mobile seed dispersal agents. We therefore test the assumptionthat flying foxes significantly influence seed distribution, having a high potential for long distancedispersal. Direct observations at three fruiting tree species (C. cordifolia, Syzigium guineense andUapaca togoensis) suggest that flying foxes mainly feed by shuttling between resources andfeeding roosts. In an island forest, dominated by C. cordifolia, a high number of feeding roosts(46) with seeds of this species could be identified within a small area of 2 km². The distribution offeeding roosts appears to be concentrated in areas of least fruiting density. Although initial seeddeposition is high, germination rate appears to be moderate yet yielding a high density ofseedlings per area. Surprisingly, very few older age classes could be found so far. In closecooperation with BIOTA-subproject W04 we have established ten 100 m² plots in whichseedlings will be individually marked and monitored over the next years to determine seedlingestablishement, survival, and the resulting spatial pattern. In addition, we measure seed rain aswell as seedling density along transects oriented away from parent trees to assess distance effects.
1 FAHR, J., 1996. Die Chiroptera der Elfenbeinküste (unter Berücksichtigung des westafrikanischenRaumes): Taxonomie, Habitatpräferenzen und Lebensgemeinschaften. Diplomarbeit, Julius-Maximilians-Universität Würzburg. 204 pp., 36 Karten.2 FINDLEY, J. S., 1993. Bats - a Community Perspective. Cambridge University Press, Cambridge.

146
Project ID: 01LC0017 (BIOTA West Africa W11)1.1.2001-31.12.2003
PHYTODIVERSITY IN THE WEST AFRICAN SAHEL AND SUDAN ZONE –DEVELOPMENT AND EVALUATION
A. Akoegninou5, W. Andres³, T. Bierschenk4, S. Dressler², K. Hahn-Hadjali, K. König³,K. Neumann³, N. Schareika4, M. Schmidt², B. Sinsin7, A. Thiombiano6, R. Wittig, G.Zizka².1 Botanisches Institut, Universität Frankfurt. Email: [email protected]² Botanik/Paläobotanik, Universität Frankfurt und Forschungsinstitut Senckenberg,³ Institut für Physische Geographie, Universität Frankfurt4 Institut für Ethnologie und Afrika-Studien, Universität Mainz5 Faculté des Sciences et Techniques, Université Nationale du Bénin,6 Unité de Formation et de Recherche en Sciences de la Vie et de la Terre, UniversitéOuagadougou7 Faculté des Sciences Agronomiques, Université Nationale du Bénin, Bénin
Key words: Phytodiversity, land use and changes in phytodiversity, ethnobotany,archaeobotany, Poaceae, digitalization of herbarium collections.
Abstract:In the Sahel and Sudan zone of Burkina Faso and north Benin,biodiversity observatories according to BIOTA-standards,based on satellite images and on observations in the field havebeen selected and established (Fig. 1). For each of the 3principal regions (representing Sahel, North and SouthSudanian zone), a pair of plots was selected, differingprincipally in the intensity of land use, especially cattlegrazing. A floristic inventory as well as relevés of the subplotswere realized during field work. The ethnobotanical studiesconcentrated on the perception/observation of changes inflora and vegetation by local people and knowledge aboutrelevant species. The field work was performed in close co-operation with the local counterparts (see above). Therequired contracts with the relevant local institutions andcounterparts where signed, serving also as research permits. The program BRAHMS wasinstalled on a new computer (financed by the project) in the national herbarium at UniversityOuagadougou, BF. Digitalizing of collections was started and further training of staff is totake place at the end of 2001.
Results:As the field campaign will only finish in the beginning of November and the diversity dataacquired (inventories, reléves etc.) have not been analyzed yet in detail, the following resultscan only be very preliminary and fragmentary. BIOTA W11 is an interdisciplinary researchproject. Its objective is the analysis and evaluation of recent and historical changes in plantdiversity following a climatic gradient and varying land-uses. Better foundations forsustainable utilization concepts and protection measures will be developed. A furtherobjective is the extension and cross-linking of West African plant collections to make theanalysis of collection data more efficient. The project is to be executed in co-operation withcounterparts at the Universities of Ouagadougou/Burkina Faso and Cotonou/Bénin and withthe SFB (collaborative research project) 268 „Westafrican Savanna“ of the UniversityFrankfurt/Main.
Ghana
MaliNiger
Elfenbein-küste
Burkina Faso
Nigeria
Togo
0 250 500 km
Bobo-Dioulasso
Fada N'Gourma
Gorom-Gorom
Cotonou
Natitingou
Benin
Ouagadougou
Fig. 1. = Study sites

147
Especially the contacts, installations and scientific results of the SFB 268 and the excellentco-operation with the counterparts were of great help to deal effectively with the authorities aswell as to locate and design the study plots for the BIOTA project.For the selection of the plots, besides studies in the field satellite images proved to be highlyinformative and helpful. For each of the 3 principal regions (representing Sahel, North andSouth Sudanian zone), a pair of biodiversity observatories were selected. Floristic inventoriesof vascular plants and relevés of subplots were realized. In co-operation with theethnobotanists, the kind, extent and duration of human impact on the study areas weredetermined. The plots of each pair differ principally in the intensity of cattle grazing. Ofspecial interest is the fact, that we were authorized to establish one observatory in a reservestrictly protected from grazing and agriculture since about 50 years. Designation andinvestigation of plots took place in close co-operation with the scientific counterparts (seeabove). PhD students from Burkina Faso and Benin are actively involved in the project andpartly also participated in the first field campaign.The increasing intensity of grazing and agriculture have led to changes in the flora andvegetation also realized by the local people [1], [2]. As expected, alpha-diversity alone cannotserve to describe the changes: While species diversity seems to be lower in the least disturbedhabitats than in the moderately disturbed ones, the changes are considerable as far as the grasslayer is concerned. Besides the overall phytodiversity studies in the plots, special topics arethe diversity changes in selected groups assumed to be of special indicator value. While thecounterparts will in part concentrate on the woody plants, we will in the first period payspecial attention to the Poaceae. Height and structure of grass layer as well as speciescomposition vary considerably according to type and intensity of land use. Striking is thedecrease of abundance and diversity of perennial Poaceae and increase of annual grasses withincreasing grazing pressure [3], [4]. Dynamics in the distribution area of these indicatorgroups are to be detected by the investigation of (especially historical) herbarium collections.As part of this analysis, relevant material of the selected indicator groups in the herbariumcollections in Berlin (B) and Brussels (MEISE) were digitalized, and the entering of thesedata in a BRAHMS database has begun. In order to facilitate the access to and exchange ofdiversity data, the national herbarium at University Ouagadougou was provided with a newcomputer which was installed together with a version of the program BRAHMS. This is toserve the digitalizing of the herbarium collections to facilitate access to and exchange ofphytodiversity information. The same program has already been established in the course of aDutch project in the national herbarium of Benin in Cotonou. The training of the staff inBRAHMS has commenced in October and will be continued in December 2001.For the study of historical changes of the Sahelian phytodiversity, samples of charred woodfrom the Sahel of Burkina Faso were identified, recovered from 20th century archaeologicalsites. These include a large number of species, some of which have become rare in themodern flora [5]. Comparison with the modern woody vegetation is done at present.References1. Hahn-Hadjali, K. & Thiombiano, A. (2001): Perception traditionelle des espèces en voie de disparition enmilieu gourmantché: causes et solution.- Ber. Sonderforschungsbericht 268, Bd. 14: 285-298.2. Schmid, S., Braun-Yao, M., Hahn-Hadjali, K. & Werthmann, K. (2001): 10 ans plus tard: La situation socio-économique et écologique dans l´ancien projet d´installation de migrants la province de la Bougouriba (BurkinaFaso).- Ber. Sonderforschungsbericht 268, Bd. 14: 535-542.3. Wittig, R., Hahn-Hadjali, K. & Thiombiano, A. (2000): Besonderheiten von Flora und Vegetation der Chaînede Gobnangou im Südosten Burkina Fasos.- Etudes flor. veg. Burkina Faso 5: 49-64.4. Wittig, R., Hahn-Hadjali, K., Krohmer, J. & Müller, J. (2000): Nutzung, Degradation und Regeneration vonFlora und Vegetation in westafrikanischen Savannenlandschaften.- Ber. R.-Tüxen Ges. 12: 263-281.5. Neumann, K. (1999): Charcoal from West African savanna sites: questions of identification and interpretation.In: van der Veen, M. (ed.): The Exploitation of Plant Resources in Ancient Africa. New York 1999 (Kluwer /Plenum Publishers): 205-220.

148
Project ID: 01LC00221.1.2001 - 31.12.2003
THE CUVETTE CENTRALE AS RESERVOIR OF MEDICINAL PLANTS
Barbara Fruth
Max-Planck-Institut für Verhaltensphysiologie, Wicklerhaus 7, D-82319 [email protected]; http://www.mpi-seewiesen.mpg.de/~knauer/fruth/fruth.html
Key words: flora, Congo basin, Cuvette Centrale, medicinal plants, ethnobotany
Abstract:
We wish to conduct a long-term project on plant diversity in the central region of the CongoBasin, the Cuvette Centrale, Democratic Republic of Congo. During the initial phase (3years) we seek to establish the personnel and infrastructural foundations. This implies theproject-oriented education and training of technicians and young academics at the Universityof Kinshasa as well as the collection and determination of the flora of this area with specificfocus on natural medicinal plants. In order to characterise the study area in terms of it's plantdiversity and abundance, parameters such as measures of size (dominance), numbers(density) and distribution (frequency) will be investigated. In addition, the historical andcurrent significance of natural herbal plants for the medication of both humans and animalsand their phytochemical characteristics will be documented. High priority will be given to theevaluation and projection of possibilities of economic use of natural herbal plants within andby the country of origin.
Results:
In the initial proposal, data collection was planned to be conducted at Basankusu (see figure),Province of Equateur. This area was occupied by rebels opposing the official governmentbased in Kinshasa and thus became inaccessible for scientific research. Therefore, weexplored other areas within the central Congo basin and decided to establish a new site forfield research situated at the south-western border of Salonga National Park (see figure),Province of Bandundu. The Institut Congolais pour la Conservation de la Nature (ICCN) hasbeen requested to grant permission to conduct field research within Salonga National Park andthe contract is now ready to be signed by the Minister of Environment.During a pilot study in spring 2001, 15 transects (200 x 10m) were established for habitatcharacteristics using methods established during earlier studies (Boubli et al. submitted).Along these transects, 170 specimens of wild plants were collected. Reference copies of thiscollection were stored in the herbaria of Kinshasa and Brussels University for identification.Out of these, 42 species are already identified. About 50% of these plants are reported to beused as medicinal plants by the local population (Bakundu).Interdisciplinary cooperation with leading officials of Kinshasa University, environmentalorganisations and NGOs was established. By the end of 2001 a small crew of graduates willbe trained and ready to start data collection in the upcoming year. A systematic investigationof both quantitative and qualitative data in the field will put light on the floristic diversity ofthe area and filter out plants of potential pharmacological interest. Goal is to obtain base-line
149
information for long term data collection and to present a comprehensive documentation ofthe flora in form of an archive of plant taxonomy accessible to the public that will be backedup by specimens in both Congolese and European herbaria. The historical and currentsignificance of the recorded medicinal plants for both humans and animals will beinvestigated and presented as monographs.
References:
1. Boubli J.P., Eriksson J., Wich S., Hohmann G. & B. Fruth. submitted. Forest HabitatCharacteristics in the Lomako-Yekokora Interfluvium, Democratic Republic of the Congo.

150
Project ID: 01LC0025 (BIOTA East Africa E01)1.04.2001-31.03.2004
BIOTA EAST AFRICA:BIODIVERSITY IN CONVERSION – THE INFLUENCE OF FRAGMENTATIONAND MAN-MADE EXPLOITATION ON THE BIODIVERSITY OF TROPICALMONTANE FORESTS
C. M. Naumann, J. Köhler
Zoologisches Forschungsinstitut und Museum Alexander Koenig, Adenauerallee 160, 53113Bonn, [email protected], [email protected], www.biota-africa.org, ww.museumkoenig.de
IntroductionBIOTA East Africa links a set of thematically and geographically strictly coordinatedanalyses of biodiversity conversion in the East African highland and montane rain forests.Analyses of biodiversity and its change are carried out mainly at Mt. Kenya and Kakamegaforest (Fig. 1), Kenya. The latter represents the eastern-most branch of the Guineo-Congolianrain forest block, situated at 1400 to 1700 m a.s.l. in western Kenya. The area has protectedstatus as National Forest Reserve and includes both primary and secondary rain forest, as wellas several isolated fragments. It is perfectly suited to conduct comparative analyses ofbiodiversity within the mentioned habitat types. The efforts are centred on the comparison ofprimary and secondary highland rain forests with small isolated fragments. The developingopen area habitats (farm land, grazing areas, waste lands etc.) and the communities formingwithin them are included. Comparative studies are carried out in other forest regions inKenya, Tanzania and Uganda.In addition, five representative palaeo-african refugial areas have been selected to investigateplant diversity and structure of xero-tropical montane communities in southern Yemen and onSoqotra. obtained ecological data will help to estimate the present state of desertification andto develop management plans for the preservation and sustainable use of unique xero-tropicalcommunities.
Scientific goals of BIOTA East Africa
Principal goal is the establishment of biodiversity observatories for long term monitoring,with main focus on the effects of man-made changes in biodiversity. To achieve this aim,significant interfaces of the trophic network of the tropical rain forest and its replacementcommunities have been critically selected: plant-pollinator systems, forest fragmentation andseed dispersal, exchange of atmospheric compounds, regeneration of tree species, decline inanurans, diversity of coprophagous beetles, dragonflies, and Lepidoptera. The use of modernremote sensing techniques and the collection of local and historical information onenvironmental change will be used to establish a local information system in the study areas.Experiments are carried out using standardised plots and sampling methods. The potential offorest and savannah ecosystems for sustainable land use are investigated. As a result, weexpect a better understanding of the complex consequences of evolutionary change on thedifferent hierarchical levels resulting from degradation and fragmentation, especially at thelevel of critical trophic and reproductive interfaces. In addition, we expect to be able toprovide useful tools and methods for rapid assessment procedures for selected systems.

151
Steps of the first monthsThe BIOTA East Africa project started in April 2001 with preparations for the Kick-Off-Tripto Kenya which took place in May and June 2001. On this trip, all necessary formalities toreceive working and research permissions from Kenyan governmental officials were arrangedand numerous scientific cooperations with local and international institutions were establishedthrough signing bilateral Memorandums of Understanding (MoU) and contracts. At the sametime, we took the first steps towards an establishment and improvement of research logistics(field equipment, vehicles, research station, weather station, field assistants) and identifiedcertain areas to which the BIOTA East Africa research design should be applied. As a resultof these activities, all involved subprojects already conducted their first campaign of fieldresearch and data sampling (mainly at Kakamega Forest and Mount Kenya).
Established CooperationsBeside the network of internal BIOTA cooperations, BIOTA East Africa established thefollowing cooperations with local and international counterparts (e.g. exchange of data, jointeducation and training of students/para-taxonomists):
• National Museums of Kenya (NMK), Nairobi• Kenya Wildlife Service (KWS), Nairobi• International Centre for Insect Physiology and Ecology (ICIPE), Nairobi• International Centre for Research in Agroforestry (ICRAF), Nairobi• Maseno University, Maseno• Kenyatta University, Nairobi• Makerere University, Kampala• EAFRINET (East African LOOP of BioNET International)• African Pollinator Initiative (API)
In addition, internal communication with BIOLOG biodiversity informatics projects (e.g.EDIS, DORSA) revealed the necessity of linking voucher collections obtained throughBIOTA research with a sustainable data management for efficient use and long-termaccessibility of information.
Fig 1: satellite image of western Kenya, including the Nandiescarpment and Kakamega Forest, the main study site ofBIOTA East Africa.

152
Project ID: 01LC0025 (BIOTA East Africa E02)1.4.2001 – 31.3.2004
IMPACT OF BIODIVERSITY CHANGE ON THE EXCHANGE POTENTIAL OFTRACE GASES BETWEEN ECOSYSTEMS AND ATMOSPHERE
A. Awiti3, G. Schaab1,2, M. Walsh3, G. Strunz2, R. Steinbrecher1
1) Fraunhofer-Institut für Atmosphärische Umweltforschung (IFU), Kreuzeckbahnstr. 19, 82467Garmisch-Partenkirchen, Germany2) Deutsches Zentrum für Luft- und Raumfahrt (DLR), 82234 Wessling, Germany3) International Centre for Research in Agroforestry (ICRAF), Kisumu, [email protected]; http://www.ifu.fhg.de
Key words: surface fluxes, atmosphere, trace gases, VOC, agroforests, GIS, remote sensing, geo-spatial data
Abstract:Tropical ecosystems produce a variety of trace gases playing decisive roles in the atmosphere bytriggering the oxidation power as well as the warming potential. Despite their relevance for theregional and global atmosphere a sound understanding of the effect of tropical agroforest-ecosystems, showing different grades of plant diversity, on trace gas emission is lacking. To closethis gap, interdisciplinary research enterprise is necessary by coupling plant and soildocumentation with a satellite imagery-based geographic information system supplemented byecophysiological studies related to the production of the trace gases and canopy modelling. Newairborne surface flux measuring platforms will be developed and used to investigate the airsurface exchange of trace gases of ecosystems of different biocomplexity. For up-scaling from theplot scale to regional scale ground as well as airborne measurements will be used to develop atrace-gas-surface-flux model for the different land use types. The proposed activities will furtherserve to document the effect of anthropogenic activities on the biodiversity of tropical forestecosystems as well as on the chemical composition of the tropical atmosphere, taking theKakamega Forest area and its change over the last thirty years as example.
Results:(1) General tasks• For an effective project performance, cooperations with institutions in Kenya (ICRAF,
ICIPE, ILRI, NMK, DRSRS, LPR) plus setting-up of geo-spatial data sharing agreements(with ICIPE, ICRAF) supplementing memoranda of understanding were arranged. Thecontract negotiation procedures with the BMBF, DLR and ICRAF have been finalised.
• During a first visit o the test area possible experimental plots were identified by overflyingKakamega Forest and the surrounding.
(2) Development of an airborne surface flux measurement platform• For airborne surface flux measurements a platform with a 5-holes-probe coupled with a
relaxed eddy accumulation device recently developed and quality assured is underconstruction.
(3) Experiments at the Kakamega Forest site• A replicated spatial analogue survey designed to assess the impacts of forest conversion
on soil productivity (greenhouse bioassay), carbon and macronutrient (C, N, P, K, S,

153
CEC), stocks soil physical properties (bulk density, water stable aggregates, texture) hasbeen completed. One hundred geographically paired sites contrasting primary/secondaryforest with subsistence maize agriculture were surveyed across the Kakamega and SouthNandi forest ecotones in Jul-Aug. 2001. All sites were georeferenced. Diffuse reflectancemeasurements (350-2500 nm range) and physicochemical analyses of topsoil samples areunderway. A selected number of samples will also be analyzed for 210Pb (210-Lead) and137Ce (137-Cesium) inventories, to contrast soil erosion-deposition rates betweenagricultural and forest sites.
• A protocol for rapid assessment of soil productivity was tested in Kisumu. Topsoilsamples were screened in a pot study using maize as an indicator species. The spectral andphysicochemical analysis of plant and soil samples is still underway.
• A meteorological station has been set up in the Kakamega forest and is operating since mid ofSeptember.
(4) Setting-up of the BIOTA-East-GIS• First GIS-application in order to support the BIOTA-East kick-off trip to the Kakamega
Forest: combining topographic maps (1:50'000 scale), a vegetation map compiled by M.Haupt, and photographs taken from an aircraft during the IFU kick-off trip
• Intensive search for available geo-spatial data by contacting different Kenyan, Swiss andGerman institutions as well as for relevant literature, resulting in orders of relevant maps andliterature.
• Setting-up the BIOTA-East-GIS-structure considering different levels of detail: a) Africa, b)Kenya, c) Kakamega Forest and associated forest areas, d) Kakamega Forest, e) observationplots
• Integration of ready-available geo-spatial data sets of African or Kenyan extent from DCW(PSU), DLR (Oberpfaffenhofen), ESRI (Redlands), FAO (Rome), ILRI (Nairobi),UNEP/GRID (Geneva), UNEP/GRID (Nairobi), as well as of topographic maps covering theKakamega Forest and associated forest areas
• Ordering fine-resolution Landsat satellite imagery time series: 2001, 1995, 1989, 1984,1979/80, 1975, 1972/73
• Presenting the BIOTA-East subproject E02 at international conferences (see references)• Preparation and presentation of a seminar “GIS in Biological Sciences” at the NMK (Nairobi)
as a contribution to capacity building.
References:1. Schaab, G., Steinbrecher, R., Forkel, R., Junkermann, W., Strunz, G., and Walsh, M., Der
Einfluß sich ändernder Biodiversität tropischer Waldökosysteme in Kenia auf den Austauschökologisch relevanter Spurenstoffe zwischen Ökosystemen und Atmosphäre – Aufgaben undEinsatz von GIS im BIOTA-Ost-Teilprojekt E02. Poster at the Agit 2001, 4-6 July 2001,Salzburg.
2. Schaab, G., and Steinbrecher, R., Land Cover Changes and its Impact on Biogenic VOCEmission. Poster at the Global Change Open Science Conference, 10-13 July 2001,Amsterdam (Abstract: Challenges of a Changing Earth, IGBP Stockholm ,P.6.0.015, 311,2001).
3. Schaab, G., and Bernt, N., Ein GIS für Biodiversitätsforschung im Kakamega Forest(Westkenia). DFD-internal Poster.
4. Schaab, G., Comparative analyses of biodiversity in the Kakamega Forest (Western Kenya) –The use of GIS within the BIOTA-East project. Proceedings of the AfricaGIS Conference, 5-9 November 2001, Nairobi.

154
Project ID: 01LC0025 (BIOTA East Africa E03)1.3.2001 – 31.3.2004
REGENERATION OF TROPICAL MONTANE TREE SPECIES – SPATIAL ANDTEMPORAL DYNAMICS OF FEED BACK PROCESSES
H. Dalitz, D. Uster, L. Straßl, W. Musila
Department of Ecology, Faculty of Biology, University of Bielefeld, Universitätsstraße 25, 33501Bielefeld, [email protected], www.biota-africa.org
Key words: Dispersion, diversity and composition of tree species; spatial variation of canopythroughfall, nutrient elements and soil characteristics; regeneration of tree species; light climate;growth;
Abstract:The plot-based study is embedded into a research network which investigates different levels ofdisturbance/fragmentation on processes which may influence changes in biodiversity. This studyfocuses on processes which regulate the regeneration of tree species. Abiotic processes like waterand nutrient input via canopy thorughfall, biotic processes like growth of trees are intercorrelatedwith changes in canopy structure, light climate and soil properties. All these processes controllthe regeneration of trees. Necessary cooperations with other research groups investigatingdissemination, diversity of tree species on larger scales give BIOTA the change to analyse spatialand temporal changes. Close cooperations with Kenyan institutes helps to reach the aim ofobservatorial areas in East Kenya.
Results:Aims of this project can be divided into four main aspects:1. Detailled description of tree species composition, dispersion and diversity in different sites ofthe Kakamega forest in Kenya. This includes measurements of the incremental growth of treespecies measured with dendrometers, which gives the chance to analyse the influence of climateon growth of tree species over a long period of time2. Analysis of the spatial and temporal variations of rain fall and canopy throughfall as oneexample of an element flux which could influence the species composition of trees.3. Analysis of spatial variations of pH, EC, total element content and water extractable elementfraction in soils. The aim is the determination of spatial variations of soil nutrients at horizontaland vertical dimensions at different sites in the Kakamega forest.4. Analysis of the regeneration of tree species within the forest in 1 squaremeter plots.Parameters which are measured: Species composition, dispersion and diversity of tree species,incremental growth of trees, diversity and dispersion of seedlings and saplings, analysis of canopythroughfall and rain fall in high temporal resolution, analysis of canopy structure usinghemispherical photographs, analysis of soil samples; in all samples (rain water, soil samples) mainnutrient elements, pH, EC and selected trace elements are to be analysed.
All investigations in this forest are plot-based which gives us the possibility to analyse spatialpatterns. The research area is influenced by the rural population at varying levels of selectivelogging, collection of fire woods, sampling of medicinal plants and fragmentation into smallerforest compartments. This allows the analysis of processes along an desturbance gradient.

155
The main idea behind the project: Soil and forest in Kakamega have developed over a longperiod but during the last ~100 years the forest is influenced by the population. Differentlevels of influence may have caused different changes in vegetation, changes in soils. Spatialvariations in canopy throughfall, light climate and changes in soil nutrient availability mayhave resulted in changes in vegetation. All this processes are interrelated and have feed backeffects on each other. Analysing processes in water and nutrient input via canopy throughfallin dependence on the structure of the vegetation, the light climate, soil properties and treespecies regeneration can help understanding the underlying processes and can be used asbiodiversity observatorial sites of changes in climate.State of affairs (first field trip from August to November):Since project start we had fixed problems concerning research permits; initiated close
cooperations with the ICRAF soildepartment (Dr. Alain Albrecht) and withthe Herbarium and Botanical department ofthe National Museum of Kenya (Dr. J.Mutanga).Demarcation of 12 plots at four differentsites in Kakamega (GPS data available):Colobus trail, Buyangu Hill, Salazar circuit,Isecheno. Plot size 20 to 20 m (see sketsch).At 25 points per plot at 7 depths soilsamples were collected (2100 samples).Additionally 600 samples of leaf litter (twosamples per soil auger hole) were collected.In all plots tree species are determined, treeheight measured, BHD determined anddispersion measured. 198 tree now havedendrometers which allows the analysis ofincremental growth.In 6 Plots we have installed collectors forcanopy throughfall. At 12 dates we havealready collections of rain fall and canopythroughfall (daily readings).
Illustration of mainaspects of the project(ellipses), cooperationswith other BIOTA-groups (boxes) and othercooperations (roundedboxes). The project isclosley related to theDFG-project group inEcaudor, which allowscomparisions ofprocesses between CostaRica, Ecuador andKenya. In all threecountries we work withsimilar project designs
in tropical (pre-)montane rain forests.

156
Project ID: 01LC0025 (BIOTA East Africa E04)
1.4.2001-31.3.2004
INFLUENCE OF NATURAL AND ANTHROPOGENIC FRAGMENTATIONON THE VEGETATION IN MONTANE RAINFORESTS OF KAKAMEGAAND MT. KENYA
A. Althof, D. Killmann, A. Solga, E. Fischer
Institut für Biologie, Universität Koblenz-Landau, Rheinau 1, 56075 Koblenz, [email protected], www.biota-africa.org
Keywords: Kakamega Forest, Mt. Kenya, primary forest, secondary forest, vascular plants,stem epiphytes, bryophytes, lichens, Kenya
Abstract:In Kakamega Forest, 35 plots of vascular plants and 130 plots of epiphytic bryophytes andlichens have been studied in primary and secondary forest types. The primary forest stands arecharacterized by a tree layer of 25-35 m, a dense shrub layer with dominating Dracaenafragrans and a low coverage of herb layer. The secondary forests show a tree layer of usuallyless than 10 m and a dense herb layer. In primary forests, hygrophytic bryophytes aredominant on stems, while lichens are represented mainly by crustose species. Secondaryforests are characterized by the abundance of desiccation tolerant bryophytes and foliose andfruticose lichens.
Results:First field investigation in the northern part of Kakamega Forest revealed that large parts ofthe forest are degraded and primary forest stands are restricted to only a few sites. 35phytosociological relevés of vascular plant vegetation have been studied at Buyangu Hill, nearthe Colobus-Trail, along the Isiukhu-River, around the KWS-Camp-Site and in the northernKisere fragment. At Buyangu Hill and near Colobus-Trail, the forest shows rather few oldertrees but many secondary forest species. Probably there has been selective logging in the past.The upper tree layer reaches 25-30 m and the following primary forest tree species have beenobserved, e.g. Prunus africana, Diospyros abyssinica, Chrysophyllum albidum, Markhamialutea and Aningeria altissima. A rather dense shrub layer consisting mainly of Dracaenafragrans is however typical for primary forest. The herb layer generally shows only lowcoverage. Dominant taxa are Pollia condensata, Dorstenia brownii and the gap speciesDesmodium repandum. In the rather abundant gaps at Colobus-trail, additional species areBrillantaisia madagascariensis, B. vogeliana, Oplismenus hirtellus and Renealmia engleri.
The Forest in the northern Kisere fragment seems to be much more undisturbed. Herelarge trees of nearly 35-40 m including Manilkara butugi, Diospyros abyssinica, Prunusafricana could be recorded. Also a dense shrub layer of Dracaena fragrans and Acanthuseminens together with a low cover of the herb layer are characteristics for primary forest.Along the Isiukhu river typical riverine forest with dense herb layer, the abundance of gapspecies and a high bryophyte cover are typical features.
The investigated relevés in the secondary forest are characterized by an open canopyusually less than 10 m tall and a dense herb layer with a high coverage. Typical trees orshrubs for this forest type are: Rapanea melanophloeos, Acacia abyssinica, Acanthuspubescens, Polyscias fulva, Neoboutonia macrocalyx and Croton megalocarpus. In the herblayer, Aframomum zambesiacum, Achyranthes aspera, Desmodium repandum and Pavonia

157
urens are abundant. Also several terrestrial orchids like Eulophia streptopetala, Habenariamalacophylla and Disperis aphylla have been observed, which could not be recorded inprimary forest plots.
Parallel to the investigation of vascular plants 130 phytosociological plots of epiphyticbryophytes and lichens in the lower stem parts have been studied in the same areas inKakamega Forest. At Buyangu Hill and Colobus-trail, epiphytic vegetation was dominated byhygrophytic bryophytes (Hepatics like Plagiochila spp., different Lejeuneaceae, e.g.Lopholejeunea spp., Lejeunea spp. and mosses like Porothamnium spp., Neckera spp.).Lichens were represented mainly by crustose species (e.g. Lepraria ssp.). Similarobservations could be made in the Kisere fragment.
The more open habitats along the Isiukhu-River allow the occurrence of crustose andfoliose lichens as well (e.g. Sticta spp., Lobaria spp., Leptogium spp.). Among the bryophytespendant species like Pilotrichella ampullacea are dominating.
The secondary forests studied are characterized by a high diversity of foliose andfruticose lichens (e.g. Parmotrema spp., Heterodermia spp., Usnea spp., Leptogium spp.).The epiphytic bryophyte flora consists mainly of desiccation tolerant taxa like Frullania spp.,Brachymenium spp. and Orthotrichum spp..
Diversity in epiphytic relevés is much higher in secondary forest than in primaryforest. In the latter usually not more than 10 species were observed in a 50 x 20 cm plot whilein secondary forest up to 25 species occurred.
Additionally 40 relevés of epiphytic bryophytes and lichens have been recorded onMt. Kenya. The study sites were situated in the montane Olea-Podocarpus forest at 2500 ma.s.l., the bamboo-zone (Sinarundinaria alpina) at 2700 m and the Hagenia-zone at 3000 m.In contrast to Kakamega Forest the more open montane forests are dominated by large Usneaspecies. Generally natural disturbance by Buffalos (Syncerus caffer) and elephants(Loxodonta africana) occurs in all forest types.

158
Project ID: 01LC0025 (BIOTA East Africa E06)1.3.2001 – 31.03.2004
LEPIDOPTERA AS INDICATORS OF HUMAN IMPACT ON TROPICALRAINFOREST ECOSYSTEMS IN EASTERN AFRICA
L. Kühne 1, Ch. L. Häuser2, J. Holstein2
¹ Museum für Naturkunde, Humboldt-Universität, Invalidenstr. 43, D-10115 Berlin, Germany² Staatliches Museum für Naturkunde, Rosenstein 1, D-70191 Stuttgart, [email protected], [email protected], [email protected]
Key words: biodiversity, rainforest, Kenya, butterflies, moths, Lepidoptera
Objectives:Afrotropical rainforest habitats have changed rapidly due to anthropogenic influences over thelast centuries, and have become increasingly fragmented and disturbed with growing demandson land use by the local population. The effects on insect biodiversity and, in particular,Lepidoptera are poorly known, as basic faunistic data about such groups are still scarce [1,3].Diversity and subsistence of phytophagous insects like Lepidoptera depend on various abioticand biotic parameters such as microclimate, soil-characteristics, vegetation composition andstructure, and availability of specific larval and adult food resources, which all are directlyinfluenced by the different forms of historic and current land use.The project aims to describe, characterise and compare the alpha- and beta-diversity ofselected families of diurnal and nocturnal Lepidoptera in natural, semi-natural and disturbedhabitats in standardised BIOTA biodiversity observatories for the area of Kakamega Forest inwestern Kenya. This approach should allow to identify suitable sets of indicator species foreffectively assessing the relative impact of different land use patterns on insect (Lepidoptera)rainforest communities in the region. At a later stage, it is intended to expand the obtainedresults by comparative studies planned at Côte d’Ivoire (Comoe N.P.) and in combinationwith data from related BIOTA projects (E05, E07, E09, E10) to become applicable fortropical rainforest ecosystems throughout eastern and western Africa.For the starting phase of the project, the focus is to refine methodological standards for therequired field surveys, including techniques for recording, determination of taxa, and dataanalysis. For the entire duration of the project, the following steps are considered:
• Inventorying the Lepidoptera diversity (8-10 families) for selected plots• Characterisation of the Lepidoptera community for every selected plot, in relation to past
and present human impact• Comparison of Lepidoptera communities between different habitats, in relation to abiotic
and biotic parameters and human disturbance factors• Identification of indicator species for different types of forest habitats / land use impact• Development of assessment schemes for evaluation of land use patterns on the current
state of insect biodiversity in (East) African forest ecosystems
For this project, it is necessary to rely on comparatively well known groups. The followingLepidoptera families have already been identified as promising candidates in Eastern Africa:Papilionidae, Pieridae, Nymphalidae, Saturniidae, Sphingidae, and Arctiidae [2]. In addition torecording Lepidoptera, soil samples will be taken for all sample plots and jointly analysed(BIOTA E02, E03) to recognise historic changes in land use patterns.

159
Methods (field surveys):Standard protocols and techniques for recording and monitoring Lepidoptera diversity arerefined to be used in different forest habitats [4]. Intense fieldwork will be carried out at theselected plots/observatories twice in a year during a period of 4-6 weeks, around thebeginning and the end of the rainy season, which will later on be supplemented by monitoringactivities throughout the year.For recording diurnal Lepidoptera, transect walks will be conducted periodically (three timesa week) at definite times during daylight. All butterflies seen within the bounds of the trailand within a distance of 5 m ahead of the observer are recorded. For recording fruit feedingNymphalidae, baited traps will be used additionally along the transect routes.Nocturnal Lepidoptera are recorded by light using both standardised automatic (15 W) as wellas manual light traps (125 W) (figs. 1, 2). Recording occurs three times a in periods rangingfrom 3 hours to the entire night. All species of the selected Lepidoptera families are recordedquantitatively, and individual records will be stored in a central database for further statisticand biometrical analysis of diversity.Furthermore, small soil samples (100 ccm) taken from homogenous patches within theobservatories will be analysed for physical properties and chemical composition followingstandard procedures.
Fig.1: automatic light trap Fig.2: manual light trap ('tower')
References:1. Fermon, H., Waltert, M., Larsen, T.B., Dall’Asta, U. & Mühlenberg, M. 2000: Effects of forest
management on diversity and abundance of fruit-feeding nymphalid butterflies in south-eastern Côted’Ivoire. - Journal of Insect Conservation, 4: 173–188.
2. Larsen, T.B. 1996: Butterflies as indicator species in Africa - Presidential address. - TropicalLepidoptera News, 3: 1–4.
3. Mey, W., Nuß, M. & Speidel, W. 1998: Data about Lepidoptera diversity in the Afromontane region ofMalawi (Insecta, Microlepidoptera and Heterocera). - Memoires de la Société Royale Belged'Entomologie, 38: 155-170.
4. Pollard, E. & Yates, T.J. 1995: Monitoring Butterflies for Ecology and Conservation. – Chapman &Hall, London, 274 pp.

160
Project ID: 01LC0025 (BIOTA East Africa E07)01.05.2001-30.04.2004
DIVERSITY AND SPECIES COMPOSITION OF ODONATA AS INDICATORS OFBIOTOPE QUALITY OF EAST AFRICAN RAIN FORESTS AND THEIRREPLACEMENT COMMUNITIES
V. Clausnitzer
Geographisches Institut, Universität Marburg, Deutschhausstr. 10, 35037 Marburg, [email protected], www.uni-marburg.de, www.biota-africa.org
Key words: dragonflies, Odonata, biodiversity, Kenya, Tanzania, Uganda, wetlands, coastalforests
Abstract:The principal aim of this project is a comparative study of ecology, diversity and biogeography ofdragonflies (Insecta: Odonata) in primary, secondary and fragmented rain forests and differentwetland habitats in East Africa. To achieve this an identification key for the dragonflies ofeastern Africa is prepared and distribution data as well as data on species specific habitatrequirements are collected. These will be used for further applied studies, e.g. the development ofindicator systems for environmental disturbances.
Results:Utilising both, aquatic and terrestrial habitats, the large, predominantly diurnal dragonflies,who can be readily observed and in most cases easily identified in the field, can contributemuch to the evaluation of environmental quality. They are known to be very sensitive tostructural habitat quality and thus can be a valuable tool to evaluate landscape degradationand have been used as indicator species. The amphibious larvae of dragonflies are critical inregard to water quality and aquatic morphology of streams. The adults are sensitive to habitatstructure and are excellent indicators of river disturbance, e.g. changes in habitat structure.Within this project a throughout inventory of East African dragonflies in various habitats andthe collection of species specific ecological data is aimed at.
Current activitiesData collection has been started in different areas in Kenya, Uganda and Tanzania. In mostareas visited, new records for the could be made and a lot of data on various ecologicalaspects could be collected. The PhD student John Joseph Kisakye, (Makerere University,Kampala, funded through the BIOTA programme) works on dragonflies in different forests.In cooperation with a GEF project in southern Tanzania it is planned to have Master's projectson the impact of habitat degradation on dragonflies from 2002.
Coastal forests in KenyaDuring the last year coastal forests of Kenya were surveyed to a large extend. Coastal forestsare listed as important areas in terms of conservation for East Africa [1] and are a majorcentre of endemism in Africa [2, 3]. Coastal forest areas of Kenya and Tanzania are theprimary habitat for a number of highly localised dragonfly species (endemics and/or of uniquetaxonomy). Detailed studies were carried out on the ecology of the two dendrolimneticspecies Coryphagrion grandis and Hadrothemis scabrifrons. Dragonfly communities relativeto different habitat types from indigenous forests to cultivated landscapes were described.

161
Most of the forest species are confined to coastal forests of East Africa, being stenotopic andhighly sensitive to disturbance. With increasing habitat disturbance eurytopic species whichare common and widely distributed in Africa colonise the habitats. The species assemblagesbetween different habitat types (stream, swamp, pool) in the disturbed landscape are identical,the β-diversity being very low, although the diversity of single localities may increase afterhabitat disturbances.
Publicity workSpecies check-lists for several National Parks and other protected areas of Kenya have beenprepared and were presented to the corresponding authorities. Information boards on the localdragonflies were prepared for some visitor centres (Saiwa Swamp NP, Mt. Elgon NP,Arabuke Sokoke Forest, Nairobi NP). For 2002 a small dragonfly workshop is planned in theInstitute of Environmental and Natural Resources, Makerere University, Kampala.
CooperationsDragonflies are included in a Wildlife Conservation Society (WCS) project in Tanzania"Southern Highlands Conservation Programme" and in a DANIDA funded project on IBA’sin Uganda. Within all projects national reference collections are build up and local scientiststrained. First negotiations have started with Ethiopia to include dragonflies in a plannedinventory of insects. Together with BIOTA S08 species assemblages in arid habitats,colonisation strategies in seasonal habitats and population genetics of species with apanafrican distribution and different dispersal types are comparatively studied.Odonatologists working in Africa are linked through the recently started PHAON (Pinhey'sHeritage African Odonata Network), which proves to be an excellent tool to exchange viewsand data, discuss results and present projects. In cooperation with other scientists an atlas forAfrican Odonata and a mapping programme are long term aims.
PublicationsThe identification key of East African dragonflies will be published in 2002 or 2003. Aregional report on dragonflies of Eastern Africa for the IUCN Odonata Specialist Group hasbeen recently prepared. At least one new species waits for description, while a number ofother projects need more field work. Following recent manuscripts could be finished:Clausnitzer, V. 2001a. Notes on the species diversity of East African Odonata, with a checklist of
species. Odonatologica 30:49-66Clausnitzer, V. 2001b. Notes on Trithemis bifida and T. donaldsoni (Odonata: Libellulidae). The
International Journal of Odonatologogy 4:179-189Clausnitzer, V. in press. Reproductive behaviour and ecology in the dendrolimnetic Hadrothemis
scabrifrons. The International Journal of Odonatologogy.Clausnitzer, V., Lindeboom, M. submitted. Natural history and description of the dendrolimnetic
larvae of Coryphagrion grandis.Clausnitzer, V. submitted. Dragonfly communities in coastal habitats of Kenya: indication of biotope
quality and the need of conservation measures.
References1. Stuart, S. N., Adams, R.J., Jenkins, M. S. 1990. Biodiversity in sub-saharan Africa and its islands -
conservation, management and sustainable use. IUCN, Gland & Camebridge2. Fjeldsa, J., and Lovett, J.C. 1997. Geographical patterns of old and young species in African forest
biota: the significance of specific montane areas as evolutionary centres. Biodiversity andConservation 6:325-346
3. Kingdon, J. 1989. Island Africa. Princetown University Press, Princetown

162
Project ID: 01LC0025 (BIOTA East Africa E08)1.4.2001 – 31.3.2004
MEASURING AND MONITORING AMPHIBIAN DIVERSITY IN EAST AFRICA:SEPARATING GLOBAL FROM REGIONAL AND LOCAL CAUSES
S. Lötters 1, J. Kosuch1, D. Rotich2 and M. Veith1
1Institute of Zoology, University of Mainz, Saarstrasse 21, 55099 Mainz, Germany2National Museums of Kenya, Herpetology Department, P.O. Box 40658, Nairobi, [email protected]; www.gadag.org; www.biota-africa.org
Key words: global amphibian decline, community ecology, global ecotypes, indicator species,monitoring, population structure, molecular genetics, standardised methodology, Kenya
Abstract:Global amphibian decline is currently a controversial issue. However, to judge upon a world-wide phenomenon needs world-wide monitoring. The Global Amphibian Diversity AnalysisGroup (GADAG) is setting up a long-term monitoring of circum-equatorial anuran communities.It focuses on ecologically similar species in comparable habitats. Under a GADAG framework,BIOTA E08 studies amphibian communities in eastern Africa (Kenya) along a transect of isolatedrainforests of different degrees of fragmentation. This research is methodologically equivalentwith analogous projects in West-Africa (BIOTA W08) and Borneo and ally coherent with otherBIOTA projects in Kenya. Amphibian diversity is measured at three different levels: thecommunity, the species, and the population level. Species for monitoring are identified, differentfield methods are tested and compared with other projects (e.g. BIOTA W08). A moleculargenetic database of African amphibians is being established and distribution data areimplemented in a BIOTA-compatible GIS.
Results:For more than one decade now, scientists have noticed tremendous population declines inamphibians all over the world. It is controversially discussed if these observations correspond to aglobal biodiversity crises – or if local or regional causes are more responsible [1, 2].The project BIOTA E08 has the intention to contribute to resolving this controversial. It is part ofa world-wide measuring and monitoring network for amphibian diversity at different levels,incorporated in the Global Amphibian Diversity Analysis Group (GADAG). This group currentlyoperates in East and West Africa under BIOTA as well as in Borneo. Additional projects arebeing added to run long-term observations of amphibian populations at the global scale after thecurrent three projects will have provided methodical standards. Only a world-wide diversityanalysis system dealing with similar methodology on comparable species (“ecotypes”) canprovide an answer to the question “global or local?” [3]. The BIOTA framework is especiallysuitable for GADAG’s goals, because of the inter- and trans-disciplinary character of BIOLOG.BIOTA E08 focuses on three different levels of amphibian diversity, the community, species andpopulation levels. The specific goals are- to long-term monitoring of amphibian communities and relative abundance of species.- to detect anuran species and populations to be monitored (indicator species)- to define monitoring standards for amphibian populations.- to analyse the diversity change of all occurring species and the demographic and genetic
development of indicator species with respect to environmental (human-caused) change.
Main study area of BIOTA E08 is the Kakamega Forest, added by four additional sites forcomparisons: Mount Elgon, Aberdares, Mount Kenya and Runda. We share the Kakamega Forest

163
with all BIOTA projects dealing with E Africa. Mount Kenya is also shared with BIOTA projectsE04 and E06. Study sites used by all three projects are already determined.
In 2001, all study sites have been surveyed for amphibians. Data from scientific collections (i.e.National Museums of Kenya, Nairobi; California Academy of Sciences, San Francisco) wereadded. We can already tentatively provide species lists for Kakamega Forest (21 spp.), MountElgon (10), Aberdares (13), Mount Kenya (10) and Runda (15). The total number of speciesfound is 39. Relationships between the localities mentioned is shown in Fig. 1. Field and museumdata are currently made GIS-compatible.
Fig. 1: Comparison of five East Africanamphibian communities, based onpresence-absence data.
Miscellaneous life history and syn-ecological data were recorded for most species (e.g. circadianactivity, vocalisations, reproductive modes, predators, prey, climatic preferences). These data areimportant to taxa that are suitable as. We preliminarily identified potential indicator species forlong-term monitoring: Hyperolius viridiflavus, H. pantherinus, Ptychadena mascareniensis,Phrynobatrachus natalensis, Xenopus borealis. They refer to amphibian ecotypes, currently beingdefined for the entire tropics by GADAG [4].
Field methods for amphibians monitoring have been tested at Kakamega Forest, includingopportunistic, quadrate and transect sampling [5]. Field methods are being evaluated, andextrapolation of species richness of communities is tested (cooperation with BIOTA W-08 and theGADAG Borneo project).
Molecular methods are used in two ways:- Molecular taxonomy: Unambiguously IDs of hardly determinable species, species
complexes and larvae through DNA sequencing. A molecular database has beenestablished. It presently harbours DNA data of 123 African amphibian taxa.
- Monitoring of genetic population characters: Genetic erosion starts prior to populationdecline. Therefore, all occurring species are currently screened to identify taxa with highgenetic variability for later long-term observations.
A GIS for amphibian distributions is currently being set-up in cooperation with BIOTA E02 tolink former and current distributions of species with geo-spatial landscape information.
References:1. Houlahan, J. E. et al. 2000. Nature 404, 752-755.2. Ross, A.A. et al. 1999. Ann. Rev. Ecol. Syst. 30, 133-165.3. Lötters, S. et al. Measuring and monitoring amphibian diversity change: separating global from
regional and local causes. Abstract, Joint Ann. Meet. Herp. League & Soc. Stud. Amphib. Rep.,Indianapolis, 27-31 July 2001.
4. Veith et al. A global view of convergence from the frog perspective. In prep.5. Heyer, W. R. et al. 1994. Smiths. Inst. Press.
3,0 3,5 4,0 4,5 5,0 5,5
Euclidian distance
Mt. Kenya
Runda
Aberdares
Mt. Elgon
Kakamega
3,0 3,5 4,0 4,5 5,0 5,5
Euclidian distance
Mt. Kenya
Runda
Aberdares
Mt. Elgon
Kakamega

164
Project ID: 01LC0025 (BIOTA East Africa E09)01.03.2001 – 31.03.2004
THE INFLUENCE OF LAND USE MODES ON DIVERSITY, ABUNDANCE ANDGUILD STRUCTURE OF COPROPHAGOUS BEETLES IN THE AFRICANFOREST-SAVANNA-MOSAIC
F.-T. Krell1,2, D. Newman1,2,3, S. Krell-Westerwalbesloh1,2, V. Mahiva4
1 Zoologisches Forschungsinstitut und Museum Alexander Koenig, Adenauerallee 160,D-53113 Bonn, Germany
2 Department of Entomology, The Natural History Museum, Cromwell Road, London SW7 5BD,U.K.
3 Department of Agriculture, The University of Reading, Earley Gate, P.O. Box 236, ReadingRG6 6AT, U.K.
4 International Centre of Insect Physiology and Ecology (ICIPE), Duduville, Kasarani, P.O. Box30772, Nyayo Stadium, Nairobi, Kenya
E-mail: [email protected], www.biota-africa.org, , www.museum-koenig.de
Key words: dung beetles, pasture, agriculture, forestry, fire ecology, community ecology, speciesdiversity
Abstract:Dung beetles (Scarabaeoidea) show their highest global diversity and abundance in Africanforest-savanna mosaics. They are responsible for the largest portion of dung decomposition and,thus, take part in the maintenance of soil fertility. The habitats of the forest-savanna mosaic aresubject to increasing man-made change and degradation. In this project we analyse changes of a-and ß-diversity, abundance, and guild structure of coprophilous beetles caused by different modesof human land use. Based on quantitative comparative data to be collected in Kenya and Côted’Ivoire we will analyse to which extent the most important influences, i.e. decrease ofautochthonous game fauna by poaching, traditional extensive cattle farming, forestry, agricultureand bush fires, affect the species composition, guild structure and abundance of coprocenosesand, consequently, soil fertility. In addition, we will investigate the historical reasons for theorigin and the maintenance of the high diversity of Afrotropical dung beetles to develop strategiesfor the conservation of this unique, diverse fauna.
Results:Dung beetles assemblages are an ideal model system for studying effects of human influences onnatural habitats because they respond to changes of vegetation structure and microclimate. Duringthe pilot period of the project (2001-2004), we are focusing on the effects of bush fires, poachingand extensive cattle farming. Bush fires are traditionally lit to clear forest areas for agriculture andplantations, and to burn savanna areas to improve extensive cattle farming and hunting.We established study sites at the border between forest and savanna in the V-Baoulé in Côted’Ivoire (near the Bringakro research station of the Centre Suisse de Recherches Scientifiques),and in the forest-savanna mosaic region (near the research station of the Universität Würzburg inthe Parc National de la Comoé). We sampled at a savanna site that is regularly used as cattlepasture, a savanna site without cattle grazing (Imperata grassland), forest islands in the grazedsavanna, in a secondary forest, a coffee plantation (burned 17 years ago) and in a yams field in theforest (freshly burned; Fig. 1). Plots of 20 m2 protected from grazing and protected from fire (Fig.2), respectively, were installed in the grazed savanna of Bringakro. Additional unburned plots are

165
chosen in the Comoé Park to be established during the next dry season.
The first samples indicate that abundance and diversity of dung beetles is lowest in the secondaryforest, higher in the open degraded areas in the secondary forest (coffee plantation and yams field)but highest in the grazed savanna. However, compared with the Parc National de la Comoé, thathas still a rich autochthonous game fauna, the species number in the grazed savanna of Bringakroseems to be lower. Baseline data on the dung beetle fauna of the Comoé Park are alreadyavailable [1, manuscripts submitted].Our prospecting trip to Kenya is scheduled for October 2001. The sampling programme inKakamega forest (different forest types and degraded areas around) will start in 2002. Also in2002, we will start sampling the third study area in Côte d’Ivoire, the Reserve d’Aboukouamekro(a former cattle farm where native large mammals have been reintroduced). Comparing dungbeetle assemblages of a region with persistent autochthonous game, fauna with an area withsecondary introduced game and with a region where big mammals are extinct and have beenreplaced by cattle herds, may indicate the relation between mammal fauna and dung beetleassemblages. To evaluate the importance of coprophagous beetles for soil fertility, we startedmeasurements of the amount of buried dung for the species responsible for the highest percentageof dung beetle biomass. Comparisons between dung beetle assemblages of geographically distantregions of the Afrotropical forest-savanna mosaic (Kenya and Côte d’Ivoire) will indicate whichof the effects of human land-use on these organisms are general and which are determined byregional parameters.In addition, we will investigate the history of the present scarab fauna of East and West Africausing phylogenetic and biogeographic analyses. This will elucidate how taxa and zoogeographicalelements react to man-made habitat changes. Comparing dung beetle faunas of regions with andwithout species-rich autochthonous large mammal faunas we will draw conclusions on theimportance of the latter for developing the extraordinary diversity of dung beetles in Africanforest-savanna mosaics. In the long term we will develop conservation strategies for maintainingdung beetle diversity, as well as an indicator system for rapid evaluation and assessment ofbiodiversity, and for estimating the degree of its endangerment.
References:1. Krell, F. and Linsenmair, K.E. 1998. Blatthornkäfer einer westafrikanischen Savanne - Vielfalt in Kot
und Aas (Coleoptera: Scarabaeoidea). Bielefelder Ökologische Beiträge 12: 36.
Fig. 1 : Yams field (Dioscorea sp.) on arecently burned secondary forest site;Bringakro, Côte d’Ivoire.
Fig. 2 : Firebreak around a 20 m2-plot ofsavanna parkland, regularly used as cattlepasture; Bringakro, Côte d‘Ivoire.

166
Project ID: 01LC0025 (BIOTA East Africa E10)1.4.2001 – 31.3.2004
EFFECTS OF HABITAT FRAGMENTATION ON PLANT-POLLINATORINTERACTIONS IN EAST AFRICAN RAINFORESTS
M. Kraemer, Th. Bergsdorf
Alexander Koenig Research Institute and Museum of Zoology, University of Bonn,Adenauerallee 160, 53113 Bonn, [email protected]; [email protected]; http://www.museum-koenig.de
Key words: pollination, plant-pollinator interactions, habitat fragmentation, Kenya
Abstract:The project analyses the influence of forest fragmentation on plant-pollinatorinteractions. First results indicate a significant difference in flower visitation ratesbetween different forest plots for the understory shrub Acanthopale pubescens.However, a clearcut pattern between continuous forest and forest fragments cannotbe shown as yet.
Background:It is widely recognized that fragmentation of natural habitats not only affects thedistribution and the abundance of organisms, but that it may disrupt the importantbiological processes that maintain biodiversity and that are of high importance for thefunctioning and the longterm existence of ecosystems. The pollination of plants byanimals represents such an integrative process - almost 100 percent of the floweringplants in tropical rainforests are pollinated by animals. The main aim of this projectis to analyse the alterations of plant-pollinator interactions caused by habitatfragmentation and the consequences for the long-term maintenance of tropicalbiodiversity.
Cooperation:The cooperation with the National Museums of Kenya and the International Centre ofInsect Physiology and Ecology, both Nairobi, were firmly established. As pollinationis a very important integrating process within and among natural communities,ecosystems and agricultural systems, the project is highly appreciated by ourcounterparts and Kenyan officials. Furthermore, a close connection to the AfricanPollinator Initiative (C. Eardley, South Africa) was established, resulting in theparticipation in the First Pan-African Pollination Workshop in February 2002.
Results:The study is carried out at the Kakamega Forest Nature Reserve, which forms theeasternmost part of the afrotropical, guineo-congolian rainforests. During the firstfield stay (June – October 2001) we established one study site in the northern part ofthe forest, and another site in the southern part, each consisting of continuous forest(Colobus, Salazar, Isecheno) and different forest fragments (Kisere and Malawa inthe north, Yala, Kaimosi and Ikuywa in the south). In order to tightly connect thepollination project to the project on seed dispersal (E11) both groups use exactly thesame study plots. Due to logistic problems, forest fragments in the eastern part of thenature reserve are to be excluded from the study.
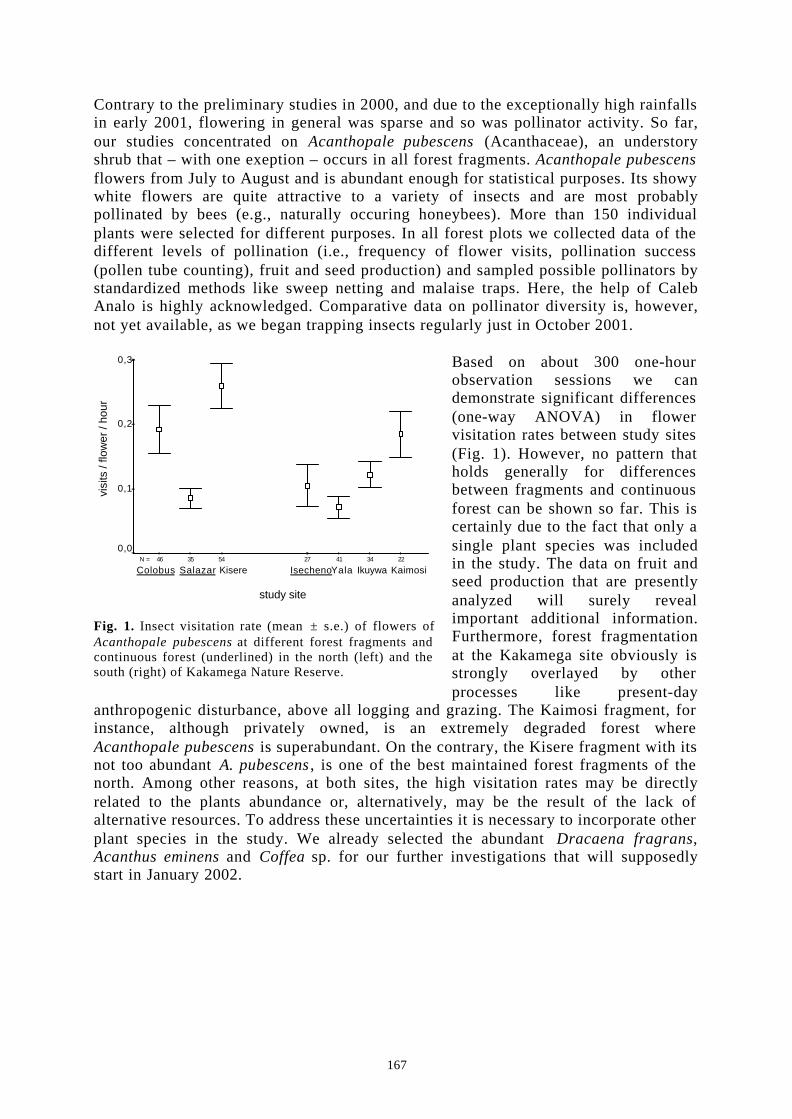
167
Contrary to the preliminary studies in 2000, and due to the exceptionally high rainfallsin early 2001, flowering in general was sparse and so was pollinator activity. So far,our studies concentrated on Acanthopale pubescens (Acanthaceae), an understoryshrub that – with one exeption – occurs in all forest fragments. Acanthopale pubescensflowers from July to August and is abundant enough for statistical purposes. Its showywhite flowers are quite attractive to a variety of insects and are most probablypollinated by bees (e.g., naturally occuring honeybees). More than 150 individualplants were selected for different purposes. In all forest plots we collected data of thedifferent levels of pollination (i.e., frequency of flower visits, pollination success(pollen tube counting), fruit and seed production) and sampled possible pollinators bystandardized methods like sweep netting and malaise traps. Here, the help of CalebAnalo is highly acknowledged. Comparative data on pollinator diversity is, however,not yet available, as we began trapping insects regularly just in October 2001.
Fig. 1. Insect visitation rate (mean ± s.e.) of flowers ofAcanthopale pubescens at different forest fragments andcontinuous forest (underlined) in the north (left) and thesouth (right) of Kakamega Nature Reserve.
Based on about 300 one-hourobservation sessions we candemonstrate significant differences(one-way ANOVA) in flowervisitation rates between study sites(Fig. 1). However, no pattern thatholds generally for differencesbetween fragments and continuousforest can be shown so far. This iscertainly due to the fact that only asingle plant species was includedin the study. The data on fruit andseed production that are presentlyanalyzed will surely revealimportant additional information.Furthermore, forest fragmentationat the Kakamega site obviously isstrongly overlayed by otherprocesses like present-day
anthropogenic disturbance, above all logging and grazing. The Kaimosi fragment, forinstance, although privately owned, is an extremely degraded forest whereAcanthopale pubescens is superabundant. On the contrary, the Kisere fragment with itsnot too abundant A. pubescens , is one of the best maintained forest fragments of thenorth. Among other reasons, at both sites, the high visitation rates may be directlyrelated to the plants abundance or, alternatively, may be the result of the lack ofalternative resources. To address these uncertainties it is necessary to incorporate otherplant species in the study. We already selected the abundant Dracaena fragrans,Acanthus eminens and Coffea sp. for our further investigations that will supposedlystart in January 2002.
N =
study site
visi
ts /
flow
er /
hour
0,3
0,2
0,1
0,022344127
KaimosiIkuywaYalaIsecheno543546
KisereSalazarColobus
0,3

168
Project ID: 01LC0025 (BIOTA East Africa E11)1.1.2001 – 31.12.2004
EFFECT OF FRAGMENTATION ON SEED DISPERSAL, REGENERATION ANDDYNAMICS OF KAKAMEGA FOREST, KENYA
B. Bleher, K. Böhning-Gaese
Institut für Zoologie, Abteilung V, Johannes Gutenberg-Universität, Becherweg 13, 55099 Mainz,[email protected], www.biologie.oekologie.uni-mainz.de, www.biota-africa.org
Key words: biodiversity, ecosystem processes, fragmentation, frugivores, fruiting plants,regeneration, seed dispersal, seedling recruitment
Abstract:This project investigates the influence of forest fragmentation on biodiversity and ecosystemprocesses related to seed dispersal and forest regeneration in Kakamega Forest. Data on thediversity of frugivorous animals as potential seed dispersers and on the diversity of fruiting trees,shrubs and lianes are currently gathered comparatively in four continuous forest and five forestfragment plots. In a first inventory, seedling recruitment of primary forest tree species wasmapped at the same sites to obtain a first understanding of the influence of fragmentation onforest regeneration. Seed dispersal, seedling recruitment and regeneration will now be studied indepth in a few selected tree species (i.e. Prunus africana, Olea capensis, Antiaris toxicaria).
Results:The greatest threats to the biodiversity of tropical forests are habitat loss and forest fragmentation.Understanding how fragmentation affects ecological processes that involve animal and plantpopulations and how this influences species and habitat persistence in the long term is particularlyimportant but hardly understood [1,2,3]. Seed dispersal by animals is one mutualistic interactionwhich is essential for the long-term maintenance of populations of tree species in tropical forests.Fragmentation disrupts animal populations and might lead to a decline in frugivore diversity inforest fragments. Consequently, this might result in a break-down of dispersal processesinfluencing the regenerative potential of forest ecosystems, leading to changes in abundance andspatial distribution of plants [4,5] with far-reaching consequences for the diversity, dynamics andstability of forest ecosystems [6,7]. However, despite its importance there has been relatively littlework on animal-plant interactions in tropical forest remnants [but see 1].
In this study we investigate seed dispersal and regeneration processes of forest treescomparatively in continous forest and forest fragments in Kakamega Forest, Western Kenya. Inour first field season 9 study plots of 1 ha each were established with four situated in the 8600 hacontinuous main forest block and five situated each in one of five peripheral forest fragments withsizes ranging from 100 ha to 1500 ha. Three of the study plots are situated within the biodiversityobservatories of BIOTA East (i.e. Buyangu Hill, Mukanku and Kisere).
Our first aim is to document changes in diversity and abundance of animal dispersersfocusing mainly on birds. Kakamega Forest is well known for its rich and unusual avifauna, withthe 194 forest-dependant bird species including 40 Guinea-Congo Forest biome species and 33Afrotropical Highland biome species [8]. A recent study showed that Kakamegas peripheralfragments have already lost several forest bird species since their separation from the main forestblock [9]. For our study we started a monthly monitoring scheme of the diversity and abundanceof the frugivore community using point counts, both in our continous forest and fragment plots.With these data we will be able to understand whether important functional groups such as certain

169
frugivorous dispersers are missing from forest fragments [10]. Parallel, we have startedmonitoring the diversity, abundance and phenology of fruiting trees, shrubs and lianas at the samesites. So far, fruiting has been low with mainly Ficus trees being a major fruit resource for thefrugivore community.
Beyond the diversity level, we secondly seek to determine whether forest fragmentationaffects seed dispersal and regeneration of tree species. Because the youngest plants in a previouslyfragmented forest are mainly the result of regeneration from seeds after the fragmentation processhas occurred, density and pattern of seedlings may provide clues to the possible effects offragmentation on tree regeneration. Therefore, a first inventory of forest regeneration was carriedout by mapping seedling recruitment and regeneration in 90 1 m2 seedling plots in each of thecontinuous forest and forest fragment plots. Seedlings were identified and counted and their ageestimated using the number of leaves, stem height and stem characteristics. First results indicatethat seedling diversity appears not to be related to fragment size or time of their isolation from themain forest block. However, there seems to be a relation to management system (with higherdiversity at sites under Kenyan Wildlife Service compared to under Forest Department) and todisturbance level with highest seedling species numbers in the low disturbance - sites Buyanguand Kisere Forest.
In a next step, we will now look behind the potential causes for differential seedlingrecruitment and study seed dispersal and regeneration of a few selected primary tree speciessuch as Prunus africana, Olea capensis and Antiaris toxicaria. We will will carry out treeobservations, identify seed dispersers, and quantify dispersal rates of individual trees in thesame continuous forest and forest fragment plots. We will also quantify seedling recruitmentand mortality due to predation and herbivory.
References:1. Laurance, W.F., and Bieregaard, R.O. 1997. Tropical forest remnants. Ecology, management, and
conservation of fragmented communities. University of Chicago Press.2. Tilman, D. 1999. Ecology 80: 455-1474.3. Chapin, F.S.III, Zavaleta, E.S., Eviner, V.T., Naylor, R.L., Vitousek, P.M., Reynolds, H.L., Hooper,
D.U., Lavorel, S., Sala, O.E., Hobbie, S.E., Mack, M.C., and Diaz, S. 2000. Nature 405: 234-242.4. Hubbell, S.P. 1979. Science 203. 1299-1309.5. Bleher, B., and Böhning-Gaese, K. 2001. Oecologia (in press)6. Bond, W.J. 1995. Pp. 131-146. in Lawton, J.H. & May, R.M. (eds). Extinction rates. Oxford University
Press, Oxford.7. Laurance, W.F., Ferreira, L.V., Rankin-De Merona, J.M., Laurance, S.G., Hutchings, R.W., and
Lovejoy, T.E. 1998. Conservation Biology 12: 460-464.8. Bennun, L., and Njoroge, P. 1999. Important bird areas in Kenya. Nature Kenya9. Brooks, T.M., Pimm, S.L., and Oyugi, J.O. 1999. Conservation Biology 13: 1140-1150.10. Symstad, A.J., Tilman, D., Willson, J., and Knops, J.M.H. 1998. Oikos 81: 389-397.

170
Project ID: 01LC0025 (BIOTA East Africa E12)01.04.2001.-31.03.2004
MONTANE FOREST ECOSYSTEMS IN KENYA - DIVERSITY AND POTENTIALFOR USE
R W. Bussmann1, S. Kiefer1, R. Njeri, G. Njoroge, S. Mathenge, Mutiso, J. Onyango2
1Lehrstuhl für Pflanzenphysiologie, Universität Bayreuth, Universitätsstr. 30, 95440 Bayreuth,Germany
2Department of Botany, Maseno University, Private Bag, Maseno, [email protected]; www.biota-africa.org
Key words: Montane forests, ecology, regeneration, vegetation, ethnobotany
Abstract:Objectives of the proposed project are: First the investigation of the dynamics and stability ofthe montane forests. How do different types of disturbance (timber extraction, and slash- andburn, high game and livestock populations) affect biodiversity. The second focus centers onthe current land-use practices and attempts to evaluate which sustainable use of theseresources would be possible. Special attention will be given to the use of non-timber products(particularly medical plants), and the carrying capacity with relation to game and livestockpopulations.
Project and first Results:Despite their enormous biological diversity and their exceptional economic and ecologicalimportance as water catchments, erosion barriers and source of timber and non-timberproducts, tropical montane forests have so far been overshadowed by the interest ofgovernments, public and scientific community in tropical lowland forests. Apart from theBiodiversity Convention, the “Mountain Agenda“ of the 1992 Rio Conference indicated thehigh priority in mountain region conservation. Little information however exists about thisecosystem. Few neotropical studies have been compiled. Most research focuses still onlowland rainforests or the alpine zones. Even large studies as „Ecoandes“ in Colombia touchthe mountain forests only marginally. Our own long-term studies about the phytosociologyand regeneration of the forests of Eastern Africa are a sound base for further ecologicalinvestigations. The research deficit in tropical mountain forests becomes apparent particularlywith regard to forest regeneration. There are almost no studies on regeneration- andsuccession processes after natural or human disturbances. Only in Eastern Africa, ourprevious work provides detailed information about forest regeneration. and the germinationand growth of important tree species.Kakamega and Mount Kenya were declared forest reserves in 1933 and 1932. At this time,there were still about 24000ha primary forest in Kakamega. Meanwhile, only 13000 ha(1988!) of fragmented forests are left. 10-20% of the fauna are regarded as endemic. KenyaIndigenous Forest Conservation Project (KIFCON) estimated a value of US$ 2.000.000 forannually harvested forest products, leading to high overharvesting. In the last 26 yearsKakamega lost about 50% of its standing timber. 60 years of absolute protection would benecessary to reach the 1965 level again – impossible in a region with high demand for forestproducts.Little knowledge exists about Kakamega. Few data on vegetation, tree species diversity andfragmentation are available, and little data on regeneration processes.

171
Research on use of non-timber products, particularly medicinal species is extremely importantto indicate alternatives to timber use and mining, and can thus provide a base for sustainableuse projects. Such studies need a high degree of trust from the side of the local population,and thus should be conducted by students from the host country.Main topic of research are the disturbance of the dynamic species composition in variousforest communities, the regeneration potential of the biocoenoses and the way of regeneration,and the use potential of the mountain forests for the local population. The successionprocesses are studied on selected plots with clear natural (treefall, wind, fire) or human(cutting, slash- and burn, pasture use) disturbance. Main focus are anthropogenousdisturbance, as natural processes have been studied by our group already in detail. Particularlyimportant will be to focus on possibilities for sustainable use – how fast can forests regenerateto a climax community? Is high species diversity necessary for maintaining stable forestcommunities? Fire is of special importance, as this is the most common mean of forestdestruction. Timber use and pasture are additional focuses. The possibility for regeneration ofdisturbed areas are studied with fencing experiments and grazing of selected plots. This willgive base data about the vegetation and its possible uses, and will lead to suggestions formanagement. Additionally, population structure, Regeneration and Ecology of importanttimber and non-timber species are investigated along altitudinal transects.
The intense use of non-timber products is extremely important for rural populations in largeparts of the tropics. Apart from forest products providing food, building materials or fodder,many species serve as medicinal plants. 80-90% of the population uses such traditionalmedicines. The knowledge about the use is handed from Generation to generation. In recenttimes, this knowledge transfer is interrupted more and more often, as old healers die, and noapprentices are available. This is particularly pitiful as this sector provides very interestingpotential for the sustainable forest use. The documentation of medicinal plants, their uses,possibly contents and their ecology are the second part of the research project. Two KenyanPh.D. students funded by the project conduct this work in the Kakamega and Mount KenyaRegions. A final goal of the work is to provide a public medicinal plant herbarium and a“medicinal plant guide“ for the training of health assistants in the local communities. This isan important focus in the establishment of a Botanical Garden at Maseno University, forwhich Maseno University has set aside 15 hectares of land. Maseno University contributes tothis task not only by providing the land but also by its pertinent botanical and horticulturalknowledge, by investment of manpower and by offering facilities for external researchers touse the plant material and other facilities in the garden. The goals of Maseno UniversityBotanical garden are to grow, study and promote tropical flora with emphasis on woodynative Kenyan species in an educationally useful setting as teaching field laboratories forstudents‘ individual ecological and conservation projects and to provide an informationresource for government, industry, science and the community for practical utilization offorest resources in human development. This is in particular necessary because of the almostcomplete deforestation of the country, leaving not more than 2% of the country as livingand/or gazetted forests. The destruction of plants with high medicinal value is taking place atfaster rate. This needs to be mitigated by ex-situ culture of medicinal plants which afterdeveloping appropriate cultivation techniques could then be made available to the traditionalhealers, thus Ethnobotany are one of the major goals of the garden.

172
Project ID: 01LC0025 (BIOTA East Africa, Yemen E13-E16)
BIOTA-EASTYEMEN-PROJECT-GROUP:
BIODIVERSITY AND ECOLOGY OF THE PALAEO-AFRICAN REFUGE AREASIN SOUTHERN ARABIA AND ON SOCOTRA (REPUBLIC OF YEMEN)
Since the Arabian Peninsula was separated from the African continent not earlier than in theOligocene, E Africa and the Arabian Peninsula had a common tropical flora, vegetation andfauna for a long time. At the same time, migration and exchange between the xerotropicalfloras and palaeotropical faunas of Asia and Africa were possible across the ArabianPeninsula. Due to the aridisation of the entire region in the late Tertiary, the palaeo-Africanvegetation and its fauna was then forced into the south of the Arabian Peninsula, isolated fromAfrica, with increasing aridity finally fragmented and delimited to refuge areas. In themonsoon-affected mountainous areas of the south coast of the Arabian Peninsula and on theisland of Socotra, such island-like relict vegetation complexes rich in endemics and with closefloristic affinities to NE tropical Africa have survived to the present. They are bioclimaticallyand ecologically conspicuously differentiated and comprise shrub formations, as well asdeciduous and semi-deciduous woodlands of various composition. Similar to the situationregarding the flora, the fauna of S Arabia consists of a unique and complex mix of widespreadafrotropical species, highly specialized endemics and remnant populations of palaearctic species.
Five representative palaeo-African refugia have been selected to investigate plant diversityand structure of xerotropical montane communities in southern Yemen and on Socotra bymeans of standardized monitoring sites. A main objective of the Yemen project group is toobtain information about the history of habitat fragmentation and its present effect on geneticdiversity in selected groups of plants, insects and vertebrates. Genetic comparison of isolatedpopulations is used to estimate the time frames of their historical dispersal and diversification.The floristic and vegetational data will provide new insights into the biodiversity andbiogeographical position of this little known but biogeographically very important region. Afirst documentation of the value, importance and ecology of these ecosystems finally will helpto estimate the present state of desertification and to develop management plans for thepreservation and sustainable use of these unique xerotropical communities.
The selected areas are:(1) Hawf Mountains, Governorate Al-Mahra: The coastal escarpments shelter the mostextended closed woodlands in Yemen, partly comprising endemic Anogeissus woodland. As aspecies rich area with considerable regional endemism it has been recommended by theEnvironmental Protection Authority of the Republic of Yermrn for protection. Important ruraleconomy.(2) Ras Fartak Mountains, Governorate Al-Mahra: A climatically transitional mountainousarea with very sparse and low woody, partly succulent vegetation cover, but with patches ofsemideciduous Anogeissus woodland in the most favourable sites in the summit area ataltitudes of 700-950 m. Extensive pasture economy.(3) Kor Seiban area, Governorate Hadhramaut: A summit plateau at altitudes of 1800-2000 mwith sea facing vertical escarpments and furrowed by numerous wadis. Remnants of semi-deciduous woodland have survived on some slopes and in wadis. Pasture economy.(4) Jabal Urays, Governorate of Abidjan: The climatically most favoured mountainous area inthe coastal SW of Yemen, with important relict communities and high local endemism.Pasture economy.(5) Socotra, Shihali Peaks, Governorate of Hadhramaut: Species-rich, climatically favouredwoodland communities. Pasture economy.
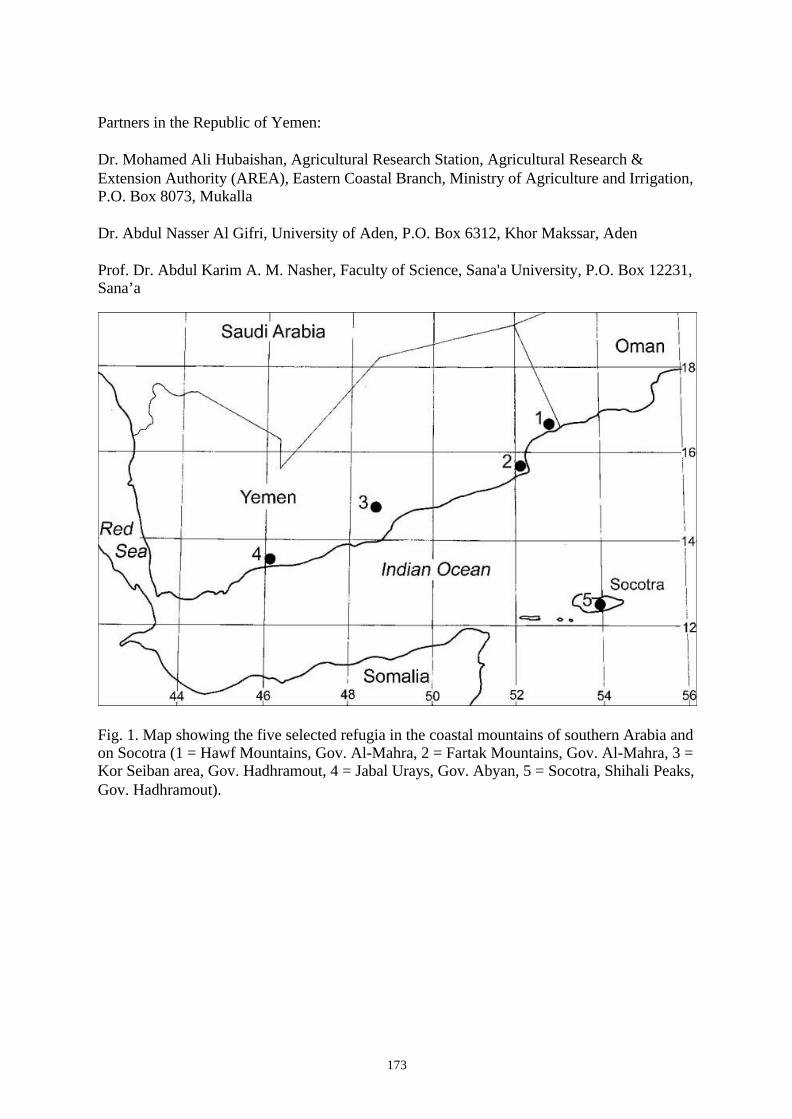
173
Partners in the Republic of Yemen:
Dr. Mohamed Ali Hubaishan, Agricultural Research Station, Agricultural Research &Extension Authority (AREA), Eastern Coastal Branch, Ministry of Agriculture and Irrigation,P.O. Box 8073, Mukalla
Dr. Abdul Nasser Al Gifri, University of Aden, P.O. Box 6312, Khor Makssar, Aden
Prof. Dr. Abdul Karim A. M. Nasher, Faculty of Science, Sana'a University, P.O. Box 12231,Sana’a
Fig. 1. Map showing the five selected refugia in the coastal mountains of southern Arabia andon Socotra (1 = Hawf Mountains, Gov. Al-Mahra, 2 = Fartak Mountains, Gov. Al-Mahra, 3 =Kor Seiban area, Gov. Hadhramout, 4 = Jabal Urays, Gov. Abyan, 5 = Socotra, Shihali Peaks,Gov. Hadhramout).

174
Project ID: 01LC0025 (BIOTA East Africa, Yemen E13)1.3.2001 – 31.3.2004
PHYTODIVERSITY AND VEGETATION TYPES OF REPRESENTATIVE PALEO-AFRICAN REFUGIA IN THE COASTAL MOUNTAINS OF SOUTHERN YEMENAND ON SOCOTRA (REPUBLIC OF YEMEN)
Norbert Kilian, Peter Hein, Mohamed Ali Hubaishan and Harald Kürschner
N. Kilian, ZE Botanischer Garten und Botanisches Museum Berlin-Dahlem, Freie UniversitätBerlin, Königin-Luise-Str. 6-8, 14191 Berlin; [email protected]. Hein, H. Kürschner, Institut für Biologie, Systematische Botanik & Pflanzengeographie,Freie Universität Berlin, Altensteinstr. 6, 14195 Berlin; [email protected],[email protected]. A. Hubaishan, AREA Mukalla, P.O. Box 8073, Mukalla, Republic of Yemen___________________________________________________________________________Key words: Yemen, Anogeissus woodlands, tropical vegetation relics, fragmentation, flora.
Abstract:As the results of the first six weeks' field campaign in September and October, firststandardized monitoring sites on which the Yemen Project Group combines their researchwere established in three of the five selected refugia, and 650 plant samples for the molecularanalyses of subproject E15 were provided. Determination, analysis and documentation of thefloristic inventory as well as the present vegetation complexes of the palaeo-African relictvegetation in the S Yemeni-Socotran refugia was started, focusing on the Bauhinia-Tarchonanthus woodland relics in the Kor Seiban refugial area in the governorate ofHadhramout and on the closed Anogeissus dhofarica woodlands in the two refugia in thegovernorate of Al-Mahra. Instructively illustrating the post-Tertiary fragmentation of thexerotropical plant cover in southern Arabia, preliminary results of the vegetational andfloristic study of the Anogeissus monsoonal woodland remnants in the Hawf and Fartakmountains are provided.
Results:The monsoonal woodland on the Arabian Peninsula is restricted to a small area along thecentral south coast. It extends from Jabal Samhan, at c. 55°E, in the province of Dhofar of theSultanate of Oman to the Ras Fartak mountains, at c. 52°10'E, in the Governorate of Al Mahraof the Republic of Yemen. The still little known main part of c. 10 000 ha in Yemen isconfined to the coastal escarpments of the Hawf Mts. between Al Fatk at 52°44'E° and theborder to Oman (our selected refugial area no. 1). To the larger extent it is an Acacia-Commiphora woodland, only on the climatically and edaphically most favoured sites, ataltitudes of 300-900 m and in about a fourth of the total woodland area, a closed woodlanddominated by the endemic Combretaceae tree species Anogeissus dhofarica and with a closedcanopy at about 7-8 m height exists. All eight species of Anogeissus are trees of rain greentropical woodlands and forests. A. leiocarpa occurs in Africa from NE Ethiopia to Senegal,five species occur from India to SE Asia, two species are endemic to S Arabia. Besides A.dhofarica this is A. bentii, which is restricted to gallery woodlands of wadis in Hadhramout.Both species are relics of the Tertiary belt of (semi)deciduous tropical forest and woodlandranging from S Asia to Africa across the south of the Arabian Peninsula.According to our preliminary results, the Anogeissus woodland is the by far most species-richwoodland community in the Arabian Peninsula, with c. 90 species per 1000 m2

175
Table 1. Tree and shrub species of Anogeissuswoodlands in Al-Mahra, Yemen.
Hawf FartakAcacia senegal + +Acacia etbaica subsp. uncinata + +Acacia nilotica subsp. indica +Acokanthera schimperi +Allophyllus rubifolius +Anogeissus dhofarica + +Azima tetracantha + +Blepharis dhofarensis +Blepharispermum hirtum +Cadia purpurea +Carissa edulis +Cocculus balfourii + +Commiphora kua + +Commiphora gileadensis + +Cordia perrottetti +Croton confertus +Delonix elata +Dichrostachys cinerea +Dodonaea angustifolia + +Euclea schimperi + +Euphorbia smithii +Ficus ingens +Ficus sycomorus +Ficus vasta + +Flueggea virosa + +Grewia bicolor +Grewia villosa + +Hildebrandtia africana + +Hybanthus durus + +Jasminum grandiflorum + +Jatropha dhofarensis +Justicia areysiana + +Lannea triphylla +Lantana sp. YP460 + +Maytenus dhofarensis + +Olea europaea subsp. cuspidata + +Pavetta longiflora + +Pistacia falcata +Premna resinosa +Rhus flexicaulis + +Rhus somalensis + +Ruttya fruticosa +Tarenna graveolens + +Woodfordia uniflora +Zygocarpum dhofarense +
on average (Table 1). Towards the upper edgeof the woodland belt, evergreen tree species aremixed into both the Acacia-Commiphora andthe Anogeissus woodland. A separate sclero-phyllous woodland belt as, e.g., the Olea-Bar-beya woodland in the Asir Mts. of Saudi Arabiais not formed in Al-Mahra. A hundred kmfarther west and entirely isolated from thesemain stands, the Fartak mountains (our refugialarea no. 2) shelter some outposts of Anogeissuswoodland. The effect of the SW monsoon isdrastically reduced there and the semi-deci-duous Anogeissus woodland type (Table 1) isconfined to small or even tiny patches (rangingfrom 80 to c. 2000 m2) in moist niches ataltitudes of 700-950 m (Fig. 1), which are,however, similarly species-rich as in the HawfMts. Initial floristic analyses of the Al-Mahrarefugia result in a number of species nothitherto known from Yemen (Table 2).
Table 2. Initial list of new records for the vascular plant flora of Yemen; 1 = Hawf area, 2 = Fartak area;vouchers at B.Alectra parasitica Hochst. (1), Becium dhofarense Sebald (1), Blepharis dhofarensis A.G. Mill. (1),Blepharispermum hirtum Oliv. (1), Centaurea dhofarica Baker (1), Ceratopteris cornuta (P. Beauv.) Lepr. (1),Cibirhiza dhofarensis P. Bruyns (1), Dhofaria macleishi A.G. Mill. (1), Dichrostachys cinerea (L.) Wight &Arn. (1), Dyschoriste dalyi A.G. Mill. (1), Euphorbia orbiculifolia S. Carter (1), E. smithii S. Carter (1),Gladiolus ukambanensis (Baker) Marais (1), Gossypium stocksii Masters (1, 2), Habenaria malacophyllaReichenb.f. (1), H. myodes Summerh. (2), Jatropha dhofarensis Radcl.-Sm. (1), Kleinia saginata P. Halliday(1), K. squarrosa Cufod. (1, 2), Lannea triphylla Engl. (1), Lavandula dhofarensis A.G. Mill. subsp. dhofarensis(1, 2), Maytenus dhofarensis Sebsebe (1, 2), Portulaca dhofarica M. G. Gilbert (1), Poskea socotrana (Balf.f.)Taylor (2), Pulicaria nobilis Gamal-Eldin (2), Remusatia vivipara Schott (1), Rungia parviflora Lindau (1),Suaeda moschata A.J. Scott (2).
Fig. 1. Small Anogeissus woodland patch on Jabal Fartak.

176
Project ID: 01LC0025 (BIOTA East Africa, Yemen E14)1.3.2001 – 31.3.2004
ECOLOGICAL CAUSES OF BIODIVERSITY IN THE COASTAL MOUNTAINS OFSOUTHERN YEMEN AND ON SOCOTRA (REPUBLIC OF YEMEN)
B. Mies, G.-B. Feige
Universität Essen FB9, Botanik/Pflanzenphysiologie, 45117 Essen, [email protected], [email protected], www.biota-africa.org
Key words: South Arabia, vegetation, relic forest, Anogeissus dhofarica, microclimate, dew-fall, ecology, ecophysiology
Abstract:The ecological analyses of the vegetation types aim at morpho-functional adaptations andgeneral patterns concerned with the xerotropical plant cover. These concern especially watereconomy, phytomass and specific productivity of the vegetation types. Pedologicalexaminations complete this view determining the influences of edaphic factors to vegetation.The causal explanation of vegetation ecology results in the explanation of the actualdesertification facing the thread of habitat destruction and loss of biodiversity. This gives ascientific basis to assist an ecologically sustainable development. An evaluation is aimed atthe ecological potential of nature resources and the intensity of human use or exploitation. Itreveals the qualification of vegetation types important to an indigenous economy and to theinfluences on specific degradation states as well as the actual risks for the ecosystems (fig. 1).
Results:During the South Arabian Field Campaign in Oct. 2001 and at the end of the monsoon season,microclimatic parameters and respective plant responses have been measured. FieldMeasurements were taken at the sights of permanent plots agreeing to the flora and vegetationstudies of Sub-Project E13. Primary data add some insight into the puzzle of understandingand approaching an ecosystem undergoing desertification. As rain-fall was exceptionally lowin 2001 - and the local population could not even yield one drop of cistern water - dew-fallseemed to provide the regular precipitation and to contribute a major supply to the vegetation.This subsidized pre-project measurements from Soqotran mountain ranges (fig. 2). Theextensive use of the chlorophyll fluorescence technique enlightened photosynthetic responseson individual and vegetation level. Diverse ecological patterns have been studied in detail atan Anogeissus dhofarica spot at Site 1 in the Hawf area. Further field data have been recordedin the coastal mountain ranges at Ras Fartaq and at Damqaut.Representative plant and soil probes have been taken for laboratory measurements. Plantmaterial was dedicated to quantify its contents of chlorophyll and minerals, soil samples totheir textures, nutrients and to the C/N analysis. Some representative specimens of dominantspecies have been transplanted to the greenhouses to give access to more sophisticatedecophysiological studies.
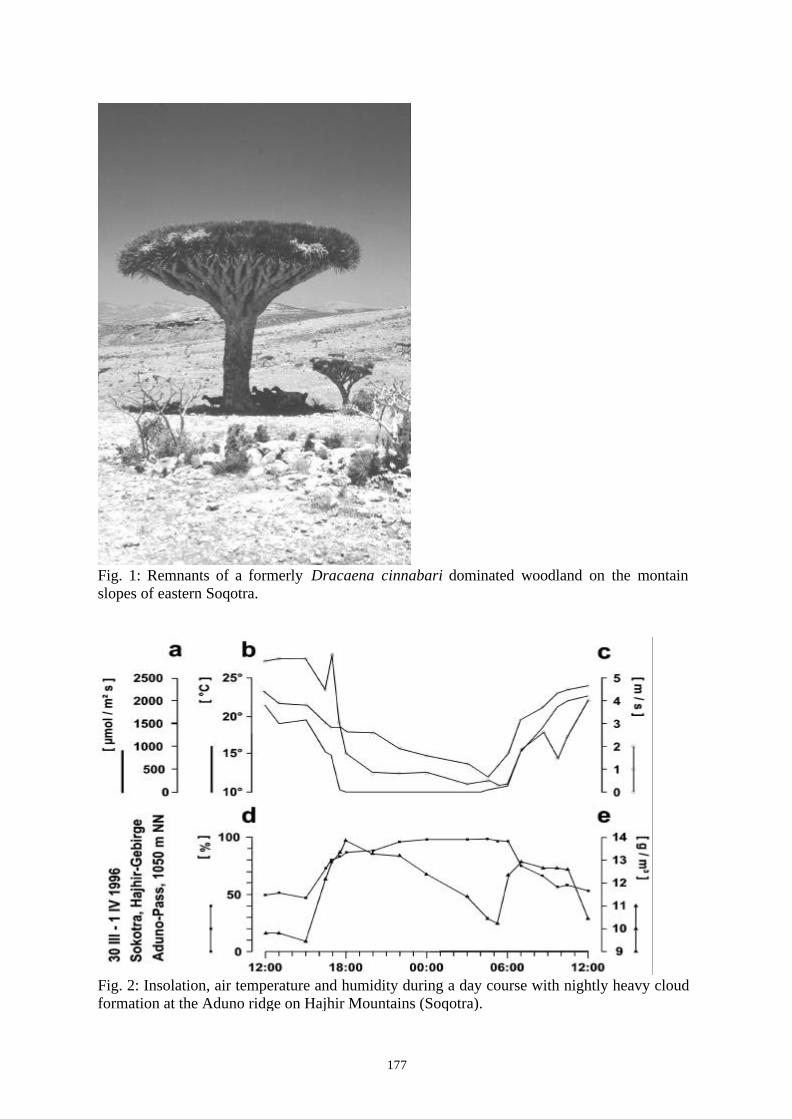
177
Fig. 1: Remnants of a formerly Dracaena cinnabari dominated woodland on the montainslopes of eastern Soqotra.
Fig. 2: Insolation, air temperature and humidity during a day course with nightly heavy cloudformation at the Aduno ridge on Hajhir Mountains (Soqotra).

178
Project ID: 01LC0025 (BIOTA East Africa, Yemen E15)1.5.2001 –31.3.2004
PHYLOGEOGRAPHIC AND POPULATION GENETIC ASPECTS OF THEPHYTODIVERSITY IN PALAEO-AFRICAN REFUGIA OF THE COASTALMOUNTAINS OF SOUTHERN YEMEN AND SOCOTRA (REPUBLIC OF YEMEN)
J. Meister1, Ch. Oberprieler2
1) Institut für Geobotanik und Botanischer Garten, Martin-Luther-Universität Halle/Wittenberg, Am Kirchtor 1, 06108 Halle/ Saale, Germany2) ZE Botanischer Garten und Botanisches Museum Berlin-Dahlem, Freie Universität Berlin,Königin-Luise-Str. 6-8, 14191 Berlin, [email protected]; [email protected], www.biota-africa.org
Key words: Arabian peninsula, phylogeography, habitat fragmentation, genetic diversity,RFLP, AFLP
Abstract:Using molecular genetic methods (PCR-RFLP analysis of the chloroplast genome, AFLP-fingerprinting) in five representative species or species groups of the palaeo-african refugia ofSouthern Yemen and Socotra, subproject E 15 aims at both the reconstruction of the temporaland spatial fragmentation processes which have led to the formation of these refugia and theeffect of habitat fragmentation on the genetic diversity of the populations involved. While for thephylogeographical part of the project the assessment of chloroplast haplotype variation asrevealed by PCR-RFLP analysis is planned, the genetic analyses of fragmented populations willbe based on the AFLP fingerprint technique. A short overview of molecular genetic methods andplant groups involved in the analyses of subproject E 15 along with first results of the PCR-RFLPscreening phase in Maytenus senegalensis (Celastraceae) is provided.
Subproject E15 of the Yemen-Project-Group within BIOTA-East uses molecular genetic methodsto reconstruct the chronological fragmentation of the palaeo-african refugia under study. Theelaboration of a hypothesis concerning the temporal and spatial development of these habitats willcontribute to a discussion of aspects of taxonomic differentiation, floristic evolution (together withsubproject E13), functional morphological adaptation (together with subproject E14) and willenable the comparison with comparable faunistic data (subproject E16). Additionally, populationgenetic analyses will be done to study the effects of fragmentation of populations on their geneticstructure and diversity.
Molecular genetic methods involved:• cpDNA PCR-RFLP: Using a set of universal primer pairs [1, 2, 3] non-coding regions
of the chloroplast genome are amplified via polymerase chain reaction (PCR) and se-quence and length polymorphisms are detected by digestion of the PCR-products witha set of restriction endonucleases followed by electrophoresis of resulting fragments.Phylogeographical interpretations are based on phylogenetic reconstructions ofchloroplast haplotype evolution.
• AFLP-fingerprinting: The digestion of a total extract of genomic DNA with specificrestriction endonucleases, followed by two selective amplifications of subsets of thepopulation of restriction fragments and their electrophoretic analysis [4, 5] yieldsindividual-specific band patterns (fingerprints) that are analysed in an population

179
genetic framework (e.g. Analysis of Molecular Variance, AMOVA [6]) to studygenetic variation within and between populations.
Species and species groups under study:• Maytenus senegalensis, M. dhofariensis (Celastraceae)• Gossypium stocksii, G. incanum, G. areysianum (Malvaceae)• Launaea crassifolia (Asteraceae)• Euryops arabicus (Asteraceae)• Euclea schimperi (Ebenaceae)• Justicia areysiana (Acanthaceae)
Results:Of the 16 pairs of cpDNA primers tested, different numbers were found to successfully amplifynon-coding regions of the chloroplast genome in each of the species involved in the study.Starting the screening process with Maytenus senegalensis, the amplified fragments were digestedwith four restriction enzymes (AluI, BsuRI, RsaI, TaqI) and the resulting fragments separated byelectrophoresis in agarose (2%) and PAA (8%) gels. While the screening is still in processpresently with a small test population of 10 individuals from different Yemeni provenances, untilnow we found two polymorphisms and consequently three chloroplast haplotypes in this species:Digestion of PCR product psbC-trnS (c. 1650 bp long) with TaqI resulted in a fragment of 370bp vs. 350 bp length in one N Yemen individual, while digestion of PCR product trnK1-trnK2 (c.2580 bp long) with TaqI revealed a difference between plants from N and S Yemen populations(240 bp vs. 280 bp fragment; Figure 1).The screening is presently continued for all other PCR products and the other species under study.Once established the relevant fragment/restriction enzyme combinations are used in analyses ofwhole populations.
Fig. 1.: Screening of chloroplast DNA variation inMaytenus senegalensis. The figured gel photographdemonstrates the occurence of two different PCR-RFLP patterns of the non-coding region trnK1-trnK2 (c. 2580 bp long) when digested with restric-tion enzyme TaqI. M: 100-bp molecular weightmarker; 1-6, 8: N Yemen populations; 7: Ras FartakMountains population; 9: Hadramaut population.
References:1. Taberlet, P. et al. 1991. Plant Molecular Biology 17, 1105-1109.2. Demesure, B. et al. 1995. Molecular Ecology 4, 129-131.3. Dumolin-Lapegue, S. et al. 1997. Molecular Ecology 6, 393-397.4. Vos, P. et al. 1995. Nucleic Acid Research 23, 4407-4414.5. Sharbel, T. F. In: Epplen, J.T. & Lubjuhn, T. (eds.), DNA profiling and DNA fingerprinting, Pp.
177-194. Basel, Boston.6. Excoffier, L. et al. 1992. Genetics 131, 479-491.

180
Project ID: 01LC0025 (BIOTA East Africa, Yemen E16)1.04.2001-31.03.2004
EVOLUTIONARY HISTORY AND PHYLOGEOGRAPHY OF SOUTH ARABIANFAUNAL ELEMENTS
C. Klütsch1, C. M. Naumann1, M. A. Hubaishan2, A. K. Nashr3
1 Zoologisches Forschungsinstitut und Museum Alexander Koenig, Adenauerallee 160, D-53113 Bonn, Germany² Director AREA Mukalla, POB 8073, Mukalla, Republic of Yemen³ University of Sanaa, Department of Biology, Chair of Zoology, Sanaa, Republic of [email protected], [email protected], [email protected]
Key words: South Arabia, biogeography, phylogeography, endemism, population genetics,evolutionary history, Lepidoptera, Reissita simonyi, Lasiommata felix, Amphibia, Hylasavignyi, Reptilia, Varanus yemenensis, DNA-microsatellites
Abstract:South Arabia is inhabited by a highly specialized fauna and flora of peculiar interest to thebiogeographer and evolutionary biologist. The rich biodiversity of the area is increasinglyendangered by various hazards, most of which are related to human population growth andits consequences. Our project aims at analysing the origin of various endemic faunalelements, the amount of gene flow between partly highly separated populations and atidentifiying viable populations that might be suited for population management and theestablishment of permanently protected areas. During two periods of field work (June andSeptember/October 2001) we have been able to 1. provide a clear picture of the presentdistribution of Reissita simonyi and closing a considerable gap in the distribution of the twosubspecies of the species (appr. 700 samples), 2. to document and sample about tenpopulations of Lasiommata felix (100 samples) and Hyla savignyi (200 samples) and 3. tocollect first samples of Varanus yemensis and related taxa from other parts of theAfrotropical region. In addition we have been able to sample about twenty populations of thelarval foodplant of Reissita simonyi, i.e. Maytenus senegalensis (Celestraceae) to be analyzedin subproject BIOTA E15. Genetic markers for population genetic analyses (DNA-microstaelliutes) of Hyla savignyi have been optimized and similar work has been started forReissita simonyi.
Results:The South Arabian fauna consists mainly of Afrotropical elements and a few Palaearcticfaunal elements. The amount of endemism is extremely high in the area (e.g. 13 endemicspecies of birds). Little is known on the temporal and spatial differentiation of these elements.Such questions therefore form the core part of our project. Of the four target species selectedfor the project two are of Afrotropical origin (Reissita simonyi, Varanus yemensis) and two ofPalaearctic origin (Lasiommata felix, Varanus yemensis). Thus, our analysis includes avertebrate and an invertebrate species of both categories.
Aims and Scopes- to analyse the faunal history of southern Arabia, i.e. processes related to local colonizationand subsequent differentiation of Afrotropical and Palaearctic faunal elements as a result oftectonic drift, post-glacial separation and subsequent habitat isolation due to regionaldesertification in the recent past.,

181
- to determine the duration of population isolation using genetic data and establishing amolecular clock for such events,- to provide basic data for the establishment of priority and target areas for natureconservation in the south-western parts of the Arabian peninsula.
Reissita simonyiSo far, two distinct and disjunct subspecies of this ecologically highly specialized taxon havebeen identified: R. s. simonyi along the southern coast (Oman, eastern governorates ofYemen; large, dark blue sheen, both sexes monomorphic) and R. s. yemenicola (along westernescarpment from the Hejjaz to Taiz; considerably smaller, forewing with greenish blue sheen,males dimorphic, females monomorphic). The disjunction (distributional gap) between thesetwo subspecies amounted to about 700 kms so far. In 2001 we have been able to discover apopulation of R. simonyi near Korseban (Prov. Hadramaut) which fills the middle of thepresent gap. Larval samples for molecular analysis have been taken from here and about 18further populations of both subspecies in Yemen (about 700 samples). The phenotype of theintermediate population is still unknown and will be the target of further analyses duringforthcoming field work. We assume that further investigations will still narrow the gapbetween the populations enabling us to estimate whether or not there is still gene flowbetween the two populations. We have also been able to document in detail the range of R. s.yemenicola and its larval host-plant, Maytenus senegalensis (Celastraceae). Of the latter,population samples have been secured for project Biota E15 (Dr. Oberprieler, Berlin)enabling our two research teams to compare data on gene flow in this phytophagous insectand its respective larval food-plant. Population samples of R. simonyi yemenicola have nowbeen documented from the border with Saudi Arabia in the north down to Jabal Sabr (nearTaiz), with maximum distances of 10-50 kilometers between locations. A number of localitieshas been recorded for the first time.
Hyla savignyi, Lasiommata felixSimilarly, we have been able to document the range of these two taxa to a rather detailedscale, again from the north (Jabal Rhaza) to Jabal Sabr in the south. The number of populationsamples taken so far from both taxa is promissing and will allow further sampling during thenext two years.
Molecular workThe development of DNA-microsatellites has been started for Reissita simonyi. In addition,DNA-microsatellitesd have een tested for Hyla savignyi and the conditions for PCR workhave been optimized.
References:1. Arens, P., Van´t Westende, W., Bugter, R., Smulders, M.J.M., Vosman, B. 2000. Molecular
Ecology 9, 1944-19462. Naumann, C.M., Edelmann, A. 1984. Fauna of Saudi Arabia 6, 473-509

182
Project ID: 01LC00011.1.2001-31.12.2003
BIODIVERSITY OF SELECTED MODEL GROUPS OF THE PHYTOPLANKTONIN STANDING INLAND WATERS OF KENYA
A. Ballot1), K. Kotut2), L. Krienitz1)
1) Leibnitz Institute of Freshwater Ecology and Inland Fisheries, D-16775 Neuglobsow, Germany2) Kenyatta University, Department of Botany, P.O. Box 43844, Nairobi, [email protected], [email protected], [email protected]
Key words: biodiversity, phytoplankton, Kenya, Rift Valley lakes, reservoirs, small water bodies
Abstract:The objective of the project is to investigate the biodiversity of phytoplankton communities instanding inland waters of Kenya with respect to the following items: (i) selection of prioritywaters representing model ecosystems of different phytoplankton biodiversity, trophic state andhuman use, (ii) investigation of the physico-chemical conditions of the waterbodies, (iii) detailedinventory of the species stock of model groups of phytoplankton with high ecosystemare relevanceand (iv) conclusions concerning their functional role in these waters. During the field workvarious Rift Valley lakes, reservoirs and small standing waters were investigated. The chemistryof the water bodies showed a great variety. Significant differences were also found in thecomposition of the phytoplankton communities. The phytoplankton biodiversity in Kenyanstanding waters is higher than expected, especially in the diversity of picoplanktic (< 3 µm) andnanoplanktic algae (< 20 µm).
Results:East Africa suffers from a deficit of high quality and easily accessible water resources, due to longdrought periods and increasing demographic pressures. Hence, there is an increasing need for theevaluation and formulation of management strategies for these water bodies. The water qualitycan be characterized by physico-chemical criteria and biological criteria like the phytoplanktoncommunity structure. The importance of the phytoplankton community structure and biodiversitycan be explained by the following circumstances: (i) being the main pelagic primary producer thephytoplankton plays a key role in standing water bodies, (ii) the structure of the phytoplanktoncommunity, i.e. its biodiversity has a principal influence on many kinds of water utilization byman, (iii) as a consequence of its high reproduction rates, phytoplankton gives a rapid response tochanges of environmental conditions, (iv) phytoplankton communities in general are more diversethan other eucaryotic populations in aquatic food webs.Despite of the importance of phytoplankton biodiversity, the investigations in this area still sufferfor various reasons: the lack of specialists of algal systematics, minor knowledge onphytoplankton diversity in different climatic regions of the world and the sometimes necessaryexpensive methods (electron microscopy and molecular biology) for correct determination. It isestimated that so far only 10-20 % of the algal taxa are known worldwide [1]. In East Africa,phytoplankton investigations have been going on for about 100 years [2]. However, theinvestigation of standing waters were mainly focused on large lakes like Lake Victoria or RiftValley lakes [3,4,5]. About reservoirs and small water bodies only few data are available [6,7,8].An extensive working program will be conducted in co-operation with: Kenyatta UniversityNairobi (KU); Kenya Wildlife Service (KWS); Kenyan Ministry of Education, Science &Technology (MEST); Tana & Athi Rivers Development Authority (TARDA); Kenya ElectricityGeneration Naivasha and Masinga (KenGen); Nairobi- Baringo- Koibatek City Councils (NCC,BCC, KCC); National Museum of Kenya (NMK); International Center for Research ofAgroforestry (ICRAF). The approaches are to link with the activities of the BIOTA-projects inAfrica and the BIOLOG-project AlgaTerra.
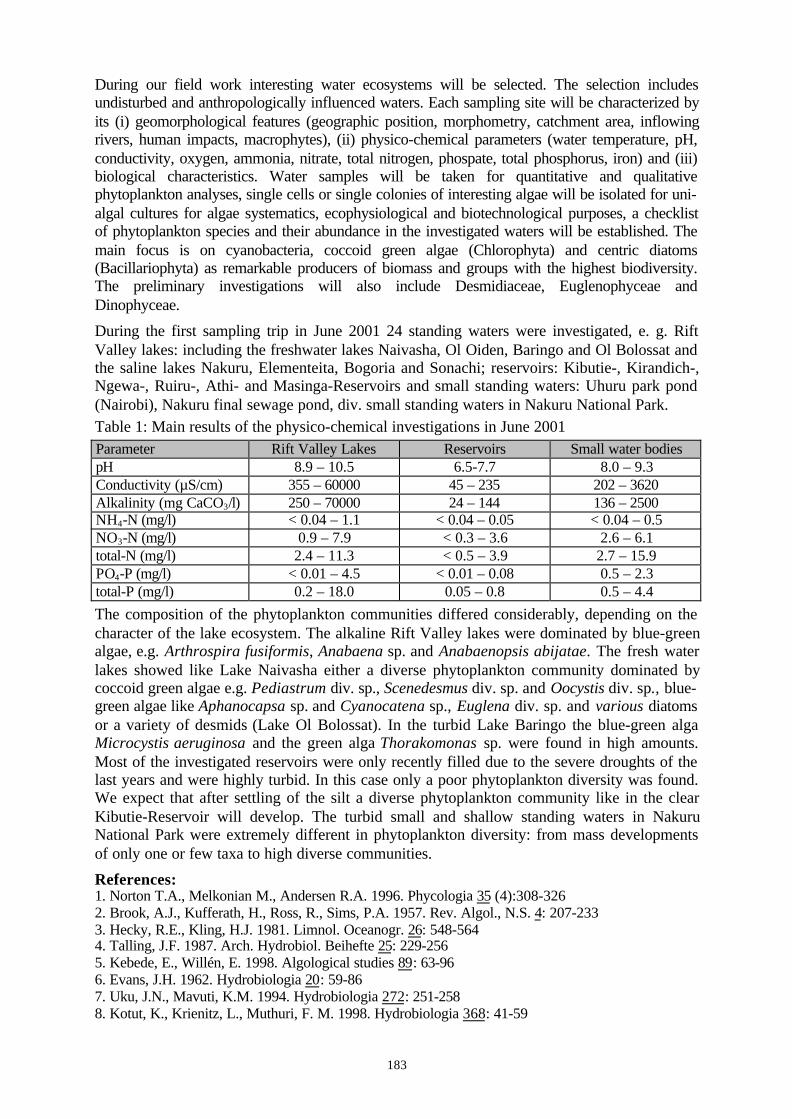
183
During our field work interesting water ecosystems will be selected. The selection includesundisturbed and anthropologically influenced waters. Each sampling site will be characterized byits (i) geomorphological features (geographic position, morphometry, catchment area, inflowingrivers, human impacts, macrophytes), (ii) physico-chemical parameters (water temperature, pH,conductivity, oxygen, ammonia, nitrate, total nitrogen, phospate, total phosphorus, iron) and (iii)biological characteristics. Water samples will be taken for quantitative and qualitativephytoplankton analyses, single cells or single colonies of interesting algae will be isolated for uni-algal cultures for algae systematics, ecophysiological and biotechnological purposes, a checklistof phytoplankton species and their abundance in the investigated waters will be established. Themain focus is on cyanobacteria, coccoid green algae (Chlorophyta) and centric diatoms(Bacillariophyta) as remarkable producers of biomass and groups with the highest biodiversity.The preliminary investigations will also include Desmidiaceae, Euglenophyceae andDinophyceae.
During the first sampling trip in June 2001 24 standing waters were investigated, e. g. RiftValley lakes: including the freshwater lakes Naivasha, Ol Oiden, Baringo and Ol Bolossat andthe saline lakes Nakuru, Elementeita, Bogoria and Sonachi; reservoirs: Kibutie-, Kirandich-,Ngewa-, Ruiru-, Athi- and Masinga-Reservoirs and small standing waters: Uhuru park pond(Nairobi), Nakuru final sewage pond, div. small standing waters in Nakuru National Park.Table 1: Main results of the physico-chemical investigations in June 2001Parameter Rift Valley Lakes Reservoirs Small water bodiespH 8.9 – 10.5 6.5-7.7 8.0 – 9.3Conductivity (µS/cm) 355 – 60000 45 – 235 202 – 3620Alkalinity (mg CaCO3/l) 250 – 70000 24 – 144 136 – 2500NH4-N (mg/l) < 0.04 – 1.1 < 0.04 – 0.05 < 0.04 – 0.5NO3-N (mg/l) 0.9 – 7.9 < 0.3 – 3.6 2.6 – 6.1total-N (mg/l) 2.4 – 11.3 < 0.5 – 3.9 2.7 – 15.9PO4-P (mg/l) < 0.01 – 4.5 < 0.01 – 0.08 0.5 – 2.3total-P (mg/l) 0.2 – 18.0 0.05 – 0.8 0.5 – 4.4
The composition of the phytoplankton communities differed considerably, depending on thecharacter of the lake ecosystem. The alkaline Rift Valley lakes were dominated by blue-greenalgae, e.g. Arthrospira fusiformis, Anabaena sp. and Anabaenopsis abijatae. The fresh waterlakes showed like Lake Naivasha either a diverse phytoplankton community dominated bycoccoid green algae e.g. Pediastrum div. sp., Scenedesmus div. sp. and Oocystis div. sp., blue-green algae like Aphanocapsa sp. and Cyanocatena sp., Euglena div. sp. and various diatomsor a variety of desmids (Lake Ol Bolossat). In the turbid Lake Baringo the blue-green algaMicrocystis aeruginosa and the green alga Thorakomonas sp. were found in high amounts.Most of the investigated reservoirs were only recently filled due to the severe droughts of thelast years and were highly turbid. In this case only a poor phytoplankton diversity was found.We expect that after settling of the silt a diverse phytoplankton community like in the clearKibutie-Reservoir will develop. The turbid small and shallow standing waters in NakuruNational Park were extremely different in phytoplankton diversity: from mass developmentsof only one or few taxa to high diverse communities.
References:1. Norton T.A., Melkonian M., Andersen R.A. 1996. Phycologia 35 (4):308-3262. Brook, A.J., Kufferath, H., Ross, R., Sims, P.A. 1957. Rev. Algol., N.S. 4: 207-2333. Hecky, R.E., Kling, H.J. 1981. Limnol. Oceanogr. 26: 548-5644. Talling, J.F. 1987. Arch. Hydrobiol. Beihefte 25: 229-2565. Kebede, E., Willén, E. 1998. Algological studies 89: 63-966. Evans, J.H. 1962. Hydrobiologia 20: 59-867. Uku, J.N., Mavuti, K.M. 1994. Hydrobiologia 272: 251-2588. Kotut, K., Krienitz, L., Muthuri, F. M. 1998. Hydrobiologia 368: 41-59

184

185
Biodiversity Informatics

186
Project ID: 01LC9901 (SysTax)1.4.2000 – 31.10.2003
SYSTAX - ELECTRONIC DATA PROCESSING FOR RECORDING ANDANALYSING BIODIVERSITY DATA WITH THE SYSTEMATIC ANDTAXONOMIC DATABASE SYSTEM SYSTAX
J. R. Hoppe and Th. Ludwig
Abteilung Systematische Botanik und Ökologie, University of Ulm, D-89069 [email protected]; http://www.biologie.uni-ulm.de/systax
Key words: SysTax, ORACLE DBMS, biodiversity data, botanical systematics, zoologicalsystematics, botanical garden, herbarium management, multimedia data, biological databasesystem, biodiversity informatics
Abstract:The SysTax system is a comprehensive biological database system which can be used for storingand retrieving biodiversity data from different sources. The data can be distributed over theWorld Wide Web. The system is available free of charge to all research institutions, botanicalgardens, and herbaria for online use. It has been developed since 1989 [1, 2, 3] based on theORACLE database management system. In this project the program modules “zoologicalsystematics”, “management of zoological collections”, and “information on taxa” (IOT, incl.“ethnobiology” and “plant-animal-interactions”) will be added to the system. The program willbe used by several BIOLOG projects (esp. “EDIS” and “InfoComp-Types”) for informationstorage to prove its usability as a common central database for biodiversity data.
Introduction. Within joint research biodiversity programs a large amount of primary andsecondary data on the biodiversity of plants, animals, and micro-organisms are accumulated. Thisincludes information relating to taxonomy and systematics, geography and geology, speciesdistribution, ecology, paleontology, and ethnobiology. Reference material, e.g. data on or picturesof collection objects, text protocols and field notes, and sound or video recordings can enormouslyincrease the amount of information to be stored and administered. The storage and retrieval of thedata in a powerful system is indispensable to promote synergistic effects, to allowmultidisciplinary evaluation, and to make the data generally available and usable for all projectparticipants and the scientific community in general.The concentration of computer resources and of database development at one site is - in ouropinion - a prerequisite for effective work and collaboration within joint projects. Thedevelopment of small local database applications often leads to systems incompatible with eachother. Because of these incompatibilities and because of the cost involved in data exchangebetween different systems, such local solutions end up being much more expensive than thedevelopment of a small number of large and powerful systems.The SysTax system has been developed at the University of Ulm and the Ruhr-University ofBochum since 1989 with the support of the DFG (Deutsche Forschungsgemeinschaft), the GTZ(Gesellschaft für Technische Zusammenarbeit), the University of Ulm, and the BMBF (within the" Federal Information System Genetic Resources - BIG; a project coordinated by Prof. Dr. Th.Stützel, Ruhr-University of Bochum). SysTax is based on the ORACLE database managementsystem. From the very beginning, the entire system was implemented not only as a multi-usersystem but also as a “multi-institutional” system. Several scientific institutions can store e.g. theircollection data separately but all the data can be queried together. Because of this feature thesystem can be used for the installation of comprehensive information systems on the World Wide

187
Web. A “security system” allows to hidesensitive data from being read by the publicso that an abuse of data can be prevented tothe greatest possible extent.Results. In this project the program modules“zoological systematics”, “management ofzoological collections”, and “information ontaxa” (IOT, incl. “ethnobiology” and “plant-animal-interactions”) are being added to thesystem. SysTax is used by several projects inthe BIOLOG-Biodiversity programme (esp.EDIS) for information storage to prove itsusability as a common central database forbiodiversity data.Up to now, the program module zoologicalsystematics has been developed. It followsthe “potential taxa” concept [4] thus pro-viding the possibility of storing several“alternative taxonomies” as well as thecorresponding “alternative synonymies” andthe so called “concept synonyms” for a taxonname. The program modules “ethnobiology”and “pictures” did already exist asprototypes. They were now combined to anew module “information on taxa” which isnow used to store any additional (multi-media) information on animal and/or plants.Data of the EDIS project as well as of theInfoComp project have been imported.
References:1. Hoppe, J. R., Boos, E., and Th. Stützel 1999: SysTax - ein Datenbanksystem für Systematik undTaxonomie. In: Begemann, F., Harrer, S., and J. D. Jiménes Krause (Hhrg.), Dokumentationen undInformationssysteme im Bereich pflanzengenetischer Ressourcen in Deutschland. Schriftenreihe zuGenetischen Ressourcen (Zentralstelle für Agrardokumentation und -information - ZADI, Informa-tionszentrum für Genetische Ressourcen - IGR), 64-79.2. Hoppe, J. R. 1998: Practical suggestions for database implementations. In: Thiéry, M., Stevens, A.-D.,Hoppe, J. R., Charles-Dominique, P., and Schuchmann, K.-L., Angiosperm pollination and seed dispersal,a review. Ecotropica, 4, 69-91.3. Hoppe, J. R. and Boos, E. 1996: Ein "Informationssystem Botanischer Gärten" auf der Basis desDatenbanksystems SysTax. Gärtnerisch-Botanischer Brief, 124, 10-15.4. Berendsohn, W. G. 1995: The concept of "potential taxa" in databases. Taxon, 44: 207-212.
SysTax –Task Profile
Botanical and zoological systematicsand taxonomyhierachical storage of names and (potential) taxa;flexible synonymy; linkage of taxa with literature andspecimens; linkage of taxa with characters andcharacter states; taxonomical reports for direct use inrevisionsLiteraturestorage and administration of references; crossreferencing literature data and taxa; storage ofliterature scans (plant descriptions)Botanical Gardenstorage and administration of garden accession data;label printing; retrieval of accession lists;creation of seed catalogues; mailing lists for seedcatalogues and seed materialHerbariumstorage and administration of herbarium label data;administration of loans; output: herbarium labels,revision labels, specimen lists, letters for loan,reminders, exchange, returnsVernacular namesAddressesInformation on Taxa (IOT)storage and retrieval of information on taxa (plantsand/or animals): texts, pictures, movies, sound, andOLE-objects; thesaurus for ethnobioloy and plant-animal-interactionsQueries by WWW-Formstaxonomic reports: taxonomy, synonymy, pictures,texts sound, movies, OLE-objects, collections (incl.pictures of specimens)

188
Project ID: 01LC9902 (ARB)1.4.2000 – 31.03.2003
AUTOMATED MAINTAINANCE OF INTEGRATED DATABASES FOR RRNA ANDOTHER RELEVANT GENE SEQUENCES; SOFTWARE DEVELOPMENT,OPTIMIZATION, AND PARALLELIZATION FOR AUTOMATED SEARCHING OFSEQUENCE IDENTIFIERS FOR ANALYZING DIVERSITY
W. Ludwig1, R. Westram1, L. Richter1, H. Meier2, and T. Ludwig3
1 Lehrstuhl für Mikrobiologie, Technische Universität München, am Hochanger 4, D-85350Freising.2 Lehrstuhl für Rechnertechnik und Rechnerorganisation, Technische Universität München,Arcistr. 21, D-80333 München. 3 Institut für Informatik, Ruprecht-Karls-Universität Heidelberg,Im Neuenheimer Feld 368, D-69120 [email protected]; http://www. [email protected]; http://www. mikro.biologie.tu-muenchen.de
Key words: databases, identification, rRNA, specific probes, software development
Abstract:Within the scope of BIOLOG the ARB project provides periodic releases of integrated databasesof processed sequence and other data for rRNA and other genes relevant in biodiversity studies.New ARB software tools are currently being developed, optimized or evaluated for automateddatabase maintenance and evaluation. Further software developments focus on the automateddesign and permanent in silico evaluation of comprehensive sets of taxon specific probes orprimers for biodiversity analyses according to the multiple probe concept. These probe sets willlater be accessible via the Internet.
Introduction. Comparative sequence analysis of informative genes nowadays plays a central rolein biodiversity studies for identification as well as phylogenetic and taxonomic analyses.Doubtless, the currently most important marker genes are those encoding ribosomal RNAsespecially 16S/18S rRNAs. This is reflected by a dataset of about 30.000 primary structuresavailable in public databases. Given that the vast majority (> 90%) of prokaryotes cannot be keptin pure culture thus far, comparative sequencing in combination with taxon specific probe and/orPCR approaches often represent the only methods for identification, detection and quantificationin environmental samples [1]. These molecular methods are also getting more and more routinetechniques for studying diversity, evaluating taxonomy, identification and detection of eukaryoticorganisms.The project. The most important prerequisites for diversity studies based upon these moleculartools are comprehensive sequence databases and powerful software for database maintaining,handling and analysis. Both aspects are in the focus of the ARB project initiated some years ago[2, 3]. The ARB program package provides a software environment for these tasks. Processed(with respect to alignment, quality, conservation, secondary structure and other criteria) sequencedata are stored in a central database. Any additional information such as descriptive data,bibliography, user comments or links to external databases can be assigned to individual sequenceentries and visualized along with a phylogenetic tree. Database access and navigation is possiblevia mouse click in that tree or by using search tools. A powerful editor for real or virtual primarystructures includes versatile tools for string searching, automated alignment and secondarystructure check as well as secondary structure visualization. Conservation profiles and columnfilters can be established according to user defined criteria. Phylogenetic treeing can be performedapplying the respective integrated tools. A special maximum parsimony approach allows

189
reconstruction and optimisation of comprehensive trees representing the full sequence data set(> 30.000 entries). The ARB PT server (positional tree) provides the basis for rapid searching ofclosest relatives or specific sequence signatures. Such signatures can be evaluated as taxonspecific probes or primers against the background of the full database. The ARB package isavailable at www.arb-home.de and in use worldwide.Results. Within the scope of the BIOLOG project ARB databases comprising processedsequences of small and large subunit rRNA, elongation and initiation factor, ATPase, RecA,HSP60, polymerases and other conserved gene sequences including additional information andanalyses are maintained and permanently updated. Periodic releases are available athttp://www.arb-home.de or upon request.In order to reduce the need of manpower and expertise for processing the rapidly increasingamount of data, software for automation of database maintenance is being developed, evaluated,and/or optimised. The focus is on further automation of data retrieval, selection, update orcorrection, quality and higher order structure check, alignment, and phylogenetic analyses as wellas automated database optimisation based upon iterative cyclic application of the former tools.The concept of integrated databases giving access to any data (molecular, physiological,morphological, ecological and other) known for an organism has been followed from thebeginnings of the ARB project. Concerning the integration of foreign data ARB tools for rapidaccess to external databases have been improved. Via mouse click and Internet browsers queriescan be performed according to ARB data field entries such as accession numbers, DNATAX(project 01LC9904/2) numbers, organism names, or other identifiers on remote databases such asEBI, GenBank, Medline, and SysTax. (project 01LC9901).Another important task in ARB biodiversity informatics is the design and evaluation of taxonspecific probes or primers as tools for studies of diversity with the ultimate goal of automateddesign and permanent evaluation of all potential probes against the background of the full data setof available sequences. In the ARB version currently available, probe design is an interactiveprocedure and can be performed for selected organisms or groups only. The ARB-PT-(positionaltree)-server which represents the central tool for probe design and in silico evaluation has beenimproved. A new tool for automated definition of specificity levels according the criteria ofpresence of potential target sites and phylogenetic relationship is currently being tested. Furthertools for finding all minimal probe sets characterizing individual taxa as well as for automateddesigning comprehensive probe sets of hierarchical specificities according to the multiple probeconcept are currently being developed.Software and database support is provided to the DNATAX project. Parts of the existing data-bases (rRNA elongation factor) can be immediately used, others (Cytochrome oxidase) have to beestablished de novo. Collaboration with respect to data exchange and software development hadalready been established between the ARB group and the BIOTA S04 project.In collaboration with the MPI for marine microbiology and the VAAM (Vereinigung fürallgemeine und angewandte Mikrobiologie) four and two ARB courses for beginners (three dayseach) and advanced users (two days each) have been performed.
References:1. Amann, R. and Ludwig, W. 2000: Ribosomal RNA-targeted nucleic acid probes for studies in microbialecology. FEMS Microbiol. Rev., 24, 555-565.2. Ludwig, W. and Strunk, O. ARB: a software environment for sequence data. http://www.arb-home.de/arb/documentation.html3. Ludwig, W. 1995: Sequence databases. In: Akkermans, A. D. L., van Elsas, J. D., de Bruijn, F. J. (eds.),Molecular microbial ecology manual, 3.3.5, 1-22. Dordrecht.

190
Project ID: 01LC9903 (ZEFOD)01.05.2000 – 30.04.2003
REGISTER FOR BIOLOGICAL RESEARCH COLLECTIONS IN GERMANY
I. Brake2, W. Braun1, D. Guttwein1, I. Groß3, F. Klingenstein3, K-H. Lampe 2, and S.Roscher1
1 Zentralstelle für Agrardodumentation und –information (ZADI), Villichgasse 17, D-53177Bonn. [email protected], http://www.zadi.de/igr/2 Zoolog. Forschungsinstitut u. Museum Alexander Koenig, Adenauerallee 160, D-53113 [email protected], http://www.uni-bonn.de/museumkoenig/3 Botanisches Institut u. Botanischer Garten Uni Bonn, Meckenheimer Allee 171, D-53115 [email protected], http://[email protected]
Key words: access system, biodiversity, biodiversity informatics, biodiversity information,biological collections, botanical gardens, botany, collection metadata, herbaria, zoologicalmuseums, zoological gardens, zoology
Abstract:Only occasional overviews and analyses of botanical and zoological research collections inGermany exist at the moment, but individual collections are in various stages of selfdocumentation. The existing data will be combined by ZEFOD, systematized and, together withnew data, presented via the Internet as an expandable, interactive information network for abroad range of users in science, administration, and the general public. Apart from an overviewof collections, the ZEFOD information system will provide a structural and content-orientateddescription of biological research collections in Germany based on metadata provided bycollection holders. To record the metadata, questionnaires have been designed, mailed to thecollection holders, and made available via the ZEFOD homepage [1]. Metadata areas coveredinclude data on the institution or collection holder (address, contacts, staff), general informationabout the collection (taxonomic and geographical foci, state of documentation), and a descriptionof the collection’s contents (number of objects, degree of computer recording, number ofnomenclatural types).ZEFOD stands for “Zentralregister biologischer Forschungssammlungen in Deutschland”.
Current state of the database of national zoological collections. The questionnaires forzoological collections in museums or universities were mailed in three batches (13.6., 3.8.,5.9.2001) to 214 institutions. The questionnaires for zoological gardens have been mailed inNovember, because these institutes usually have more time to do paper work in winter. ByOctober 9, 2001, 20% of the institutions had answered the questionnaires (42 institutions with 102collections), 13% announced that they want to participate, but need more time, and 10% do nothold the sort of collections required for participation in the ZEFOD database. In all 43% of theinstitutions responded to our initiative within the first four months. A further increase of coverageis expected, because the response rate for the first batch of questionnaires (mailed in June) hasreached 82%.Some of the museums were visited in order to clarify the aims of the project and to obtain thedata. Similarly several scientific as well as other meetings within the community of collectionholders, curators, etc. were visited. Representatives of Naturweb (Austria) and BIODIV(Belgium) were contacted in order to exchange experience with regard to online databasescarrying meta information about biological collections. A representative of ISIS (InternationalSpecies Information System), a database for zoological gardens, was contacted to check thepossibility of using some of their data for ZEFOD.

191
Current state of the database of national botanic collections. Questionnaires were mailed to 70herbaria. For Botanic Gardens data from 435 special collections in 71 gardens have already beencollected in previous projects (see, e.g., [2]). The survey for living collections will be based onthis information, so the gardens will obtain partly filled-in questionnaires to complete their data ina next step. An Access2000 database application is used for interim data collection and the dataexisting for Botanical Gardens have been transformed into the ZEFOD data format and integratedinto the data bank. The database already accepts queries for different collection characteristics liketaxonomy, geography, biotopes, and functional types. It also allows a detailed analysis and showsthe amount, variety, and diversity of living plant collections in Germany and different spectra ofthe representation of diversity in nature and in collections.To ensure support for the project and to achieve maximum data quality, the Association ofBotanical Gardens (VBG) has been informed about the objectives and working plan of ZEFOD.16 Botanic Gardens and 6 herbaria have already been visited to inform directors and staff aboutZEFOD and to give practical support in answering the questionnaire. Moreover, the project hasbeen presented in 4 meetings within the botanical collection community. Within this audienceZEFOD is now well known and the utility of its aims is widely accepted.
Information technique. The ZEFOD database itself is being implemented as a relationaldatabase, optimised to store the complex data resulting from the filled in questionnaires. Datainput from these questionnaires is done by routines reading the word files and by off-line input,using interfaces containing ‘classical’ input fields (checkboxes, drop down lists, etc.). Tools foronline-data input are also under development which will help to develop ZEFOD into apermanently sustainable information system. Acceptance of the Internet has to be improved,however, before these tools will be used by all collection holders. Scripts for checking correctnessof links to homepages of the institutions are under development to guarantee that the providedinformation is up-to-date.One important access path to the information in ZEFOD is the search for taxa by their name.Therefore a database structure to store scientific and common names is under development. Theuser will also be supported by a thesaurus for querying system.While data capture will be done through the whole phase of the project, some of the results arealready available by querying the online-database [1]. Technically this is realised by usingORACLE 8i and scripts for creating dynamic pages. Access to institution names and places is freebut there is temporary a restricted area for accessing further details. This allows collection holdersto check the data before they will be published.Biological collection information may be provided on the collection metadata level and on theunit (specimen) level. ZEFOD provides metadata for all biological research collections inGermany and will make unit level information access possible. Seen in the international contextZEFOD is a direct contribution to international initiatives like the ‘Global Taxonomic Initiative(GTI)’‚ the Convention on Biological Diversity (CBD) and the ‘Global Biodiversity InformationFacility (GBIF)’. ZEFOD is the German national node in BioCASE [3].
References:1. ZEFOD Homepage. http://www.genres.de/zefod.2. Rauer, G., v. den Driesch, M., Ibisch, P. L., Lobin, W., and Barthlott, W. 2000: Beitrag der deutschenBotanischen Gärten zur Erhaltung der Biologischen Vielfalt und Genetischer Ressourcen. Bonn.3. A Biodiversity Collection Access Service for Europe. http://www.biocase.org

192
Project ID: 01LC9904 (EDIS)1.4.2000 – 31.03.2003
EDIS – ENTOMOLOGICAL DATA INFORMATION SYSTEM
C. L. Häuser, J. Holstein, and A. Steiner
State Museum of Natural History, Rosenstein 1, D-70191 [email protected] , [email protected],[email protected]; http://www.insects-online.de
Key words: biodiversity informatics, taxonomy, type specimens, world checklists, speciesidentification, DNA sequencing, molecular systematics, image analysis, geographical informationsystems, bio-acoustics, interactive keys, insects, Collembola, Saltatoria, Lepidoptera,Hymenoptera, Coleoptera, Acari, Araneae
Abstract:The Entomological Data Information System (EDIS) is a co-operative research project jointlyundertaken by a number of leading German natural history museums and university institutions inthe area of biodiversity informatics. The purpose of EDIS is to facilitate access to informationabout insects and other arthropods for biodiversity research and the general public by developingand linking relevant databases and information tools established by EDIS partner institutions.The information provided by EDIS includes scientific data about insects from a wide variety ofsources ranging from global species inventories, molecular sequences, digital images and soundsto geographic distribution maps.
EDIS organisation:The EDIS framework project is co-ordinated at the State Museum of Natural History, Stuttgart,Germany, and currently comprises six separate projects (ABIS, DNA-TAX with sub-projectMOLART, DORSA, GART, INGE and OBIF) undertaken by different partner institutions. Thedatabases and information systems developed by EDIS are in the public domain and will all beaccessible via a central Internet gateway developed in co-operation with the Department ofSystematic Botany and Ecology at the University of Ulm, Germany, making use of the SysTaxdatabase system. The individual EDIS projects are implemented independently by the respectiveresearch institutions, and focus on specific questions in selected insect groups (see followingstatus reports of individual projects). The information provided by EDIS is primarily intended foruse by research and development, but also for other users with a general interest in insects.
EDIS goals:• establishing and extending scientific databases on insects using collection specimens,
literature, multimedial and molecular data• developing and improving new IT-tools for identification of - and information access on -
insects• linking separate databases containing insect biodiversity data of various kinds through a
common access point• providing a common public gateway to a variety of information sources about insects

193

194
Project ID: 01LC9904 (EDIS, Subproject ABIS)1.4.2000 – 31.03.2003
ABIS - AUTOMATED IDENTIFICATION OF BEE SPECIES
V. Steinhage2, T. Arbuckle2, S. Schröder1, A. B. Cremers 2, and D. Wittmann1
1 Institut für Landwirtschaftliche Zoologie und Bienenkunde, University of Bonn, Melbweg 42,D-53127 Bonn2 Institut für Informatik III, University of Bonn, Römerstr. 164, D-53117 [email protected]
Key words: species identification, taxonomy, image analysis, supervised learning, biodiversityinformatics
Abstract:Computer aided systems have become popular in zoological systematics in recent years. Forinsects, the difficult taxonomy and the lack of experts greatly hamper studies on conservation andecology. Bees in particular, as the main pollinators of food crops, represent a critical ecologicaland economical resource which needs to be carefully exploited and managed. We have developeda system for the automated identification of species that employs knowledge-based image analysisof the bee forewings. The system has been successfully deployed in Germany, Brazil and the U.S.
Results:The key idea of the Automated Bee Identification System ABIS is to identify bee species fromimages of their forewings. Using diffuse background illumination, the wings of the bees show aclear venation within a transparent skin. The structure of this venation is genetically fixed andtherefore suited to species identification [1]. Analysing each wing image gives rise to a set ofabout 300 characteristic numerical features. These features are used in a new knowledge-basedclassification approach to identify the bees’ species. The image analysis and classification areperformed in a hierarchy of consecutive operations. First, ABIS identifies the genus. In the secondstep, ABIS identifies species groups – so-called species complexes. In the final step, ABISemploys a new non-linear discriminant analysis to perform the species identification [2]. Eachstep of this hierarchical approach is guided by the knowledge extracted at the previous step [3].The result of this classification can be visualised in terms of cluster images or dendrograms. Theequipment required is easily portable and suited for work in collections or in the field [4].
Fig. 1: Use of ABIS in collections Fig. 2: Automatic image analysis Fig. 3: Use of ABIS in the field
We give two representative results of our prototype system. Figures 4 and 5 show speciesidentifications for the genera Bombus and Colletes. Figure 4 shows the genus identification withtwo clearly separated cluster centres (top: the Bombus species, bottom: the Colletes species).Figure 5 shows the identification of two species complexes of the genus Bombus. The sevenBombus species (B. cryptarum (Fabr. 1775), B. lucorum (L. 1761), B. magnus Vogt 1911, B.terrestris, B. ruderarius, B. sylvarum und B. veteranus) in the given example belong to two
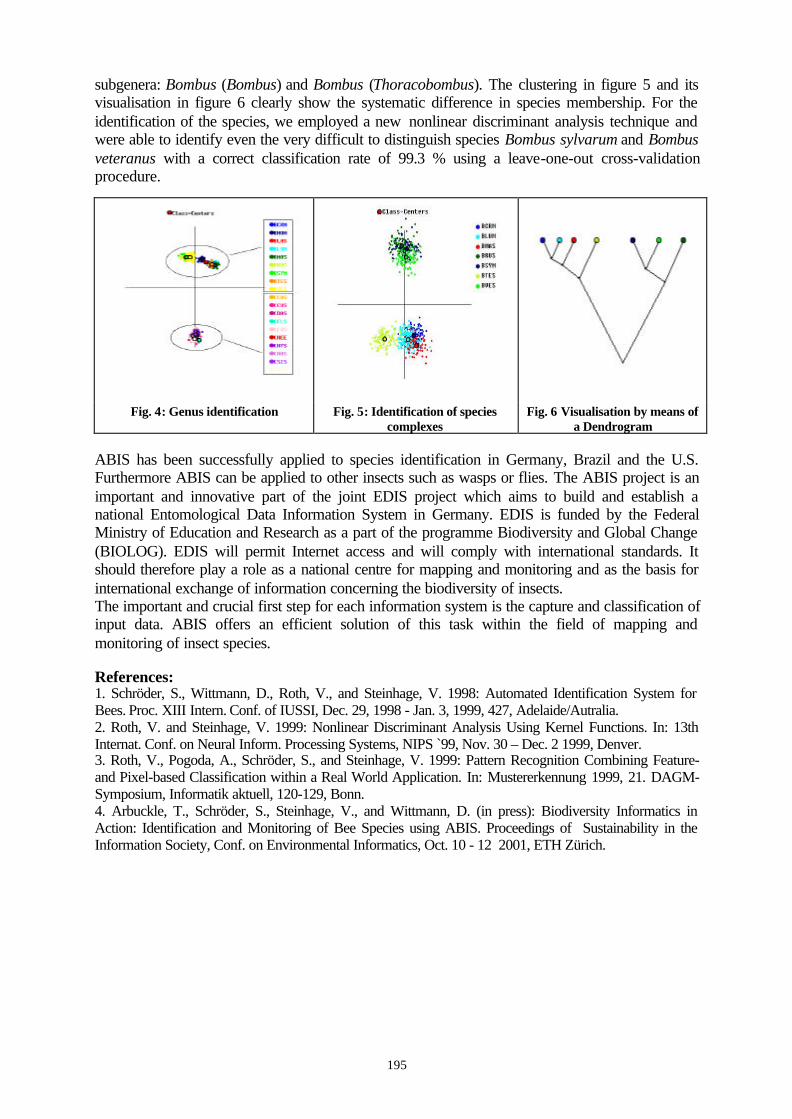
195
subgenera: Bombus (Bombus) and Bombus (Thoracobombus). The clustering in figure 5 and itsvisualisation in figure 6 clearly show the systematic difference in species membership. For theidentification of the species, we employed a new nonlinear discriminant analysis technique andwere able to identify even the very difficult to distinguish species Bombus sylvarum and Bombusveteranus with a correct classification rate of 99.3 % using a leave-one-out cross-validationprocedure.
Fig. 4: Genus identification Fig. 5: Identification of speciescomplexes
Fig. 6 Visualisation by means ofa Dendrogram
ABIS has been successfully applied to species identification in Germany, Brazil and the U.S.Furthermore ABIS can be applied to other insects such as wasps or flies. The ABIS project is animportant and innovative part of the joint EDIS project which aims to build and establish anational Entomological Data Information System in Germany. EDIS is funded by the FederalMinistry of Education and Research as a part of the programme Biodiversity and Global Change(BIOLOG). EDIS will permit Internet access and will comply with international standards. Itshould therefore play a role as a national centre for mapping and monitoring and as the basis forinternational exchange of information concerning the biodiversity of insects.The important and crucial first step for each information system is the capture and classification ofinput data. ABIS offers an efficient solution of this task within the field of mapping andmonitoring of insect species.
References:1. Schröder, S., Wittmann, D., Roth, V., and Steinhage, V. 1998: Automated Identification System forBees. Proc. XIII Intern. Conf. of IUSSI, Dec. 29, 1998 - Jan. 3, 1999, 427, Adelaide/Autralia.2. Roth, V. and Steinhage, V. 1999: Nonlinear Discriminant Analysis Using Kernel Functions. In: 13thInternat. Conf. on Neural Inform. Processing Systems, NIPS `99, Nov. 30 – Dec. 2 1999, Denver.3. Roth, V., Pogoda, A., Schröder, S., and Steinhage, V. 1999: Pattern Recognition Combining Feature-and Pixel-based Classification within a Real World Application. In: Mustererkennung 1999, 21. DAGM-Symposium, Informatik aktuell, 120-129, Bonn.4. Arbuckle, T., Schröder, S., Steinhage, V., and Wittmann, D. (in press): Biodiversity Informatics inAction: Identification and Monitoring of Bee Species using ABIS. Proceedings of Sustainability in theInformation Society, Conf. on Environmental Informatics, Oct. 10 - 12 2001, ETH Zürich.

196
Project ID: 01LC9904 (EDIS, Subproject DNATAX 1)1.7.2000 – 31.3.2003
DNATAX – DNA TAXONOMY AS A TOOL IN BIODIVERSITY RESEARCH:RECORDING, ASSESSMENT, MONITORING AND UTILIZATION OF THEGENETIC DIVERSITY OF EUKARYONT SYSTEMS
A. Hille, M. A. Miller, and G. Haszprunar
Zoologische Staatssammlung München, Münchhausenstr. 21, D-81247 Mü[email protected]; http://www.zsm.mwn.de/wiss_start.htm
Key words: insects, DNA sequencing, taxonomy, molecular systematics, database, biodiversityinformatics
Abstract:The DNATAX project aims at compiling diagnostic DNA signatures for the main orders of insects.It focuses on the elaboration of phylogenetically informative DNA sequences (e. g. for Geometridmoths, INGE). Data are organized and streamlined in a database on the World Wide Web(SysTax, Ulm) for more efficient use in taxonomic and systematic basic research.
Introduction. In species-rich groups like the insects, which are represented in the environmentand in collections by millions of specimens and tenthousands of species, accuracy of identificationoften is a serious problem for taxonomists and even more for the non-specialist scientists inapplied zoology, especially dealing with very small species or immature stages of insects (eggsand various kinds of larvae). In addition, accurate phylogenetic trees provide a sound basis formany questions of evolutionary biology concerning species discrimination and speciation,biogeographic and ancestry-descendance relationships, but also concern important applied aspectslike pest control and bio-indication. DNA-Taxonomy provides tools to cover both – identificationand phylogeny.In the initiation phase of the project a complete molecular lab including an ABI377 automatedsequencer was established at the Zoologische Staatssammlung München (ZSM), an importantinstitution holding natural history collections. The laboratory devices are intended for high-throughput sequencing which allows operating on different molecular markers such asmitochondrial (mt) or nuclear (c) DNA gene fragments. The broader relevance of molecular-aidedtaxonomy is to obtain clearly defined taxa. Extensive DNA surveys of species limits andgeographic lineage distribution, e. g. delimiting taxonomic units in species complexes are targetedby dense taxon sampling of morphologically highly variable geographic populations. A secondimportant aim is to solve long pending problems in taxonomic classification, systematics, andphylogeny.Results. At the start of the project protocols for DNA extractions from fresh and archivalbiomaterials were tested and optimised for routine DNA typing. Genomic DNA (over 600 probes)was isolated from either single legs or plugs taken from the thorax of single dried insects, or fromdead animals recovered from the field (pheromone traps, ethanol-fixed fresh specimens). To linkan individual to a source (a voucher, frozen biomaterial like tissue) by DNA typing, the repositoryof extracted DNA and its sources are curated in a resource bank at the ZSM, which allowsapplications for studying genetic variation, phylogeny, and the processes underlying diversity,such as gene flow. Laser-scanned label data on these repositories and annotation information onthe voucher specimen can simply be exported in tabular form to secondary composite databaseslike SysTax using controlled vocabulary.Since recommendations made by Caterino et al. [1] outline that “the cytochrome oxidase I, 16S,18S, and elongation factor -1∀.[...] are informative across a broad range of divergences in insects

197
[..]”, PCR-amplified fragments of these genes (more than 1200 contigs) are also used in oursequence efforts to identify species and lineages. These genes provide potent discriminators ofphylogenetic relationships for newly determined sequences for insects, and are a valuable asset tothe prospect of a global comparability with the substantial database that already exists for thesegenes in the principal DNA sequence databases like NCBI-GenBank/EMBL-EBI. In order toexploit the folding structure of RNA molecules in an approach called “molecular morphometry”,the 16S like mitochondrial ribosomal RNA is now also routinely sequenced for the studied taxa(100 alignable sequences). Sequences are stored in ARB (see this volume), and compared to itsstructural alignments preheld in the “pt_Server” to recognize sequence and fold patterns using oneof the secondary structure models of the SSU rRNA. Several sequencing projects are elaboratedso far focussing on taxon description [2], identification of critical species [3], test of conspecifityof allopatric populations [4], and molecular systematics [5]. Others are under way by cooperationwith national (Museum Koenig, Bonn, Lepidoptera: genus Zygaena, [6]; ZSM, Lepidoptera:Geometridae, Drepanidae, Diptera: Diopsidae, Hymenoptera: genus Euglossa; University ofJena, [7], University of Bremen, MOLART) and international partners (Zoological Museum, Dep.Entomol., Copenhagen, Diptera: Sepsidae, and genus Themira).Submission of sequence data and associated biological information to the EMBL database at EBIusing the WWW site WEBIN is a standard practice prior to publications [2-5]. The uniqueaccession number assigned by the database which permanently identifies the sequence submitted,is cross linked by queries made by SysTax. This interoperability using compressed versions ofGenBank’s feature table flat-file format ensures availability and distribution of new sequencedata.Phylogenetic trees as graphical representations of the putative evolutionary relationship betweengroups of organisms, e.g. as calculated from multiple nucleic acid sequence alignment can begenerated by the specialized treeing programme tools of ARB. Using its tree-building functionsaccording to an optimised maximum parsimony criterion, homologies between novel querysequences (i. e. of a newly sequenced animal) and stored database alignments are identified by asearching strategy which finds similar alignment positions in local sub trees. Nearest neighbourterminals of localized sub trees may be interchanged during the optimisation fit, while the globaltree topology is maintained.
References:1. Caterino, M. S., Soowon Cho, and Sperling, F. A. H. 2000: The current state of Insect MolecularSystematics: A thriving tower of Babel. Annu. Rev. Entomol., 45, 1-45.2. Buchsbaum, U. and Miller, M. A. (in prep.): Cerace diehli sp. n. from Sumatra (Indonesia) (Lepidoptera,Tortricidae, Tortricinae). Heterocera Sumatrana.3. Hille, A. and Miller, M. A. (in prep.): DNA sequence variation at the mitochondrial cytochrome oxidaseI subunit among pheromotypes of the sibling taxa Diachrysia chrysitis (LINNAEUS, 1758) and D. tutti(KOSTROWICKI, 1961) (Lepidoptera: Noctuidae).4. Spies, M., Sublette, J. E., Sublette, M. F., Wülker, W. F., Martin, J., Hille, A., and Miller, M. A. (inprint): Pan-American Chironomus calligraphus Goeldi (Diptera, Chironomidae): species or complex?Evidence from external morphology, karyology and DNA sequencing. Aquatic Insects, 24(4).5. Miller, M., Hausmann, A., and Trusch, R. 2001: The phylogenetic relationships in Geometrid moths. Anapproach using mitochondrial DNA (mtDNA) sequences. Spixiana, 24 (3), in print.6. Hille, A., Miller, M. A., Edelmann, A., and Naumann, C. M. (in prep.): Biochemical and MolecularSystematics of selected taxa of the genus Zygaena (Zygaenidae, Lep.).7. Erlacher, S., Trusch, R., Hille, A., and Miller, M. A. (in prep.): Phylogeny of the geometrid moth taxonDyscia inferred from morphological data and nucleotide sequences of the mitochondrial partial ND1, 16Sand COXI genes (Lepidoptera, Geometridae).

198
Project ID: 01LC9904 (EDIS, Subproject DNATAX 2 - MOLART)1.4.2000 – 31.3.2003 1
MOLART (DNATAX 2): MOLECULAR IDENTIFICATION OF COLLEMBOLA:METHOD OPTIMISATION VERIFICATION - DOCUMENTATION
J. Filser1, U. Burkhardt1, and A. Fjellberg2
1 University of Bremen, Universität Bremen, UFT - Abt. für allgemeine und theoretischeÖkologie, Leobener Str., D-28334 Bremen2 Entomological Research, Ormeletveien 53, 3145 Tjoeme, [email protected]
Key words : Collembola, Nordic species, biogeography, Protaphorura armata group, Isotomaviridis group, DNA, molecular methods
Abstract:MOLART deals with the development and standardisation of molecular methods for safe andefficient identification of species which cannot (or only with great effort and expertise) bedistinguished by morphological analysis. Different molecular techniques (such as DNAsequencing, AFLP, RAPD, allozymes) are applied, refined and evaluated according to theirdiagnostic power, their efficiency for different taxa, the necessary effort of time and therequired experience This is exemplified by two problematic species groups within theCollembola (Insecta, Apterygota), the Protaphorura armata group and the Isotoma viridisgroup. To ensure precise identification, the project is conducted in close cooperation with aninternational expert in Collembola taxonomy - Arne Fjellberg - who will also providespecimens of further Collembola groups for DNA-Tax 1, a database of his collection onNordic Collembola and volume 2 of his identification key for Collembola of Fennoscandiaand Denmark. At present, 9 genera of the family Isotomidae have been finished. Formolecular analyses, a number of sites have been sampled, the first laboratory cultures havebeen started, and the DNA techniques were established. First analyses revealed a) differenceswithin three species of the I. viridis group, b) restriction enzymes that could be used for quickspecies distinction, and c) that the power of the methods used so far differs between the twospecies groups investigated.
Results:
1. IdentificationThe first nine months of the project have mainly dealt with extending and updating theidentification key (monograph "The Collembola of Fennoscandia and Denmark"), building uplaboratory infrastructure, establishing cooperations, collecting animals and establishingcultures.Concerning the identification key, 9 genera of the family Isotomidae have been finished (incl.one n.sp. of Folsomia). The rest of the family (incl. 3 n.sp.) will be finished by the end of nextyear, together with the family Entomobryidae. The work for the remaining families of theEntomobryomorpha and Symphypleona will be conducted in 2003. The final Vol. 2 of themonography is expected to contain a total number of 223 species.Representatives of the genus Protaphorura and the Isotoma viridis group have been collectedfrom 14 sites in Germany, Denmark and Norway. The live catches are now being used forestablishing laboratory cultures to obtain sufficient animal numbers for molecular biological 1 The project started only in January 2001

199
analyses. Thus far, DNA has been extracted, amplified and purified mostly from I. viridis s.l.(whole individuals, except for the furca which has been preserved for taxonomic check).Studies with four primer pairs out of subunits of the mitochondrial cytochrome oxidase gene(CO1-fa, CO1-ru, CO2-ab, CO3-ab) revealed CO2-ab to be most appropriate for I. viridis s.l.,CO3-ab for Protaphorura sp.. The first individuals have been sequenced; new results areproduced continuously. Based on them, appropriate restriction sites have been selected whichcan be used for displaying haplotype-specific length polymorphisms on agarose gels: a firststep towards easy, quick, affordable and safe molecular identification (Fig. 1)
Fig. 1: An example for improving molecular identification of Collembola within MOLART
2. Interoperability and Linking of DatabasesThe "collection Fjellberg" comprises about 13,000 records of Nordic Collembola, all of themincluding locality, habitat, date, collector, literature source and synonyms. The data are readyfor import to Systax.
3. Life in time and space: collections as biodiversity information resourcesMOLART is specialised in Collembola (Insecta, Apterygota), with special respect to theNordic fauna. Here, one of the most complete collections worldwide is the "collectionFjellberg" which will be provided as a database by this project.
References:1. Filser, J. 1999: Habitat requirements and ecology of Isotoma viridis Bourlet, 1839 and Isotomaanglicana Lubbock, 1862 (Insecta, Collembola). Braunschweiger Naturkundliche Schriften, 5/4, 905-911.2. Fjellberg, A. 1998: The Collembola of Fennoscandia and Denmark. Part I: Poduromorpha. FaunaEntomologica Scandinavica 35, Brill, Leiden, Boston, Köln, 184pp.3. Frati, F. & Carapelli, A. 1999. An assessment of the value of nuclear and mitochondrial genes inelucidating the origin and evolution of Isotoma klovstadi Carpenter (Insecta, Collembola). AntarcticScience 11: 160-174.4. Simonsen, V., Filser, J., Krogh, P.H. & Fjellberg, A. 1999. Three species of Isotoma (Collembola,Isotomidae) based on morphology, isozymes and ecology. Zoologica Scripta 28, 281-287.5. Zabeau, M. & Vos, P. 1993. Selective Restriction Fragment Amplification: A general method for DNAfingerprinting. European Patent Application EP 0534858.
Present technique
1. Field collection2. Microscopic identification3. Establishment of laboratory cultures4. Separation of furca --> (2)5. DNA extraction, amplification, purification6. Sequencing7. Identification of haplotype-specific restriction
sites8. Design of specific restriction enzymes9. Specific fragment length polymorphism on
agarose gels
Future technique: easy and quickidentification
1. Field collection
5. DNA extraction, amplification,purification
9. Specific fragment lengthpolymorphism on agarose gels

200
Project ID: 01LC9904 (EDIS, Subproject DORSA)1.4.2000 – 31.3.2003
DORSA - GERMAN ORTHOPTERA COLLECTIONS
S. Ingrisch, K.-H. Lampe, K. Riede , and C. Dietrich
Zoologisches Forschungsinstitut & Museum Alexander Koenig (ZFMK),Adenauerallee 150-164, D-53113 [email protected]; http://www.dorsa.de
Key words: specimen database, geographical information systems, bio-acoustics, automated songrecognition, documentation of type specimens
Abstract:The DORSA subproject (abbreviation for "Deutsche Orthopteren-Sammlungen" = GermanOrthoptera Collections) is a specimen based Internet database of the Orthoptera held in Germanmuseum collections including illustrations of type specimens, sound recordings, and geographicinformation on a world-wide basis ("virtual museum"). A rapid assessment tool is developed forautomated song recognition.
Results:The aim of DORSA is to provide web access to (i) specimen information of Orthoptera inGerman collections, (ii) a virtual Orthoptera collection with link to the Orthoptera Species FileOnline [1], (iii) a photographic documentation of type specimens and other important specimens,and (iv) the wealth of acoustic data of Orthoptera in German institutions and private collections.Geo-referencing and combining the locality data with a geographical information system will betackled further on.Specimens databased up to now include type specimens (holo-, syn- and paratypes; see table 1), aselection of specimens on which important monographs were based, and voucher specimens forwhich sound recordings are available. The project covers collections held in the followingmuseums: Berlin (ZMHB), Bonn (ZFMK), Dresden (SMTD), Eberswalde (DEI), Frankfurt(SMFD), Halle (MLUH), Hamburg (ZSMH), München (ZSM), and Stuttgart (SMNS). Theprivate collections of Heller, Ingrisch, Riede and G.H. Schmidt will also be included. Databasingof specimens has been completed in the museums of Dresden, Eberswalde, and Hamburg, startedbut not yet completed in the other collections. The current state is documented in table 1. To tracethe current locality for a given type specimen is often difficult due to incompletely publishedinformation, insufficient labelling of specimens, exchange between museums, destruction ofspecimens (i.e. in war), or "prolonged" loans. DORSA helps to improve the knowledge of thecurrent type depositories. Information on the primary types of 424 species could so far be updatedas compared with the Orthoptera Species File (OSF). From the taxonomic point of view, thefocus was so far on the Tetrigoidea (354 species, 1014 specimens), Gryllacridoidea (257 [331with Halle, Stuttgart and Munich included] species, 467 [593] specimens), and Grylloidea (323[341 with Stuttgart and Munich included] species, 619 [655] specimens). The inventory data forthose three groups are ready for import into the all-biology database SysTax.12,000 photographs have so far been taken to document the specimens and the labelling of thespecimens; about half of the photographs are already processed for web publishing.For all specimens with reliable locality information, collection sites are geo-referenced bylatitude/longitude co-ordinates. This allows mapping by any geographical information system(GIS) and many additional applications, such as intersection with environmental data. A firstprototype for a Java-based graphical user interface can be found at www.groms.de. It allows
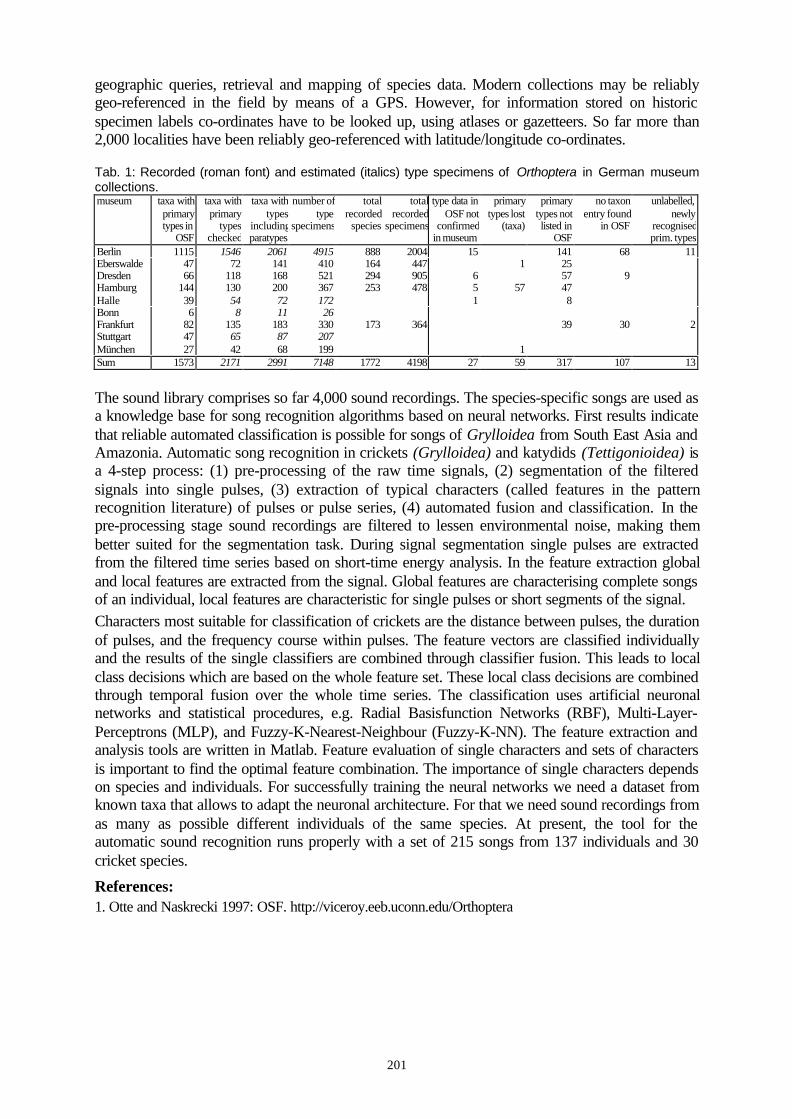
201
geographic queries, retrieval and mapping of species data. Modern collections may be reliablygeo-referenced in the field by means of a GPS. However, for information stored on historicspecimen labels co-ordinates have to be looked up, using atlases or gazetteers. So far more than2,000 localities have been reliably geo-referenced with latitude/longitude co-ordinates.
Tab. 1: Recorded (roman font) and estimated (italics) type specimens of Orthoptera in German museumcollections.museum taxa with
primarytypes in
OSF
taxa withprimary
typeschecked
taxa withtypes
includingparatypes
number oftype
specimens
totalrecorded
species
totalrecorded
specimens
type data inOSF not
confirmedin museum
primarytypes lost
(taxa)
primarytypes notlisted in
OSF
no taxonentry found
in OSF
unlabelled,newly
recognisedprim. types
Berlin 1115 1546 2061 4915 888 2004 15 141 68 11Eberswalde 47 72 141 410 164 447 1 25Dresden 66 118 168 521 294 905 6 57 9Hamburg 144 130 200 367 253 478 5 57 47Halle 39 54 72 172 1 8Bonn 6 8 11 26Frankfurt 82 135 183 330 173 364 39 30 2Stuttgart 47 65 87 207München 27 42 68 199 1Sum 1573 2171 2991 7148 1772 4198 27 59 317 107 13
The sound library comprises so far 4,000 sound recordings. The species-specific songs are used asa knowledge base for song recognition algorithms based on neural networks. First results indicatethat reliable automated classification is possible for songs of Grylloidea from South East Asia andAmazonia. Automatic song recognition in crickets (Grylloidea) and katydids (Tettigonioidea) isa 4-step process: (1) pre-processing of the raw time signals, (2) segmentation of the filteredsignals into single pulses, (3) extraction of typical characters (called features in the patternrecognition literature) of pulses or pulse series, (4) automated fusion and classification. In thepre-processing stage sound recordings are filtered to lessen environmental noise, making thembetter suited for the segmentation task. During signal segmentation single pulses are extractedfrom the filtered time series based on short-time energy analysis. In the feature extraction globaland local features are extracted from the signal. Global features are characterising complete songsof an individual, local features are characteristic for single pulses or short segments of the signal.Characters most suitable for classification of crickets are the distance between pulses, the durationof pulses, and the frequency course within pulses. The feature vectors are classified individuallyand the results of the single classifiers are combined through classifier fusion. This leads to localclass decisions which are based on the whole feature set. These local class decisions are combinedthrough temporal fusion over the whole time series. The classification uses artificial neuronalnetworks and statistical procedures, e.g. Radial Basisfunction Networks (RBF), Multi-Layer-Perceptrons (MLP), and Fuzzy-K-Nearest-Neighbour (Fuzzy-K-NN). The feature extraction andanalysis tools are written in Matlab. Feature evaluation of single characters and sets of charactersis important to find the optimal feature combination. The importance of single characters dependson species and individuals. For successfully training the neural networks we need a dataset fromknown taxa that allows to adapt the neuronal architecture. For that we need sound recordings fromas many as possible different individuals of the same species. At present, the tool for theautomatic sound recognition runs properly with a set of 215 songs from 137 individuals and 30cricket species.
References:1. Otte and Naskrecki 1997: OSF. http://viceroy.eeb.uconn.edu/Orthoptera

202
Project ID: 01LC9904 (EDIS, Subproject GART)1.4.2000 – 31.03.2003
GART – GLOBAL SPECIES REGISTER BUTTERFLIES
C. L. Häuser, J. Holstein, and A. Steiner
State Museum of Natural History, Rosenstein 1, D-70191 [email protected] , [email protected],[email protected]; http://www.insects-online.de/projfram.htm
Key words: biodiversity informatics, taxonomic databases, type specimens, world checklist,Lepidoptera, Papilionoidea, Papilionidae, Pieridae
Abstract:The GART project’s aim is to provide general access to information about the world’s butterfliesbased on global taxonomic catalogues. In a first step, global species registers were developed fortwo families of butterflies, Swallowtails (Papilionidae) and Whites (Pieridae), based oncomprehensive taxonomic databases. In addition to extending the taxonomic database, allbutterfly primary type specimens housed in the collections of the co-operating GART partnerinstitutions will be recorded, photographed, and databased. For selected taxa within thePapilionidae and Pieridae, a comprehensive species information system will be developedoffering data about distribution, ecology, and morphology of individual species as well as links toother existing data sources. The species register and the underlying taxonomic and type specimendatabases will be made publicly available through a central user interface on the Internet via theEDIS framework project.
Results:The GART project is developing a worldwide information system for butterflies (Papilionoidea).It is based on taxonomic and bibliographic databases, which will contain information on alldescribed butterfly taxa, including scans of the original descriptions, illustrations, and onphotographs of the primary type specimens in institutional collections.GART is jointly undertaken by partners from eight major German natural history museums:Staatliches Museum für Naturkunde (SMNK), Karlsruhe; Deutsches Entomologisches Institut(DEI), Eberswalde; Zoologische Staatssammlung München (ZSM); Museum für Naturkunde derHumboldt Universität (MNHU), Berlin; Forschungsinstitut und Naturmuseum Senckenberg(FIS), Frankfurt; Staatliche Naturhistorische Sammlungen Dresden, Museum für Tierkunde(SMTD); Zoologisches Forschungsinstitut und Museum Alexander Koenig (ZFMK), Bonn, andis co-ordinated at the State Museum of Natural History, Stuttgart (SMNS). The project is alsointended to provide a distinct contribution to the international 'Global Butterfly InformationSystem' (GloBIS) initiative, to be developed jointly by SMNS and several international researchcollections [1-2].In collaboration with leading butterfly taxonomists GART has developed annotated globalchecklists of the families Papilionidae and Pieridae, which include all currently recognizedspecies names and important synonyms. These lists are already available via the WWW and willbe periodically updated and maintained by a group of international specialists [3].The GART taxonomic database includes information on the original description, primary andrecent taxonomic status, taxonomic history, type locality (both original and subsequentinformation), type status as well as primary type material (including condition and currentdepository). Each record is linked to a literature database and to specimen and label photographs.
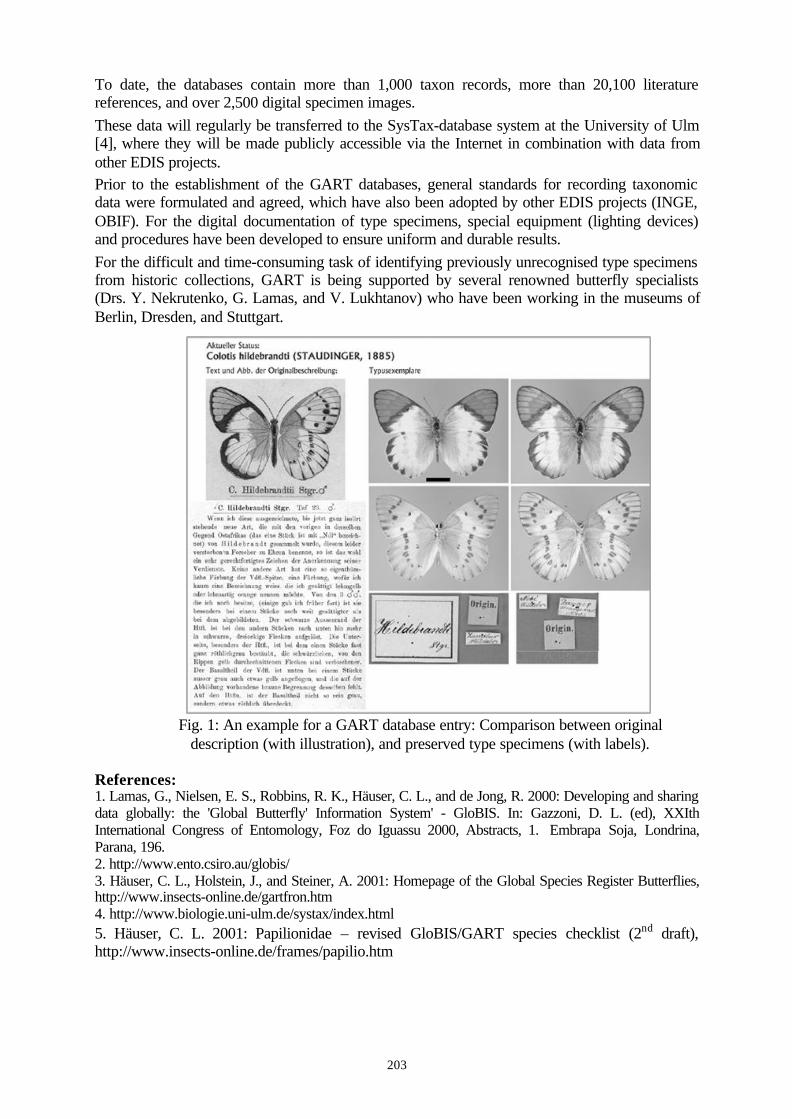
203
To date, the databases contain more than 1,000 taxon records, more than 20,100 literaturereferences, and over 2,500 digital specimen images.These data will regularly be transferred to the SysTax-database system at the University of Ulm[4], where they will be made publicly accessible via the Internet in combination with data fromother EDIS projects.Prior to the establishment of the GART databases, general standards for recording taxonomicdata were formulated and agreed, which have also been adopted by other EDIS projects (INGE,OBIF). For the digital documentation of type specimens, special equipment (lighting devices)and procedures have been developed to ensure uniform and durable results.For the difficult and time-consuming task of identifying previously unrecognised type specimensfrom historic collections, GART is being supported by several renowned butterfly specialists(Drs. Y. Nekrutenko, G. Lamas, and V. Lukhtanov) who have been working in the museums ofBerlin, Dresden, and Stuttgart.
Fig. 1: An example for a GART database entry: Comparison between originaldescription (with illustration), and preserved type specimens (with labels).
References:1. Lamas, G., Nielsen, E. S., Robbins, R. K., Häuser, C. L., and de Jong, R. 2000: Developing and sharingdata globally: the 'Global Butterfly' Information System' - GloBIS. In: Gazzoni, D. L. (ed), XXIthInternational Congress of Entomology, Foz do Iguassu 2000, Abstracts, 1. Embrapa Soja, Londrina,Parana, 196.2. http://www.ento.csiro.au/globis/3. Häuser, C. L., Holstein, J., and Steiner, A. 2001: Homepage of the Global Species Register Butterflies,http://www.insects-online.de/gartfron.htm4. http://www.biologie.uni-ulm.de/systax/index.html5. Häuser, C. L. 2001: Papilionidae – revised GloBIS/GART species checklist (2nd draft),http://www.insects-online.de/frames/papilio.htm

204
Project ID: 01LC9904 (EDIS, Subproject INGE)1.4.2000 – 31.3.2003
INGE – INVENTORY OF GEOMETRID MOTHS OF EUROPE (INSECTA,LEPIDOPTERA): PROVIDING MULTIMEDIAL DATA ON SYSTEMATICS,ZOOGEOGRAPHY AND ECOLOGY
A. Hausmann and R. Trusch
Zoologische Staatssammlung München, Münchhausenstr. 21, D-81247 Mü[email protected]; http://www.zsm.mwn.de/Lepidoptera/Biolog/index.htm
Key words: Geometridae, Europe, systematics, zoogeography, ecology, multimedial publication
Abstract:The main purposes of the INGE project are the critical examination and the coordinatedcompilation of available information on systematics, zoogeography, and ecology of the EuropeanGeometrid Moth species (Lepidoptera). The data are published using different media, such as abook series (Apollo Books, Stenstrup, Vol. 1 published in 3/2001), CD-ROM (Expert-center forTaxonomic Identification, ETI, Amsterdam, data partially submitted) and the Internet (SysTax,Ulm, data partially submitted). The results will offer easily accessible identification tools andbiodiversity databases which include a virtual type collection and will greatly reduce the need ofsearching for material scattered widely in many institutions. Geometrids being one of the largestand worldwide distributed insect groups, INGE can serve as a model for modern methods andachievements in measuring and conserving biodiversity.
Results:In the first half of the project period collection-based biodiversity data have been compiled to gainthe information needed on morphology, distribution patterns and bionomics of EuropeanGeometridae (Archiearinae-Sterrhinae). The Zoologische Staatssammlung München (ZSM)houses one of the world’s largest collections of moths including about 200,000 specimens of thesubfamilies focused on by the project. Almost half of them have been investigated so far.Microscopic slides of about 3,000 dissected specimens served as the base for morphologicalstudies and for 130 line drawings. Additionally, material from most major European collectionswas examined (about 100,000 specimens). Distribution maps reflecting collection data have beencreated for 120 species (of 240).Quality control of all taxonomic information is one of the crucial aims of this monographicproject. In this respect it overlaps with the EDIS–GART project, which established high standardsin data-collecting and data-processing. Applying those standards, original descriptions of morethan 1,000 taxa have been checked and largely databased. Interdisciplinary interaction with theDNA-TAX projects helps to evaluate the status of taxa at the species level (e.g. validity at speciesrank of Timandra comae has been confirmed), as well as on generic level (e.g. verification ofsistergroup-relationship of the tribes Cosymbiini and Timandrini; synonymy of Glossotrophia andScopula) [1]. To date, DNA-information has been provided for about 200 specimens ofGeometridae.The first volume of the series „The Geometrid Moths of Europe“ [2] has already been published(282 pp.). The project is proceeding regularly and both Vol. 2 and the interactive CD-ROMs (byETI) will be published in 2003, as planned.Taxon-based data (checklists) from these publications are transferred via the GART-database intoSysTax, where they will be made accessible via the Internet. Basic taxonomic information on thespecies under focus has already been submitted to SysTax. Furthermore, an object-database

205
contains photos and object (label) data of primary type specimens and other characteristicindividuals of all 240 European species of the subfamilies Geometrinae and Sterrhinae. 340specimens are databased this way (partially submitted to SysTax).Interactions and co-operations: The INGE project is a core activity crucial for other research.Alterations of feeding behaviour of certain Lepidoptera (e.g. Cameraria ohridella, Gracillariidae)can cause dramatic problems. DNA analysis of populations or subspecies with regional host-plantpreferences may provide a basis for the prevention and biological control of pests [3].Prerequisites are exact knowledge of regional host-plant utilisation linked with a securetaxonomy, combined with molecular genetic characterisation of properly determined specimens.In this respect INGE can benefit from the international project Geometrid Moths of Europe, theResearch Initiative Herbulot [4], and the facilities of the Molecular Research Laboratory at theZSM, a rather unique constellation in Germany.
Fig. 1: Information flow and publication activities in the INGE project
References:1. Miller, M., Hausmann, A., and Trusch, R. 2001: The phylogenetic relationships in Geometrid moths. Anapproach using mitochondrial DNA (mtDNA) sequences. Spixiana, 24 (3), in print.2. Hausmann, A., 2001: Introduction. Archiearinae, Orthostixinae, Desmobathrinae, Alsophilinae,Geometrinae. In: A. Hausmann (ed.), The Geometrid Moths of Europe, 1, 1-282.3. Hausmann, A. und Trusch, R. (in print): Proceedings of the FORUM HERBULOT 2001. Spixiana 24.4. http://www.herbulot.de

206
Project ID: 01LC9904 (EDIS, Subproject OBIF)1.4.2000 – 31.03.2003
OBIF - OPTIMIZATION OF BIODIVERSITY INFORMATION FACILITIES ONAPPLICATION-ORIENTED RESEARCH
M. Verhaagh, J. Spelda, C. Wurst, L. Beck, N. Blüthgen, F. Brechtel, W. Hanagarth, H.Höfer, F. Meyer, and S. Woas
State Museum of Natural History Karlsruhe (SMNK), Erbprinzenstr.13, D-76133 [email protected]; http://www.naturkundemuseum-karlsruhe.de/obif.htm
Key words: biodiversity information, natural history museum collections, digitalisation,interactive key, Oribatida, Araneae, Formicidae, Buprestidae
Abstract:The OBIF project aims at optimising the input of - and access to - biodiversity data from researchprojects and collections at the State Museum of Natural History Karlsruhe. It combines electronicidentification keys and morphological characters, information on collections sites anddistribution of species, ecological data, text information, references with line drawings, digitallight microscopy and REM-photos, as well as video sequences into an easy-to-use informationsystem for the scientific and public user on the Internet and in the museum.
IntroductionNatural History Museums are traditionally devoted to recording and describing, i.e. inventorying,the biological diversity on earth. Therefore, they store and administer extensive scientificcollections and distribute knowledge on natural history to the public.OBIF has the objective to optimise the in- and output of biodiversity data at the State Museum ofNatural History Karlsruhe. Scientists and technical assistants need the data internally for researchand for the administration of the collection. On the other hand, biodiversity data is also to bedistributed to external users such as scientists in other fields, NGOs, consultants, or the interestedpublic. To that end, the project improves access to existing databases in the museum. It alsocooperates in the construction of an extensive internet-accessible information system (SysTax)with contributions on several soil and wood inhabiting groups of animals. The geographical focuswas set on the Palearctic and Neotropical regions, and the taxa chosen (Oribatid mites, spiders,ants, and jewel beetles) are those where years of study have already generated a broad base ofinformation at the SMNK. The system will include information on collection sites and thedistribution of species as well as on morphological characters and data of their natural history andecology. Information will be provided as text, literature references, line drawings, digital lightmicroscopy and REM-photos, and also as video sequences where convenient. As a service to thenon-specialist user we combine the databased information with easy-to-use digital interactive keysin DELTA-format [1-4].
Acari: OribatidaThe Oribatida are a group of soil mites. Worldwide about 5,000 species have been described,with 500 of them occurring in Central Europe. Due to their detritivorous habits, they play animportant functional role in many natural ecosystems. Though of great ecological interest,identification is currently not possible without the help of a specialist. Descriptions and ecologicalinformation are scattered over a large number of publications, which are often difficult to obtain.Most of the descriptions available are not suitable for re-identification. Furthermore, the highlyspecialized terminology for their external morphology deters students from getting familiar withthis group.

207
For this reason, we provide non-verbose interactive keys based on the DELTA-System. Thefeatures of our identification system are:1. digital images on light microscopy instead of alienating scans made by electron microscopy2. colour markings of diagnostic characters in figures3. easy-to-use terminologyIn addition, taxonomic, bio-geographical, and ecological information will be provided bydatabases connecting literature data to species names. This enables an economic evaluation ofdata.
Hymenoptera: FormicidaeIn a pilot-project a database will be combined with an interactive DELTA-key to carpenter antspecies of the genus Camponotus with present and/or historical distributions in Baden-Württemberg (BW, South-West Germany). The key will be illustrated by line-drawings, REM-and light microscopy photos, and in some cases video sequences of living animals, and thus be ofeasy use for specialists as well as non-specialists. The database (SysTax) will include informationon the specimens from the collection of the SMNK (date, sex, locality, habitat, etc.); generalinformation on the ant species like its distribution in BW; literature references for the species; anddata on individual morphological characters, which can be compared with the species’ standarddescriptors used in the interactive key.
Coleoptera: BuprestidaeThe metallic wood-boring or jewel beetles are a group of insects with worldwide distribution,mainly in the tropics, but at least 80 species can be found in Central Europe. Some of the morefrequent species may cause harm to timber, as they feed on wood during their larval period, likemany species of the family. Others, however, are threatened species protected by law.Consequently, there is a high demand from foresters and conservationists for identifications ofadult beetles and their larvae.The latter can be identified with the help of their pronounced traces, i.e. characteristic courses oftunnels they originate while feeding: some species undermine the cambial layers of branches andtwigs, some bore deeply into the wood. Furthermore, the imago’s hatching hole is of characteristicoutline and enables identification on generic level. Especially if one knows about the feedingplant, its age and climatic exposition, identification even to species level is possible with the helpof these traces. However, the information about these features is scattered in the literature, and sospecialists with much field experience are needed.We present a DELTA-based key exemplifying different ways of identification in combininginformation on feeding plants, features of larval tunnels and of hatching holes, and characters ofthe beetles themselves, thus creating a tool operable by users without expert knowledge.
References:1. Dallwitz, M. J. 1980: A general system for coding taxonomic descriptions. Taxon, 29, 41–6.2. Dallwitz, M. J., Paine, T. A., and Zurcher, E. J. 1993: ‘User’s Guide to the DELTA System: a GeneralSystem for Processing Taxonomic Descriptions.’ 4th edition.3. http://biodiversity.uno.edu/delta/4. ftp://ftp.ento.csiro.au/delta/

208
Project ID: 01LC9905 (GLOPP)1.4.2000-31.3.2003
GLOPP - GLOBAL INFORMATION SYSTEM FOR THE BIODIVERSITY OFPLANT PATHOGENIC FUNGI
G. Hagedorn1, G. Deml1, D. Triebel2, M. Piepenbring3, and F. Oberwinkler4
1 Institut für Pflanzenvirologie, Mikrobiologie und Biologische Sicherheit, Biologische Bundesanstalt für Land- und Forstwirtschaft, Königin-Luise-Str. 19, D-14195 Berlin2 Botanische Staatssammlung München, Menzinger Straße 67, D-80638 München3 Botanisches Institut, Universität Frankfurt, Senckenberganlage 31-33, D-60054 Frankfurt/M.4 Lehrstuhl Spezielle Botanik/Mykologie, Botanisches Institut, Universität Tübingen, Auf der Morgenstelle 1, D-72076 Tü[email protected]
Key words: GLOPP, information systems, fungi, plant pathogens, biodiversity,DiversityWorkbench, biodiversity informatics
Abstract:The biodiversity research of several generations of scientists has produced a huge treasure ofbiological specimen collections, as well as descriptions and reports of new or known organisms.Today, this knowledge should be made readily available in the form of digital media. Anintegrated information system will greatly improve the efficiency of the work of both the scientistsand the applied users in agriculture and forestry.
Introduction:The GLOPP project ("Global Information System for the Biodiversity of Plant PathogenicFungi") aims to provide a unified view of data about fungi parasitizing higher plants. Theinformation system integrates data on host plant specificity, geographical distribution, and a coreset of descriptive characters that allow an interactive online identification. Since our resources arelimited, European pathogens will be treated with priority, to guarantee high data quality and acertain completeness, but it is planned to continue to extend the scope within a framework ofinternational, collaborative projects in the future.The information system that is currently being developed goes beyond a simple compilation. It isplanned as a critical scientific revision of the available data. Uncritical acceptance ofmisidentifications and degeneration of data quality will thus be avoided. Original information iskept strictly separate from assessments and interpretations. It will be possible to document the(possibly contradictory) assessments of multiple scientists. The system allows the onlineinteractive identification of plant pathogenic fungi and a retrieval of relevant information, as wellas analyses that increase our understanding of biodiversity. Examples are the diversity of majortaxonomic groups, the specificity of host-pathogen relations, co-evolutionary processes, ortemporal and spatial dynamics in the distribution of pathogens. The project also has aconsiderable socio-economic impact due to the large number of economically importantpathogens.The system is designed to enable farmers or the general public to identify fungal pathogenswithout having to study the complete list of characters and without access to the scientificliterature. For example, a query might be: "Show all parasites that belong to imperfect fungi,which have been found on Rhododendron in Germany, and which have oblong, aseptate spores, 4x 10 µm large" (fig. 1). The information system would try to find all fungi that fulfil the givencriteria based on past experience (i.e. information published in the literature or in herbaria). The

209
system could provide error tolerant usage, by adding a second result list, which includes allrecords from the entire region (i.e. Europe), which have been found on a species in the Ericaceae,and includes fungi that fulfil the descriptive criteria only approximately. At the end of the projectthe information system will be available on the Internet free of charge.
Fig. 1: Prototype for a web interface to use the GLOPP integrated information systemfor the identification of plant pathogenic fungi

210
Project ID: 01LC9905 (GLOPP, Subproject IT)1.4.2000 - 31.3.2003
GLOPP-IT - BIOINFORMATICS AND DEVELOPMENT OF THE INFORMATIONSYSTEM
T. Gräfenhan, M. Weiss, M. Gliech, and G. Hagedorn
Institut für Pflanzenvirologie, Mikrobiologie und Biologische Sicherheit, BiologischeBundesanstalt für Land- und Forstwirtschaft, Königin-Luise-Str. 19, D-14195 [email protected]; http://www.DiversityCampus.net
Key words: GLOPP, information systems, fungi, plant pathogens, identification, bioinformatics,biodiversity, xml-schema, DiversityWorkbench, DiversityCollection, DiversityReferences,DeltaAccess, biodiversity informatics
Abstract:GLOPP-IT is creating an integrated information system on the biodiversity of plant pathogenicfungi and their hosts by gathering descriptive and collection data, literature references, resourceinformation and taxonomic names. The project provides a set of application modules to otherprojects within GLOPP and develops the public access system on the Internet.
Results:Many current solutions to manage biodiversity data (e.g. SysTax, Specify, BioLink) are based ona highly complex, monolithic information model with different tasks tightly connected. Thesemodels are difficult to analyse and implement. The GLOPP IT project has therefore investedconsiderable time and effort to design a modular information model, where the various modulescan be implemented separately and are exchangeable with 3rd party modules. Interface definitionsallow to define associations between the central biological objects of the modules. This modularstructure (see fig. 1) provides the basis for a customisable system that can be configured to suit thedemands of scientists and other users. A data exchange format based on XML-schemata iscurrently under development. Preliminary versions can be downloaded from the Internet [5-8].DiversityCollection was designed to capture label data from specimens of plant pathogens in theherbaria of Munich and Halle/S. (GLOPP-Erysiphales). DiversityReferences manages scientificliterature citations; DiversityTaxonomy records the nomenclature and synonymy of scientificnames and organizes them into (potentially multiple) taxonomic hierarchies.The information from these three central modules is used in the two modules collecting the datafor the identification process. DiversityDescriptions (derived from DeltaAccess version 1.7)represents a complete tool to manage descriptive, especially morphological and chemical data.The SpecialIndexing module records the project specific information on host-pathogen-interactions from literature and specimen data. The identification tool (DiversityIdentify) will usemorphological data as well as geographical and host-parasite-interactions. A web interface will beimplemented during the remaining time of the project to access the identification system from theInternet.

211
Fig. 1: Overview of the modular design of the DiversityWorkbench
Support modules are: DiversityUsers for user administration and documentation, a geographicnames index (DiversityGazetteer, at present including the world-wide Getty TGN and the GermanGN250) used for geo-referencing; DiversityResources, to manage hyperlinks, images, and othermedia used to support the identification of organisms. DiversityResources indexes resources bykeywords and organism names, addresses the problem of changing URLs, and documentsintellectual property rights.All modules and information models of the DiversityWorkbench system are open source andavailable from http://www.DiversityCampus.net.
References:1. Hagedorn, G. 2000. Nachrichtenblatt des Deutschen Pflanzenschutzdienstes, 52, 149.2. Hagedorn, G., Deml, G., Burhenne, M., Guerrero Cartin, O.M., Gräfenhan, T., and Weiss, M. 2000:http://www.DiversityCampus.net/Glopp/Presentations/PflSchutzTg2000/PflSchutzTg2000en.html3. Hagedorn, G. 2001. Bocconea, 13, 261-280.4. Hagedorn, G., 2001: http://www.DiversityCampus.net/Glopp/Presentations/GLOPP2001-M/GregorHagedorn-M2001.html5. Hagedorn, G. 2001: DiversityDescriptions (version 1.8). *6. Hagedorn, G. and Gräfenhan, T. 2001: DiversityTaxonomy (version 0.6). *7. Hagedorn, G. 2001: DiversityReferences (version 0.9). *8. Hagedorn, G. and Weiss, M. 2001: DiversityCollection (beta). *
* References 5-8: All information models, including SQL DDL and xml-schemata documents can be foundunder http://www.DiversityCampus.net/Workbench/download.html

212
Project ID: 01LC9905 (GLOPP, Subproject LIT)1.4.2000 - 31.3.2003
GLOPP-LIT - DIGITIZATION OF IMPORTANT EUROPEAN HOST-PATHOGENINDICES FOR FUNGI
T. Gräfenhan, M. Burhenne , G. Hagedorn, and G. Deml
Institut für Pflanzenvirologie, Mikrobiologie und Biologische Sicherheit, BiologischeBundesanstalt für Land- und Forstwirtschaft, Königin-Luise-Str. 19, 14195 [email protected]; http://www.DiversityCampus.net
Key words: GLOPP, fungi, plant pathogens, identification, host-parasite index, Erysiphales,powdery mildews, Peronosporales, downy mildews, Brandenburger, literature capture,biodiversity informatics
Abstract:The GLOPP-LIT project focuses on capturing published or unpublished scientific information onplant pathogenic fungi. Scientists working in the particular fields of biodiversity will guaranteethe high quality standard of the database. In the framework of the collaborative GLOPP projectthe literature project provides a base layer of information for the identification of plantpathogenic fungi that is supplemented with more detailed information from the projects workingon specific groups. Eventually, scientists will not only be able to search for specific parasite-host-interactions, also a literature reference and geographic relations are given.
Results:The most important publication on plant pathogenic fungi for Europe, Brandenburger [1], hasbeen fully captured in a database of more than 11,200 records. Unfortunately, this publicationdoes not provide any information on literature references or other resources used in itscompilation. It thus has to be supplemented from index cards and manuscripts provided to theproject by Dr. Brandenburger. The original references on index cards have been scanned, partlytranscribed, and indexed in a networked database (7,600 references). The manuscript (comprising12 binders) and personal communications are currently ran through to connect the references withthe host-parasite observations. Approximately half of the work has been finished up to now.In addition two unpublished manuscripts (fig. 1) on the Erysiphales (powdery mildews) and thePeronosporales (downy mildews) are currently edited in cooperation with Dr. Brandenburger andconverted to structured documents. Based on an XML-like mark-up, they will ultimately beconverted to a relational database and integrated into the GLOPP information system.

213
Fig. 1: Plant parasite record from an unpublished manuscript by Dr. Brandenburger.The information is identified using named style tags, which are displayed in
different colours, allowing the correction of mark-up mistakes.
Other publications [2-9] have been prepared for conversion into a database. The remaining time ofthe project will be necessary to finish the conversion and integration of these sources.Subsequently specialists of the fungal groups will improve the quality level of the data.
References:1. Brandenburger, W. 1985: Parasitische Pilze an Gefäßpflanzen in Europa, Stuttgart.2. Amano, K. (Hirata) 1986: Host range and geographical distribution of the powdery mildew fungi.3. Braun, U. 1995: The powdery mildews (Erysiphales) of Europe, Jena.4. Bierlein, J. 1993: Mehltaupilze (Peronosporales, Erysiphales). Regensburger Mykol. Schriften, 2.5. Brandenburger, W. 1994: Die Verbreitung der in den westlichen Ländern der BundesrepublikDeutschland beobachteten Rostpilze (Uredinales). Regensburger Mykol. Schriften, 3.6. Flachs, K. 1931: Krankheiten und Parasiten der Zierpflanzen, Stuttgart.7. Gerlach, W. and Nirenberg, H. 1982: The genus Fusarium – a pictorial atlas. Mitteilungen aus derBiologischen Bundesanstalt für Land- und Forstwirtschaft, 209.8. Gruber, M. 1993: Uredinales. Regensburger Mykol. Schriften, 2.9. Pantidou, M. E. 1973: Fungus-host index for Greece, Athens, Kiphissia.

214
Project ID: 01LC9905 (GLOPP, Subproject Oomycetes)1.4.2000-31.3.2003
GLOPP-OOMYCETES
M. Göker1, A. Riethmüller1, H. Voglmayr2, M. Weiß1, and F. Oberwinkler1
1 Lehrstuhl Spezielle Botanik/Mykologie, Botanisches Institut, Universität Tübingen, Auf der Morgenstelle 1, D-72076 Tübingen2 Institut für Botanik, Universität Wien, Rennweg 14, A-1030 [email protected]
Key words: GLOPP, data base, biodiversity, Oomycetes, Peronosporales, molecular evolution
Abstract:A combination of classical morphology and modern molecular methods allows to achieve highquality of taxonomic data in a biodiversity information system for plant parasitic Oomycetes.
Results:The Oomycetes are a monophyletic group including obligatory parasitic and saprophytic species.Members of the Saprolegniales, Sclerosporales, Pythiales and Peronosporales may causeconsiderable economic damage on crop plants, e.g. Aphanomyces raphani, causing the radishblack dye, Phytophthora infestans, causing the late blight of potato and Plasmopara viticola,causing the downy mildew of grapevine.The GLOPP project does not aim at a simple compilation of data, so critical appraisal of the datafound in taxonomic literature is necessary. As a rule, this entails experimental re-examination ofthe published data. Current systematics of plant parasitic Oomycetes, at least above species level,are based mainly on morphological characters, so extensive light microscopical studies arerequired. As optical microscopy is the customary tool in applied phytopathology, quality dataachieved with this technique are of highest practical value.Unfortunately, the morphological data so far used for Oomycete taxonomy are often ambiguous.This is especially important where only few morphological characters are available, as it is thecase in species delimitation in most parasitic Oomycete genera. Even the total number of speciesin Peronosporales and Sclerosporales stated in the literature can vary very much depending onthe underlying species concept used by different authors [1]. Where species circumscriptions arebased mainly on the taxonomy of the respective hosts [2] and no additional infection experimentshave been carried out, a vicious circle arises with respect to conclusions on cospeciation. Thesame is true when practical questions concerning host specificity are considered.To cope with the mentioned uncertainties, the combination of morphological with modernmolecular methods is required. Application of the latter has been exemplified already in earlierwork from Tübingen [3] with emphasis on the systematics of aquatic Oomycetes. Our recentpublication [4] presenting results from sequence analyses of LSU rDNA focuses on taxonomicproblems in the field of plant parasitic Oomycetes or downy mildews. Below, we will give someexamples from these results and how they influenced our light microscopical analyses and thearrangement of the Oomycete database.Important parts of the current classification of the Peronosporomycetidae [5, 6] proved to beunsatisfactory. With regard to family concepts, we posited that Lagenidium should be placedwithin the Pythiaceae, Phytophthora and Peronophythora within the Peronosporaceae.It could also be shown that the genera Bremiella and Peronophythora should be dismissed andtransferred to Plasmopara and Phytophthora, respectively. Therefore, the character “swollenconidiophore tips” used so far for the delimitation of Bremiella, is probably of minor taxonomic

215
value. Instead, Bremiella baudysii, a species that parasitizes a member of the Apiaceae, seems tobe most closely related to the Plasmopara species infesting this plant family, a result that gives anexample of the meaning of cospeciation in plant parasitic Oomycetes.Following the work of Fraymouth [7], we had already erected a morphological typology of thehaustoria in Peronosporales to be used in the database that was clearly supported by themolecular results. Haustoria seem to be of major importance for the characterization of groups ofgenera in Peronosporales. So the Peronospora species possessing haustoria of our type D aregenetically highly diverging from the ones with type C haustoria. Additionally, the former containall Peronospora spp. infesting Brassicaceae. Therefore, we focussed microscopical research onthis group. Indeed, we could find some other morphological characters that separate it from thePeronospora spp. not specialized on Brassicaceae (unpublished data). Likewise, many of thegroups Gäumann [2] called “Formenkreise” were preliminarily supported. However, oosporecharacters of Peronospora turned out to be of negligible value for infrageneric taxonomy.If data quality of an information system about biodiversity is to be guaranteed, three maininterdependent questions arise: (1) Which taxa represent natural groups? (2) Which characterspossess taxonomic significance? (3) How should the character states be defined to achieve a clear-cut separation of taxa? Obviously, the content of the database will depend to a high degree on howthese questions are answered. In our opinion, a dual strategy employing morphological andmolecular methods not only facilitates finding the correct answer, but is the only possible way toproceed. This justifies the often time-consuming search for fresh material of Oomycete-infestedplants and for molecular methods adjusted to this material and the taxonomic problems that haveto be solved. We hope that the examples from our recent molecular and morphological resultsgiven here may support our view.
References:1. Dick, M. W. 1990: Phylum Oomycota . In: Margulis, L., Corliss, J. O., Melkonian, M., and Chapman, D.J., Handbook of Protoctista, 33, 661-685, Boston.2. Gäumann, E. 1923: Beiträge zu einer Monographie der Gattung Peronospora Corda. Beiträge zurKryptogamenflora der Schweiz, 5(4), 1-260.3. Riethmüller, A., Weiß, M., and Oberwinkler, F. 1999: Phylogenetic studies of Saprolegniomycetidae andrelated groups based on nuclear large subunit ribosomal DNA sequences. Can. J. Bot., 77, 1790-1800.4. Riethmüller, A., Voglmayr, H., Göker, M., Weiß, M., and Oberwinkler, F. 2001: Phylogeneticrelationships of the downy mildews (Peronosporales) and related groups based on nuclear large subunitribosomal DNA sequences. Mycologia (submitted).5. Hawksworth, D. L., Kirk, P. M., Sutton, B. C., and Pegler, D. N. 1995: Ainsworth & Bisby´s Dictionaryof the fungi. 8th ed. Cambridge.6. Dick, M. W. 2001: The Peronosporomycetes. In: McLaughlin, D. J., McLaughlin, E. G. and Lemke, P.A. (eds.), The Mycota, VII A, 39-72. Berlin.7. Fraymouth, J. 1956: Haustoria of the Peronosporales. Transactions of the British Mycological Society,39: 79-107.

216
Project ID: 01LC9905 (GLOPP, Subproject Erysiphales)1.4.2000 – 31.3.2003
GLOPP-ERYSIPHALES - A GLOBAL INFORMATION SYSTEM FOR THEBIODIVERSITY OF POWDERY MILDEWS (ERYSIPHALES, ASCOMYCOTA)
C. Kainz1, M. Weiss1, U. Braun2, and D. Triebel1
1 Botanische Staatssammlung München, Mycology Dept., Menzinger Str. 67, D-80638 München2 Martin-Luther-Universität Halle – Wittenberg, Institut für Geobotanik und Botanischer Garten, Neuwerk 21, D-06108 Halle (Saale)[email protected]; www.botanik.biologie.uni-muenchen.de/botsamml/
Key words: GLOPP, ColdFusion modules, DiversityCollection, Erysiphales, LabelScan, LIAS,molecular phylogeny, biodiversity informatics
Abstract:The labels of 8500 specimens of Erysiphales stored at the state-run herbaria in München andHalle were digitised using LabelScan. Approximately 20–25 % of the label data have alreadybeen transcribed to the database DiversityCollection. Three ColdFusion modules were developedfor making the data available via the Internet. The molecular studies resulted in 40 completesequences of the rDNA ITS region and partial sequences of the rDNA 28S region. An ITSsequence alignment including 50 Genbank sequences was built up for performing cladisticanalyses. Published descriptive data of all taxa of the Erysiphales have been included in the LIASinformation system and are provided as natural language descriptions. In addition, data can bequeried for interactive identification by the DeltaAccess Perl interface. All data will be evaluatedand presented in a global information system.
Results:The project Erysiphales uses a number of joined database modules mainly developed within theGLOPP project (DALI, DiversityCollection, DiversityExsiccatae, DiversityDescriptions). Thebiodiversity data were collected from herbarium labels, from original literature and images and –especially within molecular studies – derived from primary research activities. All types of dataand databases are stored in a modularised database system.Recording and evaluation of herbarium label data based on the Erysiphales collections inHalle (HAL) and München (M). In order to solve the problems of recording large amounts ofherbarium data, a concept for a standardized and user-oriented data recording in collections wasdeveloped. In a first stage the material is labelled with barcodes. These labels are scanned togetherwith the original historical labels showing all information provided by the collector such aslocality data, as well as later revisions. For this purpose the scan-software LabelScan v. 2.0 wasdeveloped to provide an automatic recording of the barcode-labels and a simplified administrationof image files [1]. In another stage the digital label images are imported into DiversityCollectionin order to establish a comfortable and structured way of transcribing the label data into this MSAccess database. The herbarium material is only needed for the scanning of the labels, allsucceeding work can be done within the database, either locally or over the Internet. Due to thegreat amount of material (8500 specimens in M and HAL) the transcription process had to bedivided into a number of stages, some of which could be carried out even by untrained persons.Actually a selective entry of data is performed in three parallel and independent procedures,namely the entry of collection data (2500 data records already completed), of host data (1200 datarecords already completed), and of location data via Getty TGM (600 data records alreadycompleted). The data input into DiversityExsiccatae, a database module with bibliographicspecifications and abbreviations of 220 exsiccata series, is now complete. A pull-up list,

217
containing the basic information of DiversityExsiccatae, was integrated into DiversityCollectionin order to facilitate data entry of the exsiccata series.Online access to collection data, images and literature of Erysiphales. Several ColdFusionmodules have been developed to give public access to the data via the Internet: “SearchExsiccatae” is a tool for searching and remote editing the entries in DiversityExsiccatae [2],“Search Collections” allows a search for host-parasite relationships and collectors inDiversityCollection [3] and the module “Search Drawings Braun 1987” [4] gives online access tomore than 300 plates with ink drawings originally published by Braun [5]. The literature databaseDALI (A Database of Ascomycete Literature) has been extended by the category Erysiphales andcontains 400 pertinent data sets [6].Data recording for molecular phylogenies (18S, ITS, 28S). The molecular studies on thephylogeny of Erysiphales resulted in 40 complete sequences of the internal spacer region (ITS) ofthe rDNA and partial sequences of the 18S and 28S rDNA. The sequences were aligned with 50sequences received from GenBank. Alignments of all sequences and several subgroups have beenmade. On the basis of these alignments cladistic calculations were performed to get new insightsinto the relationships of European taxa of powdery mildews. The analyses of the Uncinula aduncagroup are especially promising. Currently the molecular studies are extended to non-ribosomalgenes such as the beta-tubuline coding gene. After successful primer selection the first beta-tubuline sequences were obtained. The relevancy of this gene for phylogenetic studies on varioussystematic levels has still to be tested.Build-up of a LIAS associated online-system for interactive identification and characteranalysis. The descriptive data of Braun [5] were partly transferred to LIAS (A Global InformationSystem for Lichenized and Non-Lichenized Ascomycetes [7]. For this purpose the LIAS characterlist had to be extended to Erysiphales-specific characters, and a list with all species names andtheir basionyms including author abbreviations checked for correctness had to be compiled. Thedata are now accessible as natural language descriptions [8]. In order to complete the descriptivedata sets with digital illustrations of the organisms (see “Search Drawings Braun 1987”) aninterface for cross linking is under construction. The interface DeltaAccess Perl [9] is used forestablishing an online identification tool for Erysiphales within LIAS [10].
References:1. Hagedorn, G., Triebel, D., and Weiss, M. 2000: LabelScan: a tool to optimise the establishment of bigcollection information systems using selective label data transcription. Presentation and computerdemonstration. In: Anonymous (ed.), TDWG 2000: Digitising Biological collections. Taxonomic DatabaseWorking Group, 16th annual meeting, Abstracts, Mycol. Res., 10, Frankfurt.2. http://www.mycology.net/exsiccata/ExsiccataFind.cfm3. http://www.mycology.net/collections/bsm/specimens/CollectionFind.cfm4. http://www.mycology.net/lias/taxa/drawings/ braun1987/Braun87Find.cfm5. Braun, U. 1987: A monograph of the Erysiphales (powdery mildews). Bh. Nova Hedwigia, 89, 1-700.6. http://141.84.65.132/BSM-Mycology/Dali/search.html7. http://www.mycology.net/lias/index.html8. http://www.mycology.net/lias/descr/erys33s24/9. http://www.herbaria.harvard.edu/computerlab/ web_keys/delta_access_perl.html10. http://www.mycology.net/lias/online/dap.html

218
Project ID: 01LC9905 (GLOPP, Subproject Smut Fungi)1.4.2000-31.3 2003
GLOPP – SMUT FUNGI
M. Piepenbring2, C.-L. Hou1, G. Hagedorn3, G. Deml3, and F. Oberwinkler1
1 Lehrstuhl Spezielle Botanik/Mykologie, Botanisches Institut, Universität Tübingen, Auf der Morgenstelle 1, D-72076 Tübingen2 Botanisches Institut, Universität Frankfurt, Senckenberganlage 31-33, D-60054 Frankfurt/M3 Institut für Pflanzenvirologie, Mikrobiologie und Biologische Sicherheit, Biologische Bundesanstalt für Land- und Forstwirtschaft, Königin-Luise-Str. 19, D-14195 [email protected]
Key words: GLOPP, smut fungi, Tilletiales, Ustilaginales, China, DiversityDescriptions,biodiversity informatics
Abstract:Data on the morphology of smut fungi are integrated into the GLOPP information system usingthe module DiversityDescriptions. Up to now, 460 different species of smut fungi have beendocumented. Field research conducted in the Neotropics and in Yunnan, China, providedcollections of fresh material of smut fungi, which was used to study the morphology and alpha-taxonomy. These primary data significantly improved the quality of species information in thedatabase.
Results:The software DiversityDescriptions (see GLOPP-IT) presently offers 169 characters withnumerous character states for the description of species of smut fungi. These characters andcharacter states (descriptors) are based on those observed in detail by M. Piepenbring onneotropical smut fungi for which a monograph for Flora Neotropica is prepared. The monographincludes 227 species in 34 genera, or ca. 17.5 % of the about 1,300 species of smut fungi knownworld-wide. With more species of smut fungi added to the database, further characters and/orcharacter states have to be added. The database allows this at any time, without affecting existingdata sets.As many as possible species will be studied by us personally before including them into thedatabase. Further species are included based on descriptions in literature we consider reliable.European smut fungi, for example, are relatively well known by a monograph written by Vánky[1], descriptions of North American smut fungi can be found in the book of Fischer [2]. Thecharacter states of species descriptions in literature are transferred into the characters andcharacter states available in GLOPP-DiversityDescriptions. In some cases like size and colourcharacters, this is a relatively straightforward task, but e.g. for soral morphology and for sporeornamentation, different authors use different terminology. Illustrations like detailed drawings andSEM photographs of teliospores can help, but a broad knowledge of the diversity ofmorphological features of smut fungi is indispensable to avoid transcription errors.By now 460 more or less complete and revised species descriptor sets are present in theDiversityDescriptions database. We intend to include descriptions of most of the known speciesof smut fungi until the end of the project, however, present knowledge on many species of smutfungi is limited in several ways:• Details of soral morphology are only evident in good material including young states ofdevelopment.

219
• Some species are known by only one or few poor collections which do not always allow adetailed study of morphology nor the identification of the host plant.
• The identification of host plants documented in literature is not always reliable.• For many species we do not know the morphology of the basidia, which can only be observed
with recently collected teliospores placed on water agar.• There are many taxonomic problems including unresolved synonymy, problematic genericplacements, and unclear delimitation of species.In addition to these problems, we are far from knowing the really existing diversity of smut fungi,especially in large countries where few mycologists looked for this group.In order to improve the quality of data for poorly known species, field work is a constant part ofour investigations. In Bolivia, with only five weeks of field work and study of herbariumspecimens, four new species were found [3].For China the diversity of Ustilaginales is documented by Guo Lin [4]. She described 140 speciesof this order for China, which has regions with very high diversity of higher plants. This can onlybe a small proportion of the existing diversity of smut fungi, because for Europe, with lessdiversity of higher plants, we know ca. 400 species of Ustilaginales and Tilletiales, with theUstilaginales being the larger group [1]. Fieldwork and study of herbarium specimens in Yunnanduring the summer of 2001 led to 50 collections representing 25 species. These can now beexamined in detail and the respective descriptor sets can be completed in the database, includinge.g. the morphology of basidia. Among the species are new ones, new records for the region, andsome of them occur on species hitherto unknown as host. More field work of this kind, however,is necessary to come to a more realistic assessment of the diversity present in China.Recently collected specimens of smut fungi can be used for molecular analyses. Phylogenetichypotheses based on LSU rDNA provide important arguments for systematic and taxonomicconclusions. They directly improve the quality of the data in the database by indicating genericplacements of the species. The delimitation of characters and character states is also affectedbecause molecular phylogenies allow to distinguish similar character states based on homologiesfrom those based on analogies. This has recently been demonstrated for species of Cintractia byPiepenbring et al. [5].While field research on smut fungi is going on, we complete the database hoping to encouragemore mycologists and taxonomists of higher plants to look for this group of plant parasitic fungi,which is not only of economic importance but also a fascinating example on how parasitic fungicoexist with their host plants in a natural balance.
References:1. Vánky, K. 1994: European smut fungi. Stuttgart, New York.2. Fischer, G. W. 1953: Manual of the North American fungi. New York.3. Piepenbring, M. 2001: New species from the neotropics. Mycol. Res., in press.4. Lin, G. 2000: Ustilaginaceae. Flora Fungorum Sinicorum, 12. Beijing.5. Piepenbring, M., Begerow, D., and Oberwinkler, F. 1999: Molecular sequence data assess the value ofmorphological characteristics for a phylogenetic classification of the species of Cintractia. Mycologia, 91,485-498.

220
Project ID: 01LC9905 (GLOPP, Subproject Uredinales)1.4.2000-31.3.2003
GLOPP-UREDINALES - REGISTRATION OF TROPICAL AND SUBTROPICALRUST FUNGI
Reinhard Berndt, Anja Ritschel, and Franz Oberwinkler
Lehrstuhl Spezielle Botanik/Mykologie, Botanisches Institut, Universität Tübingen, Auf derMorgenstelle 1, D-72076 Tü[email protected]
Key words: biodiversity-database, Costa Rica, Prospodium, Uredinales
Abstract:Rust fungi are economically important but still insufficiently studied in tropical regions. In ourapproach we ascertain data on taxonomy, distribution and ecology of tropical rust fungi andcomplement them by basic systematic and field research. This combination yields high qualitydata that are essential for the applied research and utilization of taxonomic knowledge e.g. inagriculture as well as to solve elementary questions of rust fungal biology.
Introduction and research plan:With about 7.000 species, rust fungi are the biggest and economically most important group ofobligate plant parasites. Beside their economic importance, rust fungi have a uniquelycomplicated life cycle and offer excellent opportunities for the study of plant-parasite co-evolution. The knowledge about biodiversity, distribution and host relationship of rust fungi fromthe tropics and subtropics is still restricted and difficult to access as summarising floras andmonographs are widely lacking.Our work aims to contribute to the investigation of tropical and subtropical rust mycota(1) by providing data on the taxonomy, distribution and host selection of the parasites(2) and by ascertaining own field and systematic data.Only the permanent control of literature data by accompanying critical elementary studies canguarantee a reliable data quality.In our approach we concentrate on three subjects:• a study of the rust fungus mycota of Costa Rica• a monographic study of the neotropical rust genus Prospodium• the building of a database/checklist of tropical and subtropical rust fungi containing data
on nomenclature, distribution and host spectra.
Results:
Rust fungus mycota of Costa Rica. During our exploratory trips to Costa Rica we securedapprox. 300 rust specimens, of which 250 are determined and registered in the database. Amongthe collected specimens we found seven species that were new for science. Several speciesrepresented new reports for Costa Rica or were found on hosts that had not been recognisedbefore. In the next step the important Döbbeler-collection of Costa Rican rusts will be evaluated.So far, 220 species of rust fungi have been ascertained for Costa Rica from literature.

221
Prospodium, a neotropical genus of rust fungi. We selected the genus Prospodium for a casestudy for several reasons: with approx. 70 species the genus is exclusively neotropical and showsa strict host restriction on Bignoniaceae and Verbenaceae. Its tropical distribution, species numberand morphological diversity makes Prospodium an ideal example to combine a monographicstudy with processing the data in a character and biodiversity database. Up to now, a species- andsynonym list have been built up. Two new species, Prospodium manabii and P. kisimovae, werecollected in Ecuador and Costa Rica.
Database/checklist of tropical and subtropical rust fungi. We are working on a database thatcontains nomenclatural information as well as data on distribution and host spectra of rusts. At thepresent stage, mainly literature data on tropical and subtropical species are included and is beingcritically evaluated. The database presently contains information on approx. 13.000 parasite-hostcombinations that are represented by ca 21.000 geographical reports.

222
Project ID: 01LC9906 (INFOBOT)1.4.2000 – 31.3.2003
INFOBOT - DEVELOPMENT AND TESTING OF INFORMATION SYSTEMSCOVERING DIFFERENT ASPECTS OF BOTANICAL COLLECTIONS
W. G. Berendsohn (project coordinator) and A. Vasiliou
Department of Biodiversity Informatics and Laboratories, Botanic Garden and Botanical MuseumBerlin-Dahlem, Freie Universität Berlin, Königin-Luise-Str. 6-8, D-14191 [email protected]; http://www.bgbm.org/BioDivInf/
Keywords: information systems, botany, vascular plants, Compositae, living collection, naturalhistory collection, botanical garden, natural substances, biodiversity informatics
Abstract:INFOBOT demonstrates the application of information technology on different aspects ofbotanical collections: the management of living collections, the linking of factual information(chemical compounds) important in applied research, and access to type specimen information asone of the keys to basic taxonomic research. The resulting information systems are to bepublished on the WWW and will be integrated into information networks.
Introduction. INFOBOT is a collaborative project creating information systems about differentaspects of plant collections. Two of the three partial projects focus on the world’s largest vascularplant family, the Compositae. Although they lack the urgently needed central taxonomiccomponent (a global species database of the family), the two projects demonstrate differentapproaches towards the mobilization of taxon information in electronic databases. The thirdproject incorporates the aspect of living collections. The potential of botanic gardens as a sourceof information and materials for education and research will not be fully realized until theirinformation is available on-line. At the same time, information technology greatly assists in theinternal management and quality control of the information held and produced by suchcollections.INFOCOMP/TYPES uses digital photography (see fig. 1 and following article) in combinationwith textual input to provide access to nomenclatural type specimens (vouchers on which thespecies’ description was based and which fix the application of the scientific name). The types ofthe Compositae present in the most important German herbaria are being recorded and will bemade accessible to scientists on the WWW.INFOCOMP/CHEM , the “Bohlmann Files”, is a database of all natural substances in theCompositae. A former card index which was transferred to an ISIS/PC database is now convertedto a relational database format and will be accessible on the WWW. In this database chemical andbotanical expertise is united, providing combined information which is of great potential for awide range of users, e.g. in pharmaceutic industry and biotechnology. The project pinpoints thediversity of natural substances and will contribute to the taxonomic research attempting toelucidate the relationships and evolution within this important group of plants.INFOBOT-BoGART, the Garden Accessions Database of the Botanic Garden Berlin-Dahlem, isa model project for a complete accession and administration of the living collection of a botanicgarden. This system provides effective access to the actual living stock of vascular plants of oneof the largest living collections world-wide. It is a program for the data management of a botanicgarden and acts as a basis for further scientific research. With the linking of the database to theWWW, the information will be accessible to a wide range of users and can also be connected toother databases and information systems dealing with biodiversity information.

223
Fig. 1: A botanical type specimen (INFOCOMP/TYPES). The rules of botanical nomenclatureprescribe that any taxonomic group named like this specimen (or carrying a name based on thisname) will have to include this specimen. Type specimens thus form the essential base of theprocess of naming taxonomic concepts.

224
Project ID: 01LC9906 (INFOBOT, Subproject INFOCOMP/TYPES)1.4.2000 – 31.3.2003
COMPOSITAE TYPES IN GERMAN HERBARIA
P. Bodensteiner, A. M. R. Davies, A. Pillukat, B. Grau, F. Schuhwerk, and J. Grau
Botanische Staatssammlung München, Menzinger Straße 67, D-80838 MünchenContact: [email protected]
Key words: types, Compositae, specimen images, German herbaria, biodiversity informatics
Abstract. Of the estimated 2,700 Compositae types held in Munich, 1,533 have beenphotographed, resulting in approximately 7,600 high-resolution images. All the bibliographicinformation has been checked and linked to the images. The completed data is gradually beingmade available on the Internet, using the software "Filemaker Pro".Introduction. The Compositae are the largest family of seed plants with an estimated 25,000species [1]. They are found in all places colonised by higher plants on all continents and oftenform significant parts of the most diverse ecosystems [2]. They are not only scientificallyinteresting but also have economically important representatives. Nomenclatural types are thereference sheets of pressed plants laid down in herbaria. These are directly and permanentlyconnected to their plant names and allow species to be unambiguously identified. They thusunderpin even the most modern research techniques (see [3]). The more traditional and larger aherbarium, the richer it will be in types. For historical reasons, many types originate fromdeveloping countries and have attained value through European botanical research. With thisproject it is now possible to contribute to data repatriation by making this information globallyavailable to scientists.The project was established primarily as a pilot scheme for the digital recording of essentialcomponents of German herbaria. The project is supported by three graduate scientists and, fromtime to time, a Database/Network administrator and the current excellent co-ordination of the staffhas greatly increased the speed of progress in recent months. With the experience gained, digitalimaging in targeted spheres can now be rapidly and successfully completed.
Fig. 7: Original label Fig. 8: Capitulum
Results. Initial teething problems have now been resolved and image format and qualitystandardised. On average, five images per type are taken: the original label(s) (fig. 1) [necessarywhen determining provenance and authorship of older sheets, as well as for subsequent datacapture from the images], the entire sheet (see previous page), relevant habit detail, and macroimages of taxonomically important features - usually the capitulum(a) (fig. 2, 3). Special emphasisis placed upon the correct and accurate citation of the protologue.

225
Figure 9: Detail of capitulum necessary for species identification
Types from all seventeen of the Compositae tribes have been recorded (fig. 4). In total 350 generaand 1,533 types have been photographed, forming a database of over 7,600 images. Awaiting afully functional interface for image storage and retrieval in the database system SYSTAX, thesoftware "iView MediaPro" has been used to organize image and text data, retaining flexibility forother software options. The database software used to capture data, "Filemaker Pro", provides anintegrated web interface allowing direct access to the database via the World Wide Web. It isanticipated that the work in Munich, including the preparation of the information for Internetaccess, will be completed during the next year.
Fig. 4: Number of Compositae Types in Munich (M) photographed(as of Sept. 2001) arranged by Tribe (after [4])
References:1. Heywood, V. H. 1993: Flowering Plants of the World. London.2. Heywood, V. H., Harborne, J. B., and Turner, B. L. 1977: An overture to the Compositae. In:Heywood, V. H., Harborne, J. B., and Turner, B. L. (eds), The Biology and Chemistry of theCompositae, 1, 1–20, London.3. Hind, D. N. J., Jeffrey, C., and Pope, G. V. (eds) 1995: Advances in Compositae Systematics. Kew.4. Bremer, K. 1994: Asteraceae, Cladistics and Classification. Portland, Oregon.
Senecioneae (166)
Heliantheae (194)
Astereae (153)
Mutisieae (73)
Anthemideae (93)
Lactuceae (94)
Cardueae (141)
Plucheeae (19)Inuleae (50)Eupatorieae (63)
Calenduleae (5)Barnadesieae (6)
Liabeae (8)
Arctoteae (14)
Helenieae (46)
Gnaphalieae (196) Vernonieae (212)
Remaining Taxa (c. 1100)

226
Project ID: 01LC9906 (INFOBOT, Subproject INFOCOMP/BOHLMANN)1.8.2000 – 31.05.2002
THE BOHLMANN-FILES – A DATABASE OF NATURAL SUBSTANCES IN THECOMPOSITAE
A. Hahn, J. Jakupovic, B. Smalla, F. Tsichrintzis, C. Zdero, and W. G. Berendsohn
Department of Biodiversity Informatics and Laboratories, Botanic Garden and Botanical MuseumBerlin-Dahlem, Freie Universität Berlin, Königin-Luise-Str. 6-8, D-14191 [email protected]; http://www.bgbm.fu-berlin.de/BioDivInf/projects/bohlmannfiles/
Key words: Compositae, Asteraceae, natural substances, chemotaxonomy, chemical database,taxonomic database, secondary metabolites, biodiversity informatics
Abstract:The Compositae (Asteraceae) contain numerous chemical compounds, many of them with a highpotential in, e.g., pharmaceutical utilisation. A relational database was designed holding almost20,000 chemical structures from about 5000 taxa. The revision of plant names and chemical datais almost concluded, reviewing of synonymy is under way, and publication on the World WideWeb will follow in the near future.
Introduction. Members of the Compositae are renown for their contents of a variety ofphysiologically active compounds [1], for example, they are used as remedies such as chamomile,insecticides such as pyrethrinoids, or pharmaceuticals such as the antimalarial agent Artemisininfrom Artemisia annua. The late Ferdinand Bohlmann, professor at the Technical University ofBerlin, dedicated much of his life to the investigation of these substances [2]. Together with histechnical assistant C. Zdero he started in the beginning of the 1960ies to compile a card index onnatural substances and taxa of the Compositae, encompassing both data from original researchand from literature. About 95% of the data are published. In 1994, Zdero and one of Bohlmann’spupils, J. Jakupovic, started to transfer the data to an ISISBase chemical database, constantlyadding data from more recent publications.The project. At the outset of the present project, the database held data on about 5000 taxa, with atotal of 19,351 chemical structures. In addition, (trivial) name of the compound, molecular weightand formula, literature references, and various revision notes were on record. The ISISBaseprogram allows searching on structures and partial structures as well as on the contents of the textfields. However, ISISBase is not designed for the publication of small datasets on the World WideWeb, and it cannot easily be linked to other data sources. The Bohlmann Files project was thusproposed to meet three main objectives: (i) Convert the database into a relational system andtransfer all data, (ii) revise the chemical and nomenclatural data and fill in remaining gaps fromliterature, and (iii) make the database accessible via the WWW, including the search features of achemical database (see fig. 1).Results. The data is considered reasonably complete for secondary metabolites found up to nowin the Compositae. Providing free access to this wealth of information will not only serveapplied sciences (e.g. to identify potential sources for substances), but also enhance basicresearch, e.g. by providing additional clues to the taxonomy of groups considered to beparaphyletic (e.g. the tribe Lactuceae) or not yet assigned to a tribe (e.g. the genera Corymbiumand Centipeda ) and to elucidate end-products of gene expression and biosynthetic pathways.The project database has been set up under MS Access 2000, adapting the taxonomic module ofthe BoGART-database system (see next project) for the treatment of identification andnomenclature. Data from the flat-file format ISIS database were parsed to separate and atomiseplant names, synonyms, and literature data. Chemical structures are stored using the add-in

227
“Accord” software. Its structural search capabilities allow searches via the WWW and thus formthe core of the web publishing component which will be released before long.
Query: A (sub-)structure can bedrawn or input in textual form in theWWW query form can be made
Artemisia douglasiana BesserDendranthema maximowiczii (Komar.) TzvelevAthanasia dregeana Harv.
Result: The result consists ofsecondary metabolites containing thestructure and (among other data) thetaxa they were isolated from
Dendranthema maximowiczii (Komar.) Tzvelev
Fig. 1: Searching for a substructure in the database
Plant names have undergone a revision process including spell checking, standardisation(including author citations), and a check against major web-based databases on plant names [3, 4].They now reflect the recent classification of the Compositae [5]. The data conform to the TDWGstandard on names [6] complete with source citations for taxonomic data. Literature citations arekept in a separate reference list. Further taxonomic and bibliographic information and data on thebioactivity or use of compounds or the plant itself can be linked at a later date. Taxa from about755 genera represent all tribes within the 3 subfamilies.
References:1. Heywood, V. H., Harborne, J. B., and Turner, B. L. (eds) 1977: The Biology and Chemistry of theCompositae. 1, 1–20, London.2. Winterfeldt, E. 1994: Ferdinand Bohlmann und sein wissenschaftliches Werk. Liebigs Ann. Chem.,1994, 1-34.3. Plant Names Project 1999: International Plant Names Index. Published on the Internet;http://www.ipni.org4. Missouri Botanical Garden. VAST (VAScular Tropicos) Nomenclatural Database and AssociatedAuthority Files, Published on the Internet as W3TROPICOS release 1.5;http://mobot.mobot.org/W3T/Search/vast.html5. Bremer, K. 1994: Astereaceae – Cladistics and Classification. Portland.6. Bisby, F. 1994: Plant Names in Botanical Databases, TDWG Standard No.3, Version 1.00. 1994;http://www.tdwg.org/plants.html

228
Project ID: 01LC9906 (INFOBOT, Subproject BoGART)1.4.2000 – 30.06.2002
BOGART - THE INFORMATION SYSTEM OF THE BERLIN BOTANIC GARDEN
A. Hahn, A. Kirchhoff, U. Matthäs, A. Surma, and W. G. Berendsohn
Department of Biodiversity Informatics and Laboratories, Botanic Garden and Botanical MuseumBerlin-Dahlem, Freie Universität Berlin, Königin-Luise-Str. 6-8, D-14191 [email protected]; http://www.bgbm.fu-berlin.de/BioDivInf/Projects/Bogart-d.htm
Key words: biological collection, botanical garden, living collection, botany, taxonomy,information system, biodiversity informatics, accessions
Abstract: The “BoGART” database system of the BGBM demonstrates system development in alarge living collection. The original system has been converted to a client server environment.Existing data (electronic or in writing) were transferred to the new system and the user interfaceadapted to specific tasks in the management of the garden’s collection. A prototype XML wrapperprovides access for networks and local or remote query interfaces.
Introduction. With more than 20,000 taxa and 35,000 accessions on 43 ha, the Botanic GardenBerlin-Dahlem is one of the world’s largest gardens. It is a scientific research collection, aneducational exposition, and at the same time a recreational site for the public. Collections of thissize not only demand constant physical care but also require an elaborate system to administer theinformation flow for scientific as well as practical work (see fig. 1). Detailed tracking ofinformation on provenance and handling of accessions is an obligation with regard to the CBD’s(Convention on Biological Diversity) demand for benefit sharing with countries of origin.Adequate documentation is also a pre-requisite for the botanical garden’s role in speciesconservation [1].INFOBOT-BoGART will complete the last phase in the process of transferring this system fromthe labour intensive index card system of the past to a networked electronic information system. Itupgrades the internal prototype system to a scalable and publicly available multi-user application,based on widely used technology (SQL-Server, MS Access, WWW). It builds on several priorprojects, starting in the 1980ies, which provided a sound foundation of evaluation, modelling, andimplementation (see [2] for further historical details).Results regarding data import, input, control and standardization. The import of staticelectronic data files from existing isolated applications within the garden has been finished. Datainput of taxonomic information was completed and the data content has been checked against thecard files. The system now contains information on more than 38,000 taxon names. Likewise,almost 37,000 accession datasets from imports have been checked, supplemented and partiallyreworked, achieving a high level of detail e.g. on the original provenance of the plants. Datastandardization was a major focal point, concentrating on the areas of literature data ( ca. 1450citations), geography, and names and authors of plant taxa.Technical results. The complex data model, as well as the increased data content had severelylimited the performance of the system. True multi-user functionality was achieved with theconversion of the Access97 database into an Access2000 project. The backend resides in an MSSQL-Server2000 client-server database, and the Access-frontend’s client connection has beenconverted from ODBC to OLE DB. The new system provides efficient parallel access fromworkstations distributed throughout the Botanic Garden area. Work in progress includes furtherperformance optimisation, transferring functionality from Visual Basic code in the front end toserver-sided stored procedures and triggers.

229
Networking. An XML-DTD was prepared which covers all data items provided by theBoGART system as well as concatenated or converted items demanded by exchange standardssuch as the ITF format [3]. Based on the DTD, a prototype XML wrapper has beenprogrammed, which will be used to connect the database to national and international networks.Integration in the work process. The ongoing final period of the project is focussed on theimplementation of specific functionalities, supporting e.g. inventory processes, the scientificscrutiny of taxon names and linked data (synonyms, common names, species distribution,poisonous, medicinal, and protected plants, literature), the identification process, the collectionand documentation of herbarium specimens, etc. The functions are developed in close cooperationwith the respective user groups.
Science
BoGART
IdentificationIncorporation of taxonomic research resultsSpecies conservationResearch collections
Science
Index Seminum (international seed exchange)Supply of plant material for science and educationVisitor informationLocal network access On-line access through BGBM website and Botanical Garden Information System (VBG/Systax) and other networks
Services
Cultivation
provenance informationrequired by the CBD2000 new accessions and ~900 scientific identifications p.a.inventory data from garden departmentsvoucher collections for garden herbarium
DocumentationProcessing
Monitoringendangered plants and research collections
Fig. 1: Information domain of the BoGART program
References:1. Maunder, M., Higgens, S., and Culham, A. 2001: The effectiveness of botanic garden collections insupporting plant conservation. Biodiversity & Conservation, 10, 383-401.2. Elankovan, S. and Meyer, H. 1996: Analyse, Modellierung und Design einer taxonomischen undsammlungsbezogenen relationalen Datenbasis zur Verwaltung von Akzessionsdaten im Bereich derBotanik. Diplomarbeit TU Berlin, FB Wissensbasierte Systeme. (http://www.bgbm.org/projects/examensarbeiten/elankovanMeyer/)3. Jackson, D. (compiler) 1997: International Transfer Format for Botanic Garden Plant Records (version2.00 draft 3.2.). Richmond. [Proposed TDWG standard] [Electronic version:http://www.rbgkew.org.uk/BGCI/news.htm ]

230
Project ID: 01LC0026 (AlgaTerra)1.10.2001-30.09.2004
ALGATERRA – AN INFORMATION SYSTEM FOR TERRESTRIAL ALGALBIODIVERSITY: A SYNTHESIS OF TAXONOMIC, MOLECULAR ANDECOLOGICAL INFORMATION
R. Jahn (project coordinator)
Department of Biodiversity Informatics and Laboratories, Botanic Garden and Botanical MuseumBerlin-Dahlem, Freie Universität Berlin, Königin-Luise-Str. 6-8, D-14191 [email protected]; http://www.algaterra.net/
Key words: information system, Internet accessibility, biodiversity, algae, diatoms, green algae,cyanobacteria, types, taxonomy, nomenclature, Ehrenberg, Hustedt, collection data, literaturedata, identification, morphology, culture strains, rDNA, sequence analyses, phylogeny, ecologicaldata, biodiversity informatics
Abstract:Aim of this recently started project is to build a comprehensive information system on terrestrialmicro-algae, integrating taxon, type, name and collection data as well as ecological andmolecular information. The main target groups will be scientists pursuing basic research, such astaxonomy and biodiversity studies, as well as applied researchers, such as ecologists, limnologists[1], and public agencies, engaged in biomonitoring for public health and environmentalconservation. The system will be made available on the Internet (World Wide Web).
Background:Microscopic algae have been recognized as useful tools in global change research, e.g. formonitoring water quality indicating trophic status, acidification, and salinization, forreconstruction of paleoclimates through sediments, and for the assessment of biodiversity ofdiverse ecosystems. The taxonomy and in particular the nomenclature used to record theseobservations is often ambiguous, leading to incoherent data sets published by different researchersthat need to be synthesized for global studies. The available ecological information for these taxais thus doubtful and modern molecular information cannot be assessed correctly. The problemsare frequently due to poorly documented nomenclatural types as well as to divergent secondaryinterpretations of the taxon concept behind the name.
The network:The BGBM Berlin-Dahlem (see AlgaTerra – AT1) will be responsible for the development of thecomprehensive information system, and for data evaluation and typification of selected taxahoused at the Ehrenberg Collection. The other participants of the AlgaTerra network are theMuseum für Naturkunde Berlin (Ehrenberg Collection, see AT2), the Alfred Wegener Institute,Foundation for Polar and Marine Research (type data of the Hustedt Collection, molecular,morphological and ecological data of diatoms, see AT3), the Culture Collection of Algae at theUniversity of Göttingen (morphological and molecular data of green and bluegreen algae, seeAT4), and the University of Leipzig (ecological and applied data of aero-terrestrial algae, seeAT5). This cooperation (see fig. 1) will synthesize taxonomic, molecular and ecologicalinformation from all subprojects for worldwide accessibility.
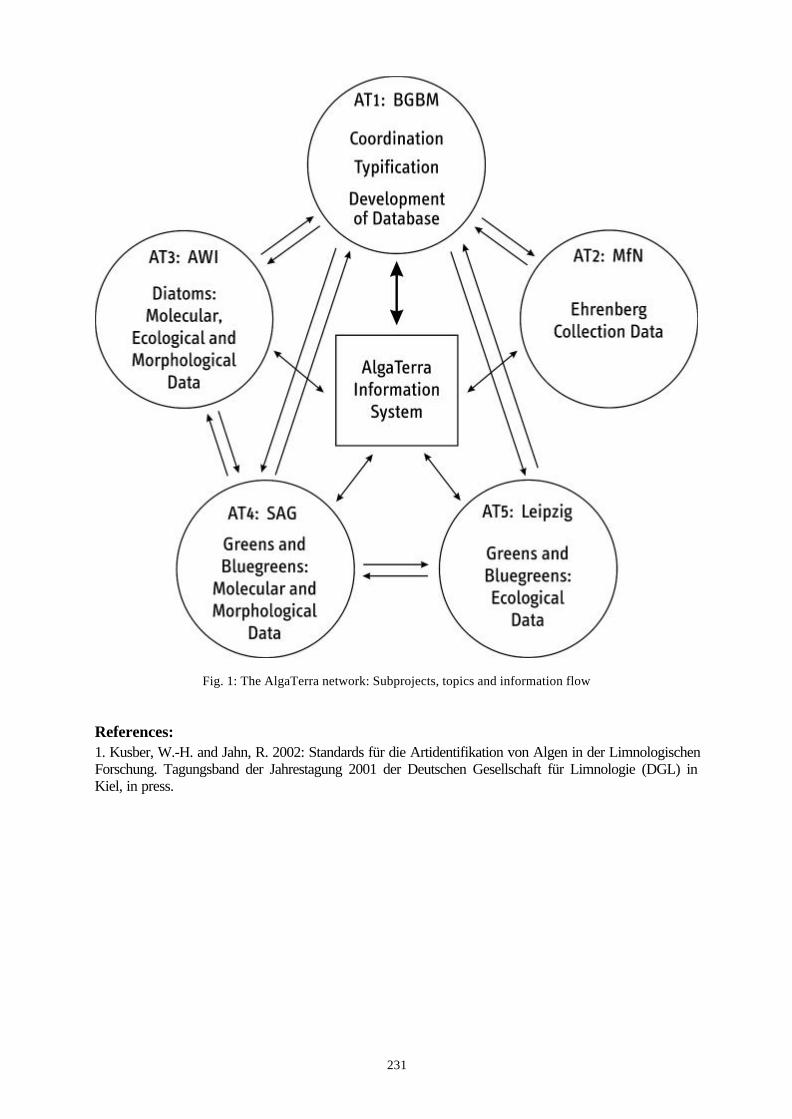
231
Fig. 1: The AlgaTerra network: Subprojects, topics and information flow
References:1. Kusber, W.-H. and Jahn, R. 2002: Standards für die Artidentifikation von Algen in der LimnologischenForschung. Tagungsband der Jahrestagung 2001 der Deutschen Gesellschaft für Limnologie (DGL) inKiel, in press.

232
Project ID: 01LC0026 (AlgaTerra Subproject 1)1.10.2001-30.09.2004
THE ALGATERRA INFORMATION SYSTEM INCLUDING THE IDEAL-MODULE
W.-H. Kusber, K. Teske , and R. Jahn
Department of Biodiversity Informatics and Laboratories, Botanic Garden and Botanical MuseumBerlin-Dahlem, Freie Universität Berlin, Königin-Luise-Str. 6-8, D-14191 [email protected], [email protected], [email protected]; http://www.bgbm.org/jahn/default.htm
Key words: types, taxonomy, nomenclature, identification, morphology, algae, diatoms,biodiversity, Ehrenberg, database, internet accessibility, biodiversity informatics
Abstract:A comprehensive Internet Information System for terrestrial algal biodiversity as a synthesis oftaxonomic, molecular and ecological information (AlgaTerra) will be developed andimplemented. The IDEAL-Module (Internet Database of Ehrenberg’s Algae Lectotypes) willsupply original data on algal types as a calibration tool for the identification of micro-algae. TheInformation System will be managed and supported over the long-term by the Botanic Gardenand Botanical Museum Berlin-Dahlem as part of its scientific information services.
Results and research plan:The BGBM Berlin-Dahlem will be responsible for the information system and for dataevaluation.The IDEAL-Module will give access to original data of selected taxa described by C.G.Ehrenberg (1795-1876). The relational database will include the validated name of the type, theoriginal diagnosis, type locality information, a digital photograph of the selected type, and, wherepossible, a SEM-analysis of the original material (for Ehrenberg’s scientific life and taxonomicconcept, see [1, 2]). The database will also supply possible synonyms, notes of experts plus linksto updated nomenclature, modern taxonomic interpretations as well as biogeographic, ecologicand molecular data.Typification of Ehrenberg-taxa, housed in the Ehrenberg-Collection ([3], see AT2) wasestablished and published for a small number of genera [4, 5] and species [6, 7, 8], but no data isaccessible via Internet.The main steps of data preparation for the IDEAL-Module are:
• digitalization of the algal name and description• translation of the original description• validity check according to the rules of nomenclature• digitalization of the published picture• digital photograph of the chosen (lecto)-typeQuality evaluation of data will include:• taxon concept of the data source (fig. 1)• type data versus interpreted data (observation data, applied data)The development of the information system [9] will include:• elaboration of an appropriate data model and database design• programming and implementation of the database• development of a user-oriented web-interface

233
• establishment of the connection between database and Internet
Fig. 1: Example of an unpublished drawing by C.G. Ehrenberg. This drawing sheet of 1834 shows his taxonomicconcept, representing at least five taxa in our current understanding. It includes manuscript names, data oncollecting dates as well as size measurements which are to be compared to his published data (fig. provided byAT2).
References:1. Jahn, R. 1998: C. G. Ehrenberg: The man and his botanical science. Special Publication of the LinneanSociety, 1, 15-29.2. Jahn, R. 1995: C. G. Ehrenberg’s concept of the diatoms. Arch. Protistenk., 146, 109-116.3. Lazarus, D. B. and Jahn, R. 1998: Using the Ehrenberg Collection. Diatom Res., 13, 273-291.4. Lange-Bertalot, H. and Jahn, R. 2001: On the identity of Navicula (Frustulia) rhomboides and Frustuliasaxonica (Bacillariophyceae). Syst. Geogr. Pl. “2000”, 70, 255-261.5. Crawford, R. M., Likhoshway, Y. V., and Jahn, R.: The morphology and identity of Aulacoseira italicaand the type of the name of the genus Aulacoseira. Diatom Res., submitted.6. Jahn, R. 1995: Ehrenberg’s marked species on five mica. Iconogr. Diatomol., 1, 101-107.7. McLachlan, J. L., Boalch, G. T., and Jahn, R. 1997: Reinstatement of the genus Exuviaella(Dinophyceae, Prorocentrophycidae) and an assessment of Prorocentrum lima. Phycologia, 36, 38-46.8. Crawford, R. M. and Likhoshway, Y. V. 1999: The frustule structure of original material of Aulacoseiradistans (Ehrenberg) Simonsen. Diatom Res., 14, 239-250.9. Jahn, R., Kusber, W.-H., and Teske, K. 2001: The project AlgaTerra: Outline of an information systemfor terrestrial algal biodiversity. Poster and Abstract TDWG, Sydney.

234
Project ID: 01LC0026 (AlgaTerra Subproject 2)1.10.2001-30.09.2004
ACCESSIBILITY OF THE EHRENBERG COLLECTION
D. Lazarus and A. Jobst
Instititut für Paläontologie, Museum für Naturkunde, Humboldt-Universität zu Berlin,Invalidenstr. 43, D-10115 [email protected]; http://www.museum.hu-berlin.de/pal/microp/david.lazarus.htm
Key words: Ehrenberg, algae, diatoms, collection data, types, original materials, internetaccessibility, biodiversity informatics
Abstract:The Ehrenberg Collection holds the scientific legacy of Christian Gottfried Ehrenberg (1795-1876), generally considered to be one of the founders of both microbiology and micro-paleontology. Ehrenberg was one of the most taxonomically prolific of all scientists, and the typespecimens for several thousand algal species and several hundred algal genera are present in thecollection. These include many of the most common species and genera names now in use, andthe collection includes the types and original material for many of the species and genera(diatoms and also other algae) that will be examined in the AlgaTerra project.
Introduction:The collection was deposited with the Museum für Naturkunde in Berlin upon Ehrenberg’s death,and has been preserved through the years in good physical condition. Experience in working withthe collection has shown that the majority of all type material is indeed present in the collection(fig. 1). However, locating these type materials in the Ehrenberg Collection is currently verydifficult, as the collection has not been adequately curated since it was deposited with theMuseum more than 100 years ago, and the original collection structure is not well suited tomodern research needs [1, 2, 3].
Fig. 1: Number of types for genera in Ehrenberg Collection
0 20 40 60 80 100 120
diatoms
green algae
phytoflagellates
radiolarians
ciliates
other protists

235
The project. The goal of this subproject is to carry out the minimum amount of basic inventory,labelling, archiving and database work (fig. 2) needed to allow other subprojects to use theEhrenberg Collection efficiently in their own work. The work will result in a database of theEhrenberg Collection; properly labelled, sorted collection objects; working image copies of fragileoriginal documents (see AT1, fig. 1); and, for those taxa selected for study by the project as awhole, data extracts from the Ehrenberg Collection database that will be imported into the mainproject database (see AlgaTerra – AT1). The Ehrenberg Collection database will be maintained atthe Museum für Naturkunde and will be accessible online via standard Internet protocols.
Fig. 2: Mica strip holding five mica preparations with original material and information; to be catalogued in thissubproject
References:1. Lazarus, D. B. 1998: The Ehrenberg Collection and its curation. In: Williams, D.M. and Huxley, R.(eds), Christian Gottfried Ehrenburg [sic] (1795-1876): The man and his legacy. The Linnean Society,Special Publication, 1, 31-48.2. Lazarus, D. B. and Jahn, R. 1998: Using the Ehrenberg Collection. Diatom Res., 13, 273-2913. Lazarus, D. B. 2000: On the value of paleontological collections. In: Berendsohn, W. G. (ed.), Resourceidentification for a biological collection information service in Europe (BioCISE), 33-36.

236
Project ID: 01LC0026 (AlgaTerra Subproject 3)1.10.2001-30.09.2004
MOLECULAR AND ECOLOGICAL DATA OF TERRESTRIAL DIATOMS
L. Medlin, R. M. Crawford, B. Berszteri, K. Hamann, F. Hinz, and S. Strieben
Alfred Wegener Institute, Foundation for Polar and Marine Research, Am Handelshafen 12, D-27570 [email protected]; http://www.awi-bremerhaven.de/Biomeer/molekulargenetische-e.html
Key words: diatoms, Hustedt, rRNA, phylogeny, ecological data, culture strains, Nitzschia,biodiversity informatics
Abstract:Type localities of the species described by F. Hustedt from terrestrial habitats in Germany andNorthern Europe will be established and re-visited to generate living cultures for deposition asepitypes in culture collections. Phylogenetic analysis of these epitype cultures will be performedusing molecular techniques. In collaboration with AlgaTerra AT1 a database will be createdincluding image-based keys to facilitate the identification of terrestrial diatoms.
Introduction. The diatoms (Bacillariophyta) are one of the most successful and ecologicallyimportant groups of microalgae, occurring in both aquatic and terrestrial habitats with over 10,000described species and potentially many more cryptic species. They are distinctly well known fortheir architecturally complex siliceous cell walls (valves and girdle bands) and their unusualpattern of reduction in cell size of one of the daughter cells following mitosis, among otherfeatures. The intricate designs of their cell wall have been the basis for their classification systemfor more than two centuries. In the last fifty years, the diatoms have been routinely used toprovide estimates of ecological perturbation, specifically eutrophication and acid rain. Currentlythey are being exploited as proxies for climate change in paleo-environments by looking at salttolerances and the changes in the diatom communities in saline lakes and some terrestrial habitatsinfluenced by salinity.Germany has had at least three renowned diatomists from the last two centuries – Kützing,Ehrenberg and Hustedt – who described many hundred new species. Kützing’s material wasdistributed to several contemporary scientists and has found its way into museums worldwide.The Ehrenberg and Hustedt collections remain more or less intact, the former at the Museum fürNaturkunde, Humboldt Universität zu Berlin (see AT2), and the latter at the Friedrich HustedtArbeitsplatz at the Alfred-Wegener-Institute in Bremerhaven, Germany. The Hustedt Collectionwill be the basis of this subproject concerning names, types, ecological and molecular data.Research plan. The Hustedt collection is fully curated [1, 2]. It is currently being catalogued intoan Internet database [2] and will be the focus of our project. The research objectives of subprojectAT3 are:• To establish the type localities of the type species described by F. Hustedt from terrestrial
habitats from Germany and Northern Europe.• To return to these localities at the same time of the year to re-establish living cultures of these
earlier described species for deposition in modern culture collections, such as the SAG(AT4). These new cultures will be designated as epitypes and epitype permanent preparationswill be prepared and deposited in the Hustedt Herbarium.
• To perform phylogenetic analysis of these epitype cultures using an analysis of their rRNAgenes or ITS regions if they are varieties of the same species.

237
• To create a database, from which an image-based key will be made to facilitate theidentification of these diatoms from both living and fixed, cleaned material using the epitypeculture and the original material in cooperation with AT1. These will be supplemented withdatabase entries for other important terrestrial diatoms in these collections that were notdescribed by Kützing, Ehrenberg and Hustedt so that a meaningful assemblage of terrestrialdiatoms will be compiled for identification purposes using our database.
Accordingly, the project is divided into 4 workpackages:• Databasing of names, pictures and type localities of terrestrial diatoms described by F.
Hustedt.• The establishment of epitype cultures based on the types and localities from work package 1
and the documentation of dominant feature of each species using both fresh and cleanedmaterial.
• The reconstruction of the phylogeny (see [3]) of dominant species of terrestrial diatoms fromGermany and Northern Europe based on 18S rRNA sequences derived from the epitypecultures established in work package 2 and from other cultures of representative diatomsdominant in our collection from type localities re-sampled throughout Germany and NorthernEurope.
• Construction of an identification key using our database of the photographs of terrestrialdiatoms.
Results. B. Beszteri came to the AWI in July 2001 on a DAAD scholarship. He beganimmediately to collect diatoms from one site on the estuary of the Geeste river near the AWIwhere Hustedt had made many collections and had described 4 species. B. Beszteri made about19 collections (16 benthic from different substrates and 3 planktonic) from this site from which heestablished 40 cultures. Among the species recovered at the time of the collection he searched forNitzschia hoehnkii, N. aestuarii, N. tryblionella f. subconstricta and Diploneis dimorpha whichwere described by Hustedt from this site. We are currently awaiting the grow up of existingcultures to see if we have obtained either of these species in our rough cultures. Preliminaryindications are that we likely have Nitzschia hoehnkii and N. aestuarii in culture collected exactlyat the same location from where they were originally described. We are growing the algae both inliquid and solid culture to facilitate their manipulation.
References:1. Simonsen, R. 1987: Atlas and Catalogue of the Diatom Types of Friedrich Hustedt, 1-3. Berlin.2. Crawford, R. and Hinz, F. 2001: Information on the Collection [http://www.awi-bremerhaven.de/Research/hustedt3.html]; List of types [http://www.awi-bremerhaven.de/Research/hustedt1.html].3. Medlin, L. K, Kooistra, W. C. H. F., and Schmid, A.-M. M. 2000: A review of the evolution of thediatoms – a total approach using molecules, morphology and geology. In: Witkowski, A. and Sieminska, J.(eds), The origin and early evolution of the diatoms, Kracow.

238
Project ID: 01LC0026 (AlgaTerra Subproject 4)1.10.2001-30.09.2004
DOCUMENTATION OF CHARACTERS OF TERRESTRIAL ALGAE INCULTURE: R-DNA SEQUENCE ANALYSES, MORPHOLOGY AND CRYO-PRESERVATION OF REFERENCE STRAINS
T. Friedl1, D. Hepperle1, and Y. van de Peer2
1 Experimentelle Phykologie und Sammlung von Algenkulturen, Albrecht-von-Haller-Institut für Pflanzenwissenschaften, Universität Göttingen, Untere Karspüle 2, D-37073 Göttingen2 Evolution Biology, Department of Biology, University of Konstanz, P.O. Box 5560 M618, D-78457 [email protected]; http://www.gwdg.de/~epsag/phykologia/sag/sag.htm
Key words: algae, cyanobacteria, rDNA, sequence analyses, phylogeny, morphology,cryopreservation, culture strains
Abstract:The project will primarily analyse molecular characters of terrestrial microalgae from cultures,and will also gather morphological data for those strains that have been sequenced. The resultswill be integrated into the comprehensive information system for the unambiguous identificationof these algae and they will be used to explore phylogenetic relationships. In addition, all strainsused in this project will be preserved by cryotechniques and thus made available for furthercomparisons and investigations. The project is closely connected with one of the largest culturecollections of algae (SAG, Sammlung von Algenkulturen Göttingen).
Research plan:Phylogeny research in terrestrial algae, in particular green algae and cyanobacteria from aero-terrestrial habitats:Molecular characters of terrestrial microalgae (excl. diatoms) will be examined by means of DNAsequence analyses of ribosomal genes (rDNA) using reference strains. This will help to clarify thephylogenetic relationships of these algae [1, 2, 3, 4]. A regularly updated gene tree will guide theuser to the sequence data as well as to all other relevant information. The DNA sequence data willbe provided as secondary structure-model based alignments ([5], e.g., in ARB format). Thesequence database will help in identifying algae even from uncultured material (e.g., soil crusts)or environmental DNA extractions, a feature which is essential for applied research.
Authentic culture and other reference strains form the basis for the documentation ofmolecular and morphological characters.This brings a large culture collection into the focus of modern research. The main source forculture strains is the “Sammlung von Algenkulturen Göttingen (SAG)” which is one of theworld’s largest living collection of algae [6]. Of the ca. 2100 strains currently in the SAG, 419 areauthentic strains (i.e. represent subcultures of the culture material used for the original taxonomicdescription of algal species) of which 369 were isolated from terrestrial habitats. These strains willbe supplemented by additional authentic culture strains, particularly from the culture collection atInnsbruck, Austria (ASIB, Dr. G. Gärtner), which mainly holds stocks of soil and airborne algae[7, 8]. Other reference strains used by the project will be carefully selected in collaboration withthe other subprojects of AlgaTerra i.e. AT1 (taxonomic identity, Ehrenberg type) and AT5(ecological importance as currently known), as well as from other culture collections needed torepresent the diversity of genera of microalgae from aero-terrestrial habitats as best as possible.

239
Further strains may be provided by the projects AlgaTerra AT5, and the BIOTA projects TP05-1and S05-1.
Preservation of reference strains by cryotechniquesAll the information as gathered by the project will be referenced to genetically stable cryo-preserved culture material which will be available to the public for further comparisons andinvestigations. Although cryopreservation is routinely used in other culture collections, newprotocols for the cryopreservation of microalgae will be developed. The establishment ofcryocultures will ensure the prolonged survival of important reference strains.
Documentation of morphological characters and culture dataAll relevant information that is available for the culture strains used in this project (e.g., origin,optimal culture conditions) will be collected and integrated into the information system.Morphological features of the strains will be documented, and a collection of image data (withattached comments that explain morphological characters) will be provided with the informationsystem.
References:1. Ben Ali, A., De Baere, R., Van der Auwera, G., De Wachter, R., and Van de Peer, Y. 2001. Int. J. Syst.Evol. Microbiol., in press.2. Fewer, D., Friedl, T., and Büdel, B. 2002. Mol. Phylogen. Evol., in press.3. Friedl, T., Besendahl, A., Pfeiffer, P., and Bhattacharya, D. 2000. Mol. Phylogenet. Evol., 14, 342-352.4. Friedl, T. in Bhattacharya D. (ed.) 1997: The Origin of Algae and Plastids. Pl. Syst. Evol., [Suppl] 11,87-101.5. Van de Peer, Y., De Rijk, P., Wuyts, J., Winkelmans, T., and De Wachter, R. 2000. Nucleic Acid Res.,28, 175-176.6. SAG: Sammlung von Algenkulturen an der Universität Göttingen [http://www.gwdg.de/~epsag/phykologia/epsag.html].7. Ettl, H. and Gärtner, G. 1995: Syllabus der Boden-, Luft- und Flechtenalgen. Stuttgart.8. Gärtner, G. 1996. Ber. Nat.-Med. Verein Innsbruck, 83, 45-69.

240
Project ID: 01LC0026 (AlgaTerra Subproject 5)1.10.2001-30.09.2004
CHARACTERISTIC DATA OF GREEN AND BLUEGREEN AERO-TERRESTRIALALGAE (CHLOROPHYTINA AND CYANOBACTERIA)
W. Reisser and M. Salisch
Institut für Botanik der Universität Leipzig, Johannisallee 21-23, D-04103 [email protected]; http://www.uni-leipzig.de/~allbot/
Key words: cyanobacteria, green algae, literature data, culture strains
Abstract:Whereas information on marine and freshwater algae is one of the standard topics ofphycological textbooks, comparably few data are available for aero-terrestrial taxa, i. e. thosealgae which live on and in soil [1, 2], cover surfaces such as rocks and tree barks or which arepart of the aero-plankton. The reasons for this information gap are manifold, but one clearobstacle for a better understanding of these algae is the difficult access to published information.Important data are scattered among journals of low citation impact or grey literature and is oftennot available in English. This subproject will contribute to overcome this impediment byassembling available information and making it accessible worldwide.
Research plan:In order to create the basis for an intensified research on aero-terrestrial algae this project willfocus on:• the search for, assembly, and processing of data on aero-terrestrial algae (blue-green and
eukaryotic algae excluding diatoms), including translation into English• axenic cultures of aero-terrestrial algae including their description and taxonomic
identification by classical standard methods and• the development of an image-based web-interface to chlorophycean aero-terrestrial algae
Progress made:Currently we are engaged in data mining (morphology, taxonomy, ecology, original descriptions)of predominantly Russian and Ukrainian sources, translation and processing of data into acompatible structure. Parallel to this, we are completing data sheets of axenic isolates of aero-terrestrial algae of our algal collection (literature data such as original descriptions,ecophysiological data, morphological documentation by drawings and photographs).
References:1. Ettl, H. and Gärtner, G. 1995: Syllabus der Boden-, Luft- und Flechtenalgen. Stuttgart.2. Wöhler, I., Mensching, A.-K., and Reißer, W. 1999: Algae in Forest soils: Distribution and taxonomicaffiliation. In: Rastin, N., Bauhus, J. (eds), Going underground. Ecological studies in forest soils. ResearchSign Post Trivandran, India, 187-200.

241

242
Appendix
Index of Names
A
Aas, G. .................................................................... 44, 46Adamczyk, J.................................................................80Ade, M. .......................................................................112Adeba, P. J. .................................................................142Agerer, R.....................................................................122Akhtar-Schuster, M...................................................110Akoegninou, A. .........................................................146Althof, A.....................................................................158Amann, R......................................................................84Andres, W. .................................................................146Anhuf, D .....................................................................130Arbuckle, T.................................................................196Audorff, V. ...................................................................38Austermühle, R..........................................................110Awiti, A.......................................................................152
B
Balkenhol, B.................................................................66Ballot, A......................................................................184Barthlott, W................................................................132Beck, E..........................................................................10Beck, L........................................................................208Becker, T.....................................................................110Behrens, T.....................................................................68Beierkuhnlein, C................................................... 44, 46Berendsohn, W. G............................ 16, 224, 228, 230Bergsdorf, Th.............................................................168Berndt, R............................................................104, 222Berszteri, B.................................................................238Bertke, E. ......................................................................76Bierschenk, T. ............................................................146Bleeker, W....................................................................30Bleher, B. ....................................................................170Blüthgen, N. ...............................................................208Bodensteiner, P..........................................................226Böhning-Gaese, K.....................................................170Bonkowski, M..............................................................90Boutros, M..................................................................140Brake, I........................................................................192Braker, G.......................................................................82Braun, G.............................................................100, 128Braun, U......................................................................218Braun, W.....................................................................192Braune, E. ...................................................................114Brechtel, F. .................................................................208Bresinsky, A.................................................................26Broll, A..........................................................................56Buchmann, N................................................................36Büdel, B. ............................................................106, 108Burhenne, M...............................................................214Burkhardt, U...............................................................200Buss, H.-J....................................................................120Bussmann, R. W........................................................172
C
Clausnitzer, V.............................................................162Conrad, R......................................................................82Crawford, R. M..........................................................238Cremers, A. B.............................................................196Cudjoe, A. A. .............................................................142
D
Dalitz, H......................................................................155Dauber, J. ......................................................................74Davies, A. M. R.........................................................226Deckert, J. ...................................................................112Deke, O. ................................................................. 92, 94Deml, G.....................................................210, 214, 220Dericks, G.....................................................................20Diedrich, K. ..................................................................76Dietrich, C..................................................................202Dojani, S. ....................................................................108Döring, R......................................................................54Dormann, W.................................................................56Dressler, S...................................................................146Dunfield, P....................................................................82Düring, C......................................................................26Durka, W.......................................................................64
E
Eberl, L..........................................................................88Ebigbo, N. M..............................................................144Ehlebe, M......................................................................28Eiseb, S........................................................................112Engels, C................................................................ 44, 48Ernst, R........................................................................142
F
Fahr, J..........................................................................144Falk, T. ........................................................................120Feige, G.-B. ................................................................178Feuerer, T....................................................................108Filser, J........................................................................200Fischer, A......................................................................76Fischer, E. ...................................................................158Fjellberg, A.................................................................200Franck-Oberaspach, S. L..................................... 92, 94Franke, T.....................................................................122Friedl, T.......................................................................240Frohschammer, S.......................................................142
G
Gebhardt, S...................................................................60Gerowitt, B. ..................................................................76Gich, F...........................................................................86Giere, P........................................................................112Gliech, M ....................................................................212Goetze, D. ...................................................................134Göker, M.....................................................................216
243
Görke, C......................................................................104Görlich, T......................................................................56Gräfenhan, T......................................................212, 214Grau, B........................................................................226Grau, J. ........................................................................226Gröngröft, A...............................................................102Groß, I. ........................................................................192Grunert, J. ...................................................................134Güth, M.........................................................................64Guttwein, D. ...............................................................192
H
Hachfeld, B.................................................................110Hadrys, H....................................................................114Hagedorn, G. ...................................210, 212, 214, 220Hahn, A..............................................................228, 230Hahn-Hadjali, K.........................................................146Hamann, K..................................................................238Hampicke, U.................................................................54Hanagarth, W.............................................................208Harbig, C.....................................................................142Haszprunar, G. ...........................................................198Häuser, C. L.......................................................194, 204Häuser, Ch. L. ............................................................160Hausmann, A..............................................................206Heimpel, B..................................................................120Hein, P.........................................................................176Held, E...........................................................................52Hellwig, F. ....................................................................42Hennenberg, K...........................................................134Hepperle, D. ...............................................................240Herrmann, B.................................................................22Hespelt, S. K.................................................................76Heyde, K. ......................................................................64Hille, A........................................................................198Hinz, F.........................................................................238Hochkirch, A................................................................56Höfer, H. .....................................................................208Hoffelner, H..................................................................86Hoffmann, A........................................................28, 112Hoffmann, I. ...............................................................120Hoffmann, S. ................................................................28Holstein, J. ................................................160, 194, 204Hoppe, J. R. ................................................................188Hörsch, B. ..........................................................100, 128Horz, H. P. ....................................................................82Hou, C.-L....................................................................220Hubaishan, M. A. .............................................176, 182Hurek, T. .......................................................................78Hurka, H........................................................................30Hüttl, R. F. ....................................................................60
I
Ikels, L...........................................................................58Ingrisch, S...................................................................202Isaacs, G. .....................................................................120Isselstein, J....................................................................76
J
Jahn, R................................................................232, 234Jakupi, A.......................................................................22Jakupovic, J. ...............................................................228Jeltsch, F. ....................................................................116
Jetschke, G....................................................................42Jobst, A. ......................................................................236Jürgens, N...................................................................110
K
Kahlheber, A..............................................................140Kahmen, A....................................................................36Kainz, C. .....................................................................218Kalko, E. K. V............................................................144Kamer, N.......................................................................56Kersting, P. .................................................................134Kiefer, S. .....................................................................172Kier, G.........................................................................132Kilian, N......................................................................176Killmann, D................................................................158Kirchhoff, A...............................................................230Kirk, M........................................................................120Klepper, G. ............................................................ 92, 94Klingenstein, F...........................................................192Klütsch, C. ..................................................................182Knight, R.....................................................................120Koch, F........................................................................112Koehler, H. ...................................................................24Köhler, J......................................................................150Köhler, W.....................................................................68König, K......................................................................146Kossmann, G. ........................................................ 44, 50Kost, C.........................................................................136Kosuch, J. ..........................................................142, 164Kotut, K.......................................................................184Kouamé, G. N. ...........................................................142Koulibaly, A...............................................................134Kowatsch, A.................................................................54Kraemer, M.................................................................168Krell, F.-T...................................................................166Krell-Westerwalbesloh, S. .......................................166Kreuzer, K. ...................................................................90Krieglsteiner, L............................................................26Krienitz, L...................................................................184Krug, M.......................................................................120Kühne, L. ....................................................................160Küper, W.....................................................................132Küppers, M.................................................................108Kürschner, H. .............................................................176Kusber, W.-H.............................................................234
L
Lampe, K.-H......................................................192, 202Langer, E.....................................................................104Lazarus, D...................................................................236Lehner, A......................................................................80Leistner, G. .................................................................142Linsenmair, K. E......................................136, 140, 142Loris, K.......................................................................108Lösch, R........................................................................20Lötters, S.....................................................................164Loy, A............................................................................80Lücking, R. .................................................................108Ludwig, T. ..................................................................190Ludwig, Th.................................................................188Ludwig, W..................................................................190
244
M
Mahiva, V. ..................................................................166Marais, E. ....................................................................112Marbach, B. ................................................................106Marggraf, R. .................................................................76Martens, A..................................................................114Mathenge, S................................................................172Mathiak, G....................................................................56Matthäs, U. .................................................................230Matzner, E. ............................................................ 44, 50Medlin, L. ...................................................................238Meier, H. .....................................................................190Meister, J.....................................................................180Mennicken, M............................................................104Mey, W. ......................................................................112Meyer, F......................................................................208Miehlich, G.................................................................102Mies, B........................................................................178Miller, M. A. ..............................................................198Mißkampf, R. ...............................................................56Mody, K......................................................................136Morzfeld, J....................................................................72Mossakowski, D. .........................................................56Motzer, T. ...................................................................130Müller, J........................................................................24Müller-Motzfeld, G.....................................................56Münzenberger, B.........................................................60Musila, W...................................................................155Mutiso .........................................................................172Mutke, J.......................................................................132
N
Nashr, A. K.................................................................182Naumann, C. M.................................................150, 182Neßhöver, C........................................................... 44, 46Neumann, K................................................................146Newman, D.................................................................166Njeri, R........................................................................172Njoroge, G. .................................................................172Nuppenau, E.-A.........................................................120
O
Oberprieler, Ch..........................................................180Oberwinkler, F.......................104, 210, 216, 220, 222Odat, N..........................................................................42Onyango, J..................................................................172Ott, S............................................................................108Overmann, J..................................................................86
P
Perner, J.........................................................................40Pernthaler, A.................................................................84Pernthaler, J..................................................................84Petersen, A. ................................................................102Piepenbring, M..................................................210, 220Pillukat, A...................................................................226Pivarci, R....................................................................134Plötner, J. ....................................................................112Porembski, S...............................................................134Purtauf, T. .....................................................................74
R
Rambold, G. ......................................................106, 108Reimer, L. .....................................................................72Reineking, B...............................................................118Reinhold-Hurek, B............................................... 12, 78Reisser, W.........................................................108, 242Reuter, A................................................................ 44, 48Richter, L. ...................................................................190Richter, O....................................................................114Richter, S. .....................................................................28Riede, K......................................................................202Riethmüller, A............................................................216Ritschel, A..................................................................222Rödel, M.-O................................................................142Roscher, S...................................................................192Rotich, D.....................................................................164Rudolf, V. ...................................................................142
S
Salisch, M..........................................................108, 242Schaab, G. ...................................................................152Schareika, N. ..............................................................146Scheu, S.........................................................................90Schickhoff, U. ..............................................................52Schmid, J.......................................................................70Schmidt, M.................................................................146Schmiedel, U..............................................................110Schmitz, U. ...................................................................20Schneiderat, U............................................................120Schorr, G.....................................................................142Schröder, S. ................................................................196Schuhwerk, F..............................................................226Schultz, C...........................................................100, 128Schwager, M...............................................................116Seiberling, S. ................................................................52Siber, R........................................................................106Sigl, K............................................................................88Sinsin, B......................................................................146Smalla, B.....................................................................228Solga, A.......................................................................158Söndgerath, D. ...........................................................114Spelda, J......................................................................208Steidle, A......................................................................88Steinbrecher, R...........................................................152Steiner, A...........................................................194, 204Steinhage, V. ..............................................................196Steinsiek, P.-M.............................................................22Stoll, P. T. .....................................................................92Stoll, P.T. ......................................................................94Straßl, L.......................................................................155Strieben, S...................................................................238Strohbach, B...............................................................110Strohbach, M..............................................................110Strunz, G. ....................................................................152Suhling, F....................................................................114Surma, A.....................................................................230Szarzynski, J...............................................................130
T
Teske, K. .....................................................................234Tews, J.........................................................................116Thies, C.........................................................................70
245
Thiombiano, A...........................................................146Thünen, T. ....................................................................78Traoré, D.....................................................................134Triebel, D...........................................................210, 218Trusch, R.....................................................................206Tscharntke, T. ..............................................................70Tsichrintzis, F.............................................................228Tute, C...........................................................................76
U
Uhlig, M......................................................................112Uhlmann, E.................................................................104Uster, D.......................................................................155
V
van de Peer, Y............................................................240Vasiliou, A..................................................................224Veith, M.............................................................142, 164Verhaagh, M...............................................................208Vidal, S..........................................................................72Vierk, R.........................................................................56Visser, U.......................................................................68Vogel, M............................................................100, 128Voglmayr, H...............................................................216Vohland, K. ................................................................112Voigtländer, K..............................................................66
W
Wagner, M....................................................................80Walsh, M. ...................................................................152Warnecke, F..................................................................84Weber, B. ....................................................................106Wegmann, M..............................................................142
Weimann, J. ..................................................................28Weiß, M......................................................................216Weiss, M............................................................212, 218Weisser, W. W...................................................... 34, 40Wendt, H.....................................................................112Werner, D. ..................................................................120Wessolek, G..................................................................58Westram, R.................................................................190Wiegand, K...................................................................68Wirth, V. .....................................................................108Wissel, C.....................................................................118Wittig, R......................................................................146Wittmann, D...............................................................196Woas, S. ......................................................................208Wöllecke, J. ..................................................................60Wolters, V.............................................................. 12, 74Wurst, C......................................................................208Wytrykush, C. ..............................................................40
X
Xylander, W.................................................................66
Z
Zdero, C. .....................................................................228Zedda, L. .....................................................................108Zeller, U. .....................................................................112Zhang, L. .......................................................................78Ziebarth, A....................................................................56Zimmermann, M..........................................................56Zimmermann, R...........................................................52Zizka, G.......................................................................146

246
BIOLOG-Coordinators
BIOLOG-Europe
Prof. Dr. Volkmar WoltersDepartment of Animal EcologyJustus-Liebig-University GiessenHeinrich-Buff-Ring 26-32 (IFZ)D-35392 GiessenPhone: ++49-(0)[email protected]
assisted byChristine [email protected]
Prof. Dr. Barbara Reinhold-HurekLaboratorium für Allgemeine MikrobiologieUniversität Bremen, FB 2Leobener Strasse NW2D-28359 BremenPhone: ++49-(0)[email protected]
BIOLOG-Africa
West Africa:Prof. Dr. Eduard LinsenmairLehrstuhl für Tierökologie und TropenbiologieUniversität Würzburg, BiozentrumAm HublandD-97074 WürzburgPhone: ++49-(0)[email protected]
assisted byDr. Minnattallah [email protected]
East Africa:Prof. Dr. Clas NaumannZoologisches Forschungsinstitut und Museum Alexander Koenig,Adenauerallee 160D-53113 BonnPhone: ++49-(0)[email protected]
assisted byDr. Jörn Kö[email protected]

247
Southern Africa:Prof. Dr. Norbert JürgensBotanical InstituteUniversity of HamburgOhnhorststr. 18D-22609 HamburgPhone: ++49-(0)[email protected]
assisted byDr. Ingo [email protected]
Biodiversity Informatics
Prof. Dr. Walter BerendsohnDepartment of Biodiversity Informatics and LaboratoriesBotanic Garden and Botanical Museum Berlin-DahlemFreie Universität BerlinKönigin-Luise-Str. 6-8D-14191 BerlinPhone: ++49-(0)[email protected]
assisted byAlexandra [email protected]
Contact BIOLOG Programme
DLR e.V.Project Management Organizationon behalf of the Federal Ministry ofEducation and ResearchEnvironmental Research and TechnologyGodesberger Allee 119D-53175 Bonn
Dr. Lothar QuinternPhone/Fax: ++49-(0)228-81996-20/[email protected]
Dr. Arndt WüstemeyerPhone/Fax: ++49-(0)228-81996-34/[email protected]





























