Embed Size (px)
Citation preview

Experimental Parasitology 123 (2009) 212–217
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr
Giardia canis: Ultrastructural analysis of G. canis trophozoites transfectedwith full length G. canis virus cDNA transcripts
Lili Cao a,c, Pengtao Gong a, Jianhua Li a,*, Xichen Zhang a,*, Xiaohuan Zou b, Wenbin Tuo c,Quan Liu b, Qiuyue Wang a, Guocai Zhang a, Lifeng Chen d, Lingdan Li a, Libo Su a
a College of Animal Science and Veterinary Medicine, Jilin University, 5333 Xi’an Road, Changchun 130062, People’s Republic of Chinab Institute of Veterinary, Academy of Military Medical Sciences, 1068 QingLong Road, Changchun 130062, People’s Republic of Chinac USDA-ARS, Animal Parasitic Diseases Laboratory, ANRI, Beltsville, MD 20705, USAd HeBei Normal University of Science and Technology, Qinhuangdao 066000, China
a r t i c l e i n f o
Article history:Received 11 August 2008Received in revised form 25 June 2009Accepted 14 July 2009Available online 18 July 2009
Keywords:Giardia canisTransfectionVirus-freeRNA transcriptUltrastructure
0014-4894/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.exppara.2009.07.001
* Corresponding authors. Fax: +86 431 87981351.E-mail addresses: [email protected] (J.
(X. Zhang).
a b s t r a c t
Giardia canis virus (GCV) is a double-stranded RNA (dsRNA) virus of the family Totiviridae. In this study, thefull length cDNA of the G. canis virus was constructed in pPoly2/sfinot vector and RNA was transcribedin vitro. Virus-free G. canis trophozoites were transfected with in vitro transcribed GCV RNA by electropor-ation. Transfected trophozoites were cultured for 12, 24, 36, 48, 60, or 72 h post transfection for analysis. Theultrastructures of the transfected trophozoites were determined by transmission electron microscopy. Theviral particles were detectable sporadically in the cytoplasm as early as 24 h post transfection, but becameevident and wide-spread 36 h post transfection. The number of viral particles increased dramatically from48 to 60 h. Viral particles were released into the culture medium starting at about 60 h and detectable innuclei 72 h post transfection. Severe vacuolization was seen in transfected G. canis trophozoites as earlyas 36 h post transfection and persisted throughout the course of this study. The results of the present studyindicate that in vitro transcribed GCV transcripts were capable of infecting Giardia trophozoites, apparentlyreplicated and packaged into mature infectious viral particles which were released from the host.
� 2009 Elsevier Inc. All rights reserved.
1 Abbreviations used: G. canis, Giardia canis; GCV, Giardia canis virus; G. lamblia,
1. Introduction
Giardia spp. is one of the flagellated, eukaryotic, and parasitic pro-tozoa. It infects a number of hosts and causes asymptomatic infec-tions and chronic disorders including persistent diarrhea. There is avery high prevalence of Giardia infection in developing countries(Adam, 2001). Giardia has a simple asexual life cycle whose develop-ing stages include motile, flagellated trophozoites parasitizing theupper intestine and the thick-walled cysts dwelling in the lower intes-tine, the latter are shed to the environment through the feces (Upcroftand Upcroft, 1998). The unique features of Giardia include apredominantly anaerobic metabolism, complete dependence on sal-vage of exogenous nucleotides, limited ability to synthesize and de-grade carbohydrates and lipids. Morphologically, it has two equalnuclei and Golgi apparatus only present in the encysting trophozoites(Adam, 1991). One unique characteristic of Giardia is that it is one ofthe few protozoan parasites that harbor an infectious virus. The Giar-dia virus is present only in some isolates of Giardia. It was reportedthat only 36% of the 76 isolates of G. lamblia from human samples werepositive for virus RNA (Wang et al., 1988). In a separate study, it was
ll rights reserved.
Li), [email protected]
confirmed that approximately 47% was positive for the virus in 38 iso-lates tested (Johan et al., 1987). Furthermore, this study also showedthat 70% of the G. lamblia virus positive isolates had virus receptors onthe parasite cell surface, providing biochemical evidence for suscepti-bility (Sepp et al., 1994). The Giardia virus is non-enveloped and icosa-hedral with a diameter of 36 nm, belonging to the family Totiviridae.The virus has a non-segmented double-stranded RNA genome ofapproximately 6.2 kb (Wang and Wang, 1986).
Since the successful cloning of the full length cDNA from dsRNAtemplate of giardiavirus (Wang et al., 1993), the modified giardia-virus RNA has been used a genetic tool for the study of possibleintervention and pathogenesis (Liu et al., 2005). Similarly, G. canisvirus has been used to construct a stable transfection system whichwas used to stably transfect the trophozoites of G. canis (Yu et al.,2001; Chen et al., 2007; Liu et al., 2008). Numerous studies showedthat transcripts of cDNA clones of viruses with a RNA genome wereused to transfect host cells for the understanding of mechanismsof infection and pathogenesis (Yanagi et al., 1997). The presentstudy was to obtain a full length cDNA clone of the GCV1 genome
Giardia lamblia; GLV, Giardia lamblia virus; RNA, ribonucleic acid; bp, base pair;dsRNA, double-stranded RNA; RT, reverse transcription; PCR, polymerase chainreaction; cDNA, complementary DNA; pPoly2/sfinot, plasmid pPoly2/sfinot; PTA, pureterephthalic acid.

L. Cao et al. / Experimental Parasitology 123 (2009) 212–217 213
and to produce RNA transcripts to be used to transfect virus-free G.canis trophozoites. The goal of this study was to determine if thein vitro transcribed transcripts from GCV cDNA were able to infectG. canis trophozoites, to replicate in and to be released from infectedtrophozoites. The formation of viral particles and their release fromthe parasite host was examined using the transmission electronmicroscopy. The Infectivity of released mature viral particles wasalso confirmed.
2. Materials and methods
2.1. Parasite and cell culture
This experiment was carried out with two G. canis isolates,GCVF and GCVNI. GCVF is a virus-free strain (Zhao et al., 2005),while GCVNI is naturally infected with a dsRNA virus (Chenet al., 2006). Both strains were isolated from Changchun in China,and maintained in a modified TYI-S-33 medium as described pre-viously (Lu et al., 1990). Infectivity of virus particles released fromtransfected trophozoites was determined using virus-free Giardiatrophozoites (Tai et al., 1991). Briefly, supernatant containing thevirus particles was collected, filtered through 0.22 lm filters(Millipore), and mixed with GCVF trophozoites in modified TYI-S-33 medium. The mixture was then incubated at 4 �C for 3 h andtransferred to 37 �C and continued to incubate overnight.
2.2. PCR primers
Three pairs of PCR primers were designed based on published se-quence of GCV (GenBank Accession No. DQ238861). The entire se-quence was divided into three overlapping fragments which werePCR amplified using the following primers. It was so designed thatthe fragments carrying EcoRI/BalI (F1/R1), BalI/ScaI (F2/R2) andScaI/PstI (F3/R3) were to be ligated to full length sequence throughthe unique restriction sites. In order to in vitro transcribe the GCVcDNA, an 18 nucleotide sequence encoding the T7 RNA polymerase
Fig. 1. Cloning strategy for the full length genome of GCV. The 6276 bp GCV genome wunique restriction sites. The three fragments individually cloned into the pMD18-T vectointo the pPoly2/sfinot vector.
promoter was included in primer F1 and followed by the 50 UTR ofGCV.
F1 (50-GAATTCAGTTAATACGACTCACTATAGGGAAGGAGTGCCAGGCCATTACC-30), R1 (50-GTAGGTTGGCCAAACGGTAGTGTCATAC-30);F2 (50-GGATACGGTCGAGCTTAGTGTACAGC-30);R2 (50-CAGAGCTCCGACCTCGCACAGTACTCG-30);F3 (50-ATTCGTTCCTGTCACTCTCAGAAGTGC-30);R3 (50-CTGCAGCGACCCCCTCGTACGCTGCCTCCTACAG-30).
2.3. Construction of full length GCV cDNA clones
Total RNA from the GCVNI trophozoites was extracted withTRIzol system (Invitrogen) and RT-PCR was performed using anAccessQuickTM RT-PCR System (Promega) according to manufac-turer’s instructions. The PCR-amplified fragments were cloneddirectly into pMD18-T vector (TaKaRa). Prior to ligation, pMD18-T-F1R1, pMD18-T-F2R2 and pMD18-T-F3R3 with inserts of expectedsizes were digested with EcoRI/BalI (pMD18-T-F1R1), BalI/ScaI(pMD18-T-F2R2) and ScaI/PstI (pMD18-T-F3R3). The pPoly2/sfinotvector plasmid (gift from Dr. Kremer, Institutde GénétiqueMoléculaire de Montpellier, Montpellier, France) was digested withEcoRI/PstI and the three PCR products were ligated into this vectorwith a molar ratio of 4:4:4:1 using T4 DNA ligase (Promega)(Fig. 1).
2.4. In vitro transcription
The pPpoly2/sfinot-GCV plasmid was linearized with EcoRI andpurified by phenol/chloroform extraction and isopropanol precipi-tation. The in vitro transcription was performed with the T7RiboMAXTM Express Large Scale RNA Production System (Promega)as described previously (Yu et al., 1995a). RNA transcript was puri-fied by phenol/chloroform extraction and isopropanol precipita-
as designed to be cloned in three fragments by PCR using primers engineered withr were excised using EcoRI + BalI, BalI + ScaI and ScaI + PstI, respectively, and cloned
214 L. Cao et al. / Experimental Parasitology 123 (2009) 212–217
tion in the presence of 1 M ammonium acetate and dissoved inRNase-free water. The purified RNA was analyzed by agarose gel(1%) electrophoresis and used in electroporation experiments.
2.5. Electroporation and analysis of GCV RNA in transfectedtrophozoites
The in vitro transcribed transcripts from full length cDNA tem-plate were electroporated into the GCVF trophozoites as describedpreviously (Singer et al., 1998; Yu et al., 1995a,b, 2005). Approxi-mately 5 � 106 trophozoites were electroporated with 100 lg ofthe RNA at 1000 V/cm for 8 ms (BTX ECM830 Electroporator).The cuvettes were placed on ice for 15 min before the cells wereadded into the fresh culture medium incubated at 37 �C. The GCVRNA was analyzed in total nucleic acid (DNA and RNA) extractedfrom trophozoites 72 h post transfection. Non-transfected tropho-zoites were used as control. The Giardia trophozoites were pelletedand washed twice in PBS, the total nucleic acid was extracted withTRIzol (Invitrogen). Samples were electrophoresed on a 0.8% aga-rose gel and stained with ethidium bromide.
Negative staining was used to detect virus-like particles intransfected trophozoites as described previously (Sherwin andGull, 1989; Robinson et al., 1991). Briefly, the trophozoites werecollected, pelleted by centrifugation 72 h after transfection andwashed twice in PBS. Trophozoites were then resuspended in0.5 ml sterilized water and subjected to three freeze/thaw cyclesin liquid nitrogen or 37 �C. The lysate was cleared by low-speedcentrifugation followed by ultracentrifugation at 50,000 � g for15 min and the resultant pellet was used for staining by 2% PTA.
2.6. Ultrastructural analysis
After transfection, the GCVF trophozoites were incubatedat 37 �C, and collected at 12, 24, 36, 48, 60, or 72 h post transfec-tion. The samples were fixed with 2.5% glutaraldehyde in 0.1 Msodium cacodylate buffer, pH 7.2, at 4 �C for 24–48 h, and post-
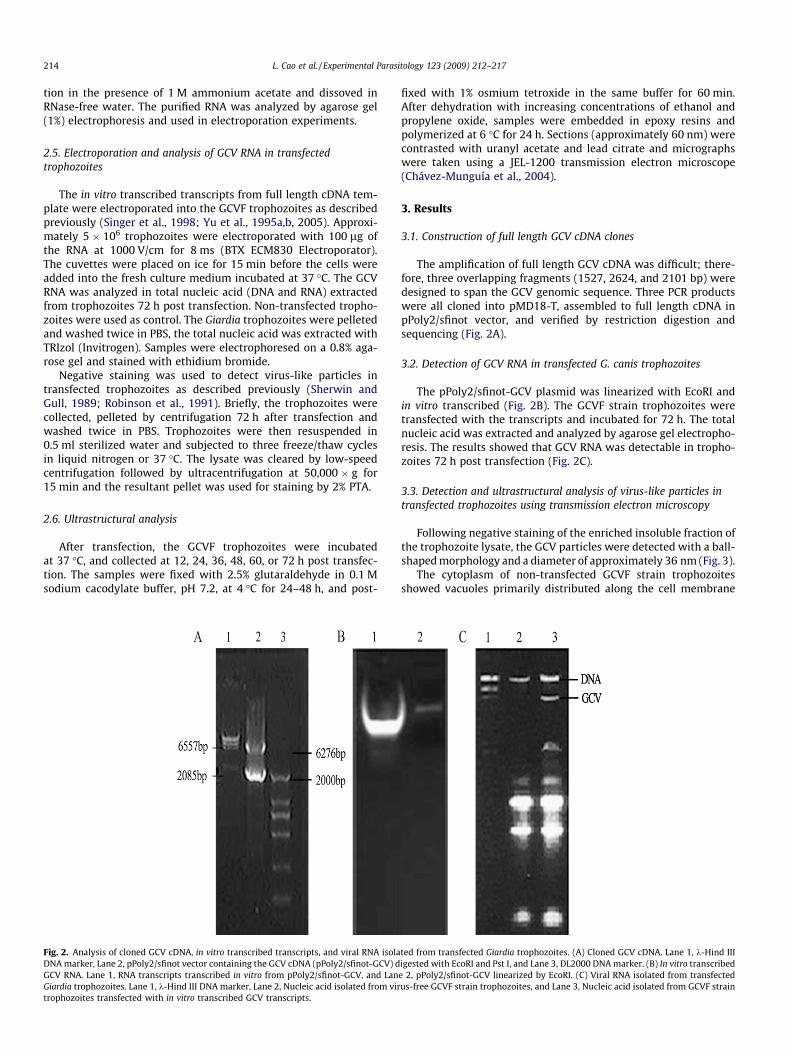
Fig. 2. Analysis of cloned GCV cDNA, in vitro transcribed transcripts, and viral RNA isolaDNA marker, Lane 2, pPoly2/sfinot vector containing the GCV cDNA (pPoly2/sfinot-GCV) dGCV RNA. Lane 1, RNA transcripts transcribed in vitro from pPoly2/sfinot-GCV, and LanGiardia trophozoites. Lane 1, k-Hind III DNA marker, Lane 2, Nucleic acid isolated from virtrophozoites transfected with in vitro transcribed GCV transcripts.
fixed with 1% osmium tetroxide in the same buffer for 60 min.After dehydration with increasing concentrations of ethanol andpropylene oxide, samples were embedded in epoxy resins andpolymerized at 6 �C for 24 h. Sections (approximately 60 nm) werecontrasted with uranyl acetate and lead citrate and micrographswere taken using a JEL-1200 transmission electron microscope(Chávez-Munguía et al., 2004).
3. Results
3.1. Construction of full length GCV cDNA clones
The amplification of full length GCV cDNA was difficult; there-fore, three overlapping fragments (1527, 2624, and 2101 bp) weredesigned to span the GCV genomic sequence. Three PCR productswere all cloned into pMD18-T, assembled to full length cDNA inpPoly2/sfinot vector, and verified by restriction digestion andsequencing (Fig. 2A).
3.2. Detection of GCV RNA in transfected G. canis trophozoites
The pPoly2/sfinot-GCV plasmid was linearized with EcoRI andin vitro transcribed (Fig. 2B). The GCVF strain trophozoites weretransfected with the transcripts and incubated for 72 h. The totalnucleic acid was extracted and analyzed by agarose gel electropho-resis. The results showed that GCV RNA was detectable in tropho-zoites 72 h post transfection (Fig. 2C).
3.3. Detection and ultrastructural analysis of virus-like particles intransfected trophozoites using transmission electron microscopy
Following negative staining of the enriched insoluble fraction ofthe trophozoite lysate, the GCV particles were detected with a ball-shaped morphology and a diameter of approximately 36 nm (Fig. 3).
The cytoplasm of non-transfected GCVF strain trophozoitesshowed vacuoles primarily distributed along the cell membrane
ted from transfected Giardia trophozoites. (A) Cloned GCV cDNA. Lane 1, k-Hind IIIigested with EcoRI and Pst I, and Lane 3, DL2000 DNA marker. (B) In vitro transcribede 2, pPoly2/sfinot-GCV linearized by EcoRI. (C) Viral RNA isolated from transfectedus-free GCVF strain trophozoites, and Lane 3, Nucleic acid isolated from GCVF strain
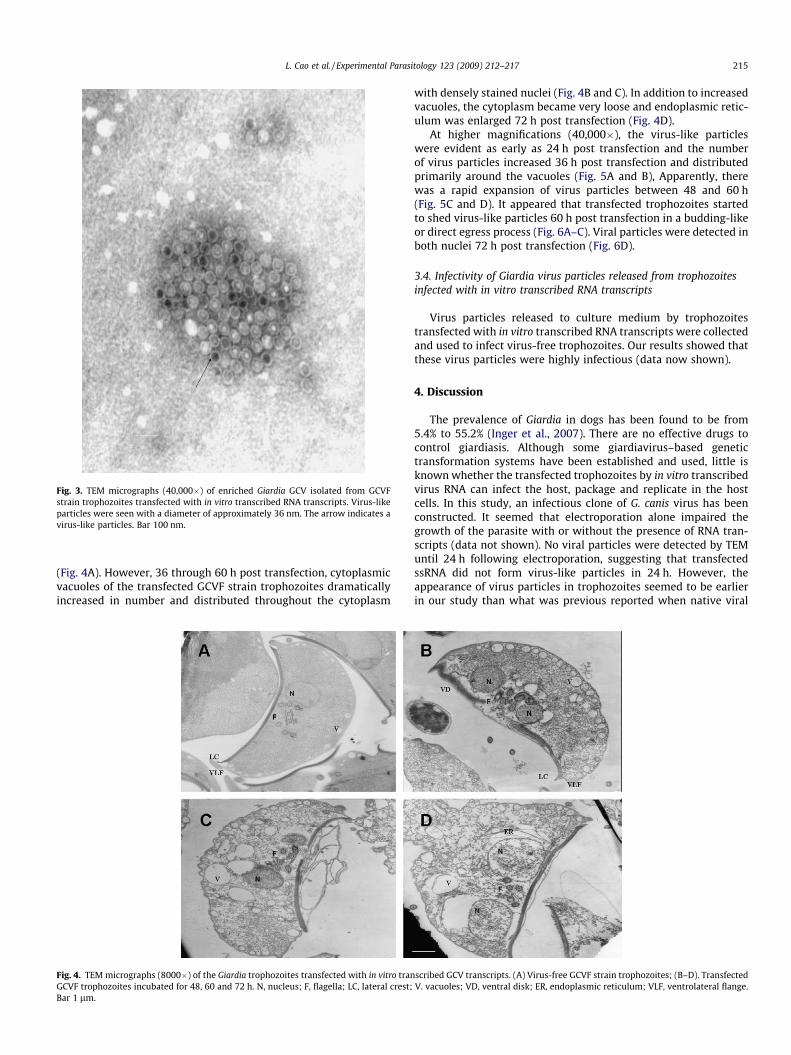
Fig. 3. TEM micrographs (40,000�) of enriched Giardia GCV isolated from GCVFstrain trophozoites transfected with in vitro transcribed RNA transcripts. Virus-likeparticles were seen with a diameter of approximately 36 nm. The arrow indicates avirus-like particles. Bar 100 nm.
L. Cao et al. / Experimental Parasitology 123 (2009) 212–217 215
(Fig. 4A). However, 36 through 60 h post transfection, cytoplasmicvacuoles of the transfected GCVF strain trophozoites dramaticallyincreased in number and distributed throughout the cytoplasm
Fig. 4. TEM micrographs (8000�) of the Giardia trophozoites transfected with in vitro tranGCVF trophozoites incubated for 48, 60 and 72 h. N, nucleus; F, flagella; LC, lateral crest;Bar 1 lm.
with densely stained nuclei (Fig. 4B and C). In addition to increasedvacuoles, the cytoplasm became very loose and endoplasmic retic-ulum was enlarged 72 h post transfection (Fig. 4D).
At higher magnifications (40,000�), the virus-like particleswere evident as early as 24 h post transfection and the numberof virus particles increased 36 h post transfection and distributedprimarily around the vacuoles (Fig. 5A and B), Apparently, therewas a rapid expansion of virus particles between 48 and 60 h(Fig. 5C and D). It appeared that transfected trophozoites startedto shed virus-like particles 60 h post transfection in a budding-likeor direct egress process (Fig. 6A–C). Viral particles were detected inboth nuclei 72 h post transfection (Fig. 6D).
3.4. Infectivity of Giardia virus particles released from trophozoitesinfected with in vitro transcribed RNA transcripts
Virus particles released to culture medium by trophozoitestransfected with in vitro transcribed RNA transcripts were collectedand used to infect virus-free trophozoites. Our results showed thatthese virus particles were highly infectious (data now shown).
4. Discussion
The prevalence of Giardia in dogs has been found to be from5.4% to 55.2% (Inger et al., 2007). There are no effective drugs tocontrol giardiasis. Although some giardiavirus–based genetictransformation systems have been established and used, little isknown whether the transfected trophozoites by in vitro transcribedvirus RNA can infect the host, package and replicate in the hostcells. In this study, an infectious clone of G. canis virus has beenconstructed. It seemed that electroporation alone impaired thegrowth of the parasite with or without the presence of RNA tran-scripts (data not shown). No viral particles were detected by TEMuntil 24 h following electroporation, suggesting that transfectedssRNA did not form virus-like particles in 24 h. However, theappearance of virus particles in trophozoites seemed to be earlierin our study than what was previous reported when native viral
scribed GCV transcripts. (A) Virus-free GCVF strain trophozoites; (B–D). TransfectedV. vacuoles; VD, ventral disk; ER, endoplasmic reticulum; VLF, ventrolateral flange.
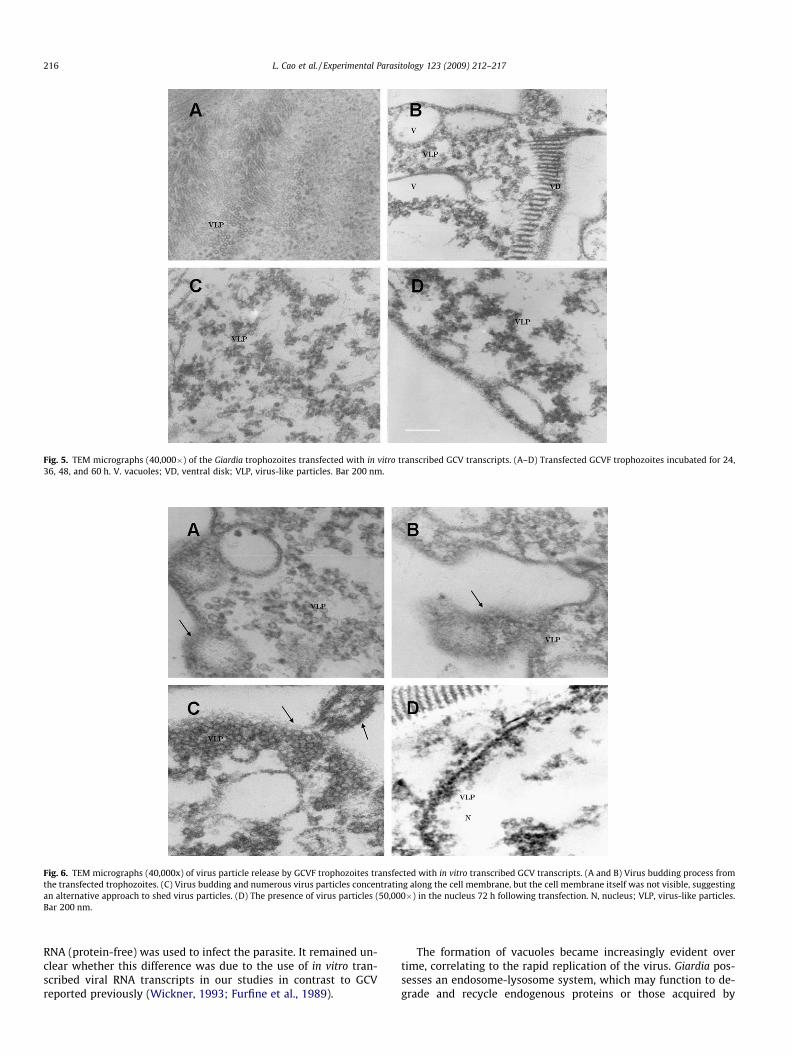
Fig. 5. TEM micrographs (40,000�) of the Giardia trophozoites transfected with in vitro transcribed GCV transcripts. (A–D) Transfected GCVF trophozoites incubated for 24,36, 48, and 60 h. V. vacuoles; VD, ventral disk; VLP, virus-like particles. Bar 200 nm.
Fig. 6. TEM micrographs (40,000x) of virus particle release by GCVF trophozoites transfected with in vitro transcribed GCV transcripts. (A and B) Virus budding process fromthe transfected trophozoites. (C) Virus budding and numerous virus particles concentrating along the cell membrane, but the cell membrane itself was not visible, suggestingan alternative approach to shed virus particles. (D) The presence of virus particles (50,000�) in the nucleus 72 h following transfection. N, nucleus; VLP, virus-like particles.Bar 200 nm.
216 L. Cao et al. / Experimental Parasitology 123 (2009) 212–217
RNA (protein-free) was used to infect the parasite. It remained un-clear whether this difference was due to the use of in vitro tran-scribed viral RNA transcripts in our studies in contrast to GCVreported previously (Wickner, 1993; Furfine et al., 1989).
The formation of vacuoles became increasingly evident overtime, correlating to the rapid replication of the virus. Giardia pos-sesses an endosome-lysosome system, which may function to de-grade and recycle endogenous proteins or those acquired by

L. Cao et al. / Experimental Parasitology 123 (2009) 212–217 217
endocytosis or phagocytosis from the extracellular space(Lanfredi-Rangelb et al., 1998). The increased formation of periph-eral and cytoplasmic vacuoles shown in this study may be a re-sult of stimulation of the rapidly replicating virus introphozoites (De et al., 1982). Similarly, the enlarged ER in viraltranscript infected trophozoites may be indicative of increasedcellular activities and it is unlikely to be related to pathologycaused by the virus since the virus has been reportedly harmlessto the host. Following transfection, the nuclei became denselystained at 36 h; however, virus particles were not detectable inboth nuclei until 72 h, which is consistent with previous studies(Tai et al., 1993).
It has been reported that the Giardia virus particles may be dis-charged into the environment(Tai et al., 1991), but very little isknown about how the virus particles are released. Our findingsindicate that Giardia virus may be released by budding. This wasshown in the TEM micrographs that there appeared to be a gradualprocess of budding formation until the virus was eventually dis-charged (Fig. 6A–C). Alternatively, the virus may also be releasedfrom the infected host by lysis of infected triophozoites. Our resultsshowed that numerous virus particles were crowded along the cellmembrane and there was an apparent loss of the cell membrane,suggesting that the Giardia virus may in fact lyse the host and bereleased (Lenstra et al., 1988).
The fact that virus particles released from in vitro transcribedRNA infected trophozoites retained high levels of infectivity indi-cate that our approach was successful and may be used formolecular manipulation of the virus. In conclusion, we have usedthe infectious RNA transcripts derived from GCV cDNA to trans-fect virus-free Giardia trophozoites. Our data showed that thein vitro transcribed Giardia RNA transcripts were able to infectthe host, rapidly replicate/package in and released from the hostcells.
Acknowledgment
This project was supported by the National Natural ScienceFoundation of China (No. 30300260).
References
Adam, R.D., 2001. Biology of Giardia lamblia. Clinical Microbiology Reviews 14, 447–475.
Adam, R.D., 1991. The Biology of Giardia spp.. Microbiology Reviews 55, 706–732.Chen, L.F., Li, J.H., Zhang, X.C., Liu, Q., Yin, J.G., Yao, L.Q., Zhao, Y.P., Cao, L.L., 2007.
Inhibition of Krr1 gene expression in Giardia canis by a virus mediatedhammerhead ribozyme. Veterinary Parasitology 143, 14–20.
Chen, L.F., Li, J.H., Zhang, X.C., Liu, Q., Zhao, Y.P., Cao, L.L., 2006. Establishment ofin vitro cultivation of Giardia canis trophozoites infected with Giardia canisvirus. Zhong guo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi 24, 261–265.
Chávez-Munguía, B., Hernández-Ramírez, V., Angel, A., Ríos, A., Talamás-Rohana, P.,González-Robles, A., González-Lázaro, M., Martínez-Palomo, A., 2004.Entamoeba histolytica: ultrastructure of trophozoites recovered fromexperimental liver lesions. Experimental Parasitology 107, 39–46.
De, B.P., Thornton, G.B., Luk, D., Banerjee, A.K., 1982. Purified matrix protein ofvesicular stomatitis virus blocks viral transcription in vitro. Proceedings of theNational Academy of Sciences of United States of America 79, 7137–7141.
Furfine, E.S., White, T.C., Wang, A.L., Wang, C.C., 1989. A single-stranded RNA copy ofthe Giardia lamblia virus double-stranded RNA genome is present in the infectedGiardia lamblia. Nucleic Acids Research 17, 7453–7467.
Inger, S.H., Bjørn, K.G., Lucy, J.R., 2007. A longitudinal study on the occurrence ofCryptosporidium and Giardia in dogs during their first year of life. ActaVeterinaria Scandinavica 49, 22.
Johan, F., De, J., Bart, G., 1987. Occurrence and transfection of a Giardia virus.Molecular and Biochemical Parasitology 23 (1), 85–89.
Liu, Q., Zhang, X., Li, J., Yin, J., Chen, L., Zhao, Y., Wei, F., Wu, T., 2005. Giardia lamblia:stable expression of green fluorescent protein mediated by giardiavirus.Experimental Parasitology 109, 181–187.
Lu, S.Q., Wang, Z.Y., Zhu, H., 1990. Establishment of an axenic culture of Giardialamblia through preliminary passage in suckling gerbil. Chinese Medical Journal103, 583–587.
Liu, C.W., Li, J.H., Zhang, X.C., Liu, Q., Liu, H., Gong, P.T., Zhang, G.C., Yao, L.Q., Zhang,X.X., 2008. Stable expression of green fluorescent protein mediated by GCV inGiardia canis. Parasitology International 57, 320–324.
Lanfredi-Rangelb, A., Attiasb, M., de Carvalhob, T.M.U., Kattenbachc, W.M., DeSouzab, W., 1998. The peripheral vesicles of trophozoites of the primitiveprotozoan Giardia lamblia May correspond to early and late endosomes and tolysosomes. Journal of Structural Biology 123, 225–235.
Lenstra, R., Samso, A., Andrieu, B., Le Bras, J., Galibert, F., 1988. Virus-like particlescontaining knob-associated histidine-rich protein are secreted into the culturemedium of Plasmodium falciparum in vitro cultures. Biochemical and BiophysicalResearch Communications 151, 749–757.
Robinson, D., Beattie, P., Sherwin, T., Gull, K., 1991. Microtubules, tubulin, andmicrotubule-associated proteins of trypanosomes. Methods in Enzymology 196,285–299.
Sepp, T., Wang, A.L., Wang, C.C., 1994. Giardiavirus-resistant Giardia lamblia lacks avirus receptor on the cell membrane surface. The Journal of Virology 68 (3),1426–1431.
Singer, S.M., Yee, J., Nash, T.E., 1998. Episomal and integrated maintenance of foreignDNA in Giardia lamblia. Molecular and Biochemical Parasitology 92, 59–69.
Sherwin, T., Gull, K., 1989. The cell division cycle of Trypanosoma brucei brucei:timing of event markers and cytoskeletal modulations. PhilosophicalTransactions of the Royal Society B: Biological Sciences 323, 573–588.
Tai, J.H., Ong, S.J., Chang, S.C., Su, H.M., 1993. Giardiavirus enters Giardia lamblia WBtrophozoite via endocytosis. Experimental Parasitology 76, 65–74.
Tai, J.H., Wang, A.L., Ong, S.J., Lai, K.S., Lo, C., Wang, C.C., 1991. The course ofgiardiavirus infection in the Giardia lamblia trophozoites. ExperimentalParasitology 73, 413–423.
Upcroft, J., Upcroft, P., 1998. My favourite cell: Giardia. BioEssays 20, 256–263.Wang, A.L., Miller, R.L., Wang, C.C., 1988. Antibodies to the Giardia lamblia double-
stranded RNA virus major protein can block the viral infection. Molecular andBiochemical Parasitology 30, 225–232.
Wang, A.L., Wang, C.C., 1986. Discovery of a specific double-stranded RNA virus inGiardia lamblia. Molecular and Biochemical Parasitology 21, 269–276.
Wang, A.L., Yang, H.M., Shen, K.A., Wang, C.C., 1993. Giardiavirus double-strandedRNA genome encodes a capsid polypeptide and a gag-pol-like fusion protein bya translation frameshift. Proceedings of the National Academy of Sciences ofUnited States of America 90, 8595–8599.
Wickner, R.B., 1993. Double-strandeded RNA virus replication and packaging.Journal of Biological Chemistry 268, 3797–3800.
Yanagi, M., Purcell, R.H., Emerson, S.U., Bukh, J., 1997. Transcripts from a single full-length cDNA clone of hepatitis C virus are infectious when directly transfectedinto the liver of a chimpanzee. Proceeding of National Academy of Sciences 94,8738–8743.
Yu, D.C., Wang, A.C., Wu, C.H., Wang, C.C., 1995a. Virus mediated expression offirefly luciferase in the parasitic protozoan Giardia lamblia. Molecular andCellular Biology 15, 4867–4872.
Yu, D.C., Wang, C.C., Wang, A.L., 1995b. Maturation of giardiavirus capsid proteininvolves posttranslational proteolytic processing by a cysteine protease. Journalof Virology 69, 2825–2830.
Zhao, Y.J., Zhang, X.C., Liu, Q., Li, J.H., Yin, J.G., Yang, J., Wu, T., 2005. Axeniccultivation of Giardia canis. Chinese Journal of Zoonoses 21, 706–709.