Embed Size (px)
Citation preview

Glucocorticoid receptor interacts with PNRC2 ina ligand-dependent manner to recruit UPF1 forrapid mRNA degradationHana Cho1,2, Ok Hyun Park1, Joori Park, Incheol Ryu, Jeonghan Kim, Jesang Ko, and Yoon Ki Kim3
Division of Life Sciences, Korea University, Seoul 136-701, Republic of Korea
Edited by Allan Jacobson, University of Massachusetts Medical School, Worcester, MA, and accepted by the Editorial Board February 11, 2015 (received forreview May 23, 2014)
Glucocorticoid receptor (GR), which was originally known to functionas a nuclear receptor, plays a role in rapid mRNA degradation byacting as an RNA-binding protein. The mechanism by which thisprocess occurs remains unknown. Here, we demonstrate that GR,preloaded onto the 5′UTR of a target mRNA, recruits UPF1 throughproline-rich nuclear receptor coregulatory protein 2 (PNRC2) in aligand-dependent manner, so as to elicit rapid mRNA degradation.We call this process GR-mediated mRNA decay (GMD). AlthoughGMD, nonsense-mediated mRNA decay (NMD), and staufen-mediatedmRNA decay (SMD) share upstream frameshift 1 (UPF1) and PNRC2,we find that GMD is mechanistically distinct from NMD and SMD.We also identify de novo cellular GMD substrates using microarrayanalysis. Intriguingly, GMD functions in the chemotaxis of humanmonocytes by targeting chemokine (C-C motif) ligand 2 (CCL2)mRNA. Thus, our data provide molecular evidence of a posttran-scriptional role of the well-studied nuclear hormone receptor, GR,which is traditionally considered a transcription factor.
glucocorticoid receptor | PNRC2 | UPF1 | glucocorticoid receptor-mediatedmRNA decay | Nonsense-mediated mRNA decay
At the cellular level, glucocorticoid receptor (GR), whichbelongs to the nuclear receptor superfamily, functions as
a transcription factor regulating various physiological processes(1–3). In the presence of a glucocorticoid, which diffuses throughthe plasma membrane into the cytoplasm, cytosolic GR bindsto the glucocorticoid. The resulting glucocorticoid–GR com-plex is activated and then enters the nucleus. Once in the nu-cleus, GR dimerizes, binds to specific cis-acting elements, andrecruits coregulatory proteins for transcriptional activation orrepression (4, 5).The majority of the coregulatory proteins commonly contain
a nuclear receptor box (NR box, also called an LXXLL motif),which is important for interactions between coregulatory pro-teins and nuclear receptors (4–6). The proline-rich nuclear re-ceptor coregulatory protein (PNRC), however, is an exceptionbecause it interacts with nuclear receptors through an SH3-binding motif [SD(E)PPSPS] rather than an NR box (7, 8). TwoPNRC paralogs, PNRC1 and PNRC2, have been identified inmammalian cells (7, 8). PNRC1 and PNRC2 are believed to playsimilar roles in nuclear receptor-mediated signaling because theyinteract with similar groups of nuclear receptors.Although PNRC2 is known to function as a coregulatory
protein for nuclear receptors, it has a distinct function in mRNAdecay pathways including nonsense-mediated mRNA decay(NMD), staufen (STAU)-mediated mRNA decay (SMD), andreplication-dependent histone mRNA degradation (HMD) (9–13). NMD serves as a mechanism of both mRNA quality controland posttranscriptional regulation by selectively recognizing anddegrading cellular transcripts that are abnormal or that containa premature translation termination codon (PTC), as reviewedelsewhere (14–16). A key NMD factor, UPF1, is recruited toa terminating ribosome at a PTC. UPF1 then recruits generalRNA-degrading enzymes via adaptor/effector proteins such as
PNRC2, suppressor with morphogenetic effect on genitalia 5(SMG5), SMG6, and SMG7, resulting in rapid mRNA degra-dation. In SMD, UPF1 is recruited to mRNA via its interaction withSTAU, which recognizes a stem-loop structure in the 3′UTR or anRNA duplex structure formed by intermolecular mRNA–mRNAor mRNA–long noncoding RNA interactions. The recruited UPF1interacts with PNRC2 and triggers rapid mRNA degradation(17–19). NMD and SMD are closely related pathways becauseboth require UPF1 and both occur in a translation-dependentmanner. HMD, another UPF1-dependent mRNA degradationpathway, also requires a translation event, as reviewed else-where (20).Here, we demonstrate a novel mRNA decay pathway induced
by ligand-bound GR loaded onto the 5′UTR of target mRNAs.We call this process GR-mediated mRNA decay (GMD). Weshow that GMD is a mechanistically unique pathway of UPF1-dependent mRNA decay because it occurs in a translation-independent manner. In addition, efficient joining of UPF1 toGMD machinery requires a glucocorticoid-induced GR–PNRC2interaction. We also provide evidence that the regulation ofGMD efficiency by a glucocorticoid is necessary for chemotaxis
Significance
Glucocorticoid receptor (GR) belongs to the nuclear receptorsuperfamily and functions as a transcription factor. GR regu-lates various physiological processes, including cell prolif-eration, energy homeostasis, and inflammation. In this study,we provide molecular evidence for the role of GR in the regu-lation of mRNA stability, which we term GR-mediated mRNAdecay (GMD). Efficient GMD requires a ligand, a GR loadedonto target mRNA, upstream frameshift 1 (UPF1), and proline-rich nuclear receptor coregulatory protein 2. GMD functions inthe chemotaxis of human monocytes by targeting chemokine(C-C motif) ligand 2 mRNA. Thus, we unravel a previously un-appreciated role of GR, which is traditionally considered atranscription factor, in posttranscriptional regulation.
Author contributions: H.C., O.H.P., J.P., I.R., J. Kim, J. Ko, and Y.K.K. designed research;H.C., O.H.P., J.P., I.R., and J. Kim performed research; H.C., O.H.P., J.P., I.R., and Y.K.K.analyzed data; and H.C., O.H.P., J. Ko, and Y.K.K. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission. A.J. is a guest editor invited by the EditorialBoard.
Freely available online through the PNAS open access option.
Data deposition: The data reported in this paper have been deposited in the Gene Ex-pression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession no. GSE49591).1H.C. and O.H.P. contributed equally to this work.2Present address: Department of Biochemistry and Biophysics, School of Medicine andDentistry, University of Rochester, Rochester, NY 14642.
3To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1409612112/-/DCSupplemental.
E1540–E1549 | PNAS | Published online March 16, 2015 www.pnas.org/cgi/doi/10.1073/pnas.1409612112

of human monocytes; these data are suggestive of the physio-logical importance of GMD.
ResultsPNRC2 Interacts with GR in a Ligand-Dependent Manner, RecruitingUPF1 and Decapping Enzyme 1a to the GR Complex. It has beenreported that GR binds directly to a subset of mRNAs (21, 22). Ithas also been reported that GR binds to PNRC2 in a ligand-dependent manner (in a yeast two-hybrid system), albeit undernonphysiological conditions (8, 23). We have shown that PNRC2physically interacts with UPF1 and decapping enzyme 1a (DCP1A;a component of the decapping complex), thereby triggering NMD(11). According to these previous reports, we hypothesized thatGR is complexed with UPF1 and DCP1A via its direct inter-action with PNRC2 in a ligand-dependent manner.To test this possibility, we first looked for differences in the
composition of the GR-containing complex either in the absenceor in the presence of dexamethasone (Dex), which is a potentsynthetic glucocorticoid (24). Immunoprecipitation (IP) experi-ments were performed using α-GR antibody and RNase A-treatedtotal extracts of HEK293T cells transiently expressing MYC-PNRC2 and FLAG-UPF1. The cells were either pretreated ornot treated with Dex before IP. The levels of transiently ex-pressed MYC-PNRC2 and FLAG-UPF1 were comparable to,and lower than, those of endogenous PNRC2 and UPF1, re-spectively (Fig. S1). RT-PCR of endogenous GAPDH mRNAusing α-[32P]-dATP and specific oligonucleotides demonstratedefficient removal of cellular RNAs by RNase A treatment beforeIPs (Fig. 1A, Lower). The IP results revealed that MYC-PNRC2,FLAG-UPF1, and endogenous DCP1A coimmunopurified withendogenous GR in nine-, seven-, and sixfold greater amounts,respectively, after Dex treatment (Fig. 1A, Upper). Other NMDfactors, such as UPF2, UPF3X, and eIF4AIII, and an SMDfactor, STAU1, were not detectably enriched in GR IP, regard-less of Dex treatment. These results suggest that Dex treatmenttriggers efficient formation of a GR complex containing PNRC2,UPF1, and DCP1A.Next, we evaluated the role of PNRC2 in the formation of the
GR-containing complex. To this end, we conducted IP experi-ments using α-GR antibody and the extracts of either undepletedor PNRC2-depleted cells. The cells were pretreated with Dexbefore IP to stimulate formation of the GR-containing complex.Transfection of PNRC2 siRNA selectively reduced the level ofendogenous PNRC2, 2% of the normal level (Fig. 1B). The IPresults showed that after the PNRC2 down-regulation, FLAG-UPF1 and endogenous DCP1A coimmunopurified with GR in5- and 10-fold smaller amounts, respectively (Fig. 1C), suggestingthat PNRC2 stabilizes or serves as a link for the associationsbetween GR and UPF1, and between GR and DCP1A. BecauseGR directly binds to the 5′UTR of specific mRNAs even in theabsence of a ligand (21, 22), it is likely that mRNA-bound GRrecruits PNRC2 or the PNRC2/UPF1/DCP1A complex tomRNA in the presence of a ligand.
GR-Mediated mRNA Decay Targets a Subset of Cellular mRNAs ina Manner Requiring PNRC2, UPF1, and GR. The results shown inFig. 1 led us to hypothesize that the recruitment of UPF1 andDCP1A to GR-bound mRNA through PNRC2 may elicit rapidmRNA degradation, which we termed GR-mediated mRNAdecay. In support of this idea, it was reported that GR bindsdirectly to the 5′UTR of chemokine (C-C motif) ligand 2 (CCL2)mRNA and CCL7 mRNA, and destabilizes these mRNAs in thepresence of Dex (21, 22). To test the above hypothesis, we firstsearched for natural GMD substrates by performing microarrayanalysis using 48K human gene chips and total transcripts puri-fied from HeLa cells that were either treated or not treated withDex. The microarray analysis revealed that 147 and 77 tran-scripts were up-regulated and down-regulated, respectively, by at
least 1.5-fold after Dex treatment (Dataset S1). The GMDsubstrates are expected to be down-regulated after Dex treat-ment. We next compared the list of transcripts down-regulatedafter Dex treatment with previously reported microarray data ontranscripts coimmunoprecipitated with GR (22), and finallyobtained four natural transcripts that were putative GMD sub-strates. Interestingly, among them was CCL2 mRNA, which isknown to be rapidly degraded as a result of Dex treatment. Theothers were transcripts encoding B-Cell CLL/lymphoma 3 (BCL3),zinc finger SWIM-type containing 4 (ZSWIM4), and pleckstrinhomology-like domain, family A, member 1 (PHLDA1).We next tested whether the putative GMD substrates re-
quire PNRC2, UPF1, and GR for efficient decay. To this end,
A Dex
mIg
Gα-
GR
mIg
Gα-
GR
IP
GR
FLAG-UPF1
1
1
6
7DCP1A
MYC-PNRC21 9
GAPDH mRNA
Before IPRNase ADex
RT-
PC
R
Before IP After IP
RNase A
β-actin
eIF4AIII
UPF3X
UPF2
STAU1
B
Con
trol
PN
RC
2
PNRC2
siRNA
β-actin
C
Con
trol
PN
RC
2
mIg
Gα-
GR
mIg
Gα-
GR
Con
trol
PN
RC
2
siRNA
IP
GR
β-actin
pcDNA3-FLAG-UPF1
Before IP After IP
Dex
1 0.2
FLAG-UPF1
1 0.1
DCP1A
Fig. 1. GR associates with UPF1 and DCP1A via PNRC2. (A) IP experimentswith endogenous GR. HEK293T cells were transiently cotransfected withplasmids expressing MYC-PNRC2 and FLAG-UPF1. Two days later, the cellswere either treated or not treated with Dex for 3 h. Cell extracts wereprepared and were either not treated or treated with RNase A for 15 minbefore IP. The IP experiments were performed using either α-GR antibody ora nonspecific mouse IgG (mIgG, control). The protein fractions before andafter IPs were analyzed using Western blotting with the indicated anti-bodies. The levels of coimmunopurified MYC-PNRC2, FLAG-UPF1, and en-dogenous DCP1A were normalized to the level of immunopurified GR. Thenormalized levels obtained from IPs from the extracts of cells not treatedwith Dex were arbitrarily set to 1.0. For quantitative analysis, threefold serialdilutions of total cell extracts that were obtained before IP were loaded inthe four leftmost lanes (Upper). Efficient digestion of cellular transcripts byRNase A treatment was demonstrated by the complete absence of cellularGAPDH mRNA according to RT-PCR using α-[32P]-dATP and specific oligo-nucleotides. For quantitative analysis, total cell RNA samples obtained be-fore IP were serially diluted twofold and analyzed using RT-PCR. The RT-PCRproducts were loaded in the four leftmost lanes (Lower). (B and C) IPexperiments of endogenous GR from the extracts of cells depleted of PNRC2.The cells were transiently transfected with either PNRC2 siRNA or a non-specific control siRNA. Two days later, the cells were retransfected witha plasmid expressing FLAG-UPF1. One day later, the cells were treated withDex for 3 h before IP. After that, cell extracts were prepared and treatedwith RNase A before IP. (B) A Western blot of PNRC2 demonstrating specificdown-regulation by siRNA. (C) IP experiments using either the α-GR antibodyor mIgG (control). The levels of coimmunopurified FLAG-UPF1 and endog-enous DCP1A were normalized to the level of immunopurified GR. Thenormalized levels obtained from IPs from the extracts of undepleted cellswere arbitrarily set to 1.0. All data represent at least two independentlyperformed transfection experiments, IPs, and RT-PCRs.
Cho et al. PNAS | Published online March 16, 2015 | E1541
BIOCH
EMISTR
YPN
ASPL
US

endogenous PNRC2, UPF1, and GR were down-regulated to15%, 8%, and 2% of the normal level, respectively, using specificsiRNA (Fig. S2 A and B). After that process, the levels and half-lives of all of the putative GMD substrates including CCL7mRNA, whose expression is known to be reduced by Dextreatment (21, 22), were analyzed using quantitative RT-PCR(RT-qPCR). The results showed that the levels and half-lives ofall of the putative GMD substrates were drastically decreasedafter Dex treatment and were significantly restored via the down-regulation of PNRC2, UPF1, or GR (Fig. 2 and Fig. S2 C andD). In contrast, the level and half-life of endogenous CCL5mRNA, which lacks a GR-binding site in its 5′UTR (22), werenot significantly affected by Dex treatment and by the down-regulation of PNRC2, UPF1, or GR. All these data indicate thatthese transcripts are bona fide substrates of GMD, which requiresPNRC2, UPF1, and GR.
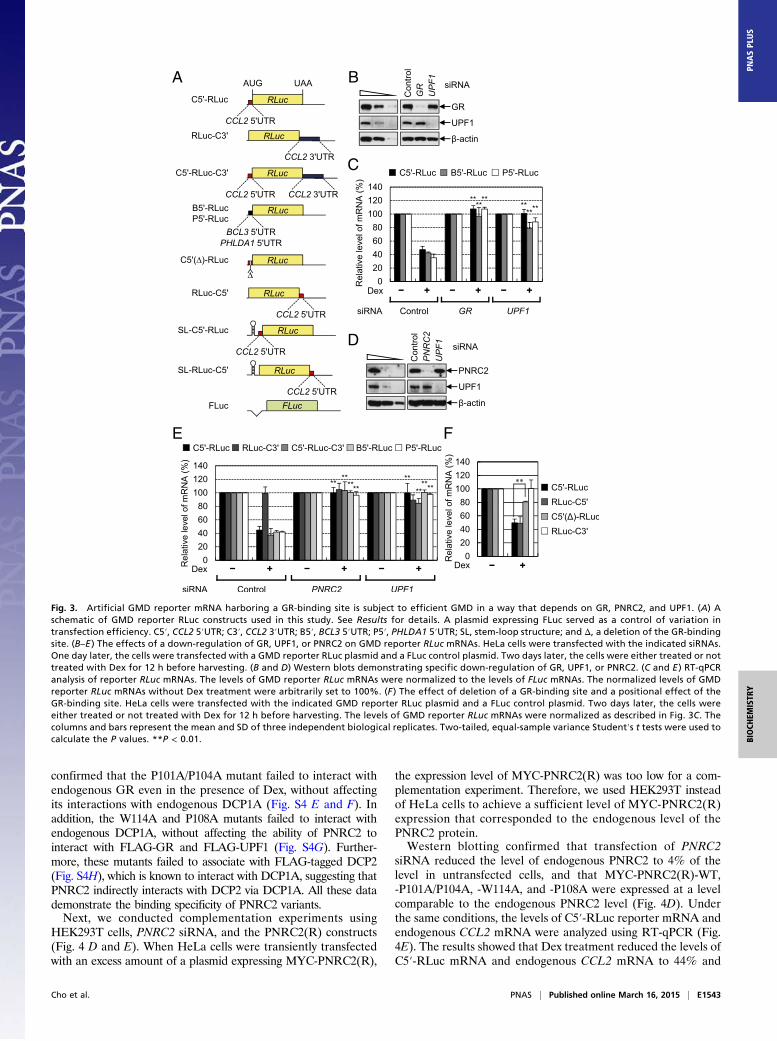
GMD Requires Binding of GR to a Target mRNA in a Position-Independent Manner. To gain molecular insight into GMD, wenext tested the positional effect of GR binding to mRNA. Forthis purpose, we designed several reporter constructs in whichthe 5′UTR, 3′UTR, or both, of CCL2 mRNA, BCL3 mRNA,or PHLDA1 mRNA were inserted immediately upstream ordownstream of the ORF of Renilla luciferase (RLuc) cDNA (Fig.3A). The levels of reporter mRNAs were monitored usingRT-qPCR. The results showed that Dex treatment reduced thelevels of C5′-RLuc mRNA, B5′-RLuc mRNA, and P5′-RLucmRNA, which contain the CCL2 5′UTR (C5′), BCL3 5′UTR(B5′), and PHLDA1 5′UTR (P5′), respectively, upstream of theRLucORF, to ∼40% of untreated levels (Fig. 3 B–E). The reduction
was almost completely reversed after a down-regulation of GR,PNRC2, or UPF1 (Fig. 3 B–E). The level of C5′-RLuc-C3′mRNA, which contains both the CCL2 5′UTR and CCL23′UTR (C3′) upstream and downstream, respectively, of theRLuc ORF, was also reduced after Dex treatment and restoredby a down-regulation of PNRC2 or UPF1 (Fig. 3E). On the otherhand, the level of RLuc-C3′ mRNA, which contains the CCL23′UTR downstream of the RLuc ORF, was not affected by Dextreatment and by down-regulation of PNRC2 or UPF1 (Fig. 3E),suggesting that the 5′UTR of GMD substrates is sufficient totrigger GMD.We further analyzed molecular features of GR binding to the
5′UTR of a GMD substrate. It is known that GR directly bindsto the region spanning nucleotides 44–60 within the 5′UTR ofCCL2 mRNA (22). Deletion of this region from C5′-RLucmRNA significantly inhibited GMD [C5′(Δ)-RLuc mRNA] (Fig.3F). We also designed RLuc-C5′ mRNA, which contains theCCL2 5′UTR downstream, rather than upstream, of the RLucORF. Intriguingly, the level of RLuc-C5′ mRNA was signifi-cantly reduced after Dex treatment to the extent observed inC5′-RLuc mRNA (Fig. 3F). The reduction was reversed by adown-regulation of GR, PNRC2, or UPF1, but not by a down-regulation of an NMD-specific factor, UPF2 (Fig. S3). These resultssuggest that loading of GR onto a GR-binding site within targetmRNA is sufficient for efficient GMD and that the position of theGR-binding site relative to the ORF is not critical for GMD.
The Loading of GR onto Target mRNA and Binding of GR to a LigandAre Crucial for GMD. Previous UV cross-linking studies using invitro-synthesized CCL2 mRNA and the extracts of cells eithertreated or not treated with Dex revealed that GR directly bindsto a GR-binding site within CCL2 mRNA (21, 22). It may bepossible, however, that Dex-induced signaling molecules orGR-related proteins other than GR itself are involved in the rec-ognition of a GR-binding site and induce GMD within the cells. Todetermine whether the loading of GR alone onto a target mRNAis sufficient for GMD, we used an artificial tethering system, thebacteriophage λN-5BoxB system (Fig. 4A). The results showedthat artificial tethering of λN-HA-GR, but not of λN-HA-GFP,to the 5′UTR of a reporter mRNA containing five tandemrepeats of the bacteriophage BoxB sequence (5BoxB) triggeredrapid degradation of the reporter mRNA in the presence of Dex,in a way that was completely reversed after down-regulation ofPNRC2 and UPF1, but not of UPF2 (Fig. 4B). The Dex-inducedrapid degradation of the reporter mRNA elicited by tetheredλN-HA-GR was almost completely blocked by either a singleamino acid substitution, V729I, which reduces the ligand-bindingability of GR (25), or by deletion of the ligand-binding domain(LBD) of GR (Fig. 4C). A specific down-regulation by siRNAand proper expression of the effectors were demonstrated bymeans of Western blotting (Fig. S4 A–C). These results suggestthat the binding of GR to the 5′UTR is sufficient for elicitingGMD and that direct interaction between GR and its ligand isnecessary for GMD.
An Interaction Between GR and DCP1A via PNRC2 Is Important forEfficient GMD. Because the ligand-dependent interaction betweenGR and PNRC2 occurs via an SH3-binding motif of PNRC2(7, 8), we further analyzed the role of the Dex-dependentGR–PNRC2 interaction in GMD using a complementation ap-proach. First, we generated siRNA-resistant PNRC2 constructs[PNRC2(R)] (Fig. S4D) that expressed: (i) MYC-tagged PNRC2WT; (ii) a double-mutant P101A/P104A, which has amino acidsubstitutions from proline to alanine at positions 101 and 104located within the SH3-binding motif and renders PNRC2 un-able to interact with GR (8) without affecting its interactionswith UPF1 (10); (iii) W114A; and (iv) P108A, both of which lackthe DCP1A-binding ability of PNRC2 (12). In this study, we also
Per
cent
rem
aini
ng (%
)
0 1 2Time (h)
BCL3 mRNA100
50
10
Per
cent
rem
aini
ng (%
)
0 1 2 3Time (h)
CCL2 mRNA100
50
10
t1/2 = 5.7t1/2 = 4.5t1/2 = 4.4t1/2 = 2.0
t1/2 = 5.3t1/2 = 4.1t1/2 = 3.1t1/2 = 1.6 Control siRNA, Dex +
PNRC2 siRNA, Dex +
UPF1 siRNA, Dex +
Control siRNA, Dex -
0 1 2 3Time (h)
CCL7 mRNA
t1/2 = 7.5t1/2 = 4.8t1/2 = 4.1t1/2 = 1.8
0 1 2 3Time (h)
CCL5 mRNA
0 1 2Time (h)
ZSWIM4 mRNA
t1/2 = 4.6t1/2 = 3.4t1/2 = 3.0t1/2 = 1.3
0 1 2Time (h)
PHLDA1 mRNA
t1/2 = 3.5t1/2 = 2.6t1/2 = 2.2t1/2 = 1.1
t1/2 = 30.1t1/2 = 24.6t1/2 = 49.5t1/2 = 30.1
Fig. 2. The half-lives of newly identified GMD substrates are increased aftera down-regulation of PNRC2 or UPF1. HeLa cells were transiently transfectedwith the indicated siRNAs. Three days after the transfection, the cells weretreated with DRB, which is a potent transcription inhibitor, and then treatedwith Dex. After that, total cell RNA and protein were prepared at the in-dicated time points after Dex treatment in the presence of DRB. Specificdown-regulation by siRNAs was demonstrated by Western blotting (Fig. S2A).The levels of the mRNAs tested, which were normalized to endogenousGAPDH mRNA, were plotted as a function of time after Dex treatment. Thenormalized levels of mRNAs at 0 h were arbitrarily set to 100%. The dots andbars represent the mean and SD of two independent biological replicates.
E1542 | www.pnas.org/cgi/doi/10.1073/pnas.1409612112 Cho et al.
confirmed that the P101A/P104A mutant failed to interact withendogenous GR even in the presence of Dex, without affectingits interactions with endogenous DCP1A (Fig. S4 E and F). Inaddition, the W114A and P108A mutants failed to interact withendogenous DCP1A, without affecting the ability of PNRC2 tointeract with FLAG-GR and FLAG-UPF1 (Fig. S4G). Further-more, these mutants failed to associate with FLAG-tagged DCP2(Fig. S4H), which is known to interact with DCP1A, suggesting thatPNRC2 indirectly interacts with DCP2 via DCP1A. All these datademonstrate the binding specificity of PNRC2 variants.Next, we conducted complementation experiments using
HEK293T cells, PNRC2 siRNA, and the PNRC2(R) constructs(Fig. 4 D and E). When HeLa cells were transiently transfectedwith an excess amount of a plasmid expressing MYC-PNRC2(R),
the expression level of MYC-PNRC2(R) was too low for a com-plementation experiment. Therefore, we used HEK293T insteadof HeLa cells to achieve a sufficient level of MYC-PNRC2(R)expression that corresponded to the endogenous level of thePNRC2 protein.Western blotting confirmed that transfection of PNRC2
siRNA reduced the level of endogenous PNRC2 to 4% of thelevel in untransfected cells, and that MYC-PNRC2(R)-WT,-P101A/P104A, -W114A, and -P108A were expressed at a levelcomparable to the endogenous PNRC2 level (Fig. 4D). Underthe same conditions, the levels of C5′-RLuc reporter mRNA andendogenous CCL2 mRNA were analyzed using RT-qPCR (Fig.4E). The results showed that Dex treatment reduced the levels ofC5′-RLuc mRNA and endogenous CCL2 mRNA to 44% and
A
C
B
D
E F
Fig. 3. Artificial GMD reporter mRNA harboring a GR-binding site is subject to efficient GMD in a way that depends on GR, PNRC2, and UPF1. (A) Aschematic of GMD reporter RLuc constructs used in this study. See Results for details. A plasmid expressing FLuc served as a control of variation intransfection efficiency. C5′, CCL2 5′UTR; C3′, CCL2 3′UTR; B5′, BCL3 5′UTR; P5′, PHLDA1 5′UTR; SL, stem-loop structure; and Δ, a deletion of the GR-bindingsite. (B–E ) The effects of a down-regulation of GR, UPF1, or PNRC2 on GMD reporter RLuc mRNAs. HeLa cells were transfected with the indicated siRNAs.One day later, the cells were transfected with a GMD reporter RLuc plasmid and a FLuc control plasmid. Two days later, the cells were either treated or nottreated with Dex for 12 h before harvesting. (B and D) Western blots demonstrating specific down-regulation of GR, UPF1, or PNRC2. (C and E ) RT-qPCRanalysis of reporter RLuc mRNAs. The levels of GMD reporter RLuc mRNAs were normalized to the levels of FLuc mRNAs. The normalized levels of GMDreporter RLuc mRNAs without Dex treatment were arbitrarily set to 100%. (F ) The effect of deletion of a GR-binding site and a positional effect of theGR-binding site. HeLa cells were transfected with the indicated GMD reporter RLuc plasmid and a FLuc control plasmid. Two days later, the cells wereeither treated or not treated with Dex for 12 h before harvesting. The levels of GMD reporter RLuc mRNAs were normalized as described in Fig. 3C. Thecolumns and bars represent the mean and SD of three independent biological replicates. Two-tailed, equal-sample variance Student′s t tests were used tocalculate the P values. **P < 0.01.
Cho et al. PNAS | Published online March 16, 2015 | E1543
BIOCH
EMISTR
YPN
ASPL
US

38%, respectively, of untreated levels and that the reductionswere almost completely reversed by a PNRC2 down-regulation.Intriguingly, when MYC-PNRC2(R)-WT was expressed, effi-cient GMD of both C5′-RLuc mRNA and endogenous CCL2mRNA was restored. On the other hand, comparable expressionof MYC-PNRC2(R)-P101A/P104A, -W114A, or -P108A failed torestore GMD. These results suggest that the abilities of PNRC2 tointeract with GR and DCP1A are necessary for efficient GMD.
GMD Is Mechanistically Distinct from NMD and SMD. BecausePNRC2 and UPF1 are commonly involved in NMD and SMD(10, 11) as well as in GMD (present study), it is possible thatthere is a cross-talk between NMD/SMD and Dex-mediatedsignaling or Dex-induced GMD. To first determine whetherNMD and SMD are affected by Dex treatment, we assessed thechanges in expression of NMD and SMD factors and measuredthe efficiencies of NMD and SMD before and after Dex treat-ment (Fig. S5). The results revealed that the abundances of alltested NMD and SMD factors were not significantly changedafter Dex treatment. In addition, NMD and SMD efficiencies
were not significantly affected by Dex treatment (see SI Resultsfor more details).Conversely, we also tested the effect of an NMD-specific
factor on GMD. To this end, UPF1 (a common factor for GMDand NMD), GR (a GMD-specific factor), and UPF2 and UPF3X(both of which are known NMD factors) were down-regulatedusing specific siRNA (Fig. 5 A and B). The results of RT-qPCRrevealed that the levels of all tested endogenous GMD sub-strates, but not of endogenous CCL5 mRNA, which lacks aGR-binding site (22), were drastically reduced upon Dex treat-ment and restored by a down-regulation of GR or UPF1, but notof UPF2 or UPF3X (Fig. 5 A and B).It is known that NMD and SMD are coupled to translation
(17, 26). Given that GMD shares common factors UPF1 andPNRC2 with NMD and SMD, it is likely that GMD is also de-pendent on translation. To test this possibility, we designedSL-C5′-RLuc and SL-RLuc-C5′ reporter constructs that harbora stem-loop (SL) structure with ΔG = −87.8 kcal/mol (Fig. 3A).It is known that an insertion of a stable SL structure (ΔG = −75kcal/mol) into the 5′UTR blocks ribosome scanning and causes
A
RLuc-5 -5BoxB
λN-HA
λN-HA-GFP
λN-HA-GR(V729I)
λN-HA-GR(∆LBD)
λN-HA-GR(WT)
RLuc
ATG TAA
5BoxBλN
GFPλN
GRλN
V729IGRλN
HA tag
λN
LBD
GR
C
**
λN-HA
Rel
ativ
e R
Luc
mR
NA
(%)
GR
GR
V72
9IG
R
LBD
GR
GR
V72
9IG
R
LBD
020406080
100120140
D siRNA
PNRC2MYC-PNRC2(R)
P101
A/P1
04A
WT
Con
trol
PN
RC
2
pCMV-MYC-PNRC2(R)
β-actinMYC-PNRC2(R)
25
15
Mr(kDa) W
114A
P108
A
Dex
E
020406080
100120140
**
Rel
ativ
e le
vel o
f mR
NA
(%)
**
**
Dex
siRNA Control
pCMV-MYC-PNRC2(R)
P101A/P104A
W114A P108A
PNRC2
******
C5 -RLucCCL2
****
B
Dex
λN-HA GFP GR GFP GR
Rel
ativ
e R
Luc
mR
NA
(%) Control PNRC2 UPF1 UPF2
020406080
100120140
***
Fig. 4. GMD requires a ligand-binding ability of GR and GR- and DCP1A-binding abilities of PNRC2. (A) A schematic diagram of (i) the tethering reporterconstruct, RLuc-5′-5BoxB, which contains five tandem repeats of 5BoxB in the 5′UTR, and (ii) the effector construct, which expresses a C-terminally HA-taggedbacteriophage λN polypeptide (λN-HA), λN-HA-GFP, λN-HA-GR, or λN-HA-GR variants. (B) Analysis of rapid mRNA degradation elicited by GR artificiallytethered to the 5′UTR. HeLa cells were transiently transfected with the indicated siRNAs and, 1 d later, transfected with a tethering reporter plasmid, aneffector plasmid, and pCI-F, which encodes FLuc cDNA and serves as a control for variation in transfection. Two days later, the cells were either treated or nottreated with Dex for 30 min before harvesting. Specific down-regulation of PNRC2, UPF1, and UPF2 (Fig. S4A), and comparable expressions of λN-HA-GFP andλN-HA-GR (Fig. S4B) were validated by Western blotting. The levels of RLuc-5′-5BoxB mRNAs were normalized to the levels of FLuc mRNAs. Normalized levelsof RLuc-5′-5BoxB mRNAs in the presence of λN-HA were set to 100%. (C) Effects of GR variants on mRNA degradation. Comparable expression of GR variantswere determined by Western blotting (Fig. S4C). (D and E) Complementation experiments using siRNA-resistant PNRC2 variants. HEK293T cells were tran-siently transfected with either PNRC2 siRNA or a nonspecific control siRNA. One day after the transfection, the cells were cotransfected with three plasmids:(i) a plasmid expressing siRNA-resistant MYC-PNRC2(R) either WT or its variant (Fig. S4D), (ii) a GMD reporter plasmid expressing C5′-RLuc mRNA, and(iii) a reference plasmid, pCI-F. Two days later, the cells were either treated or not treated with Dex for 12 h before harvesting. (D) Western blottingof endogenous PNRC2 and MYC-PNRC2(R) with either WT or its variants. (E) RT-qPCR analysis of C5′-RLuc mRNA and endogenous CCL2 mRNA. The levelsof C5′-RLuc mRNA and endogenous CCL2 mRNA were normalized to the levels of FLuc mRNAs and GAPDH mRNAs, respectively. The normalized levels ofC5′-RLuc mRNA and endogenous CCL2 mRNA in the cells not treated with Dex were arbitrarily set to 100%. The columns and bars represent the mean and SDof three independent biological replicates. **P < 0.01, *P < 0.05.
E1544 | www.pnas.org/cgi/doi/10.1073/pnas.1409612112 Cho et al.

drastic inhibition of translation (27–29). Consistent with previousstudies, translation efficiencies of SL-C5′-RLuc mRNA andSL-RLuc-C5′ mRNA were drastically inhibited compared withtranslation efficiencies of C5′-RLuc mRNA and RLuc-C5′mRNA(Fig. 5C). Nevertheless, the GMD efficiencies of SL-C5′-RLucmRNA and SL-RLuc-C5′ mRNA were comparable to those ofC5′-RLuc mRNA and RLuc-C5′ mRNA, respectively (Fig. 5D).It should be noted that when the same SL structure is insertedinto the 5′UTR of NMD reporter mRNA, NMD and translationof the reporter mRNA are inhibited by ∼3- and 10-fold, re-spectively (29). These unexpected results suggest that GMD isindependent of a translation event. All of the data shown inFig. 5 indicate that although all three pathways share commonfactors UPF1 and PNRC2, GMD is mechanistically distinct fromNMD and SMD.
GR Bound to a Target mRNA Recruits PNRC2 to the Target mRNA ina Ligand-Dependent Manner to Elicit GMD. We next characterizedthe molecular hierarchy involved in the recruitment of GMDfactors to mRNA. We performed IPs using antibody againsta GMD factor and the extracts of cells either depleted or notdepleted of GR, PNRC2, or UPF1 (Fig. 6 and Fig. S6). Afterthat, the levels of coimmunoprecipitated endogenous GMDsubstrates were analyzed using RT-qPCR. Specific IPs and se-lective down-regulations by siRNAs were demonstrated by
Western blotting (Fig. S6). The results showed that in IP of GR,endogenous GMD substrates (CCL2 mRNA, PHLDA1 mRNA,and BCL3 mRNA) were enriched approximately twofold ina way that was unaffected by Dex treatment (Fig. 6A), indicatingthat GR is loaded onto GMD substrates independently of a li-gand. On the other hand, lower amounts of the endogenousGMD substrates coimmunopurified with either MYC-PNRC2 orFLAG-UPF1 relative to GR, when the cells were not treatedwith Dex (Fig. 6A). Dex treatment, however, significantly in-creased the amounts of the coimmunopurified GMD substratesin the IPs of MYC-PNRC2 or FLAG-UPF1 (Fig. 6A). Theenrichments of GMD substrates in the IPs of FLAG-UPF1 andMYC-PNRC2 were significantly reversed by down-regulation ofGR or PNRC2 (Fig. 6B) and by down-regulation of GR or UPF1(Fig. 6C), respectively. Consistent with a recent report showingthat UPF1 promiscuously binds to mRNA (30), the CCL5 mRNAand N-acetyltransferase 9 (NAT9) mRNA, both of which are nottargeted for GMD, were also enriched in the IP of FLAG-UPF1,but not in the IP of GR or MYC-PNRC2 (Fig. 6A). However, thisenrichment was not affected by Dex treatment. All data suggest thatligand-free GR loaded onto a GR-binding site within mRNA bindsto a ligand and recruits PNRC2 and UPF1 to the mRNA.
UPF1 Facilitates the Interaction Between GR and PNRC2. With re-gard to mRNA degradation, the recruitment of PNRC2 to theGR-binding site via GR may be sufficient for mRNA degrada-tion because PNRC2 interacts directly with DCP1A (11) andstabilizes the interaction between DCP1A and DCP2 (12). If so,why does GMD require UPF1? To address this question, weperformed IPs using extracts of the cells either depleted or notdepleted of UPF1 (Fig. 6D). The cells were pretreated with Dexbefore IP to induce efficient formation of the GR-containingcomplex. The results of IP using α-GR antibody showed thatMYC-PNRC2 coimmunopurified with GR in 10-fold smalleramounts when UPF1 was down-regulated (Fig. 6E). The reciprocalIP using MYC-PNRC2 also showed decreased association betweenGR and MYC-PNRC2 after UPF1 down-regulation (Fig. S6 J andK). All of these results indicate that UPF1 promotes an interactionbetween GR and PNRC2, which leads to more efficient GMD.
Helicase Activity of UPF1, but Not Phosphorylation of UPF1, IsRequired for Efficient GMD. The above findings that UPF1 facili-tates the interaction between GR and PNRC2 led us to examinewhich abilities of UPF1 are involved in GMD. First, to test a roleof helicase activity of UPF1 in GMD, we performed the com-plementation experiments using UPF1 siRNA and siRNA-resistantMYC-UPF1-WT or -R843C, which lacks helicase activity (31, 32),under the modified conditions (Fig. 7 A and B). In Fig. 2, weobserved that the half-lives of endogenous CCL2 and CCL7mRNAs were increased to almost normal levels upon UPF1down-regulation. On the other hand, steady-state levels of en-dogenous CCL2 and CCL7 mRNAs were moderately increasedby UPF1 down-regulation (Fig. 5A). One critical differencebetween the half-life and steady-state experiments was thetreatment with a transcription inhibitor 5,6-dichloro-1-β-D-ribo-furanosylbenzimidazole (DRB) in the half-life experiment. Thesimple interpretation for the difference in the extent of GMDinhibition by UPF1 down-regulation is that Dex may be involvedin the transcription of the CCL2 gene, as well as GMD of CCL2mRNA. In support of this idea, down-regulation of UPF1 almostcompletely inhibited GMDs of artificially designed reportermRNAs, the transcription of which is driven by the CMV pro-moter (Figs. 3 C and E and 4B). Therefore, to minimize thepossible effect of Dex on transcription of CCL2 gene althoughGMD efficiency is reduced as well, the incubation time with Dexwas reduced to 1 h in complementation experiments.The results of complementation experiments using UPF1
siRNA and siRNA-resistant MYC-UPF1 revealed that Dex
B
A C
D
Fig. 5. GMD is mechanistically distinct from NMD. (A and B) Effects of NMDfactors on GMD. HeLa cells were transfected with the indicated siRNAs, andendogenous GMD substrates were analyzed using RT-qPCR. The levels ofendogenous GMD substrates were normalized to levels of endogenousGAPDHmRNA. The normalized levels of endogenous GMD substrates in Dex-untreated cells were arbitrarily set to 100%. Specific down-regulation bysiRNAs was demonstrated by Western blotting (Fig. S5 G and H). (C and D) Atest of translation independence of GMD. HeLa cells were transfected withthe indicated GMD reporter RLuc plasmid and the FLuc control plasmid. Twodays later, the cells were either treated or not treated with Dex for 12 hbefore harvesting. (C) Translation efficiency of GMD reporter RLuc mRNAs.The protein activities of RLuc translated from GMD reporter mRNAs werenormalized to the levels of the GMD reporter RLuc mRNAs. The normalizedactivities of RLuc translated from C5′-RLuc mRNA or RLuc-C5′ mRNA werearbitrarily set to 100%. (D) RT-qPCR analysis of GMD reporter RLuc mRNAs.The levels of RLuc mRNA were normalized to the levels of FLuc mRNAs. Thenormalized levels of RLucmRNA in Dex-untreated cells were arbitrarily set to100%. The columns and bars represent the mean and SD of three in-dependent biological replicates. **P < 0.01, *P < 0.05.
Cho et al. PNAS | Published online March 16, 2015 | E1545
BIOCH
EMISTR
YPN
ASPL
US

treatment for 1 h reduced the level of endogenous CCL2 mRNAto ∼50% of the untreated level, and that down-regulation ofUPF1 significantly inhibited the GMD of CCL2mRNA (Fig. 7 Aand B). Such inhibition was significantly reversed by expressionof UPF1-WT but not of UPF1-R843C, suggesting that helicaseactivity of UPF1 is involved in efficient GMD.Next, we asked if UPF1 phosphorylation contributes to GMD
(Fig. 7 C–E). In the cases of NMD and SMD, it is well knownthat the phosphorylation of UPF1 by SMG1 kinase is critical fortarget mRNA degradation (33–36). Consistent with previousreports, down-regulation of SMG1 significantly inhibited theNMDs of endogenous NAT9 and SC35 (1.6 kb) mRNAs. Onthe other hand, down-regulation of SMG1 did not signifi-cantly affect the GMDs of endogenous CCL2 and CCL7mRNAs, suggesting that GMD occurs independently of UPF1phosphorylation. In contrast, down-regulation of GR selec-tively inhibited GMD but not NMD. All these results strengthenour conclusion that GMD is mechanistically distinct from NMDand SMD (Fig. 5).
GMD Controls Chemotaxis of Monocytes by Regulating CCL2 mRNAStability. It is known that the CCL2 protein belongs to the CCchemokine family and is involved in chemotaxis of various im-mune cells toward a site of injury or infection (37). Because wefound that GMD of CCL2 mRNA is dependent on PNRC2,UPF1, and GR in this study, we tested whether CCL2-mediatedchemotaxis is influenced by PNRC2, UPF1, and GR. Accord-ingly, we first measured the protein levels of secreted CCL2using an ELISA. Dex treatment reduced the protein levels ofsecreted CCL2 to ∼50% of the untreated control (Fig. 8B and Fig.S7B). The reduced protein levels were restored when PNRC2,UPF1, and GR were down-regulated using specific siRNAs to16%, 17%, and 9% of normal levels, respectively (Fig. 8A and Fig.S7A). On the other hand, the reduced protein level of CCL2 wasnot restored by the UPF2 down-regulation (Fig. S7B).We next assessed the effects of the down-regulation of
PNRC2, UPF1, or GR on cell migration in a chemotaxis mi-crochamber that had two compartments separated by a poly-vinylpyrrolidone-treated membrane with 5-μm pores. The lowercompartments of wells were filled with a supernatant (medium)from cells that were either treated or not treated with Dex, andwere either depleted or not depleted of PNRC2, UPF1, or GR.Human acute monocytic leukemia cell line THP-1 (expressingCCL2 receptors on the plasma membrane) was seeded in theupper compartments of wells. When THP-1 cells sense CCL2that diffuses from the lower compartment, they are expected tomigrate toward the lower compartment and to consequentlyadhere to the membrane. The number of migrating cells (cellsadherent to the membrane) was counted to determine thechemotactic index. The results revealed that cell migration wasdecreased by 50–60% when the lower compartments of wellscontained supernatants from the cells treated with Dex (Fig.8 C and D and Fig. S7 C and D). Intriguingly, the rate of cellmigration was restored by a down-regulation of PNRC2,UPF1, or GR, but not by UPF2 (Fig. 8 C and D and Fig. S7 Cand D). These results suggest that GMD plays an importantrole in the chemotaxis of monocytes via regulation of CCL2mRNA stability.
DiscussionIn this study, we demonstrate a new mRNA decay pathway,GMD, which is induced by a ligand (glucocorticoid) and requiresGR, PNRC2, UPF1, and DCP1A. According to our findings, wepropose the following molecular details of GMD. GR can bindto a GR-binding site of a target mRNA even in the absence ofa ligand (Fig. 8E, Left). In the presence of a ligand (Fig. 8E,Right), on the other hand, the ligand binds to GR, which ispreloaded onto a target mRNA, and the resulting ligand-bound
A
C
B *
Rel
ativ
e en
richm
ent o
fC
CL2
mR
NA
IP
siRNA Control GR PNRC2
*
0
1
2
3
4
5
**
siRNA
Rel
ativ
e en
richm
ent o
fC
CL2
mR
NA
0
1
2
3
4
5
0
1
2
3
4
5
IP
Control GR Control UPF1
**
D
Con
trol
UP
F1
UPF1
siRNA
β-actin
E
Con
trol
UP
F1m
IgG
α-G
Rm
IgG
α-G
R
Con
trol
UP
F1 siRNA
IP
GR
Before IP After IP
Dex
1 0.1MYC-PNRC2
β-actin
****
Rel
ativ
e en
richm
ent o
fC
CL2
mR
NA
0
1
2
3
4
5
6
Rel
ativ
e en
richm
ent o
fB
CL3
mR
NA
0
1
2
3
4**
*
Rel
ativ
e en
richm
ent o
fC
CL5
mR
NA
0
2
4
6
8
10
12
Dex
Rel
ativ
e en
richm
ent o
fN
AT9
mR
NA
0
1
2
3
4
5
6
IP
Rel
ativ
e en
richm
ent o
fP
HLD
A1
mR
NA
0
1
2
3
4
5*
*
Fig. 6. GR bound to mRNA recruits PNRC2 and UPF1 to the target mRNAand the GR-PNRC2 association is stabilized by UPF1. (A) RT-qPCR of coim-munoprecipitated endogenous GMD substrates in IPs of endogenous GR,MYC-PNRC2, and FLAG-UPF1. HeLa Cells were either transfected or nottransfected with plasmid expressing MYC-PNRC2 or FLAG-UPF1. Two dayslater, the cells were either treated or not treated with Dex for 3 h. IP ex-periments were performed using α-GR, α-MYC, α-FLAG antibody, or a non-specific mouse IgG (mIgG, control). IP specificity was demonstrated byWestern blotting (Fig. S6 A–C). The levels of coimmunoprecipitated GMDsubstrates relative to GAPDH mRNA were normalized to the levels of theinput amount (GMD substrates relative to GAPDH mRNA before IP). Thenormalized levels of GMD substrates obtained in IPs using mIgG without Dextreatment were arbitrarily set to 1.0. (B) RT-qPCR of coimmunoprecipitatedendogenous CCL2mRNA in IPs of FLAG-UPF1. As performed in A, except thatHeLa cells were depleted or not depleted of GR or PNRC2. The cells weretreated with Dex for 3 h before IP. Specific down-regulation (Fig. S6D) and IPspecificity (Fig. S6E) were demonstrated by Western blotting. (C) RT-qPCR ofcoimmunoprecipitated endogenous CCL2 mRNA in IPs of MYC-PNRC2. Asperformed in A, except that HeLa cells were depleted or not depleted of GRor UPF1. The cells were treated with Dex for 3 h before IP. Specific down-regulation (Fig. S6 F and H) and IP specificity (Fig. S6 G and I) were dem-onstrated by Western blotting. (D and E) IP of endogenous GR using theextracts of cells depleted or not depleted of UPF1. HeLa cells, either depletedor not depleted of UPF1, were transiently transfected with the plasmidexpressing MYC-PNRC2. The cells were treated with Dex for 3 h before IP.After that, the cell extracts were treated with RNase A. IPs were performedusing α-GR antibody or mIgG. (D) A Western blot demonstrating specificdown-regulation of UPF1. (E) IPs using either an antibody against endoge-nous GR or nonspecific mIgG. The columns and bars represent the mean andSD of two or three independent biological replicates. **P < 0.01; *P < 0.05.
E1546 | www.pnas.org/cgi/doi/10.1073/pnas.1409612112 Cho et al.

GR would recruit PNRC2 and eventually DCP1A and UPF1,both of which directly interact with PNRC2 (11). The resultingcomplex may be displaced by the scanning ribosome. Nonethe-less, the complex may quickly reassociate with 5′UTR because ofthe intrinsic RNA-binding ability of GR (22). The recruitedUPF1 may transiently trigger a remodeling of messenger ribo-nucleoprotein particle (mRNP) via its helicase activity (Fig. 7 Aand B), consequently further stabilizing the GR-PNRC2 in-teraction (Fig. 6 D and E and Fig. S6 J and K). Then, theresulting complex at the GR-binding site would trigger decapp-ing followed by 5′-to-3′ degradation of the mRNA. The recruitedUPF1 may also repress translation initiation before mRNAdegradation, as observed in NMD (38).In the case of NMD, it has been reported that UPF1 helicase
activity is promoted by UPF2 (39–41) and that SMG1 providesa binding platform for UPF1 and UPF2, resulting in the acti-vation of UPF1 helicase activity (42). However, we observed that
UPF2 and SMG1 are not essential for efficient GMD (Figs. 4B,5B, and 7 C–E). Therefore, it is most likely that other cellularfactors rather than UPF2 may control the helicase ability ofUPF1 during GMD. In addition, another kinase rather thanSMG1 may act to activate UPF1 in GMD. Indeed, UPF1 is
A B
Rel
ativ
e le
vel o
fC
CL2
mR
NA
(%)
pCMV-MYC-UPF1(R)
R843C
Dex
ControlsiRNA UPF1
WT
**
020406080
100120140
***siRNA
UPF1MYC-UPF1(R)
R84
3CW
T
Con
trol
UP
F1pCMV-MYC-UPF1(R)
β-actin
Con
trol
siRNA
SM
G1
GR
GR
SMG1
β-actin
C
020406080
100120140
Rel
ativ
e le
vel o
f mR
NA
(%)
Dex
siRNA Control GR SMG1
**** ****
CCL2 CCL7 CCL5D
0
200
400
600
800
Rel
ativ
e le
vel o
f mR
NA
(%)
siRNA
Con
trol
GR
SM
G1
NAT9SC35 1.6kb
**
**
E
Fig. 7. GMD requires a helicase activity but not phosphorylation of UPF1.(A and B) Complementation experiments using siRNA-resistant UPF1-WT andits R843C variant. HeLa cells were transiently transfected with either UPF1siRNA or a nonspecific control siRNA. One day after the transfection, the cellswere retransfected with a plasmid expressing siRNA-resistant MYC-UPF1(R)-WTor -R843C. Two days later, the cells were either treated or not treated withDex for 1 h before harvesting. (A) Western blots demonstrating selectivedown-regulation of UPF1 and comparable levels of exogenously expressedUPF1(R) and endogenous UPF1. (B) RT-qPCR analysis of endogenous CCL2mRNA. The levels of endogenous CCL2 mRNA were normalized to the levelsof endogenous GAPDH mRNAs. The normalized levels of endogenous CCL2mRNA in the cells not treated with Dex were arbitrarily set to 100%. (C–E)GMD is not dependent on SMG1 activity. HeLa cells were transiently trans-fected with GR siRNA, SMG1 siRNA, or a nonspecific control siRNA. Threedays later, the cells were either treated or not treated with Dex for 3 h be-fore harvesting. (C) Western blots demonstrating selective down-regula-tion of GR and SMG1 by siRNAs. (D) RT-qPCR analysis of endogenous CCL2,CCL7, and CCL5 mRNAs. The levels of endogenous GMD substrates werenormalized to the levels of endogenous GAPDH mRNAs. (E) RT-qPCR analysisof endogenous NAT9 and SC35 (1.6 kb) mRNAs. The columns and bars rep-resent the mean and SD of three independent biological replicates. **P <0.01; *P < 0.05.
PN
RC
2si
RN
AC
ontro
l siR
NA
UP
F1si
RN
A
Dex
THP-1 chemotaxisC
B
D
Che
mot
axis
inde
x(%
)
020406080
100120140160
siRNA
Dex
**
**
Rel
ativ
e le
vel o
fC
CL2
pro
tein
(%)
siRNA
Dex0
20406080
100120140160
**
**
A siRNA
Con
trol
UP
F1P
NR
C2
PNRC2
UPF1
β-actin
E
Fig. 8. Chemotaxis of monocytes is mediated by GMD. The lower and uppercompartments of each well in a chemotaxis microchamber were separatedon a polyvinylpyrrolidone-treated membrane with 5-μm pores. The lowercompartments of wells were filled with the supernatants of HeLa cells de-pleted of either PNRC2 or UPF1 and either not treated or treated with Dexfor 3 h. The upper compartments were filled with THP-1 cells (a monocytecell line). After incubation of the microchamber for 3 h at 37 °C, the mem-branes were removed from the wells. Then, the migrating cells that adheredto the underside of the membranes were stained and counted under a mi-croscope in six randomly selected visual fields in each well. (A) A Westernblot of PNRC2 and UPF1, demonstrating specific down-regulation by siRNAs.(B) ELISA for the CCL2 protein concentration in the supernatants of the HeLacells depleted of either PNRC2 or UPF1. The levels of CCL2 protein werenormalized to the levels of total protein in the supernatants. The normalizedlevels of CCL2 without Dex treatment were arbitrarily set to 100%.(C) Analysis of chemotaxis of THP-1 cells. The migrating cells were stainedand images were acquired to demonstrate the effects of either the PNRC2 orUPF1 down-regulation on cell migration. (Scale bars, 20 μm.) (D) The che-motactic index. The stained cells were counted under a microscope in sixrandomly selected visual fields in each well to determine the chemotacticindex. The numbers of stained cells without Dex treatment in each visualfield were arbitrarily set to 100%. The columns and bars represent the meanand SD of at least three independent transfection experiments and ELISA orchemotaxis assay. **P < 0.01. (E) A model of GMD-mediated chemotaxis. Inthe absence of a glucocorticoid, GMD is inactive and its target mRNA (CCL2mRNA) is stable and abundant. Consequently, the expressed CCL2 proteininduces chemoattraction of monocytes (Left). In the presence of a glucocor-ticoid, GMD is activated, destabilizes CCL2 mRNA, and thereby causes in-efficient chemoattraction of monocytes (Right). The details are describedin Discussion.
Cho et al. PNAS | Published online March 16, 2015 | E1547
BIOCH
EMISTR
YPN
ASPL
US

known to be phosphorylated by ATR and DNA-PK during HMD(43, 44).Recently, genome-wide analysis showed that UPF1 associates
with mRNAs throughout the entire sequences and is displacedfrom mRNA by an elongating ribosome (30). Therefore, it ispossible that the ligand-bound GR-PNRC2 complex bound tothe GR-binding site of mRNA recruits either a preexisting UPF1within close proximity of the GR-binding site or mRNA-freeUPF1. Our data demonstrate that the latter mechanism is pre-dominant because the amount of coimmunopurified GMDsubstrate in the IP of UPF1 is significantly increased by Dextreatment (Fig. 6A). The resulting complex elicits efficientGMD, possibly in the 5′-to-3′ direction because of the en-hancement of decapping by DCP1A. The possible 3′-to-5′ deg-radation of a GMD substrate remains to be tested. Conse-quently, GMD substrates would be down-regulated, affectingvarious physiological and metabolic pathways, such as chemo-taxis of monocytes (Fig. 8).Other groups as well as ours have shown that the C terminus
of PNRC2 commonly interacts with GR, DCP1A, and UPF1,although the critical residues in the C terminus involved in theinteractions between PNRC2 and its binding partners are char-acterized (8, 10, 11, 23). Because of the common binding site,once one or two binding partners interact with PNRC2, thethird binding partner may not be easily recruited to thecomplex because of steric hindrance. In the present study,however, we showed that PNRC2 is complexed with DCP1A,UPF1, and GR. Therefore, one possible explanation is thatthe steric hindrance would be overcome by PNRC2 di-merization, which may provide a sufficient binding platformfor loading of its binding partners. In support of this idea, it hasrecently been reported that PNRC1, an isoform of PNRC2,forms a dimer (45).It is generally considered that cytosolic GR enters the nucleus
when it binds to a ligand. However, we observed that althoughmost of the cytosolic GR moved to the nuclear fraction uponDex treatment, a significant amount of GR remained in the cy-toplasmic fractions (Fig. S8). Moreover, the intracellular distri-bution of PNRC2 was only marginally affected by Dex treatment.Therefore, the GR and PNRC2 remaining in the cytoplasm uponDex treatment might be sufficient for eliciting GMD.Here, we also demonstrate that GMD is mechanistically dis-
tinct from other UPF1-dependent mRNA decay pathways—NMD, SMD, and HMD—although all these pathways sharesome common factors: UPF1, PNRC2, and DCP1A. One majordifference is the method of recruitment of UPF1 to the workingsite on mRNA and the method of stabilization of the UPF1-containing complex. In the case of NMD, either mRNA-unboundor prebound UPF1 is recruited to a terminating ribosome andis stabilized by an exon junction complex downstream of PTC.In the case of SMD and HMD, UPF1 is also recruited to aterminating ribosome during translation and is stabilized bydownstream STAU and SL binding protein, respectively. Alter-natively, UPF1 could be recruited to mRNA via direct in-teraction with STAU or SL binding protein and then could jointhe termination complex. Therefore, NMD, SMD, and HMD aretightly coupled to translation events, although the exact timing ofUPF1 recruitment is not clear at this point. In the case of GMD,however, UPF1 recruitment to the working site on mRNA occursindependently of translation (Fig. 5 C and D). Instead, the UPF1recruitment is mediated by its direct interaction with PNRC2,which bridges mRNA-bound GR and UPF1 in a Dex-dependentmanner. In addition, GMD is not affected by down-regulationof known NMD-specific factors (Fig. 5). Therefore, GMDis a previously unidentified translation-independent and UPF1-dependent mRNA decay pathway.Another novel feature of GMD is that the presence, rather
than a relative position, of the GR-binding site within mRNA is
crucial for eliciting GMD after Dex treatment. Nonetheless, allGMD substrates tested in this study were shown to containa functional GR-binding site in the 5′UTR (Fig. 3). Althoughmore GMD substrates should be tested, it seems that there is anevolutionary selective pressure to maintain the GR-binding sitein the 5′UTR. This observation points to the possibility of anadditional regulatory mechanism of gene expression through theGR-binding site, aside from GMD. A genome-wide survey shouldhelp to uncover a number of additional natural GMD substratesand will identify further molecular details and biological rele-vance of GMD.
Materials and MethodsConstruction of Plasmids. The details of plasmid construction are provided inSI Materials and Methods.
Cell Culture, Transfection, and the siRNA Sequences. The details of cellculture, transfection, and the siRNA sequences are provided in SI Materialsand Methods.
The Tethering Assay. HeLa cells were transiently cotransfected with 0.1 μgof pRL-5′-5BoxB, 0.2 μg of an effector plasmid, and 0.01 μg of pCI-F usingLipofectamine 2000 (Invitrogen). Two days later, the cells were treated with100 nM of Dex for 30 min. The cells were then harvested, and total cellRNA and protein were purified using TRIzol Reagent (Life Technologies).
Quantitative Real-Time RT-PCR and Semiquantitative RT-PCR Using α-[32P]-dATPand Specific Oligonucleotides. RT-qPCR analyses were performed as describedpreviously (10, 11, 46). The oligonucleotides used in our study are listed inDataset S2.
RT-PCR using α-[32P]-dATP and specific oligonucleotides was performed asdescribed previously (11, 17). The 32P-labeled PCR products were analyzedusing 5% polyacrylamide gel electrophoresis, visualized with Phosphor-Imaging (BAS-2500; Fuji Photo Film), and then quantitated using Multi-Gauge (Fuji Photo Film).
Immunoprecipitation. HeLa cells and HEK293T cells were transiently trans-fected with the indicated plasmids using Lipofectamine 2000 and the calciumphosphate precipitation method, respectively. The IPs and RNA IPs wereperformed as described previously (9, 11, 46).
Western Blotting. Antibodies against the following proteins or peptides wereused in this study: FLAG and β-actin (Sigma-Aldrich); MYC (Calbiochem), UPF1(a gift from Lynne E. Maquat, University of Rochester, Rochester, NY), PNRC2(11), and human STAU1 (47); DCP1A, UPF2, eIF4AIII, phospho-S1078-UPF1,and phospho-S1096-UPF1 (10); UPF3X (48), eIF4E (Cell Signaling Technology),CBP80 (49), CBP80/20-dependent translation initiation factor (CTIF; ref. 46),GR (BD Biosciences and Santa Cruz Biotechnology), snRNP70 (Santa Cruz),and SMG1 (Bethyl). The antibody against UPF1 phosphorylated at threonine28 was raised in rabbits using the synthetic peptide LGADpTQGSEF (AbClon).
Microarray Analysis. Microarray analysis was performed by Macrogen, asdescribed previously (9, 10, 50). The microarray data were deposited in theNational Center for Biotechnology Information Gene Expression Omnibusweb-based data repository (series ID: GSE49591).
CCL2 Measurements and Chemotaxis Assay. The details of CCL2 measurementsand the chemotaxis assay are provided in SI Materials and Methods.
Statistical Analysis. Two-tailed, equal-sample variance Student’s t-tests wereused to calculate the P-values. Differences with P < 0.05 were consideredstatistically significant.
ACKNOWLEDGMENTS. We thank Dr. Lynne E. Maquat for providing nonsense-mediated mRNA decay reporter plasmids and the α-UPF1 antibody; Jens Lykke-Andersen for pcDNA3-FLAG-DCP2; and Juan Ortín for α-human STAU1 antibody.This work was supported by the National Research Foundation of Korea grantfunded by the Korea government (MSIP) (2012R1A2A1A01002469 and2014R1A2A1A11050412); O.H.P. was in part supported by the Global PhDFellowship Program through the National Research Foundation funded bythe Korean Government.
E1548 | www.pnas.org/cgi/doi/10.1073/pnas.1409612112 Cho et al.

1. Vandevyver S, Dejager L, Libert C (2012) On the trail of the glucocorticoid receptor:Into the nucleus and back. Traffic 13(3):364–374.
2. Oakley RH, Cidlowski JA (2011) Cellular processing of the glucocorticoid receptorgene and protein: new mechanisms for generating tissue-specific actions of gluco-corticoids. J Biol Chem 286(5):3177–3184.
3. Santos GM, Fairall L, Schwabe JW (2011) Negative regulation by nuclear receptors: Aplethora of mechanisms. Trends Endocrinol Metab 22(3):87–93.
4. Lonard DM, O’Malley BW (2012) Nuclear receptor coregulators: Modulators of pa-thology and therapeutic targets. Nat Rev Endocrinol 8(10):598–604.
5. Kato S, Yokoyama A, Fujiki R (2011) Nuclear receptor coregulators merge transcrip-tional coregulation with epigenetic regulation. Trends Biochem Sci 36(5):272–281.
6. Heery DM, Kalkhoven E, Hoare S, Parker MG (1997) A signature motif in transcrip-tional co-activators mediates binding to nuclear receptors. Nature 387(6634):733–736.
7. Zhou D, et al. (2000) PNRC: A proline-rich nuclear receptor coregulatory protein thatmodulates transcriptional activation of multiple nuclear receptors including orphanreceptors SF1 (steroidogenic factor 1) and ERRalpha1 (estrogen related receptoralpha-1). Mol Endocrinol 14(7):986–998.
8. Zhou D, Chen S (2001) PNRC2 is a 16 kDa coactivator that interacts with nuclear re-ceptors through an SH3-binding motif. Nucleic Acids Res 29(19):3939–3948.
9. Cho H, et al. (2013) SMG5-PNRC2 is functionally dominant compared with SMG5-SMG7 in mammalian nonsense-mediated mRNA decay. Nucleic Acids Res 41(2):1319–1328.
10. Cho H, et al. (2012) Staufen1-mediated mRNA decay functions in adipogenesis. MolCell 46(4):495–506.
11. Cho H, Kim KM, Kim YK (2009) Human proline-rich nuclear receptor coregulatoryprotein 2 mediates an interaction between mRNA surveillance machinery and de-capping complex. Mol Cell 33(1):75–86.
12. Lai T, et al. (2012) Structural basis of the PNRC2-mediated link between mrna sur-veillance and decapping. Structure 20(12):2025–2037.
13. Choe J, Ahn SH, Kim YK (2014) The mRNP remodeling mediated by UPF1 promotesrapid degradation of replication-dependent histone mRNA. Nucleic Acids Res 42(14):9334–9349.
14. Karam R, Wengrod J, Gardner LB, Wilkinson MF (2013) Regulation of nonsense-mediated mRNA decay: implications for physiology and disease. Biochim Biophys Acta1829(6-7):624–633.
15. Schoenberg DR, Maquat LE (2012) Regulation of cytoplasmic mRNA decay. Nat RevGenet 13(4):246–259.
16. Schweingruber C, Rufener SC, Zünd D, Yamashita A, Mühlemann O (2013) Nonsense-mediated mRNA decay—Mechanisms of substrate mRNA recognition and degrada-tion in mammalian cells. Biochim Biophys Acta 1829(6-7):612–623.
17. Kim YK, Furic L, Desgroseillers L, Maquat LE (2005) Mammalian Staufen1 recruits Upf1to specific mRNA 3’UTRs so as to elicit mRNA decay. Cell 120(2):195–208.
18. Park E, Maquat LE (2013) Staufen-mediated mRNA decay. Wiley Interdiscip Rev RNA4(4):423–435.
19. Gong C, Maquat LE (2011) lncRNAs transactivate STAU1-mediated mRNA decay byduplexing with 3′ UTRs via Alu elements. Nature 470(7333):284–288.
20. Marzluff WF, Wagner EJ, Duronio RJ (2008) Metabolism and regulation of canonicalhistone mRNAs: Life without a poly(A) tail. Nat Rev Genet 9(11):843–854.
21. Dhawan L, Liu B, Blaxall BC, Taubman MB (2007) A novel role for the glucocorticoidreceptor in the regulation of monocyte chemoattractant protein-1 mRNA stability.J Biol Chem 282(14):10146–10152.
22. Ishmael FT, et al. (2011) The human glucocorticoid receptor as an RNA-binding pro-tein: Global analysis of glucocorticoid receptor-associated transcripts and identifica-tion of a target RNA motif. J Immunol 186(2):1189–1198.
23. Zhou D, Ye JJ, Li Y, Lui K, Chen S (2006) The molecular basis of the interaction be-tween the proline-rich SH3-binding motif of PNRC and estrogen receptor alpha.Nucleic Acids Res 34(20):5974–5986.
24. Schäcke H, Döcke WD, Asadullah K (2002) Mechanisms involved in the side effects ofglucocorticoids. Pharmacol Ther 96(1):23–43.
25. Malchoff DM, et al. (1993) A mutation of the glucocorticoid receptor in primarycortisol resistance. J Clin Invest 91(5):1918–1925.
26. Ishigaki Y, Li X, Serin G, Maquat LE (2001) Evidence for a pioneer round of mRNAtranslation: mRNAs subject to nonsense-mediated decay in mammalian cells arebound by CBP80 and CBP20. Cell 106(5):607–617.
27. Kozak M (1986) Influences of mRNA secondary structure on initiation by eukaryoticribosomes. Proc Natl Acad Sci USA 83(9):2850–2854.
28. Vassilenko KS, Alekhina OM, Dmitriev SE, Shatsky IN, Spirin AS (2011) Unidirectionalconstant rate motion of the ribosomal scanning particle during eukaryotic translationinitiation. Nucleic Acids Res 39(13):5555–5567.
29. Choe J, et al. (2014) eIF4AIII enhances translation of nuclear cap-binding complex-bound mRNAs by promoting disruption of secondary structures in 5’UTR. Proc NatlAcad Sci USA 111(43):E4577–E4586.
30. Zünd D, Gruber AR, Zavolan M, Mühlemann O (2013) Translation-dependent dis-placement of UPF1 from coding sequences causes its enrichment in 3′ UTRs. Nat StructMol Biol 20(8):936–943.
31. Sun X, Perlick HA, Dietz HC, Maquat LE (1998) A mutated human homologue to yeastUpf1 protein has a dominant-negative effect on the decay of nonsense-containingmRNAs in mammalian cells. Proc Natl Acad Sci USA 95(17):10009–10014.
32. Kurosaki T, et al. (2014) A post-translational regulatory switch on UPF1 controls tar-geted mRNA degradation. Genes Dev 28(17):1900–1916.
33. Ohnishi T, et al. (2003) Phosphorylation of hUPF1 induces formation of mRNA sur-veillance complexes containing hSMG-5 and hSMG-7. Mol Cell 12(5):1187–1200.
34. Yamashita A, Kashima I, Ohno S (2005) The role of SMG-1 in nonsense-mediatedmRNA decay. Biochim Biophys Acta 1754(1-2):305–315.
35. Yamashita A, Ohnishi T, Kashima I, Taya Y, Ohno S (2001) Human SMG-1, a novelphosphatidylinositol 3-kinase-related protein kinase, associates with components ofthe mRNA surveillance complex and is involved in the regulation of nonsense-medi-ated mRNA decay. Genes Dev 15(17):2215–2228.
36. Cho H, Han S, Park OH, Kim YK (2013) SMG1 regulates adipogenesis via targeting ofstaufen1-mediated mRNA decay. Biochim Biophys Acta 1829(12):1276–1287.
37. Myers SJ, Wong LM, Charo IF (1995) Signal transduction and ligand specificity of thehuman monocyte chemoattractant protein-1 receptor in transfected embryonic kid-ney cells. J Biol Chem 270(11):5786–5792.
38. Isken O, et al. (2008) Upf1 phosphorylation triggers translational repression duringnonsense-mediated mRNA decay. Cell 133(2):314–327.
39. Clerici M, et al. (2009) Unusual bipartite mode of interaction between the nonsense-mediated decay factors, UPF1 and UPF2. EMBO J 28(15):2293–2306.
40. Chamieh H, Ballut L, Bonneau F, Le Hir H (2008) NMD factors UPF2 and UPF3 bridgeUPF1 to the exon junction complex and stimulate its RNA helicase activity. Nat StructMol Biol 15(1):85–93.
41. Chakrabarti S, et al. (2011) Molecular mechanisms for the RNA-dependent ATPaseactivity of Upf1 and its regulation by Upf2. Mol Cell 41(6):693–703.
42. Melero R, et al. (2014) Structures of SMG1-UPFs complexes: SMG1 contributes toregulate UPF2-dependent activation of UPF1 in NMD. Structure 22(8):1105–1119.
43. Kaygun H, Marzluff WF (2005) Regulated degradation of replication-dependenthistone mRNAs requires both ATR and Upf1. Nat Struct Mol Biol 12(9):794–800.
44. Müller B, Blackburn J, Feijoo C, Zhao X, Smythe C (2007) DNA-activated protein kinasefunctions in a newly observed S phase checkpoint that links histone mRNA abundancewith DNA replication. J Cell Biol 179(7):1385–1398.
45. Zhou D, et al. (2007) PNRC is a unique nuclear receptor coactivator that stimulatesRNA polymerase III-dependent transcription. J Mol Signal 2:5.
46. Kim KM, et al. (2009) A new MIF4G domain-containing protein, CTIF, directs nuclearcap-binding protein CBP80/20-dependent translation. Genes Dev 23(17):2033–2045.
47. Marión RM, Fortes P, Beloso A, Dotti C, Ortín J (1999) A human sequence homologueof Staufen is an RNA-binding protein that is associated with polysomes and localizesto the rough endoplasmic reticulum. Mol Cell Biol 19(3):2212–2219.
48. Kim KM, Cho H, Kim YK (2012) The upstream open reading frame of cyclin-dependentkinase inhibitor 1A mRNA negatively regulates translation of the downstream mainopen reading frame. Biochem Biophys Res Commun 424(3):469–475.
49. Choe J, et al. (2012) Translation initiation on mRNAs bound by nuclear cap-bindingprotein complex CBP80/20 requires interaction between CBP80/20-dependent trans-lation initiation factor and eukaryotic translation initiation factor 3g. J Biol Chem287(22):18500–18509.
50. Cho H, Ahn SH, Kim KM, Kim YK (2013) Non-structural protein 1 of influenza virusesinhibits rapid mRNA degradation mediated by double-stranded RNA-binding protein,staufen1. FEBS Lett 587(14):2118–2124.
Cho et al. PNAS | Published online March 16, 2015 | E1549
BIOCH
EMISTR
YPN
ASPL
US

![Glucocorticoid-induced Cell Death Requires …...[CANCER RESEARCH 59, 1378–1385, March 15, 1999] Glucocorticoid-induced Cell Death Requires Autoinduction of Glucocorticoid Receptor](https://img.pdfslide.net/doc/110x75/5e5646d0314f24389e233453/glucocorticoid-induced-cell-death-requires-cancer-research-59-1378a1385.jpg)