Embed Size (px)
Citation preview

Glycolysis II

Substrates for Gluconeogenesis
• Lactate, pyruvate• Carbon skeletons of amino
acids, except leucine and lysine
• Citric acid cycle intermediates• Anything that can be converted
to oxaloacetate
• Substrates that can’t contribute:– Acetyl CoA– Fatty acids – since fatty
acid breakdown generates acetyl CoA
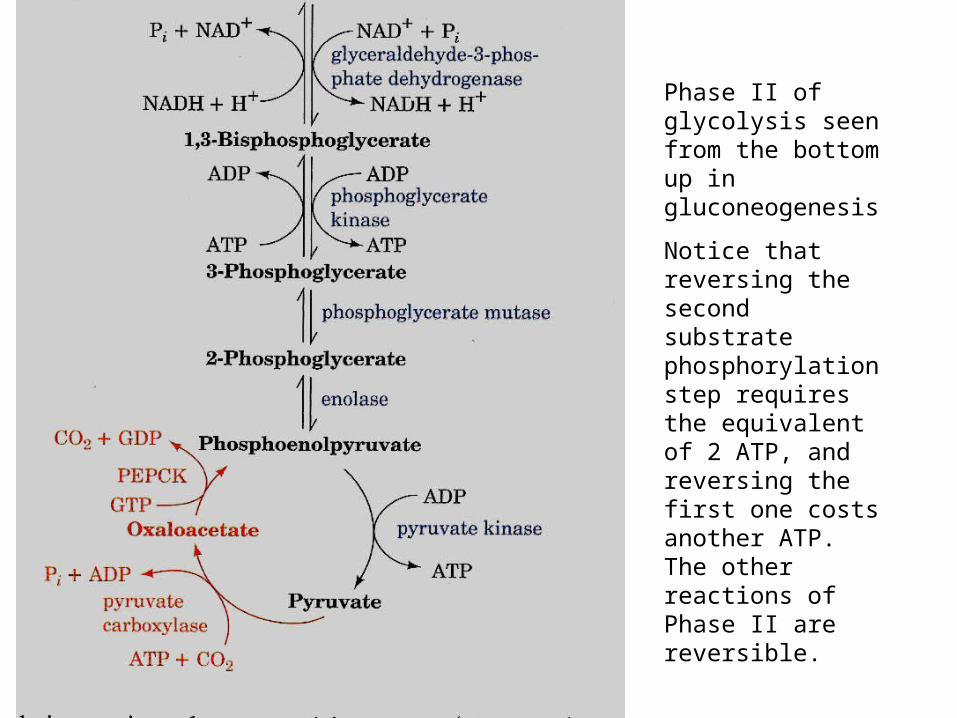
Phase II of glycolysis seen from the bottom up in gluconeogenesis
Notice that reversing the second substrate phosphorylation step requires the equivalent of 2 ATP, and reversing the first one costs another ATP. The other reactions of Phase II are reversible.
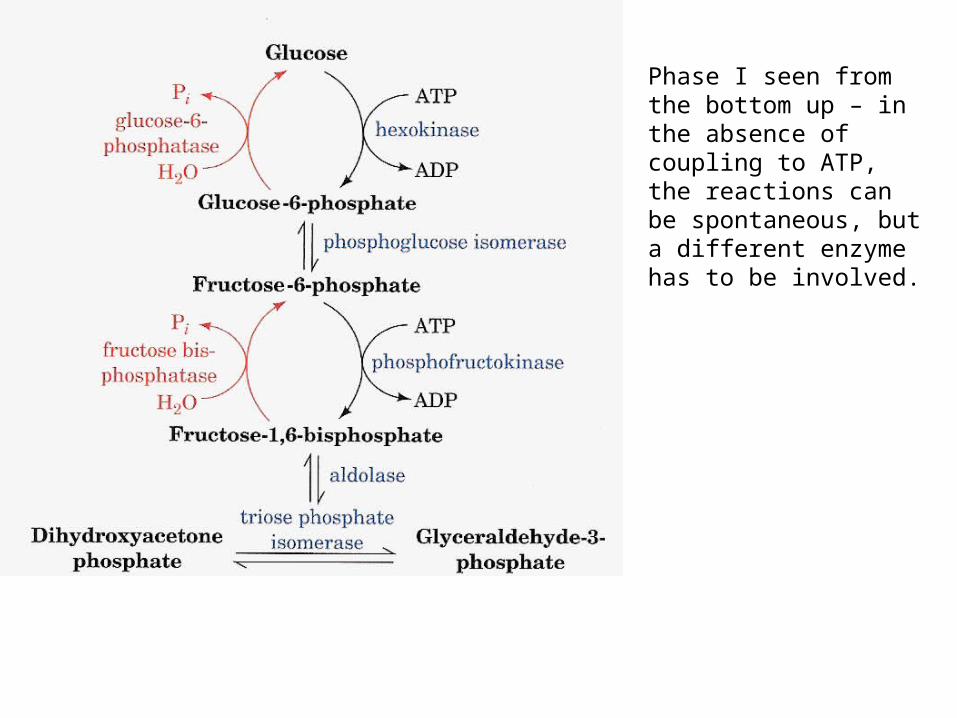
Phase I seen from the bottom up – in the absence of coupling to ATP, the reactions can be spontaneous, but a different enzyme has to be involved.

No futile cycles
• Obviously, the bypass routes of gluconeogenesis and the main route of glycolysis could not be allowed to operate at the same time, or futile cycles would result. This is prevented by the fact that most cells simply don’t express the gluconeogenic enzymes. For the cell types that do…
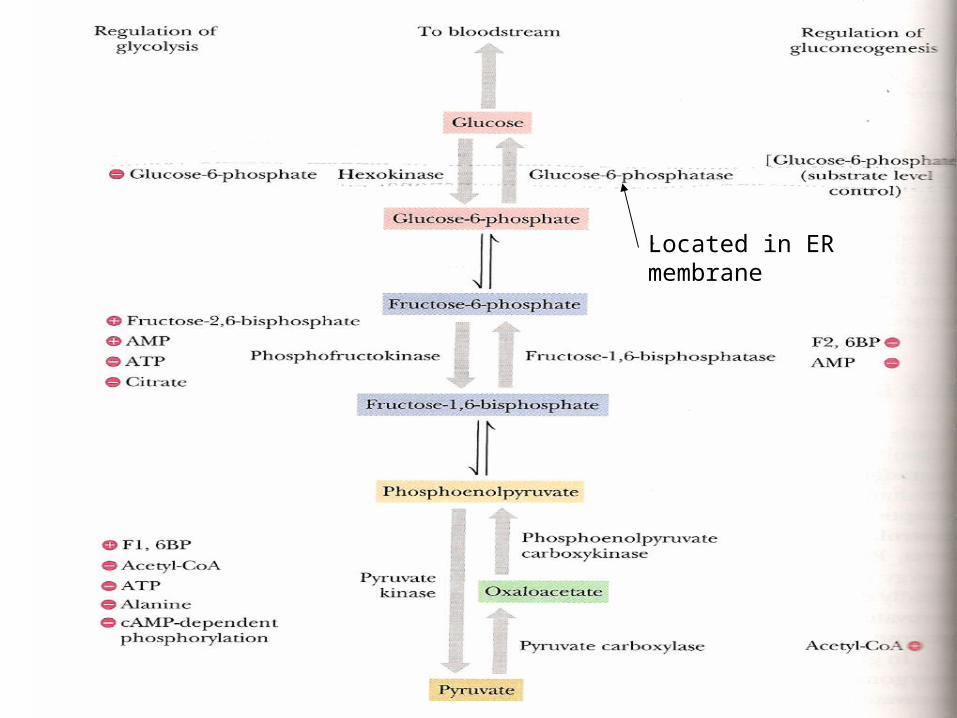
Located in ER membrane
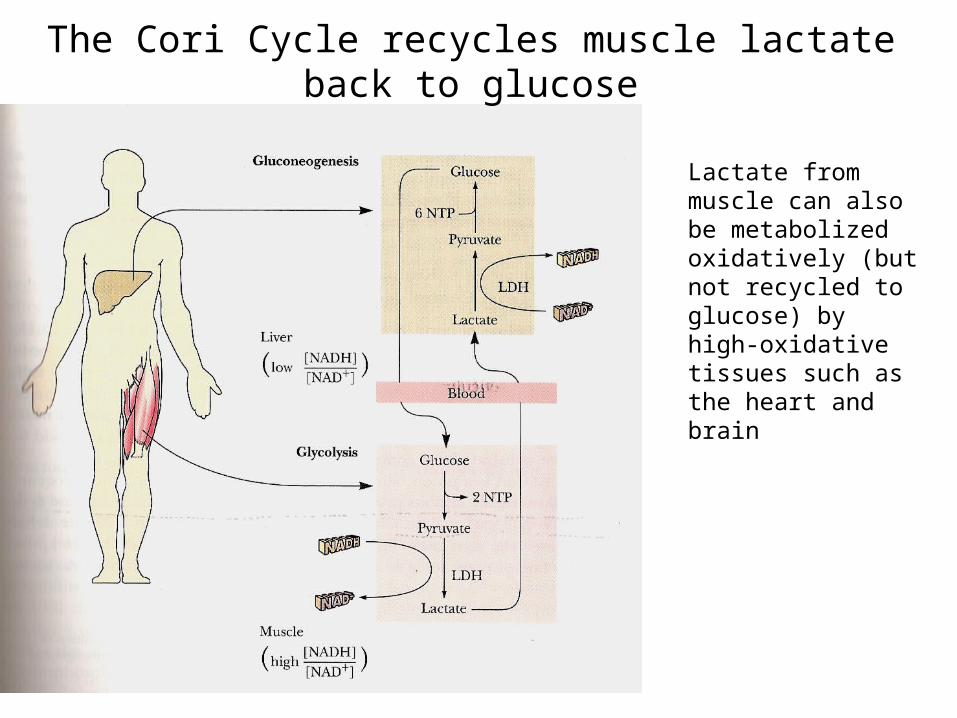
The Cori Cycle recycles muscle lactate back to glucose
Lactate from muscle can also be metabolized oxidatively (but not recycled to glucose) by high-oxidative tissues such as the heart and brain

To do all of this, we need more than one form of lactate dehydrogenase
• Heart and brain need a form that favors conversion of lactate to pyruvate
• Non-oxidative muscle needs the opposite – a form that can efficiently convert pyruvate to lactate
• Liver and kidney need an intermediate form, because they may play the game both ways
bbnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnngggggggggggggggggggggggggg

How to do this?
• LDH gene was duplicated and the two sister genes underwent separate evolution to become H (heart) type and M (muscle) types
• LDH enzyme is a tetramer that can be made up of some mixture of both H and M types. Combinatorial math gives us 5 different combos: M4H0, M3H1, M2H2, H3M1, H4M0 with a corresponding range of kinetic properties.

The solution
• So, non-oxidative skeletal muscle expresses only the M gene
• Heart and brain express only the H gene
• Kidney and liver (and other tissues) express some mix of H and M genes

Other sources of Acetyl CoA

Beta oxidation of fatty acids
• Each FA molecule undergoes an initial activation step to become a fatty acyl-CoA – this is energized by hydrolysis of an ATP to AMP
• After activation, a repeating cycle of 4 reactions splits off acetyl Co-A until the end of the fatty acid is reached
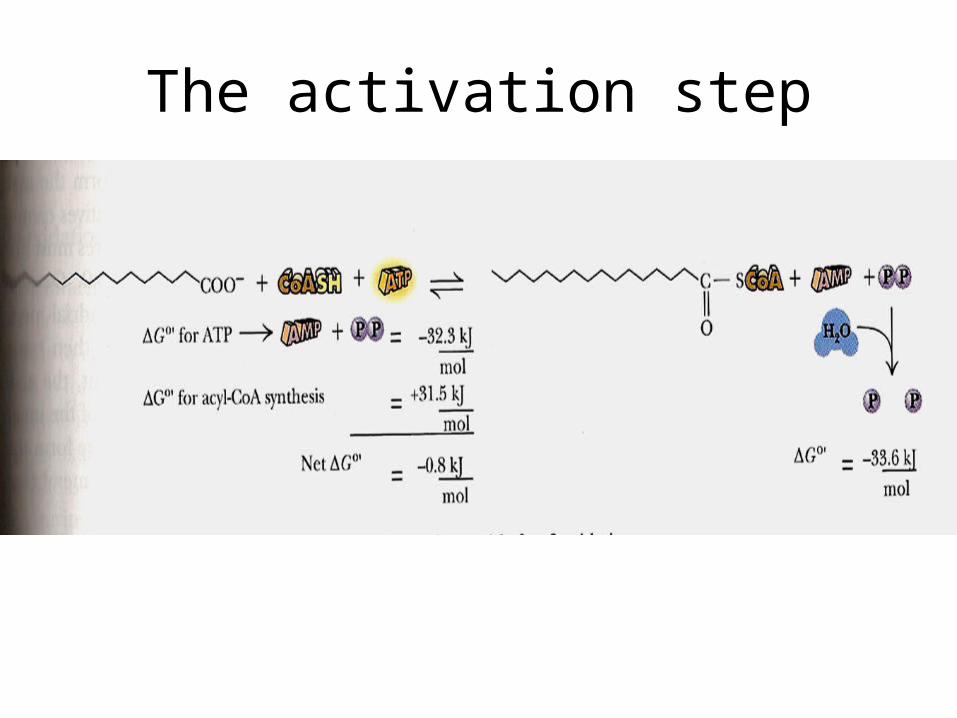
The activation step
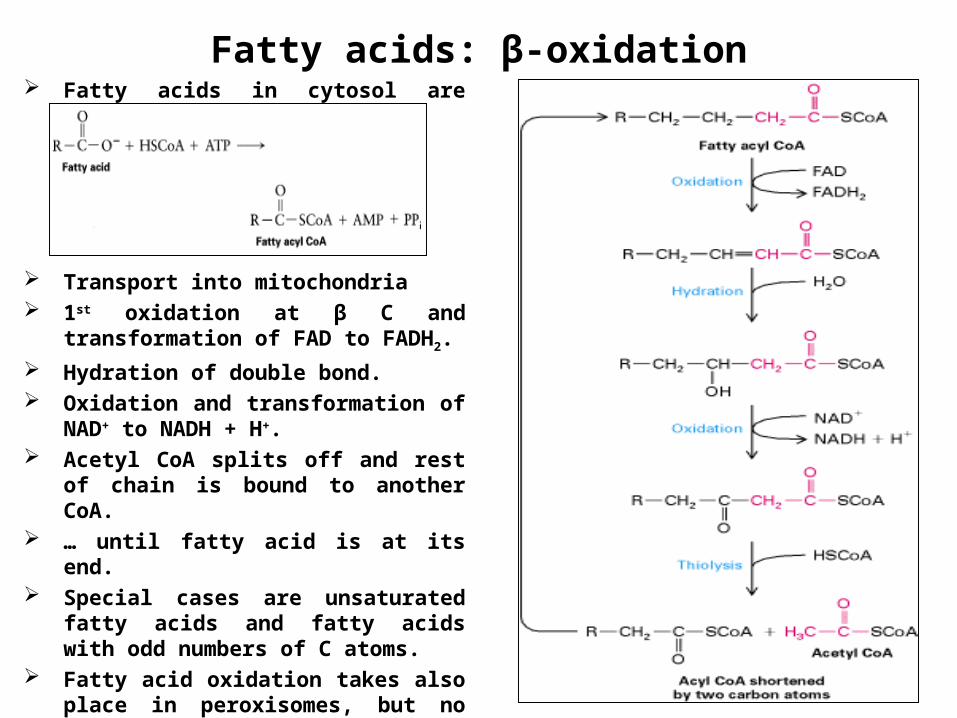
Fatty acids: β-oxidation Fatty acids in cytosol are bound to
CoA.
Transport into mitochondria 1st oxidation at β C and transformation
of FAD to FADH2.
Hydration of double bond. Oxidation and transformation of NAD+
to NADH + H+. Acetyl CoA splits off and rest of chain
is bound to another CoA. … until fatty acid is at its end. Special cases are unsaturated fatty
acids and fatty acids with odd numbers of C atoms.
Fatty acid oxidation takes also place in peroxisomes, but no ATP generation.
Acetyl-CoA back to cytosol (synthesis)
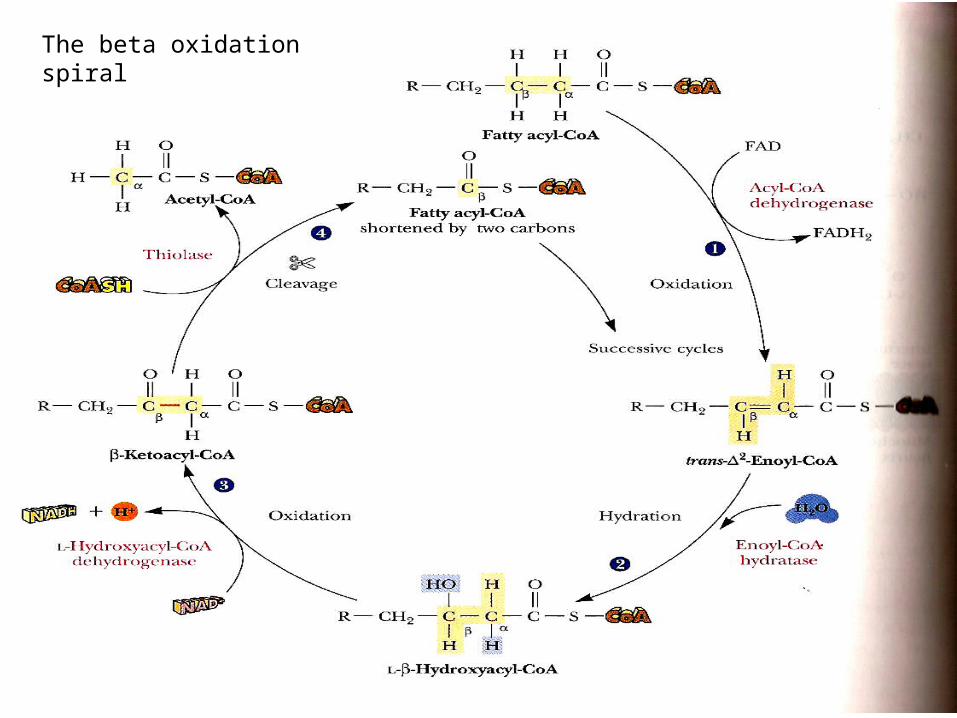
The beta oxidation spiral

Mitochondria do beta-oxidation
• Beta oxidation takes place inside mitochondria – so without mitochondria a tissue cannot metabolize fat for energy.

Ketone bodies and ketoacidosis
• High rates of fat oxidation tend to leave us with an overload of acetylCoA, which the liver converts to acetate, acetone, acetoacetate, and beta-OH butyrate. These are the so-called ketone bodies that appear in the blood, sweat and breath of individuals that are starving, or suffering from untreated diabetes mellitus. Some of them are acids, so their appearance in the blood causes ketoacidosis. Some of them are smelly, so they can be detected by a practitioner without the use of chemical analysis.

Odd vs even-numbered fatty acids
• Since fatty acids are taken apart (and also assembled) in 2-C pieces, some cells find it harder to deal with odd-numbered fatty acids – they are unusual in mammals but common in plants and marine organisms. The tail-end of an odd-numbered fatty acid turns out to be 3-C propionyl-CoA, which is decarboxylated in a multistep process to form pyruvate. So, we can eat oysters, after all.
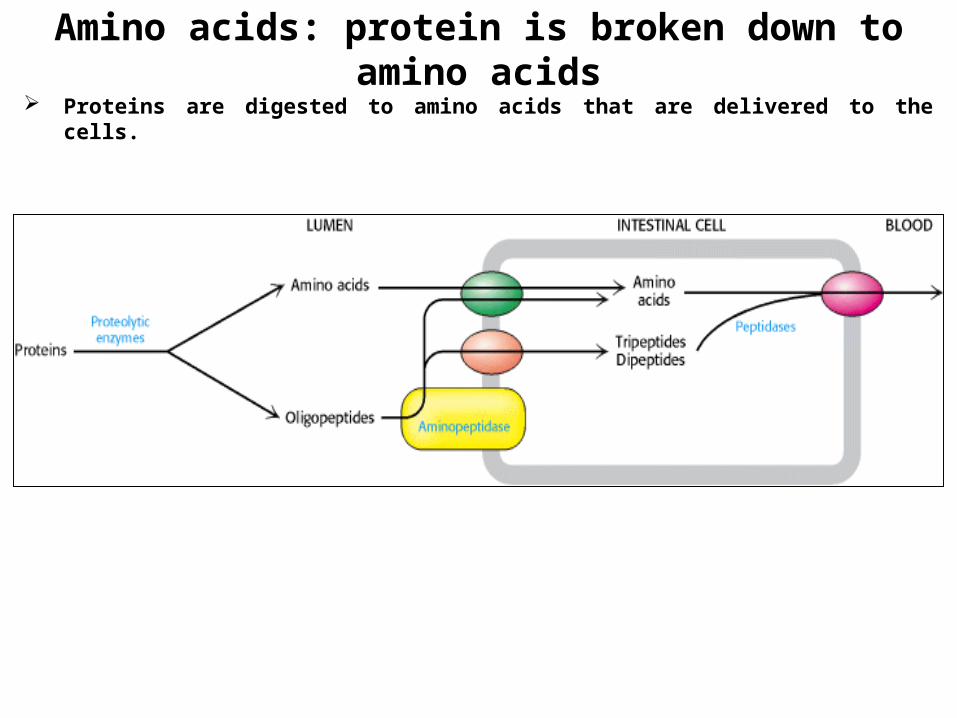
Amino acids: protein is broken down to amino acids Proteins are digested to amino acids that are delivered to the cells.
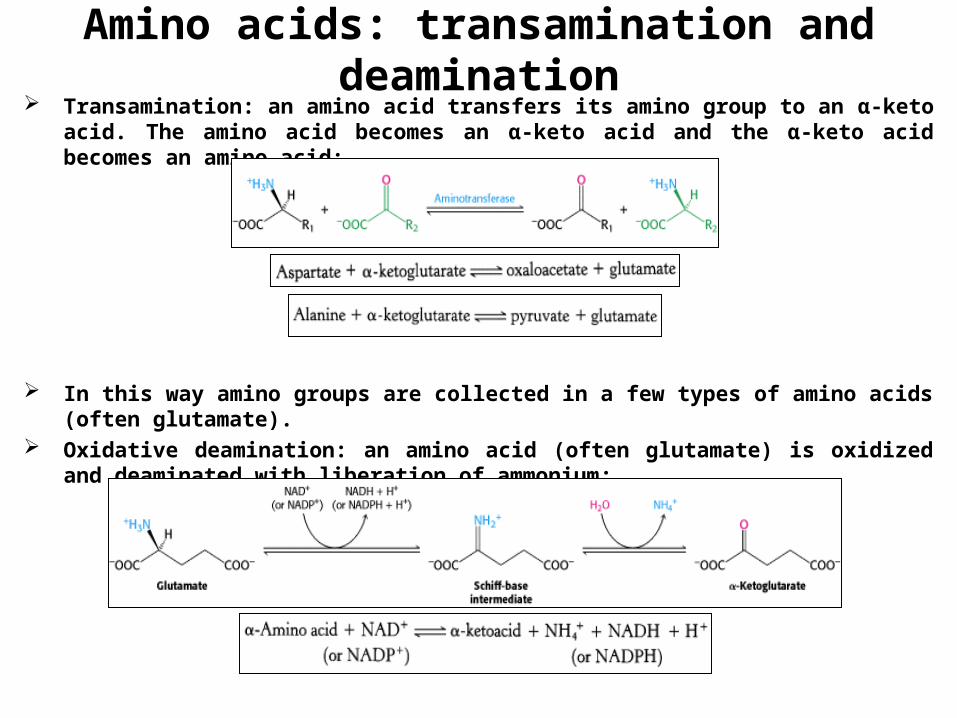
Amino acids: transamination and deamination Transamination: an amino acid transfers its amino group to an α-keto acid. The amino
acid becomes an α-keto acid and the α-keto acid becomes an amino acid:
In this way amino groups are collected in a few types of amino acids (often glutamate).
Oxidative deamination: an amino acid (often glutamate) is oxidized and deaminated with liberation of ammonium:
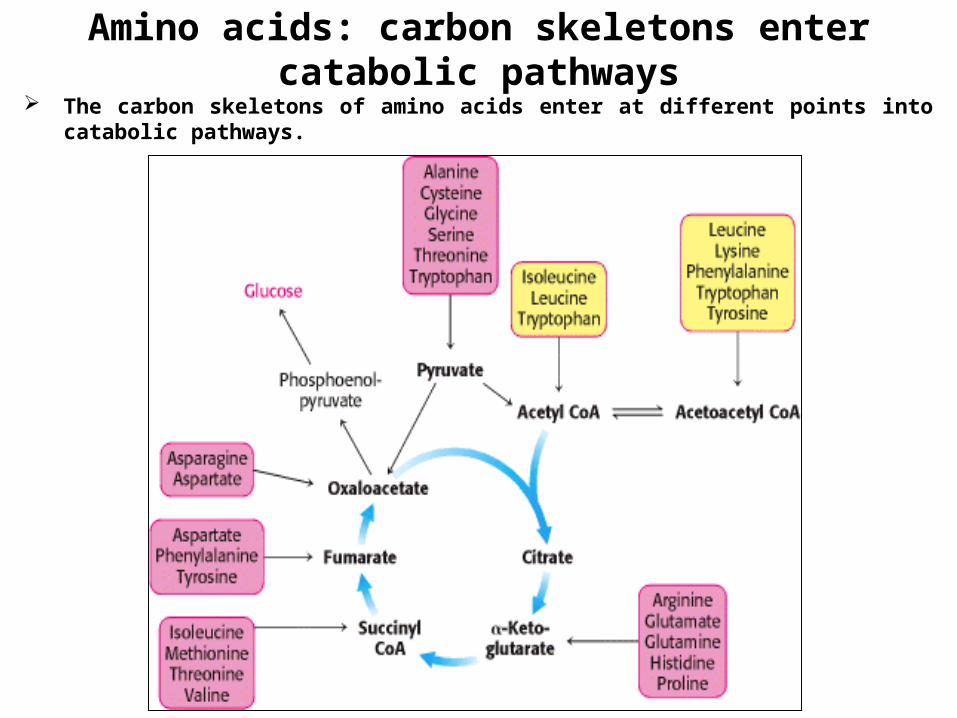
Amino acids: carbon skeletons enter catabolic pathways
The carbon skeletons of amino acids enter at different points into catabolic pathways.
















![Biochem [Gluconeogenesis]](https://img.pdfslide.net/doc/110x75/577c82b31a28abe054b1e4af/biochem-gluconeogenesis.jpg)