Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Apr. 1973, p. 378-389Copyright © 1973 American Society for Microbiology
Vol. 114, No. 1Printed in U.S.A.
Gram Characteristics and Wall Ultrastructure ofArthrobacter crystallopoietes During
Coccus-Rod MorphogenesisC. M. WARD, JR.,' AND G. W. CLAUS
Department of Microbiology, The Pennsylvania State University, University Park, Pennsylvania 16802
Received for publication 3 January 1973
Arthrobacter crystallopoietes growing exponentially as cocci were changed torods by adding succinate to the medium. Cells were sampled before, during, andafter this transition for Gram-staining and ultrastructural studies. Cells were
Gram stained by the standardized method of Bartholomew, and all sampleswere fixed and prepared for thin sectioning in an identical manner. Cocci weregram positive, and thin sections demonstrated a gram-positive type of cell wallhaving an average thickness of 31 nm. Cells sampled during morphogenesisappeared as cocci with most having a single rodlike projection. The coccusportion of these transition cells was gram positive and bound by a gram-positivetype of wall having an average thickness of 29 nm. The rodlike projection of thetransition cells appeared to be gram negative; it was also surrounded by a
gram-positive type of wall, but its average thickness was only 22 nm.
Gram-negative rods of the type species, Arthrobacter globiformis, were alsoexamined and found to produce a gram-positive type of wall with a 19-nmaverage thickness. Evidence for the trilaminar region, characteristic of mostgram-negative bacterial cell walls, was totally lacking in both species. Theseresults suggest that variations in cell wall thickness may be an importantcontributing factor to the variable Gram-staining characteristics of this genus.
Thin sections of gram-positive bacterial cellwalls usually reveal a thick, electron-dense,rather homogenous structure immediatelyoverlaying the plasma membrane (12). Al-though the gram-negative wall is depicted asseveral layers, the most distinctive layer ap-pears as a unit membrane on'the outer portionof the wall (7, 12). These structural differencesclosely parallel large differences in cell wallchemistry. One might assume that these grossstructural and chemical differences are reflec-tions of distinct and complex genetic dissimi-larities; therefore, changes from one Gram-staining characteristic to another duringgrowth of a pure culture seems very unlikely.On the other hand, certain bacteria show
marked changes in Gram-staining characteris-tics during growth. Gram-staining variability ischaracteristic of the genus Arthrobacter (6, 31),and these variations are often accompanied bychanges in cell shape. The type species, Ar-throbacter globiformis, is characterized as
I Present address: Department of Cell Biology, RocheInstitute, Nutley, N.J. 07110.
gram-positive cocci that change to gram-nega-tive rods during exponential growth and thenback to gram-positive cocci when exponentialgrowth ceases (8). Ensign and Wolfe (10) dem-onstrated that morphogenesis in A.crystallopoietes parallels that of the type spe-cies and can be experimentally controlled bynutritional manipulation. A. crystallopoietescocci are described as gram-variable cocci thatshift to gram-positive rods during exponentialgrowth (9).This report examines both the Gram-stain-
ing characteristics and the cell wall ultrastruc-ture of A. crystallopioetes before, during, andafter coccus-to-rod morphogenesis. These re-sults are compared with an examination of theArthrobacter type species and contrasted withthe generalization concerning the ultrastruc-ture of gram-positive and gram-negative cellwalls.
MATERIALS AND METHODSCultures. A. crystallopoietes ATCC 15481 was
received from the American Type Culture Collection378
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.

ARTHROBACTER CELL WALLS
in 1970 and maintained on slants containing 0.5%peptone and 1.5% agar. A. globiformis ATCC 4336was received from the Department of Microbiology,Pennsylvania State University (PSU) culture collec-tion in 1970 and also maintained on peptone-agarslants. Both Arthrobacter stock cultures were storedat 4 C and transferred every 3 months. Acetobactersuboxydans ATCC 621 was obtained from the Ameri-can Type Culture Collection in 1968, maintained at-20 C in 66% glycerol (13), and transferred every 4 to6 months. Staphylococcus aureus ATCC 10390 wasreceived from the Department of Microbiology, PSUculture collection in 1970 and maintained at 4 C onnutrient agar slants. Bacillus megaterium PSU 41and Pseudomonas aeruginosa PSU 191 were receivedin 1971 from the PSU culture collection and main-tained at 4 C on brain-heart infusion agar slants.
Growth. Subcultures of A. crystallopoietes or A.globiformis were grown at 28 C in 500-ml Nepheloculture flasks (Bellco Glass Inc., Vineland, N.J.)containing 50 ml of 1% peptone (Difco Laboratories,Detroit, Mich.) reciprocating at 200 strokes per minwith a 3.8-cm amplitude (standard shaking). Cellsfrom 2 ml of late exponential- or early stationary-phase cultures were centrifuged and washed threetimes with 0.9% saline buffered at pH 7.2 with 0.01 Mpotassium phosphate. Cells were then suspended in1.0 ml of buffered saline, and the entire volume wasused to inoculate 500-ml Nephelo flasks containing50 ml of a glucose-salts medium containing nitrilo-triacetic acid (10). Cells were grown in nitrilotriaceticacid-glucose-salts (NGS) medium at 28 C with stan-dard shaking. In order to initiate morphogenesis of A.crystallopoietes, a sterile solution of sodium succi-nate was added 28 h after inoculation to achieve a 1%(wt/vol) concentration in the NGS medium.
A. suboxydans was grown at 28 C on the basalcomplex medium (3) containing 5% glycerol, and S.aureus was grown on 0.8% nutrient broth. Whenthese cultures reached mid-exponential growth, theywere fixed by the addition of 20% (wt/vol) paraform-aldehyde. After 30 min, 10% (vol/vol) formaldehydewas used to wash each culture and to store the cells at4 C. These formaldehyde-fixed cultures were used asstandards for the Gram reaction.
B. megaterium and P. aeruginosa were grown in amedium containing 0.05% glucose, 0.5% tryptone,0.5% yeast extract, and 0.5% potassium phosphate(22). Cultures were harvested by centrifugation after10 h of growth and immediately were used asstandards for the lysozyme-EDTA experiment.Gram staining and light microscopy. Each mi-
croscope slide contained the experimentalArthrobacter cultures and separate smears of theformaldehyde-fixed gram-positive and gram-nega-tive standards. The Gram-staining procedure wascarefully standardized according to the procedure ofBartholomew (1). Reaction times were, in seconds: 60for crystal violet, 60 with Lugol iodine, 30 with 95%ethanol, and 30 with safranine. Observations weremade on Arthrobacter cultures only if 98 to 100% ofthe formaldehyde-fixed standards yielded the properappearance. A Zeiss model WL microscope wasadjusted for Kohler illumination, and all photo-
graphic records were made by using a 0.5 neutraldensity filter, a 350- to 600-nm band-pass filter, a 1.4numerical aperture achromatic condenser with theiris adjusted to the same position for each observa-tion, a 100x, 1.3 numerical aperture planapochro-matic objective, and a 1.0 Optovar setting. Kodakhigh-contrast copy film was used to record all images,and this was developed in Kodak D-76.
Electron microscopy. All Arthrobacter sampleswere prepared for electron microscopy observation inan identical manner. One milliliter of 1% OSO4 wasallowed to react with 9 ml of culture for 30 min. Cellswere centrifuged at 2,200 x g for 10 min, suspendedin 1 ml of 1% OSO4 and 0.1 ml of a solution containing1% tryptone with 0.5% NaCl (18), and then wereallowed to stand at 24 C for about 16 h. The fixedcells were removed by centrifugation, mixed withwarm 4% Noble agar, allowed to solidify, and thenwere cut into two mm3 blocks. The blocks wereplaced into a pH 6.1 solution containing 0.5% uranylacetate, 0.4% sodium acetate, 0.6% sodium barbitol,0.7% sodium chloride and 0.01% calcium dichloride,were allowed to remain in the 0.5% uranyl acetate for2 h, and then were dehydrated for 30 min in each ofthe following percentages of ethanol: 20, 40, 60, 95,and 100. Cells were infiltrated for 30 min each in50:50 and 25:75 mixtures of ethanol-Spurr epoxy(35) and then for 16 h with 100% Spurr epoxy, andthis was polymerized for 24 h at 70 C. Sectionsprepared with the LKB Ultrotome III were stainedwith lead citrate (37) and viewed with a PhillipsEM300 by using a 300-gm condenser aperture and a60-kV accelerating potential. Specimen images wereexposed on Kodak electron microscope film (Estarthick base) and were developed in D-19.
RESULTSGrowth and controlled morphogenesis. On
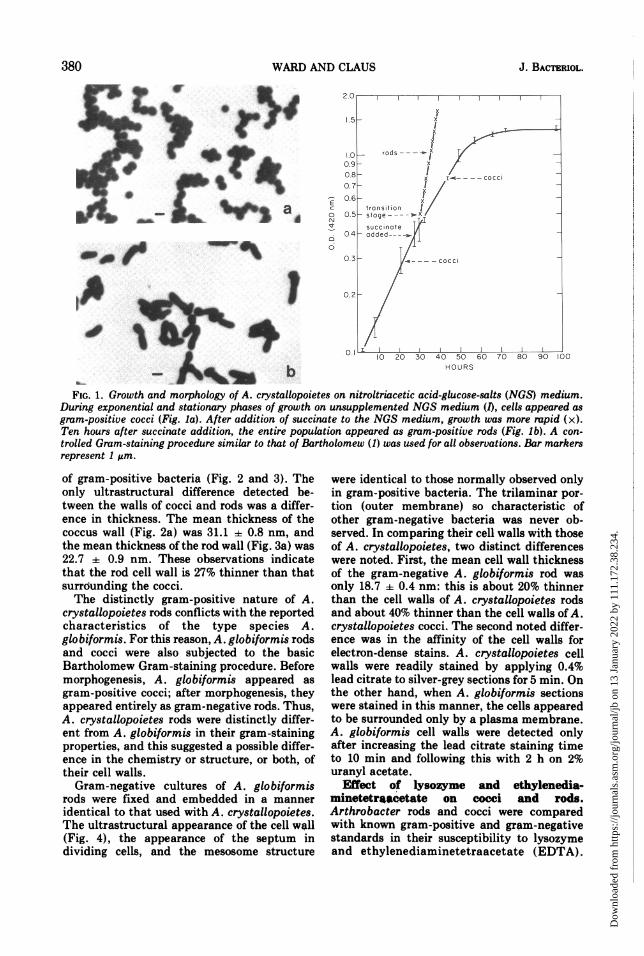
unsupplemented NGS medium, cells grew ex-ponentially with about a 13-h doubling time,and all cells were observed as cocci during eachgrowth phase (Fig. 1). On the other hand, whensuccinate was added to a 28-h culture of coccigrowing on the NGS medium, the doublingtime was reduced to about 5 h, and the coccibegan to form rods. Ten hours after succinateaddition, all cells in the culture appeared aslong rods (Fig. 1). These results are consistentwith those of Ensign and Wolfe (10) andconfirm that succinate experimentally initiatesthe coccus-to-rod morphogenesis.
Characteristics of cocci and rods. Undercarefully controlled staining conditions, A.crystallopoietes appeared as gram-positivecocci through all phases of growth on unsupple-mented NGS medium. A. crystallopoietes rods,formed after addition of succinate to the NGSmedium, also stained gram positively.Thin sections ofA. crystallopoietes harvested
during exponential growth as cocci or rodsconsistently exhibited a cell wall characteristic
379VOL. 114, 1973
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.
WARD AND CLAUS
ad
As
c
02
I
do;-_ ^ss ~b
2-0 --r-r
1.5 7 .
1.0 rods---0.9 X
0.8 J -- oc
0.7 _
0.6 /
transition |0.5 stage -w-
succ note0.4 added----,..
0.3 /--- cocci
0.2
10 - 1 ____ 1 120 30 40 50 60 70 80 90 100
HOURS
FIG. 1. Growth and morphology of A. crystallopoietes on nitroltriacetic acid-glucose-salts (NGS) medium.During exponential and stationary phases of growth on unsupplemented NGS medium (1), cells appeared asgram-positive cocci (Fig. la). After addition of succinate to the NGS medium, growth was more rapid (x).Ten hours after succinate addition, the entire population appeared as gram-positive rods (Fig. Ib). A con-trolled Gram-staining procedure similar to that of Bartholomew (1) was used for all observations. Bar markersrepresent 1 gm.
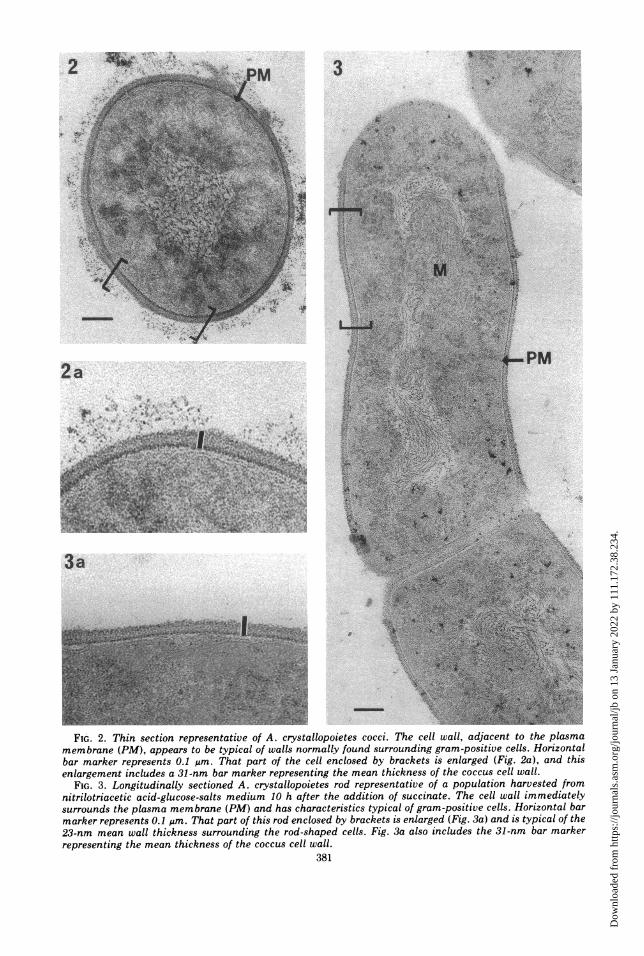
of gram-positive bacteria (Fig. 2 and 3). Theonly ultrastructural difference detected be-tween the walls of cocci and rods was a differ-ence in thickness. The mean thickness of thecoccus wall (Fig. 2a) was 31.1 ± 0.8 nm, andthe mean thickness of the rod wall (Fig. 3a) was22.7 ± 0.9 nm. These observations indicatethat the rod cell wall is 27% thinner than thatsurrounding the cocci.The distinctly gram-positive nature of A.
crystallopoietes rods conflicts with the reportedcharacteristics of the type species A.globiformis. For this reason, A. globiformis rodsand cocci were also subjected to the basicBartholomew Gram-staining procedure. Beforemorphogenesis, A. globiformis appeared asgram-positive cocci; after morphogenesis, theyappeared entirely as gram-negative rods. Thus,A. crystallopoietes rods were distinctly differ-ent from A. globiformis in their gram-stainingproperties, and this suggested a possible differ-ence in the chemistry or structure, or both, oftheir cell walls.
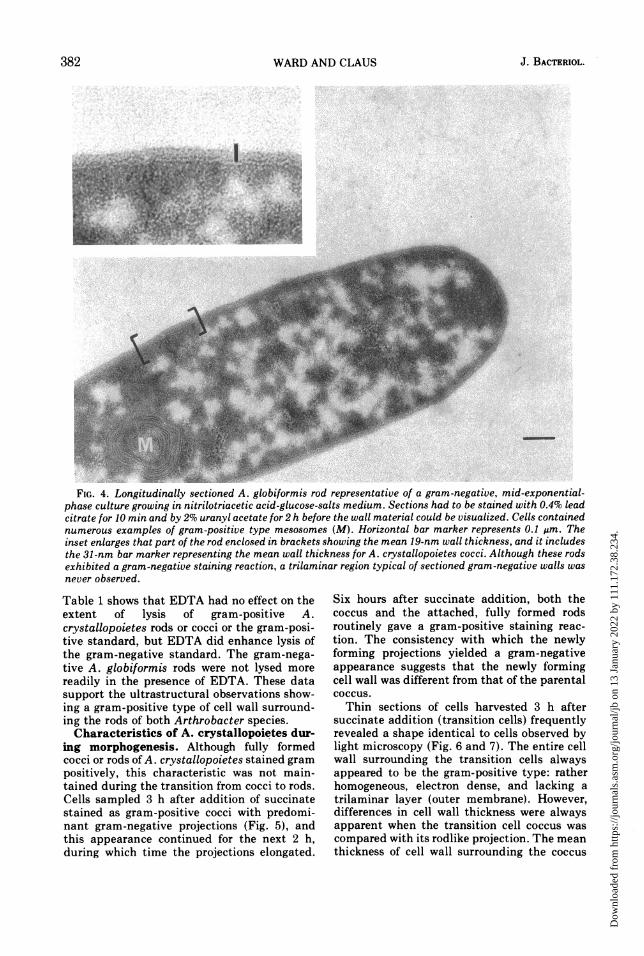
Gram-negative cultures of A. globiformisrods were fixed and embedded in a manneridentical to that used with A. crystallopoietes.The ultrastructural appearance of the cell wall(Fig. 4), the appearance of the septum individing cells, and the mesosome structure
were identical to those normally observed onlyin gram-positive bacteria. The trilaminar por-tion (outer membrane) so characteristic ofother gram-negative bacteria was never ob-served. In comparing their cell walls with thoseof A. crystallopoietes, two distinct differenceswere noted. First, the mean cell wall thicknessof the gram-negative A. globiformis rod wasonly 18.7 ± 0.4 nm: this is about 20% thinnerthan the cell walls of A. crystallopoietes rodsand about 40% thinner than the cell walls of A.crystallopoietes cocci. The second noted differ-ence was in the affinity of the cell walls forelectron-dense stains. A. crystallopoietes cellwalls were readily stained by applying 0.4%lead citrate to silver-grey sections for 5 min. Onthe other hand, when A. globiformis sectionswere stained in this manner, the cells appearedto be surrounded only by a plasma membrane.A. globiformis cell walls were detected onlyafter increasing the lead citrate staining timeto 10 min and following this with 2 h on 2%uranyl acetate.
Effect of lysozyme and ethylenedia-minetetrnacetate on cocci and rods.Arthrobacter rods and cocci were comparedwith known gram-positive and gram-negativestandards in their susceptibility to lysozymeand ethylenediaminetetraacetate (EDTA).
380 J. BACTERIOL.
,i
U .1,
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.
S't V ." 'I ., =
2~~~~~vM . ,:a~~_P:~~~~~~~~ ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~4
4~~~~S
~~Y~~:r~~2~~>.?¶~~~ A~~': -'p
membrane (PM), appears to be typical of walls normally found surrounding gram-positive cel.-oiznabar marker represents 0.1 ,im. That part of the cell enclosed by brackets is enlarged (Fig. 2a), and thisenlargement includes a 31-nm bar marker representing the mean thickness of the coccus cell wall.
FIG. 3. Longitudinally sectioned A. crystallopoietes rod representative of a population harvested fromnitrilotriacetic acid-glucose-salts medium 10 h after the addition of succinate. The cell wall immediatelysurrounds the plasma membrane (PM) and has characteristics typical of gram-positive cells. Horizontal barmarker represents 0.1 ,um. That part of this rod enclosed by brackets is enlarged (Fig. 3a) and is typical of the23-nm mean wall thickness surrounding the rod-shaped cells. Fig. 3a also includes the 31-nm bar markerrepresenting the mean thickness of the coccus cell wall.
381
wme I-. 1.
I- AW;, -.K -.-
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.
WARD AND CLAUS
FIG. 4. Longitudinally sectioned A. globiformis rod representative of a gram-negative, mid-exponential-phase culture growing in nitrilotriacetic acid-glucose-salts medium. Sections had to be stained with 0.4% leadcitrate for 10 min and by 2% uranyl acetate for 2 h before the wall material could be visualized. Cells containednumerous examples of gram-positive type mesosomes (M). Horizontal bar marker represents 0.1 MAm. Theinset enlarges that part of the rod enclosed in brackets showing the mean 19-nm wall thickness, and it includesthe 31-nm bar marker representing the mean wall thickness for A. crystallopoietes cocci. Although these rodsexhibited a gram-negative staining reaction, a trilaminar region typical of sectioned gram-negative walls wasnever observed.
Table 1 shows that EDTA had no effect on theextent of lysis of gram-positive A.crystallopoietes rods or cocci or the gram-posi-tive standard, but EDTA did enhance lysis ofthe gram-negative standard. The gram-nega-tive A. globiformis rods were not lysed morereadily in the presence of EDTA. These datasupport the ultrastructural observations show-ing a gram-positive type of cell wall surround-ing the rods of both Arthrobacter species.Characteristics of A. crystallopoietes dur-
ing morphogenesis. Although fully formedcocci or rods of A. crystallopoietes stained grampositively, this characteristic was not main-tained during the transition from cocci to rods.Cells sampled 3 h after addition of succinatestained as gram-positive cocci with predomi-nant gram-negative projections (Fig. 5), andthis appearance continued for the next 2 h,during which time the projections elongated.
Six hours after succinate addition, both thecoccus and the attached, fully formed rodsroutinely gave a gram-positive staining reac-tion. The consistency with which the newlyforming projections yielded a gram-negativeappearance suggests that the newly formingcell wall was different from that of the parentalcoccus.Thin sections of cells harvested 3 h after
succinate addition (transition cells) frequentlyrevealed a shape identical to cells observed bylight microscopy (Fig. 6 and 7). The entire cellwall surrounding the transition cells alwaysappeared to be the gram-positive type: ratherhomogeneous, electron dense, and lacking atrilaminar layer (outer membrane). However,differences in cell wall thickness were alwaysapparent when the transition cell coccus wascompared with its rodlike projection. The meanthickness of cell wall surrounding the coccus
382 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.
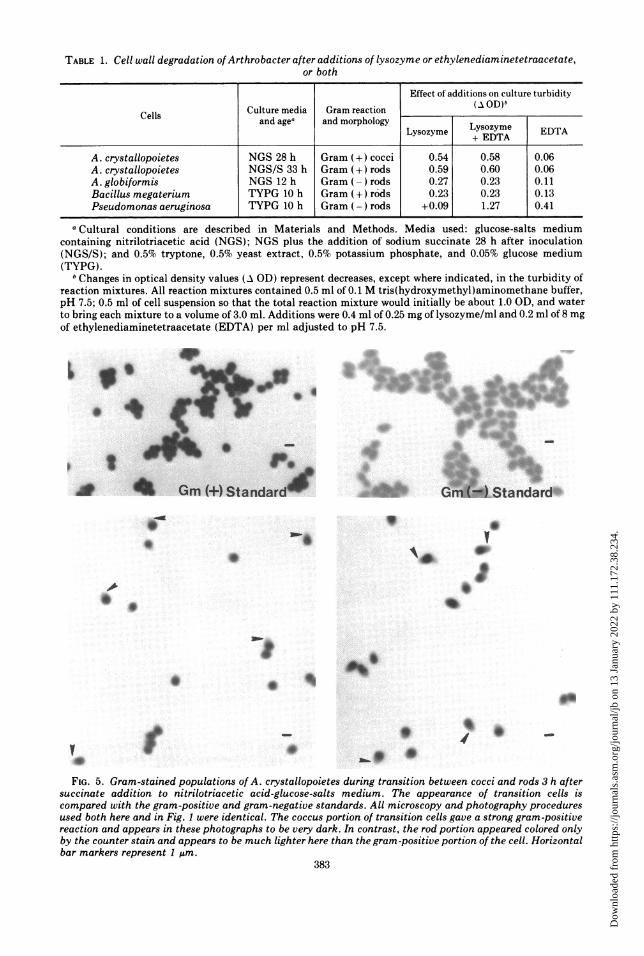
TABLE 1. Cell wall degradation of Arthrobacter after additions of lysozyme or ethylenediaminetetraacetate,or both
Effect of additions on culture turbidityCulture media Gram reaction (AOD)bCells and agea andmorphologyLyome ET
Lysozyme +yszyeDTA T
A.crstalopoitesNS 28h Grm (+)cocc 0.5 0.5 0.06A. crystallopoietes NGS/ 33h Gram c+odsi 0.59 0.60 0.06
A.globiformis NGS 12 h Gram ()rods 0.27 0.23 0.11Bacillus megaterium TYPG 10 h Gram (+ r6ds 0.23 0.23 0.13Pseudomonas aeruginosa TYPG 10 h Gram (-) rods +0.09 1.27 0.41
aCultural conditions are described in Materials and Methods. Media used: glucose-salts mediumcontaining nitrilotriacetic acid (NGS); NGS plus the addition of sodium succinate 28 h after inoculation(NGS/S); and 0.5% tryptone, 0.5% yeast extract, 0.5% potassium 'phosphate, and 0.05% glucose medium(TYPG).
bChanges in optical density values (A GD) represent decreases, except where indicated, in the turbidity ofreaction mixtures. All reaction mixtures contained 0.5 ml of 0.1 M tris(hydroxymethyl)aminomethane buffer,pH 7.5; 0.5 ml of cell suspension so that the total reaction mixture would initially be about 1.0 GD, and waterto bring each mixture to a volume of 3.0 ml. Additions were 0.4 ml of 0.25 mg of lysozyme/ml and 0.2 ml of 8 mgof ethylenediaminetetraacetate (EDTA) per ml adjusted to pH 7.5.
dr '4
Gn)Standard-
I
0 w
.9
Sb
*0*
0V
FIG. 5. Gram-stained populations of A. crystallopoietes during transition between cocci and rods 3 h aftersuccinate addition to nitrilotriacetic acid-glucose-salts medium. The appearance of transition cells iscompared with the gram-positive and gram-negative standards. All microscopy and photography proceduresused both here and in Fig. I were identical. The coccus portion of transition cells gave a strong gram-positivereaction and appears in these photographs to be very dark. In contrast, the rod portion appeared colored onlyby the counter stain and appears to be much lighter here than the gram-positive portion of the cell. Horizontalbar markers represent 1 rim.
383
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.
WARD AND CLAUS
a b
A
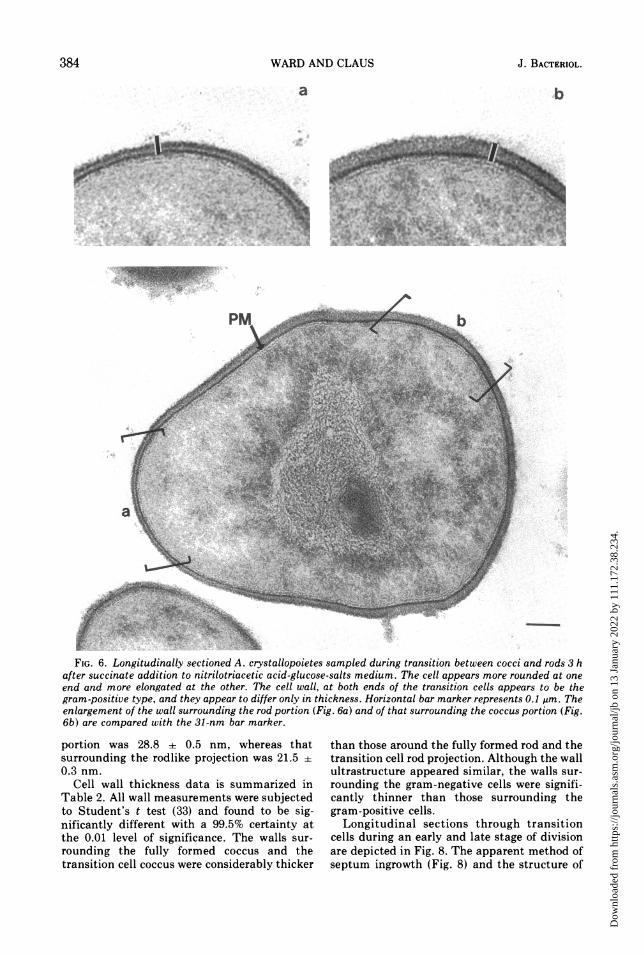
FIG. 6. Longitudinally sectioned A. crystallopoietes sampled during transition between cocci and rods 3 hafter succinate addition to nitrilotriacetic acid-glucose-salts medium. The cell appears more rounded at oneend and more elongated at the other. The cell wall, at both ends of the transition cells appears to be thegram-positive type, and they appear to differ only in thickness. Horizontal bar marker represents 0.1 um. Theenlargement of the wall surrounding the rod portion (Fig. 6a) and of that surrounding the coccus portion (Fig.6b) are compared with the 31-nm bar marker.
portion was 28.8 + 0.5 nm, whereas thatsurrounding the rodlike projection was 21.5 +0.3 nm.
Cell wall thickness data is summarized inTable 2. All wall measurements were subjectedto Student's t test (33) and found to be sig-nificantly different with a 99.5% certainty atthe 0.01 level of significance. The walls sur-rounding the fully formed coccus and thetransition cell coccus were considerably thicker
than those around the fully formed rod and thetransition cell rod projection. Although the wallultrastructure appeared similar, the walls sur-rounding the gram-negative cells were signifi-cantly thinner than those surrounding thegram-positive cells.
Longitudinal sections through transitioncells during an early and late stage of divisionare depicted in Fig. 8. The apparent method ofseptum ingrowth (Fig. 8) and the structure of
384 J. BACTERIOL.
'r . ; ,c
I
-t - . ".'i t
I ftN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.
ARTHROBACTER CELL WALLS
--
--
rF
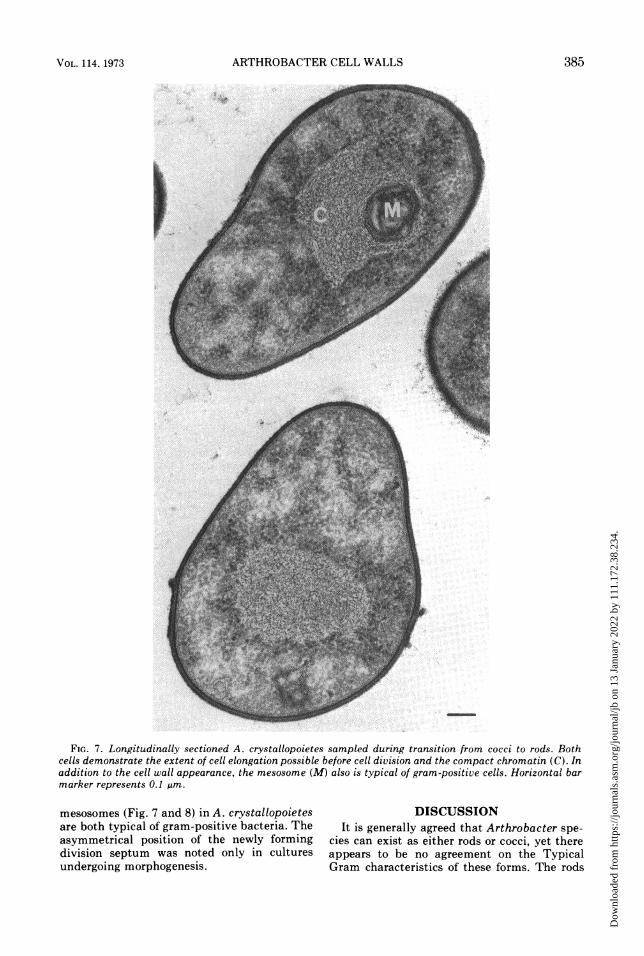
FIG. 7. Longitudinally sectioned A. crystallopoietes sampled during transition from cocci to rods. Bothcells demonstrate the extent of cell elongation possible before cell division and the compact chromatin (C). Inaddition to the cell wall appearance, the mesosome (M) also is typical of gram-positive cells. Horizonta' barmarker represents 0.1 gim.
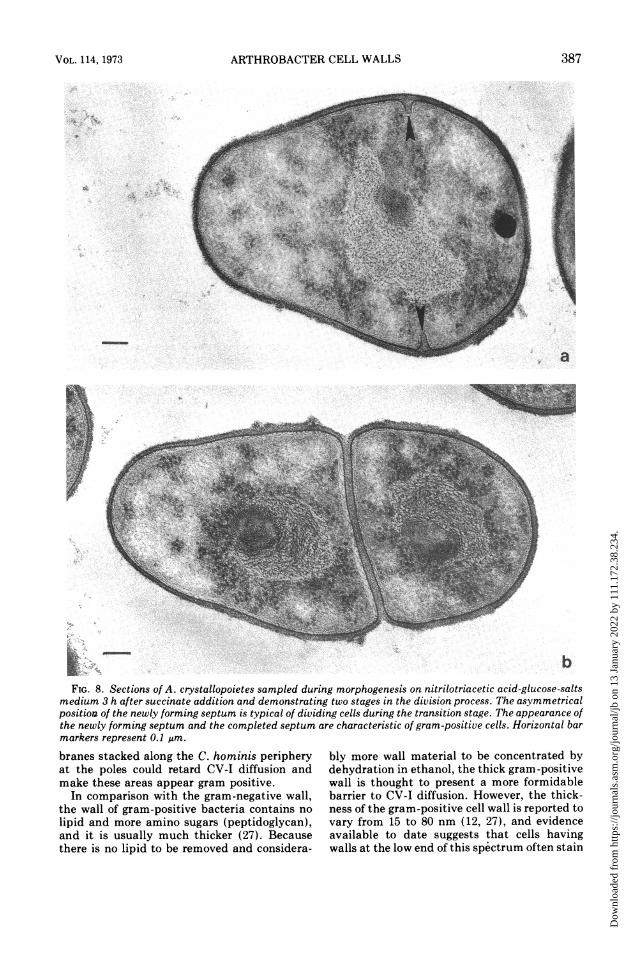
mesosomes (Fig. 7 and 8) in A. crystallopoietesare both typical of gram-positive bacteria. Theasymmetrical position of the newly formingdivision septum was noted only in culturesundergoing morphogenesis.
DISCUSSIONIt is generally agreed that Arthrobacter spe-
cies can exist as either rods or cocci, yet thereappears to be no agreement on the TypicalGram characteristics of these forms. The rods
VOL. 114, 1973 385
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.

TABLE 2. Cell-wall thickness and Gram reaction ofArthrobacter crystallopoietes and A. globiformis
Medium (culture No. of No. of Average wall GramCells M e )a Morphology cells measure- thickness reactiontmeasured ments (nm)b r
A. crystallopoietes NGS (28 h) Cocci 12 66 31.1 ± 0.8 +NGS/S (31 h) Coccus portion of tran- 48 297 28.8 ± 0.5 +
sition cellsNGS/S (31 h) Projection portion of tran- 48 350 21.5 ± 0.3
sition cellsPeptone (8 h) Rods 29 121 22.7 ±0.9 +
A. globiformis NGS (12 h) Rods 50 226 18.7 ± 0.4
a Cultural conditions are described in Materials and Methods. Media used: glucose-salts mediumcontaining nitrolotriacetic acid (NGS); NGS plus the addition of sodium succinate 28 h after inoculation(NGS/S); and 1% peptone.
bMeasurements on enlarged prints of thin-sectioned cells were taken only where the plasma membrane wasslightly pulled away from the inside surface of the cell wall. Transition cells were divided in thirdsperpendicular to their long axis, and measurements were taken randomly along the wall and along all but themiddle third of the cell. By using Student's t test (33), it was found that each measurement listed above wassignificantly different from all others with a 99.5% certainty at the 0.01 level of significance.
' Gram reaction method was the carefully controlled procedure of Bartholomew (1).
of various species have been described as gramnegative (8, 23), gram negative with gram-posi-tive granules (21, 29), gram negative to gramvariable (5), "predominately" gram positive(29), and gram positive (9, 16, 17). Cocci havebeen described as "generally" gram negative(23), gram variable (9), gram variable to grampositive (5), gram positive to gram negativewith positive granules (17), "predominately"gram positive (29), and gram positive (8, 16,21). On the basis of this literature, it seemsthat the Gram reaction does not reflect a stablecharacteristic of this genus, nor does it justifyassumption of a set of ultrastructural charac-teristics for Arthrobacter that is normally as-sociated with either gram-positive or gram-negative bacteria.Four observations presented here suggest
that A. globiformis, the type species, and A.crystallopoietes are cytologically typical of thegram-positive bacteria. First, the appearanceof sectioned walls is like that of other gram-positive bacteria in that it consistently lacksthe distinct outer membrane that is typical ofgram-negative bacteria (7, 12). Secondly, crosswalls formed during division of both speciesresemble those found only in gram-positivebacteria (15, 36). Thirdly, lysozyme easilydegrades the wall and allows lysis in theabsence of EDTA. Finally, the mesosomesfound within these species are structurallysimilar to the mesosomes of gram-positivebacteria (26, 36). The only other known fine-structural study on Arthrobacter is that ofStevenson (35) who demonstrated that A.pascens also exhibits cytological characteristics
typical of gram-positive bacteria.The evidence presented here demonstrates
that Arthrobacter is surrounded by an ultra-structurally typical, gram-positive cell wall.Nevertheless, carefully controlled Gram-stain-ing procedures show that certain forms ofArthrobacter are gram negative. In order toexplain these results, one must consider what isknown about the chemistry and anatomy of thebacterial cell wall and their relation to theGram reaction.
In contrast to gram-positive walls, typicalgram-negative walls contain a lipoproteinmembrane and much less peptidoglycan (27).Treatment with 95% ethanol presumablyremoves the wall lipid and concentrates thewall peptidoglycan through dehydration. Thealtered wall that remains does not preventrapid outward diffusion of the solubilized crys-tal violet-iodine complex (CV-I), and thereforethe CV-I is rapidly lost to the surrounding etha-nol. Any alterations in the cell's peripheralstructure that would slow or prevent this out-ward diffusion might allow part or all of the cellto appear gram positive. For example, Cardio-bacterium hominis (32) stains as a gram-nega-tive rod with gram-positive regions occurringparticularly at the cell poles. Ultrastructuralstudies show C. hominis surrounded by a typi-cal gram-negative wall, but these cells also con-tain peripheral layers of membrane inside theplasma membrane, especially at the poles (24).Most experimental results confirm that theGram reaction is dependent upon the relativerates of CV-I diffusion from the cell in ethanol(2, 28, 30); therefore, the additional mem-
386 WARD AND CLAUS J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.
ARTHROBACTER CELL WALLS 387
_ ~~~~a
~~ ~ ~ ~ ~ ~ ~ ~ ~ W~~~~~~~~~~~~~~bFIG. 8. Sections of A. crystallopoietes sampled during morphogenesis on nitrilotriacetic acid-glucose-salts
medium 3 h after succinate addition and demonstrating two stages in the division process. The asymmetricalposition of the newly forming septum is typical of dividing cells during the transition stage. The appearance ofthe newly forming septum and the completed septum are characteristic of gram-positive cells. Horizontal barmarkers represent 0.1 pm.
branes stacked along the C. hominis peripheryat the poles could retard CV-I diffusion andmake these areas appear gram positive.
In comparison with the gram-negative wall,the wall of gram-positive bacteria contains no
lipid and more amino sugars (peptidoglycan),and it is usually much thicker (27). Becausethere is no lipid to be removed and considera-
bly more wall material to be concentrated bydehydration in ethanol, the thick gram-positivewall is thought to present a more formidablebarrier to CV-I diffusion. However, the thick-ness of the gram-positive cell wall is reported tovary from 15 to 80 nm (12, 27), and evidenceavailable to date suggests that cells havingwalls at the low end of this spectrum often stain
VOL. 114, 1973
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.

WARD AND CLAUS
gram negative. For example, Haemophilusvaginalis has an ultrastructurally typical gram-positive wall that is only 15-nm thick, andthese cells stain gram negative to gram varia-ble (25). Our data show a similarly appearing19-nm wall around A. globiformis, and thesecells consistently stain gram negative. On theother hand, our studies on the structurallytypical gram-positive wall of A.crystallopoietes rods and cocci demonstrate a23- and 31-nm wall thickness, and these cellsconsistently stain gram positive (Table 2).These data suggest that the thickness of thetypical gram-positive cell wall will influence itsability to retard CV-I diffusion during theGramr-staining procedure.The rate of CV-I diffusion from the bacterial
cell is not an all-or-none phenomenon, butrelatively fast or slow depending upon the typeof cell being stained (2, 28, 30). This rate isusually constant for all cells in a pure culture,but certain bacteria will exhibit both gram-posi-tive and gram-negative cells on the same slide,and these cultures are called "gram variable."This characteristic is often given to older cul-tures that were uniformly gram positive duringtheir exponential growth. Young cultures ofcertain bacteria have also been described asgram variable, but there are only a few of thesecases where we also have information on theirwall ultrastructure or chemistry. A. globiformisrods and H. vaginalis are described as gramnegative to gram variable (5, 11, 25), and bothare surrounded by very thin walls of the gram-positive type (Table 2, 25). Our results sug-gest that these wall thicknesses may be nearor below that necessary to consistantly yielda gram-positive appearance. At this lowerlimit, slight variations in wall thickness fromcell to cell may be enough to account for agram-variable appearance in young cultures.Our data showing a 7-nm (25%) difference inwall thickness between the coccus and rodlikeprojection of transition cells, and a correspond-ing difference in gram reaction, support thispossibility. Nevertheless, there is a question ofwhether Arthrobacter species are gram variable(4, 5, 8, 9, 31). Our results show that A.globiformis rods stain uniformly gram nega-tive, that A. crystallopietes rods and cocci areconsistently gram positive, and that the stand-ard deviation for the mean wall thickness ofeach cell population is very small (Table 2).
In addition to its effect on the gram reaction,cell wall thickness in Arthrobacter may becorrelated with growth rate. Ensign and Wolfe(10) demonstrated that growth of A.crystallopoietes rods is faster than their growth
as cocci. Our results confirm this (Fig. 1) andalso show that rods maintaining this fastergrowth rate are surrounded by a thinner wallthan that which surrounds the cocci (Table 2;Fig. 2, 3). Other Arthrobacter species formrods only during periods of exponential growth,and this report demonstrates that rods of theactively growing type-species are very thin.Since less wall material surrounds the fastergrowing rods, it appears that cell wall synthesisis not directly proportional to growth rate.Others have reported that nongrowing gram-positive bacteria have thicker walls than thosegrowing exponentially (12).
It is generally concluded that the wallstrength needed to maintain a rod-shaped cellis greater than that necessary for maintenanceof a sphere within an environment of lowosmolarity. Yet in this study we found thatArthrobacter rods are surrounded by thinnerwalls than those surrounding their cocci. If weassume that the observed decrease in thicknessduring coccus-to-rod morphogenesis is also ac-companied by a corresponding decrease inpeptidoglycan concentration, then either thepeptidoglycan remaining around the thinnerwalls has sufficient strength to maintain therod shape or the rod peptidoglycan may bechemically changed to compensate in strengthfor the reduced quantity. Krulwich et al. (19,20) have found chemical differences betweenthe rod and coccus peptidoglycan of A.crystallopoietes. Although the interpeptidecross-linking within the polymer from each celltype was similar, they found that the polysac-charide backbone of the peptidoglycan isolatedfrom the rod wall was three to four times longerthan that isolated from the coccus wall. Theysuggested that this chemical difference couldmake the rod walls more rigid than those of thecocci. If this chemical change accompaniesmorphogenesis, then it could account for theextra strength needed to maintain the thinnerrod wall.
Cultures sampled shortly after initiatingmorphogenesis showed many transition cellsthat lacked a division septum (Fig. 6, 7). Thus,it appears that the beginning of coccus-to-rodmorphogenesis precedes the initiation of celldivision. When a division septum is evident(Fig. 8), the shape of the transition cell sug-gests that the newly forming rod is a linearextension of the coccus perpendicular to theplane of the newly forming septum. The lead-ing edge of this rodlike projection is surroundedby a wall that is thinner than that around otherareas of the transition cell. Higgins and Shock-man (14, 15) demonstrated that the newly
388 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.

ARTHROBACTER CELL WALLS
forming wall in dividing Streptococcus is adja-cent to the invaginating septum, that it isthinner than the older wall, and that its linearextension is followed by wall thickening. It willbe interesting for future investigators to deter-mine whether the leading edge of Arthrobactertransition cells is also a region active in wallbiosynthesis.
ACKNOWLEDGMENTSThis investigation was supported in part by grant
GB-17493 from the National Science Foundation.We gratefully acknowledge Carol Ann Baker for her
technical assistance in preparing this manuscript.
LITERATURE CITED
1. Bartholomew, J. W. 1962. Variables influencing results,and the precise definition of steps in gram staining as a
means of standardizing the results obtained. StainTechnol. 37:139-155.
2. Bartholomew, J. W., and T. Mittwer. 1952. The Gramstain. Bacteriol. Rev. 16:1-29.
3. Batzing, B. L., and G. W. Claus. 1971. Biphasic growthof Acetobacter suboxydans on a glycerol-limiting me-
dium. J. Bacteriol. 108:592-595.4. Breed, R. S., B. G. D. Murray, and M. R. Smith. 1957.
Bergey's manual of determinative bacteriology, 7thed. The Williams & Wilkins Co., Baltimore.
5. Chaplin, C. E. 1957. Life cycles in Arthrobacter pascensand Arthrobacter terregens. Can. J. Microbiol.3:103-106.
6. Clark, J. B. 1972. Morphogenesis in the genusArthrobacter. Crit. Rev. Microbiol. 1:521-544.
7. Claus, G. W., and L. E. Roth. 1964. Fine structure of theGram-negative bacterium Acetobacter suboxydans. J.Cell Biol. 20:217-233.
8. Conn, H. J., and I. Dimmick. 1947. Soil bacteria similarin morphology to Mycobacterium and Corynebacterium. J. Bacteriol. 54:291-303.
9. Ensign, J. C., and S. C. Rittenberg. 1963. A crystallinepigment produced from 2-hydroxypyridine by Ar-throbacter crystallopoietes n. sp. Arch. Mikrobiol.47:137-153.
10. Ensign, J. C., and R. S. Wolfe. 1964. Nutritional controlof morphogenesis in Arthrobacter crystallopoietes. J.Bacteriol. 87:924-932.
11. Gillespie, D. C. 1963. Composition of cell wall mucopep-tide from Arthrobacter globiformis. Can. J. Microbiol.9:515-521.
12. Glauert, A. M., and M. J. Thornby. 1969. The topogra-phy of the bacterial cell wall. Ann. Rev. Microbiol.23: 159-198.
13. Greenfield, S., and G. W. Claus. 1969. Isocitrate dehy-drogenase and glutamate synthesis in Acetobactersuboxydans. J. Bacteriol. 100:1264-1270.
14. Higgins, M. L., and G. D. Shockman. 1970. Model forcell wall growth of Streptococcus faecalis. J. Bacteriol.101:643-648.
15. Higgins, M. L., and G. D. Shockman. 1971. Procaryoticcell division with respect to wall and membranes. Crit.Rev. Microbiol. 1:29-72.
16. lisuka, H., and K. Komagata. 1965. Microbiologicalstudies on petroleum and natural gas. III. Determina-tion of Brevibacterium, Arthrobacter, Micrococcus,Sarcina, Alcaligenes, and Achromobacter isolatedfrom oil-brines in Japan. J. Gen. Appl. Microbiol.11:1-14.
17. Kaneko, T., K. Kitamura, and Y. Yamamoto. 1969.
Arthrobacter luteus, nov. sp. isolated from brewerysewage. J. Gen. Appl. Microbiol. 15:317-326.
18. Kellenberger, E., A. Ryter, and J. Sechaud. 1958.Electron microscopic study of DNA-containingplasms. III. Vegative and mature phage DNA as
compared with normal bacterial nucleoids in differentphysiological states. J. Biophys. Biochem. Cytol.4:671-675.
19. Krulwich, T. A., J. C. Ensign, D. J. Tipper, and J. L.Strominger. 1967. Sphere-rod morphogenesis in Ar-throbacter crystallopoietes. I. Cell wall compositionand polysaccharides of the peptidoglycan. J. Bacteri-ol. 94:734-740.
20. Krulwich, T. A., J. C. Ensign, D. J. Tipper, and J. L.Strominger. 1967. Sphere-rod morphogenesis in Ar-throbacter crystallopoietes. II. Peptides of the cell wallpeptidoglycan. J. Bacteriol. 94:741-750.
21. Kuhn, D. A., and M. P. Starr. 1960. Arthrobacteratrocyaneus, n. sp., and its blue pigment. Arch.Mikrobiol. 36:175-181.
22. Lichstein, H. C., and E. L. Oginsky. 1965. Protoplastsand spheroplasts, p. 9-11. In Experimental microbialphysiology. W. H. Freeman and Co., San Francisco.
23. Lochhead, A. G., and M. 0. Burton. 1953. An essentialbacterial growth factor produced by microbial synthe-sis. Can. J. Bot. 31:7-21.
24. Reyn, A., A. Birch-Anderson, and R. G. E. Murray.1971. The fine structure of Cardiobacterium haminis.Acta Pathol Microbiol. Scand. Sect. B 79:51-60.
25. Reyn, A., S. P. Lapage, and A. Birch-Anderson. 1966. Anelectron microscope study of thin-sections of Haemo-philus vaginalis (Gardner and Dukes) and some possi-bly related species. Can. J. Microbiol. 12:1125-1136.
26. Ryter, A. 1968. Association of the nucleus and themembrane of bacteria: a morphological study; Bacter-iol. Rev. 32:39-54.
27. Salton, M. R. J. 1964. The bacterial cell wall. ElsevierPublishing Co., Amsterdam.
28. Salton, M. R. J. 1963. The relationship between thenature of the cell wall and the Gram stain. J. Gen.Microbiol. 30:223-235.
29. Schippers-Lammertse, A. F., A. 0. Muijsers, and K. B.Klaster-Oedekerk. 1963. Arthrobacter poly-chromogenes sp. nov., its pigments and a bacterio-phage of this species. Antonie van Leeuwenhoek J.Microbiol. Serol. 29:1-15.
30. Shugar, D., and J. Baranowska. 1958. QuantitativeGram staining with labelled iodine. Nature (London)181:357-358.
31. Skerman, V. B. D. 1959. A guide to the identification ofthe genera of bacteria. The Williams & Wilkins Co.,Baltimore.
32. Slotnick, I. J., and M. Dougherty. 1964. Further charac-terization of an unclassified group of bacteria causingendocarditis in man: Cardiobacterium hominis gen. etsp. nov. Antonie van Leeuwenhoek J. Microbiol.Serol. 30:261-272.
33. Spiegel, M. R. 1961. Schaum's outline series: theory andproblems of statistics. McGraw-Hill Book Co., NewYork.
34. Spurr, A. R. 1969. A low-viscosity epoxy resin embed-ding medium for electron microscopy. J. Ultrastruct.Res. 26:31-43.
35. Stevenson, I. L. 1968. Fine structure of Arthrobacterpascens and the development of mesosomes during thegrowth cycle. Can. J. Microbiol. 14:1029-1034.
36. Van Iterson, W. 1965. Symposium on the fine structureand replication of bacteria and their parts. II. Bacte-rial cytoplasm. Bacteriol. Rev. 29:299-325.
37. Venable, J. H., and R. Coggeshall. 1965. A simplifiedlead citrate stain for use in electron microscopy. J. CellBiol. 25:407-408.
VOL. 114, 1973 389
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
13
Janu
ary
2022
by
111.
172.
38.2
34.









![Practice For May: Cell Ultrastructure [114 marks]blogs.4j.lane.edu/.../2018/02/Cell-Ultrastructure-Test-1.pdfPractice For May: Cell Ultrastructure [114 marks]1. Which structure found](https://img.pdfslide.net/doc/110x75/5eda4db5b3745412b5711d9c/practice-for-may-cell-ultrastructure-114-marksblogs4jlaneedu201802cell-ultrastructure-test-1pdf.jpg)



















