Embed Size (px)
Citation preview

REGULAR ARTICLE
Granzyme B-dependent and perforin-independent DNAfragmentation in intestinal epithelial cells induced by anti-CD3mAb-activated intra-epithelial lymphocytes
Masaki Ogata & Yuta Ota & Takaji Matsutani &Masanobu Nanno & Ryuji Suzuki & Tsunetoshi Itoh
Received: 2 October 2012 /Accepted: 3 December 2012 /Published online: 30 January 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract We previously found that an i.p. injection of anti-CD3 monoclonal antibody (mAb) into mice caused DNAfragmentation in the intestinal villous epithelial cells(IVECs) of the duodenum and the jejunum. In this study,in order to elucidate the mechanism of DNA fragmentationin IVECs, we searched for the inducer(s) of DNA fragmen-tation by using immunohistochemistry. The release ofcytoplasmic granules from intraepithelial lymphocytes(IELs) and the formation of large gaps between IELsand IVECs were observed electron microscopically afterantibody administration. The presence and distributionpattern of Granzyme B (GrB), a serine protease in cytolyticgranules present in cytotoxic T lymphocytes and naturalkiller cells and considered to be the responsible moleculefor DNA fragmentation in target cells, was examined indetail in intestinal villi by immunohistology. GrB was
detected in cytoplasmic granules in nearly all IELs. Thetime-kinetics of granule release from IELs after mAbinjection coincided not only with that of the extracellulardiffusion of GrB, but also with that of DNA fragmentationin IVECs. On the other hand, perforin (Pfn), assumed tocooperate with GrB in DNA fragmentation, could not bedetected in IELs, and its release was not confirmed after theanti-CD3 mAb injection. Anti-CD3 mAb injection alsoinduced DNA fragmentation in IVECs in Pfn-knockoutmice. These results support the notion that DNA fragmentationin IVECs by the stimulated IELs in the present study is inducedby a mechanism involving GrB, but independent of Pfn.
Keywords Intraepithelial lymphocytes . DNAfragmentation . Perforin . GranzymeB .Mouse (female,C57BL/6)
Introduction
The mucosal surface of the gastrointestinal tract consists ofa single layer of epithelial cells that provide a protectivebarrier against pathogenic microbes and other macromole-cules. This defensive task of the intestine is based on twoessential constituents: the mucosal barrier and the localimmune system (i.e., the gut-associated lymphoid tissue;Kiyono and Fukuyama 2004; Moreto and Perez-Bosque2009; Thompson-Chagoyan et al. 2005). Several studieshave shown that mutually functional relationships existbetween the gut epithelium and the mucosal immune system(MacDonald and Spencer 1988, 1990).
A substantial fraction of differentiated T cells are presentin the vast epithelial surface of the body where they arebelieved to contribute to local immunosurveillance (Haydayet al. 2001; Mueller and Macpherson 2006). A distinct
This work was in part supported by a Grant-in-aid for ScientificResearch from the Ministry of Education, Science and Culture, Japan(20590181 to M.O. and 21590207 to T.I.).
M. Ogata (*) :Y. Ota : T. ItohDivision of Immunology and Embryology, Department of CellBiology, Tohoku University School of Medicine,980-8575 Sendai, Japane-mail: [email protected]
T. MatsutaniLaboratory of Immune Regulation, Wakayama Medical University,Osaka, Japan
M. NannoDepartment of Applied Research, Division of Immunology, YakultCentral Institute for Microbiological Research, Tokyo, Japan
R. SuzukiDepartment of Rheumatology and Clinical Immunology,Clinical Research Center for Allergy and Rheumatology,Sagamihara National Hospital, Sagamihara, Japan
Cell Tissue Res (2013) 352:287–300DOI 10.1007/s00441-012-1549-7

population of lymphocytes located between the intestinalvillous epithelial cells (IVECs) above the basementmembrane is composed of cells called intraepithelial lym-phocytes (IELs; Asarnow et al. 1988; Nanno et al.1994; Rocha et al. 1994; Shiohara et al. 1993; Suzukiet al. 2001). IELs are distributed throughout the epithe-lium that overlies small intestinal villi (average of oneIEL for every six to ten epithelial cells; Beagley andHusband 1998; Tamura et al. 2003). A large numberof γδ T-cell receptor-bearing IELs (γδ-IELs) inhabitthe intestinal epithelial barriers (Beagley et al. 1995;Hayday et al. 2001; Hayday 2000; Lefrancois 1991).Although rare in the circulation, γδ T cells are promi-nent at intestinal surface, where they are endowed witha number of properties that distinguish them from con-ventional T cells. γδ-IELs intercalate between epithelialcells and are thus ready to provide a first line ofdefense against environmental challenges (Chen 2002;Haas et al. 1993; Ismail et al. 2011; Roberts et al. 1996). In ourprevious studies, we have shown that the administration ofanti-CD3 monoclonal antibody (mAb) to a mouse causesDNA fragmentation in the IVECs of the duodenum and thejejunum, resulting in diarrhea accompanied by the detachmentof epithelial cells and leading to the shortening of the villi(Tamura et al. 2003; Yaguchi et al. 2004).
Although tumor necrosis factor α (TNFα) has beenfound to exert a profound effect on the detachment of theIVECs after the anti-CD3 mAb injection, its postulatedeffect on DNA fragmentation has not been confirmed inexperiments with knockout mice for TNF receptor (Yaguchiet al. 2004). We have also found that DNA fragmentation ofthe IVECs is repairable (i.e., the fragmented DNA is repaired)and that, in the nuclei of the IVECs, repair-related moleculescan be clearly detected by immunohistochemistry, withvirtually the same time-kinetics as that of the DNAfragmentation (Ogata et al. 2009). Hence, the DNA fragmen-tation itself does not directly lead to cell death; it is repairable,i.e., the cell is alive, even after DNA fragmentation.
Perforin (Pfn) and Granzyme B (GrB) have been consid-ered to induce DNA fragmentation in most cases of cytoly-sis in which natural killer (NK) cells and cytotoxic T cells(CTLs) are involved (Barry and Bleackley 2002; Choy et al.2005; Fehniger et al. 2007; Kagi et al. 1994; Lowin et al.1994; Shi et al. 1997; Trapani 2012; Trapani et al. 2000; Weiet al. 1998).
To elucidate the mechanism of DNA fragmentation inIVECs of the mouse small intestine, we have sought theinducer(s) of in vivo DNA fragmentation in this study, mainlyby immunohistochemistry. Although GrB has been detectedin intestinal IELs and is released with the same time-kineticsas those of DNA fragmentation when anti-CD3 mAb isinjected, Pfn has not been observed in IELs, and the releaseof Pfn is not induced, even after the anti-CD3 mAb injection.
In Pfn-knockout (Pfn-KO) mice, the anti-CD3 mAb injectioninduces DNA fragmentation in IVECs in an identical way tothat in wild-type (wt) mice, as shown in our previous studies.The lack of Pfn in the intestinal villi, especially in IELs, theabsence of the release of Pfn from IELs even after anti-CD3mAb stimulation, and finally, the induction of DNAfragmentation in IVECs of Pfn-KO mice, together stronglysuggest a novel Pfn-independent mechanism of DNAfragmentation in this mouse experimental system.
Materials and methods
Animals
Female C57BL/6 J and C57BL/6-Prf1tm1Sdz/J (Pfn-KO)mice (6–10 weeks) were obtained from Charles River(Yokohama, Japan) and Jackson Laboratory (Bar Harbor,Me., USA), respectively. Pregnant (at 9 days post coitum[dpc]) female C57BL/6 J mice were purchased from CLEAJapan (Tokyo, Japan). These mice were maintained underspecific pathogen-free conditions and were used accordingto the guidelines of the Institutional Animal Care and UseCommittee established at Tohoku University. Animals hadaccess to food and water ad libitum and were kept under a12-h light–dark cycle.
Administration of anti-CD3 mAb for activation of IELs
For stimulation of IELs, mice were injected i.p. with ham-ster anti-mouse CD3 (145-2C11) mAb at a dose of 200 μgper mouse. Mice were killed at various times after theinjection, and the jejunum was dissected according to thedefinition of the small intestinal regions as described else-where for histology and immunohistochemistry (Tamura etal. 2003; Yaguchi et al. 2004).
Tissue preparation of the jejunum
For each tissue sampling, at least four mice were killed bycervical dislocation under anesthesia, and the small intestinewas immediately dissected. Several pieces (each 3–4 mm inlength) of the jejunum from four mice were pooled for lateruse. Since TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling; Ogata et al. 2009),immunostaining with antibodies to GrB (Guerriero et al.2011) and to Pfn (Hu and Cross 2011; Kuhla et al. 2009),and the staining of nuclei with 4′,6′-diamidino-2-phenyl-indole dihydrochloride (DAPI, 0.1 μg/ml, Sigma-Aldrich;Imanguli et al. 2009) have been reported as being possibleon ordinarily prepared paraffin sections, for all stainingprocedures used in this study, specimens were fixed with4 % formaldehyde for 4 h at 4 °C, washed three times in
288 Cell Tissue Res (2013) 352:287–300

phosphate-buffered saline (PBS), subjected to the regularprocessing protocol for light microscopy, and finally em-bedded in paraffin. For immunohistology, pooled segmentswere chosen randomly, and serial sections were made fromsamples of distinct regions.
Images of stained sections were captured by using a ProvisAX80TR microscope equipped with an DP71 digital cameraand detected by using a DP manager system (Olympus,Tokyo, Japan).
Tissue preparation of placenta
For preparation of samples of the placenta, pregnant femaleC57BL/6 J mice (at 12 dpc) were killed by cervical dislocationunder anesthesia. Placentas were separated into two halves,which were then fixed in 4 % formaldehyde for paraffinembedding. Paraffin sections (4 μm thick) were prepared bycutting transversely through the central region of the placenta.
TUNEL method
DNA fragmentation was detected according to the methoddescribed by Gavrieli et al. (1992). In brief, paraffin sections(4 μm thick), after deparaffination, rehydration, and washes,were incubated with a reaction medium for 60 min at 37 °C.The medium consisted of 100 mM sodium cacodylate bufferat pH 7.0, 1 mM CoCl2, 50 mg/ml gelatin, 10 nM biotin-16-dUTP (Boehringer Mannheim Biochemicals, Mannheim,Germany), and 100 U/ml terminal deoxynucleotidyl trans-ferase (TdT; Takara, Osaka, Japan; Nakamura et al. 1995).After being washed, the sections were incubated with anavidin-biotin-complex by using the Vectastain-ABC kit(Vector Laboratories, Burlingame, Calif., USA) for30 min. The sections were washed, incubated with 3-3′diaminobenzidine tetrahydrochloride (DAB; Wako, Osaka,Japan) solution, and counterstained with methylgreen. Forfluorescent TUNEL staining, sections were incubated withTUNEL staining buffer containing TdT and biotin-16-dUTPas stated above. Biotin-16-dUTP incorporated into DNAwas reacted with streptavidin-Alexa Fluor 488 (1:100,Jackson Immunoresearch) for 30 min. Following washesin PBS, all tissue sections were counterstained with 3 minwith DAPI (0.1 μg/ml, Sigma-Aldrich). Sections were fur-ther rinsed with PBS and mounted in Vectashield mountingmedium (Vector Laboratories, Burlingame, Calif., USA).
All images were captured with a charge-coupled device(CCD) camera (DP-71; Olympus, Tokyo) mounted on anOlympus AX80TR epifluorescence microscope (Olympus).
Transmission electron microscopy
Before or after the administration of anti-CD3 mAb, thespecimens (jejunum) were fixed in 2 % formaldehyde and
2.5 % glutaraldehyde in PBS for 2 h, washed with PBS, andpost-fixed in 2 % osmium textroxide (OsO4) for 2 h. Allsamples were washed twice in PBS, dehydrated in ascend-ing grades of ethanol, and embedded in TAAB Epon 812(TAAB, Reading, UK). Serial sections (70 nm in thickness)were cut on an Ultracut UCT ultramicrotome (Leica Micro-systems, Wetzlar, Germany). After being stained with uranylacetate and lead citrate, they were examined on a JEM 1400transmission electron microscope (Tokyo, Japan) operatedat 80 kV. Digital images were collected by means of a JeolCCD camera.
Immunohistochemical staining of cytotoxic factors
For light-microscopic observations, paraffin sections wereincubated with 10 % normal donkey serum for 30 min (forthe blocking of the second antibody [Ab]), goat anti-GrBpolyclonal Ab (R&D Systems, Minneapolis, Minn., USA),rabbit anti-Pfn polyclonal Ab (Cell Signaling, Danvers,Mass., USA), or rat anti-Pfn mAb (CB5.4, Fitzgerald Indus-tries, Concord, Mass., USA) for 24 h at 4 °C. At the sametime, relevant serial sections incubated with non-immune goatserum (Invitrogen, Calif., USA), non-immune rabbit immu-noglobulin (Dako, Carpinteria, Calif., USA), or isotype-matched irrelevant monoclonal rat IgG (eBioscience, Calif.,USA), instead of the primary antibodies but at the sameconcentrations as those of the corresponding primary anti-bodies, served as negative controls. The sections were thenincubated with biotinylated donkey anti-goat, rabbit, or ratIgG (Jackson ImmunoResearch, West Grove, Pa., USA)for 1 h. Immunoreaction was visualized with DAB, andthe sections were counterstained with methylgreen.
For immunofluorescence, after the blocking step, sec-tions immunoreacted with goat anti-GrB polyclonal Ab,rabbit anti-Pfn polyclonal Ab, or rat anti-Pfn mAb wereincubated with Alexa-Fluor-488-coupled donkey-antigoat, rabbit, or rat IgG (Molecular Probes, Eugene,Ore., USA) for 1 h. Negative controls were preparedas above. Finally, slides were counterstained with DAPIand mounted in Vectashield mounting medium (VectorLaboratories).
To examine whether the two cytotoxic factors co-localized within the same IEL, double-labeling was per-formed with the polyclonal or monoclonal anti-Pfn andthe anti-GrB Abs. Briefly, after deparaffination and re-hydration, all slides were incubated with a blockingreagent. Sections were then incubated for 24 h in acocktail of goat anti-GrB polyclonal Ab, rabbit anti-Pfn polyclonal Ab, or rat anti-Pfn mAb, followed byseveral washes in PBS. Sections were then incubated for1 h in a cocktail of Alexa-Fluor-594-coupled donkey anti-goatIgG and Alexa-Fluor-488-coupled donkey anti-rabbit IgGor rat IgG.
Cell Tissue Res (2013) 352:287–300 289

Preparation of jejunum for immunoelectron microscopy
Tissue samples were fixed with 4 % formaldehyde for 4 h at4 °C, washed with PBS, and immersed in 5 % sucrose/PBSfor 1 h, in 15 % sucrose/PBS for 3 h, and in 30 % sucrose/PBS overnight at 4 °C. Subsequently, the samples wereembedded in Tissue-Tek OCT compound 4583 (SakuraFinetek, Tokyo, Japan) and quick-frozen in a mixture ofacetone and dry ice. Frozen sections (10 μm) were cut in acryostat and affixed to Matsunami adhesive silane (MAS)-coated glass slides (Matsunami, Osaka, Japan). Immuno-staining of GrB was performed as described above.
Immunoreaction of GrB was visualized by the ABC meth-od with DAB. The sections were then washed, postosmicated(1 % OsO4 for 1 h), and dehydrated in ascending grades ofethanol. Once the areas in which GrB-positive IELs werepresent were identified, the sections were covered with a dropof 100 % Epon 812. A gelatin capsule filled with Epon 812was then positioned over the areas of interest. After polymer-ization in a 60 °C oven for 48 h, the glass slides were removed,and ultrathin sections were cut on a Ultracut UCT ultramicro-tome (Leica Microsystems). Micrographs were captured at3000× with a Jeol JEM-1400 transmission electron micro-scope equipped with a Jeol CCD camera.
Multiple-color fluorescence imaging
Double- and triple-immunostained sections were examinedwith an Olympus AX80TR epifluorescence microscope(Olympus) by using the appropriate filter-set for the variousfluorochrome characteristics.
Digital images from the different excitation wavelengthswere captured with a DP71 digital camera (Olympus) andco-registered (produced) by using Adobe Photoshop CS5(Adobe Systems) to combine multiple colors into a singlefinal image (a merged image) showing the overlapping ofmultiple fluorescent colors.
Results
DNA fragmentation in jejunal IVECs after treatment with anti-CD3 mAb in C57BL/6 mice
Without stimulation of IEL by anti-CD3 mAb, TUNELstaining detected only a few positive cells at the tip of villi(Fig. 1a). As early as 10 min after the injection of anti-CD3mAb, TUNEL-positive cells became noticeable and in-creased in number thereafter untill 20 min after the mAbinjection, when virtually all IVECs were TUNEL-positive(Fig. 1b–d).
By 1 h after the injection of anti-CD3 mAb, however,most IVECs became TUNEL-negative (Fig. 1e), and 2 h
later, all the remaining enterocytes in each shortened villusremained TUNEL-negative (Fig. 1f). However, most de-tached enterocytes were found to be TUNEL-positive, prob-ably because of degradation process progressing in theintestinal lumen, which is rich in various digestive enzymes.
Although DNA fragmentation in IVECs of C57BL/6mice occurred a little earlier than that of Balb/c mice (Ogataet al. 2009; by 10–20 min), the reactions attributable to anti-CD3 mAb injection were essentially the same in their extentand magnitude in both strains of mice.
Transmission electron microscopy of enterocytesafter treatment with anti-CD3 mAb
Before mAb injection, the jejunal IELs tightly faced adja-cent enterocytes, and many granules were observed in thecytoplasm of the IELs (Fig. 2a, d). By 20 min after IELstimulation, large gaps began to form between the IELs andenterocytes, and consequently, a substance of a granularnature, which appeared to be released from IEL, was ob-served in the interstice between the IELs and enterocytes(Fig. 2b, e). On the other hand, the granules rapidly disap-peared from the cytoplasm of the IELs. Sixty minutes later,the gaps extended greatly, and the joining regions of epithe-lial cells and the basal membrane appeared to be mostlydestroyed (Fig. 2c, f).
Immunohistochemical detection of GrB in IELs
Sections stained with an irrelevant goat IgG did not showany immunoreactivity (Fig. 3a, d, e). Most IELs presentbetween the enterocytes showed positive immunoreactionfor GrB (Fig. 3b, c). The GrB was discerned in the cyto-plasm as a dot-pattern immunoreaction (Fig. 3f–i), suggest-ing that GrB was contained within the granules of IELsobserved by transmission electron microscopy (see Fig. 2).
Release of GrB from IEL after treatment with anti-CD3mAb: examination at the light microscopic level
GrB was observed only in the cytoplasm, especially withinthe granules, of IELs before mAb injection (Fig. 4a, f, k, p).At 10 min after mAb injection, immunoreaction to GrBexpanded and diffused into the gaps newly formed betweenenterocytes and IELs (Fig. 4b, g, l, q; see also Fig. 2). Theexpansion and diffusion of immunoreaction of GrB towardthe gaps suggested that GrB was released extracellularlyfrom the granules of IELs. The release of GrB into theextracellular spaces peaked at 20 min (Fig. 4c, h, m, r),and the immune reaction to GrB was detected until 30 min(Fig. 4d, i, n, s). Diffusion of GrB was no longer detected at1 h, and its weak signal was barely detected in the cytoplasmof only a few IELs (Fig. 4e, j, o, t).
290 Cell Tissue Res (2013) 352:287–300

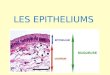
Release of GrB from IEL after treatment with anti-CD3mAb: examination at the electron-microscopic level
The immunoreaction products of GrB were observed exclu-sively within the cytoplasm of IEL before mAb injection(Fig. 5a, b). By 20 min after anti-CD3 mAb injection,however, the reaction products were detected not only with-in the cytoplasm, but also outside the IEL, scattering towardthe gap newly formed after mAb injection (Fig. 5c, d).
Immunohistochemical examination of GrB and Pfnexpression in placenta and jejunal villus
Many NK cells, which are characterized by their secretion ofcytotoxic factors, Pfn, and GrB (Burnett and Hunt 2000; King
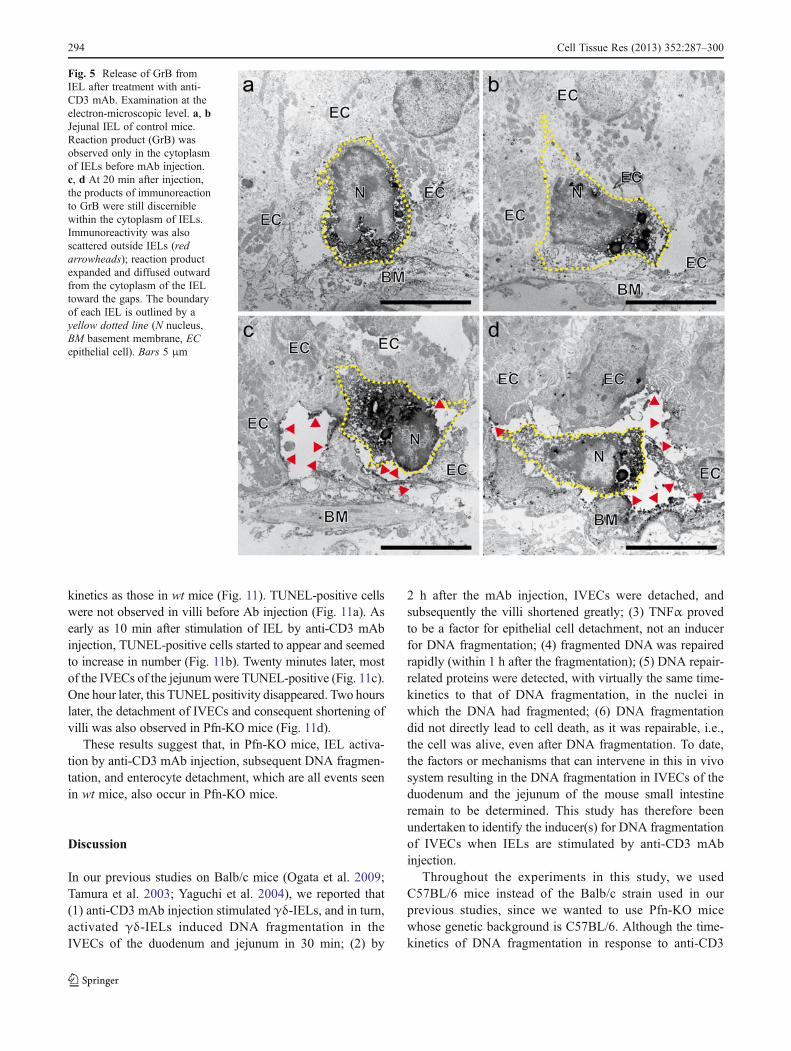
et al. 1993; Kusakabe et al. 1999; Nakashima et al. 2008;Tayade et al. 2005), can be identified in the uterus of pregnantmouse (uterine NK [uNK] ) by periodic acid-Schiff (PAS)staining (Burnett and Hunt 2000). As shown by immunohis-tochemical analyses, uNK cells in the decidua basalis regionclearly showed positive reaction to Pfn in their cytoplasm(Fig. 6e-h, j, k, m, n). On double-staining for GrB (red) andPfn (green), the co-localization of both factors was evidencedby merged immunostaining (yellow) in the uNK cells in theplacenta.
On the contrary, immunostaining of Pfn was completelynegative in the jejunal villus (Fig. 7); Pfn was observed inneither the epithelial layer nor the lamina propria of the mucousmembrane (Fig. 7a-d, f, g, i, j). Only GrB was detected in IELs(Fig. 7e, g, h, j), as also shown in Figs. 3, 4. No merged
Fig. 1 TUNEL (terminal deoxynucleotidyl transferase-mediateddUTP nick-end labeling) staining of jejunal epithelium after i.p. injec-tion of anti-CD3 monoclonal antibody (mAb; 200 μg/mouse). Jejunumof control mice given phosphate-buffered saline (PBS) alone (a).Jejunum from mice killed at 10 min (b), 20 min (c), 30 min (d),60 min (e), and 120 min (f) after anti-CD3 mAb injection. Sectionswere processed for TUNEL assay (3-3′ diaminobenzidine tetrahydro-chloride [DAB] staining). Only one or two enterocytes at the tip of thevilli were TUNEL-positive in the control jejunum (a). At 10 min after
treatment, TUNEL-positive cells began to increase in number (b), andat 20–30 min, most intestinal villous epithelial cells (IVECs) showedpositive reaction to TUNEL staining (c, d). By 60 min after treatment,however, most IVECs of the jejunum became TUNEL-negative (e).Two hours later, the length of villi shortened to about half that of thenormal villi (f). No TUNEL-positive enterocytes remained in the villi,and a large number of enterocytes could be seen in the lumen as a resultof considerable exfoliation of epithelial cells from the villi into thelumen. Bars 100 μm
Cell Tissue Res (2013) 352:287–300 291

Fig. 2 Transmission electronmicrographs of intestinal villousepithelial cells (IVECs) and intra-epithelial lymphocytes (IELs).Control (a, d). Jejunal mucosa at20 min (b, e) and 60 min (c, f)after antibody treatment (N nu-cleus,M mitochondrion, BMbasement membrane, G granule,EC epithelial cell). An imagefrom control mice (a) showstypical IVECs and an IEL at thebasement membrane. The IELcontains several distinct granulesin its cytoplasm (a, d). Remark-ably extensive gaps betweenneighboring enterocytes were ob-served when examined at 20 minafter the stimulation of IEL byanti-CD3 antibody injection, andgranular substance was observedin the gaps (b, e). At 60 min, gapsconsisted of large holes at thebasal zone, and granule-likestructures were still observed inthe gaps (c, f). Bars 5 μm (a-c),1 μm (d-f)
Fig. 3 Immunohistochemical staining of Granzyme B (GrB) in jejunalIELs. a, d, e Controls. An irrelevant goat IgG was used instead of theprimary antibody. A number of IELs were observed between the enter-ocytes, and most of them showed positive immunoreaction for GrB (b,
c). GrB immunoreactivity was identified in the cytoplasm as a dot-pattern immunoreaction (f–i). Pairs of boxed regions in a–c are shownat higher magnification in d–i, respectively. Bars 100 μm (a-c), 10 μm(d-i)
292 Cell Tissue Res (2013) 352:287–300

immunofluorescent images could be obtainedwith Pfn andGrBdouble-staining. Furthermore, no immunoreaction for Pfn wasdetected in the jejunal villus after mAb injection (Fig. 8); therelease of Pfn could not be confirmed.
Immunohistochemical analysis of GrB expressionin the jejunal villi of Pfn-KO mice
Numerous IELs were present in the jejunal villus of thePfn-KO mouse, and most of them showed positive
immunoreaction for GrB (Fig. 9a, b). Immunostaining forGrB was detected in the cytoplasm as a dot-pattern immuno-reaction (Fig. 9c–f). Release of GrB from IEL into theextracellular region was also observed by administration ofAb to Pfn-KO mice (Fig. 10) and to wt mice (see Fig. 4).
DNA fragmentation and subsequent events in Pfn-KO mice
TUNEL analysis demonstrated intense DNA fragmentation injejunal IVECs of Pfn-KO mice with mostly the same time-
Fig. 4 Release of GrB from IEL after treatment with anti-CD3 mAb.Examination at the light-microscopic level. Jejunum of control mice (a, f).Jejunum at (b, g) 10 min, (c, h) 20 min, (d, i) 30 min, and (e, j) 60min afteranti-CD3 mAb injection. At 10 min after mAb injection, the products ofimmunoreaction to GrB diffused outward from the cytoplasm of the IEL.Outward diffusion of GrB peaked at 20 min, although diffusion continued
till 30 min after anti-CD3 mAb injection. Diffusion of GrB was no longerdetected at 60min, and only an extremely weak signal was detectable in thecytoplasm of a few of IEL. Boxed regions in a–j are shown at highermagnification in k–t, respectively. a-j Alexa Fluor 488 (green GrB), k-tAlexa Fluor 488 (greenGrB), DAPI (4′,6′-diamidino-2-phenylindole dihy-drochloride; blue nuclei). Bars 50 μm (a-j), 10 μm (k-t)
Cell Tissue Res (2013) 352:287–300 293
kinetics as those in wt mice (Fig. 11). TUNEL-positive cellswere not observed in villi before Ab injection (Fig. 11a). Asearly as 10 min after stimulation of IEL by anti-CD3 mAbinjection, TUNEL-positive cells started to appear and seemedto increase in number (Fig. 11b). Twenty minutes later, mostof the IVECs of the jejunumwere TUNEL-positive (Fig. 11c).One hour later, this TUNEL positivity disappeared. Two hourslater, the detachment of IVECs and consequent shortening ofvilli was also observed in Pfn-KO mice (Fig. 11d).
These results suggest that, in Pfn-KO mice, IEL activa-tion by anti-CD3 mAb injection, subsequent DNA fragmen-tation, and enterocyte detachment, which are all events seenin wt mice, also occur in Pfn-KO mice.
Discussion
In our previous studies on Balb/c mice (Ogata et al. 2009;Tamura et al. 2003; Yaguchi et al. 2004), we reported that(1) anti-CD3 mAb injection stimulated γδ-IELs, and in turn,activated γδ-IELs induced DNA fragmentation in theIVECs of the duodenum and jejunum in 30 min; (2) by
2 h after the mAb injection, IVECs were detached, andsubsequently the villi shortened greatly; (3) TNFα provedto be a factor for epithelial cell detachment, not an inducerfor DNA fragmentation; (4) fragmented DNA was repairedrapidly (within 1 h after the fragmentation); (5) DNA repair-related proteins were detected, with virtually the same time-kinetics to that of DNA fragmentation, in the nuclei inwhich the DNA had fragmented; (6) DNA fragmentationdid not directly lead to cell death, as it was repairable, i.e.,the cell was alive, even after DNA fragmentation. To date,the factors or mechanisms that can intervene in this in vivosystem resulting in the DNA fragmentation in IVECs of theduodenum and the jejunum of the mouse small intestineremain to be determined. This study has therefore beenundertaken to identify the inducer(s) for DNA fragmentationof IVECs when IELs are stimulated by anti-CD3 mAbinjection.
Throughout the experiments in this study, we usedC57BL/6 mice instead of the Balb/c strain used in ourprevious studies, since we wanted to use Pfn-KO micewhose genetic background is C57BL/6. Although the time-kinetics of DNA fragmentation in response to anti-CD3
Fig. 5 Release of GrB fromIEL after treatment with anti-CD3 mAb. Examination at theelectron-microscopic level. a, bJejunal IEL of control mice.Reaction product (GrB) wasobserved only in the cytoplasmof IELs before mAb injection.c, d At 20 min after injection,the products of immunoreactionto GrB were still discerniblewithin the cytoplasm of IELs.Immunoreactivity was alsoscattered outside IELs (redarrowheads); reaction productexpanded and diffused outwardfrom the cytoplasm of the IELtoward the gaps. The boundaryof each IEL is outlined by ayellow dotted line (N nucleus,BM basement membrane, ECepithelial cell). Bars 5 μm
294 Cell Tissue Res (2013) 352:287–300
mAb injection was found to be slightly faster than those ofBalb/c mice in our preliminary experiments, the first task ofthis study was to examine in detail whether the resultsobtained from C57BL/6 mice could be considered identicalto those from Balb/c mice. The findings that DNA fragmen-tation in IVECs (Fig. 1), epithelial cell detachment (Fig. 1),
repair of fragmented DNA (data not shown), and detectionpatterns of DNA-repair-related proteins (γ-H2AX,phospho-ATM substrate, and Nbs1; data not shown), all ofwhich we observed in Balb/c mice (Ogata et al. 2009), wereclearly reproduced, even in C57BL/6 mice, although withslightly faster time-kinetics, strongly suggested that
Fig. 6 Immunohistochemicalexamination of perforin (Pfn)and GrB expression in the pla-centa. a-h Single-staining withanti-Pfn Ab. a-d Stained onlywith irrelevant rabbit IgG (a, b)or rat IgG (c, d) followed by thesecond Ab staining (negativecontrols). e-h Stained with anti-Pfn Ab. e, f Stained with anti-Pfn polyclonal Ab alone. g,h Stained with anti-Pfn mAbalone. a, c, e, g Low magnifica-tion micrographs reveal thedecidua basalis region of theplacenta. Abundant Pfn-positivecells (uterine natural killer [uNK]cells) were present in the deciduabasalis. b, d, f, h Higher magni-fication micrographs show indi-vidual Pfn-positive uNK cells. i-n Double-staining with anti-GrBpolyclonal Ab conjugated withAlexa Fluor 594 (red in i, k, l, n)and anti-Pfn Ab conjugated withAlexa Fluor 488 (green poly-clonal Ab in j, k; green mono-clonal in m, n). Merged imageswith GrB staining (red) and Pfnstaining (green) are shown in k,n. Overlapping of dual fluores-cence colors (yellow) was con-firmed in most uNK cells (k, n),which were detected by eachsingle fluorescent color witheither anti-GrB (i, l) or anti-Pfn(j, m). Pfn and GrB appeared toco-localize in uNK cells in theimplantation site of the uterus.Bars 1 mm (a, c, e, g), 200 μm(b, d, f, h), 100 μm (i-n)
Cell Tissue Res (2013) 352:287–300 295

Fig. 7 Immunohistochemicalexamination of Pfn and GrBexpression in the jejunal villus.a-d Single-staining with anti-Pfn Ab. a, b Stained with anti-Pfn polyclonal Ab. c, d Stainedwith anti-Pfn mAb. No positivecells were detected in the jeju-nal villus. e-j Double-stainingwith anti-GrB polyclonal Abconjugated with Alexa Fluor594 (red) and anti-Pfn Ab con-jugated with Alexa Fluor 488(polyclonal or monoclonal;green). e, g, h, j Stained withanti-GrB polyclonal Ab (red).Positive cells were detected. f,g, i, j Stained with anti-Pfn Ab.No green fluorescence wasdetected (f, g polyclonal Ab; i, jmAb). g, j Merged images withGrB staining (red) and Pfnstaining (green). No overlap-ping of fluorescence colors wasdetectable in IELs of the jejunalvillus (g, j). Pfn and GrB didnot co-localize in IELs in thejejunal villus. Bars 1 mm (a, c),200 μm (b, d-j)
Fig. 8 Immunohistochemicaldetection of Pfn expression injejunal villi after the treatmentwith anti-CD3 mAb. Jejunumof control mice (a, f). Jejunalvilli from mice killed at 10 min(b, g), 20 min (c, h), 30 min(d, i), and 120 min (e, j) afteranti-CD3 mAb injection. a-eStained with anti-Pfn polyclonalAb. f-i Stained with anti-PfnmAb. No Pfn-positive IELs werepresent, and the release ofPfn from the IELs was notconfirmed. No Pfn-positive cellswere present in the laminapropria. Bars 200 μm
296 Cell Tissue Res (2013) 352:287–300
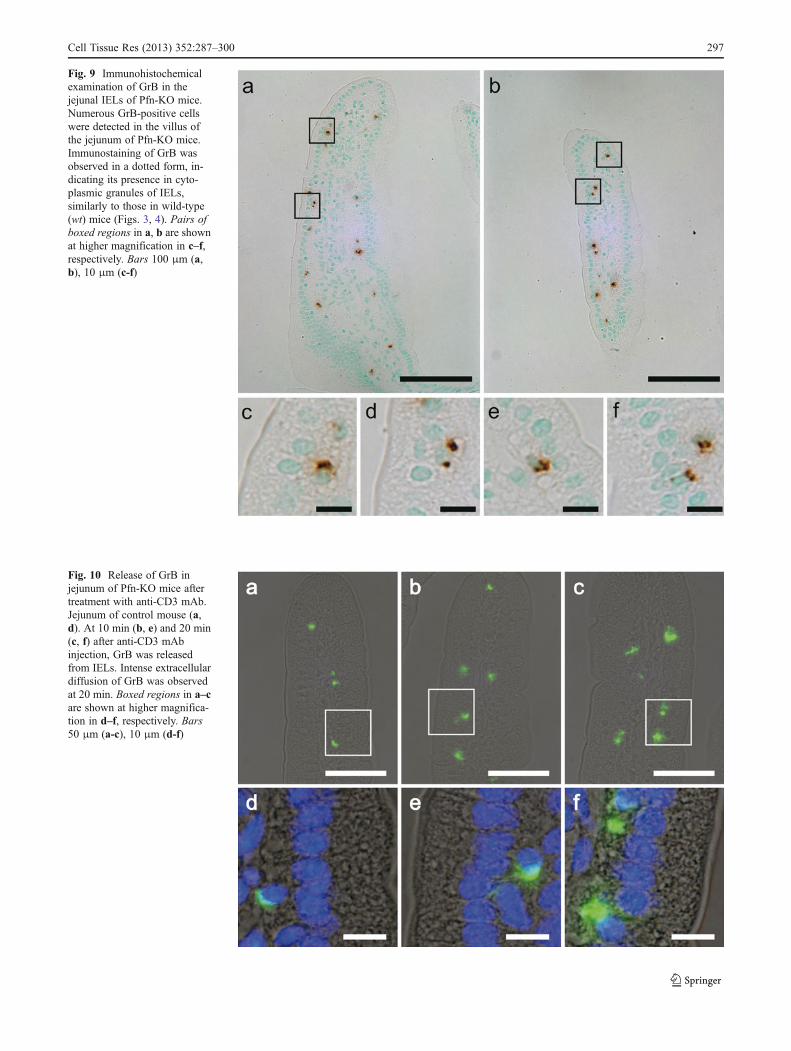
Fig. 9 Immunohistochemicalexamination of GrB in thejejunal IELs of Pfn-KO mice.Numerous GrB-positive cellswere detected in the villus ofthe jejunum of Pfn-KO mice.Immunostaining of GrB wasobserved in a dotted form, in-dicating its presence in cyto-plasmic granules of IELs,similarly to those in wild-type(wt) mice (Figs. 3, 4). Pairs ofboxed regions in a, b are shownat higher magnification in c–f,respectively. Bars 100 μm (a,b), 10 μm (c-f)
Fig. 10 Release of GrB injejunum of Pfn-KO mice aftertreatment with anti-CD3 mAb.Jejunum of control mouse (a,d). At 10 min (b, e) and 20 min(c, f) after anti-CD3 mAbinjection, GrB was releasedfrom IELs. Intense extracellulardiffusion of GrB was observedat 20 min. Boxed regions in a–care shown at higher magnifica-tion in d–f, respectively. Bars50 μm (a-c), 10 μm (d-f)
Cell Tissue Res (2013) 352:287–300 297

essentially the same responses were induced by the anti-CD3 mAb injection in both strains of mice.
Since the factors capable of exerting the cytotoxic effectthat causes DNA fragmentation were previously reported tobe Pfn and GrB in NK cells and CTLs (Barry and Bleackley2002; Choy et al. 2005; Fehniger et al. 2007; Kagi et al.1994; Lowin et al. 1994; Shi et al. 1997; Trapani 2012;Trapani et al. 2000; Wei et al. 1998), and since IELs in ourmouse system were found to have distinct granules in theircytoplasm that could be released extracellularly by the anti-CD3 mAb injection (Fig. 2), we considered it mandatory toexamine whether these factors (Pfn and GrB) were presentin granules of IELs. The polyclonal Ab to GrB gave apositive reaction in the granules of IELs by enzyme immu-nohistology (Fig. 3), in accord with other reports onthe presence of GrB-positive IELs (Muller et al. 2000;Yamazaki et al. 2005). The observations that the numberof GrB-positive cells was nearly the same as that of IELs(Tamura et al. 2003), and that GrB-positive cells werelocated between epithelial cells on the basement membrane(Fig. 3), indicated GrB was present in IELs in C57BL/6mice. Moreover, the finding that GrB-positive reaction ex-panded and diffused toward the gaps that were newlyformed between epithelial cells and IELs, and that wereattributable to the stimulation of IELs by anti-CD3 mAb(Figs. 4, 5) strongly suggested that granules together withtheir contents were released extracellularly. Pfn-positivecells, however, could not be found in the intestinal villi inthis study (Figs. 7, 8).
Positive controls for staining with both anti-GrB and anti-Pfn Abs were necessary. Sections were thus taken from theplacenta in which the presence of NK cells have beenrepeatedly reported (Burnett and Hunt 2000; King et al.1993; Kusakabe et al. 1999; Nakashima et al. 2008; Tayadeet al. 2005) in order to confirm whether the methodologyapplied in this study was appropriate. Both Abs used in this
study (to GrB and to Pfn) gave, by enzyme and fluorescentimmunohistology, positive reactions in NK cells of theplacenta (the decidua basalis region; Fig. 6). Double-staining with these Abs clearly delineated the co-localization of both factors within single placental NK cells(Fig. 6k, n). Thus, the staining procedures including the Absused were applicable for the detection of both GrB and Pfn,and additionally, the results presented here confirmed thatplacental NK cells were equipped with both cytotoxic fac-tors. With this staining technique, GrB-positive IELs wereonce again discernible, as previously shown in Fig. 3,whereas Pfn positivity was never detected in the villi(Fig. 7a-d, f, i). Co-localization of Pfn with GrB was alsonot confirmed (Fig. 7g, j). Pfn was not released, even afterstimulation with anti-CD3 mAb (Fig. 8).
Since, so far, we could not detect Pfn in IELs of themouse jejunal villi, we strongly hypothesized that DNAfragmentation caused by the activated IELs might be Pfn-independent. To examine whether Pfn is necessary in this invivo experimental system, essentially the same series ofexperiments were carried out in Pfn-KO mice. The resultsshown in Figs. 9, 10 demonstrated that, in Pfn-KO mice,GrB-positive IELs were present in the jejunal villi, and that,upon activation, GrB was released extracellularly with theidentical time-kinetics to that of wt mice. Finally, we alsoused Pfn-KO mice to determine whether DNA fragmen-tation and subsequent changes could be induced by thetreatment of anti-CD3 mAb. DNA of IVECs was frag-mented, and epithelial cells detached from the villi, withvirtually the same time-kinetics to that demonstrated inwt mice (Fig. 11).
The results presented in this study strongly argue for a Pfn-independent mechanism for DNA fragmentation by granule-containing IELs. Although several studies have previouslydemonstrated that GrB can exert cytotoxicity by CTLs in aPfn-independent manner (Chamberlain et al. 2010; Choy et al.
Fig. 11 DNA fragmentation in jejunal IVECs of Pfn-KO mice afterthe treatment with anti-CD3 mAb. Sections were processed for fluo-rescent TUNEL assay. Jejunum of control mice given PBS alone (a).Jejunum from mice killed at 10 min (b), 20 min (c), and 120 min (d)after anti-CD3 mAb injection. At 10 min after Ab injection, TUNEL-positive IVECs began to appear (b). By 20 min after Ab injection,
DNA fragmentation was induced in most jejunal IVECs of the Pfn-KOmice (c). The TUNEL-positive cells were no longer observed 120 minlater in the jejunal villus (d), and at this time, the villi appeared to haveshortened their length substantially (d). Alexa Fluor 488 (greenTUNEL-positive cells), DAPI (blue nuclei). Bars 200 μm
298 Cell Tissue Res (2013) 352:287–300

2004; Dressel et al. 2004; Lee et al. 2004), only a few reportshave described DNA fragmentation in target cells withGrB in the absence of Pfn (Froelich et al. 1996; Motykaet al. 2000). These reports, together with our currentstudy, consequently indicate a cytotoxic mechanism de-pendent on GrB, but independent of Pfn. Efforts shouldtherefore urgently be directed at searching for the pos-sible alternate factor, used instead of Pfn, for the cyto-toxicity that is exerted by GrB and that can result inDNA fragmentation in target cells. In particular, thefinding that the possible alternate factor in this study doesnot kill target cells (IVECs) is of special interests. GrBmust beincorporated by IVECs by a mechanism that is so farunknown.
Another point to note is that, IELs, delineated as largegranular lymphocytes (LGL) as shown in this study (Fig. 2),should be recognized as a distinct type different from typicalLGL, such as conventional CTLs and NK cells (Liu et al.1995; Ojcius et al. 1991; Parr et al. 1990; Vollenweider andGroscurth 1991), as IELs do not need Pfn to induce DNAfragmentation in target IVECs. Whether Pfn-independentDNA fragmentation is an immune function unique to the localimmune system of the small intestine or is one a rather gener-alized immune system for IELs, is also unknown at present.Therefore, more work should be carried out to elucidate theimmune system operating in the small intestine, especially inthe duodenum and the jejunum, in terms of IEL function.
References
Asarnow DM, Kuziel WA, Bonyhadi M, Tigelaar RE, Tucker PW,Allison JP (1988) Limited diversity of gamma delta antigenreceptor genes of Thy-1+ dendritic epidermal cells. Cell55:837–847
Barry M, Bleackley RC (2002) Cytotoxic T lymphocytes: all roadslead to death. Nat Rev Immunol 2:401–409
Beagley KW, Husband AJ (1998) Intraepithelial lymphocytes: origins,distribution, and function. Crit Rev Immunol 18:237–254
Beagley KW, Fujihashi K, Lagoo AS, Lagoo-Deenadaylan S, BlackCA, Murray AM, Sharmanov AT, Yamamoto M, McGhee JR,Elson CO et al (1995) Differences in intraepithelial lymphocyte Tcell subsets isolated from murine small versus large intestine. JImmunol 154:5611–5619
Burnett TG, Hunt JS (2000) Nitric oxide synthase-2 and expression ofperforin in uterine NK cells. J Immunol 164:5245–5250
Chamberlain CM, Ang LS, Boivin WA, Cooper DM, Williams SJ,Zhao H, Hendel A, Folkesson M, Swedenborg J, Allard MF,McManus BM, Granville DJ (2010) Perforin-independent extra-cellular granzyme B activity contributes to abdominal aortic an-eurysm. Am J Pathol 176:1038–1049
Chen ZW (2002) Comparative biology of gamma delta T cells. SciProg 85:347–358
Choy JC, Hung VH, Hunter AL, Cheung PK, Motyka B, Goping IS,Sawchuk T, Bleackley RC, Podor TJ, McManus BM, GranvilleDJ (2004) Granzyme B induces smooth muscle cell apoptosis inthe absence of perforin: involvement of extracellular matrix deg-radation. Arterioscler Thromb Vasc Biol 24:2245–2250
Choy JC, Cruz RP, Kerjner A, Geisbrecht J, Sawchuk T, Fraser SA,Hudig D, Bleackley RC, Jirik FR, McManus BM, Granville DJ(2005) Granzyme B induces endothelial cell apoptosis and con-tributes to the development of transplant vascular disease. Am JTransplant 5:494–499
Dressel R, Raja SM, Honing S, Seidler T, Froelich CJ, von Figura K,Gunther E (2004) Granzyme-mediated cytotoxicity does not in-volve the mannose 6-phosphate receptors on target cells. J BiolChem 279:20200–20210
Fehniger TA, Cai SF, Cao X, Bredemeyer AJ, Presti RM, French AR,Ley TJ (2007) Acquisition of murine NK cell cytotoxicityrequires the translation of a pre-existing pool of granzyme Band perforin mRNAs. Immunity 26:798–811
Froelich CJ, Orth K, Turbov J, Seth P, Gottlieb R, Babior B, Shah GM,Bleackley RC, Dixit VM, Hanna W (1996) New paradigm forlymphocyte granule-mediated cytotoxicity. Target cells bind andinternalize granzyme B, but an endosomolytic agent is necessaryfor cytosolic delivery and subsequent apoptosis. J Biol Chem271:29073–29079
Gavrieli Y, Sherman Y, Ben-Sasson SA (1992) Identification ofprogrammed cell death in situ via specific labeling of nuclearDNA fragmentation. J Cell Biol 119:493–501
Guerriero JL, Ditsworth D, Catanzaro JM, Sabino G, Furie MB,Kew RR, Crawford HC, Zong WX (2011) DNA alkylatingtherapy induces tumor regression through an HMGB1-mediated activation of innate immunity. J Immunol 186:3517–3526
Haas W, Pereira P, Tonegawa S (1993) Gamma/delta cells. Annu RevImmunol 11:637–685
Hayday AC (2000) [Gamma][delta] cells: a right time and a right placefor a conserved third way of protection. Annu Rev Immunol18:975–1026
Hayday A, Theodoridis E, Ramsburg E, Shires J (2001) Intraepitheliallymphocytes: exploring the third way in immunology. Nat Immunol2:997–1003
Hu D, Cross JC (2011) Ablation of Tpbpa-positive trophoblast precur-sors leads to defects in maternal spiral artery remodeling in themouse placenta. Dev Biol 358:231–239
Imanguli MM, Swaim WD, League SC, Gress RE, Pavletic SZ, HakimFT (2009) Increased T-bet+ cytotoxic effectors and type Iinterferon-mediated processes in chronic graft-versus-host diseaseof the oral mucosa. Blood 113:3620–3630
Ismail AS, Severson KM, Vaishnava S, Behrendt CL, Yu X, BenjaminJL, Ruhn KA, Hou B, DeFranco AL, Yarovinsky F, Hooper LV(2011) Gamma delta intraepithelial lymphocytes are essentialmediators of host-microbial homeostasis at the intestinal mucosalsurface. Proc Natl Acad Sci USA 108:8743–8748
Kagi D, Ledermann B, Burki K, Seiler P, Odermatt B, Olsen KJ,Podack ER, Zinkernagel RM, Hengartner H (1994) Cytotoxicitymediated by T cells and natural killer cells is greatly impaired inperforin-deficient mice. Nature 369:31–37
King A, Wooding P, Gardner L, Loke YW (1993) Expression ofperforin, Granzyme A and TIA-1 by human uterine CD56+ NKcells implies they are activated and capable of effector functions.Hum Reprod 8:2061–2067
Kiyono H, Fukuyama S (2004) NALT- versus Peyer’s-patch-mediatedmucosal immunity. Nat Rev Immunol 4:699–710
Kuhla A, Eipel C, Abshagen K, Siebert N, Menger MD, Vollmar B(2009) Role of the perforin/granzyme cell death pathway in D-Gal/LPS-induced inflammatory liver injury. Am J Physiol Gastro-intest Liver Physiol 296:G1069–G1076
Kusakabe K, Okada T, Sasaki F, Kiso Y (1999) Cell death of uterinenatural killer cells in murine placenta during placentation andpreterm periods. J Vet Med Sci 61:1093–1100
Lee SH, Bar-Haim E, Machlenkin A, Goldberger O, Volovitz I, VadaiE, Tzehoval E, Eisenbach L (2004) In vivo rejection of tumor
Cell Tissue Res (2013) 352:287–300 299

cells dependent on CD8 cells that kill independently of perforinand FasL. Cancer Gene Ther 11:237–248
Lefrancois L (1991) Extrathymic differentiation of intraepithelial lym-phocytes: generation of a separate and unequal T-cell repertoire?Immunol Today 12:436–438
Liu CC, Walsh CM, Eto N, Clark WR, Young JD (1995) Morphologicand functional characterization of perforin-deficient lymphokine-activated killer cells. J Immunol 155:602–608
Lowin B, Hahne M, Mattmann C, Tschopp J (1994) Cytolytic T-cellcytotoxicity is mediated through perforin and Fas lytic pathways.Nature 370:650–652
MacDonald TT, Spencer J (1988) Evidence that activated mucosal Tcells play a role in the pathogenesis of enteropathy in humansmall intestine. J Exp Med 167:1341–1349
MacDonald TT, Spencer J (1990) Gut immunology. Baillieres ClinGastroenterol 4:291–313
Moreto M, Perez-Bosque A (2009) Dietary plasma proteins, the intes-tinal immune system, and the barrier functions of the intestinalmucosa. J Anim Sci 87:E92–E100
Motyka B, Korbutt G, Pinkoski MJ, Heibein JA, Caputo A, HobmanM, Barry M, Shostak I, Sawchuk T, Holmes CF, Gauldie J,Bleackley RC (2000) Mannose 6-phosphate/insulin-like growthfactor II receptor is a death receptor for Granzyme B duringcytotoxic T cell-induced apoptosis. Cell 103:491–500
Mueller C, Macpherson AJ (2006) Layers of mutualism with commensalbacteria protect us from intestinal inflammation. Gut 55:276–284
Muller S, Buhler-Jungo M, Mueller C (2000) Intestinal intraepitheliallymphocytes exert potent protective cytotoxic activity during anacute virus infection. J Immunol 164:1986–1994
Nakamura M, Yagi H, Kayaba S, Ishii T, Ohtsu S, Gotoh T, Itoh T(1995) Most thymocytes die in the absence of DNA fragmenta-tion. Arch Histol Cytol 58:249–256
Nakashima A, Shiozaki A, Myojo S, Ito M, Tatematsu M, Sakai M,Takamori Y, Ogawa K, Nagata K, Saito S (2008) Granulysinproduced by uterine natural killer cells induces apoptosis ofextravillous trophoblasts in spontaneous abortion. Am J Pathol173:653–664
Nanno M, Matsumoto S, Koike R, Miyasaka M, Kawaguchi M,Masuda T, Miyawaki S, Cai Z, Shimamura T, Fujiura Y et al(1994) Development of intestinal intraepithelial T lymphocytes isindependent of Peyer’s patches and lymph nodes in aly mutantmice. J Immunol 153:2014–2020
Ogata M, Oomori T, Soga H, Ota Y, Itoh A, Matsutani T, Nanno M,Suzuki R, Itoh T (2009) DNA repair after DNA fragmentation inmouse small intestinal epithelial cells. Cell Tissue Res 335:371–382
Ojcius DM, Zheng LM, Sphicas EC, Zychlinsky A, Young JD (1991)Subcellular localization of perforin and serine esterase inlymphokine-activated killer cells and cytotoxic T cells by immu-nogold labeling. J Immunol 146:4427–4432
Parr EL, Young LH, ParrMB, Young JD (1990) Granulated metrial glandcells of pregnant mouse uterus are natural killer-like cells thatcontain perforin and serine esterases. J Immunol 145:2365–2372
Roberts SJ, Smith AL, West AB, Wen L, Findly RC, Owen MJ, HaydayAC (1996) T-cell alpha beta + and gamma delta + deficient micedisplay abnormal but distinct phenotypes toward a natural, wide-spread infection of the intestinal epithelium. Proc Natl Acad SciUSA 93:11774–11779
Rocha B, Vassalli P, Guy-Grand D (1994) Thymic and extrathymicorigins of gut intraepithelial lymphocyte populations in mice. JExp Med 180:681–686
Shi L, Mai S, Israels S, Browne K, Trapani JA, Greenberg AH (1997)Granzyme B (GraB) autonomously crosses the cell membrane andperforin initiates apoptosis and GraB nuclear localization. J ExpMed 185:855–866
Shiohara T, Moriya N, Hayakawa J, Arahari K, Yagita H, NagashimaM, Ishikawa H (1993) Bone marrow-derived dendritic epidermalT cells express T cell receptor-alpha beta/CD3 and CD8. Evidencefor their extrathymic maturation. J Immunol 150:4323–4330
Suzuki H, Jeong KI, Doi K (2001) Regional variations in the distribu-tions of small intestinal intraepithelial lymphocytes (IELs) inBALB/c +/+, nu/+, and nu/nu mice. Comp Med 51:127–133
Tamura A, Soga H, Yaguchi K, Yamagishi M, Toyota T, Sato J, Oka Y,Itoh T (2003) Distribution of two types of lymphocytes (intra-epithelial and lamina-propria-associated) in the murine small in-testine. Cell Tissue Res 313:47–53
Tayade C, Fang Y, Black GP, Paffaro VA Jr, Erlebacher A, Croy BA(2005) Differential transcription of eomes and T-bet during mat-uration of mouse uterine natural killer cells. J Leukoc Biol78:1347–1355
Thompson-Chagoyan OC, Maldonado J, Gil A (2005) Aetiology ofinflammatory bowel disease (IBD): role of intestinal microbiotaand gut-associated lymphoid tissue immune response. Clin Nutr24:339–352
Trapani JA (2012) Granzymes, cytotoxic granules and cell death: theearly work of Dr. Jurg Tschopp. Cell Death Differ 19:21–27
Trapani JA, Davis J, Sutton VR, Smyth MJ (2000) Proapoptoticfunctions of cytotoxic lymphocyte granule constituents in vitroand in vivo. Curr Opin Immunol 12:323–329
Vollenweider I, Groscurth P (1991) Ultrastructure of cell mediatedcytotoxicity. Electron Microsc Rev 4:249–267
Wei S, Gamero AM, Liu JH, Daulton AA, Valkov NI, Trapani JA,Larner AC, Weber MJ, Djeu JY (1998) Control of lytic functionby mitogen-activated protein kinase/extracellular regulatory ki-nase 2 (ERK2) in a human natural killer cell line: identificationof perforin and Granzyme B mobilization by functional ERK2. JExp Med 187:1753–1765
Yaguchi K, Kayaba S, Soga H, Yamagishi M, Tamura A, Kasahara S,Ohara S, Satoh J, Oka Y, Toyota T, Itoh T (2004) DNA fragmen-tation and detachment of enterocytes induced by anti-CD3 mAb-activated intraepithelial lymphocytes. Cell Tissue Res 315:71–84
Yamazaki K, Shimada S, Kato-Nagaoka N, Soga H, Itoh T, Nanno M(2005) Accumulation of intestinal intraepithelial lymphocytes inassociation with lack of polymeric immunoglobulin receptor. EurJ Immunol 35:1211–1219
300 Cell Tissue Res (2013) 352:287–300