Embed Size (px)
Citation preview

GROWTH AND DIAGNOSTIC STUDIES
WITH NEISSERIA GONORRHOEAE
ROBERT LESLIE HOLLAND, B.S, IN E.E.
A THESIS
IN
MICROBIOLOGY
Submitted to the Graduate Faculty of Texas Tech University in Partial Fulfillment of the Requirements for
the Degree of
MASTER OF SCIENCE
Approved
// Accepted
August 1976

t
f)t ^û'i
T"5 í/^jL, l^O'l'H lOy"'^
ACKNOWLEDGEMENTS
I am greatly indebted to Professors Clsurence L.
Baugh, Jack Hayes and Jack Sevall for their tirae,
patience and helpful criticism in the direction and
preparation of this thesis.
ii

TABLE OF CONTENTS
ACKNOWLEDGEMENTS ii
LIST OF TABLES • . . . . . • iv
LIST OF FIGURES V
CHAPTER
I . INTRODUCTION • • • • • • • 1
I I . MATERIALS AND METHODS • • • • • 18
l . RESULTS • • • • • • • • • 25
IV. DISCUSSION 51
V SUMMARY • . . . . . . . . 57
LITERATURE CITED 60
APPENDIX . . . . . . . . . . . . . . 70
iii

LIST OF TABLES
Table Page
1« Sites of infection and local
and general complications • • • • • . • • • • 5
2. Components of Neicarb medium . • • 19
J. Semi-solid medium for initial growth studies . • • • • . . • • • . . . . . 2 0
^. Liquid medium for growth curve
studies . • • • • . • . • • . . . . . • • • • 2 1
5. Carbon/Energy sources studied • • • • • • . . 2 2
6. Growth on semi-solid media . • • . • • . . . 2 6
7. Temperature effects on growth on semi-solid media . • • • . . • • . • • • • 2 7
8. Survival time with different carbon/energy sources . • • . . . . . • • • • 2 9
9. Papanicalaou and gram stain comparison • • • • • • . • • . . . . • • • . 5 0
iv

LIST OF FIGURES
Page
Figure 1. Growth of clinical isolate (2957) of Neisseria gonorrhoeae in a liquid growth medium with varying concentrat-ions of V-C-N inhibitor . • 30
Figure 2. Growth of clinical isolates (3092 and 3698) of Neisseria gonorrhoeae in a liquid growth medium with varying concentrations of glucose • • . . • • • • 3 ^
Fig\ire 3. Growth of clinical isolates (3092 and 3698) of Neisseria gonorrhoeae in liquid growth medium with glucose, sucrose and lactate as carbon/energy source . . . . . . . . . . . . . . . . . 3 ^
Figure k» Growth of clinical isolates (3092, 3225 and 3^13) of Neisseria ^onorrhoeae in liquid growth medium with glucose, fructose, propionate, raffinose, succinate or no addition as carbon/ energy source • • . • • . . . . • • • • • 3 8
Figure 5. Growth of clinical isolates (3092, 3225 and 3^13) of Neisseria gonorrhoeae in liquid growth mediura with glucose, rham-nose, xylose, aurabinose, malonate or inositol as carbon/energy source . • . • 40
Figure 6. Growth of clinical isolates (3092, 3225 and 3413) of Neisseria gonorrhoeae in liquid growth medium with combinations of glucose or sucrose with lactate as carbon/energy source • • • • • • • • • • 4 2
Figure 7» Growth of clinical isolates (3092, 3225 and 3413) of Neisseria ^onorrhoeae in liquid growth medium with combmations of glucose v/ith sucrose and with prop-ionate as carbon/energy source • • • • • 44

Figure 8^ Growth of clinical isolate (76I6) of Neisseria gonorrhoeae in a liquid growth medium with glucose or the combinations glucose plus xylose, glucose plus succinate or glucose plus lactate plus succinate • • • • 46
Figure 9. Growth of clinical isolate (7676) of Neisseria gonorrhoeae in liquid growth medium with glucose or glucose plus fructose as carbon/energy source • • • • • • • 48
vi

CHAPTER I
INTRODUCTION
Objectives. The purpose of this thesis was
essentially two fold. The first objective was to examine
the in vitro growth requirements and characteristics of
Neisseria gonorrhoeae, the etiologic agent of gonorrhea.
In particular, potential carbon and energy sources were
explored. A variety of different compounds were tried as
carbon/energy sources, first in semi-solid media, then
in broth cultures. In addition, combinations of sources
were studied. The effects of temperature on the bacteria's
growth with the different carbon/energy sources was also
studied.
The intent of this portion of the research was
both to more fully characterize the organism's growth
characteristics and to suggest a better clinical medium
for longer term survivability such as is necesseiry for
diagnosis following weekend incubation in small labor-
atories. Also, the organism's sensitivity to the inhib-
itor V-C-N, utilized in most clinical media, was studied.
The survivability of organisms in frozen storage was
also investigated.
The second objective was to evaluate the practic-

ality of using the Papanicalaou Smear as a screening tool
for gonorrhea. Cervical smears were compared by using
both the Papanicalaou and Gram stains.
The Disease. Gonorrhea is an acute infectious
disease involving primarily the mucous membranes of the
genitourinairy tract, rectum, cervix and occasionally the
eye with possible hematogenic spread to serous and
synovial membranes in other parts of the body (47)•
Gonorrhea is a disease of both sexes which is usually
transmitted by sexual intercourse (9).
It is an exceptionally prevalcnt disease throughout
the world, with the annual number of new cases exceeding
60-65 million (14, 38). In the United States nearly a
million cases are reported each year^ with possibly two
or three times as many unreported (13» 79)» Sexual
distribution for reported cases of gonorrhea is about
40^ female and 60^ male with the majority of these cases
occuring in the 15-24 year group (13)»
In the male, gonorrhea usually remains localized
in the urethra and connecting structures. Within 24
hours a purulent urethral discharge begins. If the
infection remains untreated, in 10 to 14 days the patient
may begin to experience dysuria and perhaps strangury.
In severe attacks there may be a few drops of blood at
the end of micturition (14, 47).
The most common site of infection in the feraale is

3
the cervix uteri (14, 47)• Ninety to ninety-five per-
cent of female patients with gonorrhea heæbor organisms
in the columnar epithelium of the cervix (11, 58). Ureth«
ral infection invariably spreads to the emall urethral
glands but rarely to the bladder. The vaginal membrane
is usually resistant to infection, Sometimes the mucous
membranes of the vagina are reddened and bathed in pus
from which the gonococci can be recovered.
The cervix shows endocervicitis which often results
in the formation of infected retention cysts, the
follicles of Naboth, which may be seen on the vaginal
portion of the cervix. The infection probably extends
over the surface of the endometrium to the Fallopian
tubes, although lymphatic spread is possible. In the
puerperium, the infecting organism may spread directly
to the uterine wall at the site of placental attachment,
causing parametratis (inflammation of the cellular tissue
around the ligaments near the uterus).
Once the infection reaches the Fallopian tubes,
the organism invades the lining epithelium and spreads
to the Bubmucosa. The tubes can then fill with pus,
restricting and distending them. The presence of the
infecting organism and the pus can lead to a variety of
other pelvic coraplications, including peritonitis and
pelvic abscess. In spite of treatment, often the
pelvic infections have permanent residual effects.

The incubation period may vary from 2-10 days,
though it is often less than five days. Occasionally,
incubation periods of a month or more have been described
but this could be because the symptoms did not attract
the patient's attention.
Symptoms are absent or insufficient to attract
the patient's attention is up to 50-60^ of cases of
acute uncomplicated gonorrhea in the female (9, 14, 47,
56). Many of these women never seek treatment and are
often diagnosed only because their male contacts have
developed the disease. It is because of these cases
that a mass screening program or the incorporation of
gonorrhea testing into other routine work is essential.
The sites of infection and of local and general
complications are shown in Table 1.
Organism. Although first thought to be a symptom
of syphilis by Paracelsus (12), gonorrhea is caused by
Neisseria gonorrhoeae, first described by Albert Neisser
in 1879» He demonstrated the organism consistantly in ad-
ults and in smears from acute cases of vaginitis and
urethritis in adults and in smears from acute conjunctivit-
is of newboms (83). The organism was first cultivated by
Leistikow and Loeffler in 1882 (12) and three years later
by Bumm (14, 83). Bumm also successfully innoculated the
male urethra, producing characteristic symptoms and signs
of gonorrhea (14). The gonococcal complement-fixation test

TABLE 1
SITES OF INFECTION AND LOCAL AND GENERAL COMPLICATIONS^
Classification Sites of Infection
Local General Complications Complications
In Men
Anterior Urethra
Urethra
Posterior Urethra
Anorectum Conjunctivae Oropharynx
Tysonitis Paraurethr-al duct infection
Littritis Periurethr-al abscess Urethral stricture Cowperitis Prostatitis Prostatic abscess Vesiculitis Epididymitis Trigonitis
Arthritis Anterior uveitis Myocard-itis
Endocard-itis
Pericard-itis
In Women
In Children
Urethra
Cervix uteri
Anorectum Conjunctivae Oropharynx
Skenitis Bartholin-itis
Salpingo-oophoritis Parametritis Pelvic Abscess Pelvic peritonitis
Arthritis Anterior uveitis Myocard-itis
Endocard-itis
Mengitis
Vulva and Vagina Anorectum Conjunctivae Suppurative
panophthalraitis and ^ blindness * Data taken from King and Nicôl {i).

(GFT) was introduced by Muller and Oppenheim in I906.
Neisseria gonorrhoeae is a non-motile, non-spore-
forming, gram-negative coccus which grows in pairs or, on
occasion, in tetrads or clusters. The organisms are small
(ca. 0^8 by 0.6 micrometers) oval or spherical cocci with
flattened or slightly concave adjacent sides and resemble
a pair of kidney beans. They are parasites of the mucous
membranes of mammals (7). When observed in exudates or
cervical smeairs the organisms are often found inside poly-
morphonuclear leucocytes or associated with the surface
of polymorphs or other tissue, often in small matrix
arrangements (26, 75)«
The ultrastructure of Neisseria gonorrhoeae is
similar to other Neisseria species. The cell wall
contains a dense layer probably consisting of the peptid-
oglycan and an outer membrane. No capsule has been
convincingly demonstrated on gonococci. Fimbriae and
pili raay be present (71» 82). Pili appear:, to promote the
attachraent of virulent gonococci to huraan epithelial cells
and erythrocytes, enhancing the organisra's ability to \.
evade the host's defenses in tissue (6I).
Like most gram-negative bacteria, Neisseria
gonorrhoeae has an isoelectric point of about 5«0 (43).
An acidic change, greater than in most bacterial species,
occurs when iodine is added during Gram staining. This
change, attributed to the reaction of iodine with lipids

or reactive radicals such as the sulfhydryl groups of
proteins, probably accounts for some variability in the
decolorizing during Burke-Kopeloff-Beerman Gram staining
of laboratory cultures. The gonococcus can also be stain-
ed by polychrome stains such as Pappenheim-Saathoff methyl
green-pryonine (4). Eosin followed by methylene blue alone
yields good results. Intracellular granules can be seen in
stained preparations, but generally young cultures stain
evenly while older cultures (greater than 24 hours)
contain more involution forms which stain poorly.
On semi-solid media, colonies of gonococci are
small, round, convex, translucent, finely granular with
lobate margins and are greyish, with pearly opalescense
when viewed by transmitted light after 24-^8 hours incub-
ation.
After further incubation ajid serial passage, the
gonococci exhibit a number of colonial forms (32).
Kellog (35, 36) recognized four genetically determined
colonial types under standardized conditions. A trans-
parent medium with diffuse, angled light transmitted thr-
ough the medium from the bottora is necessary to observe
colonial characteristics. Type 1 colonies are small
(0.05 mm dia.), round, convex with entire edges, translu-
cent, dark colored, amorphous, slightly viscid and easily
emulsified. Type 2 colonies are similar to type 1, but
have sharper edges, are slightly crenated, have increased

8
intemal granularity, a friable consistancy, and are
better able to reflect light due to thicker reflective
surface film, Type 3 are larger (1 mm dia.), low convex,
flat edged, granular, viscid and brown colored. Type 4
colonies are similar but are amorphous and colorless.
Type 5 colonies have been described by Jephcott (32) to
be very shiny like type 2 but as big as a type 3. Easily
seen granules are present and they are brown in color.
Often a series of consecutive rings can be seen on the
surfaces. The edges aire clearly irregulaur and coarse.
Type 1 and Type 2 have been demonstrated to be virulent
orgsmisms, while the others appear avirulent (35» 36).
Five serological types have been demonstrated by
Hutchinson (30) in a manner similar to Lancefield's typ-
ing of streptococci but this method doesn't seem to have
much value beyond epidemiological tracings as subcultivat-
ion causes the loss of serological specificity.
Growth Characteristics. Neisseria gonorrhoeae is a
fastidious organism and is susceptible to raany toxic sub-
stances. Cultivation of the gonococcus on primary isolat-
ion is often difficult. The organism is fragile and does
not survive 24 hours in 12-79% of primary isolations (15,
28). Many types of transport and growth media have been
evaluated. Most of the existing knowledge of the organism's
metabolism has come from attempts to develop suitable
clinical media.

9
Neisseria gonorrhoeae can be grown on semi-solid
media with incubation temperatures between 30-38.5® C,
with optimura growth reported at 35-36® C (7)» The pres-
ence of moisture is important for good growth. Most
Neisseria gonorrhoeae strains require an atmosphere of
2-10% carbon dioxide or sraall quantities of suppleraental
bicarbonate for growth (24, 31, 72).
Early media were enriched by the addition of blood,
serum, ascitis and hydrocele fluid. Later modifications
included the use of peptone base, replacement of ascitis
fluid by yeast extract, the development of "chocolate
agar" to reraove some of the toxic blood products and the
use of purified agar (2, 8, 37). Other media were devel-
oped utilizing pea meal extract agar (21, 22), starch and
casamino acid (54), beef extract and heart infusion (51)»
and other growth enhancing factors (78).
Specific growth factors have been identified for
many strains. Glutamine and cocarboxylase (thiamine pyro-
phosphate) were found to be essential for growth in 10-15%
of the strains tested by Lankford and Snell (45) and
Lankford and Skaggs (44). Some strains were found to
require glutathione according to Gould (23). Griffin and
Racker (24) demonstrated a requirement for hypoxanthine
while showing that the use of yeast extract could be repl-
aced by a mixture of hypoxanthine, uracil and oxalacetate.
Boor (5) reported than some strains of Neisseria pjonorrhoeae

10
require a cystine concentration of 0.025-0.075%.
Many of these facts were linked together in the
development of chemically defined media. Kenny et al. (37)
utilized a medium using Medium 199 without bicarbonate
(GIBCO), cocarboxylase, L-glutamine, glucose, and ferric
chloride, Hunter and McVeigh (29) have developed one using
inorganic salts, glucose, guanine, L-cystine, L-isoleucine,
L-leucine, L-proline, L-threonine, L-valine, cytosine,
B-vitamin supplement, L-arginine and L-aspartic acid. They
also showed conventional agar to be inhibitory to growth
but purified agar was suitable. Similar media have been
developed by other workers (16).
Catlin (8) has presented a nutritional profile for
Neisseria gonorrhoeae with the ultimate goal of auxotyping
different strains. The mediura was similar to that of
Hunter and McVeigh above. Catlin also showed that all
strains tested grew in the absence of hemin and histidine,
asparagine, tryptophan, tyrosine and phenylalanine, threo-
nine, glutamine and spermine. In sorae strains, one or
more of them stimulated growth.
Elmros (15) has shown that low pH (5»0-6,0), Cu'*"*',
Mn**"*", Mg"'"'*', Ca**"*", and sucrose (10%) increases survivability
by preventing autolysis, although these cannot be consid-
ered true essentials for growth.
The present most coramonly used laboratory mediura was
developed by Thayer and Martin (48, 50, 73. 74). Their

11
medium was a modification of Christensen and Schoenlein's
(10) "chocolate agar" to which they added the antibiotics
vancomycin, nystatin, and colistin. The addition of these
antibiotics was the main advantage to the Thayer-Martin
medium.
Vancomycin inhibits the incorporation of amino acids
into the peptidoglycan and causes accumulation of UDP-Mur-
NAc-peptides in Gram-positive organisms (20, 66)^ inter-
fering directly with the release of the phospholipid carrier
preventing transport of new material during cell-wall
synthesis, Nystatin alters the permeability of the membr-
ane inhibiting the uptake of glycine (18). Nystatin is
active against yeasts, many fungi and other eucaryotic
cells. Colistin (polymyxin E) affects the membrane function
through interactions with the lipopolysaccharide comp-
onents of Gram-negative cell walls (19). Colistin is eff-
ective against Pseudomonas, Escherichia, Enterobacter,
Salmonella, Klebsiella, Hemophilus and Shigella species.
A slight modification of Thayer-Martin media,
Transgrow, is used in most diagnostic facilities for the
transport of primary cultures. Transgrow incorporates a
carbon dioxide supplemented atmosphere within the culture
bottle (49). It also has aji increased agar concentration
(1% to 2%), the glucose concentration was increased from
0.1% to 0.25%, and triraethoprim lactate is added to reduce
the growth of Proteus species.

ÍP'
12
Talley and Baugh (72) have improved on the Thayer-
Martin and Transgrow media in the development of Neicarb
which eliminates the carbon dioxide atmosphere by the
incorporation of sodium bicarbonate.
Metabolism. In all of these studies mentioned thus
far, the primary energy source has been glucose. Glucose
is catabolized aerobically by the Enter-Doudoroff and
Pentose Phosphate pathways (84% and 16% respectively) in
this organisra (53)» The Embden-Meyerhof-Pamas pathway
appears not to function. The metabolism of glucose app-
arently occurs in two steps. During active growth, glucose
is broken down to acetate and carbon dioxide. Following
glucose depletion, acetate is oxidized to TCA intermed-
iates. During this period little or no growth occurs.
According to Morse (52), only glucose, pyruvate,
and lactate are utilized efficiently as a source of energy
^y Neisseria gonorrhoeae. His studies make no mention,
though, of the compounds which were tested and found unsuit-
able as energy sources.
Tonkazy and Pelczar (76) showed, in 1953» that
araong the TCA cycle corapounds only those from -keto-
glutaric acid through pyruvate were oxidized in whole cell
experiments, although fumerate sometiraes showed a high
rate. They also showed that of all known amino acids only
the two forms of glutamic acid were oxidized rapidly.
Hebler and Morse (25) have since taken Neisseria gonorr-

^
13
hoeae grown in a glucose-lactate medium and, before
assaying for enzymes, allowed the cells to metabolize
acetate for a short time. In these cells, all of the
normal TCA enzymes were found. He also demonstrated that
glutamine entered the cycle at -ketoglutaric acid but
that this was not reversible.
Winter and Morse (81) have demonstrated several
non-heme iron centers, cytochrome c, two b cytochromes,
and cytochrome o as the terminal oxidase in the respiratory
chain of Neisseria gonorrhoeae.
Bergey's Manual (7) describes Neisseria gonorrhoeae
as able to grow sinaerobically although Williams and Wende
(80) and James-Holmquest et al. (31) were unable to
obtain growth under anaerobic conditions. In these studies,
an atmosphere high in hydrogen and carbon dioxide could
account for the lack of growth.
Diagnosis. Diagnosis of gonorrhea in the male is
based on history, clinical signs and the demonstration of
gram-negative diplococci, intracellular in white blood
cells in exudates from obvious lesions. In the female,
culture is normally necessary to elucidate the presence of
Neisseria Ronorrhoeae in addition to history and signs.
Cultures and smears are first examined microscopic-
ally for the presence of grara-negative, "coffee-bean"
shaped diplococci. There is some debate as to the efficacy
of using Gram's staining as a diagnostic test in addition

14
to clinical history and evidence in female gonorrhea.
There are many who will base a positive or negative diagn-
osis of gonorrhea on symptoras and direct microscopy only
(42, 69). On subsequent controls of therapeutic results,
the absence of symptoras in combination with a negative
direct smear has usually been considered sufficient to
declare the patient non-infectious (39, 42). Many consider
that although this procedure may be partly justified in
male patients, it has not been satisfactory in females (55,
59» 63). In fact, it has been highly unproductive (34, 64).
According to Juhlin (33)» the discrepancy between these
viewpoints is the reason why, lately, culture has been
little used for the confirmation of successful treatment
of gonorrhea.
Lodin ( 6) presented data in 1964 on 107 females
where direct microscopy was positive and culture negative
in 24.3%, Prasier (59) in 1968 conducted a study in which
he showed only 5% of the 211 diagnosed positive female
gonorrhea cases had positive gram-stained smears with
negative culture growth where 31% bad culture growth with
negative smears. In 1972, Phillips (60) studied 2557 cases
of female gonorrhea in all common sites and found only
8% where culture was negative and gram-staining indicated
positive. With 2 ^ of the cases, the organism could not
be identified microscopically with the gram stain but were
culture positive.

15
The difficulty with microscopic examinations is that
the gonococcus is either not present in smear preparations
or because of the variety of bacteria including Morexella
urethralis, Neisseria sicca and other gram-negative diplo-
cocci may be present in the female cervix creating a
microscopic picture resembling gonorrhea (57).
The use of fluorescent antibody techniques (FAT)
have proved valuable in diagnosis, especially in women,
though less sensitive than culture (65). Although rauch
has appeared in the literature in recent years, the use of
FAT has not increased significantly in clinical laboratories.
Further identification of Neisseria gonorrhoeae is
achieved through characteristics of the cultured organism,
presence of certain enzymes, sugar fermentation reactions
or fluorescent antibody staining. Neisseria gonorrhoeae
produce acid from glucose but not from maltose, fructose,
sucrose, or starch as opposed to other Neisseria which
present different fermentation pattems.
Neisseria gonorrhoeae and other Neisseria species
produce tho enzyme oxidase or indophenol oxidase which in
the presence of air acts on certain aromatic amines to
produce color changes. The color change, from pink to raag-
enta to purple or black, produced by an indicator such as
N, N, N', N'-tetramethyl-£-phenylenediamine dihydrochloride
(1%) is readily observed and used as a confirmatory test.

16
With federal funding for public health prograras
declining (1), support for routine culture of women in
public clinics is diminishing. For this reason, a simpler
and less expensive method of screening for gonorrhea is
necessary (3)» The center of screening programs will
probably be the asymptomatic female carrier, although there
are some asymptomatic males. At present, no true screening
test exists^
In developing better diagnostic tools, Lagerholm
and Lodin (41) analized the cytology of scrapings from
urethral smears in male gonorrhea patients. Ward sind
Watt (77) also examined the relationship between infecting
bacteria and mucosal cells. These studies and those by
Heller (25) showed that the organism is often found in
close association with the mucosal cells of the urethra
and vaginal duct. Ward and Watt (77) consider the adher-
ance to these cells to be directly related to virulence.
Since there is a close relationship between the
organism and mucosal tissue, iraproved diagnostic samples
are obtained by scraping the surface with an endometrical
currette. Shapiro (68) demonstrated this by inserting a
Novak endometrical currett into the cervical os to the
endocervix in non-pregnant women. Both by scraping and
deeper sampling he was able to increase his diagnostic
sensitivity by 13-20%.

17
Some VD clinics, in Great Britain, include screening
for cervical carcinoma as a routine in addition to gonorr-
hea testing (17). This screening is accomplished by taking
cervical scrapings with wooden spatulas, staining them by
the method of Papanicolaou, and examining them with a
standard light raicroscope (27). Heller (25) took advantage
of the simultaneous testing and the fact that metaplastic
and parabasal cells have been described in association
with gonorrhea infections ( O) to look for gonococci in
Papanicolaou smears. In this prelirainary study, about 70%
of the slides examined had raetaplastic cells present with
gonococci in a mosaic arrangement situated on a different
plane of focus from the nucleus of the cells. This would
suggest that the Papsinicolaou sraear could be used for
screening woraen for gonorrhea or the presence of the carrier
state during routine pelvic examinations. Since Heller's
work appears to be a promising screening tool, this study
further investigated its potential by comparing Gram and
Papanicolaou staining for the presence of gram-negative
intracellular diplococci in white blood cells.

CHAPTER II
MATERIALS AND METHODS
Cultures. Primary cultures of Neisseria gonorrhoeae
were obtained from clinical specimens from the Lubbock
(Texas) City-County Health Department. All cultures were
confirmed as Neisseria gonorrhoeae by Gram stain, positive
oxidase reaction and fermentation of glucose. Primary
cultures were transferred to either Neicarb (below) or
liquid culture medium and incubated at 35^ C for 24-it'tt hours.
Cultures for growth studies were taken from these first-
pass subcultivations.
Samples for the diagnostic studies were taken by
staff members at the Venereal Disease Clinic of the Lubbock
(Texas) City-County Health Department in accordance with a
mutually agreeable protocol (see Appendix). Tissue scrap-
ings were taken from female patients using established
procedures for cervical sampling for Papanicolaou smears.
Tissue from the cervix uteri was obtained with a wooden
"mutton-bone" spatula. Samples were then smeared on two
prelabelled glass slides. Two samples were taken and
altemately placed on slides for gram staining and for
Papanicolaou staining. Those to be stained by the method
of Papanicolaou were immediately sprayed with a cytology
fixative. Both were then refrigerated until staining.
18

19
Women selected for this sample were mostly positive
gonorrhea patients, confirmed by culture and clinical
observation.
Cultures for frozen storage studies were prepaured
by first transferring a suitable colony to 1 ml of liquid
growth medium and then treating it on a Vortex-Geni
(Model K-550-6) to disperse clumps. One-half milliliter of
this suspension was then added to 4 ml of Skim Milk Medium
(DIFCO) in one dram screw-cap vials and frozen at -90® C.
Growth Media. Cells were grown on a variety of media.
Neicarb was prepared in 1 or 2-ounce tightly capped pre-
scription bottles (Table 2).
TABLE 2
COMPONENTS OF NEICARB MEDIUM *
Component Amount/liter
GC Agar Base (DIFCO) 72.0 gm Hemoglobin 20.0 gm IsoVitaleX 10.0 ml V-C-N 10.0 ml NaHCO-j (7.5%) 30.0 ml
^lsoViialeX (BBL), V-C-N (BBL) and NaHCO^ were added aseptically after basic medium was auto-claved.
Initial growth studies were accomplished on hemoglobin
and glucose-free Neicarb suppleraented with the carbon/energy
source being studied. Glucose-free medium was achieved by

20
using sterile distilled water in place of the diluent
fluid for reconstituting the IsoVitaleX. The diluent
fluid is a 10% glucose in water solution (Table 3)« Growth
was studied in 1 or 2-ounce prescription bottles.
TABLE 3
SEMI-SOLID MEDIUM FOR INITIAL GROWTH STUDIES *
Components Amount/liter
GC Agar Base (DIFCO) 72.0 gm IsoVitaleX (Glucose-free) 10.0 ml V-C-N 10.0 ml NaHCO^ (7.5%) 30.0 ml
*lsoVitaleX (Glucose-free)(BBL), V-C-N (BBL), NaHCOo and the sterile carbon/energy source were added aseptically after basic medium was auto-claved. Final pH was adjusted to 7»0 with 2N NaOH.
Further growth studies were accomplished in a liquid
mediura described in Table 4. One hundred milliliters of
medium were used in 500 ml growth flasks for growth studies.
The different carbon/energy sources studied in both
semi-solid and liquid cultures are listed in Table 5.
Combinations of these sources were also studied.
Growth Studies. For initial studies, cell suspen-
sions of Neisseria gonorrhoeae were prepared in liquid
growth medium and treated on a Vortex-Geni (Model K-550-G)
to disperse any cell clumps.

21
TABLE ^
LIQUID MEDIUM FOR GROWTH CURVE STUDIES *
Components
Polypeptone (BBL) Corn Starch K HPOzi. KH2POZ1 NaCl IsoVitaleX (Glucose-V-C-N NaHCO^ (7*5%)
-free)
Amount/liter
15.0 gm 1.0 gm ^.0 gm 1.0 gm 5tO gm 10.0 ml 5.0 ml 10.0 ml
NaJÍCO and sterile carbon/energy source were added aseptically after basic mediura was auto-claved. Final pH was adjusted to 7.0 with 2N NaOH.

22
TABLE 5
CARBON/ENERGY SOURCES STUDIED
Components
Glucose (IX) Glucose (0.1 X) Glucose (0.01 X) Glucose (0.001 X) Sucrose Fructose Lactose Mannitol Sorbitol Inositol Arabinose Xylose Pyruvate (Sodium Salt) Galactose Lactate (Sodium Salt)(l X) Lactate (Sodium Salt)(0.1 X) Maltose Tartrate (Potasium sodium salt) Malonic Acid Propionate (Sodium Salt) Succinate (Sodium Salt)
Amount/liter
5.0 0.5 0.05 0.005 5.0 5.0 5.0 5.0 5.0 5.0 5.0 5.0 5.0 5.0 5.0 0.5 5.0 5.0 5.0 5.0 5.0
gm gm gm gm gm gm gm gm gm gm gm gm gra gm gm gm gra gm gm gm gm

23
Two loopfuls were inoculated on previously prepared
aemi-solid media which had been preheated to 35® C. The
organisms were spread over the surface of the media with
the inoculating loop. The cultures were then incubated
at 35® C (or as specified). Growth was based qualitatively
on surface growth.
For growth maintenance studies, one loopful of the
cell suspension was placed on the lower surface of pre-
pared preheated (35® C) medium in 2-ounce prescription
bottles and incubated at 35® C at the vertical position.
Subsequent streaks from the initial growth were made at
Z^ hour intervals for five days.
For later studies, cell suspensions were prepared
by transferring primary cells to liquid growth medium with
glucose (0.5%) and growing for 24 hours. One milliliter
of this suspension was inoculated into 500 ml screw-top
nephlo-culture flasks containing 100 ml of preheated (35® C)
liquid media. The cultures were incubated at 35° C on a
controlled environmental incubator-shaker at 200 RPM
(New Brunswiclc Scientific Model G25). Growth was measured
turbidimetrically with a Klett-Summerson Colorimeter
(Model 900-3) with a number 42 filter.
Papanicolaou Stainin^« Slides for staining using
Papanicolaou's method were sprayed with a cytology fix-
ative at the time the samplc was taken. Subsequent stain-
ing was a four minute hematoxylin regressive Papanicolaou

24
stain (62). After staining, slides were cover-slipped,
preserved in Histoclad (Clay Adams) and screened with a
standard light microscope.
Gram Staining. Slides prepared for gram staining were
refrigerated and stained within 48 hours. staining was
accomplished using the Burke-Kopeloff-Beerman modification
of the gram stain (34). After staining, slides were
cover-slipped, preserved in Histoclad (Clay Adams) and
screened with a standard light raicroscope.

CHAPTER III
RESULTS
Growth Studies on Semi-Solid Media. Studies with H C W M M U a
the various carbon/energy soxirces were first performed on
semi-solid media. Clinical isolates 1735» 1773 and 76I6
were used for preliminary studies followed by isolates
2326, 3001 and 7676. Growth obtained with these organisms
is summarized in Table 6. Especially good growth was
obtained at 24 hours on media using glucose, sucrose,
xylose and pyruvate alone and the combinations of pyruvate
plus sucrose plus 0.05% glucose, pyruvate plus 0.5%
glucose and pyruvate plus succinate. Poor growth was noted
with no additional carbon/energy source, mannitol and
tartrate. In all cases, no visable growth was noted at
24 or 48 hours on media with fructose as the priraary carbon/
energy source.
Further studies with semi-solid media investigated
the ability of the organism to grow at various temperatures.
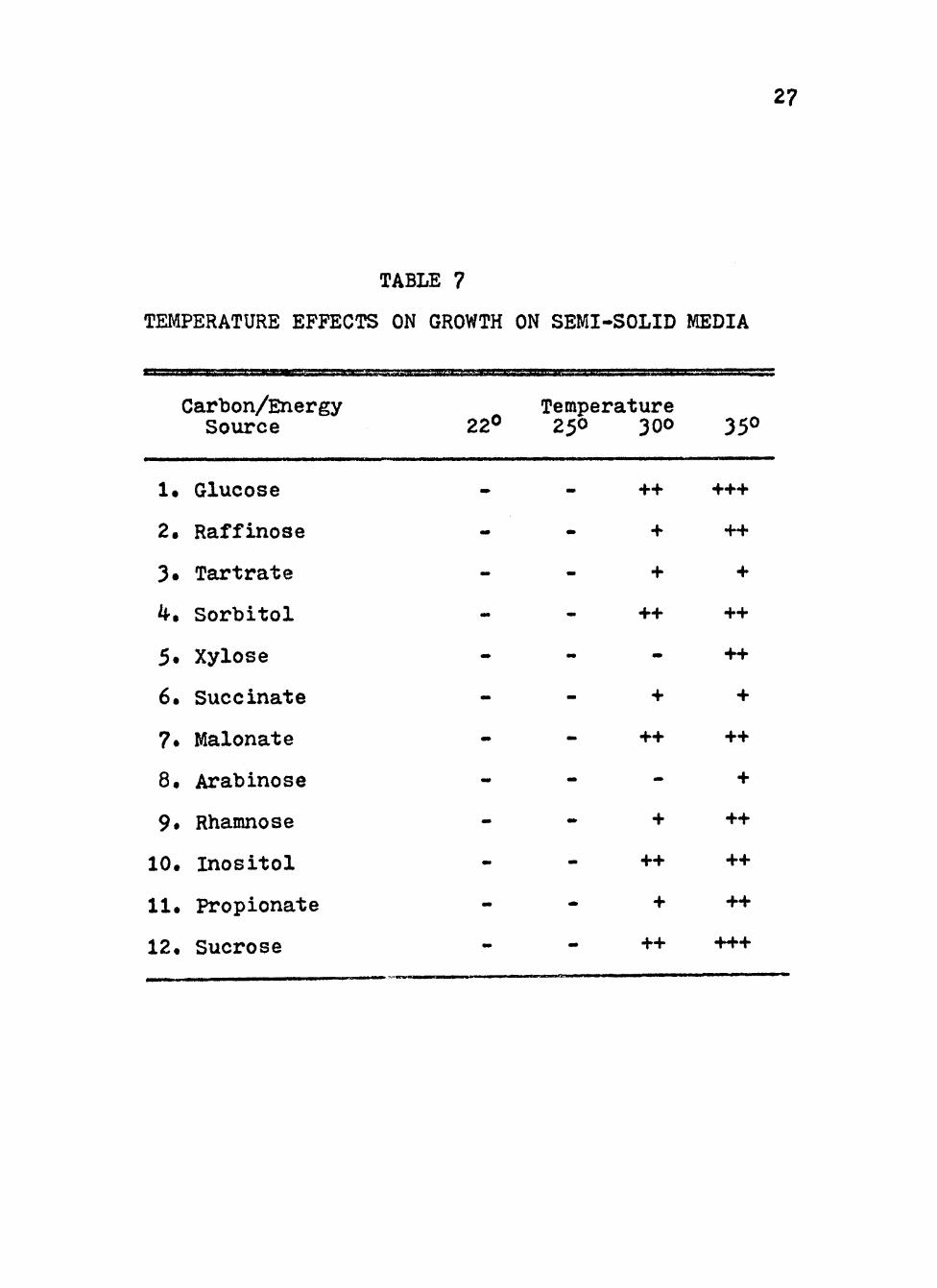
A summary of these studies is found in Table 7« In all
cases, the best growth was obtained at 35® C. At 30® C
media with sorbitol, inositol, succinate and tartrate
supported growth the same as at 35® C. Media with the rest
of the carbon/energy sources showed less growth at 30^ C
than at 35° C, with no growth observed for xylose and
25

TABLE 6 26
GROWTH ON SEMI-SOLID MEDIA •
Carbon/Energy Sources
Growth on Isolate 1735 1773 7676 2326 3001 7616
V.
1. None 2. Glucose 3. Glucose (0.05%) 4. Sucrose 5. Lactate 6. Maltose 7. Maltose +
Glucose (0.05%) 8. Pyruvate 9. Rhamnose 10. Tartrate 11. Malonaté 12. Arabinose 13. Inositol 14. Fructose 15. Propionate 16. Propionate (0.25%) 17. Propionate (0.75%) 18. Propionate (1.0%) 19. Propionate (0.5%) + Glucose (0 20. Propionate (0.25%) + Glucose ( 21. Propionate (0.75%) + Glucose ( 22. Propionate (1.0%) + Glucose (0 23. Raffinose 24. Succinate 25. Mannitol 26. Xylose 27. Sorbitol 28. Pyruvate + Sucrose + Glucose ( 29. Pyruvate + Glucose (0.05%) ++ 30. Pyruvate + Lactate + Succinate 31. Pyruvate + Succinate 32. Succinate + Lactate 33« Pyruvate + Lactate
+ +++ +++ ++ ++ ++
+++ ++
«
+ +++
++ ++ ++
+++ ++
-
+ +++
+++
++ +
++ +
++
++
+ +++
+++ ++ ++
+++ ++ ++ ++ ++ ++
-
+
+ +++
+++ ++ ++
+++ ++ +
++ ++ ++ -
++
+++
++ + + +
0.5%) 0.5%) .5%)
0.05%) ++
++ +
++ ++
++ +++ ++
+++
++ + +
+++ +
+++ +++ +++ ++ ++ ++
+
++ -
+ +++ ++
+++ +++
• Incubated at 35° C. + Poor Growth, ++ Moderate Growth, +++ Excellent Growth.
27
TABLE 7
TEMPERATURE EFFECTS ON GROWTH ON SEMI-SOLID MEDIA
Carbon/Energy Source
Temperature 22® 250 300 35®
1. Glucose
2. Raffinose
3» Tartrate
4. Sorbitol
5. Xylose
6. Succinate
7. Malonate
8. Arabinose
9. Rhamnose
10. Inositol
11. Propionate
12. Sucrose
++
+
+
++
-
+
++
-
+
++
+
++
+++
++
+
++
++
+
++
+
++
++
++
+++

28
arabinose at 30® C.
Survival Time with Different Carbon/Energy Sources.
Clinical isolates 7618 and 7619 were studies on semi-solid
raedia with the different carbon/energy sources for maximum
survival time at 35° C. The results aire summarized in
Table 8. Isolates grown on media containing lactate,
pyruvate, sucrose, inositol, sorbitol and tartrate remained
viable longer than media with glucose, with the best
results obtained from lactate and sucrose. Isolates grown
on media containing malonate as carbon/energy source
remained viable for a shorter time than those grown on
glucose. The remainder of the carbon/energy soxirces
demonstrated survivability comparable to glucose.
V-C-N Concentration Studies. The effects of various
concentrations of V-C-N inhibitor is summarized in Figure 1*
Concentrations greater than 5 ml/liter have significant
effect of the growth of Neisseria gonorrhoeae. At
10 ml/liter, maximum growth was rauch less and was achieved
a few hours earlier. With 20 ml/liter, maximum growth was
very much lower and death phase began almost immediately
after log growth.
Growth StudJes in Liquid Media. Those carbon/energy
sources which supported good growth on semi-solid media
were studied in broth cultures with clinical isolates 3092,
3225, 3413. 3698 and 7616.

29
TABLE 8
SURVIVAL TIME WITH DIFFERENT CARBON/ENERGY SOURCES
Carbon/Energy Source
1. Glucose
2. Lactate
3. Pyruvate
4. Sucrose
5. Succinate
6. Arabinose
7. Sorbitol
8. Inositol
9. Tartrate
10. Rhamnose
11. Propionate
12. Raffinose
13. Xylose
14. Malonate
Days 1
4+
4+
4+
4+
4+
4+
4+
4+
4+
4+
4+
4+
4+
4+
Since 2
4+
4+
4+
4+
4+
4+
4+
4+
4+
4+
4+
4+
4+
2+
Initial 3
2+
4+
4+
4+
2+
2+
4+
4+
4+
2+
2+
2+
2+
2+
.•L.H- - ± J
Inoculation 4 5
m>
4+
2+
4+
-
-
2+
2+
2+
-
•»
-
-
TJ'i" L - . ..
-
2+
2+
2+
-
-
-
•w
-
«»
-
-
-

30

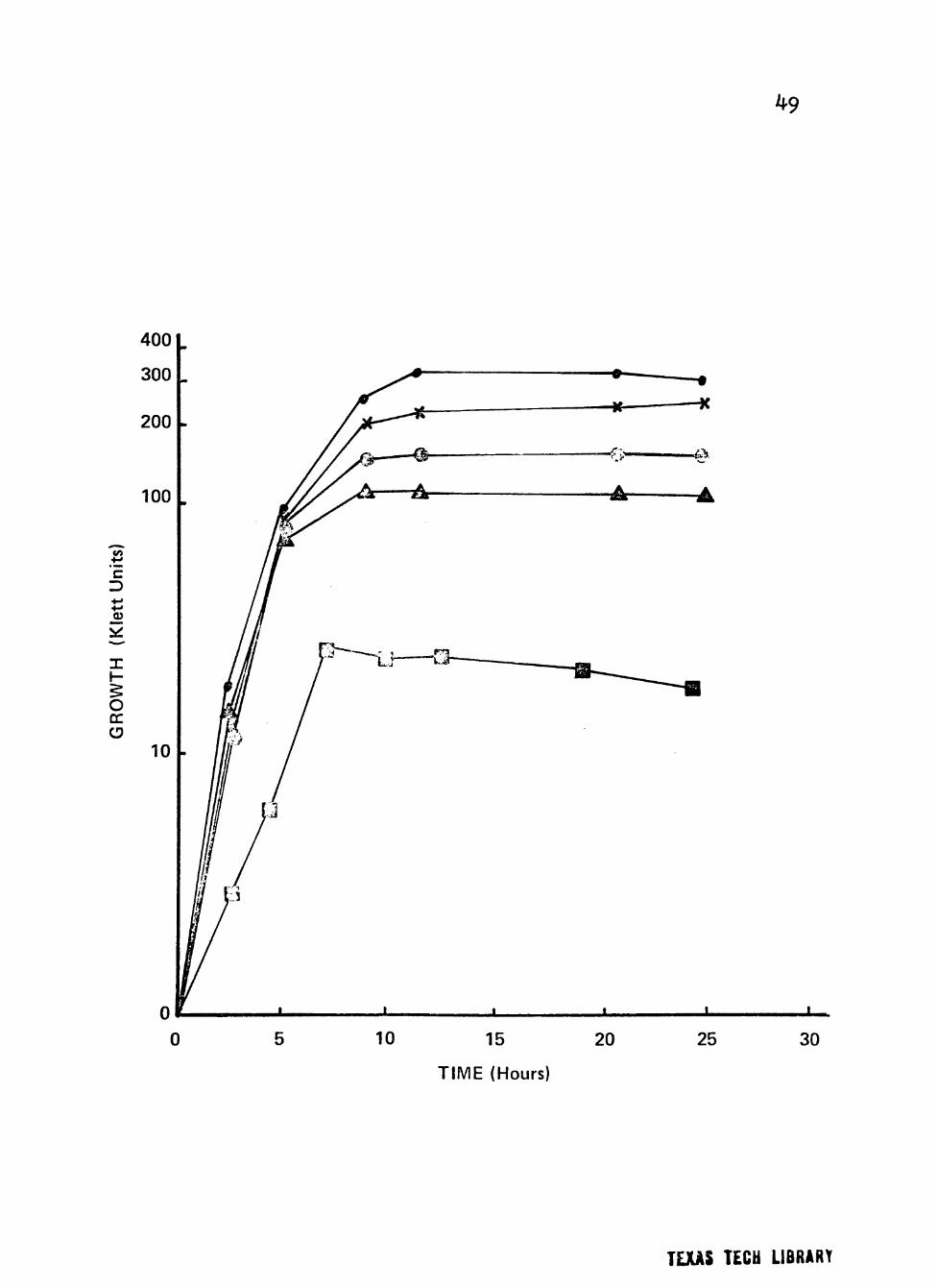
Figure 1. Growth of clinical isolate (2957) of Neisseria p;onorrhoeae in a liquid growth medium with varymg concentrations of V-C-N inhibitor.
Cultures were incubated at 35® C in screw-cap nephlo-culture flasks in a controlled environmental shaker-incubator at 200 RPM.
r
Symbolsi • 0 ml/liter
^ 2 ml/liter
I © 5 ml/liter
Q 10 ml/liter
A 20 ml/liter

31
400
300
200
100
'E
••-•
O cc CJ
10
0 8 16 24 32
TIME (Hours)

32
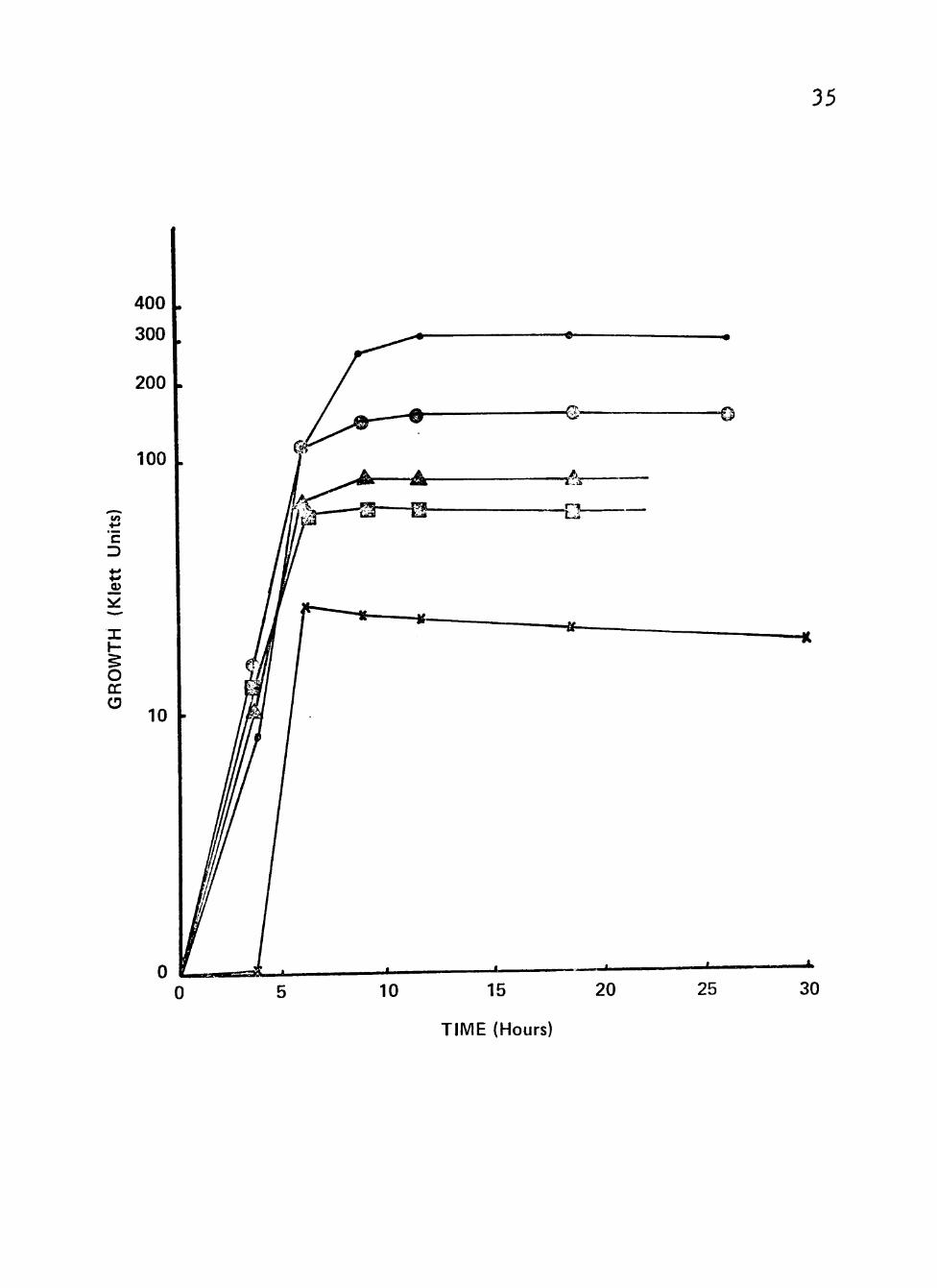
Growth with various concentrations of glucose was
explored with isolate 3698. The best growth was obtained
with a final glucose concentration of 0.5%. Successively
lower maximal growth was obtained down to 0.0005% glucose,
as shown in Figure 2.
With the other isolates the growth with various
other carbon/energy sources was studied. The best growth
was achieved using glucose, sucrose or lactate. These
results are found in Figure 3. The other energy sources
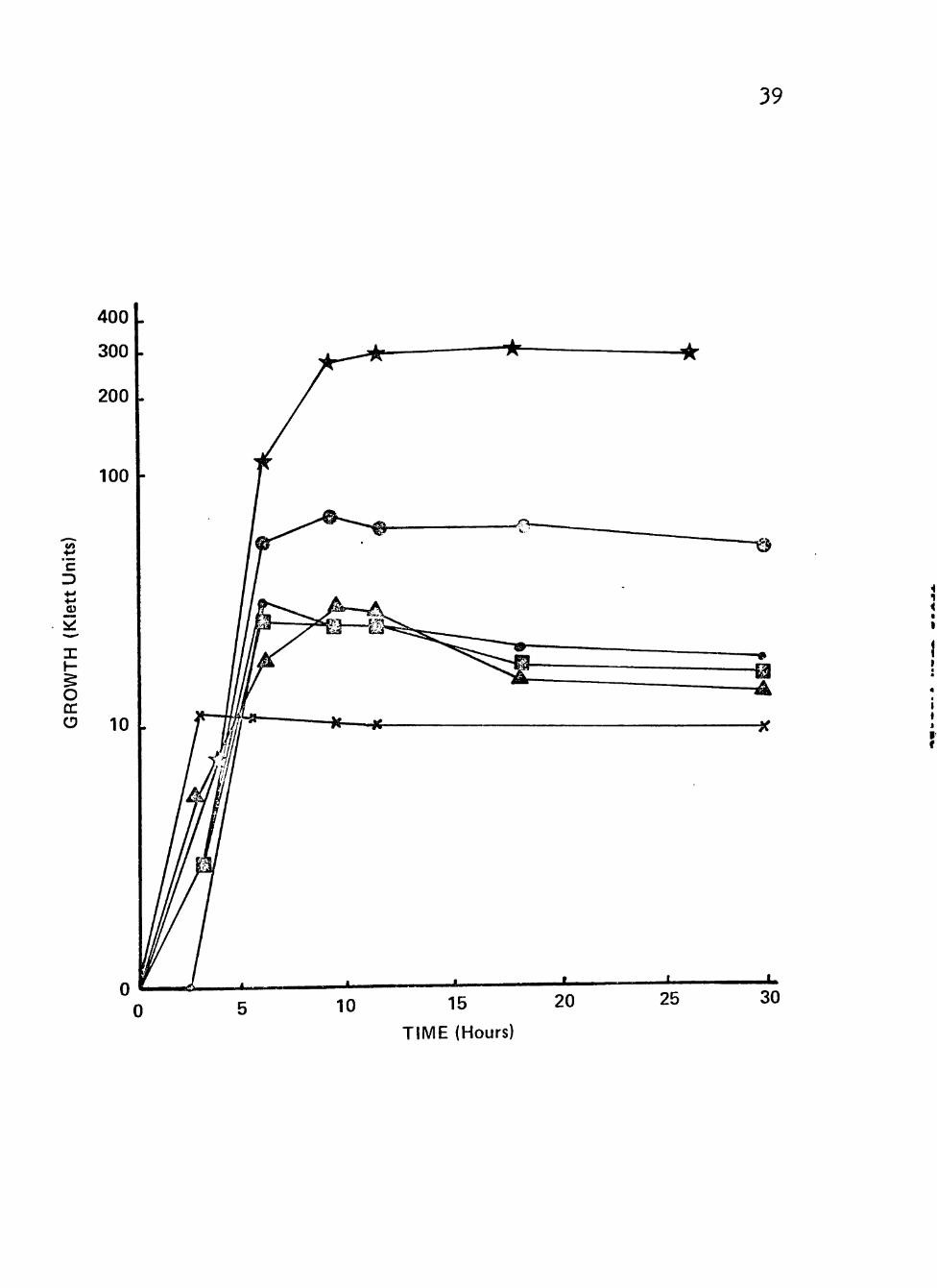
all had similar growth patterns as shown in Figures 4 and 5.
The maximum growth obtained was much less than with
glucose, sucrose or lactate. Maximum growth from the
greatest to the least with the various carbon/energy
sources wasi raffinose, rhamnose, malonate, arabinose,
xylose, inositol, propionate, no additional source,
fructose and succinate. Xylose and propionate both showed
log growth similar to the others, but the stationary
phase was much shorter and the declining death phase began
sooner. The rate of growth during log phase was similar
with all the carbon/energy sources. The lag phase was
similar for all, except with arabinose aná with no addit-
ional carbon/energy source which exhibited lag phases
about 2 hours longer than the others.
Figures 6, 7, 8 and 9 show the growth obtained with
combinations of carbon/energy sources, All the combinations
used grew about the same with the exception of propionate

33
plus 0.05% glucose and fructose plus 0.5% or 0.05%
glucose. The growth obtained in all combinations
paralleled growth with 0.05% glucose alone, except when
used with propionate which had growth less than with
' either alone and with fructose which showed a slight
reduction of growth with the combination.
Papanicolaou and Gram Staining Comparisons. Cervical
smears from twenty-six positive female gonorrhea patients
were stained by both Gram's stain and the method of
Papanicolaou. Of those gram stained, seventeen (65.4%)
smears had an overwhelming number of small, gram-negative
diplococci, both intracellular and extracellular. In
eight cases (30.8%) there was a wide variety of micro-
flora with some gram-negative diplococci, some intra-
cellular. In one case (3.8%) no gram-negative diplococci
were convincingly demonstrated.
Smears stained by Papanicolaou's method were of
poorer resolution which made identification of any micro-
flora difficult. Bacteria either stain poorly or not at
all. By adjusting the plane of focus away from the nucleus
of the tissue cells present in the smear, small areas of
what appeared to be groupings of diplococci could be seen
in eight (30.8%) smears. These areas usually appeared
to be associated with the tissue or polymorphonuclear
leucocytes. In the remainder of the slides, evidence of
Neisseria ^onorrhoeae was not present. These results

34
•'J:

Figure 2. Growth of clinical isolates (3092 and 3698) of Neisseria gonorrhoeae in a liquid growth medium v/ith varymg concentrations of glucose.
Cultures were incubated at 35® C in screw-cap nephlo-culture flasks with the addition of varying concentrations of glucose in a controlled environmental shaJker-incubator at 200 RPM.
r >
• Symbolsi • 0.5% glucose m
r
s ® 0.05% glucose r
1 A 0.005% glucose a
@ 0.0005% glucose
^ 0.0% glucose
35
C
h-
o cc
TIME (Hours)

36

Figure 3. Growth of clinical isolates (3092 and 3698) of Neisseria gonorrhoeae in liquid growth medium with glucose, sucrose or lactate as carbon/energy source.
Cultures were incubated at 35° C in screw-cap nephlo-culture flasks with the addition of different carbon/energy sources in a
5 controlled environmental shaker at 200 RPM.
- Symbolsi • Glucose (0.5%)
l 0 Sucrose (0.5%)
» Q Lactate (0.5%)
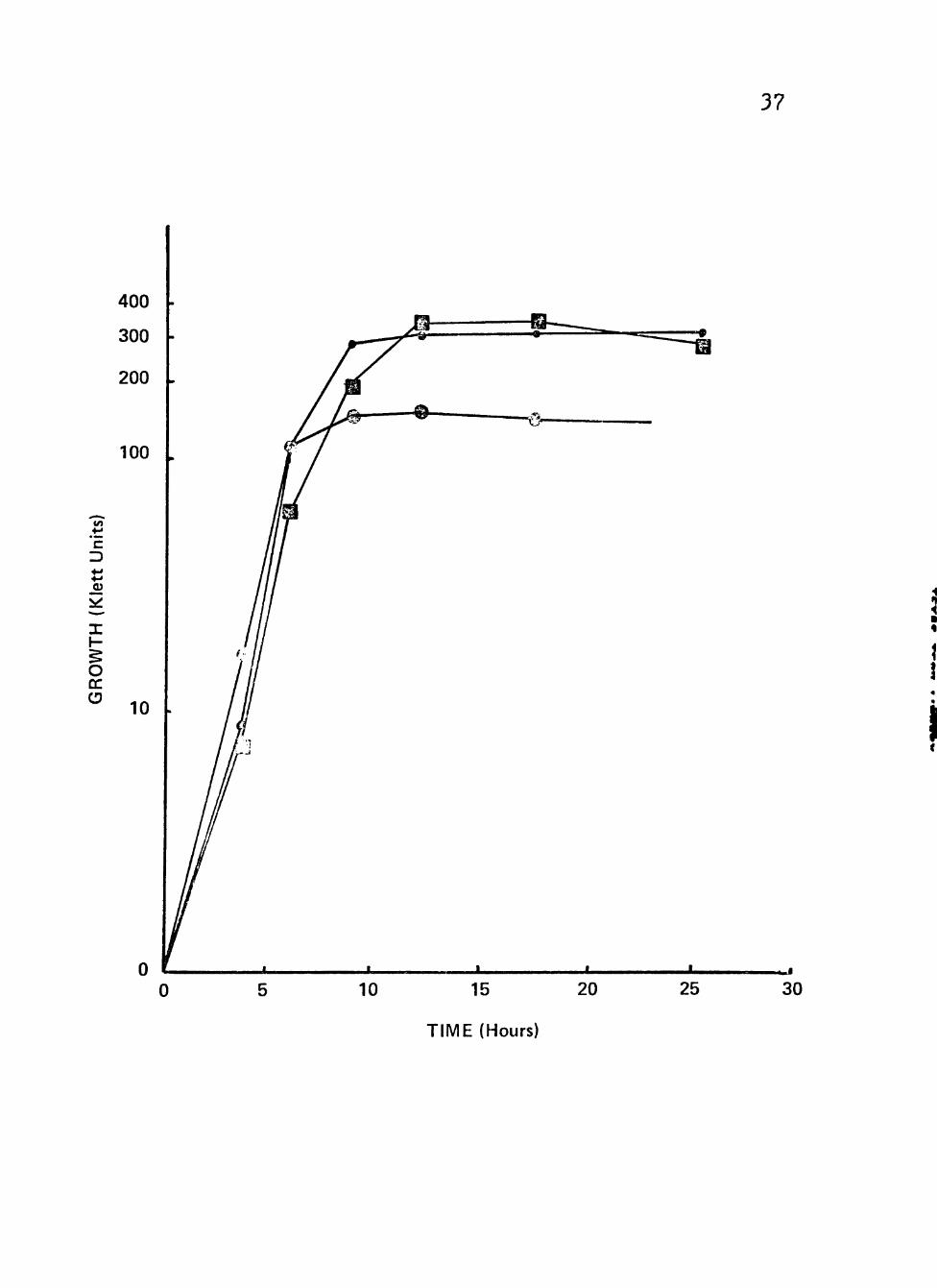
37
400
300
200
100
c
O QC O
10 .
0
I
TIME (Hours)

38
t

Figure 4. Growth of clinical isolates (3092, 3225 and 3413) of Neisseria gonorrhoeae in liquid growth medium with glucose, fructose, propionate, raffinose, succinate or no addition as carbon/energy source.
Cultures were incubated at 35® C in screw-cap nephlo-culture flasks with the addition
r of different carbon/energy sources in a H controlled environraental shaker at * 200 RPM. n c Symbolsi ^ Glucose (0.5%)
a Fructose (0.5%)
A Propionate (0.5%)
^ Raffinose (0.5%)
K Succinate (0.5%)
• No Addition
39
to +j 'E D 4-»
O tr
0 10 15
TIME (Hours)
20 25

• ; ' . ; •.
40
> 1 .• s * » ^. Í ( . 1 . . ' 1
• • ' ! .• • i
V {
I

r > >
n C 3
Figure 5. Growth of clinical isolates (3092, 3225 and 3413) of Neisseria gonorrhoeae in liquid growth medium with glucose, rhamnose, xylose, arabinose, malonate or inositol as carbon/energy source.
Cultures were incubated at 35® C in screw-cap nephlo-culture flasks with the addition of different carbon/energy sources in a controlled environmental shaker-incubator at 200 RPM.
Symbolsi x Glucose (0.5%)
• Rhamnose (0.5%)
1 ® Xylose (0.5%)
Arabinose (0.5%)
Malonate (0.5%)
Inositol (0.5%)
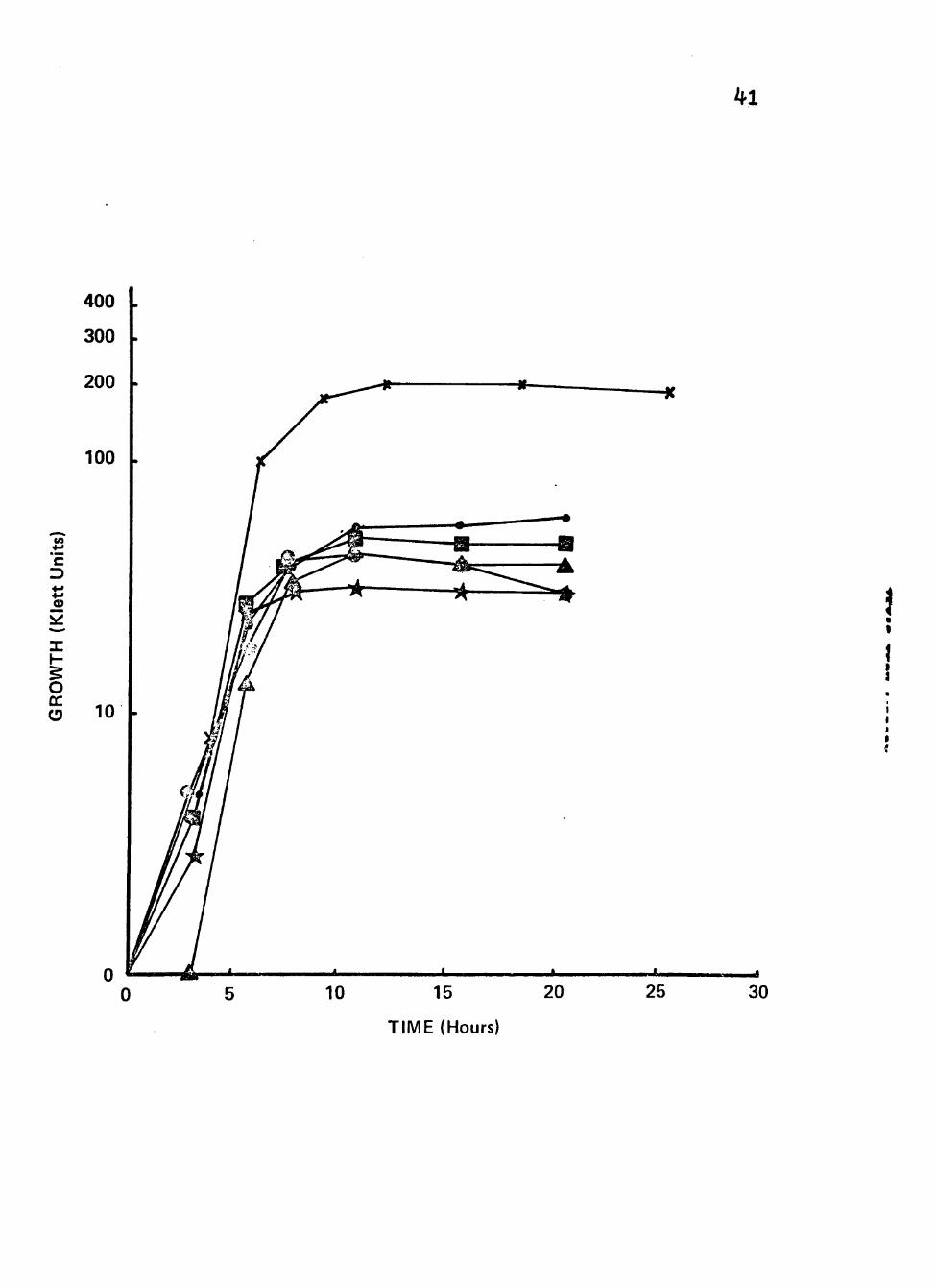
41
400
300
200
100
tn
"E D 00
X
O ÛC (D 10
10 15
TIME (Hours)

42

Figure 6. Growth of clinical isolates (3092, 3225 and 3413) of Neisseria gonorrhoeae in liquid growth medium with combinations of glucose or sucrose with lactate as carbon/energy source.
Cultures were incubated at 35® C in screw-cap nephlo-culture flasks with the addition of combinations of different carbon/energy sources in a controlled environmental
Í incubator-shaker at 200 RPM. >
5 Symbolsi • Lactate (0.5%) + Glucose (0.05%)
3.
Lactate (0.5%) + Glucose (0.005%)
Lactate (0.5%) + Sucrose (0.05%)
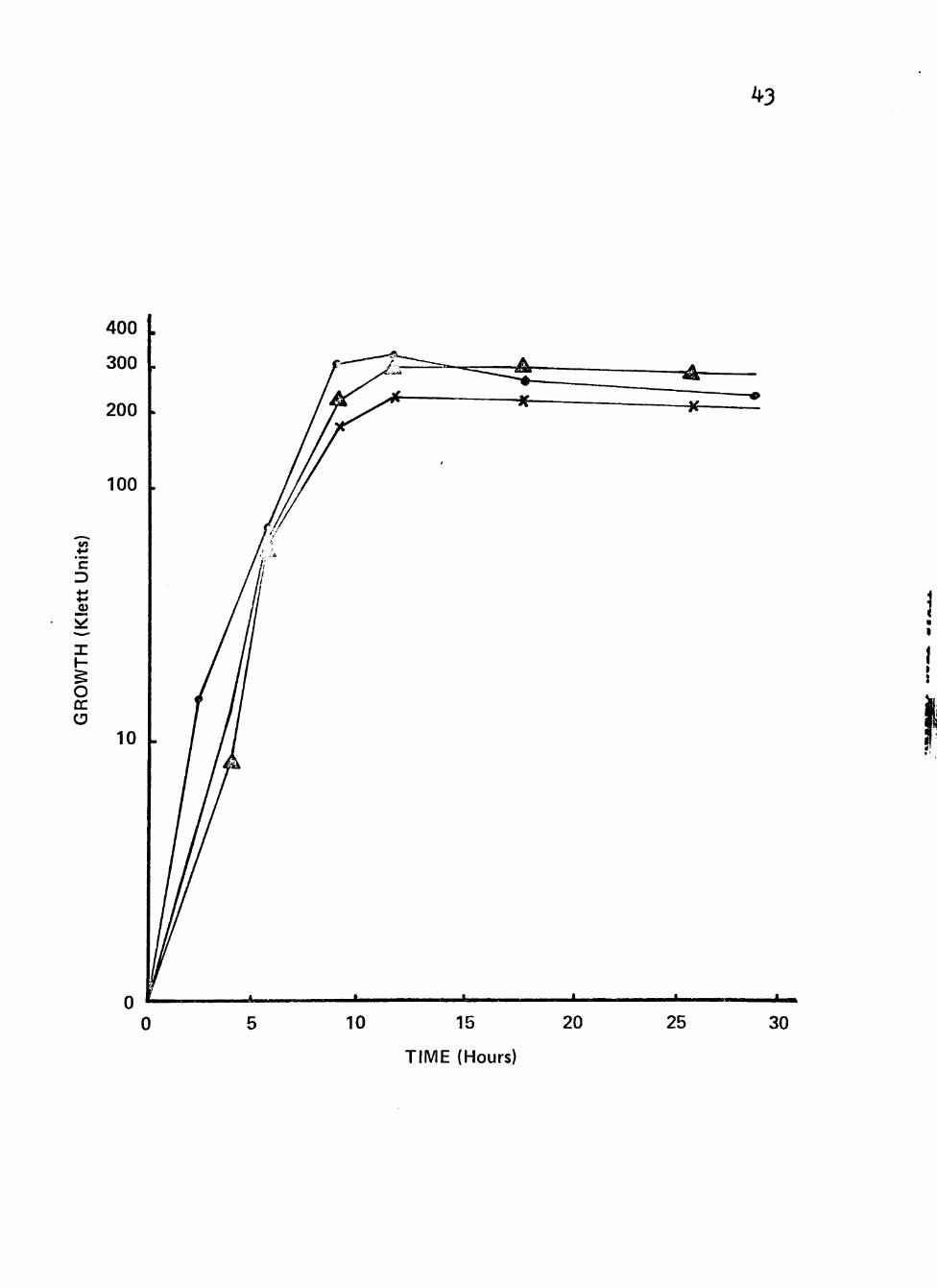
^ 3
C
+•>
.2
X h-
o cc o
15
T I M E (Hours)

r ..
' . , . ' • • •
• I ,
k V i. • J í.
44

m
Figure 7. Growth of clinical isolates (3092, 3225 and 3413) of Neisseria gonorrhoeae in liquid growth medium with combinations of glucose with sucrose and with propionate as carbon/energy source.
Cultures were incubated at 35® C in screw-cap nephlo-culture flasks with the addition of combinations of different carbon/energy sources in a controlled environmental
^ incubator-shaker at 200 RPM.
1'
Symbolsi # Sucrose (0.5%) + Glucose (0.05%)
^ Propionate (0.5%) + Glucose (0.05%)
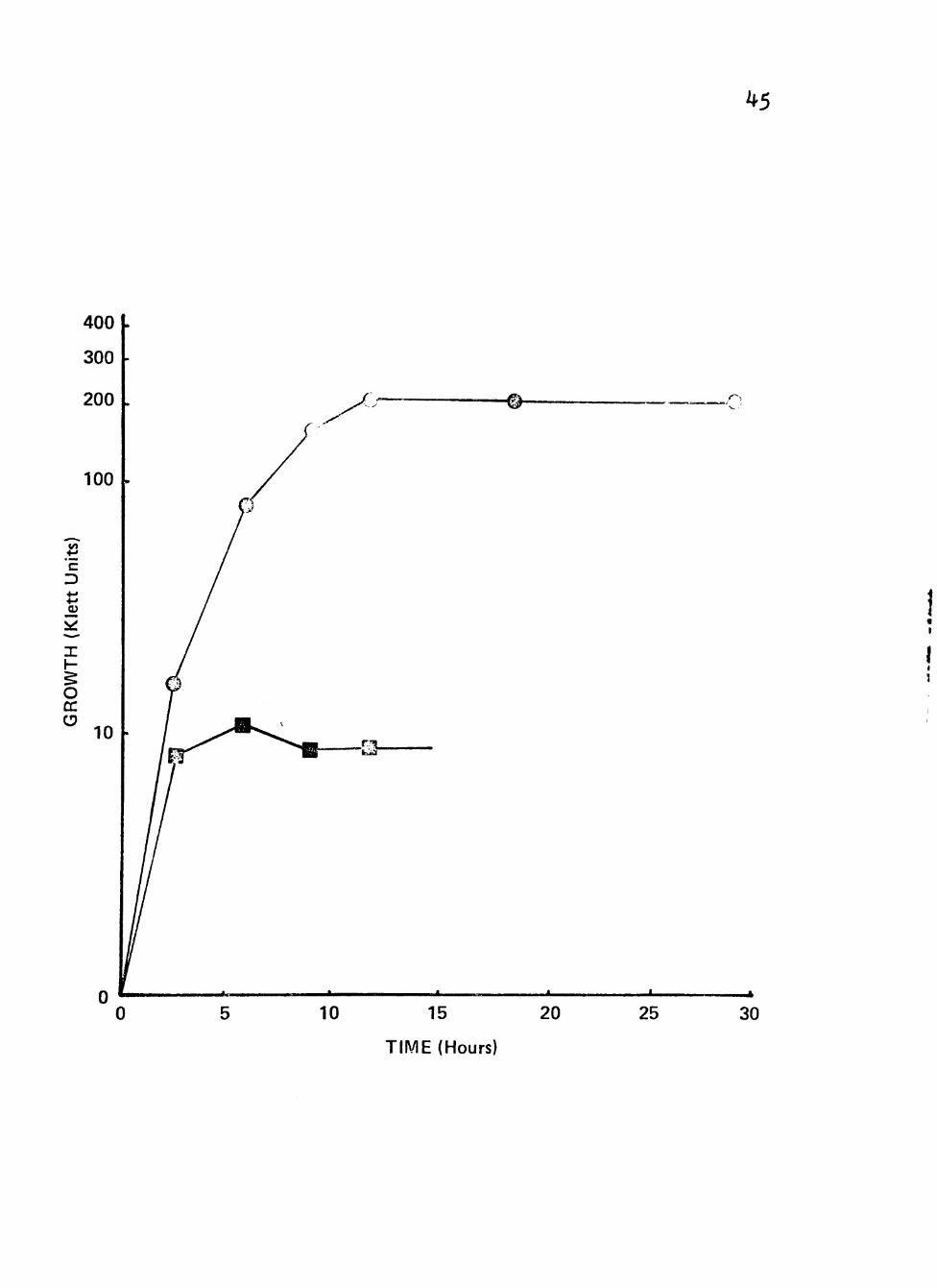
45
0 10 15 20
TIME(Hours)
30

46
. 4
r. ' i
'* r< 't
l. i:i \ "
^
. - . f
T.í •• .'.n
•"*
f . , • > < ? . y \
=f". •• s -

Figure 8. Growth of clinical isolate (76I6) of Neisseria gonorrhoeae in liquid growth medium with glucose or the combinations glucose plus xylose, glucose plus Buccinate or glucose plus lactate plus succinate.
Cultures were incubated at 35^ C in screw-cap nephlo-culture flasks with the addition
;* of combinations of different carbon/energy > sources in a controlled environmental •* incubator-shaker at 200 RPM. m S Symbolsi • Xylose (0.5%) + Z Glucose (0.05%)
< 5 @ Succinate (0.5%) +
Glucose (0.05%)
B Glucose (0.05%)
• Lactate (0.5%) + Succinate (0.5%) + Glucose (0.05%)
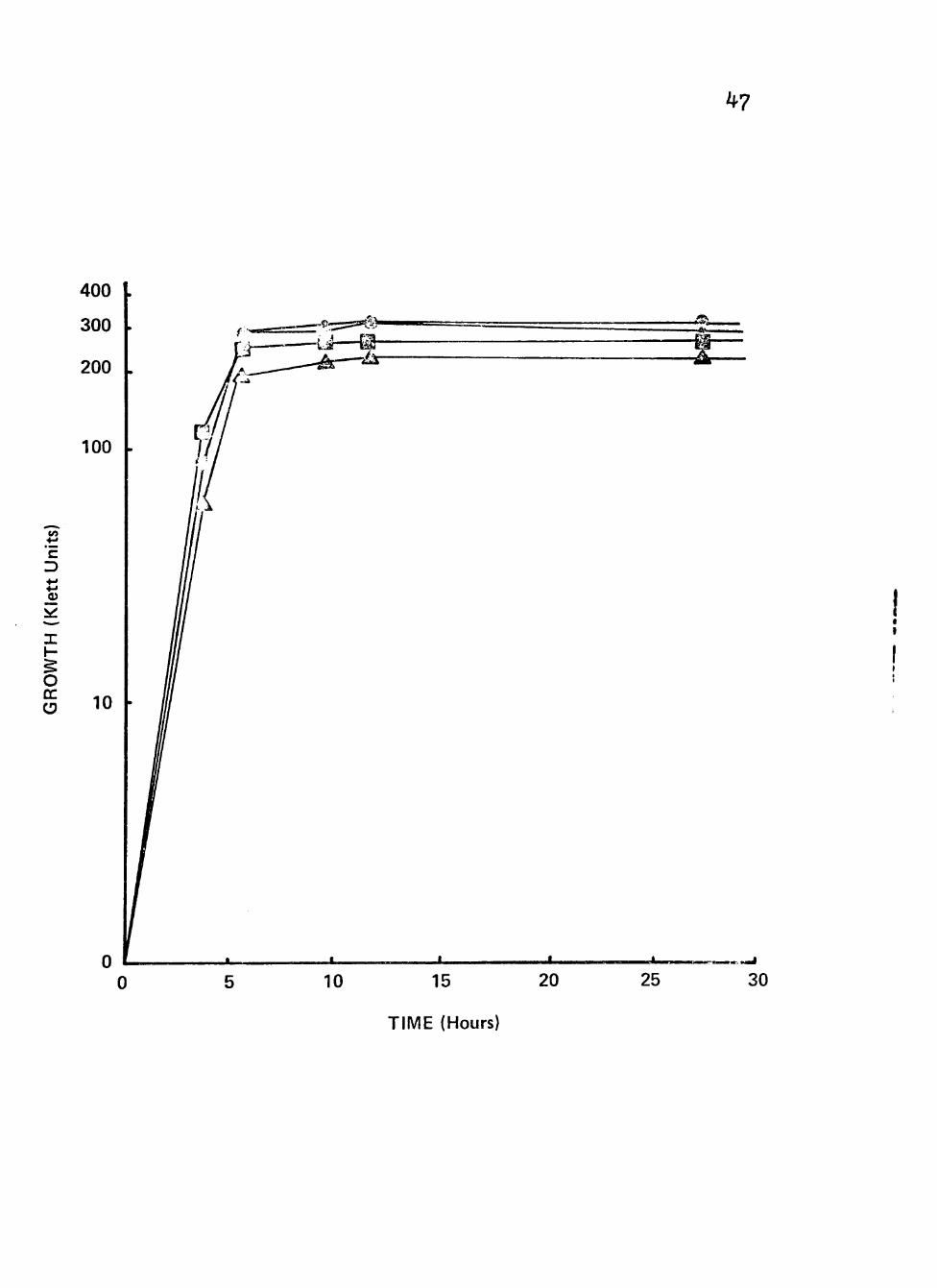
c
O cc o 10
47
100
JQ)
10 15 20 25 30
TIME (Hours)

48

0) asoq.onaj Q
(%^*0) asoq.onjtá + (5^^0*0) osoonxo ^
(%í*0) 9soq.onjy[ + (2éí*0) asoonxo x
($éíO*0) asoonxo ©
(%í'0) asoonxo • isioqni/Cs
•MH 002 íí- ae3{Bt{s-aoíi.BqnouT XT3q.U9uiuoaT«Aue paxxoaq.uoo B UT saoanos
iC9j8ua/uoqaBO q.uaaej:jTp jo suoTí).BÛTqraoo jo uoxq.TppB auq. iiq.TM SJISBXJ aanq.x^ô-0IM<Í9U dBO
-Maâos UT 0 Q C Q- peq.BqnouT eaaM saanq.xno
•aoanos iCSaaua/uoqaBo SB asoq.onaj snxd osoonx^ ao asoonxS q-îM mnTpam
i(q.Moa3 p-cnbTX u" aBaoqaaouoa BTaassTaM j o (9<i9^) *aq.Bxos X^^TUTXO JO Mq.Moao »6 aan^xá
49
c
I H
O cc o
0 10 15
TIME (Hours)
20 25 30
TEXAS TECU LIBRARY

50
are summarized in Table 9.
TABLE 9
PAPANICOLAOU AND GRAM STAIN COMPARISON
Presence of Gonorrhea-like Stain Organisms
Gram Stain 17 (65.4%) 8 (30.8%) 1 (3.8%) Papanicolaou Stain 0 8 (30.8%) 18 (69.2%)
Oxidase Testing. Attempts to incorporate a test
for oxidase into the grara staining procedure using
N, N, N*I N'-tetramethyl-£-phenylenediamine dihydrochloride
were unsuccessful. The test reagent was tried in all
steps of the Burke-Kopeloff-Beerman and Hoeker modificat-
ion of the gram stain but was either washed off or
crystalized, destroying a useful stain. At no time was a
color change indicating the presence of oxidase observed.
Frozen-Storage Studies. Two clinical isolates,
2959 and 3001, were frozen and recovered successfully at
intervals spanning over two months. Colonies recovered
appeared to be of a different morphology than those taken
for freezing. Colonies selected from Neicarb v/ere small,
convex and easily emulsified. Frozen cultures regrovm on
Ncicarb had colonies which v/ere much larger, convex and
mucoid.

CHAPTER IV
DISCUSSION
Initial growth studies on semi-solid media demon-
strated that Neisseria gonorrhoeae can grow with a variety
of compounds as the primary carbon/energy source.
Excellent growth occured with glucose, sucrose, pyruvate
and xylose on semi-solid media. Good growth occured with
a number of other carbon/energy sources including lactate,
rhamnose and raffinose. The growth curves established
from growth in liquid media quantified the growth from
these carbon/energy sources. As previously reported by
Morse and Bertenstein (52), excellent growth occured using
glucose, lactate and pyruvate. Surprisingly good growth
also occured using sucrose. Sucrose (0.5%) supported
growth comparable to 0.05% glucose. Glucose comtaraination
could not account for this level of growth, as the
maximum glucose present, according to the manufacture
(Matheson, Coleman and Bell), in the sucrose used would
result in a final glucose concentration of only 0.00025%.
Growth on peptone without any additional carbon/
energy source was also demonstrated. Although growth
was poor using peptone alone, it was better than with
fructose or succinate.
51

52
In media with fructose plus 0.5% and 0.05% glucose,
growth of Neisseria gonorrhoeae is reduced due to an
inhibitory effect of the fructose (see Figiire 9). The
inhibition is not as profound as found in liquid media with
propionate plus 0.05% glucose (see Figure 7). In this case
the propionate had the effect of reducing growth to levels
lower than when grown with either alone. In fact growth
with propionate plus 0.05% glucose showed that the propion-
ate inhibited growth to a level lower than growth on pept-
one alone. When grown on semi-solid media with varying
concentrations of propionate (see Table 6), Neisseria
^onorrhoeae was inhibited by concentrations greater than
0.75% when used along with 0.5% glucose.
The dependance on temperature for in vitro growth
oí Neisseria ^onorrhoeae was also studied. On semi-solid
media using glucose, raffinose, tartrate, malonate or
inositol as carbon/energy source, growth was the same at
both 30° C and 35° C. For all the other carbon/energy
sources growth was best at 35^ C. Isolates using xylose
or arabinose failed to grow at 30° C. No growth occured
using any carbon/energy source at 25° C or 23° C.
Bergey's Manual (7) records the optimum growth temperature
for Neisseria ^onorrhoeae as 35-36 C, but this optimum
refers to growth using only glucose as the primary
carbon/energy source. Results from these studies suggest

53
that for carbon/energy sources such as tartrate, malonate
or inositol, the optimum could be slightly lower or have
a slightly wider range.
When cell suspensions of Neisseria gonorrhoeae,
inoculated onto semi-solid media containing the various
carbon/energy sources, are restreaked from the initial
growth at one day intervals the length of time the
organisms remained viable was examined. On semi-solid
media using glucose as the primary carbon/energy source,
as found in most clinical media, the organisms remain
viable for only 2-3 days. This time prevents accurate
diagnosis in small laboratories for cultures taken on
Friday when the laboratory is to be closed over the week-
end. On media with lactate, pyruvate or sucrose as the
carbon/energy source, survivability time increases to
4-5 days. It appears that media for primary isolation of
Neisseria gonorrhoeae could be more useful, especially for
small laboratories or clinics, if they utilized lactate,
pyruvate or sucrose in place of glucose. Growth in liquid
growth media for these same carbon/energy sources
indicate that the lag phase and rate of growth during log
phase is comparable to a medium using glucose as carbon/
energy source. Thus a semi-solid media with lactate,
pyruvate or sucrose replacing glucose would also be suitable
for larger laboratories or small laboratories which read
cultures at 24-48 hours.

54
In another part of this study, the effects of
different concentrations of the commonly used inhibitor
V-C-N was studied. Concentrations of 0-5.0 ml/liter of
V-C-N have little effect on the growth of Neisseria
gonorrhoeae. At 10 ml/liter of V-C-N inhibitor growth
is significantly reduced. At 20 ml/liter, growth is poor
with the death phase beginning almost immediately
following log growth. It appears that the concentration
of 5.0 ml/liter of liquid growth media, as used in all
broth cultures in these studies, is the maximum amount
of V-C-N that ghould be used for good recovery of
Neisseria gonorrhoeae.
During frozen-storage studies, the use of Skim
Milk medixjm without the addition of glycerine appeared
to be satisfactory for storing cultures of Neisseria
gonorrhoeae over extended periods. Although colonial
morphology changed from small, convex and easily
emulsified colonies to ones which were large, convex and
mucoid when frozen cell suspensions were regrown, no
attempt was made to properly type either colony.
Results of these studies also indicate that it is
not feasable to use the Papanicolaou smear as a screening
tool for gonorrhea. The Papanicolaou staining procedure
does not adequately stain bacteria and results in a slide
of much lower resolution. The simplier gram stain is
far more valuable and much more clearly shows the micro-

55
flora on the cervix. Twenty-five cases (96.2%) showed
gram-negative diplococci using gram staining while only
eight (30.8%) Papanicolaou slides had any diplococci
present. Diplococci seen in the Papanicolaou smears
were difficult to observe and identify.
It would be more valuable for clinical screening to
make two slides at the time the Papanicolaou smear is made,
just as in this study, and stain one by the Burke-
Kopeloff-Beerman modification of the gram stain than it
would be to use the Papanicolaou stain to identify diplo-
cocci. Although this does not directly incorporate gon-
orrhea screening into an already routine procedure, it
would provide a quick technique to be performed during
the same sampling procedure. Praiser (59) and Phillips
(60) showed, in clinical diagnosis, between 24 and 31%
of the positive feraale gonorrhea cases tested v/ere
culture positive but were negative using gram staining
of cervical smears. Thus it is indicated that a raore
sensitive screening test is necessary. Data from these
investigations indicate that screening Papanicolaou
smears is not more sensitive technique, as suggested by
Heller (25). Although gram staining of cervical smears
appears more useful than the Papanicolaou staining
procedures, a more valuable microscopic tool should
involve specific fluorescent antibody procedures (65).
Also from the studies presented here, no practical

56
method was found for incorporating into the gram stain
a test for the enzyme oxidase. No location in, or
modification of, the procedure was successful in
detecting oxidase. Since the enzyme is normally detected
on the cell surface or extracellularly, the washing
procedure of the gram staining procediire tends to remove
most of the enzyme. If the enzyme was present on the
cell surface and reacted with the indicator, the cell
would most likely have been darkened, thus masking the
proper gram reaction. Microscopic techniques using
oxidase for confirmation of the presence of Neisseria
gonorrhoeae does not appear practical.

CHAPTER V
SUMMARY
This study demonstrated that in vitro Neisseria
gonorrhoeae can utilize a wide variety of compounds as
its primary carbon/energy source. The organism used
glucose, lactate and pyruvate with excellent growth.
Good growth also was obtained with sucrose as the carbon/
energy source. Other compounds found usable as carbon/
energy sources in order of maximum growth werei raffinose,
malonate, arabinose, xylose, inositol, propionate,
peptone alone, fructose and succinate.
Fructose, when used in conj\Jinction with glucose as
the carbon/energy source, was found to have a mild
inhibitory effect on the growth of Neisseria gonorrhoeae.
Propionate was found to be more inhibitory when used in
conjunction with glucose for the carbon/energy source.
A profound inhibition of the growth of Neisseria
gonorrhoeae results when propionate is added.
Preliminary studies on the dependance on temperature
for in vitro growth of Neisseria f!:onorrhoeae indicated
that the optimum temperature for growth on semi-solid
media utilizing most carbon/energy sources is near 35° C.
Growth with sorbitol, malonate, succinate, tartrate and
57

5Q
inositol was about the same at 30^ C and 350 C at 24 hours.
This implied that for these carbon/energy sources the
optimum may be slightly lower or have a broader range
than is reported for media with glucose.
With a medium utilizing glucose as the primary
carbon/energy source, Neisseria gonorrhoeae underwent
autolysis within 2-3 days. In these studies, it was
demonstrated that the use of sucrose, pyruvate or
lactate as the primary carbon/energy source in semi-solid
media prevented autolysis for about 5 days. This pro-
longed viability would provide small clinics and
laboratories the ability to collect samples on Friday
and read them on Monday without reducing diagnostic
accuracy. The lag and log growth phases are nearly
identical for in vitro growth with glucose, sucrose,
pyruvate and lactate, so there would be no loss in
diagnostic accuracy if the culture were read at shorter
intervals.
This study also demonstrated the effects of the
inhibitor V-C-N on in vitro growth of Neisseria
gonorrhoeae indicating that concentrations greater than
5 ml/liter in liquid growth media cause significant
reduction in maximum growth.
During frozen storage studies, it was deterrained
that Neisseria ^onorrhoeae can be stored at -90° C in
Skim Milk raedium without the addition of glycerine or

59
any other additives for prolonged periods with good
survivability.
The Papanicolaou smear was shown not to be a
valuable tool for the detection of gonorrhea in women. *
Bacteria in Papanicolaou smears are poorly stained and
difficult to identify. Two slides made from cervical
scrapings, one stained by Papanicolaou*s method for
cervical carcinoma and one stained by the Burke-Kopeloff-
Beerman modification of Gram's stain would provide a
more useful technique for carcinoma and gonorrhea
screening. The gram stain indicates the presence of
diplococci intracellular in white blood cells much more
clearly, and is a simple, quick procedvire.

LITERATURE CITED
1. American Public Health Association. 1976. Budget Supplement, pages 5-8. In American Public Health Association, The Nation's Health March 1976.
2. Bacigalupi, B. A., and J. Lawson. 1973. Defined physiological conditions for the isolation of the L-form of Neisseria gonorrhoeae. J. Bacteriol. Il6i778-7ô^.
3» Barlows, A. 1972. CDC program for diagnosis of gonorrhea. J. Am. Med. Assoc. 222i1557.
4, Barritt, M. M. 1944. An improved Pappenheim stain for gonococci. Brit. Med. J. 1|494.
5. Boor, A. K. 1942. Glutathione as an essential growth factor for certain strains of Neisseria gonorrhoeae. J. Biol. Chem, 153i142-150.
6. Bradford, W. R. 1973* Gonorrhea screening. J. Reproductive Med. 2i82-84.
7. Buchanan, R. E., and N. E. Gibbons. 1974. Bergey's Manual of Determinative Bacteriol-ogy Eighth Edition. Williams and Wilkins Company, Boston, Mass.
8. Catlin, B. W. 1973. Nutritional profiles of Neisseria /^onorrhoeae, Neisseria meningitidis, and NeiiTs'eria lactanrica in chemically defined media and the use of growth requirements for gonococcal typing. J. Infect. Dis. 128i179-194.
60

61
Caterall, R. D. 1974. Gonorrhea, pages 14-37. In R. D. Catterall, A Short Textbook of Venereology, 2nd Edition. J. B. Lippincott, Philadelphia, Pa.
10. Christensen, C. W., and H. W. Schoehlein. 1940. The culture method for detection and isolation ^^ Neisseria gonorrhoeae. Importance and methods of supplying carbon dioxide. J. Bacteriol. 40t162-166.
11. Clark, D. 0. 1973. Gonorrheai Changing concepts in diagnosis and management. Clinical Obstet. and Gynec. 16i2-24.
12. Davis, B. D., R. Dulbecco, H. N. Eisen, H. S. Ginsberg, W. B. Wood. 1973. The Neisseriae, pages 742-752. In B. D. Davis, R. Dulbecco, H. N. Eisen, H. S. Ginsberg and W. B. Wood, Microbiology, 2nd Edition. Harper and Row, Hagerstovm, Maryland.
13. Department of Health Education and Welfare. 1974. Reported Morbidity and Mortality in the United States 1974, pages 13-14. U. S. Department of Health Education and Welfare, U. S. Public Health Service, Atlanta, Georgia.
14. Editorial. 1971. Venereal disease. Lancet I169I.
15. Elmros, T., L. G. Burman and G. D. Bloom; 1976. Autolysis of Neisseria gonorrhoeae. J. Bact-eriol. 1261969-975.
16. Flynn, J. and S. A. Waitkins. 1972. A serum-free medium for testing fermentation reactions in Neisseria f!:onorrhoeae. J. Clin. Path. 25t52^-^^7.
17. Frarrer, C. J. and P. H. Tatham. I962. Screening for carcinoma of the uterine cervix in a YD clinic. Brit J. Ven. Dis. 38123O-23I.

62
18, Gale, E. P., E. Cundliffe, R. F. Reynolds, M. A. Richmond and M. J. Waring. 1972. Drugs affecting the function of the cytoplasmic membrane, pages 137-140. In E. F. Gale, E. Cundliffe, R. F. Reynolds, M. A. Richmond and M. J. Waring, (eds), The Molecular Basis of Antibiotic Action. John Wiley and Sons, Ltd., New York, N. Y.
19t Gale, E. F., E. Cundliffe, R. F. Reynolds, M. A. Richmond and M. J. Waring. 1972. Drugs affecting the function of the cytoplasmic membrane, pages 140-145. in E. F. Gale, E. Cundliffe, R. F. Reynolds, M. A. Richmond and M. J. Waring, (eds), The Molecular Basis of Antibiotic Action. John Wiley and Sons, Ltd., New York, N. Y.
20. Gale, E. F., E. Cundliffe, R. F. Reynolds, M. A. Richmond and M. J. Waring. 1972. Inhibition of bacterial cell wall synthesis, pages 102-116. In E. F. Gale, E. Cundliffe, R. F. Reynolds, M. A. Richmond and M. A. V/aring, (eds), The Molecular Basis of Antibiotic Action. John Wiley and Sons, Ltd., New York, N. Y.
21. Gordon and Hine (1916, cited by Difco Manual, 1953) pages 93-94. In Difco Manual, 9th Edition. Difco Laboratories, Inc. Detroit, Michigan.
22. Gordon and Hine (1916, cited by Difco Manual, 1953) Brit. Med. J. 2t678.
23. Gould, R. G. 1944. Glutathione as an essential growth requirement for certain strains of Neisseria gonorrhoeae. J. Biol. Chem. I53tl^3-150.
24. Griffin, R. J. and E. Racker. 1956. The carbon dioxide requirement of Neisseria gonorrhoeae. J. Bacteriol. 71i717-72 T

63
25» Hebler, B. H. and S. A. Morse. 1976. Tricarboxy-lic acid cycle activity in Neisseria gonorrhoeae. Abstracts of the annual meeting of the American Society of Microbiology, Washington, D. C.
26. Heller, C. J. 1974. Neisseria gonorrhoeae in Papanicolaou smearsl Ac?a Cytologica 18i338-340.
27. Holvey, D. N. 1972. Tissue pathology-cytology, pages 1825-1826. In The Merck Manual, Twelfth Edition. Merck, Sharp and Dohme Research Laboratories, Rahway, N. J.
28. Hosty, T. S., M. A. Freear, C. Baker, and J. Holston. 1975. Autolysis of Neisseria gonorrhoeae J. Bacteriol. 122138^-392.
29. Hunter, K. M. and I. McVeigh. 1970. Development of a chemically defined medium for growth of Neisseria gonorrhoeae. Antonie van Leeuween-hoek 361305-3Í6.
30. Hutchinson, R. I. 1970. Typing the Gonococcus. Brit. Med. J. 3il07.
31. James-Holmquest, A. N., R. D. Wende, R. L. Mudd and R. P, Wiliiams. 1973. Comparison of atmosphere conditions for culture specimens of Neisseria gonorrhoeae. Appl. Microbiol. 26i46é-^99.
32. Jephcott, A. E. and A. Reyn. 1971. Neisseria gonorrhoeae. Colony variation I. Acta path. micrôbiol. scand. Section B 79»609-614.
33. Juhlin, I. and G. Krook. 1965» Problems in diagnosis, treatment and control of gonorrheal infections. I. A coraparison of direct microscopy and culture. Acta Dermatovener (Stockholra) 45i142-147.

64
34. Kellogg, D. S. 1974. Neisseria gonorrhoeae. pages 124-130. In E. H. Lennette, E. H. Spauling and J. P. Truant (Eds), Manual of Clinical Micro-biology, Second Edition. American Society for Microbiology, Washington, D. C.
35. Kellogg, D. S., I. R. Cohen, L. C. Norins, A. L. Schroeter and G. Resing. 1968. Neisseria gonorrhoeae II. Colonial variation and the pathogenicity during 35 months in vitro. J. Bacteriol. 96i596-605.
36. Kellogg, D. S., W. L. Peacock, W. E. Duncon, L. Brown and C. I. Pirkle. 1964. Neisseria gonorrhoeae I. Virulence genetically linked to colonal variation. j. Bacteriol. 8511274-1279.
37* Kenny, C. P., F. E. Ashton, B. B. Diena and L. Green-berg. 1967. A chemically defined protein-free liquid medium for the cultivation of some species of Neisseria. Bull. W. H. 0. 321569-573.
38. King, A. and C. Nicol. 1975- Gonorrhea, pages 162-166. In Venereal Diseases, Third Edition. The Williams and Wilkins Company, Baltimore, Maryland.
39. Kjellander, J. 0. and M. Finland. I963. Penicillin treatment of gonorrheal urethritis. Effects of penicillin susceptibility of causative organism and concoraitant presence of penicillin-ase-producing bacteria on results. New England J. Med. 2691834-836.
40. Koss, L. G. 1961. Diagnostic Cytology, pages 143-164. J. B. Lippincott Company, Philadelphia, Pa.
41. Lagerholm, B. and A. Lodin. I966. Cytotnorpholic anaiysis of smears from the male urcthra in urethritis of gonococcal and non-gonococcal origin. Acta derm-venereol. 46i456-459.

65
42. Laird, S. M. 1963. Some current aspects of the epidemiology of gonorrhea. Brit. J. Vener. Dis. 39t101-104.
43. Lamanna, C. and M. F. Mallette. 1965. Mechanism of the gram stain, pages 17O-I7I. In C. Lamanna and M. F. Mallette, Basic Bacteriologyi Its Biological and Chemical Background. The Williams and Wilkins Company, Baltimore, Maryland.
44. Lankford, C. E. and R. K. Skaags. 1946. Cocarboxy-lase as a growth factor for certain strains of Neisseria gonorrhoeae. Arch. Biochem. 9i265-203.
45. Lankford, C. E. and E. E. Snell. 1943. Glutamine as a growth factor for certain strains of Neisseria gonorrhoeae. J. Bacteriol. 45t4l0-4ll.
46. Lodin, A., B. Nystrom and S. Rosengren. 196^. Gonorrhea in I962. Cases treated at the depart-ment of dermatology, Karolinska sjukhuset. Acta derm-venereol. 4ifi 127-129.
47. Lyght, C. E. (Ed.). 1966. Gonorrhea, pages 1455-1458. In The Merck Manual, Eleventh Edition. Merck, Sharp and Dohme Research Laboratories, Rahway, N. J.
48. Martin, J. E., T. E. Billings, J. F. Hackney and J.D. Thayer. I967. Primary isolation of Neisseria gonorrhoeae with a new commercial raedlum. •pub. Health Rep. 82i36l-363.
49. Martin, J. E. and A. Lester. 1971. Transgrow, a medium for transport and gro\Tth of Neisseria gonorrhoeae and Neisseria meninp;iticUs.'" HSMHA HeaÍth Reports 86i30-33.
50. Martin, J. E., W. L. Peacock and J. D. Thayer. 1965. Further studies with a selective medium for

66
cultivating Neisseria gonorrhoeae. Brit. J. Vener. Dis. li 199-201.
51. Moller, V. and A. Reyn. I965. A new solid medium for isolation of N. gonorrhoeae. Bull. W. H. 0. 321471-476. ""
52. Morse, S. A. and L. Bartenstein. 1974. Factors affecting autolysis of Neisseria gonorrhoeae. Proc. Soc. Exp. Biol. Med. l tl ití-lífZl.
53» Morse, S. A., S. Stein and J. Hines. 1974. Glucôse metaboiism in Neisseria gonorrhoeae. J. Bacteriol. 120t?0^-7l4.
54. Mueller, J. H. and J. Hinton. 194l. A protein-free medium for primary isolation of the gonococcus and meningococcus. Proc. Soc. Exp. Biol. Med. 481330-333.
55» Nargaard, 0. 1956. Gonorrhea treated in venereolog-ical outpatient department during the years 1950-55. Acta derm-venereol. 361150-153.
56. Nicol, C. S. 1948. Gonococci infections of the rectum in females. Brit. J. Ven. Dis. 24126-39.
57. O'Connell, C. J. 1973* The venereal diseases, pages 84-85. In 0. J. O'Donnell, Laboratory Diagnosis of Infectious Diseases. Medical Examiner's Publication Company, Inc, Flushing, N. Y.
58. Olsen, G. A. 1971. Value of vaginal and rectal cultures in the diagnosis of gonorrhea with special reference to areas with limited medical facilities. Brit. J. Vener. Dis. 47i102-106.
59. Pariser, H. and A. D. Farmer. I968. Diagnosis of gonorrhea in the asymptomatic femalet compar-' ison of slide and culture techniques. South. Med. J. 6lt505-506.

67
60. Phillips, J., D. Humphrey, A. Middelton and C. S. Nicol. 1972. Diagnosis of gonorrhea by cul-ture on a selective medium containing vancomycin, colistan, nystatin and trimethyl-prim (VCNT)t a comparison with gram staining and immunofluorescence. Brit. J. Vener. Dis. 481287-292.
61. Punsalang, A. P. and W. D. Sawyer. 1973. Role of pili in the virulence of Neisseria gonorrhoeae. Infec. and Immunity 8t255-263.
62. Putt, F. A. 1972. The Papanicolaou method, pages 7-9. In F. A. Putt, Manual of Histopathol-ogic Staining Methods. John Wiley and Sons, Ltd., New York, N. Y.
63. Reyn, A. I96I. Current problems in the diagnosis and therapy of gonorrhea from the laboratory point of view. Acta derm-venereol. 43t380-401.
64. Reyn, A. 1965. Laboratory diagnosis of gonococcal infections. Bull. W. H. 0. 32i^^9-469.
65. Reyn, A. I969. Recent developments in the labor-atory diagnosis of gonococcal infections. Bull. W. H. 0. 4O1245-255.
66. Reynolds, P. E. I96I. Studies on the mode of action of vancomycin. Biochem. Biophys. Acta 52t403-405.
67. Romano, A. H., S. J. Eberhard, S. L. Dingle and T. D. McDowell. 1970. Distribution of the phosphoenolpyruvateglucose phosphotransferase system in bacteria. J. Bacteriol. 104|800-813.
68. Shapiro, L. H. 1962. Endocervical curettementi a newer technique in the diagnosis of gonorrhea in women. Am. J. Obstet. and Gynec. 841 1806-1808.

68
69» Simpson, W. G. and W. J. Brown. 1962. Current status of the diagnosis and management of gonorrhea. J. Amer. Med. Assoc. 182t63-66.
70. Swanson, J. 1973» Studies on gonococcus infection. IV. Pilit Their role in attachment of gonococci to tissue culture cells. J. Exp. Med. 137t571-589.
71. Swanson, J., S. J. Kraus and E. C. Gotschlich. 1971. Studies on gonococcus infection. I. Pili and zones of adhesiont their relation to gonococcal growth patterns. J. Exp. Med. 134t8tí6-905.
72. Talley, R. S. and C. L. Baugh. 1975. Effects of bicarbonate on growth of Neisseria gonorrhoeae t replacement of gaseous carbon dioxide atmosph-ere. Appl. Microbiol. 29t^69-471.
73. Thayer, J. D. and J. E. Martin. 1964. A selective medium for cultivation of Neisseria gonorrhoeae ^^ Neisseria meningitidis. Pub. Health. Rep. 791^9-57.
74. Thayer, J. D. and J. E. Martin. 1966. Improved medium selective for the cultivation of Neisseria fíonorrhoeae and Neisseria menin^itidis. Pub. Health Rep. 81i559-56FI
75. Thayer, J. D., M. I. Perry, H. J. Magnuson and W. Garson. 1957. Failure of penicillin to kill phagocytized Neisseria gonorrhoeae in tissue culture. Antibiotics ajid Chemotherapy 7i3 l-3l4.
76. Tonhazy, N. E. and M. J. Pelczar. 1953. Oxidation of amino acids and compounds associated with the tricarboxylic acid cycle by Keisseria gonorrhoeae. J. Bacteriol. 651368-37?.
77. Ward, M. E. and P. J. Watt. 1972. Adherence of Neisseria ^onorrhoeae to urethral mucosal

69
cellsi An electron microscopic study of human gonorrhea. J. Infec. Dis. 126t601-605.
78. Watt, P. J., A. A. Glynn and M. E. Ward. 1972. Maintenance of virulent gonococci in laborat-ory culture. Nature (New Biology) 236tl86.
79. Wilcox, R. P. 1972. A world look at venereal diseases. Med. Clin. North. Am. 56tl057-1071.
80. Williams, R. P. and R. D. Wende. 1972. "Anaerobic" growth of gonococci and candle jar. J. Am. Med. Assoc. 222t212.
81. Winter, D. B. and S. A. Morse. 1975. Physiology and metabolism of pathogenic Neisseriat Partial characterization of the respiratory chain of Neisseria gonorrhoeae. J. Bacteriol. 1231631-633;
82. Wistreich, G. A. and R. F. Baker. 1971. The presence of fimbriae (pili) in three species oí" Neisseria. J. Gen. Microbiol. 651I67-I73.
83. V/olfgang, K., D. P. Joklik and D. T. Smith. 1972. Neisseria gonorrhoeae and gonoccal infections, pages 409-^15. In K. Wolfgang, D. P. Joklik and D. T. Smith (eds.), Microbiology, Fifteenth Edition. Appleton-Century Crofts, New York, N. Y.

••mmmv^
APPENDIX
70

71
APPENDIX
PROTOCOL FOR PREPARATION OF PAPANICOLAOU SMEARS
Introduction. This study is designed to evaluate
the efficacy of combining the routine Papanicolaou smear
with a microscopic examination for Neisseria gonorrhoeae.
It is felt that the routine nature of the Papanicolaou
Test makes it an ideal time for simultaneous screening
for gonorrhea. These trials will serve as data for the
development of rapid, easily performed screening test in
asymptomatic OB patients.
Suggested Procedure. You will be given packets
of two slides, one labelled PAP and one G. For female
patients presenting themselves for gonorrhea testing,
an additional two specimens will be taken. Using stand-
ard procedures for taking cervical smears with a wooden
"mutton-bone" spatula, collect two samples and alternat-
ely place them on the two glass slides. Before the
sample on the slide labelled PAP can dry, spray with a
cytology fixative. Allow the G slide to air dry. Place
both slides back into the cardboard packets and place in
the refrigerator. Slide packets should be labelled v ith
an identification number corresponding to the patient so
culture results can be compared with the microccopic
findings.

/





























