Embed Size (px)
Citation preview

Plant Physiol. (1988) 87, 50-570032-0889/88/87/0050/08/$O1 .00/0
Growth of the Maize Primary Root at Low Water Potentials'I. SPATIAL DISTRIBUTION OF EXPANSIVE GROWTH
Received for publication September 17. 1987 and in revised form January 13, 1988
ROBERT E. SHARP*2, WENDY KUHN SILK, AND THEODORE C. HsIAODepartment of Land, Air and Water Resources, University of California, Davis, California 95616
ABSTRACT
Seedlings of maize (Zea mays L. cv WF9 x Mo 17) were grown invermiculite at various water potentials. The primary root continued slowrates of elongation at water potentials which completely inhibited shootgrowth. To gain an increased understanding of the root growth response,we examined the spatial distribution of growth at various water potentials.Time lapse photography of the growth of marked roots revealed thatinhibition of root elongation at low water potentials was not explained bya proportional decrease in growth along the length of the growing zone.Instead, longitudinal growth was insensitive to water potentials as low as-1.6 megapascal close to the root apex, but was inhibited increasinglyin more basal locations such that the length of the growing zone decreasedprogressively as the water potential decreased. Cessation of longitudinalgrowth occurred in tissue of approximately the same age regardless ofspatial location or water status, however. Roots growing at low waterpotentials were also thinner, and analysis revealed that radial growthrates were decreased throughout the elongation zone, resulting in greatlydecreased rates of volume expansion.
anism by which osmotic adjustment occurs in roots growing atlow q. Specifically, our aim is to determine the extent to whichthe maintenance of lower 4i, in the growing zone under waterdeficits can be attributed to increased rates of solute deposition,or to reduced growth and hence slower rate of osmoticum di-lution by volume expansion. In this paper, attention is focusedon the effects of low qf,y on the spatial distribution of expansivegrowth rate. Although the spatial growth pattern at high q,, forroots of maize and other species has been well characterized formany years (8, 11), the extent to which the pattern may changeat low q is not known. Indeed, despite early recognition thatknowledge of how plant growth patterns may be altered by en-vironmental variation facilitates the opportunities to understandthe regulation of the growth response (11), relatively little in-formation of this kind is available. Here, we show that bothlongitudinal and radial growth patterns are altered markedly inroots growing at low qi,. In a forthcoming paper (RE Sharp, TCHsiao, WK Silk, unpublished data), we combine this informationwith profiles of qi5 and component solutes to determine effectsof low q on solute deposition rates in the root growing zone,and evaluate the relationship of the growth and solute depositionresponses to osmotic adjustment.
Plant growth is generally decreased when soil water is limited.Root growth is often less inhibited than shoot growth (2, 21),however. A recent study of maize has shown that root growthis intrinsically less sensitive than growth of the aerial plant partsto low water potentials (q') of the growing region (30), indicatingsome form of internal regulation. Root elongation is of obviousadvantage to plants in drying soil, and may be particularly im-portant for seedling establishment because of the vulnerabilityof surface soil layers to drying. Therefore, the mechanisms reg-ulating the expansive growth of roots at low +,, are importantprocesses to understand.
Previous studies have shown that, as water deficits develop,there is a substantial capacity for osmotic adjustment in the grow-ing zone of roots (12, 21, 30). This process helps to maintainturgor as q decreases in the growing cells, and is thereforeconsidered to be an important factor in the maintenance of rootelongation in drying soil. In a recent report, Silk et al. (25) appliedprinciples of growth kinematics (22) to determine the relationshipbetween the distribution of elongation growth and the profilesof osmotic potential (4i) and osmoticum deposition rate in pri-mary roots of maize. Their results, confined to roots at high qi",show that osmoticum deposition is locally synchronized with growthto maintain constant fr, along the length of the growing zone.
Using a similar approach, the present study addresses the mech-
I Supported by National Science Foundation Grant, DCB 8417504 toW. K. S. and T. C. H.
2 Present address: Department of Horticulture, University of Missouri,Columbia, MO 65211.
MATERIALS AND METHODS
Plant Culture. Seeds of Zea mays L. (cv WF9 x Mo 17) werewashed in 0.26% NaOCl for 10 min, rinsed for several minutes,and germinated at 29°C in vermiculite well moistened with 10'M CaCl2. Approximately 30 h after planting, seedlings with ra-dicles about 5 mm long were transplanted into Plexiglas boxeslined with transparent plastic film and containing vermiculitemixed with various amounts of 10-4 M CaCl to give vermiculiteq ranging from -0.03 to -2.0 MPa. The wetted vermiculitewas thoroughly mixed in a closed plastic bag prior to transplant-ing. For most experiments, four vermiculite water contents wereused (Table I). Seedlings were positioned against a face angledat 30 from the vertical, and grown at 29°C in a humid chamber(near saturation humidity) in darkness, except for exposure todim light during marking as described below.Growth Measurements. Primary root elongation rates after
transplanting were determined by periodically marking the po-sition of the root apices on the Plexiglas surface. When approx-imately 5 cm long, roots selected for uniformity of elongationrate (within + 15% of the mean per box of 20 seedlings) weremarked for time-lapse photographic growth analysis. Inside ahumid chamber, the Plexiglas face against which the roots weregrowing was removed, and a flap was cut in the plastic liner andcarefully pulled back to expose the roots. This procedure min-imized disturbance of the roots and vermiculite, whereas in theabsence of the liner roots growing at low , were found to adhereto the Plexiglas. The roots were then marked gently with anultrafine ball pen ('Uni-Ball,' Faber Castell, West Germany) atapproximately 1 mm intervals for 12 mm from the root apex.
50
Dow
nloaded from https://academ
ic.oup.com/plphys/article/87/1/50/6083204 by guest on 15 February 2022
SPATIAL DISTRIBUTION OF ROOT GROWTH AT LOW WATER POTENTIALS
Table I. Dry Weights of Roots and Shoots Grown at Different Vermiculite Water ContentsData from seedlings sampled 40 h after transplanting. The different treatments represent the vermiculite water contents as percentages of the
water content at high water potential. Water potentials are means ± 1 SD of 4 to 8 experiments. Dry weights are means ± 1 SD of 10 samples.Root: Shoot
Treatment Vermiculite Vermiculite Root Dry Shoot Dry Dry WeightWater Content Water Potential Weight Weight Ratio
go g-g-gl MPa mg100 3.72 -0.03 ± 0.00 21.8 ± 4.4 15.0 ± 1.3 1.4516 0.60 -0.20 ± 0.01 14.4 ± 1.3 4.5 ± 1.2 3.204 0.15 -0.81 ± 0.06 8.1 ± 2.1 1.4 ± 1.0 5.792 0.08 -1.60 ± 0.15 6.6 ± 2.0 1.2 ± 0.9 5.50
The liner and Plexiglas face were replaced, and the boxes werereturned to the incubator. After 1 h, a series of five photographswere taken at 15 min intervals using a green 'safe-flash' andKodak Technical Pan film. Elongation rates after marking werecalculated from the transparencies by projecting the image andmeasuring the displacement of the root tips over the 1 h period.For analysis of spatial growth patterns, enlarged prints weremade of the apical 10 mm of five roots of each treatment whichshowed minimal effects of the marking procedure on elongation.For the determination of dry weights, whole roots and shoots
were excised at the seed junction and oven-dried to constantweight.
Numerical Methods. Methods for spatial growth analysis wereas described by Silk et al. (23) and Silk and Abou Haidar (24).Computer assisted digitization of mark displacement on eachseries of photographs provided longitudinal growth velocities, v,(mm h- 1), as a function of distance from the root tip, z (mm).Longitudinal strain rates, dv,l8z (h-1), were calculated usingErickson's five-point differentiating formula (7) applied to valuesof velocity interpolated to equally spaced positions, 0.5 mm apart,using cubic splines. Radial and tangential components of thestrain rate were calculated as (2/R)(v_)(OR/dz), where R is rootradius (mm). Radii were measured directly from the photo-graphic prints. Addition of the strain rate components gave thetotal strain rate, or local relative elemental rate of volume in-crease (calculated as mm3 mm-3 h- 1). Multiplication by rootvolume per unit length (mm3.mm- 1) gave the local rate of rootvolume increase (mm3.mm -h- 1). Volumes were calculated forroot segments of 0.5 mm length assuming each segment to be asection of a circular cone. Growth trajectories (plot of elementposition versus time) were estimated by numerical integration,as described by Silk et al. (23).
Vermiculite f,. At the end of the experiments, the vermiculitein each box was thoroughly remixed and sampled for duplicatemeasurements of qi,, by isopiestic thermocouple psychrometry(3). Limited tests showed that the decrease in vermiculite u/,from the time of transplanting to the end of the experiments wasless than 0.1 MPa at the lowest water contents used, and neg-ligible at high water contents.
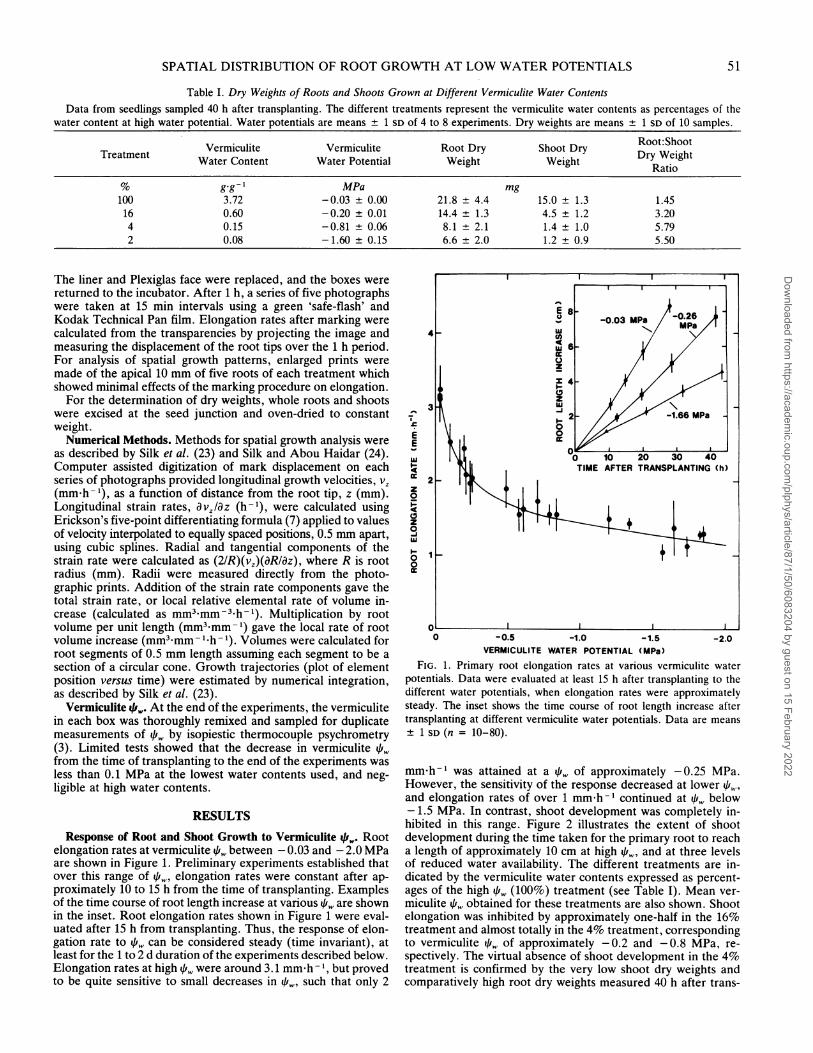
RESULTSResponse of Root and Shoot Growth to Vermiculite 4k, Root
elongation rates at vermiculite /i, between - 0.03 and - 2.0 MPaare shown in Figure 1. Preliminary experiments established thatover this range of pw, elongation rates were constant after ap-proximately 10 to 15 h from the time of transplanting. Examplesof the time course of root length increase at various /i, are shownin the inset. Root elongation rates shown in Figure 1 were eval-uated after 15 h from transplanting. Thus, the response of elon-gation rate to i can be considered steady (time invariant), atleast for the 1 to 2 d duration of the experiments described below.Elongation rates at high fr, were around 3.1 mmh-h, but provedto be quite sensitive to small decreases in 4if, such that only 2
A - 1\ 2 < -1.66 MPa -
E0I/.I
E o V 'LU 4 0 10 20 30 40
TIME AFTER TRANSPLANTING (h)
z20z40-i
0 -0.5 -1.0 -1.5 -2.0VERMICULITE WATER POTENTIAL (MPa)
FIG. 1. Primary root elongation rates at various vermiculite waterpotentials. Data were evaluated at least 15 h after transplanting to thedifferent water potentials, when elongation rates were approximatelysteady. The inset shows the time course of root length increase aftertransplanting at different vermiculite water potentials. Data are means± 1 SD (n = 10-80).
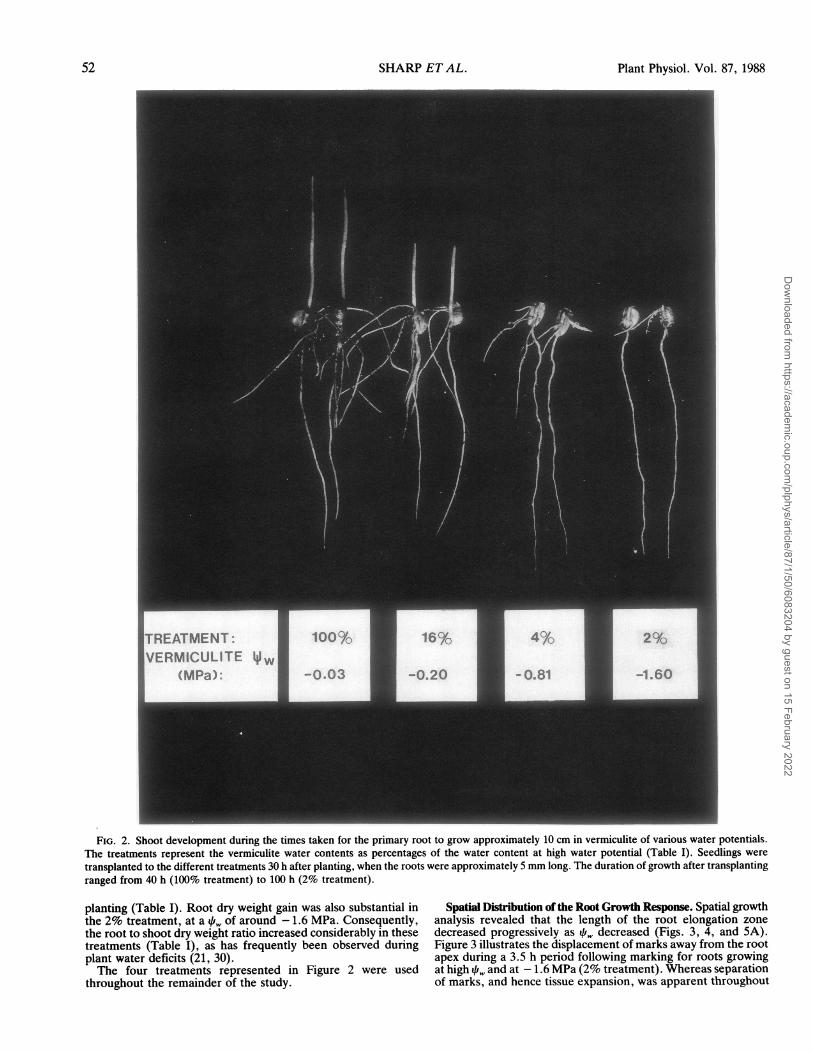
mm h-I was attained at a qi,i of approximately -0.25 MPa.However, the sensitivity of the response decreased at lower 11!and elongation rates of over 1 mm h-I continued at qi, below- 1.5 MPa. In contrast, shoot development was completely in-hibited in this range. Figure 2 illustrates the extent of shootdevelopment during the time taken for the primary root to reacha length of approximately 10 cm at high i, and at three levelsof reduced water availability. The different treatments are in-dicated by the vermiculite water contents expressed as percent-ages of the high 4fr, (100%) treatment (see Table I). Mean ver-miculite q,w obtained for these treatments are also shown. Shootelongation was inhibited by approximately one-half in the 16%treatment and almost totally in the 4% treatment, correspondingto vermiculite /i, of approximately -0.2 and -0.8 MPa, re-spectively. The virtual absence of shoot development in the 4%treatment is confirmed by the very low shoot dry weights andcomparatively high root dry weights measured 40 h after trans-
51
Dow
nloaded from https://academ
ic.oup.com/plphys/article/87/1/50/6083204 by guest on 15 February 2022
Plant Physiol. Vol. 87, 1988
FIG. 2. Shoot development during the times taken for the primary root to grow approximately 10 cm in vermiculite of various water potentials.The treatments represent the vermiculite water contents as percentages of the water content at high water potential (Table I). Seedlings weretransplanted to the different treatments 30 h after planting, when the roots were approximately 5 mm long. The duration of growth after transplantingranged from 40 h (100% treatment) to 100 h (2% treatment).
planting (Table I). Root dry weight gain was also substantial inthe 2% treatment, at a q of around - 1.6 MPa. Consequently,the root to shoot dry weight ratio increased considerably in thesetreatments (Table I), as has frequently been observed duringplant water deficits (21, 30).The four treatments represented in Figure 2 were used
throughout the remainder of the study.
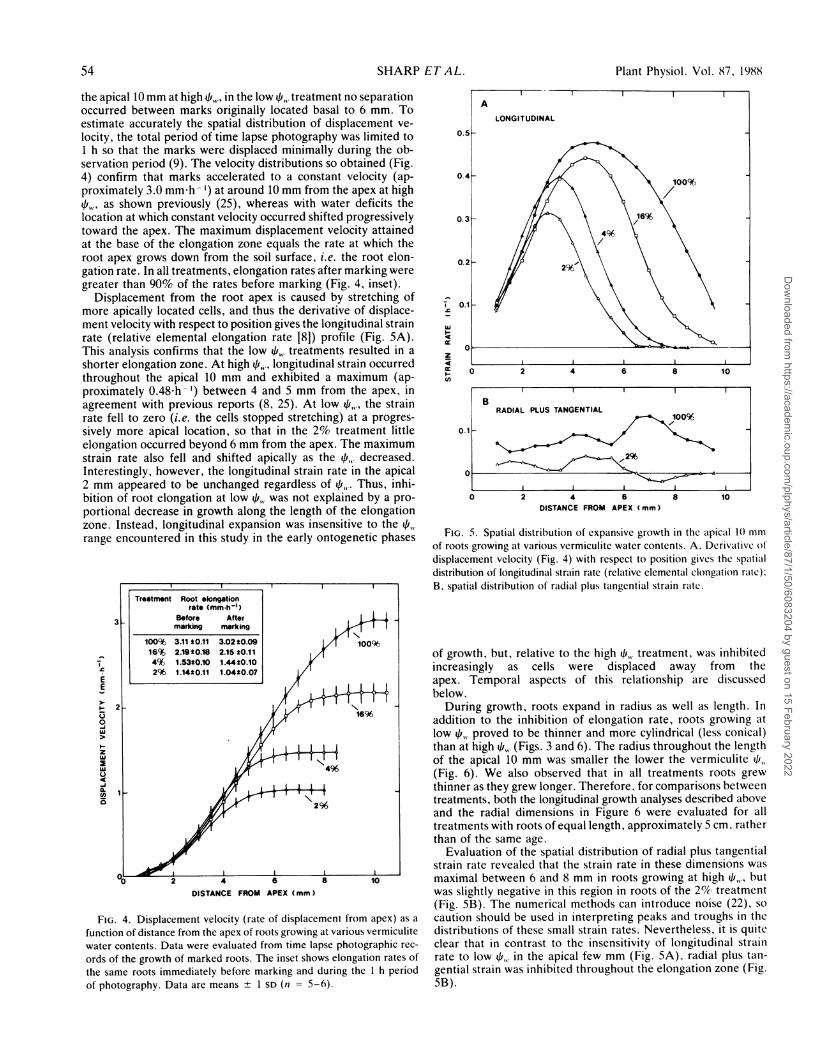
Spatial Distribution of the Root Growth Response. Spatial growthanalysis revealed that the length of the root elongation zonedecreased progressively as qi, decreased (Figs. 3, 4, and 5A).Figure 3 illustrates the displacement of marks away from the rootapex during a 3.5 h period following marking for roots growingat high , and at - 1.6 MPa (2% treatment). Whereas separationof marks, and hence tissue expansion, was apparent throughout
52 SHARP ET AL.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/87/1/50/6083204 by guest on 15 February 2022

SPATIAL DISTRIBUTION OF ROOT GROWTH AT LOW WATER POTENTIALS
VERMICULITE Yw -1.60 MPa
FIG. 3. Primary roots growing at high (100% treatment) and low (2% treatment) vermiculite water contents immediately after marking (leftphotographs) and 3.5 h after marking (right photographs). Mean vermiculite water potentials obtained for these treatments are indicated. Thewhite lines illustrate the vertical displacement of the root apices and of marks originally located 5 and 10 mm from the apices.
53
Dow
nloaded from https://academ
ic.oup.com/plphys/article/87/1/50/6083204 by guest on 15 February 2022
Plant Physiol. Vol. 87, 1988
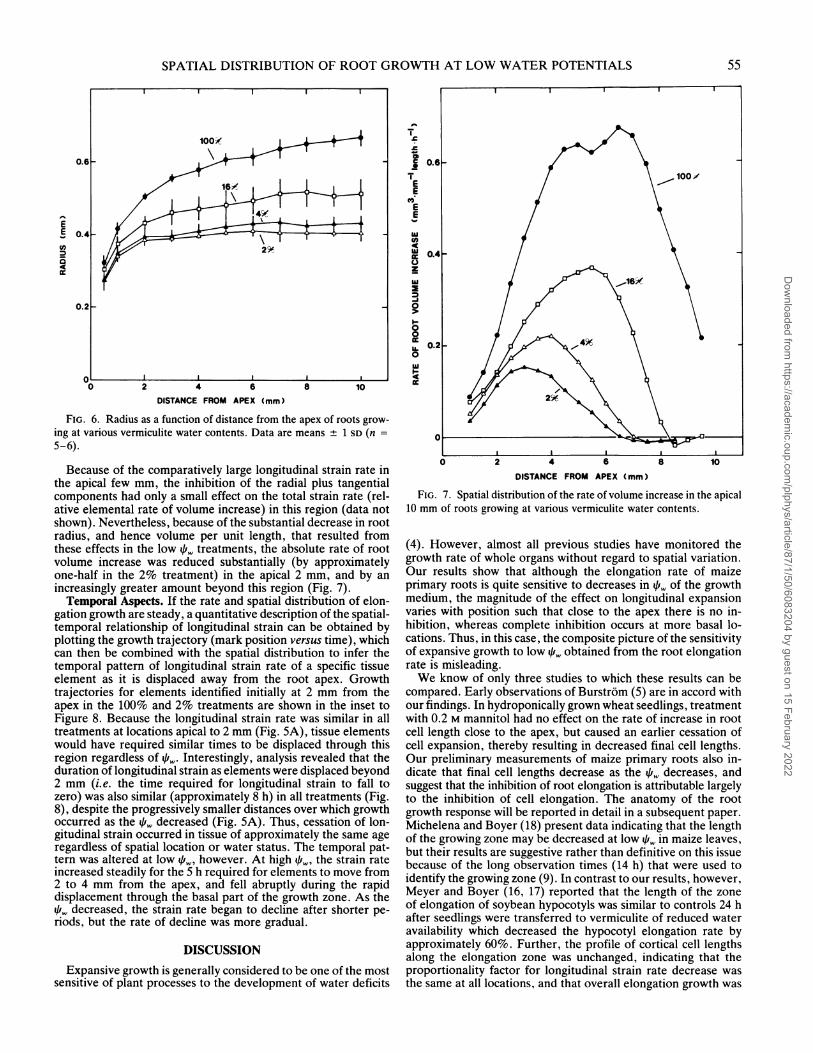
the apical 10 mm at high 41,, in the low qf,,. treatment no separationoccurred between marks originally located basal to 6 mm. Toestimate accurately the spatial distribution of displacement ve-locity, the total period of time lapse photography was limited to1 h so that the marks were displaced minimally during the ob-servation period (9). The velocity distributions so obtained (Fig.4) confirm that marks accelerated to a constant velocity (ap-proximately 3.0 mm-h ') at around 10 mm from the apex at high41k4, as shown previously (25), whereas with water deficits thelocation at which constant velocity occurred shifted progressivelytoward the apex. The maximum displacement velocity attainedat the base of the elongation zone equals the rate at which theroot apex grows down from the soil surface, i.e. the root elon-gation rate. In all treatments, elongation rates after marking weregreater than 90% of the rates before marking (Fig. 4, inset).
Displacement from the root apex is caused by stretching ofmore apically located cells, and thus the derivative of displace-ment velocity with respect to position gives the longitudinal strainrate (relative elemental elongation rate [8]) profile (Fig. SA).This analysis confirms that the low i,,. treatments resulted in ashorter elongation zone. At high 41k, longitudinal strain occurredthroughout the apical 10 mm and exhibited a maximum (ap-proximately 0.48h -') between 4 and 5 mm from the apex, inagreement with previous reports (8, 25). At low 41,., the strainrate fell to zero (i.e. the cells stopped stretching) at a progres-sively more apical location, so that in the 2% treatment littleelongation occurred beyond 6 mm from the apex. The maximumstrain rate also fell and shifted apically as the 41,,. decreased.Interestingly, however, the longitudinal strain rate in the apical2 mm appeared to be unchanged regardless of 4,,.. Thus, inhi-bition of root elongation at low 4,,, was not explained by a pro-portional decrease in growth along the length of the elongationzone. Instead, longitudinal expansion was insensitive to the 41,.,range encountered in this study in the early ontogenetic phases
i
Treatment Root elongationrate (mm-h-1)
Before Aftermarking marking
100% 3.11 t0.11 3.02 t0.09 100%616% 2.19t0.18 2.15 20.114% 1.53t0.10 1.44t0.10
F | 296 1.1420.11 1.04*0.07EE
> 0 22
0
FIG. 4.Dslcmn4vlct9rt6oipaeet rmae)a
I-~~ ~ ~ ~ ~ ~~~-
~~~~~~~~~~296
0 2 4 6 8 10
DISTANCE FROM APEX (mm)
FIG. 4. Displacement velocity (rate of displacement from apex) as a
function of distance from the apex of roots growing at various vermiculitewater contents. Data were evaluated from time lapse photographic rec-
ords of the growth of marked roots. The inset shows elongation rates ofthe same roots immediately before marking and during the 1 h periodof photography. Data are means + I SD (n = 5-6).
f-w4
I-U,
IB
RADIAL PLUS TANGENTIAL
0.1 -
IOI0 2 4 6
DISTANCE FROM APEX (mm)8 10
FIG. 5. Spatial distribution of expansive growth in the atpicial 1() mmof roots growing at various vermiculite water contents. A, Derivativc ofdisplacement velocity (Fig. 4) with respect to position givcs the spatialdistribution of longitudinal striain rate (relative elementail clongaltion ralte):B, spatial distribution of radiail plus taingential strain raitc.
of growth, but, relative to the high 41,,. treatment, was inhibitedincreasingly as cells were displaced away from theapex. Temporal aspects of this relationship are discussedbelow.During growth, roots expand in radius as well as length. In
addition to the inhibition of elongation rate, roots growing atlow 41,. proved to be thinner and more cylindrical (less conical)than at high 1,. (Figs. 3 and 6). The radius throughout the lengthof the apical 10 mm was smaller the lower the vermiculite ,1.(Fig. 6). We also observed that in all treatments roots grewthinner as they grew longer. Therefore, for comparisons betweentreatments, both the longitudinal growth analyses described aboveand the radial dimensions in Figure 6 were evaluated for alltreatments with roots of equal length, approximately 5 cm, ratherthan of the same age.
Evaluation of the spatial distribution of radial plus tangentialstrain rate revealed that the strain rate in these dimensions wasmaximal between 6 and 8 mm in roots growing at high 4,,, butwas slightly negative in this region in roots of the 2% treatment(Fig. SB). The numerical methods can introduce noise (22), socaution should be used in interpreting peaks and troughs in thedistributions of these small strain rates. Nevertheless, it is quiteclear that in contrast to the insensitivity of longitudinal strainrate to low 1,, in the apical few mm (Fig. SA), radial plus tan-gential strain was inhibited throughout the elongation zone (Fig.SB).
54 SHARP ETAL.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/87/1/50/6083204 by guest on 15 February 2022
SPATIAL DISTRIBUTION OF ROOT GROWTH AT LOW WATER POTENTIALS
0.6e
EE 0.40U)a
0 2 4 6DISTANCE FROM APEX (mm)
8 10
FIG. 6. Radius as a function of distance from the apex of roots grow-
ing at various vermiculite water contents. Data are means + 1 SD (n =
5-6).
Because of the comparatively large longitudinal strain rate inthe apical few mm, the inhibition of the radial plus tangentialcomponents had only a small effect on the total strain rate (rel-ative elemental rate of volume increase) in this region (data notshown). Nevertheless, because of the substantial decrease in rootradius, and hence volume per unit length, that resulted fromthese effects in the low qi,, treatments, the absolute rate of rootvolume increase was reduced substantially (by approximatelyone-half in the 2% treatment) in the apical 2 mm, and by anincreasingly greater amount beyond this region (Fig. 7).Temporal Aspects. If the rate and spatial distribution of elon-
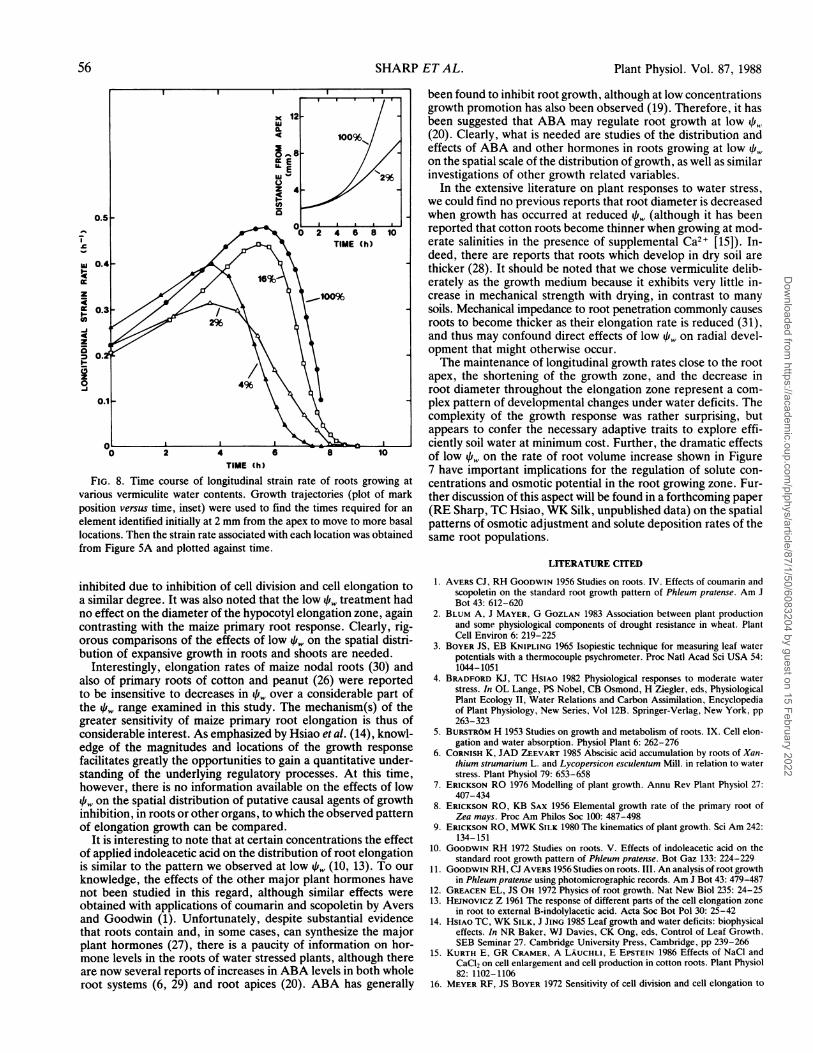
gation growth are steady, a quantitative description of the spatial-temporal relationship of longitudinal strain can be obtained byplotting the growth trajectory (mark position versus time), whichcan then be combined with the spatial distribution to infer thetemporal pattern of longitudinal strain rate of a specific tissueelement as it is displaced away from the root apex. Growthtrajectories for elements identified initially at 2 mm from theapex in the 100% and 2% treatments are shown in the inset toFigure 8. Because the longitudinal strain rate was similar in alltreatments at locations apical to 2 mm (Fig. 5A), tissue elementswould have required similar times to be displaced through thisregion regardless of 4if,. Interestingly, analysis revealed that theduration of longitudinal strain as elements were displaced beyond2 mm (i.e. the time required for longitudinal strain to fall tozero) was also similar (approximately 8 h) in all treatments (Fig.8), despite the progressively smaller distances over which growthoccurred as the 4i, decreased (Fig. SA). Thus, cessation of lon-gitudinal strain occurred in tissue of approximately the same ageregardless of spatial location or water status. The temporal pat-tern was altered at low q',, however. At high f, the strain rateincreased steadily for the 5 h required for elements to move from2 to 4 mm from the apex, and fell abruptly during the rapiddisplacement through the basal part of the growth zone. As the
decreased, the strain rate began to decline after shorter pe-riods, but the rate of decline was more gradual.
DISCUSSION
Expansive growth is generally considered to be one of the mostsensitive of plant processes to the development of water deficits
0-
T0.6
T 2 4 6 0 1
EE
E-E
0)cc 0.4 9
0i-
~~0.2 ~ ~ 2
0
0 2 4 6 8 10
DISTANCE FROM APEX (mm)
FIG. 7. Spatial distribution of the rate of volume increase in the apical10 mm of roots growing at various vermiculite water contents.
(4). However, almost all previous studies have monitored thegrowth rate of whole organs without regard to spatial variation.Our results show that although the elongation rate of maizeprimary roots is quite sensitive to decreases in ui, of the growthmedium, the magnitude of the effect on longitudinal expansionvaries with position such that close to the apex there is no in-hibition, whereas complete inhibition occurs at more basal lo-cations. Thus, in this case, the composite picture of the sensitivityof expansive growth to low /i, obtained from the root elongationrate is misleading.We know of only three studies to which these results can be
compared. Early observations of Burstrom (5) are in accord withour findings. In hydroponically grown wheat seedlings, treatmentwith 0.2 M mannitol had no effect on the rate of increase in rootcell length close to the apex, but caused an earlier cessation ofcell expansion, thereby resulting in decreased final cell lengths.Our preliminary measurements of maize primary roots also in-dicate that final cell lengths decrease as the q,, decreases, andsuggest that the inhibition of root elongation is attributable largelyto the inhibition of cell elongation. The anatomy of the rootgrowth response will be reported in detail in a subsequent paper.Michelena and Boyer (18) present data indicating that the lengthof the growing zone may be decreased at low qi,, in maize leaves,but their results are suggestive rather than definitive on this issuebecause of the long observation times (14 h) that were used toidentify the growing zone (9). In contrast to our results, however,Meyer and Boyer (16, 17) reported that the length of the zoneof elongation of soybean hypocotyls was similar to controls 24 hafter seedlings were transferred to vermiculite of reduced wateravailability which decreased the hypocotyl elongation rate byapproximately 60%. Further, the profile of cortical cell lengthsalong the elongation zone was unchanged, indicating that theproportionality factor for longitudinal strain rate decrease wasthe same at all locations, and that overall elongation growth was
I I
I ---I Iul
55
0.2 _
Dow
nloaded from https://academ
ic.oup.com/plphys/article/87/1/50/6083204 by guest on 15 February 2022
Plant Physiol. Vol. 87, 1988
0~ ~ ~ ~~~24 6 8 10TIME~~~~IM(hh
(04
z 10
0.
0.I
TIME (h)
FIG. 8. Time course of longitudinal strain rate of roots growing atvarious vermiculite water contents. Growth trajectories (plot of markposition versus time, inset) were used to find the times required for an
element identified initially at 2 mm from the apex to move to more basallocations. Then the strain rate associated with each location was obtainedfrom Figure 5A and plotted against time.
inhibited due to inhibition of cell division and cell elongation toa similar degree. It was also noted that the low qi, treatment hadno effect on the diameter of the hypocotyl elongation zone, againcontrasting with the maize primary root response. Clearly, rig-orous comparisons of the effects of low /i, on the spatial distri-bution of expansive growth in roots and shoots are needed.
Interestingly, elongation rates of maize nodal roots (30) andalso of primary roots of cotton and peanut (26) were reportedto be insensitive to decreases in qi, over a considerable part ofthe q/', range examined in this study. The mechanism(s) of thegreater sensitivity of maize primary root elongation is thus ofconsiderable interest. As emphasized by Hsiao et al. (14), knowl-edge of the magnitudes and locations of the growth responsefacilitates greatly the opportunities to gain a quantitative under-standing of the underlying regulatory processes. At this time,however, there is no information available on the effects of lowqi, on the spatial distribution of putative causal agents of growthinhibition, in roots or other organs, to which the observed patternof elongation growth can be compared.
It is interesting to note that at certain concentrations the effectof applied indoleacetic acid on the distribution of root elongationis similar to the pattern we observed at low 4i,.. (10, 13). To ourknowledge, the effects of the other major plant hormones havenot been studied in this regard, although similar effects wereobtained with applications of coumarin and scopoletin by Aversand Goodwin (1). Unfortunately, despite substantial evidencethat roots contain and, in some cases, can synthesize the majorplant hormones (27), there is a paucity of information on hor-mone levels in the roots of water stressed plants, although thereare now several reports of increases in ABA levels in both wholeroot systems (6, 29) and root apices (20). ABA has generally
been found to inhibit root growth, although at low concentrationsgrowth promotion has also been observed (19). Therefore, it hasbeen suggested that ABA may regulate root growth at low q(20). Clearly, what is needed are studies of the distribution andeffects of ABA and other hormones in roots growing at low 4f,,,on the spatial scale of the distribution of growth, as well as similarinvestigations of other growth related variables.
In the extensive literature on plant responses to water stress,we could find no previous reports that root diameter is decreasedwhen growth has occurred at reduced qi,,, (although it has beenreported that cotton roots become thinner when growing at mod-erate salinities in the presence of supplemental Ca2+ [15]). In-deed, there are reports that roots which develop in dry soil arethicker (28). It should be noted that we chose vermiculite delib-erately as the growth medium because it exhibits very little in-crease in mechanical strength with drying, in contrast to manysoils. Mechanical impedance to root penetration commonly causesroots to become thicker as their elongation rate is reduced (31),and thus may confound direct effects of low /i, on radial devel-opment that might otherwise occur.The maintenance of longitudinal growth rates close to the root
apex, the shortening of the growth zone, and the decrease inroot diameter throughout the elongation zone represent a com-plex pattern of developmental changes under water deficits. Thecomplexity of the growth response was rather surprising, butappears to confer the necessary adaptive traits to explore effi-ciently soil water at minimum cost. Further, the dramatic effectsof low qi, on the rate of root volume increase shown in Figure7 have important implications for the regulation of solute con-centrations and osmotic potential in the root growing zone. Fur-ther discussion of this aspect will be found in a forthcoming paper(RE Sharp, TC Hsiao, WK Silk, unpublished data) on the spatialpatterns of osmotic adjustment and solute deposition rates of thesame root populations.
LITERATURE CITED
1. AVERS CJ, RH GOODWIN 1956 Studies on roots. IV. Effects of coumarin andscopoletin on the standard root growth pattern of Phleum pratense. Am JBot 43: 612-620
2. BLUM A, J MAYER, G GOZLAN 1983 Association between plant productionand some physiological components of drought resistance in wheat. PlantCell Environ 6: 219-225
3. BOYER JS, EB KNIPLING 1965 Isopiestic technique for measuring leaf waterpotentials with a thermocouple psychrometer. Proc Natl Acad Sci USA 54:1044-1051
4. BRADFORD KJ, TC HsIAO 1982 Physiological responses to moderate waterstress. In OL Lange, PS Nobel, CB Osmond, H Ziegler, eds, PhysiologicalPlant Ecology II, Water Relations and Carbon Assimilation, Encyclopediaof Plant Physiology, New Series, Vol 12B. Springer-Verlag, New York, pp263-323
5. BURSTR6M H 1953 Studies on growth and metabolism of roots. IX. Cell elon-gation and water absorption. Physiol Plant 6: 262-276
6. CORNISH K, JAD ZEEVART 1985 Abscisic acid accumulation by roots of Xan-thium strumarium L. and Lycopersicon esculentum Mill. in relation to waterstress. Plant Physiol 79: 653-658
7. ERICKSON RO 1976 Modelling of plant growth. Annu Rev Plant Physiol 27:407-434
8. ERICKSON RO, KB SAX 1956 Elemental growth rate of the primary root ofZea mays. Proc Am Philos Soc 100: 487-498
9. ERICKSON RO, MWK SILK 1980 The kinematics of plant growth. Sci Am 242:134- 151
10. GOODWIN RH 1972 Studies on roots. V. Effects of indoleacetic acid on thestandard root growth pattern of Phleum pratense. Bot Gaz 133: 224-229
11. GOODWIN RH, CJ AVERS 1956 Studies on roots. III. An analysis of root growthin Phleum pratense using photomicrographic records. Am J Bot 43: 479-487
12. GREACEN EL, JS OH 1972 Physics of root growth. Nat New Biol 235: 24-2513. HEJNOVICZ Z 1961 The response of different parts of the cell elongation zone
in root to external B-indolylacetic acid. Acta Soc Bot Pol 30: 25-4214. HSIAo TC, WK SILK, J JING 1985 Leaf growth and water deficits: biophysical
effects. In NR Baker, WJ Davies, CK Ong, eds, Control of Leaf Growth,SEB Seminar 27. Cambridge University Press, Cambridge, pp 239-266
15. KURTH E, GR CRAMER, A LAUCHLI, E EPSTEIN 1986 Effects of NaCl andCaCl2 on cell enlargement and cell production in cotton roots. Plant Physiol82: 1102-1106
16. MEYER RF, JS BOYER 1972 Sensitivity of cell division and cell elongation to
56 SHARP ET AL.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/87/1/50/6083204 by guest on 15 February 2022

SPATIAL DISTRIBUTION OF ROOT GROWTH AT LOW WATER POTENTIALS
low water potentials in soybean hypocotyls. Planta 108: 77-8717. MEYER RF, JS BOYER 1981 Osmoregulation, solute distribution, and growth
in soybean seedlings having low water potentials. Planta 151: 482-48918. MICHELENA VA, JS BOYER 1982 Complete turgor maintenance at low water
potentials in the elongation region of maize leaves. Plant Physiol 69: 1145-1149
19. PILET PE, J REBEAUD 1983 Effect of abscisic acid on growth and indolyl-3-acetic acid levels in maize roots. Plant Sci Lett 31: 117-122
20. ROBERTSON JM, RP PHARIs, YY HUANG, DM REID, EC YEUNG 1985 Drought-induced increases in abscisic acid levels in the root apex of sunflower. PlantPhysiol 79: 1086-1089
21. SHARP RE, WJ DAVIES 1979 Solute regulation and growth by roots and shootsof water-stressed maize plants. Planta 146: 319-326
22. SILK WK, RO ERICKSON 1979 Kinematics of plant growth. J Theor Biol 76:481-501
23. SILK WK, RC WALKER, J LABAVITCH 1984 Uronide deposition rates in theprimary root of Zea mays. Plant Physiol 74: 721-726
24. SILK WK, S ABOU HAIDAR 1985 Growth of the stem of Pharbitis nil: analysis
of longitudinal and radial components. Physiol Veg 24: 109-11625. SILK WK, TC HsIAO, U DIEDENHOFFEN, C MATSON 1986 Spatial distributions
of potassium, solutes, and their deposition rates in the growth zone of theprimary corn root. Plant Physiol 82: 853-858
26. TAYLOR HM, LF RATLIFF 1969 Root elongation rates of cotton and peanutsas a function of soil strength and soil water content. Soil Sci 108: 113-119
27. ToRREY JG 1976 Root hormones and plant growth. Annu Rev Plant Physiol27: 435-459
28. VARTANIAN N 1981 Some aspects of structural and functional modificationsinduced by drought in root systems. Plant Soil 63: 83-92
29. WALTON DC, MA HARRISON, P COTS 1976 The effects of water stress onabscisic acid levels and metabolism in roots of Phaseolus vulgaris and otherplants. Planta 131: 141-144
30. WESTGATE ME, JS BOYER 1985 Osmotic adjustment and the inhibition of leaf,root, stem and silk growth at low water potentials in maize. Planta 164: 540-549
31. WILSON AJ, AW ROBARDS, MJ Goss 1977 Effects of mechanical impedanceon root growth in barley, Hordeum vulgare L. Effects on cell developmentin seminal roots. J Exp Bot 28: 1216-1227
57
Dow
nloaded from https://academ
ic.oup.com/plphys/article/87/1/50/6083204 by guest on 15 February 2022










![Comparative Transcriptome Profiling of Maize Coleoptilar · Comparative Transcriptome Profiling of Maize Coleoptilar Nodes during Shoot-Borne Root Initiation1[C][W][OPEN] Nils Muthreich2,](https://img.pdfslide.net/doc/110x75/5e4d3e829c5ec8732734cd8d/comparative-transcriptome-proiling-of-maize-comparative-transcriptome-proiling.jpg)


















