Embed Size (px)
Citation preview

(CANCER RESEARCH 48. 5624-5637, October 15. 1988]
Review
Hemopoietic Growth Factors: A ReviewGeorge Morstyn and Antony W. Burgess1
Melbourne Tumour Biology Branch, Ludwig Institute for Cancer Research, Australia 3050
Abstract
Several hemopoietic growth factors have now been purified, cloned,and produced in bacteria. Granulocyte colony stimulating factor andgranulocyte-macrophage colony stimulating factor are already being usedin clinical trials. Within 12 months two more hemopoietic growth factors,macrophage colony stimulating factor (also called colony stimulatingfactor 1) and interleukin 3 (also called multi-colony stimulating factor)will be used for patient treatment. This review discusses the recentadvances in our knowledge of the molecular properties and biologicalspecificities of these factors. It is now clear that these molecules are ableto modulate selectively the activity of mature blood cells as well asstimulating the production of specific lineages of blood cells. The availability of recombinant hemopoietic growth factors purified from animalor yeast cell conditioned medium or bacteria has facilitated in vivoexperiments, as well as the clinical trials. Each of the growth factors hasa unique spectrum of biological activities and it appears that the growthfactors will enhance the recovery and function of circulating white bloodcells after cancer therapy or bone marrow transplantation.
Introduction
With the present enthusiasm for the use of biological response modifiers for cancer therapy, one could be forgiven forthinking that the activation of blood cells is a new paradigm inmolecular medicine. However, Elias Metchnikoff ( 1) and Al-moth Wright (2) eloquently noted the same possibilities manyyears ago, but our incomplete knowledge of the production,activation and biology of leukocytes has delayed the effectivetesting of this hypothesis for almost a century. During the last5 years there has been a major change in our understanding ofthe molecules which control both the production and activationof leukocytes (3-10). It is already evident that some of thesemolecules are likely to have important roles in the developmentof new therapies for cancer patients (11), for the treatment ofpatients with life-threatening infections, for wound healing, andperhaps even for the prophylactic boosting of our cellulardefense systems in aged or immunocompromised patients.
In this review we will discuss the structure, biology, and likelytherapeutic indications of some of the hemopoietic growthfactors. Since many properties of both murine and humanhemopoietic growth factors have been tabulated in a number ofrecent reviews (8, 10, 12-14), we will concentrate on the morerecent publications, attempting to highlight the properties ofthe molecules which may have relevance as therapeutic agents.We will address some of the broader concepts indicated by thiswork as well as summarize some of the recent progress whichhas been made on the genetics, synthesis, and use of thehemopoietic growth factors.
Compared to other biological response modifiers, some ofwhich have already been tested in patients, the biological and
Received 12/30/87; revised 6/14/88; accepted 7/6/88.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
' To whom requests for reprints should be addressed, at Ludwig Institute forCancer Research, P. O. Royal Melbourne Hospital, Melbourne, Victoria 3050,Australia.
chemical properties of the hemopoietic growth factors havebeen studied in depth and the clinical indications for thesefactors can be predicted with some degree of confidence. Forthe first time we are confronted with biological reagents whichshould selectively "Stimulate the phagocytes" (Shaw, 1906)
( 15). Our current knowledge suggests that these reagents shouldhave immediate applications for patients with impaired phagocyte function.
Hemopoiesis
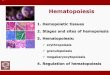
Hemopoietic Cells. The mature cells in the hemopoietic system are lymphocytes, monocytes, erythrocytes, neutrophils,eosinophils, basophils, and platelets. Each of these cells has alimited life span and only lymphocytes and monocytes arecapable of cell proliferation. All of these cells are produced asa result of the balance between self-renewal and commitmentof hemopoietic stem cells. In special situations where there isincreased loss of cells, the stem and progenitor cell compartments respond in an appropriate way by increasing the levelsof mature cells. These changes appear predominantly to beachieved by alterations in the rate of production of mature cellsand by the shifting of cells from the progenitor compartmentsto the differentiating compartments. There appears to be littlecontrol over the rate of cell destruction during normal hemo-poiesis.
There is clear evidence (Fig. 1) that the organization of thehemopoietic system is hierarchical, with multipotential stemcells leading to mature nonproliferating cells (3). The nomenclature for progenitor cells is based on the mature cells (3)which they produce. Thus, the progenitor cells are designatedas colony forming unit erythroid, BFU-e,2 GM-CFC, eosinophil
colony forming cell, and megakaryocyte colony forming cell.All of the progenitor cells (except perhaps the earliest stemcells) can be detected by in vitro culture and their proliferationis totally dependent on specific growth factors (16). The evidence for distinct subsets of progenitor cells is that they can bepartially separated from each other by velocity sedimentation,isopyknic centrifugation, and on the basis of surface markersby flow cytometry (17-22).
It is not clear whether the direction of differentiation of thestem cells is determined stochastically or whether growth regulators influence it. The studies of Koike et al. (23) and Metcalfand Burgess (24) suggest that the direction of differentiation isinfluenced by the presence and concentration of specific hemopoietic growth factors.
Progenitor cells recently identified by in vitro techniques arethe high proliferative potential colony forming cells and can bedetected by stimulation in vitro with M-CSF (also called CSF-
2The abbreviations used are: BFU-e, burst forming unit erythroid; GM-CFC,granulocyte-macrophage colony forming cell; M-CSF, monocyte colony stimulating factor; CSF, colony stimulating factor; multi-CSF, multi-colony stimulatingfactor; IL-1. -2, -3, and -4, interleukins 1, 2, 3, and 4, G-CSF, granulocytic colonystimulating factor; GM-CSF, granulocyte-macrophage colony stimulating factor;GM, granulocyte-macrophage; cDNA, complementary DNA; PWM-SCM,pokeweed mitogen stimulated spleen cell conditioned medium. HGF. hemopoieticgrowth factor; AML, acute myeloid leukemia; rG-CSF, recombinant granulocytecolony stimulating factor.
5624
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
1) and hemopoietin 1 (25-29). Nakahata et al. (30) have suggested that there is also another group of multipotential, self-renewing cells which appear to enter randomly into cycle fromGo and which can then be stimulated to produce blast cellcolonies by multi-CSF (IL-3). Perhaps hemopoietin 1 is responsible for the initial stimulation of these cells from G0 toGì(31-33). Recently it has been shown that IL-4 acts on theseearly cells (34).
Subpopulations of Granulocyte Progenitor Cells. There appears to be a hierarchy of human granulocyte-macrophageprogenitor cells with two separate growth factors stimulatingdifferent subsets of the GM-CFC. These have been operationally called Day 14 GM-CFC and Day 7 GM-CFC. There isevidence that some of the Day 14 GM-CFC are precursors ofDay 7 GM-CFC (35). Morstyn et al. (36) were able to partiallypurify these two subpopulations using lectin binding and cellsorting. The Day 14 population was enriched for myeloblastsand the Day 7 population for promyelocytes and myelocytes.Begley et al. (37), using a monoclonal antibody to mature cells,have confirmed that myelocytes and promyelocytes are a distinct subset of progenitor cells. The distinction between theDay 7 and Day 14 granulocyte progenitor cells is importantbecause they are preferentially stimulated, by G-CSF (originallycalled CSF0) and GM-CFC (formerly called CSFa) (36). Afurther complication is that Francis et al. (38) have shown thatas cells differentiate in the GM pathway they become moresensitive to conditioned media containing CSFs, i.e., to GM-CSF and G-CSF.
Hemopoietic Growth Factors
Biological Specificity. Since the discovery of the method forgrowing blood cells in culture (39, 40) four "granulocyte-macrophage" CSFs have been isolated. The first hemopoietic
growth factor was detected because of its ability to stimulatethe production of granulocytes and macrophages from murinespleen and marrow cells. The same assay system has been usedto detect the other hemopoietic growth factors (16). Whenmarrow cells are dispersed in agar at approximately 10s cells/
Stem cells
Proliferation
Progenitor cells
/ \ Proliferation
Maturationand
Activation
Mature cells
Fig. 1. Schema for the organization of the hemopoietic system. Colony stimulating factors stimulate not only the proliferation and differentiation of cells inthe myeloid lineages but also the activation of the mature cells.
ml, unless a hemopoietic growth factor is present, all of thecells die. However, if one of the hemopoietic growth factors ispresent, the early myeloid cells continue to proliferate anddifferentiate, producing a colony of mature granulocytes and/or macrophages. When human marrow is used, the coloniesrequire 7 to 14 days to develop between 50 and 5000 maturecells and the number and type of differentiated cells depend onthe concentration and species of the hemopoietic growth factor.Two names were given to the first hemopoietic growth factor:macrophage-granulocyte inducer (40) and CSF (39). Thesenames proved to be inadequate as another growth factor formurine marrow cells was also discovered (41). This moleculestimulated the production of macrophage colonies in preferenceto granulocyte colonies. Since this second hemopoietic growthfactor is also a colony stimulating factor, confusion betweenthe two molecules was inevitable. Two sets of nomenclaturewere used to distinguish between these two molecules: GM-CSF by one group (42) and CSF-2 by other groups (43); theother molecule was renamed CSF-1 or M-CSF. Most commonly, GM-CSF and CSF-1 are used to distinguish betweenthese two molecules (44).
A third hemopoietic growth factor specific for granulocyticcells was discovered in the conditioned medium from humanplacenta (45) and in the serum of mice treated with endotoxin(46). This molecule is widely known as G-CSF (47).
The fourth hemopoietic growth factor is also capable ofstimulating the production of granulocytes and macrophages.This molecule was identified by several laboratories workingon the different properties of the conditioned medium at lectinstimulated murine spleen cells (48) and the conditioned mediumfrom a "myeloid" cell line WEHI3B(D~) (49, 50). Initial obser
vations suggested that this conditioned medium induced I -lymphocyte differentiation; therefore the component responsible for this effect was named interleukin 3 (51). Subsequentinvestigations have shown that the apparent effects on T-lym-phocytes were actually associated with the production of neu-trophils and macrophages. However, it was known that thisconditioned medium stimulated the production of eosinophils,megakaryocytes, and erythroid cells as well as granulocytes andmacrophages. While Chromatographie investigations indicatedthat pokeweed mitogen stimulated spleen cell conditioned medium contained a GM-CSF, the purification data were alsocompatible with the existence of another molecule capable ofstimulating the production of all mature hemopoietic cells (48).Thus the molecule was also called multi-CSF. Indeed, it wassubsequently shown that the conditioned medium fromWEHI3B(D~) cells did not contain GM-CSF and that the
hemopoietic growth factor activity was all associated withmulti-CSF (IL-3) (50, 52). Murine multi-CSF (IL-3) was thefirst hemopoietic growth factor to be molecularly cloned (53)and the human equivalent of multi-CSF has been cloned recently (54). Its range of biological activities has not been explored in detail; however, the cDNA for gibbon IL-3 has alsobeen cloned (54) and this molecule supports the growth oferythroid and myeloid precursors in human marrow cells.
Thus, the production of granulocytes and macrophages fromthe progenitor cells of mouse marrow can be stimulated by anyone of four distinct molecules, multi-CSF (IL-3), GM-CSF, M-CSF (CSF-1), and G-CSF. There are human equivalents of allfour molecules and three of these, multi-CSF, GM-CSF, andG-CSF, have been shown to stimulate the proliferation ofleukemic progenitor cells (54-56). Other terms have been usedto describe mixtures (or impure forms) of these hemopoieticgrowth factors, e.g., CSFa, CSF/3, pluripoietin, pluripoietin a,
5625
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
and burst promoting activity (8). Usually the properties of theseconditioned media can be adequately accounted for by theproperties of one of the four CSFs. Other molecules are able tostimulate (or modify) the lineage specific proliferation and/ordifferentiation of marrow progenitor cells, e.g., interleukin 1(57), hemopoietin 1 (25), eosinophil differentiation factor (58-60), erythropoietin (61); interleukin 2 (62, 63), interleukin 4(64) and interleukin 6. We have not attempted to includedescriptions of the biology or chemistry of these molecules inthis review.
GM-CSF is known to stimulate eosinophil production (45,65) and the initial divisions of progenitor cells in the erythroidand megakaryocyte lineages (32, 66). In the mouse high concentrations of G-CSF are also able to stimulate macrophageproduction; however, only the initial proliferation of the otherhemopoietic progenitors is effected by G-CSF. M-CSF predominantly produces macrophages, but at high concentrations somegranulocytes are produced (67). M-CSF does not seem to effecteven the initial divisions of the progenitor cells in the otherhemopoietic lineages.
Another class of hemopoietic cell "response modulators"
needs to be mentioned, i.e., those molecules that appear toinfluence hemopoiesis either indirectly or in the presence ofone of the CSFs. Molecules such as IL-1 stimulate monocytesto produce hemopoietic growth factors (68). Although IL-1may not act directly on myelopoiesis, activation of the synthesisof M-CSF and GM-CSF will stimulate the production of granulocytes and macrophages. Indeed, depending on the number,distribution, and state of activation of the tissue monocytes (ormacrophages) molecules such as IL-1 could initiate a cascadewhich leads to high concentrations of the hemopoietic growthfactors and consequently the large scale activation, accumulation, and production of leukocytes.
At least two other mechanisms of amplifying the effects ofthe hemopoietic growth factors have been proposed. The overallproduction of mature cells depends on the number of availableprogenitor cells. By increasing the production of progenitorcells, the production of mature cells can be accelerated. Forexample, 'hemopoietin 1 stimulates the production of macro
phage progenitors (25) and in the presence of both hemopoietin1 and M-CSF, monocyte production can be increased considerably. Other molecules appear to act synergistically with hemopoietic growth factors, stimulating the rate of proliferationof progenitor cells in response to multi-CSF; e.g., interleukin 4stimulates the rate of multi-CSF dependent mast cell proliferation (64).
Granulocyte-Macrophage Colony Stimulating Factor. Themolecule known as GM-CSF has been purified to homogeneityfrom a human cell line (69) and murine lung conditionedmedium (70). To some extent the native GM-CSF has beenused to define the range of biological activities of GM-CSF.However, the purified proteins also provided amino acid sequence information which led to the identification and sequencing of cDNAs corresponding to both the human and murineGM-CSFs. It has been possible to express recombinant GM-
CSF for both species in COS cells (55, 71), yeast (72, 73), andEscherichia coli (74, 75). Indeed biologically active, recombinant GM-CSFs have been purified from each of these sourcesand these have been used to define some of the activities of thismolecule in vivo (11, 76).
The availability of purified GM-CSF, whether natural orrecombinant, will allow progress toward an understanding ofits structure and function. At present our knowledge of thestructure of GM-CSF is essentially limited to the amino acid
sequence. From a full length cDNA it has been possible todeduce the complete amino acid sequence of pro-GM-CSF (71)and a putative prepro-GM-CSF (77). The hypothetical prepro-GM-CSF would have a structure similar to that of a transmembrane molecule; however, in situations where this moleculewould be expected to be synthesized, only the secreted form ofthe protein has been detected. Initial amino acid sequencing onmurine GM-CSF from lung conditioned medium yielded NH2-terminal X-Thr-Val-Glu-Arg-Pro-Trp-Lys-His ... (78);i.e., the NH2 terminus commenced six residues after the expected cleavage site (71 ), between Ser29 and Alaao. The Ser-Alacleavage site is used for GM-CSF derived murine T-lympho-cytes stimulated by concanavalin A. It is clear that limited NH2-terminal truncations can be tolerated without influencing thebiological activity. Since E. coli synthesized human and murineGM-CSFs are fully biologically active in vitro we know that thecarbohydrate moieties are not necessary for either receptorbinding or activation. Studies on the serum half-life of bacte-rially synthesized murine GM-CSF indicate that the serumhalf-life (6-10 min) is similar for the native molecule.
Analysis of the structure of the GM-CSF mRNA has revealeda segment of nucleotide sequence in the 3'-untranslated section
of the mRNA which is important for controlling the breakdownof the mRNA (79). The GM-CSF gene is inducible and mRNAturnover is rapid. It is as though short bursts of synthesis ratherthan chronic production are used to control GM-CSF levels. Itmay be possible to manipulate the 3'-untranslated section ofthe cDNA to produce a GM-CSF mRNA with a longer half-life. This would lead to increased mRNA levels and presumablya more efficient animal cell production system for GM-CSF tobe developed.
There has been considerable interest in the range of biologicalactivities associated with the different hemopoietic growth factors (80, 81). The availability of large amounts of recombinantGM-CSF is allowing the biological specificity to be explored inmore detail both in vitro and in vivo. Purified murine GM-CSFstimulates a single cell to proliferate and differentiate to produce neutrophils, eosinophils, granulocytes, or macrophages(65, 82). The low frequency of eosinophil progenitor cellsinitially masked the detection of the eosinophils produced bymurine GM-CSF. Although it had not been purified, it wassuggested that human GM-CSF could also stimulate eosinophilprogenitors (83). The purification and cloning of sufficientquantities of both murine (32, 66) and human GM-CSF confirmed that these three cell lineages are all stimulated by GM-CSF (75, 80, 84).
The stimulation of isolated hemopoietic progenitor cells indicates that GM-CSF can initiate proliferation in both theerythroid and megakaryocytic lineages, but other molecules arenecessary to complete the production of mature cells in theselineages (32). The discovery of murine multi-CSF established
the concept that a single molecule could stimulate the production of all hemopoietic cells (48). Although earlier studies didnot detect megakaryocyte production in response to GM-CSF,it is now apparent that a proportion of megakaryocyte progenitors can be stimulated with either murine (66) or human (84,85) GM-CSF.
The search for an equivalent molecule produced by humancells initially led to the detection of mixtures of moleculeswhich were thought to be pluripoietins and multilineage growthfactors. In most cases these activities have proved to be mixturesof GM-CSF and G-CSF.
The biological specificity of human GM-CSF has proved tobe more difficult to determine than the corresponding murine
5626
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
molecule. One major reason for this problem is the ability of molecules. There is a particular exception to this situation;human bone marrow cells to produce growth factors whichinteract in the colony stimulating assays. Even small numbersof contaminating monocytes, T-cells, endothelial cells, orstrdiiial cells are likely to produce significant levels of moleculeswhich can synergize with and modulate the activity of humanGM-CSF (86-88). Thus, even a population of BFU-e which is50% pure may contain cells which will respond to GM-CSF toproduce molecules able to effect BFU-e proliferation. It isdifficult to ascertain whether a proportion of BFU-e is able torespond directly to GM-CSF or whether the GM-CSF "induces" the production of another molecule which acts on some
or all of the BFU-e.It should be emphasized that this "biological" specificity has
generally been determined by in vitro responses. Indeed, theanalytical efforts to determine which cells proliferate in directresponse to GM-CSF rather than those which proliferate as aresult of indirect stimulation need to be considered in theappropriate context. When GM-CSF is released in vivo, its fullrange of effects will be manifest (i.e., direct and indirect), andproliferative or differentiative responses can be expected tooccur in a wide range of cell lineages.
Although the discovery of GM-CSF occurred because of itsin vitro proliferative effects on hemopoietic progenitor cells inthe marrow and spleen, even in the first reports, it was recognized that this proliferation was associated with differentiation.During recent years there has been an increasing awareness ofthe action of GM-CSF on mature neutrophils and macrophages.It is now apparent that in the presence of initiator molecules,GM-CSF is capable of activating both neutrophils and eosino-phils (65, 75, 80).
Granulocyte-Colony Stimulating Factor. The conditioned media from mouse 1.-cells and human urine stimulate almostexclusively macrophage progenitors (89), whereas mouse lungconditioned medium stimulates both granulocyte and macrophage progenitors (66) and mitogen-stimulated spleen cellsproduce growth "factors" which produce granulocytes, macro
phages, erythroid, eosinophil, and megakaryocyte precursors(90). The CSFs in each of these preparations are heterogeneousand proved to be difficult to purify, but there were someindications that several distinctly different molecules couldstimulate granulocyte (91, 92) and macrophage progenitors.Two potent sources of GM-CSF for human bone marrow cells,human leukocytes stimulated by phytohemagglutinin and human placenta! conditioned medium, appear to contain severalCSFs. Phytohemagglutinin-stimulated human leukocytes wereseparated into a component capable of stimulating murinemacrophage progenitors and the "GM-CSF" which stimulated
human progenitor cells (93). Using phenyl-Sepharose chroma-tography, human placenta! conditioned medium was separatedinto two distinct colony stimulating factors, a and ß(45): CSFostimulated granulocyte, macrophage and eosinophil progenitors; whereas CSFßstimulated only granulocyte and macrophage progenitors (45). When used at low concentrations theCSFa preparation selectively stimulated macrophage progenitors, whereas at low concentrations CSFßpreferentially stimulated granulocyte progenitors. The human CSFa activity hadproperties similar to those of murine GM-CSF. At low concentrations the human CSFßactivity mimicked the biologicalactivity of the granulocytic specific murine G-CSF.
Most hemopoietic growth factor preparations are producedfrom tissues or cells cultured in vitro. Indeed, the concentrationof these molecules in fresh tissue specimens and body fluids isextremely low and it is difficult to detect and characterize these
when mice are given injections of endotoxin, several CSFs canbe detected in the blood. Two groups initiated studies designedto characterize the molecule(s) in endotoxin serum which wasresponsible for inducing leukemic cell differentiation (46, 94).Both groups discovered that most of the colony stimulatingactivity in endotoxin serum could be neutralized by antiseraraised against partially purified M-CSF (CSF-1) prepared fromL-cell conditioned medium (95). However, the ability of endotoxin serum to induce WEHI3B(D*) cells was not diminished
by this antiserum. Lotem et al. (94) did not detect any effect ofthe partially purified endotoxin serum differentiation factor(macrophage-granulocyte inducer 2) on normal cells. Thisgroup has developed a novel hypothesis concerning the linkbetween proliferation and differentiation, they propose thathemopoietic regulatory molecules either induce differentiationor stimulate proliferation (96). In contrast Burgess and Metcalf(46) found that the endotoxin serum differentiation factor forWEHI3B(D+) cells was associated with some colony stimulat
ing activity. All of the colonies stimulated by this CSF werecomposed of granulocytes. Thus, they suggested that a G-CSFwas responsible for the differentiation of the WEHI3B(D+)
cells. Although other differentiation factors may exist (97), itis likely that the ability of endotoxin serum to induce leukemiccell differentiation is due to the G-CSF. This molecule waspurified from mouse lung conditioned medium (47) and thepurified molecule stimulates the proliferation of normal granulocytic progenitor cells as well as the differentiation ofWEHI3B(D+) cells.
Molecular cloning and receptor competition studies havemade it clear that many conditioned media are likely to containmixtures of GM-CSF, G-CSF, and M-CSF. A particular example is the identity of pluripoietin with G-CSF and pluripoie-tin a with GM-CSF (98). Although GM-CSF and G-CSF arebecoming generally accepted terms, it is important to note thatthe cellular specificity of these two hemopoietic growth factorsis considerably broader than their names imply. Both GM-CSFand G-CSF are capable of stimulating a limited number ofdivisions early in the erythroid, megakaryocyte, and eosinophillineages (99). However, it is currently a matter of controversywhether either molecule can stimulate the production of maturecells in the erythroid and megakaryocyte lineages (100). In vitroand in vivo both GM-CSF and G-CSF can stimulate cell production in other hemopoietic lineages. However, these effectsmay be indirect (i.e., by inducing the production of other growthfactors or modulators) (11, 85). This phenomenon is still beinginvestigated in detail.
Human G-CSF has been synthesized by recombinant DNAmethods using bacteria and appears to be active on its relevanttarget cells (101, 102). Two mRNAs have been detected forhuman G-CSF; one of these mRNAs encodes 3 extra residues(101). It is likely that the larger molecule arises from a splicingartifact inasmuch as the shorter G-CSF is considerably moreactive in the bioassay (102). There appear to be no detectabledifference between the natural G-CSF (56) and the bacteriallysynthesized G-CSF in vivo. Thus the carbohydrate moiety doesnot appear to influence the interaction between G-CSF and itstarget cell. Apart from erythropoietin, there is no definitiveinformation which gives us a clue to the function of the carbohydrate moiety attached to the hemopoietic growth factors.
The homology between human and murine G-CSF is approximately 75% [cf., the homology between the murine and humanGM-CSF which is 56% (103)] and there is almost full biologicalcross-reactivity between the different species of G-CSF (104).
5627
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
At present there is insufficient structural information to indicate which parts of the G-CSF molecule are required for itsbiological activity. There is a single free sulfhydryl near theNH2 terminus (101) but this does not seem to be important forthe activity of the molecule. Peptide analogues may help todefine the binding domain by allowing the epitope for neutralizing antibodies to be defined. Once location of the binding sitecan be identified, analogues incorporating these residues can beprepared to study the linkage between binding and mitogenesis.
Macrophage-Colony Stimulating Factor. There is a high molecular weight hemopoietic growth factor, M-CSF [also calledCSF-1 by the group which achieved the first purification of thismolecule (89)], which has a propensity to stimulate the proliferation of macrophage progenitors. M-CSF has been detectedin a wide range of human and murine tissues where it is presentas a glycosylated homodimer (105). M-CSF has a variableamount of carbohydrate; human urinary M-CSF has an apparent molecular weight between 45,000 and 60,000 (43) and themolecular weight of the murine M-CSF homodimer varies from60,000 to 80,000 (43). M-CSF is active only in the dimer form;when it is dissociated by urea-mercaptoethanol or acidic ace-tonitrile the monomer is no longer active (67, 105).
M-CSF has been available only in microgram amounts andthe characterization of the important structural features hasbeen quite difficult. However, in 1985 human M-CSF wasmolecularly cloned (106). Interestingly, the open reading framefor the M-CSF cDNA predicts a polypeptide 224 amino acidslong (equivalent to a molecular weight of 26,000). There is aputative leader sequence of 32 amino acids in front of the NH2-terminal sequence determined from the purified human urinaryM-CSF. Since the size of the M-CSF polypeptide chain appearsto be 14,500, some posttranslational processing of the polypeptide must occur.
Attempts to isolate an equivalent cDNA for murine M-CSFhave proved to be even more difficult. However, one clone hasbeen isolated (107) and there is considerable homology at thelevel of amino acid sequence.
It is important to consider the range of biological activitiesof M-CSFs on mouse and human cells. Human urinary M-CSFdoes not stimulate human colony formation directly (108), butit is able to stimulate mouse marrow colony formation. Humanurinary M-CSF stimulates granulocytic clusters to form inunfractionated human marrow. This occurs by the stimulationof monocytes to produce G-CSF. Thus, the human urinary M-CSF seems to function as a stimulus for the final stages ofgranulocytic development in humans and a stimulator of theearly monocytic progenitor cells in mouse marrow. Murine M-CSF stimulates murine marrow cells to form macrophages, butdoes not stimulate human marrow cells. Even though murineM-CSF is predominantly a stimulator of murine macrophageand monocyte progenitors, there is still some stimulation ofneutrophil formation. Indeed, in serum-free cultures of individual hemopoietic progenitor cells, murine M-CSF appears to bean excellent stimulus for neutrophil-macrophage progenitors(23). M-CSF is also a powerful stimulus for mononuclearphagocytes (41) and murine monocytes require the presence ofeither GM-CSF or M-CSF for their survival (109).
Both human and murine M-CSF can induce proliferation ofmurine mononuclear phagocytes; however, although humanmononuclear phagocytes can be functionally activated by M-CSF, as yet human monocytes have not responded mitogeni-
cally to any agent.It will be interesting to study the biological effects of M-CSF
in vivo. There are strongly synergistic interactions between M-
CSF and other hemopoietic growth factors in vitro (23, 25,110). Many of these interactions should be manifest in vivo andeven small concentrations of Multi-CSF or G-CSF could beexpected to synergize with the effects of M-CSF on both progenitor cells and mature cells.
Multi-Colony Stimulating Factor (Multi-CSF, IL-3). The discovery of GM-CSF and M-CSF encouraged a search for othermolecules responsible for stem cell proliferation and productionof eosinophils, megakaryocytes, and erythroid cells. Numeroussources of GM-CSF and M-CSF were discovered, but the othercolony types were difficult to produce in vitro. It was knownthat the conditioned medium from mitogen stimulated mono-nuclear blood cells, together with erythropoietin, was able tostimulate granulocyte, macrophage, and erythroid colonies(111). Parker and Metcalf (112) produced PWM-SCM whichsupported the proliferation of progenitors for natural killercells, eosinophils, megakaryocytes, and erythroid cells. The onlyother source of growth factors for these cells was the conditioned medium from a murine myelomonocytic leukemiaWEHI3B(D~) (49). Since GM-CSF was a distinct molecular
entity, it was initially assumed that PWM-SCM andWEHI3B(D~) conditioned medium also contained distinct eo-
sinophil CSF, megakaryocyte CSF, and erythroid CSFs. However, attempts to separate these molecules from PWM-SCMinvariably yielded two classes of CSF: GM-CSF; and anotherbiologically active pool which contained a molecule(s) capableof stimulating all of the other progenitors (48, 90). Murinemulti-CSF (IL-3) was the first of the hemopoietic growth factorsto be molecularly cloned (53, 113). Factor dependent mast cellclones (114, 115, 116) were used to assay products secreted byXenopus oocytes previously treated with injections of subfrac-tions of WEHI3B(D") mRNA. The enriched mRNA was used
to prepare a cDNA library and the appropriate cDNA poolsfrom the library screened in the oocyte assay using hybridselection and elution. The availability of a full length cDNAand NH2-terminal sequences for multi-CSF indicated that considerable heterogeneity could occur at the NH2 terminus (presumably due to differential processing). Despite some limitedhomology at the signal peptide cleavage site (117), no othersignificant homologies have been detected between the predicted amino acid sequences for murine multi-CSF, GM-CSF,G-CSF, or M-CSF peptide chains.
Attempts to locate the human equivalent of multi-CSF usingthe murine multi-CSF cDNA were not successful. There is nooverlap in biological activity between murine and human IL-3.Even the rat multi-CSF (118) amino acid sequence is significantly different from murine multi-CSF.
Further analysis of the murine GM-CSF and multi-CSFcDNA clones indicated that as well as the secreted forms, thelonger clones could encode pre-CSF molecules with a numberof charged residues at the NH2 terminus. This NH2-terminalarrangement of these longer molecules would generate a ''trans-membrane-like'' domain allowing the GM-CSF or multi-CSF
to anchor in the plasma membrane. At present neither themRNA nor the protein corresponding to the transmembraneform of these two molecules has been detected.
Interactions between Hemopoietic Growth Factors. The HGFscan act synergistically; i.e., combinations of two or more of theHGFs can induce proliferation and differentiation more effectively than the individual growth factors. Until each of theHGFs had been purified it was difficult to study these synergistic effects definitively and even now it is important to note thatthere are many other potent modulators of hemopoiesis in vitro,e.g., fetal calf serum (23, 119), hemopoietin 1 (25), and IL-4
5628
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
(64). The molecular basis for these synergistic effects is poorlyunderstood. The situation will be even more complex in vivowhere the high density of cells in the marrow and spleen can beexpected to produce significant concentrations of other modulating agents (120-124).
It is simple to show the interactions between the differentgrowth factors. When murine marrow cells are stimulated byM-CSF (CSF-1) in medium containing fetal calf serum most of
the colonies contain only monocytes/macrophages. Using concentrations of M-CSF which maximally stimulate marrow colony formation, 95% of the colonies are composed of monocytes.However, when M-CSF is mixed with GM-CSF many granu-locytic and mixed granulocyte/macrophage colonies develop(110). Indeed, even when the GM-CSF is used at low concentration, as long as there is maximal stimulation by M-CSF,granulocytic colonies will form. Thus, as long as there is astrong proliferative stimulus, even a weak differentiation signalcan influence the pattern of hemopoietic differentiation.
In serum free cultures a combination of multi-CSF and M-CSF increased the size (i.e., number of cells) of both granulocyteand macrophage colonies (23). Furthermore, the number ofGM-progenitor cells responding to the combination of multi-CSF and M-CSF is greater than the sum of the number ofcolonies stimulated by the individual factors.
Hemopoietin 1 is a molecule which appears to stimulate theinitial proliferation of multipotential hemopoietic cells andearly macrophage progenitors (25), but it is not able to inducecomplete maturation. Mature cells are produced only in thepresence of the other hemopoietic regulators. There also appears to be a synergistic interaction between hemopoietin 1 andmulti-CSF (23). In the presence of both hemopoietin 1 andmulti-CSF the number of colonies formed by multipotentialprogenitors increases almost 30-fold and many of the coloniesstill contain multipotential progenitors.
The molecular basis of this synergism has not been studiedin detail, although in other cellular systems (fibroblast andepithelial) synergistic interactions have been reported to occuramong epidermal growth factor, platelet derived growth factor(125), and human transforming growth factor ß(126). There issome evidence that the synergistic interactions are modulatedat the level of the cell surface receptor via an hierarchicalnetwork (127). Thus it has been proposed that the activatedmulti-CSF and GM-CSF receptors transmodulate the M-CSFand G-CSF receptors and that it is these interactions which areresponsible for the production of granulocytes and macrophagesin the presence of Multi- or GM-CSF. By extrapolation, thegeneration of eosinophils, megakaryocytes, and erythroid cellsby multi-CSF and eosinophils by GM-CSF might be expectedto be mediated by the transmodulation of the receptors foreosinophil (59, 128, 129) megakaryocyte (130, 131), or erythroid (132) growth regulatory proteins. Receptor transmodulation could be a vital link between proliferation and differentiation. Normal progenitor cells stimulated by multi-CSF wouldbe expected to proliferate and those which acquire G-CSF orM-CSF receptors would then be induced to differentiate (evenin the absence of G-CSF or M-CSF). Multi-CSF and GM-CSF
might be capable of producing granulocytes and macrophagesat sites not normally endowed with sufficient concentrations ofM-CSF or G-CSF.
The interrelationship among three phases of hemopoietic cellproduction (i.e.. proliferation, differentiation, and activation)needs to be considered when analyzing leukemic cell populations. Most of the hemopoietic growth factors are probablycapable of influencing these different phases of hemopoiesis,
but there will be a different potency for each growth factor ateach phase. For example multi-CSF and GM-CSF appear tobe two of the preferred stimuli for expanding the early proliferation steps. Although both are also capable of inducing differentiation and activating end cell function, they may mediatethese effects through the transmodulation pathways via the M -CSF and G-CSF receptors.
In contrast, while G-CSF may influence the early proliferativeevents in hemopoiesis, this regulator appears to be a morepotent activator of neutrophil function than multi-CSF (133).In order to manipulate the hemopoietic system we will need tolearn how to use these regulators (or their antagonists) tostimulate (or inhibit) the different phases of hemopoiesis. Thelinkages between these three aspects of hemopoiesis are required to maintain the number of mature blood cells. In themurine system several myeloid cell lines have been isolatedwhich are still dependent on growth factor but appear to beblocked at different stages of differentiation. When this blockoccurs before the acquisition of G-CSF or M-CSF receptor, thecells will not differentiate, even in the presence of these regulators. However, in other cell lines such as WEHI3B(D+), GM-
CSF and G-CSF receptors coexist but do not appear to belinked (127), i.e., occupancy of the GM-CSF receptor fails totransmodulate the G-CSF receptor. Thus, even though GM-CSF stimulates the proliferation rate of WEHI3B(D+) it is not
a potent inducer of differentiation. These cells can still differentiate if they are treated directly with G-CSF or actinomycinD. A similar situation may occur in many primary leukemiasand it is imperative that we learn more about the biochemistryof receptor transmodulation in normal and neoplastic cells. Atpresent our knowledge of the receptors for the hemopoieticgrowth factors limits our ability to understand the biochemicalinteractions which mediate the receptor signal (72, 134-136).
Effects of Colony Stimulating Factors on Mature Cells
Studies with radiolabeled (iodinated) stimulating factors haveconfirmed that mature as well as immature cells in the granulocyte and macrophage cell series have receptors for colonystimulating factors (133). G-CSF receptors are lineage restricted to the neutrophil cell series and M-CSF receptors tothe monocyte cell series. The mature cells have more G-CSFand M-CSF receptors than the immature cells. GM-CSF andmulti-CSF receptors are present on neutrophils, monocytes,and eosinophils and the mature cells have less receptors thanthe early cells (133).
The presence of receptors for the HGFs on mature cells isconsistent with the observations that the colony stimulatingfactors have many effects on mature cells as well as on progenitor cells. The activities of GM-CSF on mature cells are listedin Table 1.
The effects of purified native and bacterially synthesizedhuman GM-CSF on mature neutrophils and on eosinophils aresimilar (75, 141). Superoxide production, phagocytosis, lyso-zyme secretion, and antibody dependent cell mediated cytotox-
Table I Effects of GM-CSF on neutrophils and macrophages
Function Ref.
ChemotaxisInhibition of locomotionAdherence phagocytosisSurvival in vitroSuperoxide generationAntibody dependent cell cytotoxicityParasite killing
9869
137,138139
140,14180,129,140,142,143
144,145
5629
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
HEMOPOIETIC GROWTH FACTORS
¡cityby "primed" neutrophils are all stimulated by natural and
bacterially synthesized GM-CSF. The concentrations of bacte-rially synthesized GM-CSF required to stimulate these functions were in the range of 1 to 10 pM. This is similar to thepotency of the COS cell derived human GM-CSFs (141).
These studies suggest that nonglycosylated GM-CSF is aseffective as glycosylated GM-CSF when stimulation of maturecell function is required. It is clear that low levels of neutrophils(<1000/mm3), but particularly levels below 500/mm,3 are as
sociated with an increased rate of infections (146). The mortality of febrile neutropenia with current treatment is low. Pizzoet al. (147) reported 22 deaths in 550 episodes of fever. Themorbidity of febrile neutropenic episodes can be significant andeven with current antibiotics the time to defervescence of thefever is prolonged (mean, 4 days; range, 1 to 35 days) (147). Itis not clear whether during infections the low numbers ofneutrophils are maximally activated or whether additional GM-CSF might cause further activation and additional antibacterialeffect. There are also reports that patients being treated withchemotherapy may exhibit diminished neutrophil function(148). This functional inhibition may be overcome by GM-CSF.This possibility is under investigation. Another question relatesto the functional status of neutrophils in the elderly and whetherGM-CSF might prove a useful activator.
There is an important distinction between GM-CSF and G-CSF; GM-CSF activates both mature neutrophils and eosino-phils (140, 149), whereas G-CSF activates only neutrophils. M-CSF activates mature monocytes and, in particular, causes theproduction of interferon (150) and plasminogen activator (151,152). These actions of M-CSF may have clinical applicationsin the treatment of infections.
In Vivo Effects of CSFs
Cranulocyte-Macrophage Colony Stimulating Factor. Untilrecently insufficient amounts of GM-CSF were available fordefinitive in vivo studies. The availability of all of the CSFs ingram quantities is now allowing the pharmacology and physiology of each to be studied in detail. Further, many laboratoriesare testing multiple combinations of the CSFs with other modulators such as the interferons and interleukins.
Murine Studies. Metcalf et al. (76) have studied the effects ofbacterially synthesized murine GM-CSF in vivo. The doses usedwere in the range 6-200 ng/animal. Since the serum half-lifeof GM-CSF is short (<5 min), multiple (up to 6) injectionswere used. These injections resulted in elevated peripheral bloodlevels of granulocytes and eosinophils. Administration i.p. produced a significant increase in the peritoneum of neutrophils,monocytes, and eosinophils and a decrease in lymphocytes.These experiments were controlled carefully to avoid effectsdue to contaminating endotoxin. A careful analysis of variousorgans revealed an increased number of granulocytes and monocytes in the liver (76). There was also an increased number ofgranulocytes in the lung (76). The basis for the accumulationof mature hemopoietic cells in the peritoneal cavity, liver, andlung is not apparent because these are not normally sites ofhemopoiesis.
Simian Studies. Donahue et al. (11) reported on the effectsof GM-CSF (1-2 x IO7 units/mg) infused continuously into 7
macaque monkeys. They found significant elevations of peripheral blood WBC counts. They also reported that GM-CSFstimulated hemopoiesis in a monkey with a virally inducedcytopenia. The effects of GM-CSF were not restricted to themyeloid system inasmuch as a reticulocytosis also developedafter the GM-CSF treatment. One animal apparently developed
antibodies which recognized the GM-CSF; these antibodies,however, did not interfere with hemopoiesis. A major problemwith this study was that the human GM-CSF was only partiallypurified from COS cell conditioned medium. Nevertheless assuming that the findings are reproducible with pure GM-CSFone can speculate that GM-CSF may be useful in conditionssuch as acquired immunodeficiency syndrome in which there isa neutropenia perhaps secondary to a deficiency of lymphokinesproduced by lymphocytes.
Multi-Colony Stimulating Factor. Kindler et al. (153) havereported the effects of multi-CSF in mice. Similar results havebeen obtained by Metcalf et al. (154) and Lord et al. (155).Kindler et al. (153) used doses of 0.03-0.1 Mg/24 h for 7 days.The total number of progenitor cells in normal mice wasincreased 2-fold and, in particular, there was a rise in theerythroid and myeloid progenitor cells in the spleen. If micewere irradiated prior to therapy there was a 10-fold increase inprogenitor cell levels compared to control animals. Multi-CSFinjected into the peritoneal cavity produced increased numbersof monocytes (154) and these monocytes appeared to functionas more effective phagocytes. The human equivalent of murineIL-3 has been cloned recently but its activity in vivo is not yetknown.
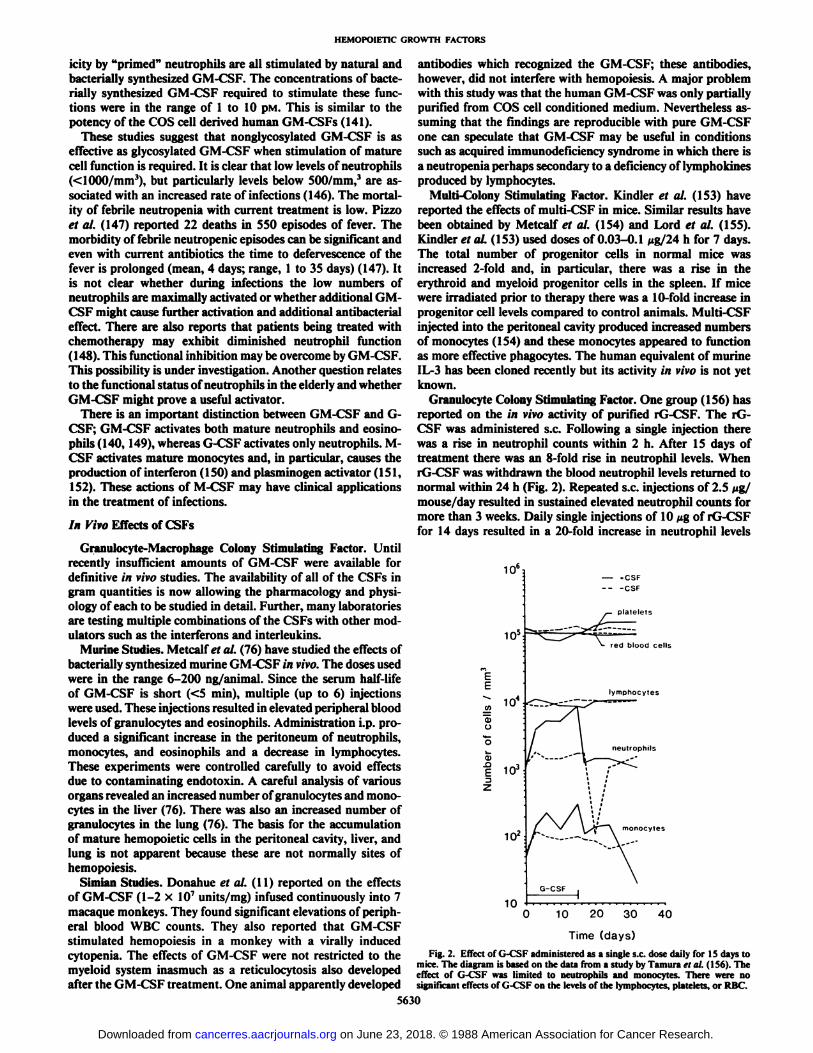
Granulocyte Colony Stimulating Factor. One group (156) hasreported on the in vivo activity of purified rG-CSF. The rG-CSF was administered s.c. Following a single injection therewas a rise in neutrophil counts within 2 h. After 15 days oftreatment there was an 8-fold rise in neutrophil levels. WhenrG-CSF was withdrawn the blood neutrophil levels returned tonormal within 24 h (Fig. 2). Repeated s.c. injections of 2.5 fig/mouse/day resulted in sustained elevated neutrophil counts formore than 3 weeks. Daily single injections of 10 Mgof rG-CSFfor 14 days resulted in a 20-fold increase in neutrophil levels
106,
E
<D0
10=
104
o£ 103
102
10
red blood cells
lymphocytes
neutrophils
10 20 30 40
Time (days)Fig. 2. Effect of G-CSF administered as a single s.c. dose daily for IS days to
mice. The diagram is based on the data from a study by Tamura <•/al. (1S6). Theeffect of G-CSF was limited to neutrophils and monocytes. There were nosignificant effects of G-CSF on the levels of the lymphocytes, platelets, or RBC.
5630
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
over the basal level. In the mouse the effects were restricted tothe myeloid system and there were no consistent changes in thelevels of lymphocytes, monocytes, platelets, or RBC. In themouse a transient effect on lymphocytes was apparent. G-CSFalso increased the number of CFU-spleen in the spleen. It islikely that this is an indirect effect, perhaps representing recruitment, since G-CSF has no in vitro effect on multipotentialprogenitor cells.
When purified G-CSF was given after an alkylating drug suchas cyclophosphamide, the neutropenia produced by this agentwas completely abrogated (156). Administration of G-CSF canpreempt the death of mice which have been infected with alethal dose of Gram negative microorganisms (Pseudomonas)(157). Pseudomonas is a common cause of infections and deathsin cancer patients being treated with cytotoxic chemotherapy.
Hemopoietic Growth Factors and Leukemia
The acute myeloid leukemias are subdivided into 7 categoriesby the French-American-British classification system (158).Each leukemia is characterized according to the degree ofproliferation of the leukemic clone in the marrow. The variantsof myeloid leukemia are further distinguished by the morphological and biochemical properties of the leukemic cells (158).
A feature of all cases of acute myeloid leukemia is that thereappears to be a block to the normal maturation of blast cells.This block may be associated with a disturbance in the biochemical pathways associated with receptor cross-modulation (126)or with the control of expression of the genes encoding differentiation factors (96). Therapy of acute myeloid leukemiasusually leads to the eradication of the leukemic clone and therecovery of normal hemopoiesis (159), although in some casesnormal differentiation and maturation of the leukemic cells areachieved (159).
Are the CSFs involved in the development or maintenanceof the myeloid leukemias? Would it be possible by manipulatingthe levels of one or more of the CSFs to improve the therapyof leukemia patients? It will be important to determine thebasis of the suppression of normal hemopoiesis by the leukemiccells and the levels of each of the CSFs in the tissues and serumof patients with AML. Further, an understanding of the dependence of the proliferation and differentiation of AML cellson the normal CSFs and other biological response modifiersshould help define the conditions to create for attacking theleukemic cells with cytotoxic drugs.
Both the autocrine and autonomous models for leukemic cellpopulation predict factor independent myeloid leukemias.However, in vitro studies suggest that at the time of diagnosismost myeloid leukemias require exogenous CSF to proliferate(160). There are a few leukemias which do not require CSF( 161). In some cases the leukemic cells may produce CSF (162).It has also been found that the dose-response curves for CSFstimulation of normal and leukemic cells are similar (38, 163).The number of G-CSF receptors on normal and leukemic cellsis similar at the equivalent stages of development. Some casesof AML, particularly during the later phases of the disease,may not be dependent on an exogenous source of CSF but thishas not been extensively tested. However, it should be notedthat many leukemias in blast crisis do not proliferate in vitro,even in the presence of the CSFs. Even those that do not growin vitro obviously proliferate in vivo, suggesting there are he-mopoietic growth factors still to be identified. It is not clearwhich growth factor is stimulating these leukemias in vivo. Itmay be that the leukemia models developed by Lang et al. (164)
represent the later stages of some cases of myeloid leukemia.However, their direct relevance to spontaneously arising leukemias is yet to be established.
Morstyn et al. ( 165) demonstrated that normal promyelocytesand myelocytes were highly responsive to partially purifiedhuman G-CSF. Recently, human G-CSF has been shown to bevery similar to murine G-CSF and I25l-Iabeled murine G-CSFcan be used to study human G-CSF receptors on leukemic cells(104); 14 patients with myeloid leukemia had near normal G-
CSF receptor levels. The data for human GM receptors (166)also demonstrate that leukemic myeloid cells have near normalreceptor levels. In many ways this evidence suggests that theseleukemias are not driven by autocrine stimulation.
Leukemogenesis. Murine marrow cells can be forced to adoptthe leukemic phenotype in several different ways. Continuouscell lines have been derived from apparently normal long-termmarrow culture. These cell lines are invariably dependent onmulti-CSF (115, 167, 168). However, cell lines dependent onG-CSF (169) and M-CSF (170) have also been obtained.
These cell lines which are absolutely dependent on the he-mopoietic growth factor for proliferation are blocked at an earlystage of differentiation but are not leukemogenic in syngeneicrecipients. They can be made independent of CSF by spontaneous mutation (168), by transformation with the Abelsonmurine leukemia virus (171, 172), or by infection with a recombinant retrovirus that encodes and expresses GM-CSF (164).The molecular lesions leading to leukemia in the case of insertion of the GM-CSF gene and the spontaneous mutations areassociated with the autocrine secretion of GM-CSF or multi-CSF. The \-abl gene appears to circumvent the requirementsfor GM-CSF and probably does this by producing a tyrosinekinase which mimics the intracellular events normally triggeredby the GM-CSF-GM-CSF receptor complex. Dunn3 has re
cently found that after the transfection of factor dependent cellswith a retrovirus which encodes and expresses the polyomamiddle T-antigen one class of cells becomes leukemic but remains dependent on GM-CSF for growth in vitro. These cellsappear to resemble human leukemic cells.
Acute Myelogenous Leukemia. It has been shown recentlythat the genes for M-CSF, GM-CSF, multi-CSF, interleukin 4,interleukin 5, and M-CSF receptor are all clustered on chromosome 5 (173, 174). There is a clinical syndrome called the5q- syndrome that involves a deletion of the long arm of
chromosome 5 (174). It is intriguing to speculate that thedisorder in these patients is due to activation of a hemopoieticgrowth factor or receptor gene. However, this has not yet beeninvestigated.
Promyelocytic leukemia has a 17/15 translocation and theG-CSF gene is on chromosome 17. The abnormality in thissituation may be the absence of a differentiation stimulus (G-CSF) leading to a maturation block at the level of the promye-locyte (175).
Production and Storage of GM-CSF
Most organs can produce the granulocyte-macrophage colonystimulating factors. The organs that have been shown to produce GM-CSF using bioassays (176) include salivary gland,lung, thymus, kidney, spleen, lymph node, pancreas, brain,heart, small intestine, testis, skeletal muscle, liver, and skin.Nicola et al. (177) have demonstrated that GM-CSF synthesized by different organs had very similar biochemical proper-
3A. Dunn, personal communication.
5631
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
ties. These studies indicate either that GM-CSF is made bycells present in all tissues such as monocytes, endothelial cells(177, 178), or fibroblasts or that cells of many different typesmake GM-CSF. This has not been clarified directly as yet.Endotoxin and tumor necrosis factor appear to stimulate CSFrelease (87, 179). The presence of GM-CSF in serum is still amatter of some contention. Undoubtedly serum contains he-mopoietic growth factors, but whether GM-CSF, multi-CSF,G-CSF, M-CSF, or IL-1 is responsible for the biological responses has not been studied with the appropriate reagents.
A recent study (180) has demonstrated GM-CSF productionby endothelial cells, smooth muscle, and fibroblasts and thatthese can be stimulated to produce CSF by tumor necrosisfactor. Broudy et al. (180) also showed that protein synthesiswas required. Koury and Pragnell (181) demonstrated CSFproduction by fibroblasts. The in vivo importance of theseobservations is unclear. The kinetics of release in vivo and thefailure of cycloheximide to reduce accumulation of CSFs inendotoxin serum suggest that at least one of the CSFs exists ina stored form (e.g., in membrane bound vesicles, in granules,or even on the surface of cells). The initial release of thesegrowth factors is probably connected with their ability to activate mature cells rather than to increase cell production.
Another approach to identifying the tissues and cell typesthat produce GM-CSF is to identify the cells and tissues whereGM-CSF mRNA is present. Recently, Chan et al. (182) usedfull length GM-CSF cDNA as a probe and screened, by dotblot hybridization analysis, 64 tissue samples including liver,colon, esophagus, rectum, stomach, breast, lung, skin, andvarious solid tumors and leukemias. There was no constitutiveproduction of detectable levels of GM-CSF mRNA in anysample studied. Since smooth muscle, endothelial cells, andfibroblasts would have been present in these tissues and othertechniques suggest they synthesize GM-CSF the approach mayhave been too insensitive. However, it is also possible that theGM-CSF mRNA only appears 8-12 hours after the tissues havebeen stimulated. Chan et al. (182) did confirm that mRNA forGM-CSF is present in activated T-lymphocytes and were ableto identify the region of the GM-CSF gene involved in regulating the transcription of the GM-CSF mRNA.
Clinical Disorders and Abnormal GM-CSF Levels
There is both direct and indirect evidence that CSFs cancontrol hemopoiesis in vivo. This evidence includes a correlationbetween urine CSF levels and WBC levels (183), a demonstration that tumors that produce CSF are associated with elevatedWBC and that the in vivo administration of purified CSFsproduces high WBC. Table 2 contains a list of clinical conditions that are associated with altered levels of CSF. In eachcase CSF was measured by the marrow bioassay. All of thesestudies need to be reassessed because the bioassay does notdistinguish among multi-CSF, GM-CSF, or mixtures of M-CSF and G-CSF. Also some studies (184, 188) on the distribution of human CSFs were undertaken with murine cells astargets. The latter assays presumably detected human G-CSFbut not human GM-CSF since only human G-CSF acts onmurine targets. It is also possible that M-CSF which actsindirectly to stimulate G-CSF release may have been detectedin some of these studies. In cases such as those associated withinfections where levels of endotoxin may have been high, indirect effects of endotoxin in the bioassay rather than GM-CSFlevels may also interfere with the assay. More direct assays(e.g., a radioimmunoassay) are required to characterize the
Table 2 Relationship between CSF levels and clinical states
ClinicalstateCyclic
neutropeniaAplastic
anemiaCongenitalneutropeniaEndotoxinadministrationInfectionInfectious
mononucleosisAcuteneutropeniaChronic
neutropeniawithoutinfectionChemotherapy
forlym-phomaTherapy
withanti-neutrophilantibodyHuman
cancersassociatedwithneutrophiliaSerum
CSFlevelCSF
levels cycle andprecederiseinWBCElevated
CSF inurineDiminishedserum CSFlevelElevated
serum andurine"GM-CSF-Elevated
serum and urineCSFLowlevels ofCSFElevated
G-CSFlevelsCSFlevelsnormalElevated
CSFlevelsElevated
CSFlevelsElevated
CSF levelsRef.183184185186,
187188.
189190191192193194195-198
Table 3 Potential clinical applications of G-CSF and GM-CSF
1. Mitigation of chemotherapy induced neutropenia.2. Improvement of host defense.3. Management of infections.4. Recovery following marrow transplantation.5. Expansion of progenitor cells prior to transplantation.6. Marrow failure.7. Improvement of granulocytc function.8. Anticancer effects.
distribution of each of the CSFs in various diseases. Studies ofCSF levels in normal and disease states are now under way andan enzyme-linked immunosorbent assay is available for monitoring exogenous bacterially synthesized GM-CSF administered to patients (199). This assay also detects native GM-CSFin serum.
Early Clinical Trials with G-CSF
The potential clinical applications of the CSFs are listed inTable 3. One possible application of the CSFs is to prevent orreverse the neutropenia that follows cytotoxic chemotherapyand thus reduce the incidence or duration of infections. Morstynet al. (200) have reported a Phase I/II study of G-CSF inpatients receiving i.v. melphalan. The patients received G-CSFfor 5 days prior to chemotherapy to assess the effect of G-CSF.The major findings were that G-CSF caused an immediatetransient fall in circulating neutrophils followed by a rise whichwas dependent on the dose of G-CSF administered. Circulatinglevels of neutrophils over 70,000/mm3 were achieved. The
neutrophils exhibited granulation and Dohle bodies. The majorchange in the marrow was an increase in the proportion ofpromyelocytes to over 30%. These changes would appear toreflect initial margination or sequestration of neutrophils followed by release and new synthesis of neutrophils after 4-5days. Following melphalan administration the period of neutropenia was reduced from 8 days to 0 days in patients who hadnot received prior cytotoxic therapy. Patients responded lesswell to G-CSF if they had been extensively pretreated withchemotherapy or radiotherapy. Bronchud et al. (201) havereported in a study of 12 patients a reduction in the neutropeniacaused by combination chemotherapy in small cell lung cancer,and Gabrilove et al. (202) have reported a reduction in neutropenia in patients receiving combination chemotherapy for bladder cancer. These studies involved the administration of G-CSFduring one cycle of chemotherapy. Studies are now required ofG-CSF administration for longer periods of time and randomized studies are required to confirm that G-CSF causes not onlya reduction in the period of neutropenia but also protectionagainst bacterial and fungal infections. The pharmacokinetics
5632
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
of G-CSF following i.v. administration reveals at least onephase of elimination of about 110 min. Morstyn et al. (200)also reported a brief first phase lasting for 3 to 8 minutes bybioassay. The best route of administration of G-CSF has notyet been defined; however, we have given 10-day continuouss.c. infusion with good effect and no local reactions. G-CSF iswell tolerated although bone pain has been reported (200). Amaximum tolerated dose has not been identified and studieshave stopped because it was considered that WBC levels above75,000/mm3 were undesirable.
Early Clinical Studies with GM-CSF
Three forms of GM-CSF are in clinical trials: mammalianderived material (Sandoz); material produced by yeast (Immu-nex); and bacterial!) synthesized material (Schering Plough).Like G-CSF, GM-CSF causes elevations in WBC levels. At lowdoses the predominant rise is in neutrophils; however, at higherdoses a rise in monocytes and eosinophils is also seen. Theimmediate effect of GM-CSF is a transient fall in the WBClevel (203) which occurs after both i.v. and s.c. administrationand which is followed by a rise. The best route of administrationof GM-CSF has not been identified. GM-CSF has been shownto reduce the period of neutropenia after autologous marrowtransplantation from 16 to 10 days (204). It has been shown tocause elevations in the WBC levels in patients with acquiredimmunodeficiency syndrome. Cebón et al. (199) have studiedthe pharmacokinetics of GM-CSF and found that after a singles.c. dose of GM-CSF of 10 Mg/kg the serum level of GM-CSFis sustained above 1 ng/ml for more than 12 h. There wouldtherefore appear to be no advantage to continuous i.v. infusionsof GM-CSF. This is particularly important because GM-CSFis a chemoattractant and might cause leukostasis and thrombosis at the catheter tip. The s.c. route of administration,however, has been associated with local rashes in some patients.Vadhan-Raj et al. (205) have reported a beneficial effect inpatients with myelodysplasia. However, Mertelsmann (206)reported at a recent meeting that in some cases where theproportion of blasts in the marrow is greater than 20% GM-CSF causes a worsening of the leukemia. An apparent differencebetween the early clinical studies using G-CSF (200-202) andGM-CSF (199, 203-205) is that no maximum tolerated doseof G-CSF has been identified, whereas there is clearly a maximum tolerated dose of GM-CSF. The dose limiting toxicity ofGM-CSF may reflect direct effects or indirect effects perhapsas a consequence of monocyte activation and release of othercytokines. The toxicities of GM-CSF include bone pain, fever,edema, rashes, and serositis, including pericarditis.
Future Directions
Anticancer Effects of GM-CSF. GM-CSF can function as amacrophage activating factor and consequently can promotethe ability of macrophages to recognize and destroy humanmelanoma cells. GM-CSF also activates antibody dependentcell cytotoxicity (140). Administered systemically, the CSFs areunlikely to have anticancer effects. However, there are clinicalsituations, e.g., ovarian cancer, in which the malignancy isconfined to the peritoneal cavity for a significant period of time.In this situation the i.p. infusion of GM-CSF and tumor directed antibodies may be able to stimulate an anticancer effectvia the activation of antibody mediated monocyte cytotoxicity.
Effects of Colony Stimulating Factors on NonhemopoieticCells. Studies on G-CSF receptors (133) and GM-CSF receptors (134) indicate that both are restricted to cells of the
5633
hemopoietic series. However, Ruff et al. (207) have reportedthat at high concentrations GM-CSF inhibits the proliferationof small cell lung cancer cells. It suggests either that malignantcells may express GM-CSF receptor aberrantly or that thesetumors are closely related to hemopoietic cells. Baldwin et al.(218) have reported G- and GM-CSF receptors on small celllung cancers. We do not know what effect the CSFs will haveon such tumors or whether we will lose the selective protectionof the marrow.
Potential Side Effects of CSF Therapy. The direct effects ofthe colony stimulating factors will be confined to cells thatcontain CSF receptors; however, the stimulated cells may beinduced to secrete other biological response modifiers whichcould influence the action of these molecules. These couldinclude IL-1 and tumor necrosis factor. Accumulation of gran-ulocytes and/or monocytes in the lungs, liver, pericardium, andperitoneum is possible. Such infiltrates have been observed inpreclinical studies (76).
Another potential side effect is the development of antibodiesto CSF. Such antibodies are theoretically more likely to developin response to glycosylated than to nonglycosylated CSF; however, even nonglycosylated CSF may prove antigenic due tominor differences in the three dimensional structure or its stateof aggregation.
Antibodies to hGM-CSF were detected in some of the monkeys treated with the COS cell derived human GM-CSF (11).However, in this situation there was a species difference betweenthe GM-CSF and the animals and the carbohydrate moietywould be quite different. The antibodies were not neutralizingand appeared to be of little pathological significance.
Role of Combinations of Hemopoietic Growth Factors. Manycombinations of biological response modifiers will be used inan attempt to modify hemopoiesis and phagocyte function andit is a little too early to predict the best combinations ofhemopoietic growth factors, modulators, and cytotoxic drugs.A considerable effort will be required to define the in vivo effectsof the hemopoietic growth factors so that appropriate baselinedata are available from which their use can be optimized. Theactions of GM-CSF and G-CSF are synergistic (209, 210).However, the testing of the combinations of these growthfactors should be delayed until we have adequate baseline dataon the individual molecules.
The hemopoietic growth factors have been characterized indetail and we have a considerable knowledge of their biology invitro. The exquisite specificity of these growth factors makesthem potentially powerful modulators of blood cell productionand function. At present we can only hope that this potentialcan be combined with new or current therapies to help transplant or cancer patients. The results of the initial animal studieshave been so encouraging that it is tempting to suggest that thehemopoietic growth factors will lead to entirely new approachesin oncology. Now that neutrophil levels can be restored morequickly, it is essential to develop similar strategies for theelevation of platelets. However, it is also clear that the controlof hemopoiesis is complex and depends on the interaction ofmany factors. We do not fully understand this network andmuch work remains before we can ensure that the hemopoieticgrowth factors can be used for the maximum benefit of thepatient.
Acknowledgments
The authors wish to thank Jean Kingett for her care and patience inthe preparation of this manuscript. We are also grateful to Henrike"»
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
Heckmann for translating Metchnikoffs paper (1) from German intoEnglish.
References
1. MetchnikofT. E. About a yeast infection of Daphnia. A contribution to thestudy about the fighting of phagocytes against organisms that cause disease.Arch. Pathol. Anal. Physiol. Clin. Med., 96: 177-195, 1884.
2. Wright, A. E.. and Douglas, S. R. An experimental investigation of the roleof the blood fluids in connection with phagocytosis. Proc. R. Soc., 72: 357-370, 1903.
3. Burgess, A. W., and Nicola, N. A. Growth Factors and Stem Cells. NewYork: Academic Press, 1983.
4. Burgess, A. W. Growth factors and cancer. Aust. NZ J. Surg., 55:105-110,1985.
5. Burgess, A. W. Growth factors and oncogenes. Immunol. Today, 6: 107-112, 1985.
6. Metcalf, D. The granulocyte macrophage colony stimulating factors. Science(Wash. DC), 299: 16-22, 1985.
7. Metcalf, D. The granulocyte-macrophage colony stimulating factor. Cell,43:5-6, 1985.
8. Metcalf, D. The molecular biology and functions of the granulocyte-macrophage colony stimulating factors. Blood, 67: 257-267, 1986.
9. Arai, K., Yokota, T., Miyajima, A., Arai, N., and Lee, F. Molecular biologyof T-cell derived lymphokines: a model system for proliferation and differentiation of hemopoietic cells. Bioessays, 5: 166-171, 1986.
10. Gough. N. M., and Burgess, A. W. The genes for granulocyte-macrophagecolony stimulating factor and multi-colony stimulating factor IL-3. In: G.Guroff (ed.), Oncogenes, Genes and Growth Factors. New York: John Wiley&Sons, Inc., 1987.
11. Donahue. R. E.. Wang, E. A., and Stone, D. K., el al. Stimulation ofhaematopoiesis in primates by continuous infusion of recombinant humanGM-CSF. Nature (Lond.), 321: 872-875, 1986.
12. Nicola, N. A., and Vadas, M. Hemopoietic colony-stimulating factors.Immunol. Today, 5: 76-80, 1984.
13. Nicola, N. A. Granulocyte colony stimulating factor and differentiation-induction in myeloid leukemic cells. Int. J. Cell Cloning, 5: 1-15, 1987.
14. Schlick, E., Kuliin.um. R., Härtung,K., and Chirigos, M. A. Modulationof myelopoiesis by CSF or CSF-inducing biological response modifiers. J.Immunopharmacol., 7: 141-166, 1985.
15. Shaw, G. B. The Doctor's Dilemma: A Tragedy. Act I. New York: Signet,
1985.16. Metcalf, D. The Hemopoietic Colony Stimulating Factors. Amsterdam:
Elsevier/North-Holland Biomedicai Press, 1984.17. Morstyn, G., Nicola, N., and Metcalf, D. Purification of hemopoietic
progenitor cells from human marrow using a fucose binding lectin and cellsorting. Blood, 56: 798-805, 1980.
18. Williams, D. E., Straneva, J. E., Shen, R.-N., and Broxmeyer, H. E.Purification of murine bone-marrow-derived granulocyte-macrophage colony-forming cells. Exp. Hematol., 3: 243-250, 1987.
19. Nicola, N. A., Burgess, A. W., Staber, F. G., Johnson, G. R., Metcalf, D.,and Battye. F. L. Differential expression of lectin receptors during hemopoietic differentiation: enrichment for granulocyte-macrophage progenitorcells. J. Cell. Physio!., 103: 217-237, 1980.
20. Nicola, N. A., Metcalf, D., von Melchner, II.. and Burgess, A. W. Isolationof murine fetal hemopoietic progenitor cells and selective fractionation ofvarious erythroid precursors. Blood, 58: 376-386, 1981.
21. Harris, R. A., Hogarth, P. M., McKenzie, I. F. C, and Penington, D. G.Differential expression of (,)a nr alloantigen on murine hemopoietic progenitor cells. Selective enrichment for megakaryocyte progenitors. Exp.Hematol., //: 527-541, 1983.
22. Kannourakis. G., and Johnson, G. R. Fractionation of subsets of BFU-Efrom normal human bone marrow. Responsiveness to erythropoietin, humanplacenta! conditioned medium or granulocyte-macrophage colony stimulating factor. Blood, 71: 758-765, 1988.
23. Koike, K., Stanley, E. R., Ihle, J. N., and Ogawa, M. Macrophage colonyformation supported by purified CSF-1 and/or interleukin-3 in serum-freeculture: evidence for hierarchical difference in macrophage colony formingcells. Blood, 67: 859-864, 1986.
24. Metcalf, D., and Burgess, A. W. Clonal analysis of progenitor cell commitment to granulocyte or macrophage production. J. Cell. Physiol., Ill: 275-283, 1982.
25. Stanley, E. R.. Bartocci, A., Patinkin, D., Rosendaal, M., and Bradley, T.R. Regulation of very primitive, multipotent, hemopoietic cells by hemo-poietin-1. Cell, 45: 667-674. 1986.
26. Bradley, T. R., and Hodgson. G. Detection of primitive macrophage progenitor cells in mouse bone marrow. Blood, 54: 1446-1450, 1979.
27. McNiece, I. K., Bradley, T. R., Kriegler, A. B., and Hodgson, G. S. Agrowth factor produced by WEHI-3 cells for murine high proliferativepotential GM-progenitor colony forming cells. Cell Biol. Int. Rep., 6: 243-251, 1982.
28. Bradley, T. R.. Hodgson, G. S., and Bertoncello, I. Characteristics ofprimitive macrophage progenitor cells with high proliferative potential.Their relationship to cells with marrow repopulating ability in 5-fluorouraciltreated mouse bone marrow. In: S. J. Baum and I . van Bekkum (eds.).Experimental Hematology Today, pp. 284-297. Basel: S. Karger, A. G.,1980.
29. Bertoncello, I., Bradley, T. R., and Hodgson, G. S. Characterization andenrichment of progenitor cells from normal and 5-fluorouracil treated mousebone marrow by unit gravity sedimentation. Exp. Hematol., 9: 604-610,1981.
30. Nakahata, T., Tsuji. K., Ishiguro, A., Ando, O., Norose, N., Koike, K., andAkabane, T. Single-cell origin of human mixed hemopoietic colonies expressing various combinations of cell lineages. Blood, 65:1010-1016, 1985.
31. Jubinsky. P. T., and Stanley, E. R. Purification of hemopoietin 1: a multi-lineage hemopoietic growth factor. Proc. Nati. Acad. Sci. USA, S3: 2764-2768, 1985.
32. Metcalf, D., Johnson. G. R., and Burgess, A. W. Direct stimulation bypurified GM-CSF of the proliferation of multipotential and erythroid precursor cells. Blood, 55: 138-147, 1980.
33. Emerson, S. G., Sieff, C. A., Wang, E. A., Wong, G. G., Clark, S. C., andNathan, D. G. Purification of fetal hematopoietic progenitors and demonstration of recombinant multipotential colony-stimulating activity. J. Clin.Invest., 76: 1286-1290, 1985.
34. Smith, C. A., and Rennick, D. M. Characterization of a murine lymphokinedistinct from interleukin 2 and interleukin 3 (IL-3) possessing a T-cellgrowth factor activity and a mast-cell growth factor activity that synergizeswith IL-3. Proc. Nati. Acad. Sci. USA, 83: 1857-1861, 1986.
35. Jacobsen, N., Broxmeyer, H. E., Grossbard, E., and Moore, M. A. S.Colony forming units in diffusion chambers (CFUd) and colony formingunits in agar culture (CFUc) obtained from normal human bone marrow apossible parent progeny relationship. Cell. Tissue Kinet., 12:113-226,1979.
36. Morstyn, G., Nicola, N. A., and Metcalf, D. Separate actions of differentcolony stimulating factors from human placenta! conditioned medium onhuman hemopoietic progenitor cell survival and proliferation. J. Cell. Physiol., 109: 133-142, 1981.
37. Begley, C. G., Metcalf, D., Lopez, A. F., and Nicola, N. A. Fractionatedpopulations of normal human marrow cells respond to both human colony-stimulating factors with granulocyte-macrophage activity. Exp. Hematol.,13: 956-962, 1985.
38. Francis, G. E., Turna, G. A., Berney, J. J., and Hoffbrand, A. V. Sensitivityof acute myeloid leukaemia cells to colony stimulating activity. Relationshipto response to chemotherapy. Br. J. Hematol., 49: 259-267, 1981.
39. Bradley, T. R., and Metcalf, D. The growth of mouse bone marrow cells invitro. Aust. J. Exp. Biol. Med. Sci., 44: 287-300, 1966.
40. Pluznick, D. H., and Sachs, L. The cloning of normal mast cells in tissueculture. J. Cell. Comp. Physiol., 66: 319-324, 1986.
41. Stanley, E. R., Hansen, G., Woodcock, J., and Metcalf, D. Colony stimulating factor and the regulation of granulopoiesis and macrophage production. Fed. Proc., 34: 2272-2278, 1975.
42. Burgess, A. W., and Metcalf, D. The effect of colony stimulating factor onthe synthesis of ribonucleic acid by mouse bone marrow cells in vitro. J.Cell. Physiol., 90: 471-484, 1977.
43. Stanley, E. R., and Guilbert, L. J. Methods for the purification, assay,characterization and target cell binding of a colony stimulating factor «SI1). J. Immunol. Methods, 42: 253-284, 1981.
44. Axelrad, A., Burgess, A., Iscove, N., and Wagemaker, G. Culture of Hae-mopoietic Cells: Workshop Report, pp. 421-425. New York: Alan R. Liss,Inc., 1981.
45. Nicola, N. A., Metcalf, D., Johnson, G. R., and Burgess, A. W. Separationof functionally distinct human granulocyte-macrophage colony-stimulatingfactors. Blood, 54:614-627, 1979.
46. Burgess, A. W., and Metcalf, D. Characterization of a serum factor stimulating the differentiation of myelomonocytic leukemic cells. Int. J. Cancer,20:647-654, 1980.
47. Nicola, N. A., Metcalf, D., Matsumoto, M., and Johnson, G. R. Purificationof a factor inducing differentiation in murine myelomonocytic leukemiacells. J. Biol. Chem., 25«:9017-9023, 1983.
48. Burgess, A. W., Metcalf, D., Russell, S. H. M., and Nicola, N. A. Granulocyte-macrophage, megakaryocyte, eosinophil and erythroid colony-stimulating factors produced by mouse spleen cells. Biochem. J., 185: 301-314,1980.
49. Williams, N., Eger, R. R., Moore, M. A. S., and Mendelsohn, N. Differentiation of mouse bone marrow cells into neutrophilic granulocytes by anactivity separable from WEHI-3 cell conditioned medium. Differentiation,//: 59-63, 1978.
50. Bazill, G. W., Haynes, M., Garland, J., and Dexter, T. M. Characterizationand partial purification of a haemopoietic cell growth factor in WEHI-3 cellconditioned medium. Biochem. J., 210: 747-759, 1983.
51. Ihle, J. N., Keller, J., Henderson, L., Klein, F., and Palaszynski, E. Procedures for the purification of interleukin-3 to homogeneity. J. Immunol.,129: 2431-2436, 1982.
52. Clark-Lewis, I., Kent, S. B. H., and Schrader, J. W. Purification to apparenthomogeneity of a factor stimulating the growth of multiple lineages ofhemopoietic cells. J. Biol. Chem., 25:7488-7494, 1984.
53. Fung, M. C., Hapel, A. J., Ymer, S., Cohen, D. R., Johnson, R. A.,Campbell, H. D., and Young, I. G. Molecular cloning of cDNA for mouseinterleukin-3. Nature (Lond.), 307: 233-237, 1984.
54. Yang, Y. C., Ciarletta, A. B., Temple, P. A., et al. Human IL-3 (multi-CSF)identification by expression cloning of a novel hematopoietic growth factorrelated to murine IL3. Cell, 47: 3-10, 1986.
55. Wong, G. G., Witek, J. S., Temple, P. A., et al. Human GM-CSF: molecularcloning of the complementary DNA and purification of the natural andrecombinant proteins. Science (Wash. DC), 22«:810-815, 1985.
56. Nomura, H., Imazeki, I., Oheda, M., et al. Purification and characterization
5634
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
of human granulocyte colony-stimulating factor (G-CSF). EMBO J., 5:871-876, 1986.
57. Tiku, K., Tiku, M. L., and Skosey, J. L. Interleukin 1 production by humanpolymorphonuclear neutrophils. J. Inumino].. 136: 3677-3685, 1986.
58. Lopez, A. F., Begley, C. G., Williamson, D. J., Warren, D. J., Vadas, M.A., and Sanderson, C. J. Murine eosinophil differentiation factor. J. Exp.Med, 163: 1085-1099, 1986.
59. Enokihara, II.. Hamaguchi, 11..Sakamaki, II., et al. Specific production ofeosinophil colony stimulating factor from sensitized T cells from a patientwith allergic eosinophilia. Br. J. Haematol., 59:85-91, 1985.
60. Warren, D. J., and Sanderson, C. J. Production of a T cell hybrid producinga lymphokine stimulating eosinophil differentiation. Immunology, 54:615-623, 1985.
61. Miyaké,T., Kung, CK.-H., and Goldwasser, E. Purification of humanerythropoietin. J. Biol. Chem., 252: 5558-5564, 1977.
62. Smith, K. A. Regulation of normal and neoplastic T-cell growth. In: J.Feramusco, B. Ozanne, and C. Stiles (eds.), Cancer Cells 3. pp. 205-210.Cold Spring Harbor, NY: Cold Spring Harbor Laboratory, 1985.
63. Robb, R. J. Conversion of low affinity interleukin 2 receptors to a high-affinity state following fusion of cell membranes. 1'me. Nati. Acad. Sci.USA, 83: 3992-3996, 1986.
64. Lee, F., Yokota, T., Otsuka, T., et al. Isolation and characterization of amouse interleukin cDNA clone that expresses BSF-1 activities and T-celland mast cell stimulating activities. Proc. Nati. Acad. Sci. USA, 83: 2061-2065, 1986.
65. Johnson, G. R., and Metcalf, D. Detection of a new type of mouse eosinophilcolony by Luxol-Fast-Blue staining. Exp. Hematol., 8: 549-561, 1980.
66. Metcalf, D., Burgess, A. W., Johnson, G. R., et al. In vitro actions onhemopoietic cells of recombinatn murine GM-CSF purified after productionin E. coli; comparison with purified native GM-CSF. J. Cell. Physiol., 128:421-431, 1986.
67. Burgess, A. W., Metcalf, D., Kozka, I. J., et al. Purification of two formsof colony-stimulating factor from mouse L-cell conditioned medium. J. Biol.Chem., 260: 16004-16011, 1985.
68. Mizel, S. B. Interleukin 1 and T cell activation. Immunol. Rev., 63: 51-72,1982.
69. Gasson, J. C., Weisbart, R. H., Kaufman, S. E., et al. Purified humangranulocyte-macrophage colony-stimulating factor: direct action on neutrophils. Science (Wash. DC), 226:1339-1342, 1984.
70. Burgess. A. W., and Nice, E. C. Murine granulocyte-macrophage colony-stimulating factor. Methods Enzymol., 116: 588-600, 1985.
71. Gough, N. M., Gough, J., Metcalf, D., et al. Molecular cloning of cDNAencoding a murine haematopoietic growth regulator, granulocyte-macrophage colony stimulating factor. Nature (Lond.), 309: 763-767, 1984.
72. Park, L. S., Friend, D., Gillis, S., and Urdal, D. L. Characterization of thecell surface receptor for granulocyte-macrophage colony-stimulating factor.J. Biol. Chem., 267: 4177-4183, 1986.
73. Miyajima, A., Otsu, K., Schreurs, J., Bond, M. W., Abrams, J. S. A., andArai, K. Expression of murine and human granulocyte-macrophage colony-stimulating factors in 5. cerevisiae: mutagenesis of the potential glycosyla-tion sites. EMBO J., 5:1193-1197, 1986.
74. DeLamarter, J. F., Mermod, J. J., Liang, C. M., Eliason, J. F., andThatcher, D. R. Recombinant murine GM-CSF from E. coli has biologicalactivity and is neutralized by a specific anti-serum. EMBO J., 4: 2575-2581, 1985.
75. Burgess. A. W., Begley, C. G., Johnson, G. R., et al. Purification andproperties of bacterially synthesized human granulocyte-macrophage colonystimulating factor. Blood. 69:43-51, 1987.
76. Metcalf. D., Begley. C. G., Williamson, D. J., et al. Hemopoietic responsesin mice injected with purified recombinant murine GM-CSF. Exp. Hematol.,15: 1-9, 1987.
77. Stanley, E., Metcalf, D., Sobieszczuk, P., Gough, N. M., and Dunn, A. R.The structure and expression of the murine gene encoding granulocyte-macrophage colony stimulating factor: evidence for utilization of alternativepromoters. EMBO J., 4: 2569-2573, 1985.
78. Sparrow, L. G.. Metcalf, D., Hunkapiller, M. W.. Hood, L. E., and Burgess,A. W. Purification and partial amino acid sequence of asialo murine granulocyte-macrophage colony stimulating factor. Proc. Nati. Acad. Sci. USA,82: 292-296, 1985.
79. Shaw, G., and Kamen, R. A conserved AU sequence from the 3' untranslatedregion of GM-CSF mRNA mediates selective mRNA degradation. Cell, 46:659-667, 1986.
80. Metcalf, D., Begley, C. G.. Johnson, G. R., et al. Biologic properties invitro of a recombinant human granulocyte-macrophage colony stimulatingfactor. Blood. 67: 37-45, 1986.
81. Nicola. N. A., and Metcalf, D. The colony stimulating factors and myeloidleukemia. Cancer Surv., 45: 789-815, 1985.
82. Metcalf. D. Clonal culture of hemopoietic cells: techniques and applications.Amsterdam: Elsevier/North-Holland Biomedicai Press. 1984.
83. Nicola, N. A., Metcalf, D., Johnson, G. R., and Burgess, A. W. Preparationof colony stimulating factors from human placenta! conditioned medium.Leuk. Res., 2:313-322, 1978.
84. Kaushanksy. K., O'Hara, P. J., Berkner, K., Segal, G. M., Hagen, F. S.,
and Adamson, J. W. Genomic cloning, characterization, multilineagegrowth-promoting activity of human granulocyte-macrophage colony-stimulating factor. Proc. Nati. Acad. Sci. USA, 83: 3101 -3105. 1986.
85. Sieff, C. A.. Emerson, S. G., Donahue. R. E.. Nathan, D. G., Wang, E. A.,Wong, G. G., and Clark, S. C. Human recombinant granulocyte-macrophage
colony-stimulating factor: a multilineage hematopoietin. Science (Wash.DC), 230: 1171-1173, 1985.
86. Motoyoshi, K., Suda, T., Kusumoto, K., Takaku, F., and Miura. Y. Granulocyte-macrophage colony-stimulating and binding activities of purifiedhuman urinary colony-stimulating factor to murine and human bone marrowcells. Blood, 60: 1378-1386, 1982.
87. Munker, R., Gasson, J.. Ogawa, M., and Koeffier. H. P. Recombinanthuman TNF induces production of granulocyte-monocyte colony-stimulating factor. Nature (Lond.), 323:79-82, 1986.
88. Piacibello, W., Lu, L., Wächter, M., Rubin, B., and Broxmeyer, H. E.Release of granulocyte-macrophage colony stimulating factors from majorhistocompatibility complex class 2 antigen-positive monocytes is enhancedby human -y-interferon. Blood, 66: 1343-1351, 1985.
89. Stanley, E. R., and Heard, P. M. Factors regulating macrophage productionand growth. Purification and some properties of the colony stimulatingfactor from medium conditioned by mouse L cells. J. Biol. Chem., 252:4305-4312,1977.
90. Metcalf, D., Russell, S., and Burgess, A. W. Production of hemopoieticstimulating factors by pokeweed-mitogen-stimulated spleen cells. Transplant. Proc., 70:91-94, 1978.
91. Tsuneoka, K., and Shikita, M. A granulocyte colony-stimulating factor fromserum-free cultures of RSP-2.P3 cells: its separation from a macrophagecolony-stimulating factor and its biological and molecular characterization.Cell Struct. Funct., 9:67-81, 1984.
92. Tsuneoka, K., and Shikita, M. A colony-stimulating factor for neutrophilgranulocytes; a marked increase of its production by the addition of sodiumbutyrate and lipopolysaccharide in serum-free culture of RSP-2.P3 cells. J.Cell. Physiol., 725:436-442, 1985.
93. Yunis, A. A., Wu, M.-C, and Miller, A. M. Colony-stimulating factor. In:G. Guroff (ed.). Growth and Maturation Factors, pp. 209-248. New York:John Wiley & Sons, Inc., 1983.
94. Lotem, J., Lipton, J. H., and Sachs, L. Separation of different molecularforms of macrophage- and granulocyte-inducing proteins for normal andleukemic myeloid cells. Int. J. Cancer, 25: 763-771, 1980.
95. Shadduck, R. K., and Metcalf, D. Preparation and neutralization characteristics of an anti-CSF antibody. J. Cell. Physiol., 86: 247-252, 1975.
96. Sachs, L. Growth, differentiation and the reversal of malignancy. Sei. Am.,254:40-47, 1986.
97. Tomida, M., Yamamoto-Yamiguchi, Y., and Hozumi, M. Purification of afactor inducing differentiation of mouse myeloid leukemic Ml cells fromconditioned medium of mouse fibroblast L929 cells. J. Biol. Chem., 259:10978-10982, 1984.
98. Gabrilove, J. L., Weite, K., Harris, P., et al. Pluripoietin a: a second humanhematopoietic colony-stimulating factor produced by the human bladdercarcinoma cell line 5637. Proc. Nati. Acad. Sci. USA, «J:2478-2482, 1986.
99. Metcalf, D., Burgess, A. W., Johnson, G., et al. In vitro actions on hemopoietic cells of recombinant murine GM-CSF purified after production inE. coli; comparison with purified native GM-CSF. J. Cell. Physiol., 725:421-431,1986.
100. Donahue, R. E., Emerson, S. G., Wang, E. A., Wong, G. G., Clark, S. C.,and Nathan, D. G. Demonstration of burst promoting activity of recombinant human GM-CSF on circulating erythroid progenitors using an assayinvolving the delayed addition of erythropoietin. Blood, 66: 1479-1481,1985.
101. Nagata, S., Tsuchiya, M., Asano, S., et al. Molecular cloning and expressionof cDNA for human granulocyte colony stimulating factor. Nature (Lond.),579:415-417,1986.
102. Souza, L. M., Boone, T. C., Gabrilove, J., et al. Recombinant humangranulocyte-colony-stimulating factor: effects on normal and leukemic myeloid cells. Science (Wash. DC), 232:61-65, 1986.
103. Tsuchiya, M., Asano, S., Kaziro, Y., and Nagata, S. Isolation and characterization of the cDNA for murine granulocyte colony-stimulating factor.Proc. Nati. Acad. Sci. USA, 83: 7633-7637, 1986.
104. Nicola, N. A., Begley, C. G., and Metcalf, D. Identification of the humananalogue of a regulator that induces differentiation in murine leukemic cells.Nature (Lond.), 314:625-628, 1985.
105. Das, S. K., and Stanley, E. R. Structure-function studies of a colonystimulating factor (CSF-1). J. Biol. Chem., 257:13679-13684, 1982.
106. Kawasaki, E. S., Ladner, M. B., Wang, A. M., et al. Molecular cloning of acomplementary DNA encoding human macrophage-specific factor (CSF-1).Science (Wash. DC), 230: 291-296, 1985.
107. DeLamarter, J. F., Hession, C., Semon, D., Gough, N., Rothenbuhler, R.,and Mermod, J.-J. Nucleotide sequence of a cDNA encoding murine CSF-1 (Macrophage-CSF). NucÃ.Acids Res., 75: 2389-2390, 1987.
108. Ishizaka, Y., Motoyoshi, K., Hatake, K., Saito, M., Takaku, F., and Miura,Y. Mode of action of human urinary colony-stimulating factor. Exp. Hematol., 14: 1-8, 1986.
109. Tushinski, R. J., Oliver, I. T., Guilbeit, L. J., et al. Survival of mononuclearphagocytes depends on a lineage-specific growth factor that the differentiated cells selectively destroy. Cell, 2Ä:71-81, 1982.
110. Johnson, G. R., and Burgess, A. W. Molecular and biological properties ofa macrophage colony-stimulating factor from mouse yolk sacs. J. Cell. Biol..77:35-47, 1978.
111. Stephenson, J. R., Axelrad, A. A., McLeod, D. L., and Shreeve, M. M.Induction of colonies of hemoglobin-synthesizing cells by erythropoietin invitro. Proc. Nati. Acad. Sci. USA, 6«:1542-1546, 1971.
112. Parker, J. W., and Metcalf, D. Production of colony-stimulating factor inmitogen-stimulated lymphocyte culture. J. Immunol., 7/2: 502-510, 1974.
5635
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
113. Yokota, T.. Lee, F., Rennick, D., et al. Isolation and characterization of amouse cDNA clone that expresses mast-cell grow th-factor activity in monkey cells. Proc. Nail. Acad. Sci. USA, 81: 1070-1074. 1984. 140.
114. Yung. Y.-P., Eger, R.. Tertian. G., and Moore, M. A. S. Long term in vitroculture of murine mast cells. II. Purification of a mast cell growth factorand its dissociation from TCGF. J. Immunol.. 127: 794-799, 1981. 141.
115. Greenberger, J. S., Sakakeeny, M. A., Humphries, R. K.. Eaves, C. J., andEckner. R. J. Demonstration of permanent factor-dependent multipotential(erythroid/neutrophil/basophil) hematopoietic progenitor cell lines. Proc. 142.Nati. Acad. Sci. USA, 80: 2931-2935, 1983.
116. Greenberger. J. S., Hapel, A.. Nabel, G., et al. Effects of murine leukemiaviruses on establishment, growth and differentiation of permanent factor 143.dependent committed and pluripotential hematopoietic stem cell lines invitro. In: S. J. Bau. G. D. Ledney, and S. Thierfelder (eds.). ExperimentalHematology Today, pp. 195-209. Basel: S. Karger AG, 1982. 144.
117. Schrader, J. W., Ziltener, H. J., and Leslie. K. B. Structural homologiesamong the hemopoietins. Proc. Nati. Acad. Sci. USA, 83:2458-2462,1986.
118. Cohen, D. R., Hapel, A. J., and Young, I. G. Cloning and expression of the 145.rat interleukin-3 gene. NucÃ.Acids Res., 14: 3641-3658, 1986.
119. Eliason, H. F. The effects of pure granulocyte-macrophage colony stimulating factors in serum free culture. In: Eleventh EMBO Annual Symposium: 146.Growth Factors, Receptors and Oncogenes, p. 85. Heidelberg: EMBL, 1985.
120. Ralph. P.. Warren, M. K., Lee, M. T., et al. Inducible production of humangrowth factor. CSF-1. Blood, 68: 633-639, 1986. 147.
121. Goldwasser, E., Ihle. J. W., Prystowsky, M. B., Rich, I., and van Zant, G.The effect of interleukin-3 on hemopoietic precursor cells, in: D. W. Goldeand P. A. Marks (eds.). Normal and Neoplastic Hemopoiesis, UCLA 148.Symposia on Molecular and Cellular Biology, pp. 301-309. New York:Alan R. Liss, Inc., 1983. 149.
122. Guilbert, L. J., Nelson, D. J., Hamilton, J. A., and Williams, N. The natureof 12-O-tetradecanoylphorbol-13-acetate (TPA)-stimulated hemopoiesis,colony stimulating factor (CSF) requirement for colony formation, and the 150.effect of TPA on |12"I-CSF-1 binding to macrophages. J. Cell. Physiol., IIS:276-282. 1983.
123. Warren, M. K., and Ralph, P. Macrophage growth factor CSF-1 stimulates 151.human monocyte production of interferon. tumor necrosis factor and mye-loid CSF. J. Immunol., 137: 2281-2285, 1986.
124. Zucali, J. R., Dinarello, C. A., Oblon, D. J., Gross, M. A., Anderson, L., 152.and Weiner, R. S. Interleukin l stimulates fibroblasts to produce granulocyte-macrophage colony-stimulating activity and prostaglandin E¡.J. Clin.Invest., 77: 1857-1863, 1986. 153.
125. Bowen-Pope, D. F.. Dicorleto, P. E., and Ross, R. Interactions between thereceptors for platelet-derived growth factor and epidermal growth factor. J.Cell. Biol., 96:679-683, 1983.
126. Massague, J. Transforming growth factor-^ modulates the high affinity 154.receptors for epidermal growth factor and transforming growth factor-a. J.Cell. Biol., 100: 1508-1514, 1985.
127. Walker, F., Nicola, N. A.. Metcalf, D., and Burgess, A. W. Hierarchical 155.down-modulation of hemopoietic growth factor receptors. Cell, 43: 269-276, 1985.
128. Ruscelli, F. W., Collins, S. J., Woods, A. M.. and Gallo, B. C. Clonal 156.analysis of the response of human myeloid leukemic cell lines to colony-stimulating activity. Blood, 58: 285-292, 1981.
129. Dessein, A. J., Vadas, M. A., Nicola, N. A., Metcalf, D., and David, J. R.Enhancement of human blood eosinophil cytotoxicity by semipurified eosin- 157.ophil colony-stimulating factors. J. Exp. Med., 156: 90-103, 1982.
130. McDonald, T. P., Clift, R., Cottrell, M., and Lond, M. D. Further studieson the purification and assay of thrombopoietin. Prog. Clin. Biol. Res., 2/5:215-219, 1986. 158.
131. Tayrien, G., and Rosenberg. R. D. Purification and properties of a mega-karyocyte stimulatory factor present both in the serum-free conditionedmedium of human embryonic kidney cells and in thrombocytopenic plasma. 159.J. Biol. Chem., 262: 3262-3268, 1987.
132. Goodman. J. W., Hall, E. A., Miller, K. L., and Shinpock, S. G. Interleukin-3 promotes erythroid burst formation in "serum free" cultures withoutdelectable erythropoietin. Proc. Nail. Acad. Sci. USA, 82:3291-3295,1985. 160.
133. Nicola, N., Vadas, M. A., and Lopez, A. F. Down modulation of receptorsfor granulocyte colony-stimulating factor on human neutrophils by granu-locyte-activating agents. J. Cell. Physiol., 128: 501-509, 1986. 161.
134. Walker, F.. and Burgess, A. W. Specific binding of radioiodinated granulocyte-macrophage colony-stimulating factor to hemopoielic cells. EMBOJ., 4: 933-939. 1985.
135. Gasson, J. C., Kaufman, S. E.. Weisbart, R. H.,Tomonaga, M., and Golde, 162.D. W. High-affinity binding of granulocyte-macrophage colony-stimulatingfactor to normal and leukemic human myeloid cells. Proc. Nati. Acad. Sci.USA, «J:699-673, 1986. 163.
136. Sherr, C. J., Rettenmier, C. W., Sacca, R., Roussel, M. F., Look, A. T., andStanley, E. R. The c-fms proto-oncogene product is related to the receptorfor the mononuclear phagocyte growth factor, CSF-1. Cell, 41: 665-676, 164.1985.
137. Stanley, I. J.. and Burgess. A. W. Granulocyte-macrophage colony stimulating factor stimulates the synthesis of membrane and nuclear proteins inmurine neutrophils. J. Cell. Biochem., 23: 241-258, 1983. 165.
138. Fleischmann, J., Golde, D. W., Weisbart, R. H., and Gasson, J. C. Granulocyte-macrophage colony-stimulating factor enhances phagocytosis of bacteria by human neutrophils. Blood, 68: 708-711. 1986.
139. Begley, C. G., Lopez, A. F., and Nicola, N. A. Purified colony stimulating 166.factors enhance the survival of human neutrophils and eosinophils in vitro;
5636
a rapid and sensitive microassay for colony stimulating factors. Blood, 68:162-166. 1986.Vadas. M. A., Nicola, N. A., and Metcalf, D. Activation of antibody-dependent cell-mediated cytotoxicity of human neutrophils and eosinophilsby separate colony-stimulating factors. J. Immunol., 130: 795-799, 1983.Weisbart, R. H., Golde, D. W., Clark, S. C., et al. Human granulocyte-macrophage colony-stimulating factor is a neutrophil activator. Nature(Lond.), 314: 361-363, 1985.Vadas, M. A., Lopez, A. F., and Williamson, D. J. Selective enhancementof the expression of granulocyte functional antigens 1 and 2 on humanneutrophils. Proc. Nati. Acad. Sci. USA, 82: 2503-2507, 1985.Lopez, A. F., Nicola, N. A., Burgess, A. W., el al. Activation of granulocytecytotoxic function by purified mouse colony-stimulating factors. J. Immunol., 131: 2983-2988, 1983.Handman. E., and Burgess, A. W. Stimulation by granulocyte-macrophagecolony stimulating factor of Leishmania tropica killing by macrophages. J.Immunol., 122: 1134-1137, 1979.v ill.ili.i. F., and Kierszenbaum, F. Effects of human colony-stimulatingfactor on the uptake and destruction of a pathogenic parasite (Trypanosomacruz/) by human neutrophils. J. Immunol., 137: 1703-1707, 1986.Pizzo, P. A., Robichand, K. J., Wesley, R., and Commers, J. A. Fever inthe pediatrie and young adult patient with cancer. Medicine (Baltimore),61: 153-165, 1982.Pizzo, P. A., Hathorn, J. W., Hiemenz, J., et al. A randomized trialcomparing ceftazidine alone with combination antibiotic therapy in cancerpatients with fever and neutropenia. N. Engl. J. Med., 315: 552-558, 1986.Pickering, L. K., Anderson. D. L., Choi, S., et al. Leukocyte function inchildren with malignancies. Cancer (Phila.), 35: 1365-1371, 1975.Vadas, M. A., Varigos, G., Nicola, N., et al. Eosinophil activation bycolony-stimulating factor in man; metabolic effect and analysis by flowcytometry. Blood, 61: 1232-1241, 1983.Fleit, H. B., and Rabinovitch, M. Interferon induction in marrow-derivedmacrophages: regulation by L cell conditioned medium. J. Cell. Physiol.,108: 347-352, 1981.Lin. U.S.. and Gordon, S. Secretion of plasminogen activator by bonemarrow-derived mononuclear phagocytes and its enhancement by colony-stimulating factor. J. Exp. Med., 150: 231-245, 1979.Hamilton, J. A., Stanley, E. R., Burgess, A. W., and Shadduck, R. K.Stimulation of macrophage plasminogen activator activity by colony-stimulating factors. J. Cell. Physiol., 103:435-445, 1980.Kindler, V., Thorens, B., De Kossodo, S., Allet, B., Eliason, J. F., Thatcher,D., Farber. N., and Vassalli, P. Stimulation of hematopoiesis in vivo byrecombinant bacterial murine interleukin 3. Proc. Nati. Acad. Sci. USA, .S1(1001-1005, 1986.Metcalf, D., Begley, C. G., Johnson, G. R., Nicola, N. A., Lopez, A. F.,and Williamson, D. J. Effects of bacterially synthesized murine multi-CSF(IL-3) on hematopoiesis in normal adult mice. Blood, 68: 46-57, 1987.Lord, B., Molineux, G., Testa, N. G., Kelly, M., Spooncer, F., and Dexter,T. M. The kinetic response of haemopoietic precursor cells in vivo to highlypurified recombinant interleukin-3. Lymphokine Res., 5: 97-104, 1986.Tamura, M., Hattori, K., Nomura, H., et al. Induction of neutrophilicgranulocytosis in mice by administration of purified human native granulocyte colony-stimulating factor (G-CSF). Biochem. Biophys. Res. Commun., 142: 454-460, 1987.Matsumoto, M., Matsubara, S., Matsuno, T., Tamura, H., Hattori, K.,Nomura, H., and Ono, M. Protective effect of human granulocyte colony-stimulating factor (G-CSF) on microbial infection in mice with neutropenia.Infect. Immun., 55: 2715-2720, 1987.Bennett, J. M., Catovsky, D., Daniel, M. T., et al. Proposals for theclassification of the acute leukemias French-American-British (FAB) Cooperative Group. Br. J. Hematol., 33: 451-458, 1976.Fearon, E. R., Burke, P. J., Schiffer, C. A„Zehnbauer, B. A., and Vogelstein,B. Differentiation of leukemia cells to polymorphonuclear leukocytes inpatients with acute nonlymphocytic leukemia. N. Engl. J. Med., 315: 15-24, 1986.Moore, M. A. S., Williams, N., and Metcalf, D. In vitro colony formationby normal and leukemic human hematopoietic cells: characterization of thecolony-forming cells. J. Nati. Cancer Inst., 50: 603-623, 1973.Griffen, J. D., Young, D., Herrmann, F., Wiper, D., Wagner, K., andSabbath, K. D. Effects of recombinant human GM-CSF on proliferation ofclonogenic cells in acute myeloblastic leukemia. Blood, 67: 1448-1453,1986.Maeda, M., and Ichikawa, Y. Production of a colony-stimulating factorfollowing differentiation of leukemic myeloblasts to macrophages. J. Cell.Physiol., ¡02:323-331, 1980.Metcalf, D., Moore, M. A. S., Sheridan, J. W., and Spitzer, G. Responsiveness of human granulocytic leukemic cells to colony-stimulating factor.Blood, 43: 847-859, 1974.Lang, R. A., Metcalf, D., Gough, N. M., Dunn, A. R., and Gonda, T. J.Expression of a haemopoietic growth factor cDNA in a factor dependentcell line results in autonomous growth and tumorigenicity. Cell, 43: 531-542, 1985.Morstyn, G., Nicola, N., and Metcalf, D. Separate actions of differentcolony stimulating factors from human placenta! conditioned medium onhuman hemopoietic progenitor cell survival and proliferation. J. Cell. Physiol., 109: 133-142, 1981.Begley, C. G., Metcalf, D., and Nicola, N. A. Binding characteristics andproliferative action of purified granulocyte colony-stimulating factor (G-
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HEMOPOIETIC GROWTH FACTORS
CSF) on normal and leukemic human promyelocytes. Exp. Hematol., 16:71-79, 1988.
167. Dexter, T. M., Garland, J., Scott, D., Scolnick, E., and Metcalf, D. Growthof factor-dependent hemopoietic precursor cell lines. J. Exp. Med., 752:1036-1047. 1980.
168. Schrader. J. W., and Crapper, R. M. Autogenous production of a hemopoietic growth factor, persisting-cell-stimulating factor, as a mechanism fortransformation of bone marrow-derived cells. Proc. Nati. Acad. Sci. USA,30:6892-6896, 1983.
169. Weinstein, Y., Ihle, J. N., Lavu, S., and Reddy, P. Truncation of the c-mybgene by a retroviral integration in an interleukin 3-dependent myeloidleukemia cell line. Proc. Nati. Acad. Sci. USA, 83: 5010-5014, 1986.
170. Schwarzbaum, S., Halpern, R., and Diamond, B. The generation of mac-rophage-like cell lines by transfection with SV-40 origin defective DNA. J.Immunol., 132: 1158-1162, 1984.
171. Cook, W. D., Metcalf, D., Nicola, N. A., Burgess, A. W., and Walker, F.Malignant transformation of a growth factor-dependent myeloid cell line byAbelson virus without evidence of an autocrine mechanism. Cell, 41: 677-683, 1985.
172. Pierce, J. H., Di Fiore, P. P., Aaronson, S. A., et al. Neoplastic transformation of mast cells by Abelson-MuLV: abrogation of IL-3 dependence bya nonautocrine mechanism. Cell, 41:685-693, 1985.
173. Barlow, D. P., Bucan, M., Lehrach, H., Hogan, B. L. M., and Gough, N.M. Close genetic and physical linkage between the murine hemopoieticgrowth factor genes GM-CSF and multi-CSF (IL-3). EMBO J., 6:617-624,1987.
174. Huebner, K., Isobe, M., Croce, C. M., Golde, D. W., Kaufman, S. E., andGasson, J. C. The human gene encoding GM-CSF is at 5q21-q32, thechromosome region deleted in the 5q—anomaly. Science (Wash. DC), 230:1282-1285, 1985.
175. Pettenati, M. J., Le Beau, M. M., Lemons, R. S., et al. Assignment of CSF-1 to 5q33.1 : evidence for clustering of genes regulating hematopoiesis andfor their involvement in the deletion of the long arm of chromosome 5 inmyeloid disorders. Proc. Nati. Acad. Sci. USA, 84: 2970-2974, 1987.
176. Knudtzon, S., and Mortensen, B. T. Growth stimulation of human bonemarrow cells in agar culture by vascular cells. Blood, 46:937-943, 1975.
177. Nicola, N. A., Burgess, A. W., and Metcalf, D. Similar molecular propertiesof granulocyte-macrophage colony-stimulating factors produced by differentmouse organs in vitro and in vivo. J. Biol. Chem., 254: 5290-5299, 1979.
178. r,ali-Ili. A., Dosne, A., Morin, A., Dubor, F., and Chedid, L. Stimulation ofhuman endothelial cells by synthetic muramyl peptides: production ofcolony-stimulating activity (CSA). Exp. Hematol., 13:1157-1163, 1985.
179. Hammer, S. M., Gillis, J. M., Groopman, J. E., and Rose, R. M. In vitromodification of human immunodeficiency virus infection by granulocyte-macrophage colony stimulating factor and y interferon. Proc. Nati. Acad.Sci. USA, 83:8734-8738, 1986.
180. Broudy, V. C., Zuckerman, K. S., Jetmalani, S., Fitchen, J. H., and Bagby,G. C. Monocytes stimulate fibroblastoid bone marrow stromal cells toproduce multi-lineage hematopoietic growth factors. Blood, 68: 530-534,1986.
181. Koury, M. J., and Pragnell, I. B. Retroviruses induce granulocyte macrophage colony stimulating activity in fibroblasts. Nature (Lond.), 299: 638-640, 1982.
182. Chan, J. Y., Shimon. D. J., Nimer, S. D., Golde, D. W., and Gasson, J. C.Regulation of expression of human granulocyte/macrophage colony stimulating factor. Proc. Nati. Acad. Sci. USA, 83:8669-8673, 1986.
183. Mangalik, A., and Robinson, W. Cyclic neutropenia: the relationship between urine granulocyte colony stimulating activity and neutrophil count.Blood, 41: 79-84, 1973.
184. Koshaki, M., Noguchi, K., Araki, K., et al. In vivo stimulation of murinegranulopoiesis by human urinary extract of patients with aplastic anemia.Proc. Nati. Acad. Sci. USA, 80:3802-3806, 1983.
185. Amato, D., Freeman, M. H., and Saunders, F. F. Granulopoiesis in severecongenital neutropenia. Blood, 47: 531-538, 1976.
186. Golde, D. W., and Cline, M. J. Endotoxin-induced release of colony-stimulating activity in man. Proc. Soc. Exp. Biol. Med., 149: 845-848,1975.
187. Hinterberger, W., Paukovits, W. R., and Mittermayer, K. Endotoxin-induced colony stimulating activity in normal and myeloid leukaemic subjects.Scand. J. Haematol., 22: 280-287, 1979.
188. Metcalf, D., Chan, S. H., Gunz, F. W., Vincent, P., and Ravich, R. B. M.Colony-stimulating factor and inhibitor levels in acute granulocytic leukemia. Blood, 38: 143-152, 1971.
189. Foster, R., Metcalf, D., Robinson, W. A., and Bradley. T. R. Bone marrowcolony stimulating activity in human sera. Br. J. Haematol., 15: 147-159,1968.
190. Metcalf, D., and Wahren, B. Bone marrow colony stimulating activity ofsera in infectious mononucleosis. Br. Med. J., 3:99-101, 1968.
191. Metcalf. D. Induction of differentiation in murine myelomonocytic leukemiacells by the serum of patients with acute myeloid leukemia and otherdiseases. Int. J. Cancer, 27: 577-584, 1981.
192. Wewerka. J. R., and Dale, D. C. Colony-stimulating factor in patients withchronic neutropenia. Blood, 47: 861-867, 1976.
193. Myers, A. M., and Robinson, W. A. Colony stimulating factor levels inhuman serum and urine following chemotherapy. Proc. Soc. Exp. Biol.Med., ¡48:694-700, 1975.
194. Shadduck, R. K., and Nagabhushan, N. G. Granulocyte colony stimulatingfactor. I. Response to acute granulocytopenia. Blood, 38: 559-568, 1971.
195. Sato, N., Asano, S., Ueyama, Y., et al. Granulopoiesis and colony-stimulating activity (CSA) produced by a human squamous cell carcinoma. Cancer(Phila.), «.-605-610, 1979.
196. Asano, S., Urabe, A., Okabe, T., et al. Demonstration of granulopoieticfactor(s) in the plasma of nude mice transplanted with a human lung cancerand in the tumor tissue. Blood, 49:845-852, 1977.
197. Yunis, A. A., Jiminez, J. J., Wu, M.-C, and Andreotti, P. E. Furtherevidence supporting an in vivo role for colony-stimulating factor. Exp.Hematol., 12: 838-843, 1984.
198. Suda, T., Miura, Y., Mizoguchi, H., Kubota, K., and Takaku, F. A case oflung cancer associated with granulocytosis and production of colony-stimulating activity by the tumour. Br. J. Cancer, 41: 980-984, 1980.
199. Cebón,J., Dempsey, P., Kannourakis, G., Fox, R., Bonnern, E., Burgess,A. W., and Morstyn, G. Pharmacokinetics of human granulocyte-macrophage colony stimulating factor (hGM-CSF) using a sensitive immunoassay.Blood, in press, 1988.
200. Morstyn, G., Campbell, L., Souza, L. M. Alton, N. K., Keech, J., Green,M., Sheridan, W., Metcalf, D., and Fox, R. Effect of granulocyte colonystimulating factor on neutropenia induced by cytoxic chemotherapy. Lancet,2:667-672, 1988.
201. Bronchud, M. H., Scarffe, J. H., Thatcher, N., Crowther, D., Souza, L. M.,Alton, N. K., Testa, N. G., and Dexter, T. M. Phase I/II study of recombinant human granulocyte colony-stimulating factor in patients receivingintensive chemotherapy for small cell lung cancer. Br. J. Cancer, 56: 809-813, 1987.
202. Gabrilove, J., Jakubowski, A., Fain, K., Scher, H., Grous, T., Sternberg, C.,Yagoda, A., Clarkson, B., Moore, M. A. S., Bonilla, M. A., Oettgen, H. F.,Alton, K., Downing, M., Weite, K., and Souza, L. M. A phase I/II study ofrhG-CSF in cancer patients at risk for chemotherapy-induced neutropenia.Proc. Am. Soc. Hematol., 394: 135a, 1987.
203. Morstyn, G., Lieschke, G., Cebón,J., Mäher,D., Villeval, J. L., Duhrsen,U., McGrath, K., Boyd, A., et al. Phase I study of bacterially synthesizedgranulocyte macrophage colony stimulating factor (rh-bs GM-CSF). In:Proceedings of the International Society of Heamtology Meeting, Milan,August 1988.
204. Brandt, S. J., Peters, R. A., Waters, K., et al. Effect of recombinant humangranulocyte-macrophage colony-stimulating factor on hematopoietic reconstitution after high dose of chemotherapy and autologous bone marrowtransplantation. N. Engl. J. Med., 318:869-876. 1988.
205. Vadhan-Raj, S., Keating, M., LeMaistre, A., Hittelman, W. N., McCredie,K., Trujillo, J. M., Broxmeyer, H. E., Henney, C., and Gutterman, J. U.Effects of recombinant human granulocyte-macrophage colony-stimulatingfactor in patients with myelodysplastic syndromes. N. Engl. J. Med., 317:1545-1552,1987.
206. Mertelsmann, R., and Cosman, D. Colony-stimulating factors in vivo andin vitro. Immunol. Today, 9: 97-98, 1988.
207. Ruff, M. R., Farrar, W. L., and Pert, C. B. Interferon y and granulocyte/macrophage colony stimulating factor inhibit growth and induce antigenscharacteristics of myeloid differentiation in small-cell lung cancer cells.Proc. Nati. Acad. Sci. USA, 83:6613-6617, 1986.
208. Baldwin, G. C., Dipersio, J., Kaufman, S. E., Quan, S. G., Golde, D. W,and Gasson, J. C. Characterization of human GM-CSF receptors on non-hematopoietic cells. Blood Suppl., 70: 166a, 1987.
209. Kelleher, C., Miyauchi, J., Wong, G., Clark, S., Minden, M. D., andMcCulloch, E. A. Synergism between recombinant growth factors GM-CSFand G-CSF, acting on the blast cells of acute myeloblastic leukemia. J. CellBiochem.. Suppl. 2A, p. 216, 1987.
210. Quesenberry, P. J. Synergistic hematopoietic growth factors. Int. J. CellCloning, 4: 3-15, 1986.
5637
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1988;48:5624-5637. Cancer Res George Morstyn and Antony W. Burgess Hemopoietic Growth Factors: A Review
Updated version
http://cancerres.aacrjournals.org/content/48/20/5624
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/48/20/5624To request permission to re-use all or part of this article, use this link
on June 23, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from