Embed Size (px)
Citation preview

Hepatic Preconditioning in Rats Is Definedby a Balance of Adenosine and Xanthine
CARMEN PERALTA, DANIEL CLOSA, CARME XAUS, EMILIO GELPI, JOAN ROSELLO-CATAFAU, AND GEORGINA HOTTER
The present work investigates the relationship betweenadenosine, nitric oxide (NO), and free radicals duringischemic preconditioning in rat liver. For this purpose, weevaluated: 1) the efficacy of different periods of precondition-ing; 2) the changes in the concentration of adenine nucleo-tides during preconditioning; 3) the importance of adeno-sine and xanthine concentrations in the induction ofpreconditioning; and 4) the possible effect of xanthineoxidase-derived superoxide anion on NO during precondi-tioning. Results show that just a 10- to 15-minute period ofischemia followed by 10-minute reperfusion prevents theischemic damage that would be induced by a subsequent 90minutes of ischemia followed by 90 minutes of reperfusion.Administration of xanthine or metabolization of endog-enous adenosine abolishes the protective effect of precondi-tioning. When rats have been subjected to a period ofpreconditioning not within the effective time window(10-15 minutes), and thus offering no protection, theadministration of a NO donor was found to restore theprotection. The dose needed to restore protection appearsto be proportional to the endogenous xanthine concentra-tion. In addition, when xanthine oxidase was inhibited,preconditioning effectively offered protection in front ofischemia and reperfusion, independently of the xanthineconcentration. Altogether, this indicates that the timewindow of ischemia capable to induce preconditioning inliver is defined by the relative tissue concentrations ofadenosine and xanthine. The lower limit of this window (10minutes) is defined by the amount of adenosine able toinduce NO generation. Its upper limit (15 minutes) isdefined by the concentration of xanthine able to remove thegenerated NO. (HEPATOLOGY 1998;28:768-773.)
In 1986, Murry et al. discovered that short periods ofischemia and reperfusion render the myocardium resistant to
a subsequent more-sustained ischemic insult.1 This phenom-enon, called ischemic preconditioning, has been commonlystudied in the heart, but few studies have been performed onintestine2 or liver.3 The mechanism underlying precondition-ing remains unknown and is currently under intense investi-gation. It has been suggested that protection depends on therelease of substances by the organ helping to protect it againstinjury. Along this line, potential mediators include nitricoxide (NO)2,4 and adenosine.5
Vegh et al.6 reported that the protection conferred bypreconditioning in dog heart in vivo disappeared after NOinhibition. Nevertheless, other authors7 found that endog-enous NO is not a mediator in the ischemic preconditioningin the isolated rat heart. Recent work by our group, using amodel of hepatic preconditioning in rat, demonstrated thatinhibition of NO abolishes the effect of preconditioning.8 Inthe same study, we suggested that NO production wasstimulated by endogenous adenosine. It is known thatpretreatment with adenosine or selective adenosine A1 recep-tor agonists mimics the effect of preconditioning in heart.10
Both adenosine and NO are generated in the endothelium andreleased into the vascular and interstitial space during theprocess of ischemia and reperfusion. It appears that during ashort ischemic period, cellular consumption of high-energyadenine phosphate nucleotides leads to accumulation ofadenosine.10 During reperfusion, NO is generated as a resultof adenosine receptor activation and triggers the precondi-tioning.
There is evidence that only very short and transient periodsof ischemia and reperfusion are able to induce the protectiveeffect of preconditioning. In the present work, we describeexperiments designed to explore the mechanisms that justifywhy only short, and limited, periods of ischemia induce thehepatic protection conferred by preconditioning.
MATERIALS AND METHODS
Adenosine, adenosine deaminase (ADA), allopurinol, and xan-thine were obtained from Sigma Chemical (St. Louis, MO). Sper-mine NONOate was purchased from Cayman Chemical (AnnArbor, MI).
Animal Mode of Hepatic Ischemia
Male Wistar rats (240-280 g body weight) were used. All animals(including controls) were anesthetized with urethane (10 mg/kgintraperitoneally) and placed in a supine position on a heating padfor maintenance of body temperature between 36°C and 37°C. Toinduce hepatic ischemia, laparotomy was performed, and the bloodsupply to the right lobe of the liver was interrupted by placement ofa bulldog clamp at the level of the hepatic artery and portal vein.Reflow was initiated by removal of the clamp.8 The studies per-formed were in compliance with the European Union regulations foranimal experiments.
Abbreviations: ADA, adenosine deaminase; AST, aspartate transaminase; ALT, alaninetransaminase; ATP, adenosine triphosphate; ADP, adenosine diphosphate; AMP, adeno-sine monophosphate; I/R, ischemia reperfusion; Prec, preconditioning.
From the Department of Medical Bioanalysis, IIBB-CSIC,* IDIBAPS,** Barcelona,Spain.
Received January 8, 1998; accepted April 21, 1998.*IIBB-CSIC-Instituto de Investigaciones Biomedicas de Barcelona-Consejo Superior
de Investigaciones Cientıficas.**IDIBAPS-Institut d’Investigacions Biomediques August Pi i Sunyer.Supported by grant 97/2076 Fondo de Investigaciones Sanitarias.Address reprint requests to: Dr. Georgina Hotter, Department of Medical Bioanalysis,
Instituto de Investigaciones Biomedicas de Barcelona, IIBB, CSIC, C/Jordi Girona,18-26, 08034 Barcelona, Spain. Fax: 34-93-2045904.
Copyright r 1998 by the American Association for the Study of Liver Diseases.0270-9139/98/2803-0025$3.00/0
768

Experimental Protocols
Ischemic Period to Induce Preconditioning. A control group (n 5 6)was subjected to anesthesia and laparotomy. A second group ofanimals (n 5 6) were subjected to 90 minutes of right-lobe hepaticischemia, followed by 90-minute reperfusion. To test the effective-ness of different preconditioning periods, six groups of animals (n 56) were each subjected to a different period of preconditioning,followed by 90-minute ischemia and 90-minute reperfusion. Thepreconditioning times tested were: 2, 5, 10, 15, 20, and 30 minutesof ischemia, followed in each case by a 10-minute reperfusion. Bloodsamples were collected at the end of the protocol and processed todetermine plasma transaminases (aspartate transaminase [AST],alanine transaminase [ALT]), used herein as markers of hepaticinjury.
High-Energy Nucleotides and Their Breakdown Products. To evaluatethe effect of a short ischemic period on high-energy nucleotides andtheir degradation products, a second series of experiments wasperformed. For this purpose, seven groups of animals (n 5 6) wereeach subjected to a different period of preconditioning ischemia (0,2, 5, 10, 15, 20, 30 minutes), and immediately after, a portion of liverwas excised and promptly frozen (within 10 seconds). These tissuesamples were used to evaluate the levels of adenosine triphosphate(ATP), adenosine diphosphate (ADP), adenosine monophosphate(AMP), adenosine, inosine, and xanthine by high-performanceliquid chromatography.
Modulation of Preconditioning. To evaluate the importance of adeno-sine and xanthine concentrations in the appearance of the precondi-tioning, rats were either infused with xanthine or administeredADA. The latter was performed to induce the metabolism ofendogenous adenosine. For this purpose, eight groups of animals(n 5 6) underwent procedures according to the following protocol:1) Control: animals subjected to anesthesia and laparotomy; 2) I/R:animals subjected to 90 minutes of right-lobe hepatic ischemia,followed by 90 minutes of reperfusion; 3) Prec. 10 min: same asgroup 2, but animals were subjected to 10 minutes of ischemia and10 minutes of reperfusion previous to I/R; 4) Prec. 10 min. 1 ADA:same as group 3, but animals were treated with a continuousintravenous infusion of ADA (0.75 U/mL) dissolved in bicarbonate-buffered saline (pH 7.4) (0.066 mL/min, portal vein)8 20 minutesbefore preconditioning and throughout the preconditioning process;5) Prec. 10 min 1 xanthine: same as group 3, but animals weretreated with a continuous intravenous infusion of 5 mmol/Lxanthine dissolved in bicarbonate-buffered saline (pH 7.4) (0.066mL/min, portal vein)11 5 minutes before preconditioning andthroughout the preconditioning process; 6) Prec. 5 min: same asgroup 2, but animals were subjected to 5 minutes of ischemia and 10minutes of reperfusion previous to ischemia and reperfusion; 7)Prec. 5 min 1 adenosine: same as group 6, but animals were treatedwith a continuous intravenous infusion of adenosine (0.1 µmol/min/kg) dissolved in bicarbonate-buffered saline (pH 7.4) (0.066 mL/min, portal vein)8 during 20 minutes before preconditioning andthroughout the preconditioning process; 8) Prec. 5 min 1 adenosine 1xanthine: same as group 6, but animals were treated with acontinuous infusion of adenosine (0.1 µmol/min/kg) dissolved inbicarbonate-buffered saline (pH 7.4) (0.066 mL/min, portal vein)during 20 minutes before preconditioning and throughout thepreconditioning process and a continuous infusion of 5 mmol/Lxanthine dissolved in bicarbonate-buffered saline (pH 7.4) (0.066mL/min, portal vein) 5 minutes before preconditioning and duringthe preconditioning process.
Blood samples were obtained after reperfusion to determine thelevels of AST and ALT activities.
Effect of Xanthine on the Protection Conferred by NO. The possibleeffect of xanthine/xanthine oxidase-derived superoxide anions onNO was evaluated by administering increasing concentrations of aNO donor in animals subjected to different periods of precondition-ing (5 or 20 minutes) previously established not to offer anyprotection in front of the ischemia-reperfusion: 9) I/R (n 5 6):
animals subjected to 90 minutes of right-lobe hepatic ischemia,followed by 90 minutes of reperfusion.
10) Prec. 2 min (n 5 24): same as group 9 but with previouspreconditioning consistent in 2 minutes of ischemia and 10 minutesof reperfusion. This group was divided into four subgroups (eachn 5 6) in which 0, 1.5, 9, or 21 mg/kg body weight of spermidineNONOate was intravenously administered immediately before thestart of the 10-minute reperfusion; 11) Prec. 20 min (n 5 24): sameas group 9 but with previous preconditioning consistent in 20minutes of ischemia and 10 minutes of reperfusion. This group wasdivided into four subgroups (each n 5 6) in which 0, 1.5, 9, or 21mg/kg body weight of spermidine NONOate was intravenouslyadministered immediately before the start of the 10-minute reperfu-sion; 12) Prec. 20 min 1 allopurinol (n 5 24): same as group 11, butwith inhibition of xanthine oxidase by administration of allopurinol(intraperitoneally, 150 mg/kg) 30 minutes before the start of theprotocol.
Blood samples were obtained after reperfusion to determine ASTand ALT.
Biochemical Assays
Nucleotide Analysis. Liver samples, weighing approximately 100mg, were placed in 1.0 mL of 3.6% perchloric acid solution and thenimmediately homogenized (S25N-8G, IKA, Labortechnik, Stauffen,Germany). Following homogenization, tissues were allowed toextract for 30 minutes at 0.5°C, and were centrifuged at 850g for 15minutes at 0.5°C. Supernatants were neutralized with a solution ofpotassium carbonate 6N/potassium hydroxide 6N: 1/1 to a pH of 6.0to 6.5 and frozen at 270°C. Frozen samples were thawed, centri-fuged at 13,000g for 2 minutes, and then 50 µL of the supernatantwas directly injected into liquid chromatographic equipment. Nucle-otide profiles were obtained using a reversed-phase Spherisorb ODS(C18, 5-µm particle size, 15 3 0.4 cm; Teknokroma, San Cugat,Spain) coupled to a 600S HPLC system (Waters, Milford, MA)equipped with a 1000S Diode Array UV detector (Applied Biosys-tems, Foster City, CA). The absorbance was monitored at 254 nm.Nucleotide separation was allowed to proceed in an isocratic fashionwith 100 mmol/L ammonium phosphate (pH 5.5), until ATP, ADP,hypoxanthine/xanthine, and AMP were separated. At this point, amixture of methanol:water (96:4) was introduced into the column,eluting inosine. A mixture of methanol:water (60:40) was intro-duced after the inosine to elute adenosine.12 Under these conditions,hypoxanthine and xanthine coelute, so that the values given hereinfor xanthine represent a combination of both metabolites. Calibra-tion chromatograms for the standards ATP, ADP, AMP, adenosine,inosine, hypoxanthine, and xanthine were generated by injecting 50µL of a mixture of known concentrations.
Enzymatic Determinations. Plasma ALT was measured by enzymaticreduction of pyruvate to lactate with oxidation of b-Nicotinamideadeninine dinucleotide reduced form, followed spectrophotometri-cally by the rate of absorption loss at 340 nm, using a commercial kitfrom Boehringer Mannheim (Munich, Germany).
Plasma AST was measured by enzymatic reduction of oxalacetateto lactate with oxidation of b-Nicotinamide adeninine dinucleotidereduced form, followed spectrophotometrically by the rate of absorptionloss at 340 nm, using a commercial kit from Boehringer Mannheim.
Statistics
Experimental results are expressed as means 6 SEM. Means ofdifferent groups were compared using a one-way ANOVA. TheStudent’s t test was performed for evaluation of significant differ-ences between groups. Significance was determined at the 5% level(P , .05).
RESULTS
Ischemic Period to Induce Preconditioning. The degree of he-patic damage, evaluated by the plasma levels of AST and ALT,induced after 90 minutes of ischemia and 90 minutes of
HEPATOLOGY Vol. 28, No. 3, 1998 PERALTA ET AL. 769
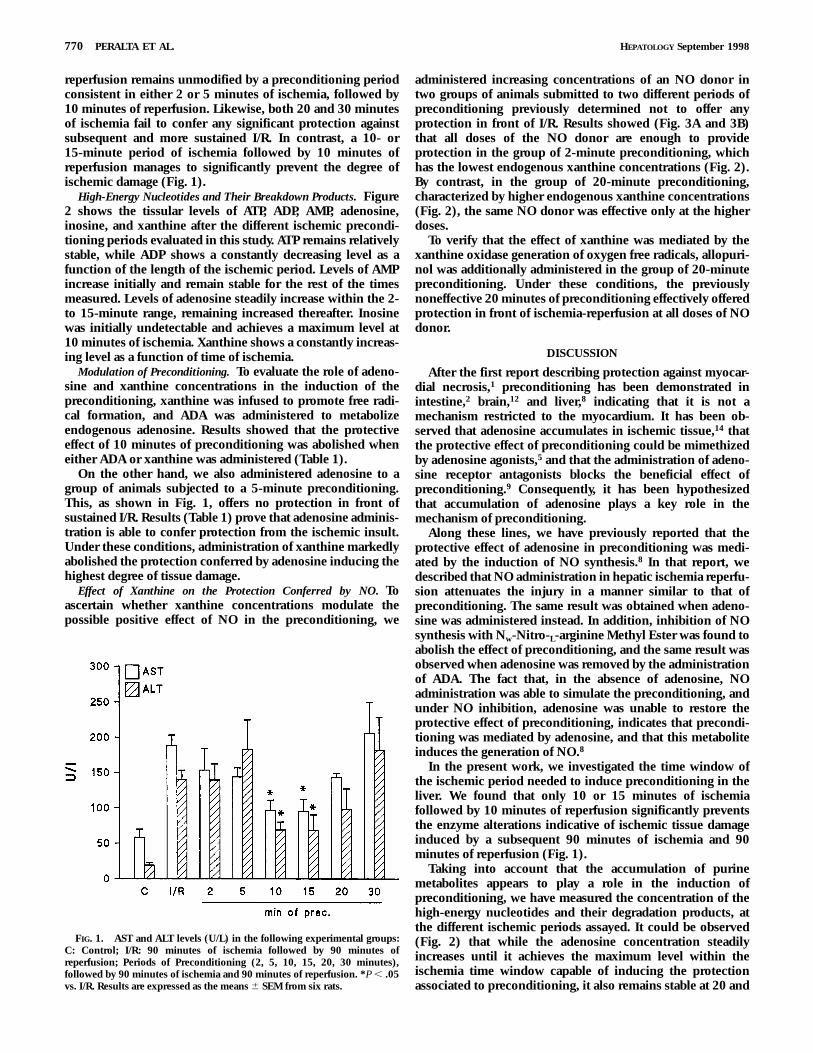
reperfusion remains unmodified by a preconditioning periodconsistent in either 2 or 5 minutes of ischemia, followed by10 minutes of reperfusion. Likewise, both 20 and 30 minutesof ischemia fail to confer any significant protection againstsubsequent and more sustained I/R. In contrast, a 10- or15-minute period of ischemia followed by 10 minutes ofreperfusion manages to significantly prevent the degree ofischemic damage (Fig. 1).
High-Energy Nucleotides and Their Breakdown Products. Figure2 shows the tissular levels of ATP, ADP, AMP, adenosine,inosine, and xanthine after the different ischemic precondi-tioning periods evaluated in this study. ATP remains relativelystable, while ADP shows a constantly decreasing level as afunction of the length of the ischemic period. Levels of AMPincrease initially and remain stable for the rest of the timesmeasured. Levels of adenosine steadily increase within the 2-to 15-minute range, remaining increased thereafter. Inosinewas initially undetectable and achieves a maximum level at10 minutes of ischemia. Xanthine shows a constantly increas-ing level as a function of time of ischemia.
Modulation of Preconditioning. To evaluate the role of adeno-sine and xanthine concentrations in the induction of thepreconditioning, xanthine was infused to promote free radi-cal formation, and ADA was administered to metabolizeendogenous adenosine. Results showed that the protectiveeffect of 10 minutes of preconditioning was abolished wheneither ADA or xanthine was administered (Table 1).
On the other hand, we also administered adenosine to agroup of animals subjected to a 5-minute preconditioning.This, as shown in Fig. 1, offers no protection in front ofsustained I/R. Results (Table 1) prove that adenosine adminis-tration is able to confer protection from the ischemic insult.Under these conditions, administration of xanthine markedlyabolished the protection conferred by adenosine inducing thehighest degree of tissue damage.
Effect of Xanthine on the Protection Conferred by NO. Toascertain whether xanthine concentrations modulate thepossible positive effect of NO in the preconditioning, we
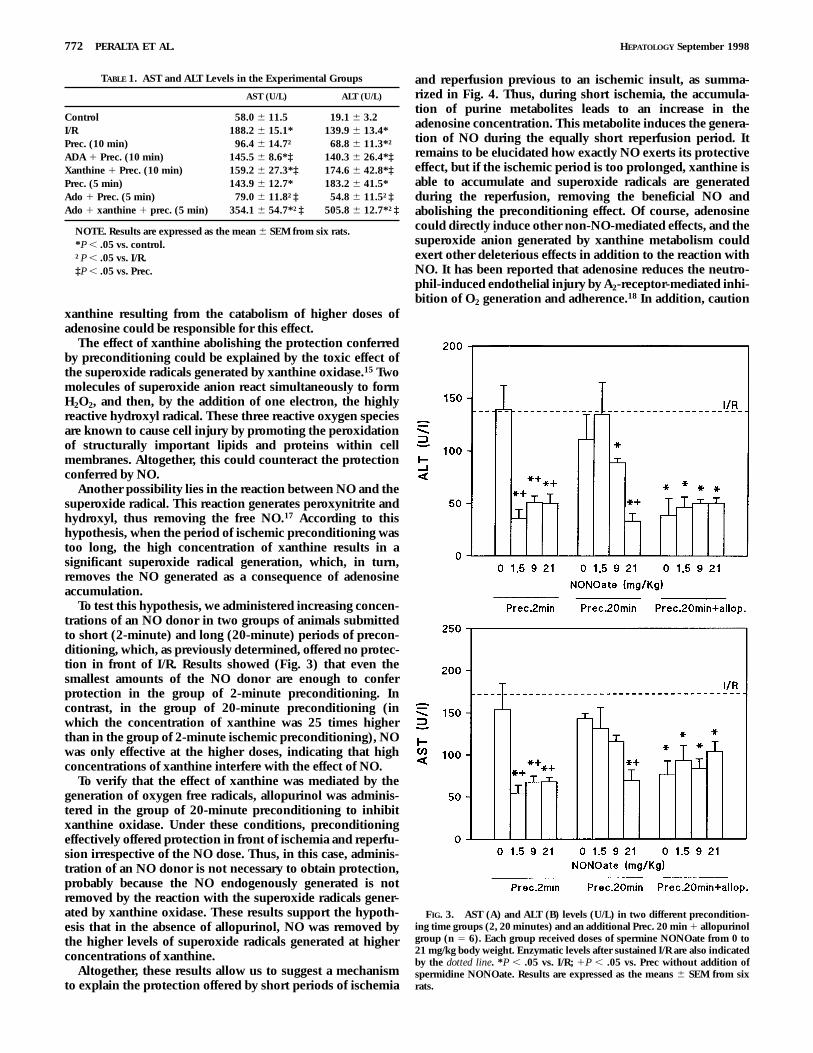
administered increasing concentrations of an NO donor intwo groups of animals submitted to two different periods ofpreconditioning previously determined not to offer anyprotection in front of I/R. Results showed (Fig. 3A and 3B)that all doses of the NO donor are enough to provideprotection in the group of 2-minute preconditioning, whichhas the lowest endogenous xanthine concentrations (Fig. 2).By contrast, in the group of 20-minute preconditioning,characterized by higher endogenous xanthine concentrations(Fig. 2), the same NO donor was effective only at the higherdoses.
To verify that the effect of xanthine was mediated by thexanthine oxidase generation of oxygen free radicals, allopuri-nol was additionally administered in the group of 20-minutepreconditioning. Under these conditions, the previouslynoneffective 20 minutes of preconditioning effectively offeredprotection in front of ischemia-reperfusion at all doses of NOdonor.
DISCUSSION
After the first report describing protection against myocar-dial necrosis,1 preconditioning has been demonstrated inintestine,2 brain,12 and liver,8 indicating that it is not amechanism restricted to the myocardium. It has been ob-served that adenosine accumulates in ischemic tissue,14 thatthe protective effect of preconditioning could be mimethizedby adenosine agonists,5 and that the administration of adeno-sine receptor antagonists blocks the beneficial effect ofpreconditioning.9 Consequently, it has been hypothesizedthat accumulation of adenosine plays a key role in themechanism of preconditioning.
Along these lines, we have previously reported that theprotective effect of adenosine in preconditioning was medi-ated by the induction of NO synthesis.8 In that report, wedescribed that NO administration in hepatic ischemia reperfu-sion attenuates the injury in a manner similar to that ofpreconditioning. The same result was obtained when adeno-sine was administered instead. In addition, inhibition of NOsynthesis with Nw-Nitro-L-arginine Methyl Ester was found toabolish the effect of preconditioning, and the same result wasobserved when adenosine was removed by the administrationof ADA. The fact that, in the absence of adenosine, NOadministration was able to simulate the preconditioning, andunder NO inhibition, adenosine was unable to restore theprotective effect of preconditioning, indicates that precondi-tioning was mediated by adenosine, and that this metaboliteinduces the generation of NO.8
In the present work, we investigated the time window ofthe ischemic period needed to induce preconditioning in theliver. We found that only 10 or 15 minutes of ischemiafollowed by 10 minutes of reperfusion significantly preventsthe enzyme alterations indicative of ischemic tissue damageinduced by a subsequent 90 minutes of ischemia and 90minutes of reperfusion (Fig. 1).
Taking into account that the accumulation of purinemetabolites appears to play a role in the induction ofpreconditioning, we have measured the concentration of thehigh-energy nucleotides and their degradation products, atthe different ischemic periods assayed. It could be observed(Fig. 2) that while the adenosine concentration steadilyincreases until it achieves the maximum level within theischemia time window capable of inducing the protectionassociated to preconditioning, it also remains stable at 20 and
FIG. 1. AST and ALT levels (U/L) in the following experimental groups:C: Control; I/R: 90 minutes of ischemia followed by 90 minutes ofreperfusion; Periods of Preconditioning (2, 5, 10, 15, 20, 30 minutes),followed by 90 minutes of ischemia and 90 minutes of reperfusion. *P , .05vs. I/R. Results are expressed as the means 6 SEM from six rats.
770 PERALTA ET AL. HEPATOLOGY September 1998
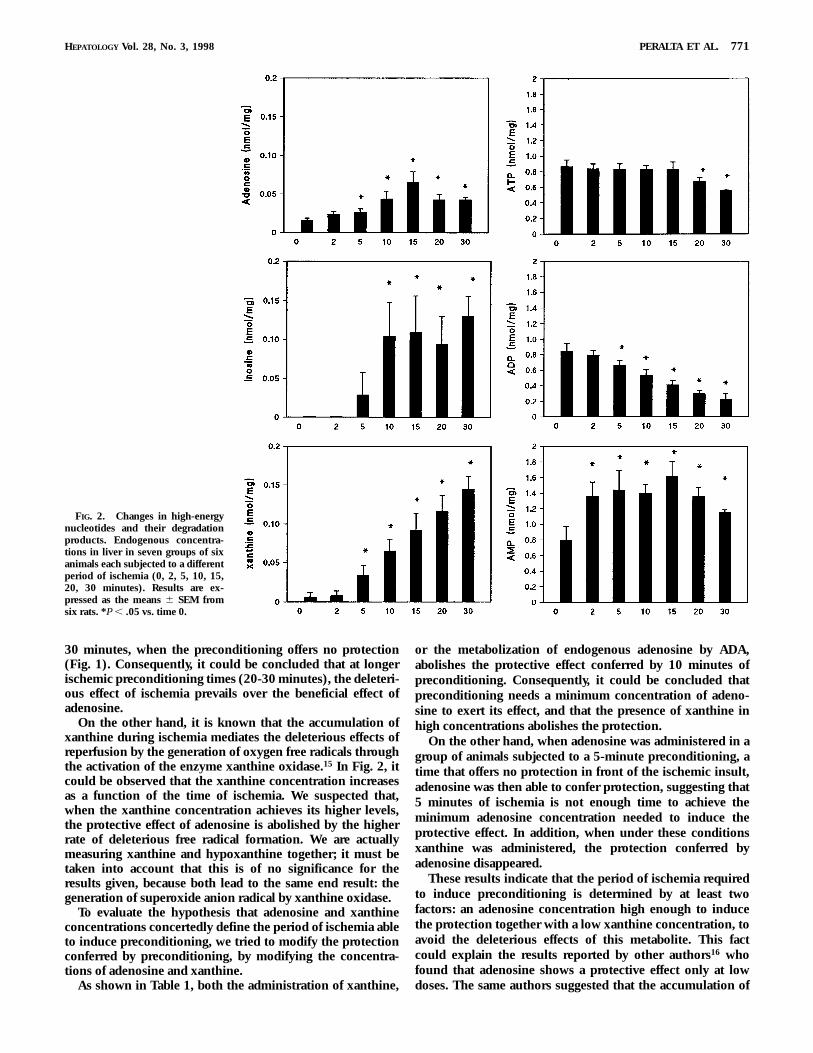
30 minutes, when the preconditioning offers no protection(Fig. 1). Consequently, it could be concluded that at longerischemic preconditioning times (20-30 minutes), the deleteri-ous effect of ischemia prevails over the beneficial effect ofadenosine.
On the other hand, it is known that the accumulation ofxanthine during ischemia mediates the deleterious effects ofreperfusion by the generation of oxygen free radicals throughthe activation of the enzyme xanthine oxidase.15 In Fig. 2, itcould be observed that the xanthine concentration increasesas a function of the time of ischemia. We suspected that,when the xanthine concentration achieves its higher levels,the protective effect of adenosine is abolished by the higherrate of deleterious free radical formation. We are actuallymeasuring xanthine and hypoxanthine together; it must betaken into account that this is of no significance for theresults given, because both lead to the same end result: thegeneration of superoxide anion radical by xanthine oxidase.
To evaluate the hypothesis that adenosine and xanthineconcentrations concertedly define the period of ischemia ableto induce preconditioning, we tried to modify the protectionconferred by preconditioning, by modifying the concentra-tions of adenosine and xanthine.
As shown in Table 1, both the administration of xanthine,
or the metabolization of endogenous adenosine by ADA,abolishes the protective effect conferred by 10 minutes ofpreconditioning. Consequently, it could be concluded thatpreconditioning needs a minimum concentration of adeno-sine to exert its effect, and that the presence of xanthine inhigh concentrations abolishes the protection.
On the other hand, when adenosine was administered in agroup of animals subjected to a 5-minute preconditioning, atime that offers no protection in front of the ischemic insult,adenosine was then able to confer protection, suggesting that5 minutes of ischemia is not enough time to achieve theminimum adenosine concentration needed to induce theprotective effect. In addition, when under these conditionsxanthine was administered, the protection conferred byadenosine disappeared.
These results indicate that the period of ischemia requiredto induce preconditioning is determined by at least twofactors: an adenosine concentration high enough to inducethe protection together with a low xanthine concentration, toavoid the deleterious effects of this metabolite. This factcould explain the results reported by other authors16 whofound that adenosine shows a protective effect only at lowdoses. The same authors suggested that the accumulation of
FIG. 2. Changes in high-energynucleotides and their degradationproducts. Endogenous concentra-tions in liver in seven groups of sixanimals each subjected to a differentperiod of ischemia (0, 2, 5, 10, 15,20, 30 minutes). Results are ex-pressed as the means 6 SEM fromsix rats. *P , .05 vs. time 0.
HEPATOLOGY Vol. 28, No. 3, 1998 PERALTA ET AL. 771
xanthine resulting from the catabolism of higher doses ofadenosine could be responsible for this effect.
The effect of xanthine abolishing the protection conferredby preconditioning could be explained by the toxic effect ofthe superoxide radicals generated by xanthine oxidase.15 Twomolecules of superoxide anion react simultaneously to formH2O2, and then, by the addition of one electron, the highlyreactive hydroxyl radical. These three reactive oxygen speciesare known to cause cell injury by promoting the peroxidationof structurally important lipids and proteins within cellmembranes. Altogether, this could counteract the protectionconferred by NO.
Another possibility lies in the reaction between NO and thesuperoxide radical. This reaction generates peroxynitrite andhydroxyl, thus removing the free NO.17 According to thishypothesis, when the period of ischemic preconditioning wastoo long, the high concentration of xanthine results in asignificant superoxide radical generation, which, in turn,removes the NO generated as a consequence of adenosineaccumulation.
To test this hypothesis, we administered increasing concen-trations of an NO donor in two groups of animals submittedto short (2-minute) and long (20-minute) periods of precon-ditioning, which, as previously determined, offered no protec-tion in front of I/R. Results showed (Fig. 3) that even thesmallest amounts of the NO donor are enough to conferprotection in the group of 2-minute preconditioning. Incontrast, in the group of 20-minute preconditioning (inwhich the concentration of xanthine was 25 times higherthan in the group of 2-minute ischemic preconditioning), NOwas only effective at the higher doses, indicating that highconcentrations of xanthine interfere with the effect of NO.
To verify that the effect of xanthine was mediated by thegeneration of oxygen free radicals, allopurinol was adminis-tered in the group of 20-minute preconditioning to inhibitxanthine oxidase. Under these conditions, preconditioningeffectively offered protection in front of ischemia and reperfu-sion irrespective of the NO dose. Thus, in this case, adminis-tration of an NO donor is not necessary to obtain protection,probably because the NO endogenously generated is notremoved by the reaction with the superoxide radicals gener-ated by xanthine oxidase. These results support the hypoth-esis that in the absence of allopurinol, NO was removed bythe higher levels of superoxide radicals generated at higherconcentrations of xanthine.
Altogether, these results allow us to suggest a mechanismto explain the protection offered by short periods of ischemia
and reperfusion previous to an ischemic insult, as summa-rized in Fig. 4. Thus, during short ischemia, the accumula-tion of purine metabolites leads to an increase in theadenosine concentration. This metabolite induces the genera-tion of NO during the equally short reperfusion period. Itremains to be elucidated how exactly NO exerts its protectiveeffect, but if the ischemic period is too prolonged, xanthine isable to accumulate and superoxide radicals are generatedduring the reperfusion, removing the beneficial NO andabolishing the preconditioning effect. Of course, adenosinecould directly induce other non-NO-mediated effects, and thesuperoxide anion generated by xanthine metabolism couldexert other deleterious effects in addition to the reaction withNO. It has been reported that adenosine reduces the neutro-phil-induced endothelial injury by A2-receptor-mediated inhi-bition of O2 generation and adherence.18 In addition, caution
FIG. 3. AST (A) and ALT (B) levels (U/L) in two different precondition-ing time groups (2, 20 minutes) and an additional Prec. 20 min 1 allopurinolgroup (n 5 6). Each group received doses of spermine NONOate from 0 to21 mg/kg body weight. Enzymatic levels after sustained I/R are also indicatedby the dotted line. *P , .05 vs. I/R; 1P , .05 vs. Prec without addition ofspermidine NONOate. Results are expressed as the means 6 SEM from sixrats.
TABLE 1. AST and ALT Levels in the Experimental Groups
AST (U/L) ALT (U/L)
Control 58.0 6 11.5 19.1 6 3.2I/R 188.2 6 15.1* 139.9 6 13.4*Prec. (10 min) 96.4 6 14.7† 68.8 6 11.3*†ADA 1 Prec. (10 min) 145.5 6 8.6*‡ 140.3 6 26.4*‡Xanthine 1 Prec. (10 min) 159.2 6 27.3*‡ 174.6 6 42.8*‡Prec. (5 min) 143.9 6 12.7* 183.2 6 41.5*Ado 1 Prec. (5 min) 79.0 6 11.8†‡ 54.8 6 11.5†‡Ado 1 xanthine 1 prec. (5 min) 354.1 6 54.7*†‡ 505.8 6 12.7*†‡
NOTE. Results are expressed as the mean 6 SEM from six rats.*P , .05 vs. control.†P , .05 vs. I/R.‡P , .05 vs. Prec.
772 PERALTA ET AL. HEPATOLOGY September 1998

must be applied before extrapolating these results to otherorgans.
In summary, this work indicates that the optimal ischemictime window to induce preconditioning in liver is defined bythe balance between the tissue concentrations of adenosineand xanthine. The lower limit is defined by the amount ofadenosine able to induce the NO generation, whereas theupper limit is defined by the concentration of xanthine thatprobably acts in removing the generated NO.
REFERENCES
1. Murry CE, Jennings RB, Reimer KA. Preconditioning with ischemia: adelay of lethal cell injury in ischemic myocardium. Circulation 1986;74:1124-1136.
2. Hotter G, Closa D, Prados M, Fernandez-Cruz L, Prats N, Gelpı E,Rosello-Catafau J. Intestinal preconditioning is mediated by a transientincrease in nitric oxide. Biochem Biophys Res Commun 1996;222:27-32.
3. Peralta C, Closa D, Hotter G, Gelpı E, Prats N, Rosello-Catafau J. Liverischemic preconditioning is mediated by the inhibitory action of nitricoxide on endothelin. Biochem Biophys Res Commun 1996;229:264-270.
4. Bilinska M, Maczewski M, Beresewicz A. Donors of nitric oxide mimiceffects of ischaemic preconditioning on reperfusion induced arrhythmiasin isolated rat heart. Mol Cell Biochem 1996;161:265-271.
5. Van Winkle DM, Downey JM, Thornton JD, Davis RF. Ischemicpreconditioning of myocardium: effect of adenosine. In: Maruyama ed.Recent Advances in Coronary Circulation. Tokyo: Springer-Verlag,1993:223-234.
6. Vegh A, Szekeres L, Parratt J. Preconditioning of the ischaemic myocar-dium: involvement of the L-arginine nitric oxide pathway. Br J Pharmacol1992;107:648-652.
7. Weselcouch EO, Baird AJ, Sleph P, Graver GJ. Inhibition of nitric oxidesynthesis does not affect ischemic preconditioning in isolated perfusedrat hearts. Am J Physiol 1995;268:H242-H249.
8. Peralta C, Hotter G, Closa D, Gelpı E, Bulbena O, Rosello-Catafau.Protective effect of preconditioning on the injury associated to hepaticischemia-reperfusion in the rat: role of nitric oxide and adenosine.HEPATOLOGY 1997;25:934-937.
9. Parrat J. Protection of the heart by ischaemic preconditioning: mecha-nisms and possibilities for pharmacological explotation. TIPS 1994;15:19-25.
10. Schoenberg MH, Beger HG. Oxygen radicals in intestinal ischemia andreperfusion. Chem Biol Interact 1990;76:141-161.
11. Hotter G, Closa D, Prats N, Pi F, Gelpı E, Rosello-Catafau J. Free radicalenhancement promotes leukocyte recruitment through a PAF and LTB4dependent mechanism. Free Radic Biol Med 1997;22:947-954.
12. Hull-Ride EA, Lewis WR, Veronee CD, Lowe JE. Simple step gradientelution of the major high-energy compounds and their catabolites incardiac muscle using high-performance liquid chromatography. J Chro-matogr Biomed App 1986;377:165-174.
13. Swenney MI. Neuroprotective effects of adenosine in cerebral ischemia:window of opportunity. Neurosci Biobehav Rev 1997;21:207-217.
14. Olsson RA. Changes in content of purine nucleoside in canine myocar-dium during coronary occlusion. Circ Res 1970;26:301-306.
15. Granger DN, Rutili G, McCord JM. Superoxide radicals in felineintestinal ischemia. Gastroenterology 1981;81:22-29.
16. Smolenski RT, Simmonds HA, Chambers DJ. Exogenous adenosine,supplied transiently during reperfusion, ameliorates depressed endog-enous adenosine production in the post-ischemic rat heart. J Mol Cardiol1997;29:333-346.
17. Freeman B. Free radical chemistry of nitric oxide. Looking at the darkside. Chest 1994;105:79S-84S.
18. Zhao Z, Sato H, Williams MW, Fernandez AZ, Vinten-Johansen J.Adenosine A2 receptor activation inhibits neutrophil mediated injury tocoronary endothelium. Am J Physiol 1996;271:H1456-H1464.
FIG. 4. Proposed mechanism of induction of ischemic preconditioning.The optimum ischemic time window to induce preconditioning is defined bythe tissue concentrations of adenosine and xanthine. These metabolitesaccumulate in tissue as a function of the time of ischemia. The lower limit isdefined by the minimum amount of adenosine able to induce the NOgeneration, whereas the upper limit is defined by the concentration ofxanthine that generates superoxide anion in amounts able to remove thegenerated NO.
HEPATOLOGY Vol. 28, No. 3, 1998 PERALTA ET AL. 773





























