Embed Size (px)
Citation preview

Accepted Manuscript
Neuroscience Forefront Review
Hippocampal estradiol synthesis and its significance for hippocampal synapticstability in male and female animals
Ricardo Vierk, Nicola Brandt, Gabriele M. Rune
PII: S0306-4522(14)00384-4DOI: http://dx.doi.org/10.1016/j.neuroscience.2014.05.003Reference: NSC 15403
To appear in: Neuroscience
Accepted Date: 4 May 2014
Please cite this article as: R. Vierk, N. Brandt, G.M. Rune, Hippocampal estradiol synthesis and its significance forhippocampal synaptic stability in male and female animals, Neuroscience (2014), doi: http://dx.doi.org/10.1016/j.neuroscience.2014.05.003
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, andreview of the resulting proof before it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.

1
Hippocampal estradiol synthesis and its significance for hippocampal synaptic stability in male and female animals Ricardo Vierk, Nicola Brandt, Gabriele M. Rune* Institute of Neuroanatomy, University Medical Center Hamburg-Eppendorf, Martinistr. 52, 20246 Hamburg, Germany Running title: Estrogen-induced synaptic plasticity *Corresponding author Dr. Gabriele M. Rune Institut für Neuroanatomie Universitätsklinikum Hamburg-Eppendorf Martinistr. 52 20246 Hamburg Germany Tel: +49-7410-53575 Fax: +49-7410-5496 e-mail: [email protected] Key words: aromatase, estrogen, sexual dimorphism, synaptic plasticity The study was supported by the Deutsche Forschungsgemeinschaft

2
Abstract Increasing evidence points to an essential role played by neuron-derived
neurosteroids, such as estrogen, on synaptic connectivity in the hippocampus.
Inhibition of local estradiol synthesis results in synapse loss specifically in
females, but not in males. Synapse loss in females, after inhibition of estradiol
synthesis in hippocampal neurons, appears to result from impairment of long-
term potentiation (LTP) and dephosphorylation of cofilin, and thereby the
destabilization of postsynaptic dendritic spines. Such clear-cut effects were
not seen in males. Cognitive deficits after inhibition of aromatase, the final
enzyme of estrogen synthesis, have been seen in women, but not in men.
Altogether, the data demonstrate distinct differences between genders in
neurosteroid-induced synaptic stability.
Introduction
Steroids are essential for brain function, from the early steps of differentiation to the
senescent brain during which they help to maintain neuronal performance and
protect against damage. The brain is equipped with all enzymes active in
steroidogenesis and is thus capable of synthesizing so-called “neurosteroids”, which
act locally to modify neural performance (Compagnone and Mellon, 2000, Shibuya et
al., 2003). Steroids, however, also enter the brain from the peripheral circulation.
Both pathways – paracrine and endocrine - are often interlinked. Thus, the precise
attribution of the individual routes to physiological processes is not easily addressed.
As to the origin of steroids in the CNS, the varying density of spines along
hippocampal dendrites of CA1 pyramidal neurons strongly supports the idea that
estrogen of ovarian origin regulates spinogenesis in the hippocampus (Woolley et al.,
1990; for review see Spencer et al., 2008). In addition, the removal of gonads has
been shown to result in reduced dendritic spine density in this brain area of males

3
and of females (Gould et al., 1990, Leranth et al., 2004). A rescue of spine loss after
gonadectomy could be achieved by injections of estradiol in females but not in males.
In gonadectomized males, injections of testosterone restored reduced spine density
after removal of the testes (Leranth et al., 2004).
Our data from recent years point to hippocampus-derived estradiol as the
main and primary player in estrogen-induced synaptic plasticity in females. We
inhibited estradiol synthesis in hippocampal cultures, thereby in the absence of any
other source of estradiol, either pharmacologically, by using inhibitors of aromatase,
the final enzyme of estradiol synthesis, or by knock-down of steroidogenic acute
regulatory protein (StAR), the rate-limiting step in estrogen synthesis. Inhibition of
aromatase in hippocampal slice cultures resulted in a significant reduction in the
number of spine synapses in the CA1 hippocampal region (Kretz et al., 2004,
Prange-Kiel et al., 2006). Similarly, knock-down of StAR, the protein that transports
cholesterol through the mitochondrial membranes, where steroidogenesis
commences, induced a downregulation of pre- and postsynaptic marker proteins
(Fester et al., 2009). Systemic treatment of female mice with letrozole also
significantly reduced spine synapse density and, most importantly, this effect was
also seen in ovariectomized animals, thus after having removed the main source of
estradiol in females (Zhou et al., 2010). The latter result strongly supports the notion
that in females, estradiol originating from neurons is essential for synaptogenesis,
rather than estrogen from peripheral sources (Prange-Kiel et al., 2013).
Nevertheless, we also found that estradiol synthesis in the female
hippocampus is linked to the hypothalamic-hypophyseal axis via the regulatory role of
Gonadotropin releasing hormone (GnRH) on hippocampal estrogen synthesis. We
showed that spine synapse density increases dose-dependently upon GnRH
stimulation in hippocampal cultures from female animals; this regulation functions via

4
stimulation of aromatase. Thus, in females, brain sex steroid levels should correlate
to circulating levels of sex hormones, which vary depending on the reproductive state
of the organism throughout life (Prange-Kiel et al., 2008, Prange-Kiel et al., 2013).
This was recently confirmed by Kato et al. (2013), showing that in fact estradiol
concentrations in hippocampal tissue correlate to estradiol concentrations in serum
during the estrous cycle. Consistently, it was previously shown that cyclic peripheral
concentrations of estradiol influence StAR expression in the brain (Meethal et al.,
2009), thus estradiol concentrations in serum influence steroidogenesis in the brain,
as the expression of StAR is the rate limiting step in estradiol synthesis.
Estrogen synthesis and spine synapse density in the hippocampus of males
and females
Although it has been known for years that the brain is equipped with all enzymes
active in steroidogenesis, our knowledge on differences in the capacity of
neurosteroid synthesis between genders is fragmentary. Naftolin (1971) was the first
to describe aromatase expression in the diencephalon in females. Aromatase is the
final enzyme of estradiol synthesis and catalyzes the conversion of testosterone to
estradiol. Ten years ago, we showed that the enzyme is actually functional. After a
couple of days in culture, hippocampal neurons had secreted estradiol into the
supernatant, which was determined by RIA (Prange-Kiel et al., 2003), which was
confirmed shortly thereafter by Hojo et al. (2004). As to differences between genders,
immunoreactivity of aromatase was seen in hippocampal neurons of male animals as
well as in those of female animals. Laser scanning microscopy and subsequent
image analysis did not reveal any difference in the expression of the protein between
genders (Fester et al., 2012). Although protein expression does not allow for any
conclusion on the activity of the enzyme, the amount of estradiol measured in the

5
supernatant of cultivated “female” and “male” neurons was similar as well (Fester et
al., 2012). In contrast to these in vitro findings we found clear-cut differences
between genders, when we measured the content of estradiol in male and female
hippocampal tissue by mass spectrometry. In hippocampal tissue of male animals
and ovariectomized animals, the amount of estradiol was extremely low and below
the sensitivity of our system. In contrast, in hippocampal tissue of females the
amount of estradiol was easily measured. The differences between males and
females in the amount of estradiol in hippocampal tissue corresponded to the
differences between genders in estradiol concentrations in plasma (Fester, 2012).
Robust functional sex differences have been identified in the hypothalamic
circuitry regulating reproductive function. Increasing levels of estradiol synthesized by
granulosa cells in the ovaries and released into the blood stream trigger the release
of gonadotropin releasing hormone (GnRH) from neurons scattered throughout the
hypothalamus, which in turn induce the mid-cycle surge in luteinizing hormone (LH)
that is essential for ovulation. In contrast, in male rats the pattern of LH release upon
GnRH stimulation is tonic and acyclic, thus leading to a steady state of testosterone,
which always exerts a negative feed back on GnRH release (for review see Gillies
and McArthur (2010). The lack of cyclic variations of GnRH/LH in males should result
in differences between genders. In fact, we found that systemic inhibition of
aromatase in mice had no effect on spine synapse density in males (Vierk et al.,
2012). In males, an increase in synapse density was even found in tendency. In this
context, it is of note that concentrations of testosterone, being the direct substrate for
the enzyme aromatase, should increase in tissue as well as in serum after inhibition
of aromatase activity. According to data by Leranth and coworkers (2004), showing
that spine loss in response to gonadectomy was rescued by testosterone, aromatase
inhibition in hippocampal neurons should actually result in an increase in synapse

6
density in males. The failure of letrozole to increase spine density in males may be
explained by insufficient activity of 5alpha reductase, which converts testosterone to
dehydrotestosterone, the active metabolite with a higher affinity to androgen
receptors. The role of hippocampus-derived testosterone in hippocampal cultures
that originate from male animals are currently under investigation in our laboratory.
Does spine synapse loss after inhibition of neuronal estradiol synthesis result
from the effects of estradiol on the spine cytoskeleton?
Both the function and stability of dendritic spines depend on an intact actin
cytoskeleton (Honkura et al., 2008, Dent et al., 2010, Hotulainen and Hoogenraad,
2010). Our results suggest that aromatase activity in hippocampal neurons is
required for the stabilization of postsynaptic spines. In stable spines, disassembly of
F-actin is prevented as soon as cofilin, an actin associated protein, becomes
phosphorylated. In hippocampal slice cultures of female animals, we found
pronounced dephosphorylation of cofilin, together with synapse loss, after 7 days of
letrozole treatment. In mice treated with letrozole, we also found enhanced
dephosphorylation of cofilin as early as after 6 hours of treatment. Accordingly,
intracellularly synthesized estradiol obviously induces phosphorylation of cofilin,
presumably via activating LIM-1 kinase, as shown by (Kramár et al., 2009b). Cofilin is
itself regulated; it becomes inactive when phosphorylated by LIM-1 kinase, thus
allowing polymerization to proceed (Bramham, 2008).
Our findings in hippocampal slice cultures confirm the studies by Kramár et al.
(Kramár et al., 2009a), insofar as we achieved opposite effects after inhibition of
estradiol synthesis in hippocampal neurons, thus, most importantly, in the absence of
any other source of estradiol. Kramár et al. (Kramár et al., 2009a) showed that

7
application of estradiol to acute hippocampal slices increases phalloidin-positive
punctae, indicating enhanced F-actin polymerization, and consequently increases
phosphorylation of cofilin via activation of LIM-1 kinase, which altogether stabilizes
spines. The time course of letrozole effects on spinogenesis and cofilin activity
suggests that spine synapse loss in response to letrozole is mediated by its effects
on the spine cytoskeleton. Dephosphorylation of cofilin due to aromatase inhibition in
female mice was seen as early as after 6 hours of treatment, whereas a significant
spine synapse loss was not seen before 48 hours of treatment had elapsed.
Obviously, destabilizing the spine cytoskeleton precedes and finally results in spine
synapse loss. This result is consistent with the finding that LIM-1 kinase knock-out
mice produce spines that lack actin and have small synapses (Meng et al., 2002).
The essential role of the actin cytoskeleton, which is regulated by numerous proteins,
in the plasticity and maintenance of spines, has been extensively shown (for review
see (Tada et al., 2007, Bramham, 2008, Kasai et al., 2010).
It is of note in this context, that in females, blocking cofilin phosphorylation by
letrozole is likely to prevent the formation of large actin-rich spines, or it reduces
spine head diameter of pre-existing spines, respectively. The proportion of large,
mushroom-shaped, actin-rich spines was particularly reduced, while thin spines were
less affected in hippocampal tissue of mice treated with letrozole; mushroom spines,
in turn, are considered to be the memory spines (Bourne and Harris, 2007, Tada et
al., 2007, Kasai et al., 2010).
Regarding the spine phenotype, the findings after application of estradiol to
acute slices, as reported in the literature, are discrepant. For instance, Kawato and
coworkers (2007) found an increase in the number of spines, but no alteration in
head diameter, thus estradiol only increases the number of thin, immature spines.
Using an almost identical experimental paradigm, Kramár et al. (2009a) found an

8
increase in the number of phalloidin-labeled spine-like profiles. The latter finding,
together with the findings by Kawato’s research group, would suggest that estradiol
primarily increases F-actin in thin spines, whereas mushroom spines are not affected,
although mushroom spines particularly depend on the actin cytoskeleton.
Furthermore, estradiol was found to rapidly enhance long-term depression (LTD)
(Mukai et al., 2007), which is known to cause shrinkage of spines (Nagerl et al., 2004,
Okamoto et al., 2004), whereas Kramár et al. (2009a) found LTP to be enhanced in a
non-genomic rapid manner, which stimulates spine formation (Yuste and Bonhoeffer,
2001, Matsuzaki et al., 2004). Along these lines, in ovariectomized mice, but not in
rats, estradiol injections failed to increase spine number; only the number of large
spines was increased (Li et al., 2004). We used rats and mice for our experiments
with consistent results. In addition, aromatase inhibition in female mice, even in
ovariectomized animals, significantly reduces the number of spine synapses (Zhou et
al., 2010). We found a clear time course of events that started 6 hours after
aromatase inhibition was induced, either in female mice and/or in “female” rat
hippocampal slice cultures, and which reached its maximum after 7 days.
In summary, application of estradiol at non-physiological concentrations, such
as 1 nM estradiol in male rats (Mukai et al., 2007, Kramár et al., 2009a), and
inhibition of local estradiol synthesis do not automatically cause opposite effects.
Since we have frequently shown that hippocampus-derived estradiol, rather than
estradiol originating from other sources, influences synaptic plasticity, namely due to
the regulatory role of GnRH on hippocampal estradiol synthesis (Kretz et al., 2004,
Zhou et al., 2007, Prange-Kiel et al., 2008, Prange-Kiel et al., 2009, Zhou et al.,
2010), we presume that inhibition of local estradiol synthesis is the more adequate
approach to study the role of estradiol in synaptic plasticity, at least in females.

9
Despite the intriguing idea that the influence of estradiol on spine formation in
the hippocampus is mediated via the effect of estradiol on the spine cytoskeleton, it
remains an open question as to why dephosphorylation of cofilin in response to
letrozole does not result in spine synapse loss in males. Although the time course
and the degree of downregulation of p-cofilin were similar in male and female mice, a
similar decrease in the synapse density seen in females was not observed in males.
Obviously, another additional mechanism protects “male” synapses from regression.
Long-Term Potentiation, cofilin, and estradiol
Our findings on spine synapse density after inhibition of neuronal estradiol synthesis
and the question as to whether this letrozole-induced synapse loss is of any
biological significance prompted us to study the effects of letrozole on long-term
potentiation (LTP) as a cellular, electrophysiological parameter for memory. In
addition, the questions as to whether the paradigm that “LTP induces spines” (Engert
and Bonhoeffer, 1999, Yuste and Bonhoeffer, 2001, 2004) is also valid vice versa,
and the paradigm “LTP is paralleled by increased phosphorylation of cofilin”
(Fukazawa et al., 2003), were addressed in the study by Vierk et al. (Vierk et al.,
2012).
It has frequently been shown that estradiol enhances LTP in the hippocampus
(for review see Spencer et al., 2008). The effects of estradiol on spine density and
LTP, however, were shown in most studies either by using ovariectomized animals,
which were treated systemically with estradiol (Gould et al., 1990, Warren et al., 1995,
Cordoba Montoya and Carrer, 1997, Smith and McMahon, 2005, 2006, Kramár et al.,
2009a), or estradiol was applied to acute slices of mostly male rats (Foy et al., 1999,
Ito et al., 1999, Mukai et al., 2007, Kramár et al., 2009a), or by using hippocampal
slice cultures indifferently prepared from male and neonatal females (Murphy and

10
Segal, 1996, Kretz et al., 2004, Mendez et al., 2011). The vast majority of results
from these studies show that exogenous application of estradiol is primarily effective
after having removed estradiol from the environment before the treatment, i.e. after
ovariectomy, and by using an in vitro experimental approach. These experimental
approaches leave the question open as to whether in females, estradiol actually
causes major alterations of non-affected pre-existing networks (Mendez et al., 2011)
or simply restores effects after estradiol depletion. In males, the biological
significance of applying estradiol at highly supraphysiological doses is a further
obvious question.
We studied LTP and synaptic density in acute slices of animals treated by
daily injections of letrozole over 7 days, and hippocampal slice cultures treated with
letrozole for 7 days. In both cases the effect was controlled by measurements of
estradiol concentrations in serum and in the supernatant resp. After 7 days of
treatment LTP could no longer be induced in females and this was paralleled by
significant synapse loss; in males, however, LTP was reduced by 20% with no loss of
synapses after treatment (Fig. 1).
Regarding time-dependency, impairment of LTP was found as early as 6
hours after a single injection of letrozole. Dephosphorylation of cofilin, was also
already apparent after 6 hours had elapsed. Spine synapse loss, however, was not
seen prior to 48 hours after the last injection, which made us assume that synapse
loss results from impaired LTP (Fig. 1;Vierk et al., 2012)) according to the paradigm
that LTP induces spines (Engert and Bonhoeffer, 1999).
In addition, in our hands effects were not seen when we applied letrozole
directly to acute slices for 3–4 hours after stable recording of LTPs for 90 min
(unpublished observations). This finding is consistent with findings by Grassi and
coworkers (Grassi et al., 2011). They found, however, that the potentiation after high

11
frequency stimulation but not the maintenance of LTP was markedly reduced in the
presence of letrozole in males compared to that induced under control conditions.
This effect was seen as early as after 1 min and progressed further up to 40 min. An
effect after 1 min and 10 min would argue for a so-called rapid membrane-bound
estrogenic effect. Given this background, it is interesting that the effects of letrozole
could be mimicked by using ICI 182780. This compound has been used to
discriminate between rapid and genomic effects (Kramár et al., 2009a). Kramár et al.
describe that the rapid effects of estradiol on LTP were not affected by ICI 182780. In
addition, ICI 182780, according to the suppliers, upregulates membrane bound
receptors, which should support rapid estradiol signalling. Furthermore, it would be
interesting to know whether a substantial reduction in tissue estradiol levels is
already achieved after 1 min of letrozole treatment, which could account for the
markedly reduced synaptic potentiation. In a recent study, Tanaka and Sokabe
(Tanaka and Sokabe, 2012) provided evidence that continuous synthesis of
neurosteroids is required for normal transmission in the hippocampus. They used
aminoglutethimide (AG), which blocks steroid biosynthesis from the beginning.
Application of AG to acute slices of male animals decreased fEPSPs by 20-30% in
the dentate gyrus, an effect that was mimicked by letrozole. Similar to our findings in
the hippocampus of male animals treated with letrozole, and after direct application
as shown by Tanaka and Sokabe (Tanaka and Sokabe, 2012) LTP was also
reduced by 20% in response to letrozole treatment in acute slices of the male medial
rat vestibular nuclei (Grassi et al., 2009). Whether 20% reduction in LTP is of any
functional significance in terms of behavioural deficits remains an open question.
Our results are consistent with studies showing that estradiol induces the
magnitude of LTP at CA3-CA1 Schaffer collaterals in the hippocampus (Foy et al.,
1999, Kim et al., 2006). The results further confirm the relationship of spines and

12
LTP: impairment of LTP in female letrozole-treated mice precedes spine synapse
loss, while in males no major effect on LTP was seen, paralleled by unchanged spine
synapse number. LTP and phosphorylation of cofilin appeared merely to be
phenomena independent from one another and are not necessarily linked. When LTP
is maximally downregulated after letrozole treatment of female mice, p-cofilin
expression is upregulated (Vierk et al., 2012). Along these lines, p-cofilin is
downregulated, together with minor changes in LTP in male hippocampi, after
treatment with letrozole.
Cognitive effects of aromatase inhibition in females and males
Regardless of discrepancies and inconsistencies of the data in the literature, overall,
the effects of estradiol on synapse density and LTP suggest that estrogen is involved
in the regulation of cognitive function.
Aromatase inhibitors, such as letrozole, are commonly used in the therapy of
postmenopausal women suffering from hormone-dependent breast cancer (Geisler et
al., 2002, Puddefoot et al., 2002). Clinical pilot studies have demonstrated that
aromatase-inhibitors affect cognition and memory deficits in women treated with
these inhibitors (Shilling et al., 2001, Dowsett et al., 2005, Shilling et al., 2005). Due
to the small charge of the molecule, letrozole is easily transported across the blood-
brain barrier after systemic application and exerts an inhibitory influence on
hippocampal estrogen synthesis, as it does in other regions of the body (Zhou et al.,
2010). Impaired LTP (as a cellular correlate of memory), together with spine synapse
loss, both parallel dephosphorylation of cofilin in spines after estradiol depletion. This
could be the underlying mechanisms for memory deficits in women treated with
aromatase inhibitors for therapeutical reasons (Lamprecht and LeDoux, 2004). This

13
appears to be of greater importance, as letrozole also induces synapse loss in
ovariectomized animals (Zhou et al., 2010). Ovariectomy is frequently regarded as an
experimental paradigm for menopause in women. Menopausal women, as compared
to women during their reproductive phase, are preferentially treated with aromatase
inhibitors, since this does not interfere with their reproductive function.
In men, it has been shown that testosterone-induced improvement of verbal memory
in older men depends on aromatization of testosterone to estradiol, while the
improvement of spatial memory in response to testosterone treatment was
independent of conversion of estradiol (Cherrier et al., 2005). Prepubertal boys
suffering from growth disorders are increasingly being treated with aromatase
inhibitors. Aromatase inhibition is considered to be a potential new treatment modality
for idiopathic short stature. In tests for cognitive impairment, blockade of estrogen
synthesis with an aromatase inhibitor did influence cognitive performance in boys
who had been treated with letrozole for two years (Hero et al., 2010).
Studies that focus on synaptic plasticity and cognitive abilities related to sexual
hormones primarily target the effects of testosterone treatment in men and in male
animals (Bimonte-Nelson et al., 2003, Sherwin, 2003, Frye et al., 2004, Gibbs, 2005,
Romeo et al., 2005, Martin et al., 2008, Cost et al., 2012) and of estrogen treatment
in women and in female animals (Gibbs, 2005, Gasbarri et al., 2012, Inagaki et al.,
2012, Nissen et al., 2012, Ryan et al., 2012, Sherwin, 2012, Stelly et al., 2012,
Velazquez-Zamora et al., 2012, da Rocha et al., 2013, McClure et al., 2013). Studies
on the role of estrogen in male animals are more frequent than studies on the effect
of testosterone in female animals (Benice and Raber, 2009, Cost et al., 2012).
Data of estrogen-regulated cognitive abilities in females originate
predominantly from studies on estrogen replacement therapy of pre- and
postmenopausal women. Many excellent reports point to a beneficial function of

14
estradiol with respect to memory (Sherwin, 1988, Phillips and Sherwin, 1992, Duka et
al., 2000, Maki et al., 2001) and risk of dementia (Yaffe et al., 1998, Nelson et al.,
2002). Other studies failed to confirm these results (Espeland et al., 2004). They
found that hormone therapy had an adverse effect on cognition for women over the
age of 65. Similar discrepancies are also found in men and male animals with respect
to the influence of testosterone on cognition. Testosterone was postulated to
decrease CA plasticity in vivo in gonadectomized male rats (Harley et al., 2000),
while others report that testosterone replacement reverses altered synaptic
transmission after gonadectomy in males (Sakata et al., 2000). Gibbs’s (Gibbs, 2005)
data revealed a dissociation between the effects of testosterone and estradiol in
cognitive performance in male rats. Female rats outperform males on an object
recognition task when circulating levels of ovarian steroids are elevated (Cost et al.,
2012). In males, in turn, testicular androgens are important for maximal levels of
working memory (McConnell et al., 2012).
In spite of the tremendous amount of data on the role of sex steroid hormones in
cognition, the inconsistencies in the literature, which may be due to different
methodological approaches, point to the necessity for further studies in order to
understand the effects of sex steroid hormones. This could lead to new therapeutical
strategies for age-related memory decline (for review see Sherwin and Henry, Maki
and Sundermann, Sherwin and Frick (Sherwin and Henry, 2008, Frick, 2009, Maki
and Sundermann, 2009, Frick, 2012, Sherwin, 2012)). Since GnRH regulates
estradiol synthesis in the hippocampus (Prange-Kiel et al., 2008), ovariectomy results
in loss of aromatase activity in the hippocampus (Fester et al., 2012), and estradiol
content in the hippocampus varies with the estrus cycle (Kato et al., 2013), synthesis
in the ovaries is synchronized with hippocampal estradiol synthesis. Thus, one would
assume that the aging hippocampal neuron synthesizes less estradiol than the young

15
neuron, which, however, has not yet been shown. If this turns out to be true, attempts
to understand, and as a consequence to manipulate the local, intraneuronal
regulation of aromatase could deliver another therapeutical approach. For instance,
aromatase activity in neurons was shown to be regulated by neuronal activity (Hojo et
al., 2004) and becomes inactive by phosphorylation via calcium transients and
calcium-dependent kinases (Balthazart et al., 2006, Fester et al., 2012). Local
stimulation of aromatase activity in neurons would avoid undesired side effects of
estrogen replacement therapy.
Conclusion
Hippocampal neurons synthesize estradiol, which maintains LTP and synapses in
females but not in males. In females, inhibition of estradiol synthesis results in
impairment of LTP, dephosphorylation of cofilin and final synapse loss. These effects
were not seen in males. The essential role of local estrogen on the stability and
maintenance of connectivity in the hippocampus is consistent with age-related
cognitive decline in women. In male animals the regulation of synaptic stability by
local sexual steroids remains to be clarified.

16
References
Balthazart J, Baillien M, Ball GF (2006).Rapid control of brain aromatase activity by
glutamatergic inputs. Endocrinology 147:359-366.
Benice TS, Raber J (2009).Testosterone and dihydrotestosterone differentially improve
cognition in aged female mice. Learn Mem 16:479-485.
Bimonte-Nelson HA, Singleton RS, Nelson ME, Eckman CB, Barber J, Scott TY,
Granholm AC (2003).Testosterone, but not nonaromatizable dihydrotestosterone,
improves working memory and alters nerve growth factor levels in aged male
rats. Exp Neurol 181:301-312.
Bramham CR (2008).Local protein synthesis, actin dynamics, and LTP consolidation. Curr Opin Neurobiol 18:524-531.
Cordoba Montoya DA, Carrer HF (1997).Estrogen facilitates induction of long term potentiation in the hippocampus of awake rats. Brain Res 778:430-438.
Cost KT, Williams-Yee ZN, Fustok JN, Dohanich GP (2012).Sex differences in object-in-place memory of adult rats. Behav Neurosci 126:457-464.
da Rocha JT, Sampaio TB, Santos Neto JS, Nogueira CW, Zeni G (2013).Cognitive effects of diphenyl diselenide and estradiol treatments in ovariectomized mice.
Neurobiology of Learning and Memory 99:17-24.
Dent EW, Merriam EB, Hu X (2010).The dynamic cytoskeleton: backbone of dendritic
spine plasticity. Curr Opin Neurobiol.
Duka T, Tasker R, McGowan JF (2000).The effects of 3-week estrogen hormone
replacement on cognition in elderly healthy females. Psychopharmacology (Berl)
149:129-139.
Engert F, Bonhoeffer T (1999).Dendritic spine changes associated with hippocampal
long-term synaptic plasticity. Nature 399:66-70.
Espeland MA, Rapp SR, Shumaker SA, Brunner R, Manson JE, Sherwin BB, Hsia J,
Margolis KL, Hogan PE, Wallace R, Dailey M, Freeman R, Hays J
(2004).Conjugated equine estrogens and global cognitive function in
postmenopausal women: Women's Health Initiative Memory Study. JAMA
291:2959-2968. Fester L, Prange-Kiel J, Zhou L, Blittersdorf BV, Bohm J, Jarry H, Schumacher M,
Rune GM (2012).Estrogen-regulated synaptogenesis in the hippocampus: sexual dimorphism in vivo but not in vitro. J Steroid Biochem Mol Biol 131:24-29.
Fester L, Prange-Kiel, J. Zhou, L., von Blittersdorf, B., Böhm, J., Jarry, H., Schumacher, M., Rune, GM. (2012).Estrogen-regulated synaptogenesis: sexual dimorphism in
vivo but not in vitro. . J Steroid Biochem Mol Biol 2012 (in press)
Fester L, Zhou L, Voets C, Ossig C, Disteldorf E, Blaute F, Prange-Kiel J, Dudzinski D,
Jarry H, Rune GM (2009).The opposing roles of estradiol on synaptic protein
expression in hippocampal cultures. Psychoneuroendocrinology 34 Suppl 1:S123-129.
Foy MR, Xu J, Xie X, Brinton RD, Thompson RF, Berger TW (1999).17beta-estradiol enhances NMDA receptor-mediated EPSPs and long-term potentiation. J
Neurophysiol 81:925-929. Frick KM (2009).Estrogens and age-related memory decline in rodents: what have we
learned and where do we go from here? Horm Behav 55:2-23.

17
Frick KM (2012).Building a better hormone therapy? How understanding the rapid
effects of sex steroid hormones could lead to new therapeutics for age-related
memory decline. Behav Neurosci 126:29-53.
Frye CA, Edinger KL, Seliga AM, Wawrzycki JM (2004).5alpha-reduced androgens
may have actions in the hippocampus to enhance cognitive performance of male
rats. Psychoneuroendocrinology 29:1019-1027.
Fukazawa Y, Saitoh Y, Ozawa F, Ohta Y, Mizuno K, Inokuchi K (2003).Hippocampal
LTP is accompanied by enhanced F-actin content within the dendritic spine that
is essential for late LTP maintenance in vivo. Neuron 38:447-460.
Gasbarri A, Tavares MC, Rodrigues RC, Tomaz C, Pom pili A (2012).Estrogen,
cognitive functions and emotion: an overview on humans, non-human primates
and rodents in reproductive years. Rev Neurosci 23:587-606. Geisler J, Haynes B, Anker G, Dowsett M, Lonning PE (2002).Influence of letrozole and
anastrozole on total body aromatization and plasma estrogen levels in postmenopausal breast cancer patients evaluated in a randomized, cross-over
study. J Clin Oncol 20:751-757. Gibbs RB (2005).Testosterone and estradiol produce different effects on cognitive
performance in male rats. Horm Behav 48:268-277. Gillies GE, McArthur S (2010).Estrogen actions in the brain and the basis for
differential action in men and women: a case for sex-specific medicines.
Pharmacol Rev 62:155-198.
Gould E, Woolley CS, Frankfurt M, McEwen BS (1990).Gonadal steroids regulate
dendritic spine density in hippocampal pyramidal cells in adulthood. J Neurosci
10:1286-1291.
Grassi S, Frondaroli A, Dieni C, Scarduzio M, Pettorossi VE (2009).Long-term
potentiation in the rat medial vestibular nuclei depends on locally synthesized
17beta-estradiol. J Neurosci 29:10779-10783.
Grassi S, Tozzi A, Costa C, Tantucci M, Colcelli E, ScarduzioM, Calabresi P, and
Pettorossi VE (2011). Neural 17beta-estradiol facilitates long-term potentiation in
the hippocampal CA1 region. Neuroscience 192:67-73.
Harley CW, Malsbury CW, Squires A, Brown RA (2000).Testosterone decreases CA1
plasticity in vivo in gonadectomized male rats. Hippocampus 10:693-697. Hero M, Maury S, Luotoniemi E, Service E, Dunkel L (2010).Cognitive effects of
aromatase inhibitor therapy in peripubertal boys. Eur J Endocrinol 163:149-155. Hojo Y, Hattori T, Enami T, Furukawa A, Suzuki K, Ishii HT, Mukai H, Morrison JH,
Janssen WGM, Kominami S, Harada N, Kimoto T, Kawato S (2004).Adult male rat hippocampus synthesizes estradiol from pregnenolone by cytochromes
P45017 alpha and P450 aromatase localized in neurons. Proc Natl Acad Sci U S A 101:865-870.
Honkura N, Matsuzaki M, Noguchi J, Ellis-Davies GC, Kasai H (2008).The subspine
organization of actin fibers regulates the structure and plasticity of dendritic
spines. Neuron 57:719-729.
Hotulainen P, Hoogenraad CC (2010).Actin in dendritic spines: connecting dynamics to function. J Cell Biol 189:619-629.
Inagaki T, Frankfurt M, Luine V (2012).Estrogen-induced memory enhancements are blocked by acute bisphenol A in adult female rats: role of dendritic spines.
Endocrinology 153:3357-3367. Ito K, Skinkle KL, Hicks TP (1999).Age-dependent, steroid-specific effects of oestrogen
on long-term potentiation in rat hippocampal slices. J Physiol 515 ( Pt 1):209-220.

18
Kasai H, Fukuda M, Watanabe S, Hayashi-Takagi A, Noguchi J (2010).Structural
dynamics of dendritic spines in memory and cognition. Trends Neurosci 33:121-
129.
Kato A, Hojo Y, Higo S, Komatsuzaki Y, Murakami G, Yoshino H, Uebayashi M,
Kawato S (2013).Female hippocampal estrogens have a significant correlation
with cyclic fluctuation of hippocampal spines. Front Neural Circuits 7:149.
Kim MT, Soussou W, Gholmieh G, Ahuja A, Tanguay A, Berger TW, Brinton RD
(2006).17beta-Estradiol potentiates field excitatory postsynaptic potentials within
each subfield of the hippocampus with greatest potentiation of the
associational/commissural afferents of CA3. Neuroscience 141:391-406.
Kramár EA, Chen LY, Brandon NJ, Rex CS, Liu F, Gall CM, Lynch G
(2009a).Cytoskeletal changes underlie estrogen's acute effects on synaptic transmission and plasticity. J Neurosci 29:12982-12993.
Kramár EA, Chen LY, Rex CS, Gall CM, Lynch G (2009b).Estrogen's Place in the Family of Synaptic Modulators. Mol Cell Pharmacol 1:258-262.
Kretz O, Fester L, Wehrenberg U, Zhou L, Brauckmann S, Zhao S, Prange-Kiel J, Naumann T, Jarry H, Frotscher M, Rune GM (2004).Hippocampal synapses
depend on hippocampal estrogen synthesis. J Neurosci 24:5913-5921. Lamprecht R, LeDoux J (2004).Structural plasticity and memory. Nat Rev Neurosci
5:45-54.
Leranth C, Hajszan T, MacLusky NJ (2004).Androgens increase spine synapse density
in the CA1 hippocampal subfield of ovariectomized female rats. J Neurosci
24:495-499.
Maki PM, Sundermann E (2009).Hormone therapy and cognitive function. Hum Reprod
Update 15:667-681.
Maki PM, Zonderman AB, Resnick SM (2001).Enhanced verbal memory in
nondemented elderly women receiving hormone-replacement therapy. Am J
Psychiatry 158:227-233.
Martin DM, Wittert G, Burns NR, McPherson J (2008).Endogenous testosterone levels,
mental rotation performance, and constituent abilities in middle-to-older aged
men. Horm Behav 53:431-441.
McClure RE, Barha CK, Galea LA (2013).17beta-Estradiol, but not estrone, increases the survival and activation of new neurons in the hippocampus in response to
spatial memory in adult female rats. Horm Behav 63:144-157. McConnell SE, Alla J, Wheat E, Romeo RD, McEwen B, Thornton JE (2012).The role of
testicular hormones and luteinizing hormone in spatial memory in adult male rats. Horm Behav 61:479-486.
Meethal SV, Liu T, Chan HW, Ginsburg E, Wilson AC, Gray DN, Bowen RL, Vonderhaar BK, Atwood CS (2009).Identification of a regulatory loop for the
synthesis of neurosteroids: a steroidogenic acute regulatory protein-dependent
mechanism involving hypothalamic-pituitary-gonadal axis receptors. J
Neurochem 110:1014-1027.
Mendez P, Garcia-Segura LM, Muller D (2011).Estradiol promotes spine growth and synapse formation without affecting pre-established networks. Hippocampus
21:1263-1267. Meng Y, Zhang Y, Tregoubov V, Janus C, Cruz L, Jackson M, Lu WY, MacDonald JF,
Wang JY, Falls DL, Jia Z (2002).Abnormal spine morphology and enhanced LTP in LIMK-1 knockout mice. Neuron 35:121-133.
Mukai H, Tsurugizawa T, Murakami G, Kominami S, Ishii H, Ogiue-Ikeda M, Takata N, Tanabe N, Furukawa A, Hojo Y, Ooishi Y, Morrison JH, Janssen WG, Rose
JA, Chambon P, Kato S, Izumi S, Yamazaki T, Kimoto T, Kawato S

19
(2007).Rapid modulation of long-term depression and spinogenesis via synaptic
estrogen receptors in hippocampal principal neurons. J Neurochem 100:950-967.
Naftolin F, Ryan KJ, Petro Z (1971).Aromatization of androstenedione by the
diencephalon. J Clin Endocrinol Metab 33:368-370.
Nelson HD, Humphrey LL, Nygren P, Teutsch SM, Allan JD (2002).Postmenopausal
hormone replacement therapy: scientific review. JAMA 288:872-881.
Nissen I, Estrada FS, Nava-Kopp AT, Irles C, de-la-Pena-Diaz A, Fernandez GJ,
Govezensky T, Zhang L (2012).Prolame ameliorates anxiety and spatial learning
and memory impairment induced by ovariectomy in rats. Physiol Behav 106:278-
284.
Phillips SM, Sherwin BB (1992).Effects of estrogen on memory function in surgically
menopausal women. Psychoneuroendocrinology 17:485-495. Prange-Kiel J, Fester L, Zhou L, Lauke H, Carretero J, Rune GM (2006).Inhibition of
hippocampal estrogen synthesis causes region-specific downregulation of synaptic protein expression in hippocampal neurons. Hippocampus 16:464-471.
Prange-Kiel J, Jarry H, Schoen M, Kohlmann P, Lohse C, Zhou L, Rune GM (2008).Gonadotropin-releasing hormone regulates spine density via its regulatory
role in hippocampal estrogen synthesis. J Cell Biol 180:417-426. Prange-Kiel J, Schmutterer T, Fester L, Zhou L, Imholz P, Brandt N, Vierk R, Jarry H,
Rune GM (2013).Endocrine regulation of estrogen synthesis in the hippocampus?
Prog Histochem Cytochem 48:49-64.
Prange-Kiel J, Wehrenberg U, Jarry H, Rune GM (2003).Para/autocrine regulation of
estrogen receptors in hippocampal neurons. Hippocampus 13:226-234.
Puddefoot JR, Barker S, Glover HR, Malouitre SD, Vinson GP (2002).Non-competitive
steroid inhibition of oestrogen receptor functions. Int J Cancer 101:17-22.
Romeo RD, McCarthy JB, Wang A, Milner TA, McEwen BS (2005).Sex differences in
hippocampal estradiol-induced N-methyl-D-aspartic acid binding and
ultrastructural localization of estrogen receptor-alpha. Neuroendocrinology
81:391-399.
Ryan J, Stanczyk FZ, Dennerstein L, Mack WJ, Clark MS, Szoeke C, Kildea D,
Henderson VW (2012).Hormone levels and cognitive function in postmenopausal
midlife women. Neurobiol Aging 33:1138-1147. Sakata K, Tokue A, Kawai N (2000).Altered synaptic transmission in the hippocampus
of the castrated male mouse is reversed by testosterone replacement. J Urol 163:1333-1338.
Sherwin BB (1988).Estrogen and/or androgen replacement therapy and cognitive functioning in surgically menopausal women. Psychoneuroendocrinology 13:345-
357. Sherwin BB (2003).Steroid hormones and cognitive functioning in aging men: a mini-
review. J Mol Neurosci 20:385-393.
Sherwin BB (2012).Estrogen and cognitive functioning in women: lessons we have
learned. Behav Neurosci 126:123-127.
Sherwin BB, Henry JF (2008).Brain aging modulates the neuroprotective effects of estrogen on selective aspects of cognition in women: a critical review. Front
Neuroendocrinol 29:88-113. Smith CC, McMahon LL (2005).Estrogen-induced increase in the magnitude of long-
term potentiation occurs only when the ratio of NMDA transmission to AMPA transmission is increased. J Neurosci 25:7780-7791.
Smith CC, McMahon LL (2006).Estradiol-induced increase in the magnitude of long-term potentiation is prevented by blocking NR2B-containing receptors. J
Neurosci 26:8517-8522.

20
Spencer JL, Waters EM, Romeo RD, Wood GE, Milner TA, McEwen BS
(2008).Uncovering the mechanisms of estrogen effects on hippocampal function.
Front Neuroendocrinol 29:219-237.
Stelly CE, Cronin J, Daniel JM, Schrader LA (2012).Long-term oestradiol treatment
enhances hippocampal synaptic plasticity that is dependent on muscarinic
acetylcholine receptors in ovariectomised female rats. J Neuroendocrinol 24:887-
896.
Tada T, Simonetta A, Batterton M, Kinoshita M, Edbauer D, Sheng M (2007).Role of
Septin cytoskeleton in spine morphogenesis and dendrite development in neurons.
Curr Biol 17:1752-1758.
Tanaka M, Sokabe M (2012).Continuous de novo synthesis of neurosteroids is required
for normal synaptic transmission and plasticity in the dentate gyrus of the rat hippocampus. Neuropharmacology 62:2373-2387.
Velazquez-Zamora DA, Garcia-Segura LM, Gonzalez-Burgos I (2012).Effects of selective estrogen receptor modulators on allocentric working memory
performance and on dendritic spines in medial prefrontal cortex pyramidal neurons of ovariectomized rats. Horm Behav 61:512-517.
Vierk R, Glassmeier G, Zhou L, Brandt N, Fester L, Dudzinski D, Wilkars W, Bender RA, Lewerenz M, Gloger S, Graser L, Schwarz J, Rune GM (2012).Aromatase
inhibition abolishes LTP generation in female but not in male mice. J Neurosci
32:8116-8126.
Warren SG, Humphreys AG, Juraska JM, Greenough WT (1995).LTP varies across the
estrous cycle: enhanced synaptic plasticity in proestrus rats. Brain Res 703:26-30.
Woolley CS, Gould E, Frankfurt M, McEwen BS (1990).Naturally-Occurring
Fluctuation in Dendritic Spine Density on Adult Hippocampal Pyramidal
Neurons. J Neurosci 10:4035-4039.
Yaffe K, Sawaya G, Lieberburg I, Grady D (1998).Estrogen therapy in postmenopausal
women - Effects on cognitive function and dementia. JAMA-J Am Med Assoc
279:688-695.
Zhou L, Fester L, von Blittersdorff B, Hassu B, Nogens H, Prange-Kiel J, Jarry H,
Wegscheider K, Rune GM (2010).Aromatase inhibitors induce spine synapse loss
in the hippocampus of ovariectomized mice. Endocrinology 151:1153-1160.

21
Legend to the figure
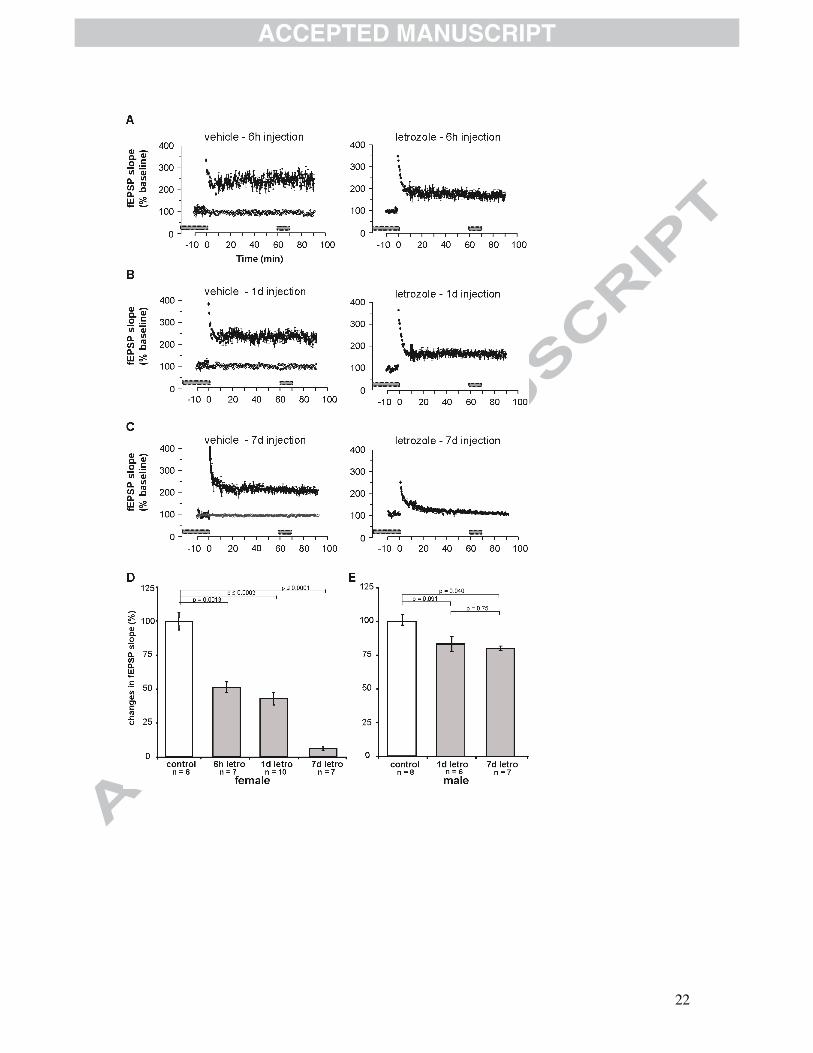
Fig. 1 LTP in acute hippocampal slices of mice treated with letrozole.
Results of female animals treated with vehicle and animals treated with letrozole. (A)
In slices of animals treated with vehicle (6 hrs), the average course of the fEPSP
slopes had significantly increased after 60 min to 247%±9% (n=8), whereas in
animals treated with letrozole (6 hrs) the slopes of fEPSPs increased only to
175%±4% (n=7). (B) After 1 day of vehicle treatment the slopes of fEPSPs increased
to 226%±8% (n=8), and after letrozole treatment for 1 day the fEPSP slopes had
changed only to 154%±5% (n=7). (C) After 7 days of vehicle injection, the average
slope of the fEPSPs increased to 213%±3% (n=6); in contrast, letrozole injections
after 7 days resulted in virtually unchanged fEPSP slopes 60 min after TBS,
107%±2% (n=7). Mean±SEM; time point 0 min represents TBS-stimulation; fEPSP
slopes before and 60-70 min after TBS were quantified and are indicated by grey
bars. (D) Quantitative examination of changes in the average course of the fEPSP
after letrozole treatment for 6 hrs, 1 day and 7 days illustrated a highly significant
reduction in LTP by 49%±4%, 57%±5% and 94%±1% respectively in females. (d:
day; n=number of acute slices of 3 animals; mean±SEM) (E) Quantitative
examination of changes in the average course of fEPSP slopes after letrozole
treatment in males. Letrozole treatment of male animals resulted in reduced LTP by
17%±5% after 1 day and 20%±2% after 7 days of treatment. (d: day; mean±SEM) (d:
day; n=number of acute slices of 3 animals; mean±SEM) (Vierk et al., 2012)
22

23
Highlights aromatase; estrogen; sexual dimorphism; synaptic plasticity





























