Embed Size (px)
Citation preview

1991) 767
ELECTROPHORETIC CHARACTERIZATION OF NORTH AMERICAN POTATO CULTIVARSl
D. S. Douches and K. Ludlam2
Abstract
A total of 112 named cultivars and· advanced breeding lines were assayed through horizontal starch electrophoresis to test the utility of isozyme loci as a means to objectively identify potato cultivars. Both leaf and tuber tissue were sampled in thirteen enzyme systems. Two buffer systems (Histidine-citrate, pH 5.7 and Lithium-borate, pH 8.3) were used to resolve 16 isozyme loci, of which 14 were scorable without progeny testing. A total of 43 scorable allozymes were detected. All cultivars or advanced breeding lines that were the result of hybridization were discriminated by their sum electrophoretic pattern, whereas, sports (i.e., Russet Burbank vs. Burbank) or line selections (i.e., Norgold Russet "M1" vs. "M2") have patterns that are identical to the original mother clone. The allelic diversity within and among cultivars indicate that horizontal starch electrophoresis offers an objective means to discriminate sexually-derived potato cultivars.
Compendio
Se analizaron un total de 112 cultivares y selecciones avanzadas a traves de electroforesis horizontal en almidon para probar la utilidad de loci de isoenzimas para la identificaci6n objetiva de cultivares de papa. Se utilizaron dos sistemas buffer (Histidina-citrato, pH 5.7 y Litio-borato, pH 8.3) para separar 16 loci de isoenzimas, de los cuales 14 pudieron ser calificados sin pruebas de progenie. Un total de 43 aloenzimas pudieron ser caracterizadas. Todos los cultivares 0 selecciones avanzadas productos de hibridaci6n fueron identificados por su patron electroforetico aditivo, mientras que los productos de mutaci6n (por ejemplo Russet Burbank vs Burbank), olfneas selectas (por ejemplo Norgold Russet "M1" vs "M2") mostraron patrones identicos al cIon materno original. La diversidad alelica dentro y entre cultivares indica que la tecnica de electroforesis horizontal en almid6n es un metodo objectivo para distinguir cultivares de papa originados sexualmente.
Introduction
Through the work of federal, state, and private breeders, over 250 potato cultivars (Solanum tuberosum ssp. tuberosum L.) have been named and
lJournal Article No. 000122. 2Professor and Graduate Student, respectively. Department of Crop and Soil Sciences, Michigan State University, East Lansing, Michigan 48224. Accepted for publication February 28, 1991. ADDITIONAL KEY WORDS: Solanum tuberosum L., isozyme, variety identification, fingerprint.

768 AMERICAN POTATO JOURNAL (Vol. 68
publicly released in North America (5). Historically, identification of these cultivars has been based upon key morphological markers and good record keeping. Primary grouping of potato cultivars is based on tuber type (i.e., skin, shape, and flesh). AdditionaJ characters such as sprout appearance, growth habit, leaf type, flower color and disease reaction are also used in identification and certification. Unfortunately, these morphological traits are inherently weak identifiers since they are subject to errors caused by environmental influences and subjectivity in observations. In addition, these traits are not all available for sampling at a single time. For these reasons, accurate identification of cultivars as well as unnamed breeding lines, has been a fundamental problem that confronts the plant breeders, certification agencies, and tissue culturists within the industry.
. With the advent of utility patents and proprietary rights being sought for new cultivars, it is imperative that methods that can generate new characters for potato identification are explored. These methods should provide a reliable, rapid, and efficient means to discriminate between potato cultivars. Numerous attempts to utilize electrophoretic techniques to separate biochemical and DNA markers for potato cultivar discrimination have been reported (7, 11, 13, 15, 18, 26). Isozymes, which are multiple molecular forms of an enzyme, have recently been used to characterize cultiv:ars in a number of crop species (2, 3, 4, 6, 20). Many isozyme loci, as revealed through starch electrophoresis, have been identified in potato and their inheritance studied (9, 14, 19, 23). These isozyme loci are valuable genetic markers because they exhibit codominance, lack epistatic and environmental effects, can be sampled nondestructively, and are expressed in various tissues (25). In, this paper, we will examine the value of 16 isozyme loci to generate genotypically unique patterns to discriminate North Ame~ican potato cultivars.
Materials and Methods
One hundred and twelve (112) named potato cultivars and advanced bre~dinglines were used in this study. Tubers were kindly supplied by Drs. N. S. (Bud) Wright (Agriculture Canada), R. W. Chase (Michigan State University) andJ. J. Pavek (USDA, Aberdeen, Idaho) and tissue culture plal~tlets of Garnet Chili by Dr. J. Bamberg (IR-l, Sturgeon Bay, Wisconsin). These clones are listed in Table 1.
Two tuber samples and two leaf samples were taken from each culti~
var for the analysis. Potato tubers were maintained in 4 C storage. One day prior to sampling the dormant tubers were moved to the laboratory and a small (150 mg) sample was extracted from the tuber eye for tissue processing. Tubers were then planted in the greenhouse, and a newly expanded leaflet (150 mg) was sampled from each plant for immediate proceSSIng.
Horizontal starch electrophoresis was employed. Procedure regarding tissue processing, electrophoresis, staining, and nomenclature has been

1991) DOUCHES AND LUDLAM: ELECTROPHORETIC CHARACTERIZATION 769
TABLE 1. - Potato cultivars and advaru;ed breeding lines survey.
~ar ~ar
Clone of Release Clone of Release
Purple Viking Bake King 1967
White Rose 1893 Centennial Russet "Colo 1" White Rose "Mettler W" Centennial Russet "Colo 2"
Columbia Russet 1919 Atlantic "WF31-4"
LaChipper 1962 LaBelle 1989 Monona 1964 F70021 1990
Garnet Chili 1853 Norchip 1968 Early Rose 1861 Norchief 1968 Early Ohio 1871 Spunta 1968 Irish Cobbler "Maine" 1876 Abnaki 1970 Irish Cobbler "BC" Raritan 1970 Burbank 1876 Hudson 1972 Russet Burbank Nooksack 1973 Beauty of Hebron 1878 Nampa 1973 Bliss Triumph 1878 Targhee 1973 Green Mountain 1885 Bison 1974 Rural New Yorker 1888 Centennial Russet 1976
Up-to-Date 1894 Tobique 1976 Bintje 1910 Butte 1978 Red McClure 1910 Jemseg 1978 Hindenburg 1916 Denali 1978 Arran Victory 1918 -Atlantic 1978
Katahdin 1932 Belrus 1978 Warba 1933 Belchip 1979 Warba "White skin" Rideau 1979 USDA 41956 1933 Shepody 1980 Chippewa 1933 Rosa 1980 Sebago 1938 Yukon Gold 1980 Menominee 1940 Rhinered 1981 Ontario 1946 Caribe 1981 Kennebec 1948 Lemhi Russet 1981 Red LaSoda 1948 Chipbelle 1981 Red Pontiac 1949 Oceania 1981 Pontiac "White skin" 1938 Sangre 1982 Pungo 1950 Conestoga 1982 Keswick "NB1" 1951 Yankee Chipper 1983 Keswick "NB2" Tolaas 1984 Saco 1954 Hampton 1985 Norland 1957 Elba 1985 Norland "Dark Red" Eramosa 1987 Norland "BC" HiLite Russet 1987 Fundy 1958 Tejon 1987 Superior 1961 Krantz 1988 Superior "Nebraska" Michigold 1988 Superior "Ontario" Minn 10162 1988 Onaway 1961 Saginaw Gold 1988 La Rouge 1962 Shasta 1988 Viking 1962 Steuben 1989

770 AMERICAN POTATO JOURNAL (Vol. 68
TABLE 1.- continued
~ar ~ar
Glone of ~lease Glone of Release
Norgold Russet "BC" 1964 NDD277-2 1990 Norgold Russet "M1" A7411-2 1990 Norgold Russet "M2" ND860-2 1990 Norgold Russet "19" MS716-15 1990 Chieftain 1966 MS702-80 1990 Desiree 1967 Spartan Pearl 1990 Wauseon 1967 Frontier Russet 1990 Alaska Frostless 1967 LA12-59 1990
previously described by Douches and Quiros (9). However, a Tris-borate, pH 8.3 buffer and gel system (24) was substituted for the Tris-citrate, pH 7.8 buffer. The two buffer systems that were utilized resolved 16 isozyme loci in 13 enzyme systems (Table 2). To facilitate the interpretation of the bandil~gpattern, two samples from each genotype was run side by side on a gel. A diploid clone from S. phureja, 84810, was used as a control on each gel to accurately identify the allozyme bands. Following interpretation, the genotypes for each locus of each cultivar were transcribed to a computer database for sorting (Lotus 123). The summation of the electrophoretic pattern for each cultivar is referred to as its "fingerprint?' These data were then analyzed using an unweighted group pairs mean (UWGPM) analysis for the personal computer (program written by Dr. K. Ritland, University of Toronto). A dendrogram was then constructed to visualize the separation of the cultivars on the basis of the fingerprints.
Results
Tuber and leaftissue"sampleswere assayed from 112 clones in 13 enzyme systems. A total of 16 enzyme loci were expressed strongly and resolved using leaf and tuber tissue (Table 2). The Sdh-l and Est-c loci generated complex patterns that could not be interpreted without progeny testing. The complexity was attributed to mu~tiple-bandedalleles and overlapping bands within each locus which precluded accurate transcription to numerical data. These loci were excluded from the UWGPM analysis; however, they were examined when identical patterns were observed between clones. Fortythree alleles were identified within the 14 scorable loci.
The value of each locus as a component of the electrophoretic fingerprint is a function of the number of alleles at each locus and their respective frequencies. Allele frequencies for each locus are described in Table 3. One locus, Tpi-l, was not polymorphic among the clones tested, therefore, it was not 'useful in developing a fingerprint. The qlost useful loci in

---TABLE 2.-Isozyrne loci assayed in potato.
Buffer System Enzyme
Histidine-Citrate, pH 5.7 "
Tris-Borate, pH 8.3
Malate dehydrogenase "
6-Phosphogluconic acid dehydrogenase Isocitric acid dehydrogenase Acid phosphatase Shikimic acid dehydrogenase Phosphoglucose isomerase Diaphorase Peroxidase Alcohol dehydrogenase Phosphoglucomutase
" Glutamate oxaloacetate transamiJ)ase
Triose phosphate isomerase Esterase
lL=leaf, T=tuber eye (including periderm).
Loci
Mdh-l Mdh-2
6-Pgdh-3
Idh-l Aps-l
Sdh-l
Pgi-l Dia..l Prx-3 Adh-l Pgm-l Pgm-2
Got-l Got-2
Tpi-l Est-c
I--L
\0 \0 I--L
0 0 c::: (1 ::tTissue expression 1 tTj r;n
L,T ~ L,T 0
~ c:::
L,T 0 ~
L ~ tTjT ~ tTj (1
L ~
0 ~
~L,T ::tL 0
~T trj
T ~ 0L,T 0
L,T ::t > ~ L,T 0
L,T tTj ~
~
L N T ~
0 Z
""'-J ""'-J I--L
'-l '-l N
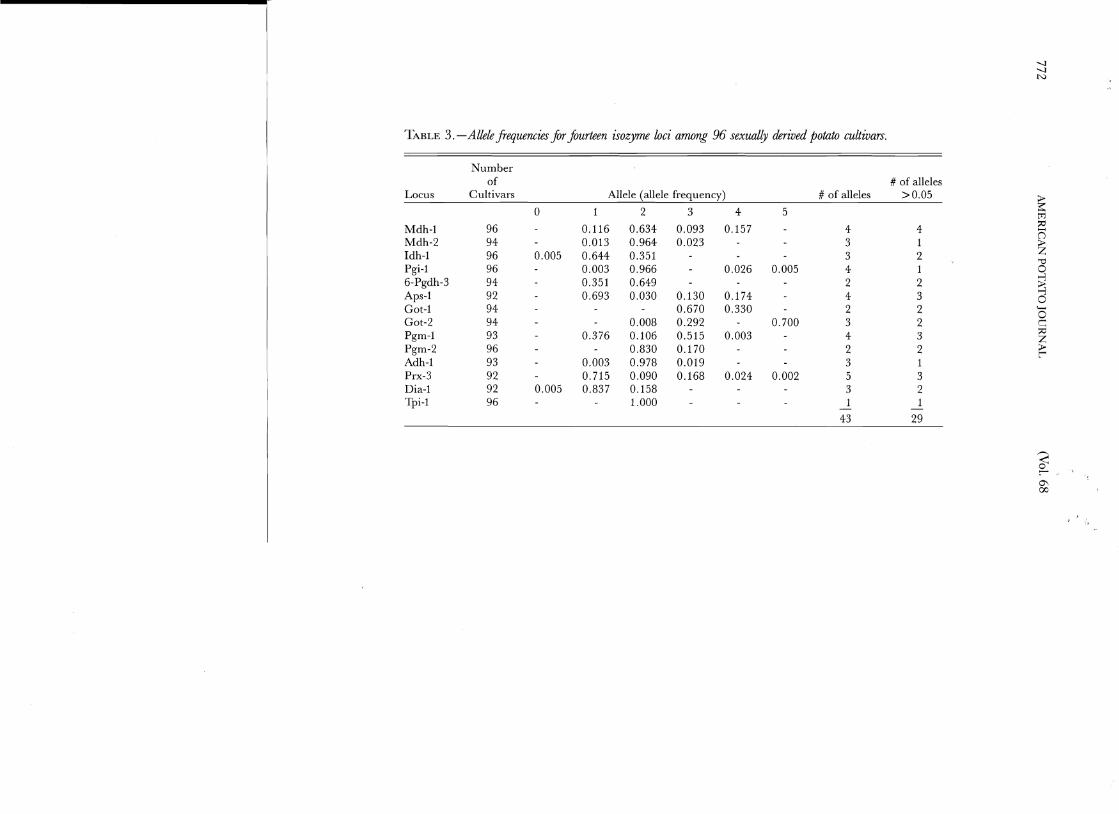
TABLE 3. -Allele frequencies for fourteen isozyme loci among 96 sexually derived potato cultivars.
Number of # of alleles
Locus Cultivars Allele (allele frequency) # of alleles >0.05 > 0 1 2 3 4 5 ~
tTJ
Mdh-l Mdh-2 Idh-1 Pgi-1
96 94 96 96
--0.005 -
0.116 0.013 0.644 0.003
0.634 0.964 0.351 0.966
0.093 0.023
--
0.157 --
0.026
---
0.005
4 3 3 4
4 1 2 1
::0 C3 >Z "'0 0
6-Pgdh-3 94 - 0.351 0.649 - - - 2 2 ~ Aps-l 92 - 0.693 0.030 0.130 0.174 - 4 3 ~
0 Got-l Got-2
94 94
--
--
-0.008
0.670 0.292
0.330 -
-0.700
2 3
2 2
~
0 c Pgm-l Pgm-2 Adh-l
93 96 93
--
0.376 -
0.003
0.106 0.830 0.978
0.515 0.170 0.019
0.003 --
--
4 2 3
3 2 1
::0 Z >r
Prx-3 92 - 0.715 0.090 0.168 0.024 0.002 5 3 Dia-l 92 0.005 0.837 0.158 - - - 3 2 Tpi-l 96 - - 1.000 - - - 1 1
43 29
i jf,
0\ 00

1991) DOUCHES AND LUDLAM: ELECTROPHORETIC CHARACTERIZATION 773
developing a unique fingerprint were Mdh-l, Pgm-l, Aps-l, and Prx-3. Each of these loci carried at least three alleles in frequencies greater than 0.05. These frequencies would lead to a greater number of observable genotypic classes at each locus and, as a conseque11ce, contribute significantly to the fingerprint. On the other hand, Pgi-l, Mdh-2, and Adh-l each have a predominant allele in a frequency greater than 95%, therefore contribute little to the fingerprint.
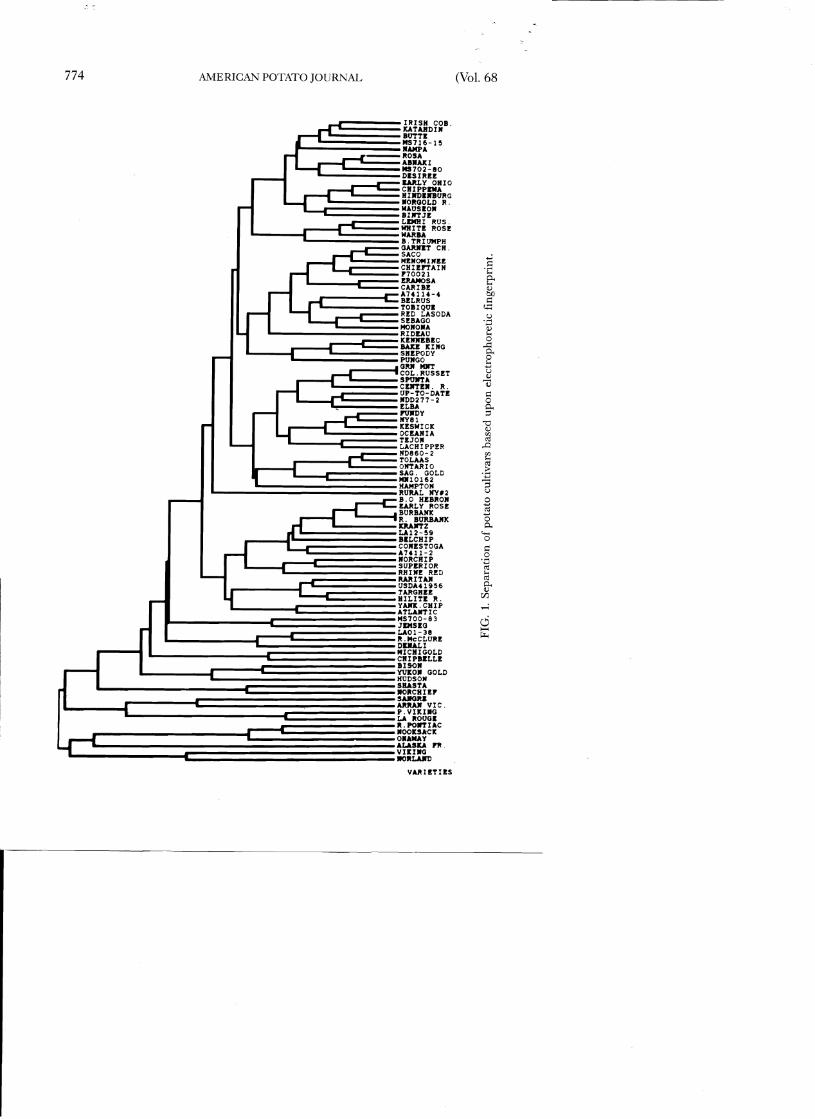
To separate the cultivars on the basis of their electrophoretic fingerprint, allele frequencies were calculated at each locus of each genotype, and then they were clustered using an UWPGM analysis. Of the 112 clones evaluated, 95 unique patterns were observed when all 13 enzyme systems
- were assayed. This discrimination between cultivars was also observed when either the array of isozyme loci from the ~eaf or tuber tissue was considered. The separation of these cultivars is expressed as a dendrogram in Figure 1. Duplicated patterns were observed for the clones listed in Table 4. In all but one case, the duplicate patterns were observed between line or clonal selections of cultivars of the same name. The other duplicated pattern was found between Green Mountain, a sexually-derived cultivar, and Columbia Russet, a selection of ambiguous origin.
All sexually-derived cultivars or advanced breeding lines were discriminated by their electrophoretic fingerprint, however, some cultivars had very similar patterns. Beauty of Hebron and Early Rose, which are sibs ofGarnet Chili, differed from each other by only one allele at Got-l, and, Belrus and Frontier Russet, unrelated russet cultivars, differed by only one allele at Mdh-l. This suggests that isozyme similarity mayor may not infer a close genetic relationship. Further inspection of the dendrogram shows that other cultivars with full-sib relationships, such as Red Pontiac and Red LaSoda or Katahdin and Chippewa, fail to share similar isozyme patterns that would give tight clustering in the dendrogram construction (Figure 1). On the other hand, Alaska Frostless, the only pentaploid (2n = 60 = 5x) cultivar examined, had four unique alleles, providing the most distinct fingerprint and unique clustering among the cultivars.
Discussion
The isozyme loci used in this study proved to be a valuable set of characters for cultivar identification in potato. This can be attributed to the allelic variation of the germplasm, the tetraploid nature of the crop, and the asexual means of propagation. However, isozyme loci, like conventional characters, also have finite limits of allelic diversity. The electrophoretic fingerprint that is generated for each cultivar is a result of the independent segregation of discreet Mendelian characters. For example, the ability to separate full sib progeny is a function of the allelic state of each locus in a cross. It is unlikely that with the high percentage of poly
774 AMERICAN POTATO JOURNAL (Vol. 68
--J...----- IRISH COB. ------- XATABDIJI
..------ BOTTI -.-------- MS716:"1 5
...--------- KAMPA-ftOSA
....--- A.WAXI ...-----MS702-aO
---------DISIR!! __~ --!AJtLY OSlO
CBIPPDA ....---- HllfDlnURG
....------WOltOOLD R . ....------NAUSEOII
-~....------811fT.]!_~----LDOII RUS. ------ WHIT! ROS!
...-----NARBA ...-------- 8. TRIUMPH
OAJUUT CR. SACO
....----MENOMIJf!! L ----- CHI!P'!'AIH
L----C===-----..-----
~~~A P70021
...-----... -- ~itA6~-·
...-------TOBIQUI ____- -- RED LASODA
SEBAGO ...-----MOIfOJlA
...------------RIDEAUK!lfJfEBIC ---__-1"---1..---==== ~O~~NG -.----------- PUJfGO
...---.. g~.~SET
...---- SPUJrTA ...-----ClJfTIJI. R. -----' -----t1P-TO-DATE (1 ---- JfDD2 77 - 2 ~-----ELBA _~--- PUlfDYr-"--- NYel ...----KESWI CK
...-----L----C===== OC!AJfIA
l:g~~PPER HD860-2 ...__-tr-----l:.:=== ~~~io
.,.------ SAG. GOLD ---......------MJUOl62
-.-------------- HAMPTON ...-----------------RURAL HY.2
B.O H!BROII EARLY ROSE
...__.... BURBANJC R. BORBA1QC
....-----OAJrTZ ...--------LA12-59 ...---1·--------- ~~~~~;~GA
---,....--------A7.11- 2 ._------- IfORCRIP
....--"'l.__=:::::::::::::: ~~==I~~D ._------- RA!'ITAJf
.---....------- t1SDA.19S6...------1 TAROSI! ----.------ HILITI R.
......;L.------c=========X~~~p,.. <===========:~~~G8 3
__---------LAOl-38 L--------t:==~========= .DUAL IIt . McCLUR!
L.----------c===========~i~i~~t~ YUKOJI GOLDL------r---~I:============ 81 SOli
·_-_-_-_-_:-[-:-:-:-:-:-:-:-:-:-:-:-:-:-:-:-:-:-::-:-:-:-:-:-:-::-:- :~:~: JIOJtCBIEP
AJtbJf VIC .
L
r------C================== SAJIGU..----.....l---------------t:==========LAP. VIJCIJIGItOCOI
r---------------[-:-:-:-:-:-~-I~,. =-:~:i~:C::-:-:-::-:-:-::-:-:-::-:-:-:-:-: O••MAY
l --:--------:------:------:------i-:-:-:-:-:-:-::-:-::-:-:-::-:-:::-:-:-::-:-:-::-:-:-::-:-:-::-:-:-=-~~ IK)ItLAJIDn.
VAIUITIIS

1991) DOUCHES AND LUDLAM: ELECTROPHORETIC CHARACTERIZATION 775
TABLE 4. - Clones with duplicate multilocus electroplwretic patterns.
Clone Duplicate
Russet Burbank Burbank Red Pontiac Pontiac (white skin) Red Warba Warba (white skin) Irish Cobbler "BC" Irish Cobbler (Maine source) Norgold Russet "BC" Norgold Russet "M1;' Norgold Russet "19"
Norgold Russet "N2" Norland Dark Red Norland, Norland "BC" Atlantic Atlantic WF-31-4 White Rose White Rose "Mettler W" Superior Superior "Neb;' Superior "Ont" Centennial Russet Centennial Russet "Colo 2"
Centennial Russet "Colo I" Keswick "NB 2" Keswick "NB 2" Green Mountain Columbia Russet
morphic loci examined, two related cultivars will have identical fingerprints. Olivier and Martinez-Zapater (18) were able to separate 68 or 74 cultivars with 9 isozymes. However they found that Katahdin and Sebago had identical phenotypes in their analysis, while we were able to distinguish these cultivars with a greater array of isozymes.
The degree of intercultivar variation has a large effect upon the utility of these isozyme loci for cultivar identification. The cultivated potato of North America is a relatively new crop and has been developed from a narrow genetic base (12). Most cultivars can be traced to a few introductions in the mid-1800's (21) and the genetic relationships are quite close (17). Despite concerns of a restricted genetic base, the allelic diversity detected in this study (Table 3) was adequate to separate the cultivars. These results are in accordance with previous electrophoretic work in potato (7, 13, 18, 26). This electrophoretic polymorphism is distributed both between the number ofloci examined and the number of alleles detected at a locus. Thirteen of the 14 loci were polymorphic with a range of 2 - 5 alleles per locus. The number of heterozygous loci varied from 5 - 11 per clone. Tri-allelic loci were detected in Mdh-l, Prx-3, and Pgm-l (Figure 2). Garnet Chili and Katahdin, progenitors of many important cultivars, are heterozygous at 9 and 8 loci, respectively. Interestingly, only diallelic states were observed at the heterozygous loci of these cultivars.
The utility of the isozymes for cultivar identification can also be attributed to the potatds polyploid nature and asexual means of propagation. A tetraploid organism can have a greater array ofheterozygous classes at a diallelic locus and also carry up to 4 alleles per locus if additional polymorphism exists. In a clonally-propagated crop, such as potato, both heterozygous and homozygous genotypes are fixed. This expands the number
776 AMERICAN POTATO JOURNAL (Vol. 68
B
c
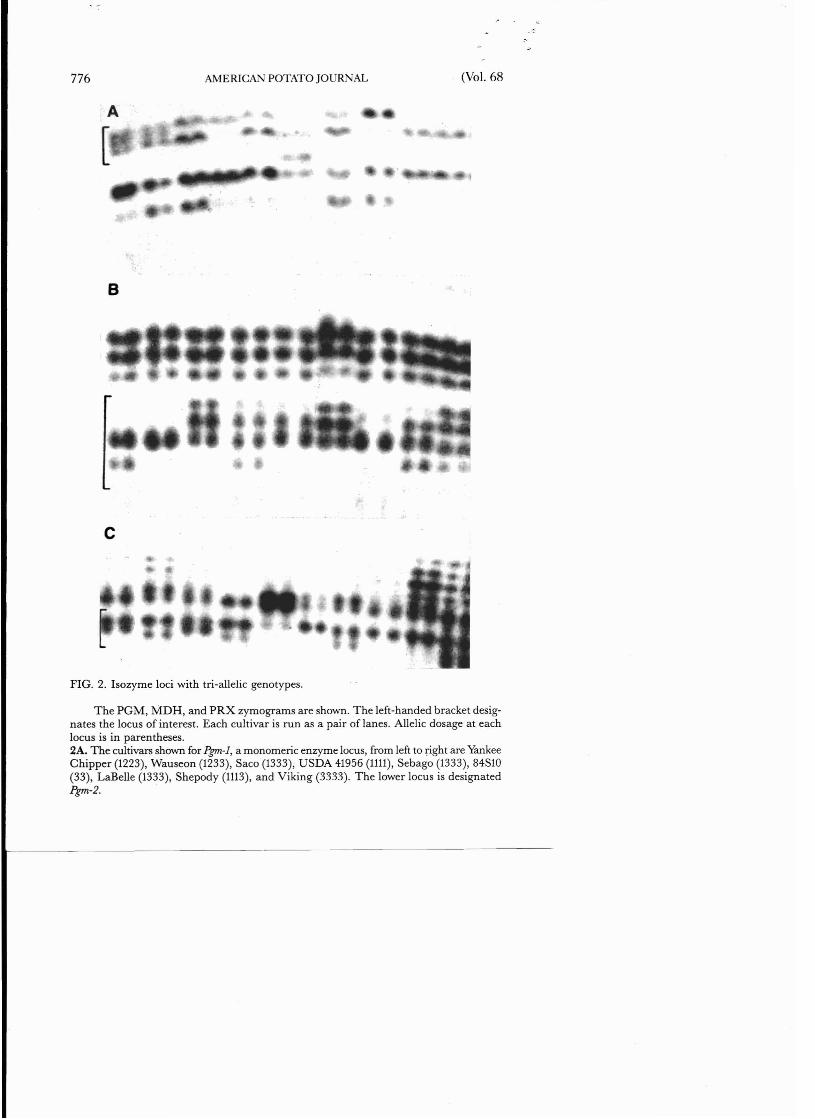
FIG. 2. Isozyme loci with tri-allelic genotypes.
The PGM, MDH, and PRX zymograms are shown. The left-handed bracket designates the locus of interest. Each cultivar is run as a pair of lanes. Allelic dosage at each locus is in parentheses. 2A. The cultivars shown for Pgm-l, a monomeric enzyme locus, from left to right are Yankee Chipper (1223), Wauseon (1233), Saco (1333), USDA 41956 (1111), Sebago (1333), 84S10 (33), LaBelle (1333), Shepody (1113), and Viking (3333). The lower locus is designated Pgm-2.

1991) DOUCHES AND LlTDLAM: ELECTROPHORETIC CHARACTERIZATION 777
FIG. 2. continued
2B. The upper locus is Mdh-2. The cultivars examined for Mdh-l, a dimeric enzyme locus, from left to right are Red McClure (2224), Rural New Yorker (2222), Columbia Russet (1122), Bliss Triumph (1224), Beauty of Hebron (1222), 84S10 (12), Irish Cobbler (2233), White Rose (1224), and Warba (1124).
2C. The Prx-3locus is a monomeric enzyme locus that is not expressed in leaf tissue. The cultivars shown are Burbank (1112), Irish Cobbler (1113), Early Rose (1122), Early Ohio (1113), 84S10 (not determined from leaf tissue), Bintje (1111), Bison (1113), F70021 (1111), and Menominee (1111).
of stable isozyme phenotypes available to classify the cultivars. Asexual propagation also eliminates the need for extensive intracultivar sampling.
We attempted to maximize our ability to identify potato cultivars by classifying all the heterozygous classes in this study. Prior knowledge of the quaternary structure, genetic basis, and range of allelic variation allows the genotype to be inferred from the observed banding pattern for most loci (8). For example, a dimeric enzyme locus, Mdh-j, can have 3 diallelic heterozygotes in a tetraploid potato: Mdh-]ljl]2j2 (duplex), Mdh-]lj2]2j2 and Mdh-jljljlj2 (simplex). These heterozygotes can be differentiated on the basis of band dosage (23). A duplex class will show a balanced 3-banded phenotype in a 1:2:1 ratio. The simplex classes are expressed as unbalanced 3-banded patterns that have a dosage relationship in a 9:6:1 ratio which favors the more frequent allele (Figure 2). For a monomeric enzyme, such as PRX or PGM, the dosage relationship at a locus is directly proportional to the allelic dosage. Genotypic inference from the banding pattern is possible if proper control genotypes are also run on the same gel and the clones are run as a paired sample.
Various line selections were compared in this study (Table 4). The isozymes we examined did not distinguish between sports or line selections of a cultivar. These 16 loci code for only a very small fraction of the entire potato genome. The likelihood that isozyme changes would be correlated with skin mutations or other physiological differences is quite low or negligible. Using isozyme analysis, May et ale (15), concluded that cultivar mixes (other red-skinned cultivars) occurred in Red Pontiac rather than a mutation or mitotic recombination. Therefore, if numerous isozyme differences exist between a cultivar and a putative line selection, it is most likely mislabeled.
Isozyme analysis can be useful when examining cultivars of unknown origin. The cultivar, HiLite Russet, was found in a field of Butte. The distinct fingerprints between the two cultivars indicates that HiLite Russet was probably a cultivar mixture in that field of Blltte. The fingerprint of Shasta, a selection of unknown origin, is unlike any other long white cultivar such as Shepody, White Rose, or Burbank. Again, Shasta could have

778 AMERICAN POTATO JOURNAL (Vol. 68
been selected from a clonal mixture. Alternately, Columbia Russet and Green Mountain have identical fingerprints. Columbia Russet, according to MacLeod and Jones (16), possibly originated from a seedling of Wee McGregor in 1919. However, their morphological description and field performance is very similar to that of Green Mountain. It is possible that a skin mutation was selected out of Green Mountain similar to other sports that were selected out of Burbank, Pontiac, LaSoda, and Warba. Electrophoretic examination of Wee McGregor could provide evidence toward this hypothesis.
One of the predominant reasons isozymes have been exploited in cultivar identification is their apparent environmental stability (7, 13,22,26). In addition, we have observed stable electrophoretic patterns for a sample of cultivars grown in various locations in North America that have identical electrophoretic patterns to our greenhouse samples (unpublished dat~).
It is of equal importance that sample preparation be standardized so that the results are reproducible. Staub et ale (22), has shown that tuber extracts can be stored at - 60 C for 70 days without loss in activity or altered mobility for 4 enzyme systems. To obtain consistently high enzyme activity, we chose two plant tissues for this study. A newly expanded leaflet or a small section of the tuber that surrounds the tuber eye will each give consistent expression for a set of 16 loci. In addition, these loci have resolvable variation under the laboratory conditions described earlier. Artifacts can occur and are usually attributed to the reduced quality of the tissue sample or a lack of adherence to standardization in the procedure. Any consistent qualitative (allelic differences) or quantitative (intralocus dosage differences) banding variation we observed between cultivar zymograms can be attributed to genotypic differences between cultivars.
To consider the potential of a character, such as a isozyme locus, in cultivar description and identification Bailey (1) lists four basic criteria. The isozy~es utilized in this study follow these criteria. Isozyme analysis ofpotatoes is reliable, simple, fast, accurate and objective. DNA-based markers are also reliable, accurate and objective, however, require greater time, cost and expertise (10). Prior to registration and release, all new cultivars should require to have a standardization electrophoretic fingerprint. This procedure would simplify problen~s regarding cultivar purity or confirmation of a cultivar in tissue culture stocks. Unfortunately, electrophoretic fingerprints, via isozyme analysis, will not be useful in distinguishing line selections or transgenic plants derived from a previously released public cultivar.
The purpose of this study was to assess the value ofisozymes as additional characters for cultivar identification. However, isozyme analysis
.should not be used to the exclusion of conventional characters. The real value of these isozyme loci is in combination with morphological characters. In practice, the standard morphological characters used in certification can be supplemented with these additional discreet characters.

1991) DOUCHES AND LUDLAM: ELECTROPHORETIC CHARACTERIZATION 779
Acknowledgments
Authors thank Mr. Ed Varga and Mrs. Katherine Cullen for technical assistance. This research was supported in part by the Michigan Agricultural Experiment Station and Michigan Potato Industry Commission.
Literature Cited
1. Bailey, D.C. 1983. Isozymic variation and plant breeders' rights. In: Isozymes in Plant Genetics and Breeding, Part A. Eds. S.D. Tanksley and T.J. Orton. Elsevier Science Publishers, Amerstdam .425-440.
2. Bassiri, A. and M.W. Adams. 1978. Evaluation of common bean cultivar relationships by means of isozyme electrophoretic patterns. Euphytica 27:707-720.
3. Bringhurst, R.S., S. Arulsekar, J.F. Hancock, Jr. and V. Voth. 1981. Electrophoretic characterization of strawberry cultivars. J Amer Soc Hort Sci 106(5):-684-687.
4. Byrne, D.H. and T.G. Littleton. 1989. Characterization of isozyme variability in apricots. J Amer Soc Hort Sci 114(4):674-678.
5. Chase, R.W. 1989. North American Potato Inventory. Certification Section of the Potato Association of America.
6. Cousineau,J.C. and D.J. Donnelly. 1989. Identification of raspberry cultivars in vivo and in vitro using isoenzyme analysis. Hortscience 24(3):490-492.
7. Desborough, S.L. and S.J. Peloquin. 1968. Potato variety identification of electrophoretic patterns of tuber proteins and enzymes. Am Potato J 45:220-229.
8. Douches, D.S. and C.F. Quiros. 1987. Use of 4x-2x crosses to determine genecentromere map distances of isozyme loci in Solanum species. Genome 29:519-527.
9. Douches, D.S. and C.F. Quiros. 1988. Additional loci in tuber-bearing Solanums: Inheritance and linkage relationships. J Hered 79:377-384.
10. Douches, D.S., R. Freyre and K. Hicks. 1990. Use ofRFLPs to fingerprint North American potato varieties. In: First Planning Conference for the Use of Molecu1ar Techniques in Potato Germplasm Enhancement. CIP pp. 166-175.
11. Gebhardt, C., C. Blomendahl, U. Schachtschabel, T. Debener, F. Salamini and E. Ritter. 1989. Identification of2n breeding lines and 4n varieties ofpotato (Solanum tuberosum ssp. tuberosum) with RFLP fingerprints. Theor Appl Genet 78:16-22.
12. Glendenning, D.R. 1983. Potato introductions and breeding up to the early twentieth century. New Phytol 94:479-505.
13. Loeschcke, V. and H. Stegemann. 1966. Proteine der Kartoffelknollen in Abhangigkeit von Sorte und Virosen (Polyacrylamide - Elektrophorese). Phytochemistry 5:985-991.
14. Martinez-Zapater, J.M. andJ.L. Olivier. 1984. Genetic analysis of isozyme loci in tetraploid potatoes (Solanum tuberosum L.. ). Genetics 108:669-679.
15. May, B., J.E. Staub and L.J. Kuhns. 1982. Potato cUltivars: genetic variation within putative clones. Am Potato J 59:179-184.
16. MacLeod, H.S. and W. Jones. 1937. A new variety from British Columbia. Am Potato J 14:93-99.
17. Mendoza, H.A. and F.L. Haynes. 1974. Genetic relationship among potato cultivars grown in the United States. Hortscience 9:328-330.
18. Olivier, J.L. andJ.M. Martinez-Zapater. 1985. A genetic classification of potato cultivars based on allozyme patterns. Theor Appl Genet 69:305-311.

780 AMERICAN POTATO JOURNAL (Vol. 68
19. Quiros, C.F. and N. McHale. 1985. Genetic Analysis of isozyme variants in diploid and tetraploid potatoes. Genetics 111:131-145.
20. Rajora, a.p. 1989. Genetic structure and identification ofPopulus deltoides clones based on allozymes. Genome 32:440-448.
21. Salaman, R.N. 1926. Potato Varieties. Cambridge University Press, Cambridge, 378 p.
22. Staub, J.E., L.J. Kuhns, P. Grun and B. May. 1982. Stability of potato under different storage regimes. J Am Soc Hortic Sci 107:405-408.
23. Staub,J.E., L.J. Kuhns, P. Grun and B. May. 1984. Genetic basis for isozyme variation for alkaline phosphate and glucosephosphate isomerase in Solanum. Theor Appl Genet 67:505-513.
24. Stuber, C.W.,J.F. Wendel, M.M. Goodman andJ.S.C. Smith. 1988. Techniques and scoring procedures for starch gel electrophoresis from maize (Zea mays L.). Technical Bull. 286, North Carolina Agricultural Research Service, NCSU, Raleigh, North Carolina.
25. Tanksley, S.D. 1983. Molecular markers in plant breeding. Plant Mol BioI Rep. 1:3-8. 26. Zwartz, J.A. 1966. Potato varieties and their protein electropherogram characteris
tics. Eur Potato J 9:111-128.



















![Scripta Mathematica [Si1756] Simson, R.,Los Seis …godfried/publications/euclid.spanish.pdfWhat is Mathematics? Oxford University Press, 1981. Oxford University Press, 1981. [Du90]](https://img.pdfslide.net/doc/110x75/5b05f0c37f8b9ad1768c1c5c/scripta-mathematica-si1756-simson-rlos-seis-godfriedpublicationseuclidspanishpdfwhat.jpg)









