Embed Size (px)
Citation preview

ADVANCES IN HEPATITIS C 1089-3261/01 $15.00 + .OO
IMMUNOPROPHYLAXIS OF HEPATITIS C VIRUS INFECTION Alfred M. Prince, MD, and Mohamed Tarek Shata, MD, PhD
Hepatitis C virus (HCV), an RNA virus related to the flaviviruses and pestiviruses, is characterized by a predominance of infections lasting lifelong. Only about 20% to 30% of infections terminate after the acute phase.6 Hepatitis C virus infections exist in humans throughout the world, with the prevalence of chronic infection in adults varying from 0.5% to as high as 25% (in Egypt).'T2r4 The number of chronically HCV- infected people has been estimated as between 200 and 700 million. These estimates are not exact, because the actual prevalence remains unknown in large parts of the world.
The medical importance of HCV infection is primarily the risk of development of cirrhosis and liver cancer in persons with chronic infections.31, 4o It has been estimated that about 25% of chronic carriers who live for at least 20 years after infection will develop cirrhosis, and that 1% to 4% of those with cirrhosis will develop hepatocellular carci- noma annually. Hepatocellular carcinoma has been estimated by the World Health Organization (WHO) to be the world's most common cancer, accounting for more than 18% of all cancers worldwide.
An epidemic of HCV infection occurred in Japan after World War I1 because of the introduction of parenterally administered antibiotics with- out the availability of disposable needles and syringes and also because of widespread pulmonary surgery for tuberculosis requiring massive transfusions. Twenty to 30 years later, the incidence of hepatocellular carcinoma increased two- to threefold, and the proportion of cases which
Supported by NIH Grant No R01-A147349
From the Lindsley F. Kimball Research Institute of the New York Blood Center, New York, New York
~~~~ ~~
CLINICS IN LIVER DISEASE
VOLUME 5 - NUMBER 4 * NOVEMBER 2001 1091

1092 PRINCE & SHATA
were positive for hepatitis B (HBV) infection dropped from 70% to 80% to 20% to 30’/0.~~, 40 The role of vaccines in prevention of hepatocellular carcinoma has been recently re~iewed.3~
EPIDEMIOLOGY OF HEPATITIS C VIRUS INFECTION
Hepatitis C virus is transmitted almost entirely by blood-to-blood contact. Previously, about 5% of cases in the United States were related to blood transfusion, but because of the great improvements that have been made in blood screening tests, posttransfusion HCV infections are now exceedingly rare. Most cases in developed countries are now related to illicit parenteral-drug use. Sexual transmission occurs but is rare, with a rate of transmission lower than 1% per year between couples in which one is chronically HCV infected. Vertical transmission is also rare, with reported frequencies of 1% to 5%, except in cases of maternal HCV-HIV coinfection, for which transmission rates of 10% to 20% have been reported. In developing countries, syringes and needles continue to be reused, often without adequate sterilization, and are probably the major route of transmission, other than illicit injecting-drug use (where preva- lent). In some parts of the world, practices such as scarification, ritual circumcision, and acupuncture also probably play a role.
IS A HEPATITIS C VIRUS VACCINE NEEDED?
The high rate of chronicity, high global prevalence, and most im- portantly, the high rate of lethal chronic liver disease occurring in chroni- cally HCV infected people would seem to provide abundant justification for giving the development of HCV vaccines the highest priority. Unfor- tunately, some argue that this conclusion is false. In the developed world the incidence of new HCV infections is undoubtedly dropping to quite low levels, resulting in part from the extraordinary increase in blood safety which has occurred during the last decade. Widespread introduc- tion of nucleic acid testing of transfused blood will probably lower this risk to the vanishing point. Furthermore, medical personnel in devel- oped countries use disposable syringes and needles, eliminating this mode of transmission. Illicit drug use will probably continue, but unfor- tunately users of illicit drugs are not readily vaccinated. The limited need for a vaccine in the developed world provides little incentive for vaccine manufacturers to enter this field. Thus it is likely that support from the public sector will be needed to develop an HCV vaccine.
In developing countries, in which most of the world’s population lives, HCV infections continue to be a major public health threat. In these countries blood screening is rarely performed, especially for HCV because of the high cost of reagents. Furthermore, reuse of injection equipment by medical and paramedical personnel is likely to continue for many years, because of the cost of disposable syringes and needles

IMMUNOPROPHYLAXIS OF HEPATITIS C VIRUS INFECTION 1093
relative to local economic realities. Regrettably, intravenous-drug abuse is also spreading in many parts of the developing world. We suggest that in developing countries, a vaccine for HCV is as important as the vaccine for HBV, and it is hoped that the two can eventually be com- bined. Because both HCV and HBV are equally responsible for the development of cirrhosis and liver cancer in a significant proportion of chronically infected persons, these vaccines must be an important goal of global public health.
IMMUNITY IN HEPATITIS C VIRUS INFECTION
In contrast to most viral infections, immunity after natural infection by HCV is weak.30, 35, 5n There is, however, growing awareness that although reinfection after reexposure is the rule, second infections are usually mild and self-limited, so an important degree of immunity does exist. No correlation was observed between anti-HCV envelope 2 (E2) antibodies and self-limitation of HCV infection^.^^, 47 By contrast, numer- ous investigators have observed that patients with self-limited infections have much higher levels of cell-mediated immunity than those who progress to chronic infections. This immunity has been found both in T- helper (Th) responses7 and in cytotoxic T-lymphocyte (CTL) respon- ses.ln, l6 These findings underscore the importance of cell-mediated im- munity in determining the outcome of this infection.
IMPEDIMENTS TO DEVELOPMENT OF HEPATITIS C VIRUS VACCINES
As previously reviewed, there are numerous potential impediments to the development of HCV vaccines.33
Genotypes
At least six major genotypes of HCV e x i ~ t . ~ , ~ ~ The prevalence of these genotypes varies markedly in different parts of the world. Thus, especially if vaccines are based on neutralizing antibodies, a complex mixture of antigens and neutralizing epitopes is needed to provide a vaccine with universal efficacy. At present, however, there is little, if any, knowledge of cross-protection among different genotypes. It is known that chronically infected animals can be reinfected with HCV of the same or different genotypes.3n, 35 It is not clear, however, whether infection by other genotypes can occur in animals or humans who have been success- fully immunized with a single genotype.

1094 PRINCE & SHATA
Quasispecies
Even within a single infected individual, HCV populations are composed of molecular clones having different nucleic acid sequences, particularly in the hypervariable 49 These variants are termed quasispecies. During chronic infections there is continuous selection of quasispecies, probably as a result of immune selection. These quasispe- cies are likely to present a problem for vaccine development, again most importantly for vaccines based on neutralizing antibodies, because these vaccines act against a limited number of epitopes. At present, however, there is no knowledge of the actual importance of quasispecies in viral immunity after natural infection or after immunization.
Lipid Association
A large proportion of HCV virions are now known to be associated with low-density lipoproteins (LDL) and very low-density lipoproteins (VLDL) of host origin.19, 37, 44 The association with these lipoproteins is sufficiently tight to impede reaction of antiviral antibodies with virions; thus the lipid-coated virions are not precipitated with anti-IgG antibod- ies even in serum containing anti-HCV antib~dies.'~, 2o The implications of these findings for the development of a neutralizing antibody-based HCV vaccine are obvious.
Protective Immunity
The type of immune response that is protective against HCV infec- tion is not yet fully understood. The mechanisms of protective immunity td HCV infection need to be further defined.
Animal Model
Another hindrance for developing an HCV vaccine has been the limitation of a suitable animal model. The chimpanzee is the only animal model for HCV infection. For many investigators, the expense of using chimpanzees for vaccine testing is prohibitive.
Statistical Problems
The number of chimpanzees required for statistical validation of a vaccine trial in chimpanzees, the only animal model for HCV infection, assuming that the vaccine produces sterilizing immunity, is shown in Table 1. Nine chimpanzees would have to be used to assure statistically significant results. The investment required for such a study, necessitated

IMMUNOPROPHYLAXIS OF HEPATITIS C VIRUS INFECTION 1095
Table 1. STATISTICAL CONSIDERATIONS FOR PROPHYLACTIC IMMUNIZATION*
Immunized: Controls: Proportion Proportion P
Infected Infected (Fisher’s Extract)
0.33 0.06 0.01 0.002
*Using infection determined by polymerase chain reaction as an end point P = statistical probability.
by the high cost of maintaining a chimpanzee colony for the life span of the chimpanzees used, which must be borne by the investigators, approaches 1 million dollars. Furthermore, the assumption underlying this analysis, that sterile immunity will be produced by the vaccine, is probably unrealistic and unnecessary, especially with vaccines that pro- vide efficacy on the basis of induction of strong cell-mediated immunity.
Table 2 analyzes the statistical requirements for a vaccine which induces a form of immunity which results in self-limited infections and prevents chronic infection. Such an outcome would be expected with a vaccine producing cell-mediated immunity. Here the statistical consider- ations become even worse, because the use of even 12 chimpanzees barely approaches statistical significance. Thus, the cost of statistical validation is a major impediment to the development of HCV vaccines.
POTENTIAL HEPATITIS C VIRUS VACCINES
Subunit Vaccines Based on Viral Proteins, Peptides, or Viruslike Particles
Viral Proteins
In 1994, in a pioneering study, Choo et a1 reported the first attempt to immunize chimpanzees against HCV using mammalian cell-derived
Table 2. STATISTICAL CONSIDERATIONS FOR PROPHYLACTIC IMMUNIZATION*
Immunized: Controls: Proportion Proportion P
Chronic Chronic (Fisher’s Extract)
0/2 1/2 0.33 0/4 1/2 0.33 0/6 2/3 0.08 0/8 2/4 0.09
‘Using chronicity as an endpoint and assuming 33% to 36% chronicity in controls. P = statistical probability.

1096 PRINCE & SHATA
El /E2 heterodimers, together with MF-59 and muramyl dipeptide adju- v a n t ~ . ~ Strong although transient humoral immune responses to these antigens were observed. The animals were challenged at the height of these responses. Five of seven animals totally resisted challenge with 10 chimpanzee infectious doses of homologous-strain HCV. When similarly immunized animals were challenged with a different strain of the same HCV genotype, however, protection was not ~bserved.’~” This finding suggested that the immunity was strain specific. More recently, Houghton reported that none of the immunized animals, challenged with either strain, developed chronic infection.lgb This finding suggested that an important degree of immunity had been induced, because the main goal of prophylactic vaccines is prevention of chronic infections. Mild, acute, self-limited infections are of no clinical significance.
Because of the high cost and limited availability of chimpanzees, many important questions remain concerning the suitability of protein- based vaccines for HCV immunization. It is not known whether chal- lenge of chimpanzees after levels of antibody declined would be effective (i.e., the duration of the immunity is not known). Furthermore, there is as yet no knowledge concerning cross-genotype immunity. The degree to which cell-mediated immunity was induced is also unclear. Nevertheless these studies have begun the investigation of prophylactic HCV immuni- zation and give encouragement for eventual success.
Peptides
The use of T-helper and CTL epitope peptides represents an interest- ing and potentially powerful approach to immunization. Shirai et a1 showed that such combinations are able to elicit strong HCV epitope- specific CTL response^.^^ Encouraging, although preliminary, results have also been reported on peptide-based immunization of chimpan- z e e ~ . ~
Viruslike Particles
Baumert et a1 have reported the generation of viruslike particles in insect cells using a recombinant baculovirus containing the genes of the HCV structural protein^.^ The particles resembled HCV in morphologic, biophysical, and antigenic properties and induced strong humoral re- sponses in experimental animals. Their ability to induce cell-mediated immunity remains unclear. Nevertheless, these easily produced particles are an interesting candidate vaccine for HCV.
DNA-based Vaccines
Cytotoxic T lymphocytes, specific for conserved viral antigens, can respond to different strains of virus, whereas antibodies tend to be strain-specific. This lack of specificity is one of major rationales for the recently growing interest in DNA-based vaccines.= DNA-based immuni-

IMTvIUNOPROPHYLAXIS OF HEPATITIS C VIRUS INFECTION 1097
zation permits cytosolic presentation to class I major histocompatibility complex (MHC) molecules in a manner closely similar to that occurring when viruses replicate in cells. Thus this form of immunization is partic- ularly well suited for the induction of cell-mediated immunity. Proof of principle that DNA-based immunization can protect against different strains of influenza A virus was provided by Ulmer et a1.46
Additional advantages of DNA-based immunization are high ther- mal stability, permitting delivery in the developing world without a cold chain, and the ease with which multiple immunogens, directed against different pathogens or diverse genotypes, can be combined in a single vaccine.
Numerous investigators have begun to explore the use of DNA- based immunization for immunization against HCV in mouse models.8, 13, 153 26, 27, 32, 39, 45 This form of immunization was confirmed to induce both cellular and humoral immunity, although in many cases the responses were suboptimal. It has been confirmed that DNA-based immunization can give CTL responses to epitopes from different gen0t~pes.l~
There is only one published report of an attempt to use DNA-based immunization in chimpanzees.12 In this study both humoral and cellular responses appeared predominantly only after challenge with live virus; however, these responses were anamnestic in type. Neither of these animals resisted challenge with live virus, although both had self-limited infections. Heile et a1 compared E l /E2 subunit protein-based immuniza- tion with DNA-based immunization and concluded that, if the aim is to elicit antibodies which interfere with binding of E2 to CD81 [NOB]), protein-based immunization is superior.I8
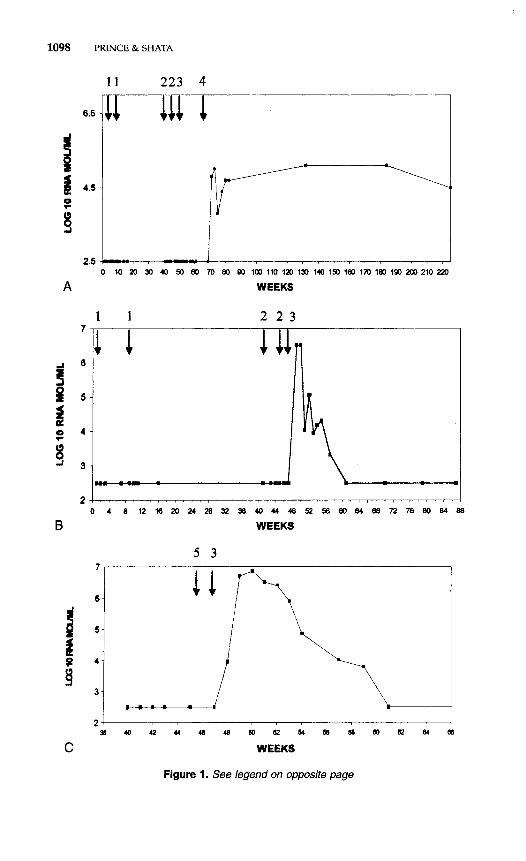
The authors have carried out a preliminary evaluation of DNA- canarypox-based immunization in chimpanzees. Two chimpanzees were immunized with plasmids expressing either HCV capsid, HCV nonstruc- tural (NS)3, or HCV NS5, each under the control of the CMV immediate early promotor, given as 0.5 mg DNA per plasmid in multiple intramus- cular sites 8 weeks apart. The HCV genes were derived from HCV genotype lb. The animals were boosted twice at 40 and 45 weeks after the initial immunization with 4 x lo8 plaque-forming units (pfu) of recombinant canarypox (ALVAC, constructed and provided by Virogene- tics-Aventis Pasteur, Toronto, Canada) given intravenously and with 4 X lo8 pfu given by multiple intramuscular injections. This regimen had given strong cell-mediated immune responses in preliminary evaluations in mice. The immunized animals, and a control animal which had received only parental ALVAC, were challenged at 48 weeks after onset of immunization with 10 chimpanzee infectious doses (CID50) of HCV-BK (genotype lb). The results are shown in Figure 1. The control animal, chimpanzee 317, became PCR-positive 1 week after challenge. Immunized chimpanzee 214 resisted challenge with 10 CID50 but became infected after challenge with 100 CID50. It is not clear whether the lack of infection after the lower-dose chal- lenge reflects partial immunity or whether the infectivity of the chal- lenge dose was borderline. Chimpanzee 214 became chronically
1098
!I a 4 3
A
r' P P
8 a L -I
B
i! i B
C
PRINCE & SHATA
1 1 223 4
0 10 M 30 40 50 60 70 80 90 100 110 IZU 130 140 19D 180 170 180 190 MD 210 2p
WEEKS
1 1 2 2 3
'7 1111 1
WEEKS
5 3
Figure 1. See legend on opposite page

IMMUNOPROPHYLAXIS OF HEPATITIS C VIRUS INFECTION 1099
Table 3. IMMUNIZING DOSES USED IN MICE AND CHIMPANZEES
DoselGM DoselGM lrnmunogen (in mice) (in chimpanzees) Ratio
DNA 5 IJG 40 ng 125 X Canarypox 2.5 x 107 PFU z x 1 0 4 ~ 1 ~ 125 X
PFU = plaque forming units
infected, but with a virus load more than 100-fold lower than that usually found in chronically infected chimpanzees. Again, this viral load may or may not indicate a degree of immunity. Immunized chimpanzee 191 became transiently infected after the low-dose challenge. Prelimin- ary determinations of cell-mediated immunity, using the y-interferon ELISPOT assay revealed a modest induction of interferon secretion (1000-4000 interferon-secreting cells/ lo6 lymphocytes after 6 days of in vitro stimulation with autologous recombinant canarypox virus) after DNA-based immunization. Surprisingly, however, the marked boost in cell-mediated immunity always seen in mice receiving canarypox boost- ers was not seen in these two chimpanzees. The results of this experi- ment are equivocal, although not particularly promising.
It is too early to draw conclusions regarding the efficacy of DNA- based immunization for HCV prophylaxis. Clearly the immune re- sponses in primates are presently weak. Doses used in chimpanzees are, on a gram body-weight basis, about 125-fold less than the doses used in mice, and this dosage level may in part explain the low level of response (Table 3). Mice clearly, so far, have stronger responses than chimpanzees. Coimmunization with cytokines,2*, 43 immunostimulatory sequences (C~G’S),~, 23, 48 or other adjuvants may be required to induce optimal responses. Furthermore, if means can be developed, the specific delivery of plasmids to antigen-presenting cells, particularly Langerhans’ and dendritic cells, would probably potentiate the efficacy of DNA-based immunization and might also radically reduce the large and impractical doses that are presently required. A reduction of required dosage is of considerable importance if DNA-based immunization is to be used in developing countries, because plasmids produced under good manufac- turing practices (cGMP) are presently quite expensive.
Figure 1. Attempted DNA-canarypox based immunization against hepatitis C virus (HCV). Arrows signify: (1) HCV recombinant plasmids encoding HCV capsid, HCV NS3 and HCV NS5, 0.5 mg of each by multiple i.m. injection; (2) recombinant (HCVcap-NS3) canary pox (ALVAC) boost with 4 x lo8 pfu i.m and 4 x lo8 pfu i.v.; (3) 10 CID50 HCV bk challenge, i.v.; (4) 100 CID50 HCV bk challenge, i.v.; (5) parental ALVAC, 4 x lo8 pfu i.m and i.v. A, CHIMP 214. 6, CHIMP 191. C, CHIMP 317.

1100 PRINCE & SHATA
Use of Viral Vectors for Immunization
Recombinant viral vectors are another important potential tool for immunization against HCV. To date most work has used recombinant pox viruses, particularly canarypox, fowlpox, and the attenuated, modi- fied vaccinia virus Ankara. None of these viruses grow in mammalian cells, but they can efficiently infect and express their carried nucleic acids into the cytosol. These vectors may be used as the sole immunizing agentz8 or as a booster after DNA-based immunization (a so-called prime- boost regimen).24, 32, 38 The latter was shown first by Leong et a1 to be a powerful means for enhancing the response to DNA-based immuniza- tion" and is now being applied to immunization against HIV in clinical t~ia1s.l~ Preliminary results have been reported for HCV recombinant sindbis virus, a potential live-virus vector." Other potential vectors, such as adenoviruses, which efficiently infect dendritic cells, need to be explored.
SUMMARY
Because hepatitis C virus is etiologically involved in about half the cases of the world's most common cancer, hepatocellular carcinoma, and because this virus is likely to continue to spread in most of the devel- oping world for many years, the authors believe that development of a prophylactic vaccine is imperative.
Numerous approaches are available to overcome the many impedi- ments which make the development of an HCV vaccine difficult. Such impediments include the many viral genotypes and quasispecies of HCV and the association of virions with host lipids. It is likely that overcom- ing these impediments will require a vaccine which induces a strong cell-mediated response. The most promising approach seems to be DNA- based immunization or a prime-boost regimen with DNA priming and boosting with a viral vector. Potentiation of responses with adjuvant strategies will probably be necessary.
Hepatitis C virus immunization is in an early stage of development. Given the explosive growth in the understanding of immunology, prog- ress should be rapid.
References
Abdel-Aziz F, Habib M, Mohamed MK, et al: Hepatitis C virus (HCV) infection in a communiw in the Nile Delta: Population description and HCV prevalence. Hepatol- ogy 32:11f-115,2000 Abdel-Wahab, MF, Zakaria S, Kame1 M, et al: High seroprevalence of hepatitis C infection among risk groups in Egypt. Am J Trop Med Hyg 51:563-567, 1994 Baumert TF, Vergalla J, Satoi J, et a1 Hepatitis C virus-like particles synthesized in insect cells as a potential vaccine candidate. Gastroenterology 1171397-1407, 1999

IMMUNOPROPHYLAXIS OF HEPATITIS C VIRUS INFECTION 1101
4. Bukh J, Miller RH, Purcell RH: Genetic heterogeneity of hepatitis C virus: Quasispe- cies and genotypes. Semin Liver Dis 15:41-63,1995
5. Choo Q-L, Kuo G, Ralston R, et al: Vaccination of chimpanzees against infection by the hepatitis C virus. Proc Natl Acad Sci U S A 91:12911-1298, 1994
6. Dienstag JL, Alter HJ: Non-A, non-B hepatitis: Evolving epidemiologic and clinical perspective. Semin Liver Dis 667-81, 1986
7. Diepolder HM, Zachoval R, Hoffmann RM, et al: Possible mechanism involving T- lymphocyte response to non-structural protein 3 in viral clearance in acute hepatitis C virus infection. Lancet 346:1006-1007, 1995
8. Encke J, Pulitz JZ, Geissler M, et al: Genetic immunization generates cellular and humoral immune responses against the nonstructural proteins of the hepatitis C virus in a murine model. J Immunol 161:49174923, 1998
9. Esumi M, Rikihisa T, Nishimura S, et al: Experimental vaccine activities of recombi- nant El and E2 glycoproteins and hypervariable region 1 peptides of hepatitis C virus in chimpanzees. Arch Virol 144:97>980, 1999
10. Ferrari C, Valli A, Galati L, et al: T cell responses to structural and non-structural hepatitis C virus antigen in persistent and self-limited hepatitis C virus infection. Hepatology 19:286-295, 1994
11. Filocamo G, Pacini L, Migliaccio G: Chimeric sindbis viruses dependent on the NS3 protease of hepatitis C virus. J Virol 71:1417-1427, 1997
12. Forns X, Payette PJ, Ma X, et al: Vaccination of chimpanzees with plasmid DNA encoding the hepatitis C virus (HCV) envelope E2 protein modified the infection after challenge with homologous monoclonal HCV. Hepatology 32:61&625, 2000
13. Foumillier A, Depla E, Karayiannis P, et a1 Expression of noncovalent hepatitis C virus envelope El-E2 complexes is not required for the induction of antibodies with neutralizing properties following DNA immunization. J Virol 73:7497-7504, 1999
14. Frank C, Mohamed MK, Strickland GT, et al: The role of parenteral antischistosomal therapy in the spread of hepatitis C virus in Egypt. Lancet 355:887-891, 2000
15. Gordon EJ, Bhat R, Liu Q, et a1 Immune responses to hepatitis C virus structural and nonstructural proteins induced by plasmid DNA immunizations. J Infect Dis
16. Gruener N, Gerlach TJ, Jung MC, et al: Association of hepatitis C virus-specific CD8 + T cells with viral clearance in acute hepatitis C. J Infect Dis 181:1528-1536, 2000
17. Hanke T, McMichael A: Design and construction of an experimental HIV-1 vaccine for a year-2000 clinical trial in Kenya. Nat Med 6:951-954, 2000
18. Heile JM, Fong Y-L, Rosa D, et al: Evaluation of hepatitis C virus glycoprotein E2 for vaccine design: An endoplasmic reticulum-retained recombinant protein is superior to secreted recombinant protein and DNA-based vaccine candidates. J Virol 74:6885- 6892, 2000
19. Hijikata M, Shimizu YK, Kato H, et al: Equilibrium centrifugation studies of hepatitis C virus: Evidence for circulating immune complexes. J Virol671953-1958, 1993
19a. Houghton NI: Progress and Problems Presented at the 6th International Meeting on Hepatitis C and Related Viruses, Bethesda, MD, June 6 9 , 1999
19b. Houghton M Progress and Problems Presented at the 7th International Meeting on Hepatitis C and Related Viruses, Gold Coast, Australia, December 3-7, 2000
20. Kanto T, Hayashi N, Takehara T, et al: Density analysis of hepatitis C virus particle population in the circulation of infected hosts: Implications for virus neutralization or persistence. J Hepatol 22440448, 1995
21. Kim JJ, Maguire HC Jr, Nottingham LK, et al: Coadministration of IL-12 or IL-10 expression cassettes drives immune responses towards a Thl phenotype. J Interferon Cytokine Res 18:537-547, 1998
22. Klinman DM, Bamhart KM, Conover J: CpG motifs as immune adjuvants. Vaccine
23. Krieg AM: The CpG motif Implication for clinical immunology. BioDrugs 10:341- 346,1998
24. Leong KH, Ramsey AJ, Morin MJ, et a1 Generation of enhanced immune responses by consecutive immunization with DNA and recombinant fowlpox virus. In Brown
181:42-50,2000
1719-25, 1999

1102 PRINCE & SHATA
F, Chanock R, Ginsberg H, et a1 (eds): Vaccines 95. Cold Spring Harbor, NY, Cold Spring Harbor Laboratory Press, 1996, pp 327-331
25. Lowrie DB, Whalen RG (eds): DNA Vaccines. Methods and Protocol. Totowa, NJ, Humana Press, 2000
26. Major ME, Vitviski L, Mink MA, et al: DNA-based immunization with chimeric vectors for the induction of immune responses against the hepatitis C virus nucleocap- sid. J Virol 69:5798-5805, 1995
27. Nakano I, Maertens G, Major ME, et a1 Immunization with plasmid DNA encoding hepatitis C virus envelope E2 antigenic domains induces antibodies whose immune reactivity is linked to the injection mode. J Virol 71:7101-7109, 1997
28. Ockenhouse CF, Sun P-F, Lanar DE, et al: Phase I/IIa safety, immunogenicity, and efficacy trial of NYVAC-Pf7, a pox-vectored, multiantigen, multistage vaccine candi- date for Plasmodium falciparum malaria. J Infect Dis 1771664-1673, 1998
29. Ogata N, Alter HJ, Miller RH, et al: Nucleotide sequence and mutation rate of the H strain of hepatitis C virus. Proc Natl Acad Sci U S A 88:3392-3396, 1991
30. Okamoto H, Mishiro S, Tokita H, et al: Superinfection of chimpanzees carrying hepatitis C virus of genotype II/lb with that of genotype III/2a or I/la. Hepatology
31. Okuda K, Fujimoto I, Hanai A, et al: Changing incidence of hepatocellular carcinoma in Japan. Cancer Res 474967, 1987
32. Pancholi P, Liu Q, Tricoche N, et al: DNA prime-canarypox boost with polycistronic hepatitis C virus (HCV) genes generates potent immune responses to HCV structural and nonstructural proteins. J Infect Dis 182:18-27, 2000
33. Prince AM: Challenges for development of hepatitis C virus vaccines. FEMS Microbiol Rev 14273-278,1994
34. Prince AM: Prevention of liver cancer and cirrhosis by vaccines. Clin Lab Med
35. Prince AM, Brotman B, Huima T, et al: Immunity in hepatitis C infection. J Infect Dis 165:438-443, 1992
36. Prince AM, Brotman B, Lee D-H, et al: Significance of the anti-E2 response in self- limited and chronic hepatitis C virus infections in chimpanzees and in humans. J Infect Dis 180:987-991, 1999
37. Prince AM, Huima-Byron T, Parker TS, et al: Visualization of hepatitis C virions and putative defective interfering particles isolated from low-density lipoproteins. J Virol Hepatitis 3:ll-17, 1996
38. Robinson HL, Montefiori DC, Johnson RP, et a1 Neutralizing antibody-independent containment of immunodeficiency virus challenges by DNA priming and recombinant pox virus booster immunization. Nat Med 5:526-534, 1999
39. Saito T, Sherman GJ, Kurokohchi K, et al: Plasmid DNA-based immunization for hepatitis C virus structural proteins: Immune responses in mice. Gastroenterology
40. Sakamoto M, Hirohashi S, Tsuda E, et al: Increasing incidence of hepatocellular carcinoma possibly associated with non-A, non-B hepatitis in Japan, disclosed by hepatitis B virus DNA analysis of surgically resected cases. Cancer Res 48:7294, 1988
41. Shirai M, Chen M, Arichi T, et al: Use of intrinsic and extrinsic helper epitopes for in vivo induction of anti-hepatitis C virus cytotoxic T lymphocytes (CTL) with CTL epitope peptide vaccines. J Infect Dis 173:24-31, 1996
42. Simmonds P, McOmish F, Yap PL, et al: Sequence variability in the 5 prime non- coding region of hepatitis C virus: Identification of new virus type and restrictions on sequence diversity. J Gen Virol 74:661-668, 1993
43. Sin JI, Kim JJ, Arnold RL, et al: IL-12 gene as a DNA vaccine adjuvant in a herpes mouse model: IL-12 enhances Thl-type CD4 + T cell-mediated protective immunity against herpes simplex virus-2 challenge. J Immunol 162:2912-2921, 1999.
44. Thomssen R, Bonk S, Propfe C, et al: Association of hepatitis C virus in human sera with P-lipoprotein. Med Microb Immunol (Bul) "293-300, 1992
45. Tokushige K, Watika T, Pachuk C, et al: Expression and immune response to hepatitis C virus core DNA-based vaccine constructs. Hepatology 2434-20, 1996
20~1131-1136, 1994
16~493-505, 1996
112:1321-1330, 1997

IMMUNOPROFHYLAXIS OF HEPATITIS C VIRUS INFECTION 1103
46. Ulmer JB, Donnelly JJ, Parker SE, et a1 Heterologous protection against influenza by injection of DNA encoding a viral protein. Science 259:1745-1749, 1993
47. Wang Y-F, Brotman B, Andrus L, et al: Immune responses to epitopes of hepatitis C virus (HCV) structural proteins in HCV-infected humans and chimpanzees. J Infect Dis 173:808-821, 1996
48. Weeratna RD, McCluskie MJ, Xu Y, et al: CpG DNA induces stronger immune responses with less toxicity than other adjuvants. Vaccine 18:1755-1762, 2000
49. Weiner AJ, Brauer M, Rosenblatt MJ, et al: Variable and hypervariable domains are found in the regions of HCV corresponding to the flavivirus envelope and NSI proteins and the pestivirus envelope glycoproteins. Virology 180:842-848, 1991
50. Wyatt CA, Andrus L, Brotman B, et al: Immunity in chimpanzees chronically infected with hepatitis C virus: Role of minor quasispecies in reinfection. J Virol 72:1725- 1730, 1998
Address reprint requests to Alfred M. Prince, MD
The Lindsley F. Kimball Research Institute of the New York Blood Center 310 East 67th St
New York, NY 10021
e-mail: [email protected]





























