Embed Size (px)
Citation preview

ARTICLE IN PRESS
0142-9612/$ - se
doi:10.1016/j.bi
�CorrespondUniversity, Sha
fax: +86754 29
E-mail addr1Both author
Biomaterials 28 (2007) 3896–3903
www.elsevier.com/locate/biomaterials
In vitro effect of oligo-hydroxyalkanoates on the growth of mousefibroblast cell line L929
Jie Suna,1, Zhongwei Daia,1, Yan Zhaob, Guo-Qiang Chena,b,�
aMultidisciplinary Research Center, Shantou University, Shantou 515063, Guangdong, ChinabProtein Science Laboratory of Ministry of Science, Dept of Biological Science and Biotechnology, Tsinghua University, Beijing 100084, China
Received 12 January 2007; accepted 21 May 2007
Available online 26 May 2007
Abstract
The cellular responses to polyhydroxyalkanoates (PHA) degradation products oligo-hydroxyalkanoates (OHAs) are very important
factors that control the biocompatibility of these polymers when they are used in tissue-engineering applications. In this study, oligo
(3-hydroxybutyrate) (OHB, Mn 2000), oligo(3-hydroxybutyrate-co-4-hydroxybutyrate) (O3HB4HB, Mn 2100, 6mol% 4HB), oligo
(3-hydroxybutyrate-co-3-hydroxyhexanoate) (OHBHHx, Mn 2800, 12mol% 3HHx) and medium-chain-length oligo(3-hydroxyalkanoates)
(OmclHAs, Mn 2300, 2mol% 3-hydroxyhexanoate (3HHx), 25mol% 3-hydroxyoctanoate, 71mol% 3-hydroxydecanoate and 3mol%
3-hydroxydodecanoate) were prepared as insoluble particles in aqueous solution. Liposomes were employed to encapsulate OHAs and
facilitate their transfer into the cytosol. The methylthiazol tetrazolium (MTT) assay, a cell apoptosis study and the flow cytometry were
used to evaluate cell viability of mouse fibroblast L929. OHAs in concentration lower than 20mg/l did not significantly affect cell
viability, while OHAs over 40mg/l reduced cell viability with more cell apoptosis, more cell death, delayed cell cycle and reduced cell
proliferation. The cytotoxicity of OHAs decreased with increasing OHAs side chain length. It is proposed that medium-chain-length
OHAs containing PHA, such as poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) (PHBHHx) and medium-chain-length PHA Poly(3-
hydroxyalkanoates) (mcl PHA), are preferred for development and application as PHA-based tissue-engineering biomaterials.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: PHB; Polyhydroxyalkanoates; Oligo-hydroxyalkanoates; Tissue engineering; Biomaterials; Liposome
1. Introduction
Polyhydroxyalkanoates (PHA) are polyesters producedby microorganisms under unbalanced growth conditions[1]. PHA have attracted increasing interest as tissue-engineering materials due to their biodegradability andbiocompatibility, together with other adjustable mechan-ical properties based on monomer structures and contents[2–4]. The applications of PHA in medicine are now beingexpanded to include wound management, vascular systemdevices, bone regeneration and drug delivery system [5,6].
Some studies have attributed the PHA biocompatibilityto their physical properties such as porosity, surface
e front matter r 2007 Elsevier Ltd. All rights reserved.
omaterials.2007.05.011
ing author. Multidisciplinary Research Center, Shantou
ntou 515063, Guangdong, China. Tel.: +86 754 2901186;
01175.
ess: [email protected] (G.-Q. Chen).
s contributed equally to this research.
hydrophilicity, surface energy [7–10] and degradationproducts of the polymers [11,12]. It has been reported that3-hydroxybutyrate (3HB), which is the most commondegradation product of PHA, plays an important role inbiocompatibility [11,12]. It has been indicated that in vivo
nutritional or therapeutic compositions are sustained byproviding a source of ketone bodies in the form of linear orcyclic oligomers and/or derivatives of 3HB to increaseketone body levels in the blood of mammals [13]. Previousresults also showed that 3HB could reduce the death of cellline L929 when cultured at high density [11]. There arestudies demonstrating that 3HB could overcome thedefects in mitochondrial energy generation and providetherapeutic protection against many diseases [14–16].However, with the exception of oligo(3-hydroxybutyrate)(OHB) [17–20], the preparation and cellular responses ofoligo-hydroxyalkanoates (OHA), which are all degradationproducts of the PHA implants, have not been studied.

ARTICLE IN PRESSJ. Sun et al. / Biomaterials 28 (2007) 3896–3903 3897
Reusch [17] reported that OHB acted not only as an inertstorage polymer in certain bacteria but also as aubiquitous, interactive, solvating biopolymer involved inimportant physiological functions. She went on to find thatlow-molecular-weight poly(3-hydroxybutyrate) (PHB)(OHB) integrating with other macromolecules are widelydistributed in nearly all representative organisms [18],OHB combined with some proteins to form complex PHB(cPHB), cPHB and CaPPi could form another complexwhich functions as an ion channel in genetically competentEscherichia coli [19]. At least two different super-molecular structures, one for the non-ion-selective trans-port system which is affected by the OHB alone, andanother for the ion-selective, OHB/CaPPi complex channelwere found [20].
In this study, OHAs including OHB, oligo(3-hydroxy-butyrate-co-4-hydroxybutyrate) (O3HB4HB), oligo(3-hy-droxybutyrate-co-3-hydroxyhexanoate) (OHBHHx) andmedium-chain-length oligo(3-hydroxyalkanoates) (Omcl-HAs) were prepared and encapsulated in liposomes. Theirin vitro effects on cell growth and the cell cycle of mousefibroblast L929 were investigated.
2. Experiment
2.1. Materials
PHB (Mw 400,000), poly(3-hydroxybutyrate-co-4-hydroxybutyrate)
(P3HB4HB, 6mol% 4HB, Mw 800,000) and poly(3-hydroxybutyrate-co-
3-hydroxyhexanoate) (PHBHHx, Mw 400,000) consisting of 12mol%
3-hydroxyhexanoate (3HHx), were donated by Nantian Co., Ltd. (Jiangsu,
China), Guoyun Biotech. Co., Ltd. (Tianjin, China) and Lianyi Biotech
Co., Ltd. (Guangdong, China), respectively. Medium-chain-length poly
(3-hydroxyalkanoates) (mcl PHA) was produced by Pseudomonas stutzeri
1317 as described by Xi et al. [21]. The mcl PHA consists of 1.6mol%
3HHx, 24.6mol% 3-hydroxyoctanoate, 71.2mol% 3-hydroxydecanoate
and 2.6mol% 3-hydroxydodecanoate. L-a-phosphatidylcholine (PC) fromsoybean and cholesterol were purchased from Sigma (USA) and Biosharp
(USA), respectively. Dubecco’s Modified Eagle Medium (DMEM) was
purchased from Gibco-BRL (Gaithersberg, MD), fetal bovine serum
(FBS) from Sijiqing Co., Ltd. (Zhejiang, China), Annxin V-FITC/PI
apoptosis kit from Biosea Co., Ltd. (Beijing, China). All reagents were of
analytical grade.
2.2. Preparation and characterization of oligomers OHAs
OHB, O3HB4HB, OHBHHx and OmclHA were obtained by
methanolysis of corresponding polymers [22], without detectable toxic
crotonic acid formation which is toxic to cells [23]. Particles of these
oligomers were formed by their precipitation in cold methanol from OHAs
chloroform solutions.
The OHAs oligomers were characterized using nuclear magnetic
resonance spectroscopy (NMR, Brucker 400, Germany). The precipitated
OHAs particle sizes were studied using scanning electron microscope
(SEM, JEOL JSM-6360LA, Japan). The thermodynamics characters of
the oligomers were determined by differential scanning calorimetry (DSC,
TA-Q100, USA).
2.3. Preparations of liposomes encapsulating OHAs
OHAs containing liposomes were prepared using sonication [24].
Briefly, 5mg OHAs, 250mg PC (from soybean) and 100mg cholesterol
were dissolved in 20ml chloroform and carefully dried to produce a thin
film. The film was then hydrated using 20ml N-(2-hydroxyethyl)
piperazine-N-ethane-sulfonic acid (HEPES, 10mM, pH 7.4) and was
followed by sonication in ice bath for 60min and centrifugation at
10,000 g. The OHAs-liposome suspensions were filtered using a filter paper
with a diameter of 0.45mm to obtain liposomes with narrow size
distribution for further experiments. Fresh liposomes were prepared each
time. The diameters of liposomes were determined by transmission
electron microscope (TEM, Hitachi 300, Japan).
2.4. Cell culture of mouse fibroblast cell line L929
The mouse fibroblast cell line L929 (Chinese Academy of Preventive
Medical Sciences, Beijing, China) was cultured in DMEM medium
supplemented with 10% FBS, 100U/ml penicillin and 100mg/ml
streptomycin. Cells were incubated at 37 1C in a 5% CO2 incubator and
the medium was changed every 2 days. When the cells reached the stage of
confluence, they were harvested using 0.25% trypsin, followed by addition
of fresh culture medium to create a new single cell suspension for further
inoculation [25].
2.5. Cell survival assay
Cells in an initial concentration of 5� 103/well were seeded in 96-well
plate in DMEM supplemented with 10% FBS. After 24 h, the culture
medium was changed into DMEM medium plus 10% FBS, and OHAs
particles or OHAs-liposomes of various concentrations were added into
the culture medium, respectively. Cells added with no oligomer and blank
liposomes (prepared only with PC and cholesterol) acted as control and
blank, respectively. After inoculation for 24 and 48 h, methylthiazol
tetrazolium (MTT) assay [26] was performed to evaluate cell activity.
Briefly, the culture medium was removed, and the cultures were washed
with PBS twice. About 100ml serum-free DMEM medium and 10 ml MTT
solution (5mg/ml in PBS) were added to each sample, followed by
incubation at 37 1C for 4 h to allow MTT formazan formation. The
medium andMTT were replaced by 100ml of dimethylsulfoxide to dissolve
the formazan crystals. After 30min, the optical density (OD) at 550 nm
was determined using a microplate reader (ThermoLabsystems, Multiskan
Mk3, Finland). The ratios of OD550 from each parallel experimental
group (n ¼ 6) to the OD550 of control group were used to assess the cell
activities.
2.6. Flow cytometry analysis
OHAs-liposomes containing OHB, O3HB4HB, OHBHHx and Omcl-
HAs were added to L929. Flow cytometry analysis (Argon excited wave
488nm Beckman Coulter Epics XL, USA) using apoptosis kit Annexin
V-FITC (Beijing Baosai biotechnologic Co. Ltd.) was performed to
evaluate the effect of OHAs on cell L929 apoptosis [7]. Briefly, cells in an
initial concentration of 1� 104 cm�2 were seeded in 6-well plates with
slides in DMEM medium supplemented with 10% FBS for 24 h. Then the
culture medium was replaced by DMEM medium. After 24 h, the blank
liposomes and OHAs-liposomes were all added to the culture medium.
The concentration of the blank liposomes was 40mg/l and the content of
the OHAs in the medium (not in OHAs-liposomes) was 40mg/l. After
cultivation for 24 and 48 h, respectively, the cells were harvested with
0.25% trypsin and concentrated. Cells were resuspended with 200mlbinding buffer and then incubated with 10 ml Annexin V-FITC and 5 ml PIfor 15min at room temperature. Subsequently, 300ml binding buffer was
added again into the solution. Samples were detected using flow cytometry
in an hour.
DNA contents were also measured to evaluate the effect of OHAs on
cell apoptosis [7]. Conditions of cell culture and OHAs-liposomes
treatment were the same as in the apoptosis assay described above. After
24 and 48 h OHAs treatment, respectively, the cells were harvested and
washed with cold PBS twice, followed by fixing in 75% ethanol at 4 1C

ARTICLE IN PRESSJ. Sun et al. / Biomaterials 28 (2007) 3896–39033898
overnight. Fixed cells were washed twice with PBS and treated with
10mg/ml DNase-free RNase at 37 1C for 30min. DNA was stained using
10mg/ml PI at room temperature for 30min in the dark [7]. Samples were
analyzed using a flow cytometery with excitation at 488 nm and emission
at 620 nm. In each experiment, 10,000 cells were recorded. The
proportions of cells in the G0/G1, S and G2/M were determined using
MultiCycle software (Beckman Coulter Epics XL, USA). The prolifera-
tion index (PI) was calculated using the following equation:
Proliferation index ð%Þ ¼SþG2=M
G0=G1þSþG2=M� 100% ½27�.
Fig. 1. Transmission electron microscopy and scanning electron micro-
scopy study of OHAs particles and OHAs-liposomes. (a) Particles were
obtained by OHAs precipitation in cold methanol, washed in distilled
water for five times, dried in vacuum at 40 1C for 48 h. The dried particles
were coated with gold in a sputtering device for SEM observation.
OmclHAs was oily, not suitable for particles preparation. Since all OHAs
particles appeared similar, only one representative picture of OHB is
shown. (b) OHAs-liposomes were prepared by sonication and diluted in
PBS. TEM images were taken after the samples were dried at room
temperature. Since all OHAs-liposomes appeared similar, only one
representative picture of OHB-liposomes is shown.
2.7. Statistical analysis
All data were presented as the mean value7standard deviation of each
group. Variation between groups was evaluated using the Students t-test,
with a confidence level of 95% (po0.05) considered statistically
significance and 99% (po0.01) considered very significant.
3. Results
3.1. Characterization of OHAs and OHAs-liposomes
The PHAs particles were prepared through methanolprecipitation. All OHAs particles were crystalline exceptOmclHAs, which was oily and amorphous. Results of SEMindicated that the particles had irregular shape withdiameters of 1–10 mm (Fig. 1a). OHAs can be incorporateddirectly into the membranes of pure PC vesicles throughtheir inclusion in the organic solvent, along with phospho-lipids during vesicle preparation. A OHAs/PC ratio (5mg:250mg PC) was chosen to prepare liposomes after PCcapabilities were examined. After centrifugation, theliposomes containing OHAs were collected. Gas chroma-tography (GC) analysis was performed to evaluate contentsof OHAs in the liposomes. Results of GC indicated that allthe OHAs were encapsulated into the liposomes (data notshown). OHAs-liposomes and blank liposomes showed thesame diameter (Fig. 1b). TEM showed that the OHAs-liposomes had oval shapes with sizes of 200–500 nm(Fig. 1b). Diameters of liposomes remained similarregardless of what OHAs were encapsulated.
NMR was used to characterize OHAs. No alkenyl bondwas found in all OHAs (data not shown), indicating thehydrolysis process did not generate unsaturated bonds thatmay be toxic for cells. Number average molecular weights(Mn) of all OHAs were found to be in the range between2000 and 3000 based on gel permeation chromatography(Table 1). DSC studies showed that Tg, Tm and DH ofOHAs decreased with increasing monomer chain length(Table 1).
3.2. Viability of mouse fibroblast L929
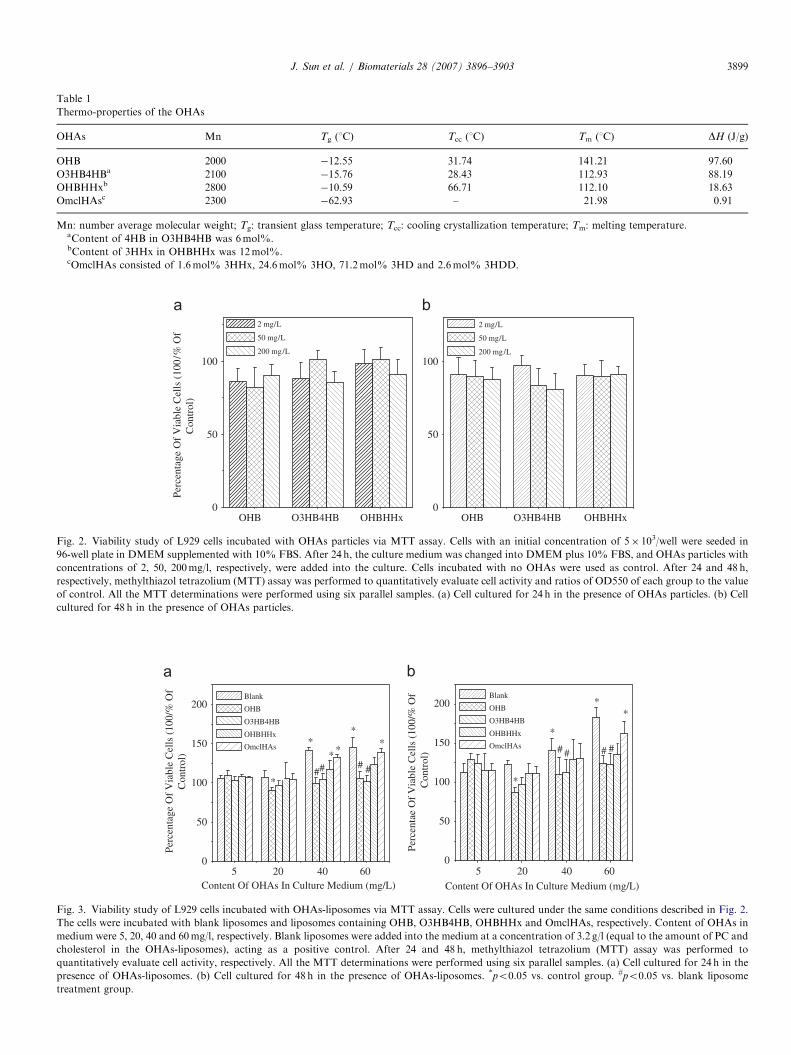
MTT assay was performed to evaluate cell viabilityaffected by OHAs and OHAs encapsulating liposomes.Results of MTT assay indicated that all OHAs particleswith concentration lower than 200mg/l did not signifi-
cantly affect cell viability (Fig. 2). In comparison, OHAs-liposomes affected cell viability differently (Fig. 3). Cellviability was slightly reduced in the presence of OHB- andO3HB4HB-liposomes, with OHB-liposomes showing asignificant effect at a content of 20mg/l OHB in themedium (NOT liposomes) compared with the controlgroup. At higher content (40–60mg/l OHAs in themedium), inhibitions of OHB and O3HB4HB were notsignificant compared with the control group. Interestingly,although blank liposomes, OHBHHx and OmclHAs allhad a significant growth promotion effect on the cells(po0.05) in concentration and OHAs content of 40–60mg/l,respectively, blank liposomes showed the strongest effect
ARTICLE IN PRESS
Table 1
Thermo-properties of the OHAs
OHAs Mn Tg (1C) Tcc (1C) Tm (1C) DH (J/g)
OHB 2000 �12.55 31.74 141.21 97.60
O3HB4HBa 2100 �15.76 28.43 112.93 88.19
OHBHHxb 2800 �10.59 66.71 112.10 18.63
OmclHAsc 2300 �62.93 – 21.98 0.91
Mn: number average molecular weight; Tg: transient glass temperature; Tcc: cooling crystallization temperature; Tm: melting temperature.aContent of 4HB in O3HB4HB was 6mol%.bContent of 3HHx in OHBHHx was 12mol%.cOmclHAs consisted of 1.6mol% 3HHx, 24.6mol% 3HO, 71.2mol% 3HD and 2.6mol% 3HDD.
OHB O3HB4HB OHBHHx0
50
100
2 mg/L
50 mg/L
200 mg/L
2 mg/L
50 mg/L
200 mg/L
OHB O3HB4HB OHBHHx0
50
100
Perc
enta
ge O
f V
iabl
e C
ells
(10
0/%
Of
Con
trol
)
Fig. 2. Viability study of L929 cells incubated with OHAs particles via MTT assay. Cells with an initial concentration of 5� 103/well were seeded in
96-well plate in DMEM supplemented with 10% FBS. After 24 h, the culture medium was changed into DMEM plus 10% FBS, and OHAs particles with
concentrations of 2, 50, 200mg/l, respectively, were added into the culture. Cells incubated with no OHAs were used as control. After 24 and 48 h,
respectively, methylthiazol tetrazolium (MTT) assay was performed to quantitatively evaluate cell activity and ratios of OD550 of each group to the value
of control. All the MTT determinations were performed using six parallel samples. (a) Cell cultured for 24 h in the presence of OHAs particles. (b) Cell
cultured for 48 h in the presence of OHAs particles.
5 20 400
50
100
150
200 Blank
OHB
O3HB4HB
OHBHHx
##
#
60
Perc
enta
ge O
f V
iabl
e C
ells
(10
0/%
Of
C
ontr
ol)
Content Of OHAs In Culture Medium (mg/L) 5 20 40
0
50
100
150
200
60
Perc
enta
e O
f V
iabl
e C
ells
(10
0/%
Of
C
ontr
ol)
Content Of OHAs In Culture Medium (mg/L)
*
***
**#
OmclHAs
Blank
OHB
O3HB4HB
OHBHHx
OmclHAs
*
# #
*
**
##
Fig. 3. Viability study of L929 cells incubated with OHAs-liposomes via MTT assay. Cells were cultured under the same conditions described in Fig. 2.
The cells were incubated with blank liposomes and liposomes containing OHB, O3HB4HB, OHBHHx and OmclHAs, respectively. Content of OHAs in
medium were 5, 20, 40 and 60mg/l, respectively. Blank liposomes were added into the medium at a concentration of 3.2 g/l (equal to the amount of PC and
cholesterol in the OHAs-liposomes), acting as a positive control. After 24 and 48 h, methylthiazol tetrazolium (MTT) assay was performed to
quantitatively evaluate cell activity, respectively. All the MTT determinations were performed using six parallel samples. (a) Cell cultured for 24 h in the
presence of OHAs-liposomes. (b) Cell cultured for 48 h in the presence of OHAs-liposomes. *po0.05 vs. control group. #po0.05 vs. blank liposome
treatment group.
J. Sun et al. / Biomaterials 28 (2007) 3896–3903 3899
ARTICLE IN PRESSJ. Sun et al. / Biomaterials 28 (2007) 3896–39033900
which did not fluctuate after 24 and 48 h (Figs. 2a, b, 3aand b).
Normally, L929 cells showed a typical spindle morphol-ogy, which remained unchanged in the presence of blankliposomes and low content of OHAs in OHAs-liposomes(5mg/l). The cells turned round and became detachedfrom the culture dishes when exposed to 40–60mg/l ofOHB- and O3HB4HB-liposomes, respectively. However,OHBHHx- and OmclHAs-liposomes treatment did notlead to this morphological change, indicating the less toxiceffect of longer chain length OHAs to the cells (photos notshown).
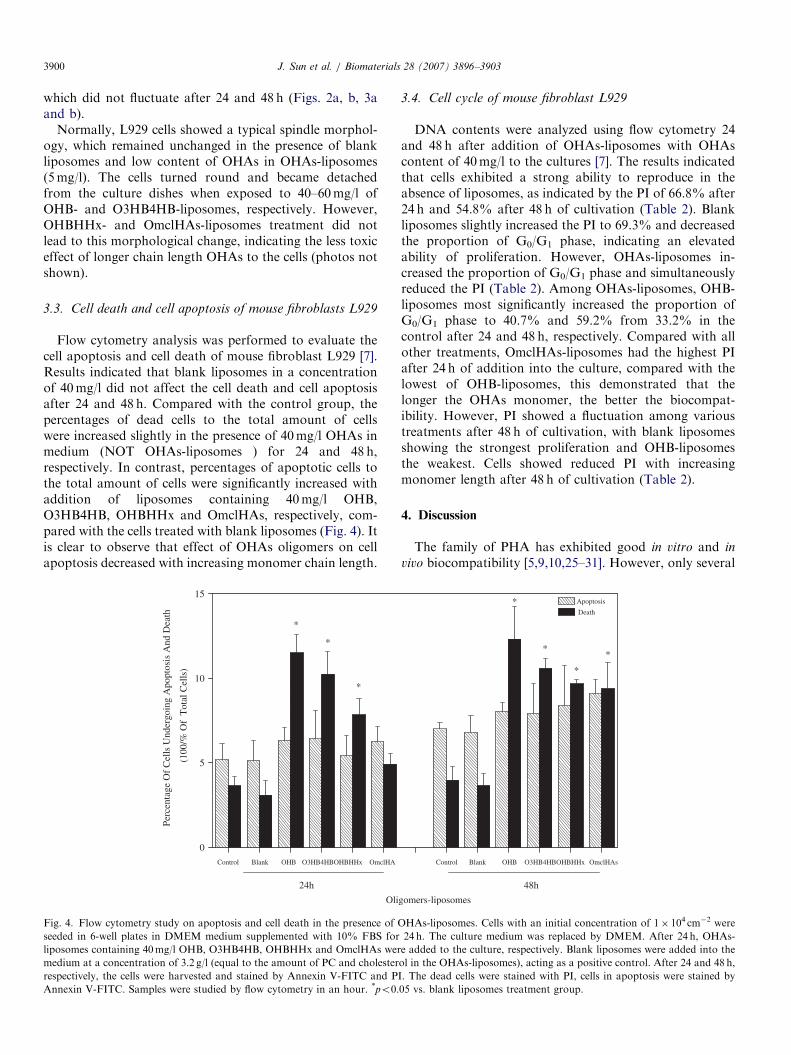
3.3. Cell death and cell apoptosis of mouse fibroblasts L929
Flow cytometry analysis was performed to evaluate thecell apoptosis and cell death of mouse fibroblast L929 [7].Results indicated that blank liposomes in a concentrationof 40mg/l did not affect the cell death and cell apoptosisafter 24 and 48 h. Compared with the control group, thepercentages of dead cells to the total amount of cellswere increased slightly in the presence of 40mg/l OHAs inmedium (NOT OHAs-liposomes ) for 24 and 48 h,respectively. In contrast, percentages of apoptotic cells tothe total amount of cells were significantly increased withaddition of liposomes containing 40mg/l OHB,O3HB4HB, OHBHHx and OmclHAs, respectively, com-pared with the cells treated with blank liposomes (Fig. 4). Itis clear to observe that effect of OHAs oligomers on cellapoptosis decreased with increasing monomer chain length.
Control Blank O3HB4HBOHBHHx OmclHA
0
5
10
15
Oli
24h
*
*
*
Perc
enta
ge O
f C
ells
Und
ergo
ing
Apo
ptos
is A
nd D
eath
(100
/% O
f T
otal
Cel
ls)
OHB
Fig. 4. Flow cytometry study on apoptosis and cell death in the presence of O
seeded in 6-well plates in DMEM medium supplemented with 10% FBS for
liposomes containing 40mg/l OHB, O3HB4HB, OHBHHx and OmclHAs wer
medium at a concentration of 3.2 g/l (equal to the amount of PC and cholester
respectively, the cells were harvested and stained by Annexin V-FITC and P
Annexin V-FITC. Samples were studied by flow cytometry in an hour. *po0.
3.4. Cell cycle of mouse fibroblast L929
DNA contents were analyzed using flow cytometry 24and 48 h after addition of OHAs-liposomes with OHAscontent of 40mg/l to the cultures [7]. The results indicatedthat cells exhibited a strong ability to reproduce in theabsence of liposomes, as indicated by the PI of 66.8% after24 h and 54.8% after 48 h of cultivation (Table 2). Blankliposomes slightly increased the PI to 69.3% and decreasedthe proportion of G0/G1 phase, indicating an elevatedability of proliferation. However, OHAs-liposomes in-creased the proportion of G0/G1 phase and simultaneouslyreduced the PI (Table 2). Among OHAs-liposomes, OHB-liposomes most significantly increased the proportion ofG0/G1 phase to 40.7% and 59.2% from 33.2% in thecontrol after 24 and 48 h, respectively. Compared with allother treatments, OmclHAs-liposomes had the highest PIafter 24 h of addition into the culture, compared with thelowest of OHB-liposomes, this demonstrated that thelonger the OHAs monomer, the better the biocompat-ibility. However, PI showed a fluctuation among varioustreatments after 48 h of cultivation, with blank liposomesshowing the strongest proliferation and OHB-liposomesthe weakest. Cells showed reduced PI with increasingmonomer length after 48 h of cultivation (Table 2).
4. Discussion
The family of PHA has exhibited good in vitro and in
vivo biocompatibility [5,9,10,25–31]. However, only several
Control Blank OHB O3HB4HBOHBHHx
gomers-liposomes
Apoptosis
Death
48h
*
**
*
OmclHAs
HAs-liposomes. Cells with an initial concentration of 1� 104 cm�2 were
24 h. The culture medium was replaced by DMEM. After 24 h, OHAs-
e added to the culture, respectively. Blank liposomes were added into the
ol in the OHAs-liposomes), acting as a positive control. After 24 and 48 h,
I. The dead cells were stained with PI, cells in apoptosis were stained by
05 vs. blank liposomes treatment group.

ARTICLE IN PRESS
Table 2
Effects of OHAs-liposomes on cell cycle of mouse fibroblast L929
Group 24 h 48 h
G0/G1 (%) PI (%) G0/G1 (%) PI (%)
Control 33.271.2 66.871.2 45.270.8 54.870.8
Blank liposome 30.771.6 69.371.6 43.071.1 57.071.1
OHB-liposomes 40.772.1� 59.372.1� 59.271.9� 40.871.9�
O3HB4HB-liposomes 40.371.9� 59.771.9� 45.572.8 54.572.8
OHBHHx-liposomes 37.672.7 62.472.7 50.171.4 49.971.4
OmclHAs-liposomes 33.870.9 66.270.9 51.573.6 48.573.6
The cells were cultured under the same conditions as described in Fig. 4. After 24 and 48 h of OHAs incubation, respectively, cells were harvested and
washed with cold PBS twice, then fixed in 75% ethanol at 4 1C overnight. Fixed cells were washed twice with PBS and treated with 10mg/ml DNase-free
RNase at 37 1C for 30min. DNA was stained using 10 mg/ml PI at room temperature for 30min in the dark. Samples were analyzed using a flow
cytometery with excitation at 488 nm and emission at 620 nm. In each experiment, 10,000 cells were recorded
Proliferation Index ðPIÞ ð%Þ ¼SþG2=M
G0=G1þSþG2=M� 100%
�po0.05 vs. blank liposomes treatment group.
J. Sun et al. / Biomaterials 28 (2007) 3896–3903 3901
PHA including PHB, P3HB4HB, poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV), PHBHHx and mcl PHAare available in sufficient quantity for study so far [5] and ithas been reported that PHBV, PHBHHx and mcl PHA hadbetter biocompatibility than PHB did [5].
Biocompatibility for tissue engineering not only requiresbiocompatibility from implant polymers themselves butalso from the degradation products of the polymersincluding oligomers and monomers. Cellular behaviorsaffected by the degradation products must be consideredfor a comprehensive biocompatibility evaluation of theimplant polymers. 3HB, the most common PHA degrada-tion product, had reported to have various therapeuticeffects both in vitro and in vivo [7,11,14–16]. It was alsodemonstrated that dimer and trimer of 3HB could beconverted rapidly to monomers in rat and human tissues asan energy substrate for injured patients [32].
Although phagocytosis of PHB fragments has beenreported with fibroblasts, this may lead to cell death. Thiswas not the case with the present study.
OHB, O3HB4HB, OHBHHx and OmclHAs in the formof particles were found to have no significant effect on cellviability (Fig. 2). Although there was report that phago-cytosis of PHB polymer fragments did happen withfibroblast, this may lead to cell death [33], it was not thecase with our studies here. The different results may be dueto the difference on PHA molecular weight and morphol-ogy of the OHAs [33,34].
OHAs were amphiphilic molecules containing hydro-phobic methyl groups alternated with hydrophilic estercarbonyl oxygens along the polyester chain [35]. Thus,OHAs may be able to across the cell phospholipid bilayermembranes and affect cellular growth behaviors. To betterimitate the in vivo conditions, liposome encapsulation wasapplied to transport extracellular OHAs to the cells.Liposomes composed of phospholipid bilayer can trans-port foreign substances to the target cells by fusion andendocytosis processes [36,37]. With the help of liposomes,
OHAs encapsulated in liposomes would be taken up by thecells and be released into the cytosols [38,39]. It isobservable that the blank liposomes could elevate cellviability at concentrations of 40–60mg/l, which was notsurprising as PC and cholesterol-containing liposomes arethe basis of the cell membrane. The liposomes containingOHB, O3HB4HB, OHBHHx and OmclHAs seemed tohave less positive effect on cell viability of mouse fibroblastL929 compared with the cells grown in the presence ofblank liposomes, respectively (Fig. 3). It is remarkable thatthe inhibitory effect of OHAs on cell growth was decreasedwith increasing OHAs chain length.The cells incubated with OHAs-liposomes showed an
elevated apoptosis level, along with increased percentage ofdead cells. Noticeably, the promotion on cell apoptosis andcell death decreased significantly as OHAs monomer chainlength increased (Fig. 4), this result agrees well with the cellviability assay (Fig. 3).Cell cycle was also found to be influenced by OHAs.
A checkpoint exists between G1 and S phases which controlthe passage of eukaryotic cells into state of DNAduplication [40]. If environments permit, cells will crossthe point and start DNA synthesis. Otherwise, cells will beblocked at G1 phase. It was clearly observed thatpercentages of cells in G1 phase (G0/G1%) fluctuated inthe presence of OHAs, especially for the OHB-liposomes,which significantly increased the percentage of cells in G1
phase and decreased the PI% compared with cells treatedwith blank liposomes (Table 2). Thus, cell cycle of L929was postponed and cell proliferation was inhibited byOHB-liposomes. Such phenomenon was in good agreementwith the cell viability assay (Fig. 3). It could also explainthe cell damage and necrotic cell death induced by OHB ifmore cells could not pass the G1/S checkpoint (Fig. 4).It is obvious that the extents of inhibition on cell growth
induced by OHAs were different. OHAs with shortmonomer chain length, including OHB and O3HB4HBhad stronger inhibition than OHAs with longer monomer

ARTICLE IN PRESSJ. Sun et al. / Biomaterials 28 (2007) 3896–39033902
chain length, including OHBHHx and OmclHAs. Suchphenomena could be explained based on the physical andthermodynamic properties of OHAs (Table 1). Comparedwith OHB and O3HB4HB, OHBHHx and OmclHAs havelonger pendant side chains, which allow the oligomers to bemore flexible [2,41] (Table 1) and accessible to thebiomolecules in the cells, as well as impart the oligomersfaster degradation rate [5,42]. In comparison, OHB andO3HB4HB had higher enthalpy and crystallinity thanOHBHHx and OmclHAs (Table 1), this crystallinity notonly led to rigidness and brittleness of the chains [39,43],but also made the oligomers hard to be degraded.Therefore, it could be reasonably explained that OHBand O3HB4HB had negative effect on cell viability (Fig. 3),they promoted cell death and cell apoptosis, they alsopostponed cell cycle (Fig. 4, Table 2).
Besides the chain length of OHAs, the amount of OHAsadded to the cells affected the cell behavior. It is noticeablethat the highest amount of OHAs in medium was 60mg/l,much lower than around 200mg/l for the OHAs particles.However, the inhibition induced by OHAs-liposomes wasmore obvious than OHAs particles (Figs. 2 and 3). Thiscan be explained as limited OHAs particles were phagocy-tosed by fibroblasts L929, others exerted their effectextracellularly. On the contrary, OHAs encapsulated inliposomes could be more easily taken up, passed thecytosolic membrane and be released into the cytosol.
To be successfully applied as tissue-engineering bioma-terials, PHA must exhibit biocompatibility and be de-graded to release non-toxic degradation products [2].In vitro results of lower toxicity of longer-chain OHAscompared with the short-chain OHB in this studydemonstrated better biocompatibility of PHBV, PHBHHxand OmclHAs compared with that of PHB.
5. Conclusion
It was found that OHAs particles negatively affected cellviability. Liposomes containing OHAs (OHAs-liposomes)generally had positive effects on cell viability depending ontheir side chain length. OHAs-liposomes affected cellapoptosis and cell death and they delayed cell cycle. Thecytotoxicity of OHAs decreased with increasing OHAs sidechain length. Based on the better biocompatibility ofmedium-chain-length OHAs, it is proposed that medium-chain-length OHAs containing PHA, such as PHBHHxand mcl PHA, are preferred for development and applica-tion as PHA-based tissue-engineering biomaterials.
Acknowledgment
The research was supported by Li Ka Sheng Foundationand National High Tech 863 Grant (Project No.2006AA02Z242). Thanks to Professor Cai Yingmu in LiKa Sheng Hospital attached to Medical School of ShantouUniversity for discussion in flow cytometry study.
References
[1] Doi Y, Steinbuchel A. Biopolymers. Weinheim, Germany: Wiley-
VCH; 2002.
[2] Williams SF, Martin DP, Horowitz DM, Peoples OP. PHA
applications: addressing the price performance issue I. Tissue
engineering. Int J Biol Macromol 1999;25:111–21.
[3] Khanna S, Srivastava AK. Recent advances in microbial polyhy-
droxyalkanoates. Process Biochem 2005;40:607–19.
[4] Steinbuchel A. Perspective for the biotechnological production and
utilization of biopolymers: metabolic engineering of polyhydroxyalk-
anoate biosynthesis pathways as successful example. Macromol
Biosci 2001;1:1–24.
[5] Chen GQ, Wu Q. The application of polyhydroxyalkanoates as tissue
engineering materials. Biomaterials 2005;26:6565–78.
[6] Misra SK, Valappil SP, Roy I, Boccaccini AR. Polyhydroxyalk-
anoate (PHA)/inorganic phase composites for tissue engineering
applications. Biomacromolecules 2006;7:2249–58.
[7] Zheng Z, Bei FF, Deng Y, Tian HL, Chen GQ. Effects of
crystallization of polyhydroxyalkanoate blend on surface physico-
chemical properties and resulting biocompatibility for chondrocytes.
Biomaterials 2005;26:3537–48.
[8] Zhao K, Deng Y, Chen JC, Chen GQ. Polyhydroxyalkanoate (PHA)
scaffolds with good mechanical properties and biocompatibility.
Biomaterials 2003;24:1041–5.
[9] Yang XS, Zhao K, Chen GQ. Effect of surface treatment on the
biocompatibility of microbial polyhydroxyalkanoates. Biomaterials
2002;23:1391–7.
[10] Zhao K, Deng Y, Chen JC, Chen GQ. Polyhydroxyalkanoate (PHA)
scaffolds with good mechanical properties and biocompatibility.
Biomaterials 2003;24:1041–54.
[11] Cheng S, Wu Q, Yang F, Xu M, Leski M, Chen GQ. Influence of
DL-b-hydroxybutyric acid on cell proliferation and calcium influx.
Biomacromolecules 2005;6:593–7.
[12] Cheng S, Chen GQ, Leski M, Zou B, Wang Y, Wu Q. The effect of
D,L-b-hydroxybutyric acid on cell death and proliferation in L929
cells. Biomaterials 2006;27:3758–65.
[13] Martin DP, Peoples OP, Williams SE, Zhong LH. Nutritional and
therapeutic uses of 3-hydroxyalkanoate oligomers, US Patent Appl
359086, 1999.
[14] Zou Z, Sasaguri S, Rajesh KG, Suzuki R. DL-3-hydroxybutyrate
administration prevents myocardial damage after coronary occlusion
in rat hearts. Am J Physiol—Heart Circ Physiol 2002;283(5):
H1968–74.
[15] Tieu K, Perier C, Caspersen C, Teismann P, Wu DC, Yan SD, et al.
() D-beta-hydroxybutyrate rescues mitochondrial respiration and miti-
gates features of Parkinson disease. J Clin Invest 2003;112:892–901.
[16] Kashiwaya Y, Takeshima T, Mori N, Nakashima K, Clarke K,
Veech RL. D-beta-hydroxybutyrate protects neurons in models of
Alzheimer’s and Parkinson’s disease. Proc Natl Acad Sci USA
2000;97:5440–4.
[17] Reusch RN. Transmembrane ion transport by polyphosphate/poly-(R)-
3-hydroxybutyrate complexes. Biochemistry (Moscow) 2000;65:280–95.
[18] Reusch RN. Low molecular weight complexed poly (3-hydroxybu-
tyrate): a dynamic and versatile molecule in vivo. Can J Microbiol
1995;41(Suppl. 1):50–4.
[19] Reusch RN, Sadoff HL. D-(-)-Poly-b-hydroxybutyrate in membranes
of genetically competent bacteria. J Bacteriol 1983;156:778–88.
[20] Seebach D, Fritz MG. Detection, synthesis, structure, and function of
oligo (3-hydroxyalkanoates): contributions by synthetic organic
chemists. Int J Biol Macromol 1999;25:217–36.
[21] Xi JZ, Wu Q, Chen GQ. Hyperproduction of polyesters consisting of
medium-chain-length hydroxyalkanoate monomers by strain Pseu-
domonas stutzeri 1317. Anton Leeuw J Microbiol 2000;78:43–9.
[22] Li S, Vert M. Biodegradation of aliphatic polyesters. In: Scott G,
Gilead D, editors. Degradable polymers. London: Chapman & Hall;
1985. p. 43.

ARTICLE IN PRESSJ. Sun et al. / Biomaterials 28 (2007) 3896–3903 3903
[23] Karlsson S, Sares C, Renstad R, Albertsson A-C. Gas chromato-
graphic, liquid chromatorgraphic and gas chromatographic-mass
spectrometric identification of degradation products in accelerated
aged microbial polyhydroxyalkanoates. J Chromatogr A 1994;669:
97–102.
[24] Fritz MG, Walde P, Seebach D. Oligoesters of (R)-3-hydroxybuta-
noic acid: transmembrane transport of Ca2+ across vesicle bilayers.
Macromolecules 1999;32:574–80.
[25] Wang YW, Yang F, Cheng YC, Yu PHF, Chen JC, Wu Q, et al.
Effect of composition of poly (3-hydroxybutyrate-co-3-hydroxylhex-
anoate) on growth of fibroblast and osteoblast. Biomaterials 2005;
26:755–61.
[26] Deng Y, Zhao K, Zhang XF, Hu P, Chen GQ. Study on the three-
dimensional proliferation of rabbit articular cartilage derived
chondrocytes on polyhydroxyalkanoate scaffolds. Biomaterials 2002;
23:4049–56.
[27] Su J, Tian HM, Liu R, Liang JZ. Inhibitive effects of glucose and free
fatty acids on proliferation of human vascular endothelial cells
in vitro. Chinese Med J 2002;10:1486–90.
[28] Miller ND, Williams DF. On the biodegradation of poly-beta-
hydroxybutyrate homopolymer and poly beta-hydroxybutyrate-
hydroxyvalerate copolymers. Biomaterials 1987;8:129–37.
[29] Wang YW, Wu Q, Chen GQ. Increased hydrophilicity of microbial
polyhydroxyalkanoates via hyaluronan coating reduced growth of
mouse fibroblast L929 cells. Biomaterials 2003;25:4621–9.
[30] Deng Y, Lin XS, Zheng Z, Deng JG, Chen JC, Ma H, et al.
Poly(hydroxybutyrate-co-hydroxyhexanoate) promoted production
of extracellular matrix of articular cartilage chondrocytes in vitro.
Biomaterials 2003;24(23):4273–81.
[31] Sendil D, Gursel I, Wise DL, Hasirci V. Antibiotic release from
biodegradable PHBV microparticles. J Control Rel 1999;59:207–17.
[32] Tasaki O, Hiraide A, Shiozaki T, Yamamura H, Ninomiya N,
Sugimoto H. The dimer and trimer of 3-hydroxybutyrate oligomer as
a precursor of ketone bodies for nutritional care. JPEN—Parenter
Enteral 1999;23:321–5.
[33] Saad B, Ciardelli G, Matter S, Welti M, Uhlschmid GK,
Neuenschwander P, et al. Cell response of cultured macrophages,
fibroblasts, and co-cultures of Kupffer cells and hepatocytes to
particles of short-chain poly [(R)-3-hydroxybutyric acid]. J Mater
Sci—Mater Med 1996;7:56–61.
[34] Saad B, Ciardelli G, Matter S, Welti M, Uhlschmid GK,
Neuenschwander P, et al. Characterization of the cell response of
cultured macrophages and fibroblasts to particles of short-chain poly
[(R)-3-hydroxybutyric acid]. J Biomed Mater Res A 1996;30:429–39.
[35] Reusch RN. Non-storage poly-(R)-3-hydroxyalkanoates (complexed
PHA) in prokaryotes and eukaryotes. In: Doi Y, Steinbuchel A,
editors. Biopolymers. Weinheim, Germany: Wiley-VCH; 2002.
[36] Duzgunes N, Nir S. Mechanisms and kinetics of liposome–cell
interactions. Adv Drug Deliv Rev 1999;40:3–18.
[37] Kendall DA, MacDonald RC. A fluorescence assay to monitor
vesicle fusion and lysis. J Biol Chem 1982;257:13892–5.
[38] Liu F, Huang L. Development of non-viral vectors for systemic gene
delivery. J Control Rel 2002;78:259–66.
[39] Gagnon KD, Lenz RW, Farris RJ, Fuller RC. The mechanical
properties of a thermoplastic elastomer produced by the bacterium
Pseudomonas oleovorans. Rubber Chem Technol 1992;65:761–7.
[40] Hartwell LH, Weinert TA. Checkpoint: controls that ensure the order
of cell cycle events. Science 1989;246:629–34.
[41] Gassner F, Owen AJ. Some properties of poly (3-hycroxubutyrate)
and poly (3-hydroxyvalerate) blends. Polym Int 1999;39:215–9.
[42] Hasirci V. Biodegradable biomedical polymers: a review of degrada-
tion of and in vivo response to polylactides and polyhydroxyalk-
anoates. In: Wiae DL, editor. Biomaterials and bioengineering
handbook. New York: Marcel Dekker; 2000.
[43] Tokiwa Y, Calabia BP. Degradation of microbial polyesters.
Biotechnol Lett 2004;26:1181–9.





























