-
7/26/2019 In vitro reconstitution of wool-IF.pdf
1/7
n v i t r o r e c o n s t i t u t i o n o f w o o l
i n t e r m e d i a t e f i l a m e n t s
Helga Thomas, Andrea Conrads,
K i m - H b P h a n ,
Monika van de Li icht and
H e l m u t
Z a h n t
D e u t s c h e s W o l l f o r s ch u n g s i n st i t u t a n
d e r R W T H A a c h e n V e l t m a n p la t z 8 D - 5 1 0 0 A a
c h e n F R G
(Received 7 February 1986; revised 7 May 1986)
Al thou yh the hard a-kerat ins o f woo l are recognized as mem
bers oJ the in termediate j i lame nts by sequence
comparison thus far a l l a t t empts on recons t i tu t ion o f
woo l a-kerat in f i lamen ts
in vitro
have f i~iled. Here we show
that ox idat ive sulphito lys is rather than the prev ious ly
used S-carboxy methy lat ion i s the method Of choice to
prepare a -kerat in der ivatives sui table ., (or assemb ly
exper ime nts . O nce the protec t ing S-sulpho grou p i s
removed
by 2 - m e r c ap t oe thano l in vitro . fi lament form at ion
can be induced. Elec t ron micrographs show f i laments wi th a
diameter o f ~ 1 1 nm as in a l l o ther in termediate . fi
laments . Thu s , f i lament Jormation o/ c t -kerat ins does
not
require the presence o f m atr ix prote ins .
Keywords: Intermediate filaments: in vitro reconstitu tion:
negative staining; oxidative sulphitolysis
Introduction
I n t e r me d ia t e f i l a me n t s ( I F ) a r e a c o n s t
i t u e n t o f t h e
c y to s k e l e to n p r e s e n t i n a lmo s t a l l v e r t
e b r a t e c e l l s . T h e y
w e r e o r i g in a l l y d e s c r i b e d a t t h e e l e c t
r o n m ic r o s c o p i c a l
l e v e l a s f i l a me n to u s s t r u c tu r e s w i th a c
h a r a c t e r i s t i c
d i a m e t e r o f 7 - 1 n m w h i c h is in t e r m e d i a t
e b e t w e e n th a t
o f th e a c t i n - c o n t a i n i n g m i c r o f i l a m e n
t s ( 5 -7 n m ) a n d t h e
t u b u l i n - c o n t a i n i n g m i c r o t u b u l e s (2 5
n m ) 1. A l t h o u g h I F
a r e a g r o u p o f s t r u c tu r a l l y s imi l a r f il a
me n t s s h o w in g
p a r t ia l s e q u e n c e h o m o l o g y a n d a h i g h p e
r c e n t a g e o f ~-
h e l ic a l c o n f o r m a t io n , t h e y c a n b e d iv id
e d i n to f iv e d i s t i n c t
c la ss e s a c c o r d in g t o t h e i r i m m u n o l o g i c
a l a n d b i o c h e m i c a l
c r i t e r i a : k e r a t i n s , v ime n t in , d e s min , g
l i a l f i l a me n t s a n d
n e u r o f i l a m e n t s 2. C o m p a r e d w i t h t h e o t
h e r f o u r g r o u p s
th e k e r a t i n s r e p r e s e n t t h e mo s t c o m p le x
c l a s s 6 . R e c e n t l y ,
the mic ro f ibr i l s o f ha r d c~-ke ra tins hav e been inc
lude d in to
th e c l as s o f I F b a s e d o n t h e i r s e q u e n c e a
n d s t r u c tu r a l
h o m o lo g i e s w i th t h e o th e r I F p r o t e in s 4
8.
G e n e r a l l y , a m in o a c id se q u e n c e d a t a h a v
e r e v e a l e d
t h a t I F s u b u n i t s a r e c o m p o s e d o f a c e n t
r a l s -h e l ic a l
d o m a i n o f c o n s e r v e d l e n g th a n d s e c o n d a
r y s t r u c t u re a n d
tw o n o n c t - h e l i c a l p a r t s a t t h e a min o a n d
c a r b o x y
t e r m i n u s , s h o w i n g i n c o n t r a s t t o t h e c
e n tr a l d o m a i n w i d e
v a r i a t i o n s i n s i z e a n d p r im a r y s t r u c tu
r e 5 '9 t l
A l t h o u g h w o o l f i l a m e n t o u s s t r u c t u r e
s s i m i l ar t o t h o s e
o f I F h a v e b e e n o b s e r v e d i n t h e e le c t r o n
mic r o s c o p e a f t e r
me c h a n i c a l d i s r u p tu r e o f w o o l f ib r e s l
2, i n t a c t h a i r k e r a t i n
f i l a me n t s h a v e b e e n i s o l a t e d o n ly f r o m
f o l l i c l e s o f r a t
v ib r a s s a e a n d t h e i n n e r r o o t s h e a th o f g
u in e a - p ig
h a i r t 3 't * . A t t e mp t s h a v e b e e n m a d e t o r
e c o n s t i t u t e
mic r o f ib r i l l a r p r o t e in s f r o m h a r d k e r a
t i n f i b r e s ( w o o l )
a f t e r d ig e s t i o n o f t h e n o n - h e l i c a l t a i
ls tS ' t 6 , w h e r e a s i n t a c t
p r o t e in m a te r i a l f r o m th e o th e r I F - c o n t
a in in g t i s s u e s, e. g .
* Partly presented at the EMBO-Workshop Intermediat e
Filaments:
Structure, Function and Patho logy' , Irsee, 27-30 April 1985 as
well as a
poster demonstrat ion at the 3rd Meeting of the European
Cytoskeletal
Club, 'The Cytoskeleton in De velopment and Pathogenesis' ,
Bielefeld,
7-10 September 1985.
+ To whom correspondence should be addressed.
0141-8130/86/050258~7503.00
~ 1986 Butterworth & Co. [Publishers) Ltd
2 5 8 I n t . J . B io l . M a c r o mo l . , 1 9 86 , V o l 8 ,
O c to b e r
a s t r o c y t e s , s mo o th mu s c l e c e l l s , k e r a t
i n i z in g a n d n o n -
k e r a t i n i z in g e p i t h e l i a h a v e b e e n i s o l
a t e d a n d
r e c o n s t i t u t e d
in v i t ro 17.
T h e s e d i f f ic u l ti e s w i th h a r d k e r a t i n s a
r e d u e t o t h e
e x t r e me in s o lu b i l i t y o f t h e w o o l mic r o f
ib r i l l a r p r o t e in s ,
r e s u l t i n g fr o m h ig h c o n t e n t s o f c y s t i n
e b r id g e s a n d t h e
e x i s t e n c e o f a s u lp h u r - r i c h i n t e r mic r o
f ib r i l l a r ma t r i x i n
w o o l a n d h a i r , l e a d in g t o a c o - e x t r a c t i
o n o f b o th
c o mp o n e n t s a f t e r c l e a v a g e o f t h e c y s t i
n e l i n k a g e s .
U p t il l n o w e x t r a c t i o n o f w o o l k e r a t i n s
i n v o lv in g a
s e p a r a t i o n i n to mic r o f ib r i l l a r a n d m a t
r i x p r o t e in s h a s
b e e n p e r f o r m e d u s i n g e i t h e r S - c a r b o x
y m e t h y l a t i o n o r
o x id a t i o n p r o c e d u r e s 1 8't 9. T h e s e l e d ,
h o w e v e r , t o n e w
in t r o d u c e d c h a r g e s . S o f a r n o
i n v i t r o
r e c o n s t i t u ti o n o f
w o o l mic r o f ib r i l s b a s e d o n t h e s e i s o l a t
i o n me th o d s h a s
b e e n d e s c r i b e d . W i th a mo d i f i e d o x id a t i
v e s u lp h i t o ly s i s
p r o c e d u r e a c c o r d i n g t o B a i l e y 2 ' 2 1 w e
h a v e a l r e a d y
s h o w n 2 2-2 4 t h e p o s s ib i l i ty o f e x t r a c t i
n g w o o l k e r a t i n s i n
th e S - s u lp h o f o r m, w h ic h a l l o w s a s e p a r a
t i o n i n to
mic r o f ib r i l l a r a n d ma t r i x p r o t e in s a s w e
l l a s t h e
r e f o r m a t io n o f c y s t i n e b ri d g e s . I t w a s
t h e r e f o r e o b v io u s t o
u s e t hi s t e c h n i q u e in o r d e r t o a t t e m p t -
- a s i n t h e c as e o f
t h e o t h e r I F t y p e s - - a r e c o n s t i t u ti o n o
f w o o l i n t e r m e d i a t e
f i l a me n t s w h ic h c a n b e c h a r a c t e r i z e d b
y e l e c t r o n
mic r o s c o p y . W e s h o u ld l i k e t o d e s c r i b e h
e r e t h e
e x t r a c t i o n p r o c e d u r e a s s u c h , i n v o lv
in g s o lu b i l i z a t i o n
a n d f r a c t io n a t i o n s t e ps , a n d t h e
i n v i t r o
r e c o n s t i t u t i o n o f
wool mic rof ibr i l s .
Mater ia l s and methods
S t a r t i n g m a t e r i a l
U n d a m a g e d L in c o ln w o o l (~b 3 8 / a m, w i th o u
t t ip s ).
E x t e r n a l l i p id w a s r e mo v e d b y a S o x h l e t
e x t r a c t i o n
p r o c e d u r e u s i ng d i c h l o r o m e t h a n e f o r 2
h.
E x t r a c t i o n o f S - s u l p h o - k e r a t e i m
O n e g r a m s n i p p et s o f L i n c o l n w o o l ( a p p r
o x i m a t e l y
2 m m ) w e r e e x t r a ct e d w i t h 1 0 0 m l o f t h e f
ol l o w in g
s u lp h i t o ly s i s r e a g e n t : 0 . 2 M N a 2 S O 3 , 0
. I M N a z S a O 6 i n
-
7/26/2019 In vitro reconstitution of wool-IF.pdf
2/7
In v i t ro
r e c o n s t i t u t i o n o f w o o l i n t e r me d i a t e f
i l a me n t s : H . T h o ma s
et al.
8
M
urea, 0 .1 MTri s , pH 9 .5 , fo r 24 h a t room temp era tu re
.
The inso lub le res idue was removed af t e r cen t r i fuga t
ion
( 4 0 0 0 r e v / mi n , r o o m t e mp e r a t u r e , 3 0 mi n
) a n d t h e
superna tan t was d ia lysed aga ins t de ion ized water fo r
2
days (Visking dialys is tube 27/32).
Separa t ion o f matr ix and micro f ibr i l lar pro te ins
After concen t ra t ing the S-su lpho-kera t e in -con ta in
ing
so lu t ion to 100 ml , 2 ml o f a 1 M z inc ace ta t e so lu t
ion w ere
added ( resu l t ing pH 6 .045 .5 ) and the p rec ip i t a t
ed
pro te ins separa t ed by cen t r i fuga t ion (4000 rev /min ,
roo m
t e mp e r a t u r e , 3 0 mi n ) . T h e p e l l e t w a s s u
s p e n d e d i n 1 %
s o d i u m c i t r a t e s o l u t i o n t o a p r o t e i n c
o n c e n t r a t i o n o f
approx imate ly 1% and d ia lysed aga ins t 30 - fo ld excess o
f
d i s ti l led wa ter and 0 .05 M sod iu m te t rab ora te so lu
t ion .
The p ro te ins were aga in p rec ip i t a t ed wi th z inc ace
ta t e ,
red i s so lved in sod ium c i t ra t e , d i a lysed aga ins t
d i s t i l l ed
wa ter a nd f ina lly f reeze-d r i ed .
In v i t ro
r e c o n s ti t u ti o n o f w o o l f i l a me n t s
The p rec ip i t a t ed f reeze-d r i ed p ro te ins were d i s
so lved in
the fo l lowing so lu t ion (2 mg /ml ) an d kep t fo r 12-15 h
a t
6C: 8 M urea , 8 % f l -m ercap toe thano l , 0 .05 M Tr i s -HC
1
pH 7 .5 . Af te r cen t r i fuga t ion (10 0000 g , 4C, 10m
in)
f i lamen t s were recons t i tu t ed b y d ia lys ing the
supern a tan t
a t 6 C o v e r n i g h t a g a i n s t 4 M u r e a , 1 0 mM T r
i s - H Cl p H
7.5 , 25mM f l -mercap toe thano l . Fur ther d i a lys i s
was
per fo rmed aga ins t 10mM Tr i s -HC1 pH 7 .5 , 10mM f l -
me r c a p t o e t h a n o l f o r a n o t h e r p e r i o d o f
1 2 - 2 4 h a t 6 C .
So lu t ions o f recon s t i tu t ed w ool kera t in f i lamen t
s were
d i lu t ed to approx imate ly i mg/ml wi th the l as t d i a
lys i s
buf fer and a d ro p o f th i s so lu t ion p laced on a g l as
s p l a t e
covered wi th paraf i lm. A carbon coa ted g r id (Cu ,
3 .05 mm , 200 mesh T ed Pe l l a Inc .) was p laced w i th
the
c a r b o n s i de o n t o t h e s u r f a c e o f th e d r o p
, a l lo w i n g t h e
p r o t e i n s to a d s o r b d u r i n g 2 - 5 m i n . T h e s
a mp l e s w e r e th e n
i mme d i a t e l y c o n t r a s t e d w i t h u r a n y l a c
e t a t e ( 1 - 2 % ,
f i l t e red so lu t ion) dur ing 60 s and a i r d r i ed .
Elec tron microscopy
Neg at ive ly s ta ined spec im ens were examined in a Ze i s
s
E M 1 0 9 e l e c t r o n mi c r o s c o p e t h a t w a s o p e
r a t e d a t a n
acce le ra t ing vo l t age o f 50 kV. E lec t ron m icrograph s
w ere
r e c o r d e d a t e i t h e r 2 0 0 0 0 o r 5 0 0 0 0 t i me s
n o mi n a l
ma g n i f i c a ti o n o n A G F A O r t h o 2 5 f ilm a n d d
e v e l o p e d f o r
17min a t 20C in I l fo rd Percep to l . Magn i f i ca t ion
ca l ib ra t ion was per fo rmed us ing a l i ne g r id .
Amino ac id ana lys i s
Sa mp l e s o f th e S - s u l p h o - p ro t e i n s w e r e h
y d r o l y s e d w i t h
c o n s t a n t b o i l i n g 6 M H C I a t 1 0 8C u n d e r v a
c u u m f o r 2 4 h
a n d a n a l y s e d o n a B i o t r o n i c L C 6 0 0 0 E a mi
n o a c i d
ana lyser .
Prepara t ion o f rad io label led S -carboxymethy la ted
kera te ins
T h e r e d u c t i v e c l e a v a g e o f th e S - s u l ph o
g r o u p s a n d t h e
fo l lowing S-carbox ym et hy la t ion s t ep ( iodo[2-14C]_ace
t ic
a c i d ) w e r e c a r r i e d o u t u n d e r t h e s a me c o
n d i t i o n s a s
descr ibed by M arsha l l and Gi l l esp ie 26.
Polyacrylamide gel electrophoresis
So l u t i o n s o f r a d io l a b e l le d p r o t e i n s w e
r e e x a mi n e d b y
t w o - d i me n s i o n a l p o l y a c r y l a mi d e g e l e
l e c t r o p h o r e s i s
accord ing to Ma rsha l l and Gi l l esp ie 26. P ro te in
separa t ion w as car r i ed ou t a t p H 8 .9 ( l s t d im. : 8
M urea ,
7 . 5 % s e p a r a t i o n g e l ; 2 n d d i m . : 0 . 1 % SD
S, 1 0 %
separat ion gel) .
Af te r e l ec t rophores i s , rad io labe l l ed p ro te ins
were
l o c a t e d b y f l u o ro g r a p h y . T h e g e l w a s i
mp r e g n a t e d w i t h
2 . 5 - d ip h e n y l o x a z o le a n d p la c e d o n a K o d
a k - X - O m a t A R
film a t - 35C fo r 7 h a f t e r d ry ing the ge l a t 80C
under
v a c u u m.
Pro te in prepara t ion for l aC-n .m.r s tud ies
Freeze-d r i ed S-su lpho-pro te in f rac t ions were d i s so
lved
in the fol lowing solut ion (1 mg /ml) for 12-15 h at 6C: 8
~o
f l -merca p toe thano l , 8 M urea , 0 .05 M Tr i s -H Cl , pH
7 .5 .
Af te r cen t r i fuga t ion (4000 rev /min , room temp era tu
re ,
1 0 mi n ) t h e s u p e r n a t a n t w a s d i l u t e d w i t
h 8 M u r e a t o
appro x im ate ly 1 mg/m l . P ro te ins w ere f reeze-d r ied a
f te r
d i a ly s in g a g a i n s t a b u f fe r c o n ta i n i n g 0
.0 5 M N H 4 H CO 3 /
N H 3 O H , p H 8 .5 f o r 1 0 d a y s .
Re su l t s
Figure 1
s h o w s s c h e ma t i c a ll y t h e e x t r a c t io n o f w
o o l
kera t ins us ing ox ida t ive su lph i to lys i s . Af t e r
reduc t ive
c leavage o f the cys t ine b r idges wi th so d ium su lph i te
,
c y s t e i n e g r o u p s a r e c o mp l e t e l y c o n v e r
t e d i n t o t h e S -
s u l p h o f o r m b y t h ei r r e a c t i o n w i t h s o d i
u m t e t r a t h io n a t e
and sod ium su lph i t e . A separa t ion in to micro f ib r il
l a r and
mat r ix p ro te ins can be ach ieved accord ing to the i r
d i f fe ren t so lub i l i t y behav iour us ing an i soe lec t
r i c
p rec ip i t a t ion p rocedu re . Af te r rem oval o f the so
lub le
ma t r i x c o mp o n e n t s t h e f i l a me n t o u s S - s u
l p h o p r o t e i n s
can b e t rans fo rm ed in to the th io l fo rm us ing an excess
o f a
reducing agen t .
W i t h t h e o x i d a t i v e s u lp h i to l y s is p r o c e
d u r e m o r e t h a n
50% of the s t a r t ing mater i a l (L inco ln wool ) can b
e
so lub i li zed . Scann ing e l ec t ron m icrosc opy sh ow s 25
tha t
P e l l e t : m i c r o f i b r i l l a r p r o t e i n s
( L S - S S O 3 )
R e d u c t i o n
s s s o s
R e c o n s t i t u t i o n o f I F
( L S - S S - L S )
W o o l
K e r a t i n
e x t r a c t i o n
o x i d a t i v e s u l p h i t o l y s i s )
k e r - S S - k e r ~ 2 k e r - S S 0 3~
I s o e l e c tr ic p r e c i p i t a t i o n
S u p e r n a t a n t : m a t r ix p r o t e in s
( H S - S S 0 3 0 )
Figure 1 W ool keratin extraction schem e. Protein
extraction
procedure performing oxidative sulphitolysis followed by an
isoelectrical precipitation of the microfibrillar, low
sulphur
proteins (LS-SSO~ ); matrix, high sulphur proteins (HS-SSO
3)
remain in solution. Cleavage of the S-sulpho groups can be
achieved by reaction with an excess of reducing agent. Low
sulphur proteins so prepared are the starting material for
the
in
vitro
reconstitution experiments of wool IF
in t . j . Bio l . Macromol . , 1986 , Vol 8 , October
259
-
7/26/2019 In vitro reconstitution of wool-IF.pdf
3/7
I n v i t r o
reconst i tu t ion of w ool in termediate . fi laments: H. Tho
ma s
et a l .
t h e r e m a i n i n g r e s i d u e is c o m p o s e d o f a m
e m b r a n e - l i k e
n e t w o r k o f f i b re f r a g m e n t s a s w el l a s n e
a r l y u n d a m a g e d
w o o l f i b r e s i n m i n o r a m o u n t s .
T h e m a t e r i a l s s ol u b i li z e d w i t h s o d i u m
s u l p h i t e / s o d i u m
t e t r a t h i o n a t e a r e c o m p o s e d o f m i c r o f
i b r il l a r a n d m a t r i x
p r o t e i n m a t e r i a l , a s s h o w n b y c o m p a r i
s o n o f o u r t w o -
d i m e n s i o n a l p o l y a c r y l a m i d e g el e l e c
tr o p h o r e s i s ( 2 D -
P A G E ) p r o t e i n p a t t e r n s w i t h th o s e o f M a
r s h a l l f o r S -
c a r b o x y m e t h y l a t e d k e r a t e i n s Figure
2) 26.
A c c o r d i n g t o t h e e x t r a c t i o n s c h e m e a n
i s o e l e c t r i c
p r e c i p i t a ti o n u s i n g zi n c a c e ta t e w a s p e
r f o r m e d i n o r d e r t o
s e p a r a t e m a t r i x f r o m m i c r o f i b r i l l a r
k e r a t e in s 2 7 ; 6 4 9/0 o f
t h e s o l u b i l i z e d S - s u l p h o p r o t e i n s a l
w a y s p r e c i p i t a t e d ,
i n d e p e n d e n t l y o f t h e e x t r a c t i o n r a t e
. T o t e s t w h e t h e r t h i s
s e p a r a t i o n w a s c o m p l e t e , a m i n o a c i d a
n a l y s i s a s w e l l a s
2 D - P A G E , w e r e c a r ri e d o u t . Table 1 s h o w s t
h e a m i n o
a c i d c o m p o s i t i o n o f th e p r e c i p i ta t e a n
d t h e s u p e r n a t a n t ;
t h e r e a r e m a r k e d d i f fe r e n c e s i n t h e c o n
t e n t o f c e r t a i n
a m i n o a c i d s , e sp e c i a l ly in t h e c y s t i n e c
o n t e n t . T h e
p r e c i p i t a t e d p r o t e i n s a r e e n r i c h e d i
n a - h e l i x s t a b i l i z i n g
a m i n o a c i d s ( A s p , G l u , L y s , L e u , A l a ) w
h i l e t h e o t h e r
~ ~ / ~ i ~ / ~ ~ : / i ~ : ~ ~
F igure 2 2D -P A GE of a wool p ro te in ex trac t The s am ple
i s
com posed of h igh su lphur (HS ) and low su lphur (LS ) p ro te
ins .
Af te r reduc t ive c leavage of the p ro tec t ing S -su lpho
groups
sam ples were rad io labe l l ed by S -ca rboxym ethy la t ion o
f the
resulting thiol groups using iodo(2-14C)acetic acid.
Electrophores is according to Marshal l and Gil lespie26;
charge
sepa ra t ion (u rea ) f i r s t d im ens ion , fo l lowed by S
DS -P AGE a t
r ight angles
Table 1 Am ino ac id com p os i t ion (m ol 9 'o ) o f wool
low/h igh
su lphur p ro te ins . The a r row s ind ica te an inc reased
(1" ) o r
dec reased (~ ) concen t ra t ion in the low su lphur p ro te
ins in
com par i son wi th the h igh su lphur f rac t ion
Low su lphur High su lphur
fract ion fract ion
As p 9.4 ~ 3.1
Th r + 5.5 11.3
Ser I 9.9 14.5
G lu 16.3 T 9.6
Pro ~ 4.9 14.5
Gl y 7.5 6.9
Ala 7.2 T 3.2
(Cys)2~ 2.9 6.3
Val 5.8 6.4
Met 0.6
Ile 3.2 3.6
Leu 10.4 ~ 4.2
Ty r 2.9 2.7
Phe 2.6 2.2
Ly s 1.9 i" 0.3
Hi s 0.8 0.8
Ar g 7.8 8.3
" A low cy stine content is detected because of the fact that
S-sulpho-
cysteine is not completely detectable by amin o acid
analysis
S I
L 8 H S
F igures 3 /4 2D -P A GE of m ic rof ib r i ll a r , low su
lphur (LS ) and m a t r ix , h igh su lphur (HS ) p ro te ins . S
am ple p repa ra t ion a s we ll a s
pe r form ance of the e lec t rophores i s , s ee Figure 2 . The
e lec t rophore t i ca l expe r im ents c lea r ly dem ons t ra te
the pos s ib i l i ty o f a
quan t i t a t ive s epa ra t ion in LS and HS pro te ins accord
ing to the wool ex t rac t ion s chem e
Figure 1)
2 6 0 I n t . J . B i o l . M a c r o m o l . , 1 9 86 , V o l
8, O c t o b e r
-
7/26/2019 In vitro reconstitution of wool-IF.pdf
4/7
I n v i t r o reconstitution of wool intermediate filaments: H
Thomas: et a l .
C H
M i c r o f i b r i l l a r p r o t e i n s i
. / L e u
a h ~ A l a ~ /
I I L P ro . /
I
I I I I i
2 0 0 1 5 0 1 0 0 5 0 0
6 ( p p m )
M a t r i x p r o te i n s
ehl
\
C H
P h e / ~ ~ / / V ~ i a
I I I I i
1 2 0 0 1 5 0 1 0 0 5 0
0
6 ( p p m )
Figures 5/6 t3C-n.m.r, spectra of wool LS/H S prote ins. 75.5 M
Hz ~aC-n.m.r . CP /MA S spectrum o f sol id fract ions of Lincoln
wool .
Measu rements an d signal assignments were conducted accord ing
to K richeldo rf and Mii llera4; for each spectrum abou t 12000
transients were accum ulated. P rote in samples w i th recom
bined cyst ine bridges, received af ter reduct ive c leavage of the
S-sulpho groups
followed by oxidat ive dia lysis, w ere examined. I n the sh ort
range o rder microfibr i l lar prote ins show st rong ~-hel ical
(~h) and weak f l-
st ructure (f ls) signals whereas in the case of the m atr ix
prote ins the st rength of the signals i s
vice versa
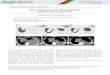
Figure 7 Elect ron microg raph of protofi lamen tous st ructures
of wool microfibri llar mater ia l . After reduct ive cleavage of
the S-sulpho
groups and 100 000 g centr i fugat ion wool microfibr i l lar
prote ins were dia lysed for 16 h di rect ly against a no n-urea co
ntaining T ris-H Cl
buffer . Samples w ere negatively sta ined wi th 1% uranyl aceta
te . Sho rt protofi lame ntous st ructures in a diam eter range
of
approx imately 2-4 nm can be observed. Bar , 100 nm; x
140000
s o lu b l e c o m p o n e n t is m a i n l y c o m p o s e d o
f am i n o a c id s
d i s t u r b i n g ~ t- he li ca l c o n f o r m a t i o n ( P
r o , S e r , T h r ) .
Figures 3 a n d 4 s h o w t h e 2 D p a t t e r n s o f t h e s
e p a r a t e d
p r o t e i n f r a c t i o n s w i t h t h e p r e c i p i t a
t e d p r o t e i n s c l e a r l y
c o r r e s p o n d i n g t o m i c r o f i b r i l l a r , l o
w s u l p h u r w o o l
p r o t e i n s a n d t h e s u p e r n a t a n t o n e s t o t
h e h i g h s u l p h u r
m a t r i x c la s s , a s s h o w n b y c o m p a r i s o n o f
th e s e r e s u l ts w i t h
t h o se a l r e a d y e s t a b l i sh e d i n t h e l i t e r
a t u r e 2 6.
I n t . J . B i o l . M a c r o m o l . , 1 9 8 6, V o l 8 , O c
t o b e r 2 6 1
-
7/26/2019 In vitro reconstitution of wool-IF.pdf
5/7
In vitro
reconst i tu t ion o f wool in termediate f i lamen ts: H. Tho
ma s
et al.
Figure
8 Effectof prolonged dialysis ime on filamentreconstitution of
wool LS proteins. In contrast to the upper sample preparation
(16 h dialysis) a prolonged dialysis (24 h) was performed. Quite
long filamentous structures can be observed with the filaments
having
the tendency to twist around each other. Bar, 100 nm; x
105000
Figures 5
and 6 show ~3C-n.m.r. studies of the protein
fractions after cleavage of the protecting groups and
recombination of cystine linkages. Both keratin classes
show high degrees of short-range order. Besides 10-20%
of totally amorphous parts there is a relation of 8/2 (~-
helical//~-structure) in the microfibril lar proteins while
the
matrix components show a relation of 3/7, indicating a
strongly diminished a-helical content.
The 13C-n.m.r. studies as well as the results of amino
acid analysis, 2D-PAGE and the constant precipitating
ratio of 64 ~o shows that a complete separation in the two
wool protein classes (low sulphur/high sulphur) can be
achieved by the use of the S-sulpho technique.
The low sulphur wool proteins thus isolated were used
as starting material for the
in vitro
reconstitution of wool
microfibrils.
Directly after the reductive cleavage of the protecting
groups, reconst itution of the intermediate filaments in t
he
disulphide form was performed and the filamentous
proteins were negatively stained with uranyl acetate.
After dialysing the samples for 16 h against a non-urea-
containing Tris buffer, relatively short protofilamentous
structures could be observed with a diameter of 2 4 nm,
as shown in Figure 7 .
After a prolonged dialysis (total 24 h) under the same
conditions, filament assembly was more or less
completed, resulting in 7-11 nm IF, with the proteins
having the tendency to aggregate by twisting around each
other
Figure 8).
Obviously the conditions chosen were not yet optimal.
With the help of an additional dialysis step (against 4 M
urea) further improvement in IF reaggregation could be
achieved
Figure 9).
The filaments are no longer twisted;
they appear to be longer and the outlines are sharper.
D i s c u s s i o n
Keratin extraction procedures using oxidative
sulphitolysis have been performed to obtain pure wool
microfibrillar proteins as starting material for
in vitro
reconstitution experiments. Only about 50~ of the
starting material can be solubilized. This is less than with
commonly used reduction procedures 2s and could be
explained by the extraction step as such being very gentle,
resulting in single fibres still resistant against the
chemical
attack, as revealed by scanning electron microscopy.
However, the described method fulfils the following
conditions in order to reconstitute wool-IF:
(1) representative isolation of protein material
throughout the fibre,
(2) prior condition for a complete separation into
microfibrillar and matrix proteins, and
(3) a cleavage of the S-sulpho groups can be easily
performed29.30.
262 Int. J. Biol. Macromol. , 1986, Vol 8, October
-
7/26/2019 In vitro reconstitution of wool-IF.pdf
6/7
In vitro r e c o n s t i t u t i o n o f w o o l i n t e r m e d
i a t e f i l a m e n t s : H . T h o m a s et al.
Figure 9 Electronmicrographs of
intermediate-sizedfilamentsreconstituted from wool microfibrillar
proteins. After cleavage of the
S-sulpho group and 100 000 g centrifugation filament
reconstitution was achieved by two-step dialysis. First, samples
were dialysed
against 4 M urea bufferand then dialysed against the filament
buffer (I0 mM Tris-HC1, 10 mM fl-mercaptoethanol,pH 7.5). This
two-
step procedure obviously led to the best results. Wool
microfibrillar proteins can be reconstituted to long filaments
showing the
diameter range of 7-11 nm being typical for intermediate
filaments. Bar, 100 nm; x 78 000
The latter point represents the main difference in
comparison to the other keratin extraction and
separation procedures; e.g. a reduction followed by S-
carboxymethylation leads to an irreversible introduction
of charged groups into the polypeptide chains. In contrast
to these methods the keratin isolation procedure using
oxidative sulphitolysis allows the reformation o f cystine
bridges and and is therefore an approach to the 'native
structure'.
On the basis of this method it has been possible for the
first time to reconstitute IF from hard keratin fibres in
vi tro showing the typical diameter of 7-11 nm.
Obviously the filament formation is not spontaneous.
A prolonged dialysis time is therefore necessary, possibly
because the reconstitution of IF proceeds via a succession
of intermediate states involving thiol-disulphide
interchange.
With the successful IF reconstitution, hard ~-keratin
microfibrils can be classified as members of the IF family
not only on the basis of sequence data but also on their
electron microscopical appearance, this being the original
criterion.
Our results differ from those obta ined by Campbell e t
al.15 and Ahrnadi et al. 16 who investigated only ~-helical
enriched fragments of low-sulphur keratinous proteins.
However, according to the current state of knowledge, the
non-helical terminal domains of IF proteins play an
essential role in filament assembly and stabili ty 17'a1'32
As these are particulary rich in cysteine9,33 it is not
surprising that previous attempts on S-carboxy-
methylated wool keratin failed to provide in v i tro
filament
formation.
Starting with the in v i tro reconstitution of wool IF,
further studies could elucidate the common properties of
the IF family of proteins. In the case of wool the chemical
and physical behaviour of isolated microfibrils can be
investigated by, e.g. using X-ray techniques to detect a
possible fibre diagram after orientation of IF proteins.
This would complement the 13C-n.m.r. data.
It must also be mentioned that an in vitro
reconstitution of wool IF cannot be achieved with the
same ease as in the case of other IF proteins, e.g. from
human skin. As we have performed IF reconstitution of
callus keratins even without separation of the keratins
from other cellular proteins (A. Conrads, unpublished
data) parallel to the wool microfibrillar proteins, we could
observe that the renatured wool IF often do not show the
more 'normal' appearance compared with the skin
Int. J. Biol. Macromol., 1986, Vol 8, October 263
-
7/26/2019 In vitro reconstitution of wool-IF.pdf
7/7
I n v i t r o reconstitution o f wool intermediate filaments. H.
Th om as et a l .
kerat in IF wh ich are very smo oth an d long. In order to
explain the observed phenomenon a further advance in
the kno wled ge o f the respect ive prote ins i s necessary.
A c k n o w l e d g e m e n t s
We are grate ful to Profes sor H. R . K riche ldorf for
carrying o ut the ~3C-n .m.r . s tudies and for perm iss ion
to
publ i sh the data and to Dr J . F6h les for performing the
am i n o ac i d an a ly s is . F u r t h e r m or e w e w i s h
to t h an k D r R .
C. Marshal l , Dr R . A. Quin lan and E. Bartn ik for
v a l u ab l e d i s c u s s i on s an d t h e D e p ar t m e n
t o f P l an t
P h y s i o l og y a t t h e R W T H A ac h e n f or t h e p e r
for m an c e o f
the 100000 g centr i fugat ion . In addi t ion , we thank
Profes sor Dr K. Weber , Drs E. F innimore and J . F /Sh les
for their critical reading of the man uscript.
R e f e r e n c e s
1 Ishikaw a, H., Bischoff, R. and Ho l tzer, R J Ce l l B i o l
1968, 38,
538
2 Lazaride s, E. A n n R e v B i o c h e m 1982, 51 ,219
3 Mo l l , R. , Fra nke , W. W. and Schi ller, D. L. C e l l
1982, 31, i1
4 Web er , K . and Gei s l e r, N . E M B O J 1982, 1, 1155
5 Gei s l e r , N . and Web er , K . E M B O J 1982, 1, 1649
6 Hanu koglu , I . and Fuchs , E . C e l l 1982, 31 ,243
7 Dow l ing , L . M. , Parry , D . A . D . and Sparrow, L . G .
B i o s c i R e p
1983, 3, 73
8 Fraser , R . D . B . and Mac Rae , T . P . B i o s c i R e p
1983, 3, 517
9 Hanu koglu , I . and Fuchs , E . C e l l 1983, 33, 915
10 Crewther , W. G . , Dow l ing , L . M. , S t e iner t , P .
M. and Parry , D .
A D I n t J B i o l M a c r o m o l 1983, 5, 267
11 Steinert , P. M., Rice, R. H., Roop , D. R., Trus , B. L. and
Steven,
A C N a t u r e 1983, 302, 794
12 Zahn , H . M e l l i a n d T e x t i l b e r 1941, 22,
305
13 Jones , L . N . B i o c h e m B i o p h y s A c t a 1975,
412, 91
14 Steinert , P. M., Dye r, P. Y. and Rogers, G E J I n v e s
t
D e r m a t o l 1971, 56, 49
15 Camp bel l , M. E. , Do bb, M. G . , H i lburn , M. E . , Loh
, P ., Lot ay ,
S . S . , Speakman, P . T . , S t a insby , G . and Yarwood, R .
E . i n
'Proceedings 5 th In t e rna t i ona l Wool Text i l e
Research
Con ference Aa chen ' , (Ed. K. Ziegler), 1976, Vol . II , p.
243
16 Ahm adi , B . , Bos ton , N . M. , Dob b, M. G . and
Speakman, P . T . i n
'Fibrous Proteins: Scient i fic, Indust rial and Medical Aspects
' ,
(Eds D . A . D . Parry and L . K . Creamer) , Academic Press
,
Lon don , 1980, p. 161
17 S t e iner t, P . M. i n 'E l ec t ron Microscopy of Pro t e
ins ' , (Ed . J . R .
Harris), Academic Press, London, 1981, p. 126
18 God dard , D . R . and Michae l i s , L J B i o l C h e m
1935, 11 2, 361
19 Alexander , P . and Ear l and , C . N a t u r e 1950, 166,
396
20 Bailey, J. L. B i o c h e m J 1957, 67, 21
21 Harrap , B . S . and Wo ods , E . F . B i o c h e m J 1964,
92, 8
22 Spei, M. and Thom as , H . C o l l o i d a n d P o l y m S c
i 1983, 261,96 8
23 Spei, M. and Thom as , H . M e l l i a n d T e x t i l b e r
1984, 65, 266
24 Thom as , H . PhD Thesi s, RW TH Aachen , 1984
25 Thom as , H . , Greven , R . and Spei, M. M e l l i a n d T e
x t i l b e r 1983,
64, 297
26 Mars hal l , R. C. and Gi l lespie, R M J F o r e n s S c i S
o c 1982, 22,
377
27 Gillespie, J. M. A u s t J B i o l S c i 1957, 10, 105
28 Greven , R . PhD Thesi s, RW TH , Aachen , 1982
29 Fo otne r, H. B. and Smiles, S J J C h e m S o c 1925, 127,
2887
30 Thom as , H . and Spei, M. M e l l i a n d T e x t i l b e r
1984, 65, 208
31 Sauk , J . J . , Krum mweide , M. , Cocking -Johnson , D .
and Whi t e ,
J G J Ce l l B i o l 1984, 99, 1590
32 Gei s l e r, N . , Kaufm ann, E . and Weber , K . C e l l
1982, 30, 277
33 Fraser , R . D . B ., MacR ae , T . P . and Rogers , G . E .
i n 'Kera t i ns ,
t he i r Composi t i on , S t ruc ture and Biosynthes is ' , C .
C . Thom as ,
Springfield, Il l inois, 1972
34 Kri che ldorf , H . R . and Muel l e r , D . M a c r o m o l
e c u l e s 1983, 16, 615
2 6 4 I n t . J . B i o l . M a c r o m o l . , 1 9 8 6, V o l 8
, O c t o b e r