Embed Size (px)
Citation preview

Kidney International, Vol. 57 (2000), pp. 949–958
Induction of proliferation and apoptotic cell death via P2Y andP2X receptors, respectively, in rat glomerular mesangial cells
HITOSHI HARADA, CHOONG MENG CHAN, ANDREW LOESCH, ROBERT UNWIN,and GEOFFREY BURNSTOCK
Autonomic Neuroscience Institute, Royal Free and University College Medical School, London, England, United Kingdom;Department of Environmental Biochemistry & Toxicology, School of Pharmaceutical Sciences, University of Shizuoka,Shizuoka, Japan; Centre for Nephrology, Royal Free and University College Medical School, Middlesex Hospital, andDepartment of Anatomy and Developmental Biology, Royal Free and University College Medical School, London, England,United Kingdom
Induction of proliferation and apoptotic cell death via P2Y and Cell surface receptors for adenosine 59-triphosphateP2X receptors, respectively, in rat glomerular mesangial cells. (ATP; P2 receptors) have been classified into two families:
Background. Cell surface receptors for adenosine 59-tri- (1) ligand-gated ion channels, P2X receptors (P2X127),phosphate (ATP; P2 receptors) have been subdivided into two
and (2) G-protein–coupled P2Y receptors (P2Y128) [1].families: ligand-gated ion channels (P2X127) and G-protein–P2Y receptors are seven-transmembrane domain recep-coupled (P2Y128) receptors. We investigated the potential roletors that are coupled to G proteins and linked to inositolof P2 receptors on rat glomerular mesangial cells.
Methods. To investigate cell proliferation, DNA synthesis 1,4,5-trisphosphate generation and calcium mobilizationwas assayed by measuring [3H]thymidine incorporation into from intracellular stores [2, 3]. P2X receptors possessDNA. For detecting apoptosis, morphological features, DNA two transmembrane domains and can form multimericfragmentation, and exposure of phosphatidylserine on the out-
channels or pores by polymerization of P2X receptorside surface of the cell membrane were investigated. Expres-subunits. ATP binding gates the opening of these P2Xsion of mRNA and distribution of receptors were detected by
reverse transcription-polymerase chain reaction and immuno- channels to sodium and calcium ions, resulting in mem-histochemistry, respectively. brane depolarization and further calcium influx [4]. The
Results. ATP triggered a dose-dependent increase in DNA seventh member of the P2X receptor family is P2X7synthesis. This response was also induced by uridine triphos- and was formerly known as the P2Z receptor. The P2Zphate (UTP), an agonist equipotent with ATP at P2Y2 andreceptor was the name for the receptor that mediatedP2Y4 receptors; both P2Y2 and P2Y4 mRNA are expressed inthe permeabilizing action of extracellular ATP on mastglomerular mesangial cells and isolated glomeruli. In contrast,
the P2X7 receptor agonist 29-839-O-(4-benzoyl benzoyl) ATP cells [5] and was implicated in extracellular ATP-induced(BzATP) caused a decrease in cell number. BzATP produced cell death [6]. This protein is structurally related to otherDNA cleavage and exposure of phosphatidylserine on the out- members of the P2X family; there is a 35 to 40% aminoside of the cell membrane. P2X7 receptors were distributed
acid identity in the region of homology, but the cyto-heterogeneously in unstimulated cells. The expression of P2X7plasmic C terminal portion is 239 amino acids long inmRNA was maintained at a low level, but was induced bythis receptor compared with 27 to 120 amino acids intumor necrosis factor-a.
Conclusions. Stimulation of glomerular mesangial cells via the other P2X subtypes [7, 8]. Stimulation of the P2X7P2Y2 and/or P2Y4 and via P2X7 receptors can induce prolifera- receptor by ATP opens a small cationic channel, whichtion and apoptotic cell death, respectively. The balance be- then activates a large and nonselective membrane poretween proliferation and apoptosis will depend on the relative
permeable to high molecular weight molecules (betweenstimulation and expression of these P2 receptor subtypes, and600 and 900 D).could play an important role in normal and abnormal glomeru-
Glomerular mesangial cells are important in normallar function.and abnormal glomerular function. Mesangial cell prolif-eration plays a key role in promoting glomerular scarringbecause these cells can elaborate extracellular matrixKey words: cell proliferation, apoptosis, P2 receptors, glomerular cell
function. components, secrete proinflammatory cytokines, andregulate glomerular blood flow and filtration [9]. Apo-Received for publication September 10, 1998ptosis of mesangial cells is an important mode of celland in revised form September 29, 1999
Accepted for publication October 12, 1999 death that maintains normal morphology and functionwithin the glomerulus during development [10] and in 2000 by the International Society of Nephrology
949

Harada et al: P2 receptors in mesangial cells950
normal cell turnover [11]. Cell proliferation was thought vol). The cells incubated for another 10 minutes in 0.5to be the major feature of inflammatory glomerular dis- N NaOH. The dissolved pellet was neutralized with 1.0eases leading to glomerulosclerosis, but recent studies N HCl, 40% TCA (vol/vol) added, and it was incubatedhave shown that apoptosis of mesangial cells also plays for 20 minutes. The samples were rapidly passed throughan important role in the pathological processes associ- a glass fiber filter (GF/C, Whatman) under vacuum andated with glomerular injury, repair, and scarring [12, 13]. rinsed twice with 2 mL of ethanol. The radioactivity re-Since platelets are a rich source of ATP, which is stored tained on the filter was measured by scintillation counting.in their dense bodies [5], it is possible that high local The effect of BzATP on cell number was estimatedconcentrations of ATP are present in the inflamed glo- using the colorimetric alamarBluee dye assay. The cellsmerulus in vivo. Rat mesangial cells proliferate in re- were seeded into 24-well tissue culture plates at a densitysponse to comparable concentrations of ATP and uridine of 5 3 104 cells per well in 400 mL of DMEM/F-12/triphosphate (UTP), and this response profile is charac- 10% FCS. After 24 hours, the medium was changed toteristic of the P2Y receptor [14, 15]. DMEM/F-12/0.5% FCS. After 24 hours, the cells were
In the present study, we have investigated the potential treated for 20 hours with BzATP at various concentra-roles of P2 receptors expressed by glomerular mesangial tions. The alamarBluee solution (40 mL) was added tocells. Extracellular ATP showed dual and opposite ac- all wells, and plates were incubated for four hours attions on glomerular mesangial cell growth, mediating 378C to allow cleavage of alamarBluee by living cells.both cell proliferation and cell death. The receptor sub- The absorbance of the cleavage product was then mea-types involved and their mechanisms of action were ex- sured using a test wavelength of 570 nm and a referenceamined. Our results suggest that extracellular ATP may wavelength of 600 nm. We observed a linear relationshiphave both physiological and pathophysiological roles in between the absorbance of the dye and cell number.the regulation of glomerular mesangial cell function, This linearity extended over almost the entire rangewhich depend critically on the relative expression and tested, from 5000 to 5 3 105 cells/well (data not shown).activation of the P2Y and P2X receptor subtypes. All experiments were performed in triplicate.
Morphological analysisMETHODSFollowing treatment with drugs, microscopic analysisCells
was performed by phase-contrast microscopy. ApoptosisRat mesangial cells from isolated glomeruli of adult was identified using morphological criteria, including
male Sprague-Dawley (SD) rats were kindly donated bycondensation of nuclei and shrinkage of cytoplasm [18].
Dr. M. Kitamura [16, 17]. The cells were maintained inDulbecco’s modified Eagle medium/Ham F-12 (DMEM/ DNA cleavage detection assayF-12; GIBCO-BRL, Gaithersburg, MD, USA) supple-
Cleavage of oligonucleosomal chromatin, a biochemi-mented with 100 U/mL penicillin G, 10 mg/mL strepto-cal hallmark of apoptosis [18], was examined by a laddermycin, 0.25 mg/mL amphotericin B, and 10% fetal calfdetection assay [12]. After treatment with drugs, cellsserum (FCS; GIBCO-BRL) in an atmosphere of 5%(about 1 3 105/dish) were harvested, washed with PBS,CO2/95% air at 378C. Cells were used for experimentsand lyzed with 150 mL of hypotonic lysis buffer [10at passages 20 to 30.mmol/L tris(hydroxymethyl)aminomethane (Tris) hy-drochloride, pH 8.0, 1 mmol/L ethylenediaminetetraace-Cell proliferation assaytic acid (EDTA), 0.5% Triton X-100] for 20 minutes onTo investigate the effect of P2 receptor agonists on cellice. Following the addition of polyethylene glycol andproliferation, DNA synthesis was assayed by measuringNaCl to a final concentration of 2.5% and 1 mol/L, re-[3H]thymidine ([3H]dThd) incorporation into DNA andspectively [19], samples were placed on ice for 10 minutesby the colorimetric alamarBluee assays. The rat mesan-and then centrifuged at 12,000 g for 15 minutes. The super-gial cells were seeded at a density of 5 3 104 cells pernatants were incubated in the presence of deoxyribo-well on a 24-well tissue culture plate and were incubatednuclease-free ribonuclease (400 mg/mL; Sigma, St. Louis,for 24 hours in DMEM/F-12/10% FCS, and the mediumMO, USA) at 378C for one hour and subsequentlywas changed to DMEM/F-12/0.5% FCS. After a 24-hourtreated with proteinase K (400 mg/mL; Sigma) for oneincubation in medium containing 0.5% FCS, the cellshour. They were then precipitated with equal volumeswere incubated with various ATP analogues for 20 hoursof isopropanol at 2208C overnight. After centrifugation,and then treated for 4 hours with [3H]dThd (0.5 mCi/assay)each pellet was dissolved in 10 mL of Tris-EDTA (pHfor incorporation into DNA. The cells were washed twice8.0) buffer and electrophoresed on 1.5% agarose gelwith ice-cold phosphate-buffered saline (PBS), incu-containing ethidium bromide. Ladder formation of oligo-bated with 10% trichloracetic acid (TCA; vol/vol) for 20
minutes at 48C and washed twice with 5% TCA (vol/ nucleosomal DNA was visualized under ultraviolet light.

Harada et al: P2 receptors in mesangial cells 951
Detection of apoptosis by flow cytometry turation) for one minute; 578C, 628C, and 568C for P2X7,P2Y2, and P2Y4, respectively (annealing), for one minute;To detect the early membrane changes of apoptosis,and 728C (extension) for two minutes. Finally, the sam-the annexin V-FITC apoptosis detection kit (Oncogeneples were incubated at 728C (final extension) for sevenResearch Products, Cambridge, MA, USA) was used forminutes. The PCR reactions were performed to 40 cycles.flow cytometric detection of phosphatidylserine exter-The PCR products were analyzed by gel electrophoresis.nalization on the outside of the cell membrane [20]. InThe products were visualized by ultraviolet illuminationbrief, mesangial cells were gently trypsinized, suspendedof the ethidium bromide stained gel.in DMEM/F-12/0.5% FCS, and treated with drugs for
the indicated time. The cells were washed with annexin- Immunohistochemical localization of P2X7 receptor inbinding buffer consisting of 10 mmol/L HEPES, pH 7.4, normal rat kidney150 mmol/L NaCl, 2.5 mmol/L CaCl2, 1 mmol/L MgCl2,
Normal rat kidneys were used and fixed as previouslyand 4% bovine serum albumin (BSA) and incubated fordescribed for the immunolocalization of P2X1 [22]. Ten-15 minutes at room temperature with 0.25 mg/mL ofmicrometer thick sections of renal tissues were incubatedFITC-annexin V in the presence of annexin-bindingin the primary rabbit polyclonal P2X7 antibody (5 mg/mL)buffer. After washing with annexin-binding buffer, apo-in 10% normal horse serum/PBS overnight at room tem-ptosis was assessed by flow cytometry. At least 10,000 cellsperature. The nickel-diaminobenzidine tetrahydrochlo-were analyzed using Coulter Elite workstation software.ride (DAB) enhancement technique was used, whichproduced a black precipitate in the sections. The P2X7Detection of P2X7, P2Y2, and P2Y4 mRNAantibody used was generated as previously describedusing RT-PCRfor the P2X receptor subtypes [23]. A synthetic peptideIn these experiments, total RNA was extracted usingcomprising amino acid residues 394 to 429 correspondingthe technique of Chomczynski and Sacchi [21] with Trizolto the C-terminal 15 amino acids of the rat P2X7 receptor(GIBCO BRL) from glomeruli isolated from renal corticalprotein (NH2-TWRFVSQDMADFAIL-COOH; Gen-
tissue by a standard microsieving technique and frombank Accession number X95882) was covalently linked
cultured mesangial cells. In another series of experiments to KLH (keyhole limpet hemocyanin). Control experi-for detection of P2X7 mRNA, mesangial cells were incu- ments were performed to establish specific immunoreac-bated with tumor necrosis factor-a (TNF-a; 50 ng/mL) tivity: Sections were incubated with P2X7 antibody pre-for 0, 1, 3 and 24 hours, and total RNA was similarly treated with an excess of the homologous peptideextracted. Reverse transcription (RT) was performed antigen; the primary antibody was replaced with nonim-using total RNA extracted from five glomeruli and 5 mg mune rabbit antiserum or without primary antibody.of mesangial cell RNA in a 20 mL RT reaction containing0.5 mg oligo(dT)12-18 primer (GIBCO BRL), 20 units Ultrastructural localization of P2X7 receptor inof RNasin (Promega, Madison, WI, USA), 2 mL dTT mesangial cells(0.1 mol/L), 1 ml dNTP (10 mmol/L), 5 3 First Strand Mesangial cells were grown on Melonex and werebuffer (GIBCO BRL), and 1 mL of Superscripts II Rnase rendered quiescent 24 hours before P2X7 receptor immu-H-reverse transcriptase (GIBCO BRL). The P2X7, P2Y2, nolocalization, as described [23]. In brief, the mesangialand P2Y4 primers were designed from the rat cDNA se- cells were fixed in 4% formaldehyde-0.03% picric acidquences (sense, 59-CGTGCGTTTTGACATCCTGG-39, in 0.1 mol/L phosphate buffer (pH 7.4) for 10 minutes.antisense, 59-ACCATCCAAATGTGGGGCTC-39 for After blocking the endogenous peroxidase activity inP2X7; sense, 59-GGT GTG CGT GCT CGG GCT GTG- 50% methanol/0.3% hydrogen peroxide (H2O2) for 1539, and antisense, 59-TAG GCT CTG TGG CGG GCT minutes, the specimens were incubated in 10% normalTGG-39 for P2Y2; sense, 59-TGT TCC ACC TGG CAT horse serum/PBS to block the nonspecific binding sites.TGT CAG-39, and antisense, 59-AAA GAT TGG GCA The specimens were then incubated in the primary P2X7CGA GGC AG-39 for P2Y4) and produced a polymerase antibody (5 mg/mL) in 10% normal horse serum/PBSchain reaction (PCR) product of 294, 851, and 294 bp, overnight at room temperature. The secondary antibodyrespectively. PCR reactions were performed in 20 mL was a biotinylated donkey antirabbit IgG (Jacksonvolume containing 2 mL 10 3 PCR buffer (Promega), ImmunoResearch, Westgrove, PA, USA), and the Avi-0.4 mL dNTP (10 mmol/L), 0.4 mL MgCl2 (25 mmol/L), din-Biotin Complex technique (extravavidin-horse per-1 unit Taq Polymerase (Promega), and 5 mL of first oxidase; Sigma) was used to increase the sensitivity ofstrand cDNA (Promega). RNase-free water was added antigen localization. DAB and H2O2 were used as theto make up to a final volume of 20 mL. The PCR reaction enzyme substrate that produced a reddish brown precipi-was conducted using a thermal cycler (Hybaid, omn E) tate in the sections. The specimens were postfixed inand incubated at 948C for two minutes (initial denatur- 1% osmium tetroxide, stained with 7% aqueous uranyl
acetate, dehydrated in graded series of ethanol, and em-ation) and then cycled for 35 times through 948C (dena-
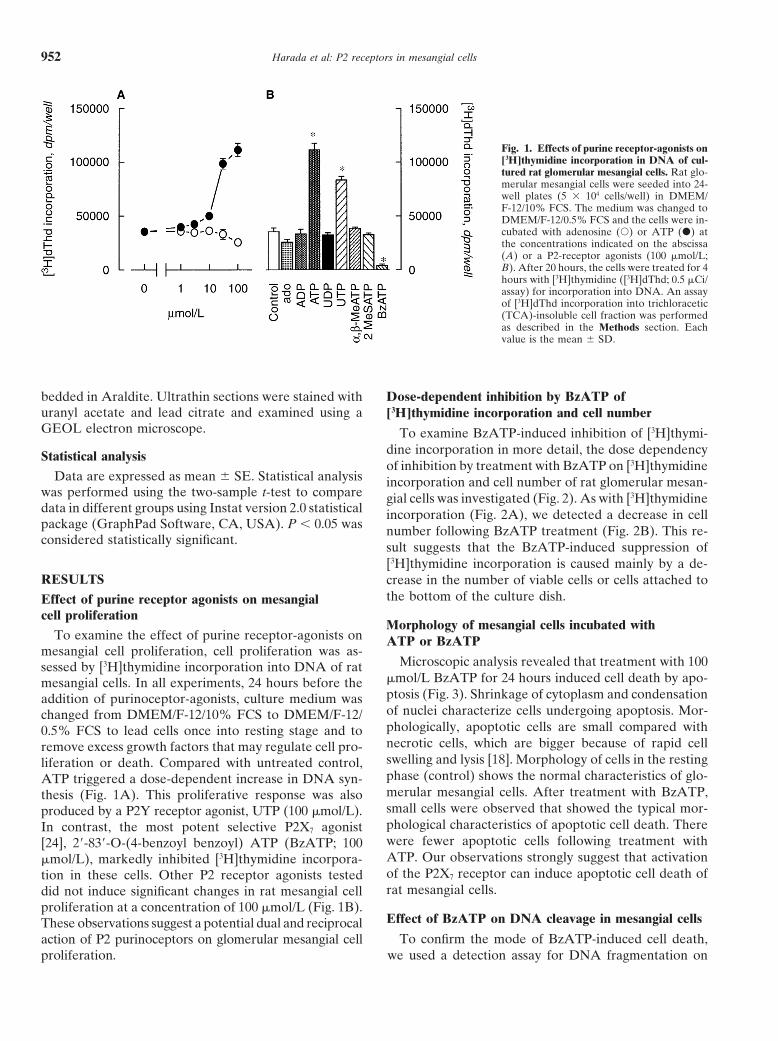
Harada et al: P2 receptors in mesangial cells952
Fig. 1. Effects of purine receptor-agonists on[3H]thymidine incorporation in DNA of cul-tured rat glomerular mesangial cells. Rat glo-merular mesangial cells were seeded into 24-well plates (5 3 104 cells/well) in DMEM/F-12/10% FCS. The medium was changed toDMEM/F-12/0.5% FCS and the cells were in-cubated with adenosine (s) or ATP (d) atthe concentrations indicated on the abscissa(A) or a P2-receptor agonists (100 mmol/L;B). After 20 hours, the cells were treated for 4hours with [3H]thymidine ([3H]dThd; 0.5 mCi/assay) for incorporation into DNA. An assayof [3H]dThd incorporation into trichloracetic(TCA)-insoluble cell fraction was performedas described in the Methods section. Eachvalue is the mean 6 SD.
bedded in Araldite. Ultrathin sections were stained with Dose-dependent inhibition by BzATP ofuranyl acetate and lead citrate and examined using a [3H]thymidine incorporation and cell numberGEOL electron microscope. To examine BzATP-induced inhibition of [3H]thymi-
dine incorporation in more detail, the dose dependencyStatistical analysisof inhibition by treatment with BzATP on [3H]thymidine
Data are expressed as mean 6 SE. Statistical analysis incorporation and cell number of rat glomerular mesan-was performed using the two-sample t-test to compare gial cells was investigated (Fig. 2). As with [3H]thymidinedata in different groups using Instat version 2.0 statistical incorporation (Fig. 2A), we detected a decrease in cellpackage (GraphPad Software, CA, USA). P , 0.05 was number following BzATP treatment (Fig. 2B). This re-considered statistically significant. sult suggests that the BzATP-induced suppression of
[3H]thymidine incorporation is caused mainly by a de-RESULTS crease in the number of viable cells or cells attached to
the bottom of the culture dish.Effect of purine receptor agonists on mesangialcell proliferation
Morphology of mesangial cells incubated withTo examine the effect of purine receptor-agonists on ATP or BzATP
mesangial cell proliferation, cell proliferation was as-Microscopic analysis revealed that treatment with 100sessed by [3H]thymidine incorporation into DNA of rat
mmol/L BzATP for 24 hours induced cell death by apo-mesangial cells. In all experiments, 24 hours before theptosis (Fig. 3). Shrinkage of cytoplasm and condensationaddition of purinoceptor-agonists, culture medium wasof nuclei characterize cells undergoing apoptosis. Mor-changed from DMEM/F-12/10% FCS to DMEM/F-12/phologically, apoptotic cells are small compared with0.5% FCS to lead cells once into resting stage and tonecrotic cells, which are bigger because of rapid cellremove excess growth factors that may regulate cell pro-swelling and lysis [18]. Morphology of cells in the restingliferation or death. Compared with untreated control,phase (control) shows the normal characteristics of glo-ATP triggered a dose-dependent increase in DNA syn-merular mesangial cells. After treatment with BzATP,thesis (Fig. 1A). This proliferative response was alsosmall cells were observed that showed the typical mor-produced by a P2Y receptor agonist, UTP (100 mmol/L).phological characteristics of apoptotic cell death. ThereIn contrast, the most potent selective P2X7 agonistwere fewer apoptotic cells following treatment with[24], 29-839-O-(4-benzoyl benzoyl) ATP (BzATP; 100ATP. Our observations strongly suggest that activationmmol/L), markedly inhibited [3H]thymidine incorpora-of the P2X7 receptor can induce apoptotic cell death oftion in these cells. Other P2 receptor agonists testedrat mesangial cells.did not induce significant changes in rat mesangial cell
proliferation at a concentration of 100 mmol/L (Fig. 1B).Effect of BzATP on DNA cleavage in mesangial cellsThese observations suggest a potential dual and reciprocal
To confirm the mode of BzATP-induced cell death,action of P2 purinoceptors on glomerular mesangial cellproliferation. we used a detection assay for DNA fragmentation on
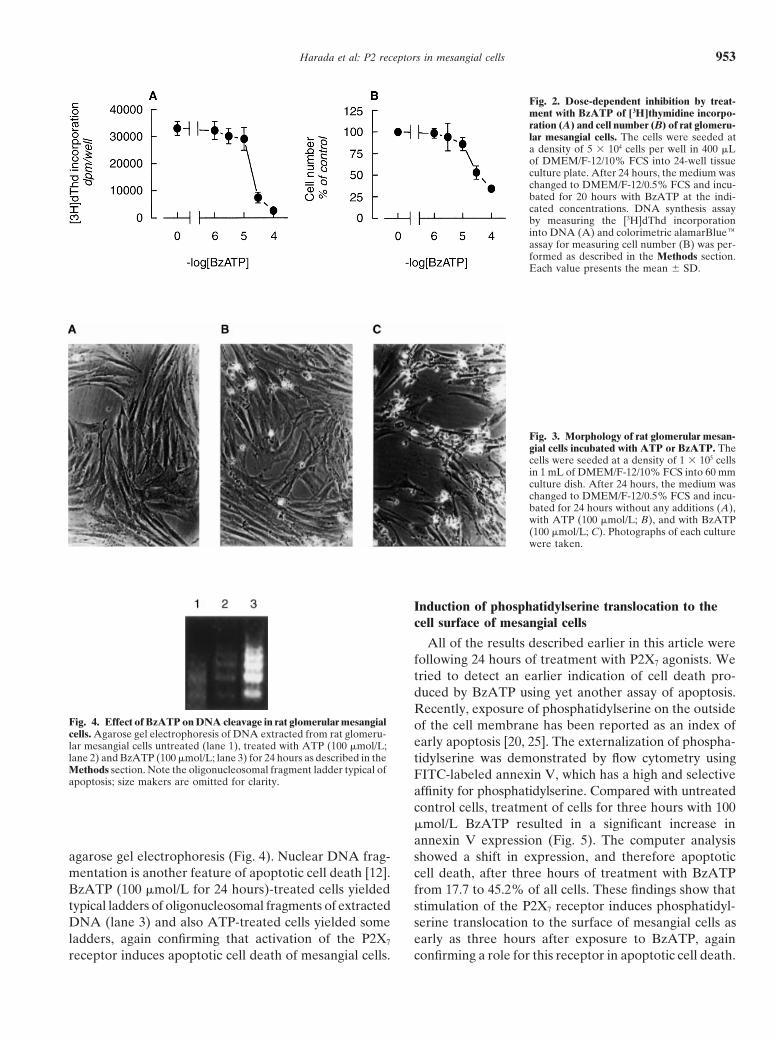
Harada et al: P2 receptors in mesangial cells 953
Fig. 2. Dose-dependent inhibition by treat-ment with BzATP of [3H]thymidine incorpo-ration (A) and cell number (B) of rat glomeru-lar mesangial cells. The cells were seeded ata density of 5 3 104 cells per well in 400 mLof DMEM/F-12/10% FCS into 24-well tissueculture plate. After 24 hours, the medium waschanged to DMEM/F-12/0.5% FCS and incu-bated for 20 hours with BzATP at the indi-cated concentrations. DNA synthesis assayby measuring the [3H]dThd incorporationinto DNA (A) and colorimetric alamarBlueeassay for measuring cell number (B) was per-formed as described in the Methods section.Each value presents the mean 6 SD.
Fig. 3. Morphology of rat glomerular mesan-gial cells incubated with ATP or BzATP. Thecells were seeded at a density of 1 3 105 cellsin 1 mL of DMEM/F-12/10% FCS into 60 mmculture dish. After 24 hours, the medium waschanged to DMEM/F-12/0.5% FCS and incu-bated for 24 hours without any additions (A),with ATP (100 mmol/L; B), and with BzATP(100 mmol/L; C). Photographs of each culturewere taken.
Induction of phosphatidylserine translocation to thecell surface of mesangial cells
All of the results described earlier in this article werefollowing 24 hours of treatment with P2X7 agonists. Wetried to detect an earlier indication of cell death pro-duced by BzATP using yet another assay of apoptosis.Recently, exposure of phosphatidylserine on the outside
Fig. 4. Effect of BzATP on DNA cleavage in rat glomerular mesangial of the cell membrane has been reported as an index ofcells. Agarose gel electrophoresis of DNA extracted from rat glomeru-
early apoptosis [20, 25]. The externalization of phospha-lar mesangial cells untreated (lane 1), treated with ATP (100 mmol/L;lane 2) and BzATP (100 mmol/L; lane 3) for 24 hours as described in the tidylserine was demonstrated by flow cytometry usingMethods section. Note the oligonucleosomal fragment ladder typical of FITC-labeled annexin V, which has a high and selectiveapoptosis; size makers are omitted for clarity.
affinity for phosphatidylserine. Compared with untreatedcontrol cells, treatment of cells for three hours with 100mmol/L BzATP resulted in a significant increase inannexin V expression (Fig. 5). The computer analysis
agarose gel electrophoresis (Fig. 4). Nuclear DNA frag- showed a shift in expression, and therefore apoptoticmentation is another feature of apoptotic cell death [12]. cell death, after three hours of treatment with BzATPBzATP (100 mmol/L for 24 hours)-treated cells yielded from 17.7 to 45.2% of all cells. These findings show thattypical ladders of oligonucleosomal fragments of extracted stimulation of the P2X7 receptor induces phosphatidyl-DNA (lane 3) and also ATP-treated cells yielded some serine translocation to the surface of mesangial cells asladders, again confirming that activation of the P2X7 early as three hours after exposure to BzATP, again
confirming a role for this receptor in apoptotic cell death.receptor induces apoptotic cell death of mesangial cells.
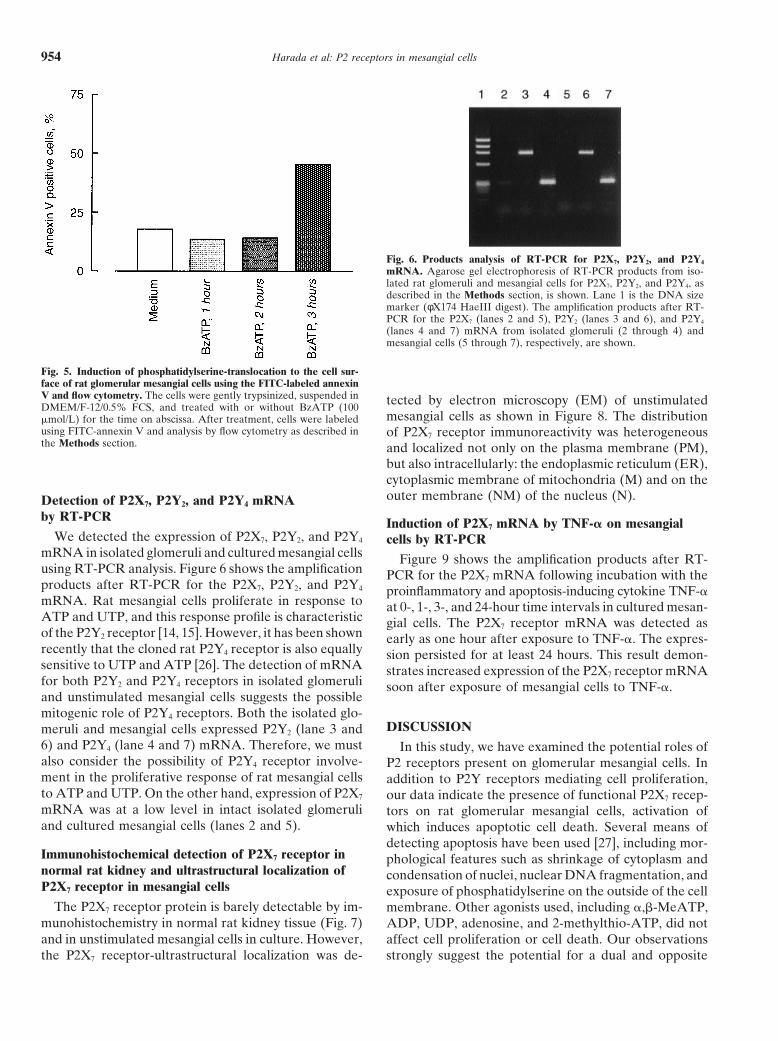
Harada et al: P2 receptors in mesangial cells954
Fig. 6. Products analysis of RT-PCR for P2X7, P2Y2, and P2Y4
mRNA. Agarose gel electrophoresis of RT-PCR products from iso-lated rat glomeruli and mesangial cells for P2X7, P2Y2, and P2Y4, asdescribed in the Methods section, is shown. Lane 1 is the DNA sizemarker (φX174 HaeIII digest). The amplification products after RT-PCR for the P2X7 (lanes 2 and 5), P2Y2 (lanes 3 and 6), and P2Y4
(lanes 4 and 7) mRNA from isolated glomeruli (2 through 4) andmesangial cells (5 through 7), respectively, are shown.
Fig. 5. Induction of phosphatidylserine-translocation to the cell sur-face of rat glomerular mesangial cells using the FITC-labeled annexinV and flow cytometry. The cells were gently trypsinized, suspended in tected by electron microscopy (EM) of unstimulatedDMEM/F-12/0.5% FCS, and treated with or without BzATP (100
mesangial cells as shown in Figure 8. The distributionmmol/L) for the time on abscissa. After treatment, cells were labeledusing FITC-annexin V and analysis by flow cytometry as described in of P2X7 receptor immunoreactivity was heterogeneousthe Methods section. and localized not only on the plasma membrane (PM),
but also intracellularly: the endoplasmic reticulum (ER),cytoplasmic membrane of mitochondria (M) and on theouter membrane (NM) of the nucleus (N).Detection of P2X7, P2Y2, and P2Y4 mRNA
by RT-PCR Induction of P2X7 mRNA by TNF-a on mesangialWe detected the expression of P2X7, P2Y2, and P2Y4 cells by RT-PCR
mRNA in isolated glomeruli and cultured mesangial cells Figure 9 shows the amplification products after RT-using RT-PCR analysis. Figure 6 shows the amplification PCR for the P2X7 mRNA following incubation with theproducts after RT-PCR for the P2X7, P2Y2, and P2Y4 proinflammatory and apoptosis-inducing cytokine TNF-amRNA. Rat mesangial cells proliferate in response to at 0-, 1-, 3-, and 24-hour time intervals in cultured mesan-ATP and UTP, and this response profile is characteristic gial cells. The P2X7 receptor mRNA was detected asof the P2Y2 receptor [14, 15]. However, it has been shown early as one hour after exposure to TNF-a. The expres-recently that the cloned rat P2Y4 receptor is also equally sion persisted for at least 24 hours. This result demon-sensitive to UTP and ATP [26]. The detection of mRNA strates increased expression of the P2X7 receptor mRNAfor both P2Y2 and P2Y4 receptors in isolated glomeruli soon after exposure of mesangial cells to TNF-a.and unstimulated mesangial cells suggests the possiblemitogenic role of P2Y4 receptors. Both the isolated glo-
DISCUSSIONmeruli and mesangial cells expressed P2Y2 (lane 3 and6) and P2Y4 (lane 4 and 7) mRNA. Therefore, we must In this study, we have examined the potential roles ofalso consider the possibility of P2Y4 receptor involve- P2 receptors present on glomerular mesangial cells. Inment in the proliferative response of rat mesangial cells addition to P2Y receptors mediating cell proliferation,to ATP and UTP. On the other hand, expression of P2X7 our data indicate the presence of functional P2X7 recep-mRNA was at a low level in intact isolated glomeruli tors on rat glomerular mesangial cells, activation ofand cultured mesangial cells (lanes 2 and 5). which induces apoptotic cell death. Several means of
detecting apoptosis have been used [27], including mor-Immunohistochemical detection of P2X7 receptor in phological features such as shrinkage of cytoplasm andnormal rat kidney and ultrastructural localization of condensation of nuclei, nuclear DNA fragmentation, andP2X7 receptor in mesangial cells exposure of phosphatidylserine on the outside of the cell
The P2X7 receptor protein is barely detectable by im- membrane. Other agonists used, including a,b-MeATP,munohistochemistry in normal rat kidney tissue (Fig. 7) ADP, UDP, adenosine, and 2-methylthio-ATP, did notand in unstimulated mesangial cells in culture. However, affect cell proliferation or cell death. Our observations
strongly suggest the potential for a dual and oppositethe P2X7 receptor-ultrastructural localization was de-

Harada et al: P2 receptors in mesangial cells 955
Fig. 7. Immunolocalization of P2X7 receptor in normal rat kidney. A view of the low-intensity P2X7 immunoreactivity in the glomeruli (G) ofthe normal rat kidney (magnification 3800) is shown.
action of P2 receptors on rat glomerular mesangial cell trolled so as to prevent inappropriate and unregulatedapoptotic or necrotic cell death.proliferation and death.
The profile of cell proliferation in response to compa- ATP was significantly less potent than BzATP in caus-ing cell death by apoptosis. This probably accounts forrable concentrations of ATP and UTP was characteristic
of the P2Y2 receptor [14, 15]. Recently, it has been shown the predominant mitogenic effect of ATP on mesangialcells via P2Y receptors; only under conditions of massivethat the cloned rat P2Y4 receptor is also equally sensitive
to UTP and ATP [26]. We also detected the expression ATP release and/or up-regulation of P2X7 receptors canATP induce mesangial cell death. Humphreys and Du-of the P2Y2 and P2Y4 mRNA in rat glomerular mesangial
cells and the isolated glomeruli. Thus, there is also the byak reported the induction of the P2X7 receptor bylipopolysaccharide and interferon-g in the human THP-1possibility of P2Y4 receptor involvement in the prolifera-
tive response of rat mesangial cells to ATP and UTP. monocytic cell line [28]. Their report suggests that P2X7
receptor expression might increase in damaged tissueThe P2X7 mRNA has a more limited expression, atleast in normal and healthy tissue, including kidney [27]. following ischemic injury. In glomerulonephritis (GN),
immune deposits attract inflammatory cells and result inIn fact, expression of P2X7 mRNA is normally at a lowlevel in glomeruli and is undetectable in unstimulated local release of TNF-a; therefore, we investigated the
effect of TNF-a on P2X7 receptor mRNA expression.mesangial cells. Although the intensity of P2X7 immuno-reactivity in the glomeruli of the normal rat kidney is Treatment with TNF-a induced P2X7 receptor mRNA
in a time-dependent manner in rat mesangial cells. Thislow, we could detect the localization of P2X7 receptorprotein in unstimulated rat mesangial cells by EM. The finding suggests that expression of P2X7 mRNA is nor-
mally maintained at a low level in vivo, but can increaseP2X7 receptors were distributed heterogeneously andpresent not only on the PM, but also intracellularly, in significantly in pathological states.
The characteristic pathological features of GN are in-the ER, cytoplasmic membrane of M, and on the outernuclear membrane. These findings raise the possibility flammatory cell infiltration, extracellular matrix accumu-
lation, and glomerular cell proliferation, all resulting inof an intracellular role for the cytolytic P2X7 receptor(perhaps in relation to the apoptotic pathway) and/or its glomerulosclerosis and scarring. Because mesangial cell
expansion within the glomerulus can herald glomerulartrafficking to the PM and surface expression under cer-tain conditions. It is perhaps not surprising that the sur- scarring, turnover of mesangial cells may be important.
In some forms of glomerular injury, mesangial cell prolif-face expression of such a receptor should be tightly con-
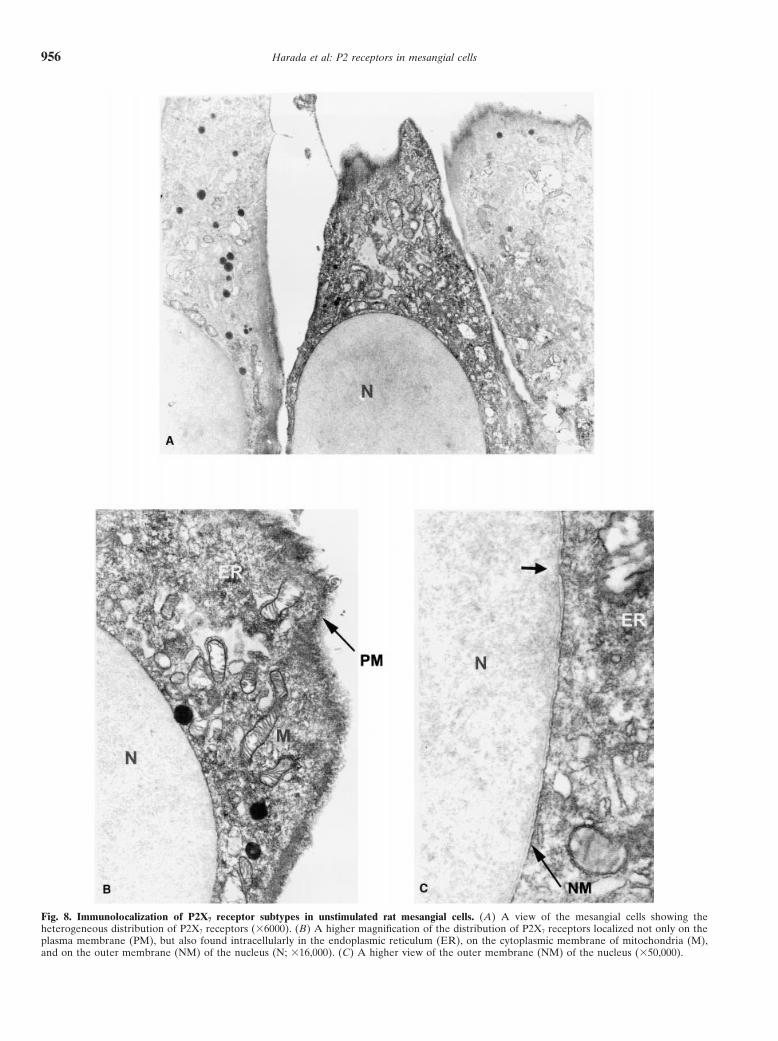
Harada et al: P2 receptors in mesangial cells956
Fig. 8. Immunolocalization of P2X7 receptor subtypes in unstimulated rat mesangial cells. (A) A view of the mesangial cells showing theheterogeneous distribution of P2X7 receptors (36000). (B) A higher magnification of the distribution of P2X7 receptors localized not only on theplasma membrane (PM), but also found intracellularly in the endoplasmic reticulum (ER), on the cytoplasmic membrane of mitochondria (M),and on the outer membrane (NM) of the nucleus (N; 316,000). (C) A higher view of the outer membrane (NM) of the nucleus (350,000).

Harada et al: P2 receptors in mesangial cells 957
and vascular smooth muscle cells [5, 38, 39]. We speculatethat both the regulation of local concentrations of extra-cellular ATP and the different affinities of receptor sub-types for ATP are important in determining mesangialcell proliferation or death. Thus, our observations sug-gest that P2 receptors may have a role in normal andabnormal glomerular mesangial cell function. The bal-ance between proliferation and apoptosis will dependon the stimulation of the various P2 receptors subtypesexpressed on these cells. The P2X7 receptor mediationof apoptosis may be open to manipulation and couldprovide a novel means of therapy in glomerular disease.
Fig. 9. Products analysis of RT-PCR for P2X7 mRNA following expo-sure to TNF-a. Agarose gel electrophoresis of RT-PCR products from ACKNOWLEDGMENTSrat glomerular mesangial cells without (lane 2) after exposure to TNF-a
This work was also supported in part by a grant-in-aid for encourage-for 1, 3 and 24 hours (lane 3 through 5), respectively, as described inment of young scientists from the Ministry of Education, Science, andthe Methods section. Lane 1 is the DNA size marker (φX174 HaeIIICulture, Japan (H. Harada), and by the Medical Research Council,digest).UK (C.M. Chan, R. Unwin, G. Burnstock). We thank Dr. M. Kitamurafor his advice and comments and for his help with the cultured mesan-gial cells and Roche Bioscience, Palo Alto, CA, USA, for support.
Reprint requests to Dr. Hitoshi Harada, Department of Environmen-eration resolves, and glomerular structure and functiontal Biochemistry and Toxicology, School of Pharmaceutical Sciences,return to normal. In experimental mesangial prolifera-University of Shizuoka, 52-1 Yada, Shizuoka 422-8526, Japan.
tive GN, mesangial cell apoptosis is a major repair mech- E-mail: [email protected] in the resolution of mesangial hypercellularity[29, 30]. Apoptosis is also a feature of healing and scar REFERENCESformation in progressive crescentic GN [31]. These data
1. Abbracchio MP, Burnstock G: Purinoceptors: Are there familiessuggest that apoptosis is important in the process of of P2X and P2Y purinoceptors? Pharmacol Ther 64:445–475, 1994
2. Webb TE, Simon J, Krishek BJ, Bateson AN, Smart TG, Burn-glomerular inflammation and healing, but that its dysreg-stock G, Barnard EA: Cloning and functional expression of aulation may lead to scarring and progression to glomeru-brain G-protein-coupled ATP receptor. FEBS Lett 324:219–225,
losclerosis. For example, in glomerular diseases such as 19933. Barnard EA, Burnstock G, Webb TE: G protein-coupled recep-IgA and lupus nephritis, cell depletion is by apoptosis,
tors for ATP and other nucleotides: A new receptor family. Trendswhich subsequently progresses to sclerosis [13]. A bal-Pharmacol Sci 15:67–70, 1994
ance between proliferation and apoptosis may be an im- 4. Valera S, Hussy N, Evans RJ, Adami N, Surprenant A, BuelG: A new class of ligand-gated ion channel defined by P2X receptorportant mechanism of glomerular remodeling during andfor extracellular ATP. Nature 371:516–519, 1994after injury. Thus, manipulating these pathways could
5. Gordon JL: Extracellular ATP: Effects, sources and fate. Biochemreduce the hypercellularity seen in proliferative GN and J 233:309–319, 1986
6. Chow SC, Kass GN, Orrenius S: Purines and their roles in apopto-limit glomerular scarring [32].sis. Neuropharmacology 36:1149–1156, 1997Apoptosis-inducing signals lead to an organized and
7. Rassendren F, Buell GN, Virginio C, Collo G, North RA: Theself-amplifying cascade that involves a variety of enzymes permeabilizing ATP receptor, P2X7: Cloning and expression of a
human cDNA. J Biol Chem 272:5482–5486, 1997known as caspases [related to interleukin-1b–converting8. Surprenant A, Rassendren F, Kawashima E, North RA, Buellenzyme, (ICE)]. Through a complex series of protein
G: The cytolytic P2Z receptor for extracellular ATP identified askinase and transcription factor activation, the biochemi- a P2X receptor (P2X7). Science 233:309–319, 1996
9. Lovett DH, Sterzel RB: Cell culture approaches to the analysiscal sequelae and morphological changes of apoptosis andof glomerular inflammation. Kidney Int 30:246–254, 1986cell death are induced [33, 34]. ICE is responsible for
10. Oppenheim RW: Cell death during development the nerve system.the generation of mature IL-1b; in apoptosis as opposed Annu Rev Neurosci 14:453–501, 1991
11. Hall PA, Coates PJ, Ansari B, Hopwood D: Regulation of cellto necrosis, there is an increase in release of IL-1b [35].number in the mammalian gastrointestinal tract: The importanceExtracellular ATP, acting on the P2X7 receptor, has beenof apoptosis. J Cell Sci 107:3569–3577, 1994
shown to be a powerful stimulus for ICE activation. In 12. Savill JS, Wyllie AH, Henson JE, Walport MJ, Henson PM,Haslett C: Macrophage phagocytosis of aging neutrophils in in-macrophage and microglial cells, activation of the P2X7flammation: Programmed cell death in the neutorphil leads to itsreceptor causes a large and rapid release of mature IL-1brecognition by macrophages. J Clin Invest 83:865–875, 1989
[36, 37]. 13. Sugiyama H, Kashihara N, Makino H, Yamasaki Y: Apoptosisin glomerular sclerosis. Kidney Int 49:103–111, 1996Extracellular ATP is normally in the low micromolar
14. Huwiler A, Pfeilschifter J: Stimulation by extracellular ATPrange (5 to 20 mmol/L), but in hypoxia or ischemia, stressand UTP of the mitogen-activated protein kinase cascade and
injury, or membrane damage, ATP can be released in proliferation of rat renal mesangial cells. Br J Pharmacol 113:1455–1463, 1994large quantities (mmol/L) from platelets, endothelial,

Harada et al: P2 receptors in mesangial cells958
15. Heidenreich S, Tepel M, Schluter H, Harrach B, Zidek W: RA, Buell G: Tissue distribution of the P2X7 receptor. Neuro-pharmacology 36:1277–1283, 1997Regulation of rat mesangial cell growth by diadenosine phosphates.
28. Humphreys BD, Dubyak GR: Induction of the P2z/P2X7 nucleo-J Clin Invest 95:2862–2867, 1995tide receptor and associated phospholipase D activity by lipopoly-16. Kitamura M, Mitarai T, Maruyama N, Nagasawa R, Yoshidasaccharide and IFN-g in the human THP-1 monocytic cell line.H, Sakai O: Mesangial cell behavior in a three dimensional extra-J Immunol 157:5627–5637, 1996cellular matrix. Kidney Int 40:653–661, 1991
29. Baker AJ, Mooney A, Hughes J, Lombardi D, Johnson RJ, Savill17. Kitamura M, Burton S, English J, Kawachi H, Fine LG: TransferJ: Mesangial cell apoptosis: The major mechanism for resolutionof a mutated gene encoding active transforming growth factor-b1of glomerular hypercellularity in experimental mesangial prolifera-suppresses mitogenesis and IL-1b response in the glomerulus. Kid-tive nephritis. J Clin Invest 94:2105–2116, 1994ney Int 48:1747–1757, 1995
30. Shimizu A, Kitamura H, Masuda Y, Sugisaki Y, Yamanaka N:18. Areds MJ, Morris RG, Wyllie AH: Apoptosis: The role of theApoptosis in the repair process of experimental proliferative glo-endonuclease. Am J Pathol 136:593–608, 1990merulonephritis. Kidney Int 47:114–121, 199519. Ioannou YA, Chen FW: Quantitation of DNA fragmentation in
31. Shimizu A, Masuda Y, Kitamura H, Sugisaki Y, Yamanaka N:apoptosis. Nucleic Acids Res 24:992–993, 1996Apoptosis in progressive crescentic glomerulonephritis. Lab Invest20. Koopman G, Reutelingsperger CP, Kuijten GA, Keehnen RM,74:941–951, 1996Van Oers MH: Annexin V for flow cytometric detection of phos-
32. Savill J: Apoptosis: Will cell death add life to nephrology? Nephrolphatidylserine expression on B cells undergoing apoptosis. BloodDial Transplant 10:1977–1979, 199584:1415–1420, 1994
33. Alnemri ES: Mammalian cell death proteases: A family of highly21. Chomczynski P, Sacchi N: Single-step method of RNA isolation byconserved aspartate specific cysteine proteases. J Cell Biochemacid guanidinium thiocyanate-phenol-chloroform extraction. Anal64:33–42, 1997Biochem 162:156–159, 1987
34. Zhivotovsky B, Burgess DH, Vanags DM, Orrenius S: Involve-22. Chan CM, Unwin RJ, Bardini M, Oglesby IB, Ford APDW, ment of cellular proteolytic machinery in apoptosis. Biochem Bio-Townsend-Nicholson A, Burnstock G: Localization of the P2X1 phys Res Commun 230:481–488, 1997purinoceptors by autoradiography and immunohistochemistry in 35. Hogquist KA, Nett MA, Unanue ER, Chaplin DD: Interleukinthe rat kidney. Am J Physiol 274:F799–F804, 1998 1 is processed and released during apoptosis. Proc Natl Acad Sci
23. Groeschel-Stewart U, Bardini M, Robson T, Burnstock G: USA 88:8485–8489, 1991Localisation of P2X5 and P2X7 receptors by immunohistochemistry 36. Boersma AW, Nooter K, Oostrum RG, Stoter G: Quantificationin rat stratified squamous epithelium. Cell Tissue Res 296:599–605, of apoptotic cells with fluorescein isothiocyanate-labeled annexin1999 V in Chinese hamster ovary cell cultures treated with cisplatin.
24. Soltoff SP, McMillian MK, Talamo BR: ATP activates a cation- Cytometry 24:123–130, 1996permeable pathway in rat parotid acinar cells. Am J Physiol 37. Perregaux D, Gabel CA: Interleukin-1b maturation and release262:C934–C940, 1992 in response to ATP and nigericin: Evidence that potassium deple-
25. Fadok VA, Voelker DR, Campbell PA, Cohen JJ, Bratton DLH: tion mediated by these agents is a necessary and common featureExposure of phosphatidylserine on the surface of apoptotic lym- of their activity. J Biol Chem 269:15195–15203, 1994phocytes triggers specific recognition and removal by macrophages. 38. Born GV, Kratzer MA: Source and concentration of extracellularJ Immunol 148:2207–2216, 1992 adenosine triphosphate during haemostasis in rats, rabbits and
26. Bogdanov YD, Wildman SS, Clements MP, King BF, Burnstock man. J Physiol 354:419–429, 1984G: Molecular cloning and characterization of rat P2Y4 nucleotide 39. Bergfeld GR, Forrester T: Release of ATP from human erythro-receptor. Br J Pharmacol 124:428–430, 1998 cytes in response to a brief period of hypoxia and hypercapnia.
Cardiovasc Res 26:40–47, 199227. Collo G, Neidhart S, Kawashima E, Kosco-Vilbois M, North





























