Embed Size (px)
Citation preview

Stoffwechsel:
1. Grundprinzipien des Metabolismus
2. Enzyme & Cofaktoren
3. Glykolyse und Gärung
4. Citratzyklus – die zentrale Drehscheibe des Metabolismus
5. Atmungskette und ATP-Synthese
6. Pentosephosphatweg – der Adapter im Stoffwechsel
7. Gluconeogenese und Cori-Zyklus
8. Biosynthese und Abbau von Glycogen
9. Fettsäuresynthese und β-Oxidation
10.Stoffwechsel von Cholesterin, Steroiden und Membranlipiden
11.Aminosäurestoffwechsel und Harnstoffzyklus
12.Stoffwechsel der Nukleotide
Inhalt der Vorlesung
1
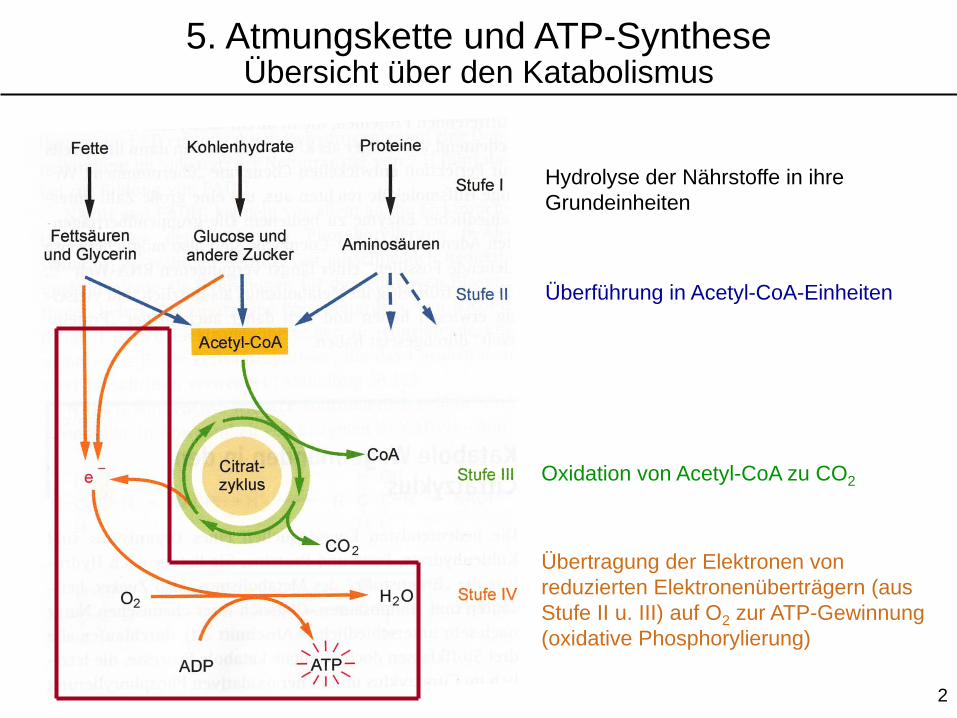
Hydrolyse der Nährstoffe in ihre
Grundeinheiten
Überführung in Acetyl-CoA-Einheiten
Oxidation von Acetyl-CoA zu CO2
Übertragung der Elektronen von
reduzierten Elektronenüberträgern (aus
Stufe II u. III) auf O2 zur ATP-Gewinnung
(oxidative Phosphorylierung)
5. Atmungskette und ATP-SyntheseÜbersicht über den Katabolismus
2
3
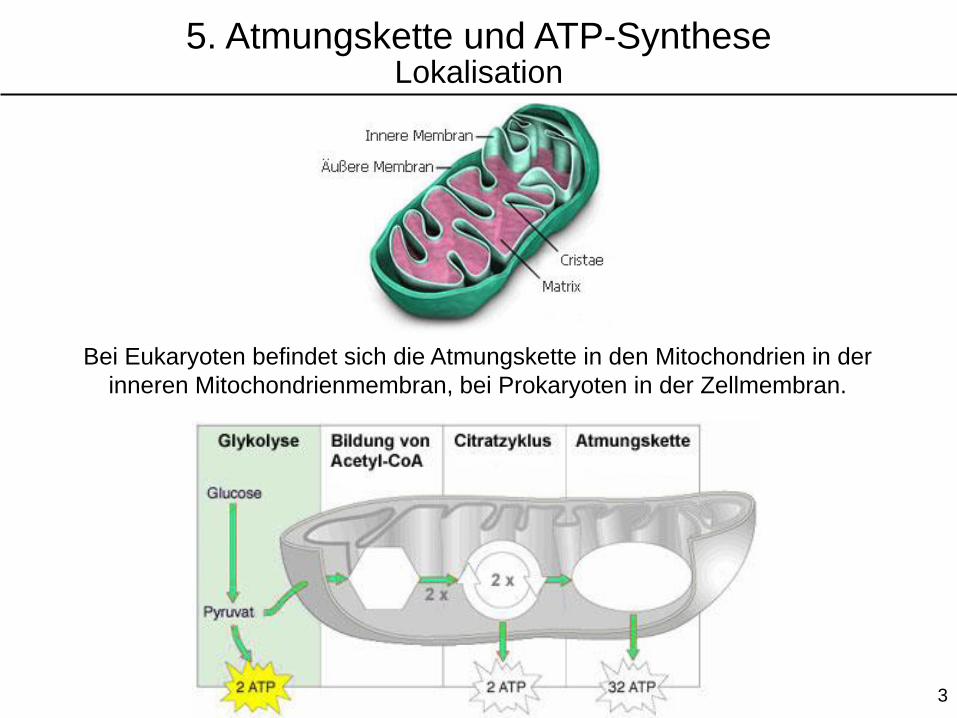
Bei Eukaryoten befindet sich die Atmungskette in den Mitochondrien in der
inneren Mitochondrienmembran, bei Prokaryoten in der Zellmembran.
5. Atmungskette und ATP-SyntheseLokalisation
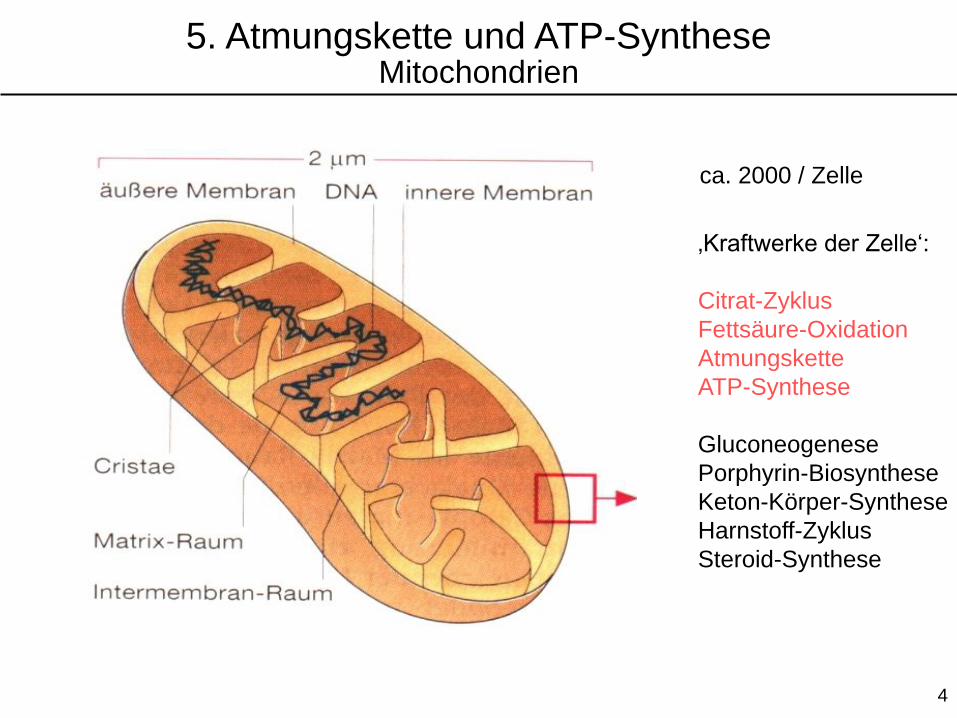
ca. 2000 / Zelle
‚Kraftwerke der Zelle‘:
Citrat-Zyklus
Fettsäure-Oxidation
Atmungskette
ATP-Synthese
Gluconeogenese
Porphyrin-Biosynthese
Keton-Körper-Synthese
Harnstoff-Zyklus
Steroid-Synthese
5. Atmungskette und ATP-SyntheseMitochondrien
4
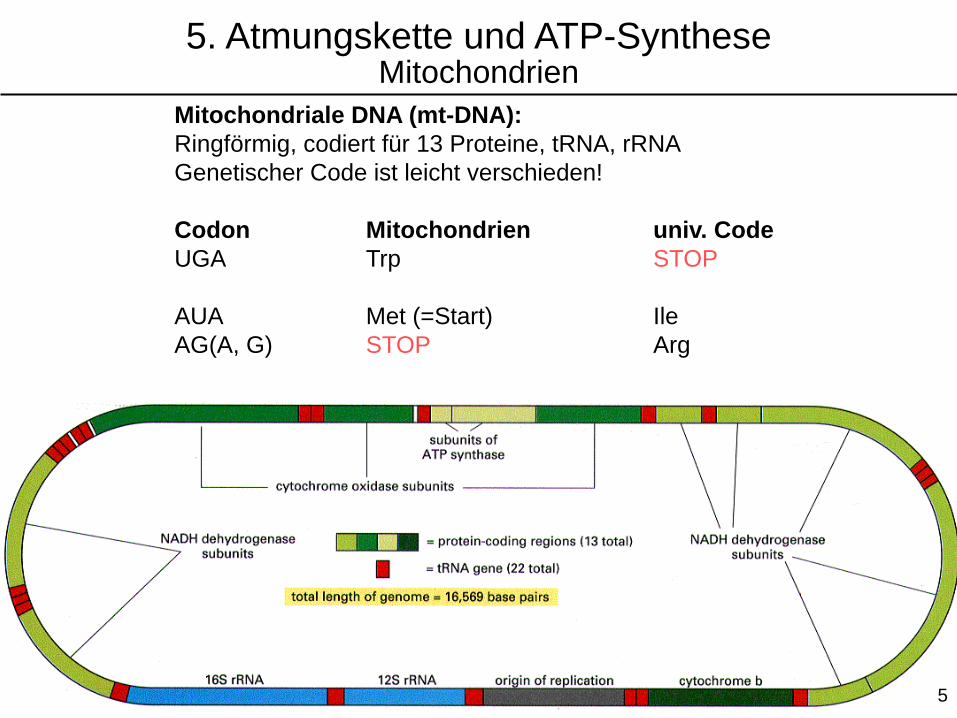
Mitochondriale DNA (mt-DNA):
Ringförmig, codiert für 13 Proteine, tRNA, rRNA
Genetischer Code ist leicht verschieden!
Codon Mitochondrien univ. Code
UGA Trp STOP
AUA Met (=Start) Ile
AG(A, G) STOP Arg
5. Atmungskette und ATP-SyntheseMitochondrien
5
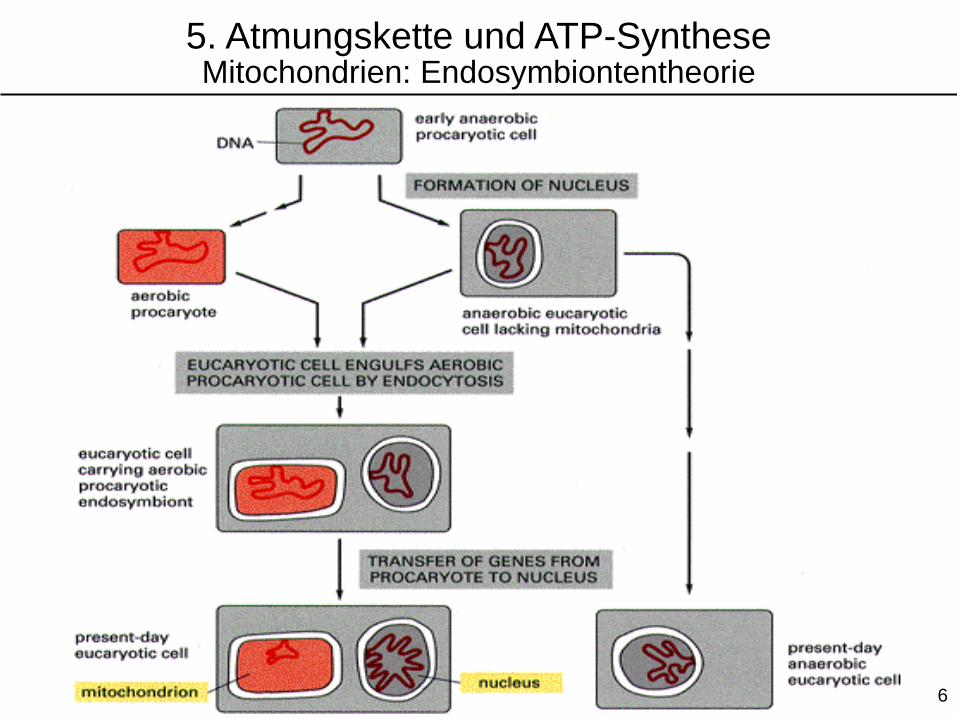
5. Atmungskette und ATP-SyntheseMitochondrien: Endosymbiontentheorie
6
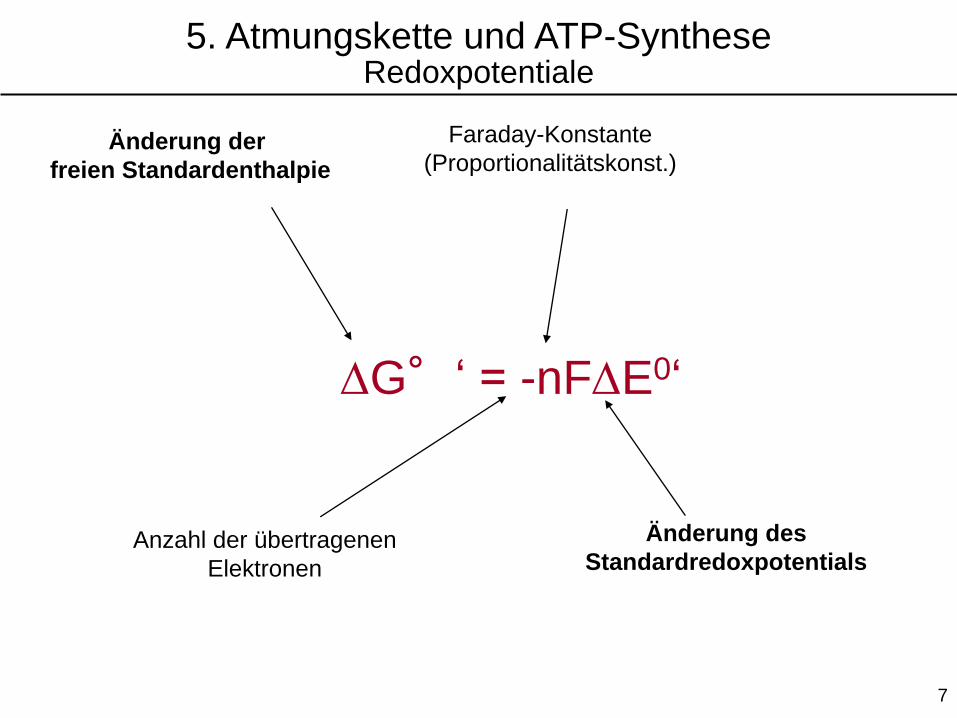
DG°‘ = -nFDE0‘
Anzahl der übertragenen
Elektronen
Faraday-Konstante
(Proportionalitätskonst.)Änderung der
freien Standardenthalpie
Änderung des
Standardredoxpotentials
7
5. Atmungskette und ATP-SyntheseRedoxpotentiale

Die Potentialdifferenz zwischen NADH und O2 ist sehr groß!
Die treibende Kraft bei der oxidativen Phosphorylierung:
Elektronentransfer von NADH (und FADH2) auf O2.
1. Teilreaktion: 1/2 O2 + 2 H+ + 2e- H2O E0` = +0,82V
2. Teilreaktion: NAD+ + H+ + 2e- NADH E0` = -0,32V
1/2 O2 + NADH + H+ H2O + NAD+ ΔE0` = +1,14V
=> NADH ist ein guter e- Donor, O2 ein guter e- Akzeptor:
Die Reaktion ist begünstigt: 1,14 Volt entspricht -220kJ/mol.
5. Atmungskette und ATP-SyntheseRedoxpotentialdifferenz zwischen NADH und O2
8
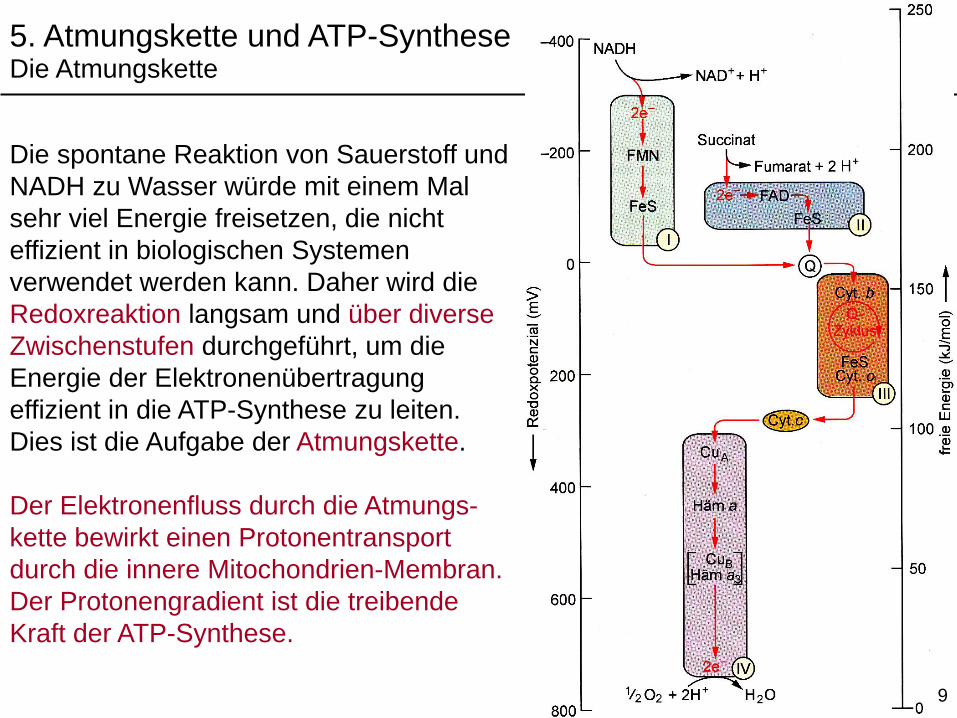
Die spontane Reaktion von Sauerstoff und
NADH zu Wasser würde mit einem Mal
sehr viel Energie freisetzen, die nicht
effizient in biologischen Systemen
verwendet werden kann. Daher wird die
Redoxreaktion langsam und über diverse
Zwischenstufen durchgeführt, um die
Energie der Elektronenübertragung
effizient in die ATP-Synthese zu leiten.
Dies ist die Aufgabe der Atmungskette.
Der Elektronenfluss durch die Atmungs-
kette bewirkt einen Protonentransport
durch die innere Mitochondrien-Membran.
Der Protonengradient ist die treibende
Kraft der ATP-Synthese.
5. Atmungskette und ATP-SyntheseDie Atmungskette
9
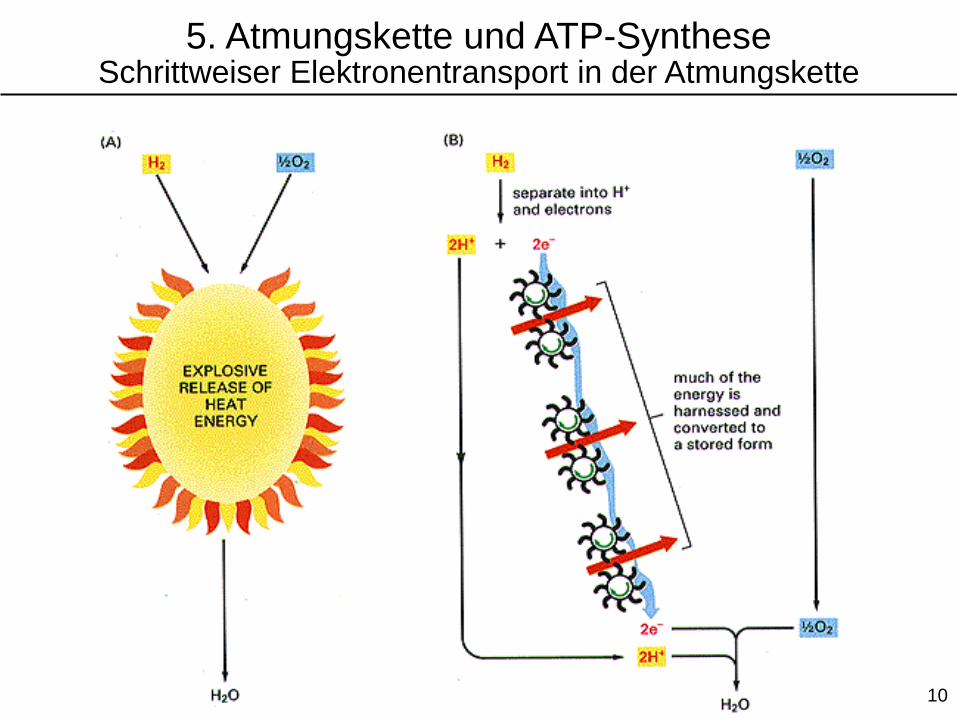
5. Atmungskette und ATP-SyntheseSchrittweiser Elektronentransport in der Atmungskette
10
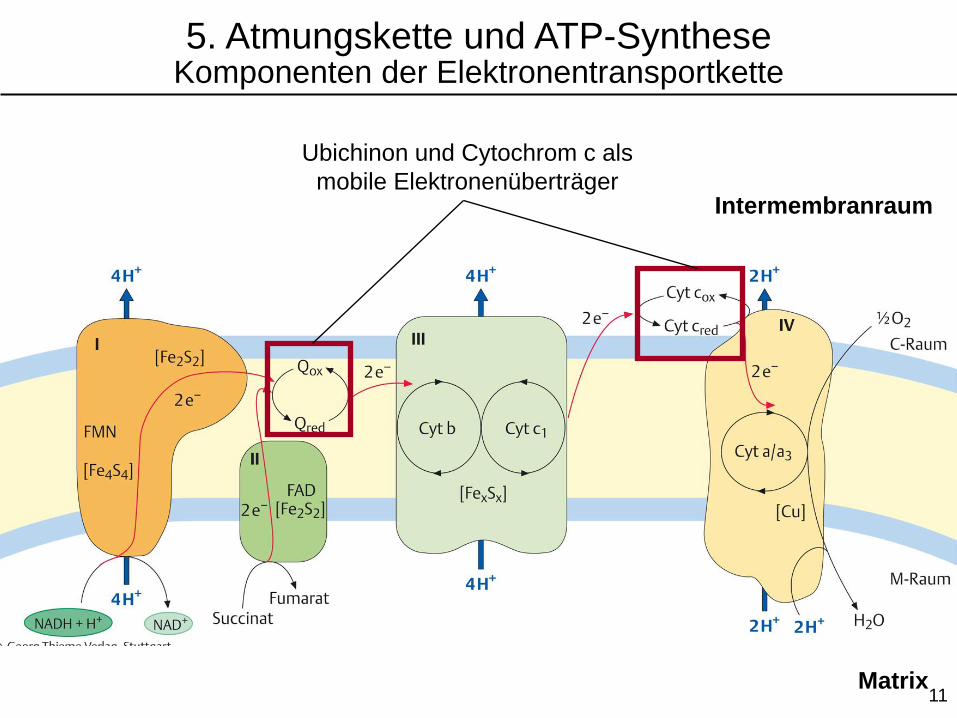
Intermembranraum
Matrix
Ubichinon und Cytochrom c als
mobile Elektronenüberträger
5. Atmungskette und ATP-SyntheseKomponenten der Elektronentransportkette
11
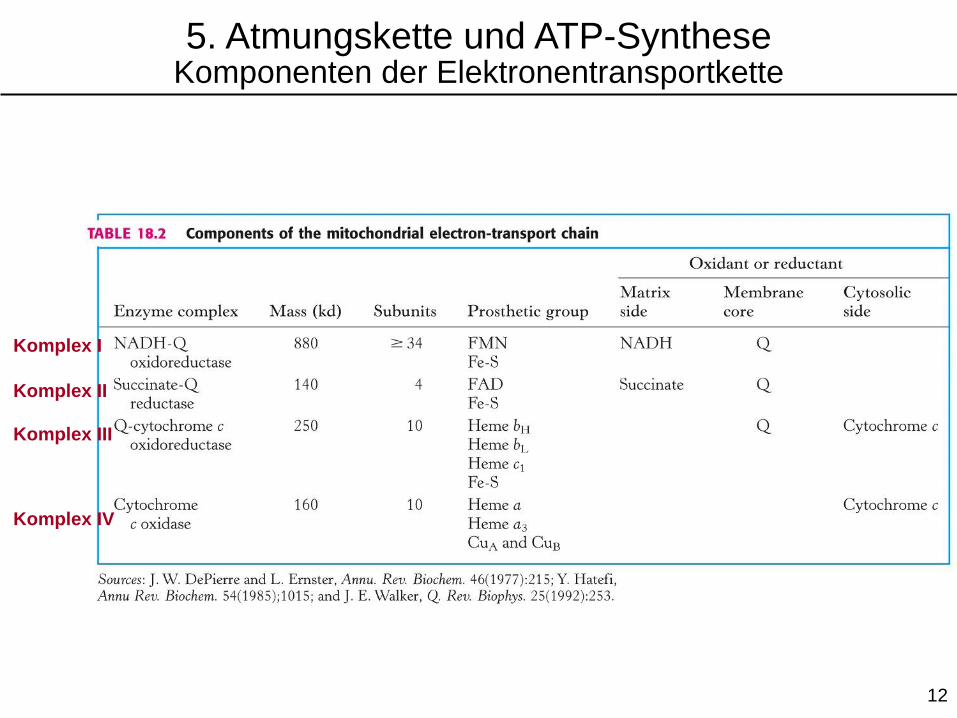
5. Atmungskette und ATP-SyntheseKomponenten der Elektronentransportkette
Komplex I
Komplex II
Komplex III
Komplex IV
12
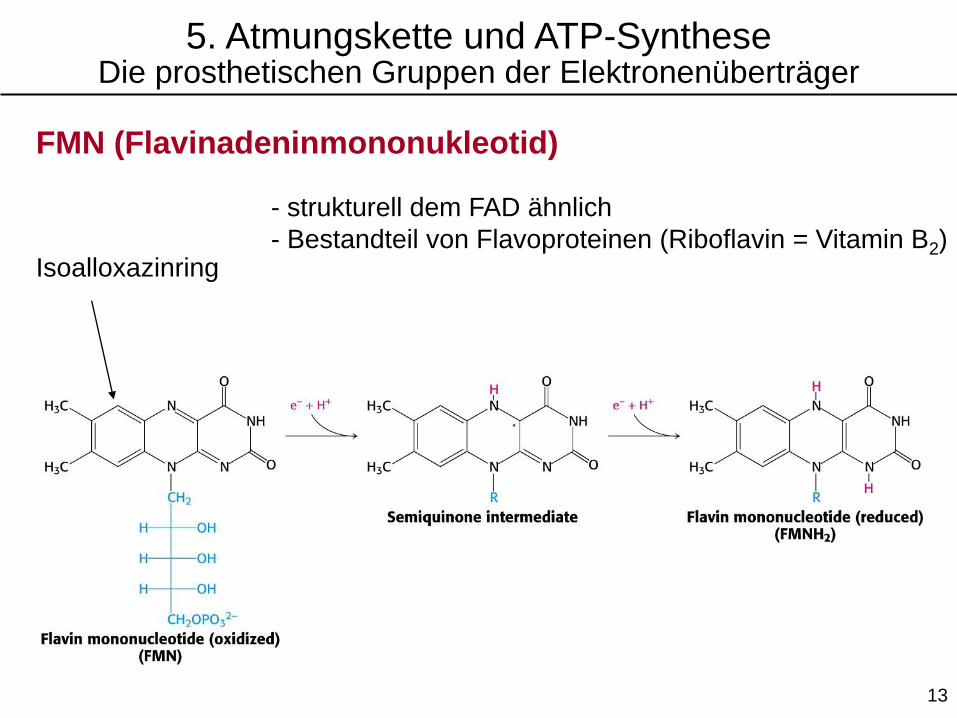
FMN (Flavinadeninmononukleotid)
Isoalloxazinring
- strukturell dem FAD ähnlich
- Bestandteil von Flavoproteinen (Riboflavin = Vitamin B2)
5. Atmungskette und ATP-SyntheseDie prosthetischen Gruppen der Elektronenüberträger
13
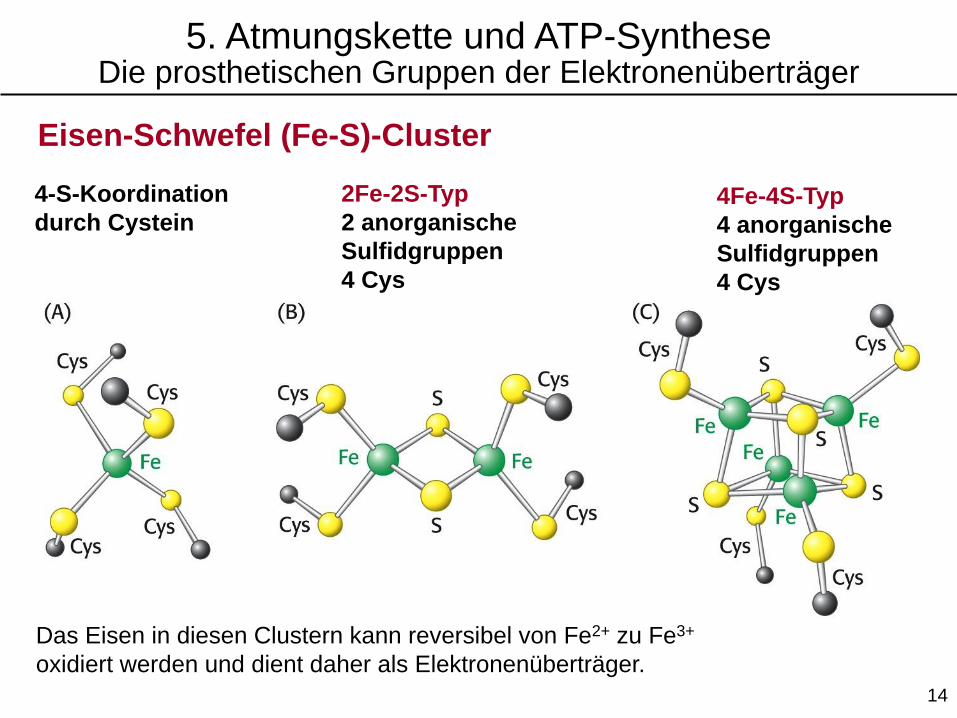
Eisen-Schwefel (Fe-S)-Cluster
4-S-Koordination
durch Cystein
2Fe-2S-Typ
2 anorganische
Sulfidgruppen
4 Cys
4Fe-4S-Typ
4 anorganische
Sulfidgruppen
4 Cys
Das Eisen in diesen Clustern kann reversibel von Fe2+ zu Fe3+
oxidiert werden und dient daher als Elektronenüberträger.
5. Atmungskette und ATP-SyntheseDie prosthetischen Gruppen der Elektronenüberträger
14
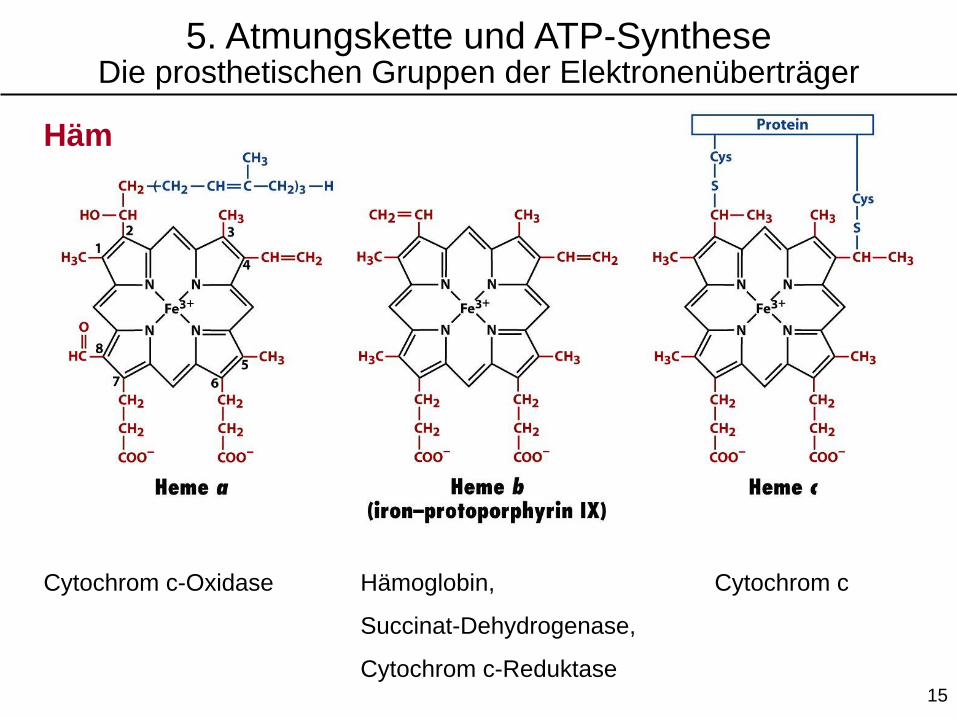
5. Atmungskette und ATP-SyntheseDie prosthetischen Gruppen der Elektronenüberträger
Häm
Cytochrom c-Oxidase Hämoglobin,
Succinat-Dehydrogenase,
Cytochrom c-Reduktase
Cytochrom c
15
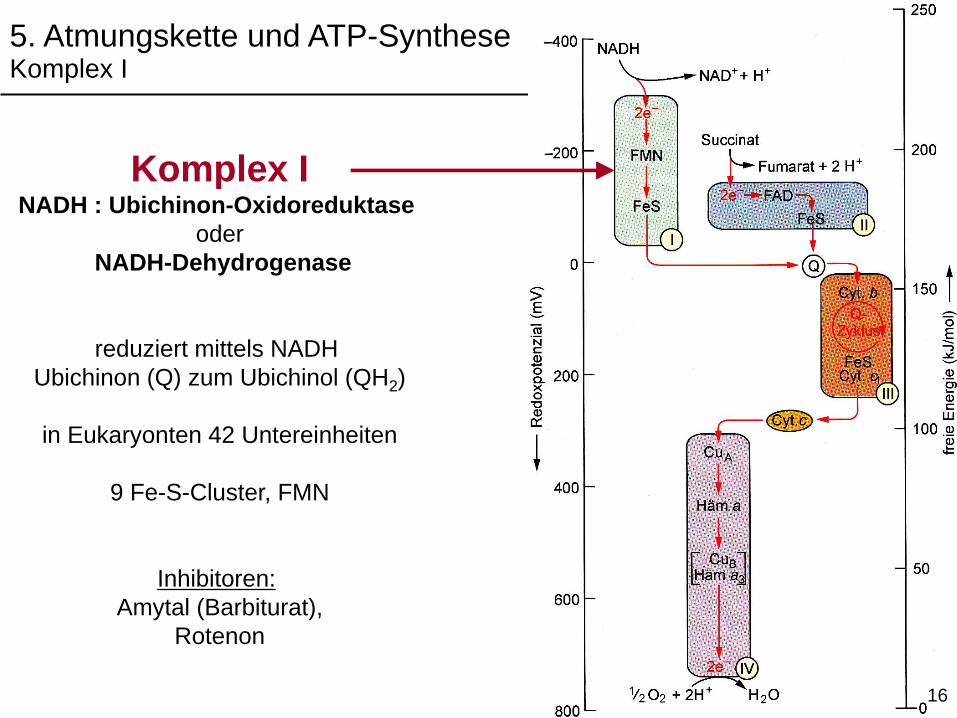
Komplex INADH : Ubichinon-Oxidoreduktase
oder
NADH-Dehydrogenase
reduziert mittels NADH
Ubichinon (Q) zum Ubichinol (QH2)
in Eukaryonten 42 Untereinheiten
9 Fe-S-Cluster, FMN
Inhibitoren:
Amytal (Barbiturat),
Rotenon
5. Atmungskette und ATP-SyntheseKomplex I
16
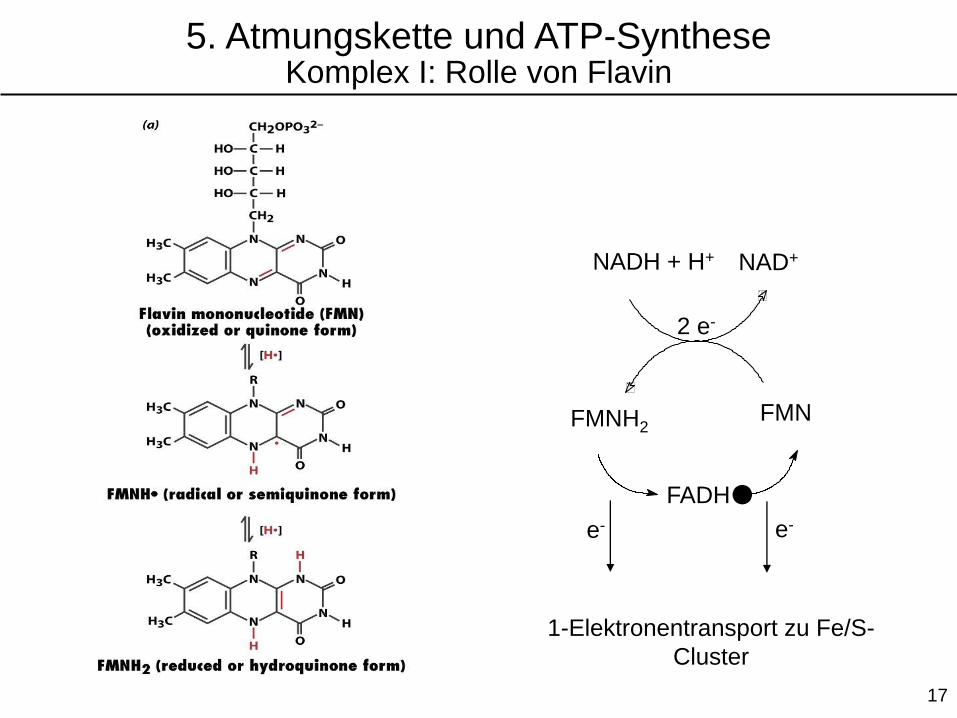
NADH + H+ NAD+
FMNFMNH2
FADH●
1-Elektronentransport zu Fe/S-
Cluster
5. Atmungskette und ATP-SyntheseKomplex I: Rolle von Flavin
17
2 e-
e- e-
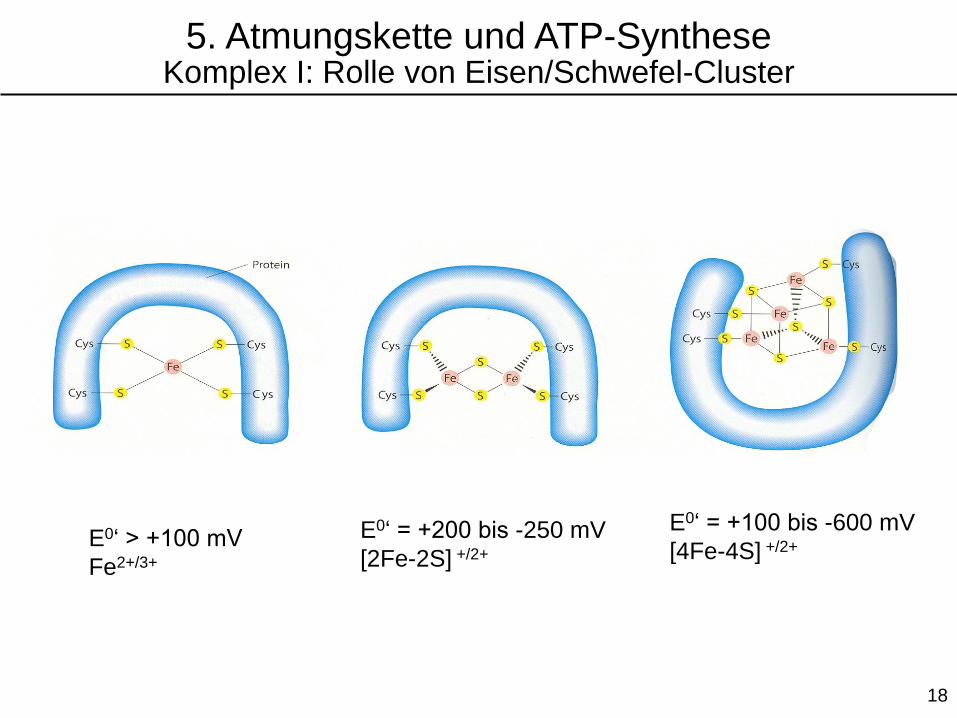
E0‘ > +100 mV
Fe2+/3+
E0‘ = +200 bis -250 mV
[2Fe-2S] +/2+
E0‘ = +100 bis -600 mV
[4Fe-4S] +/2+
5. Atmungskette und ATP-SyntheseKomplex I: Rolle von Eisen/Schwefel-Cluster
18
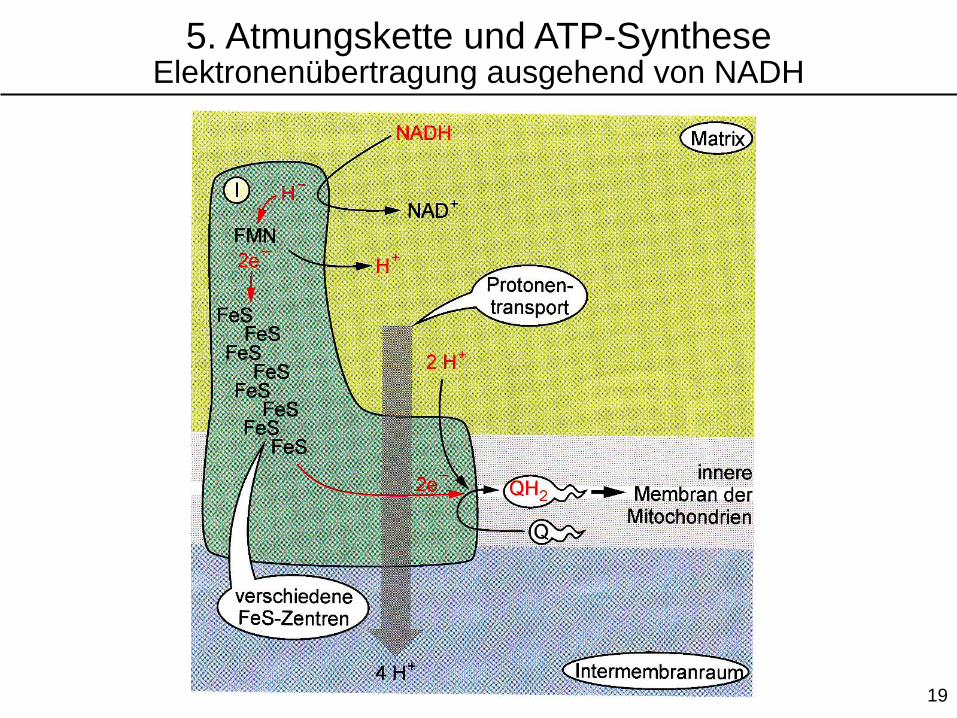
5. Atmungskette und ATP-SyntheseElektronenübertragung ausgehend von NADH
19
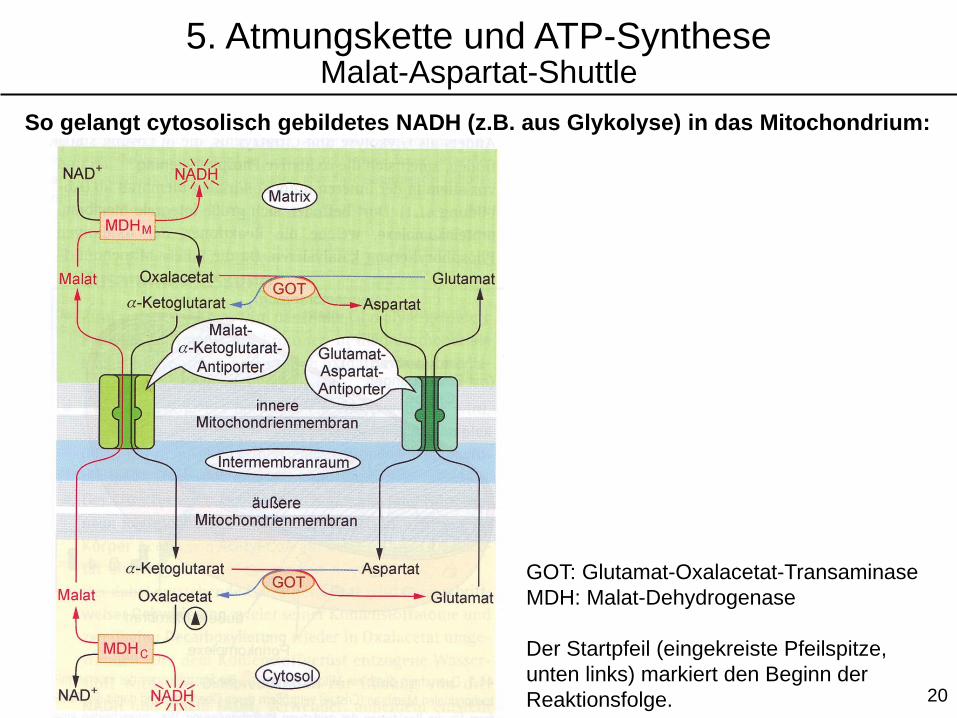
5. Atmungskette und ATP-SyntheseMalat-Aspartat-Shuttle
20
So gelangt cytosolisch gebildetes NADH (z.B. aus Glykolyse) in das Mitochondrium:
GOT: Glutamat-Oxalacetat-Transaminase
MDH: Malat-Dehydrogenase
Der Startpfeil (eingekreiste Pfeilspitze,
unten links) markiert den Beginn der
Reaktionsfolge.
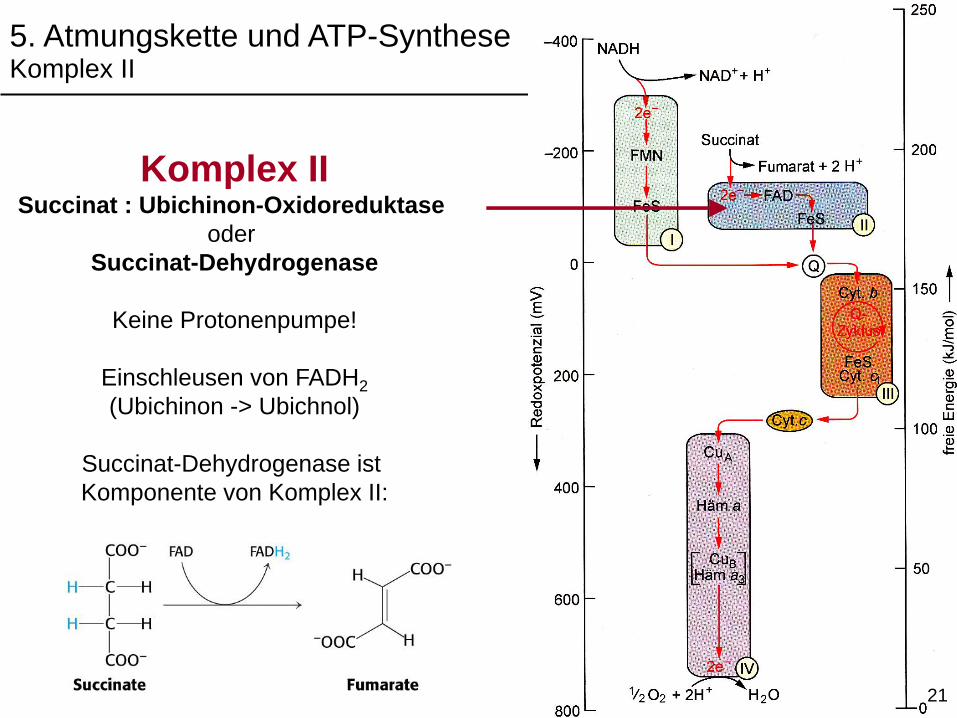
Komplex IISuccinat : Ubichinon-Oxidoreduktase
oder
Succinat-Dehydrogenase
Keine Protonenpumpe!
Einschleusen von FADH2
(Ubichinon -> Ubichnol)
Succinat-Dehydrogenase ist
Komponente von Komplex II:
5. Atmungskette und ATP-SyntheseKomplex II
21
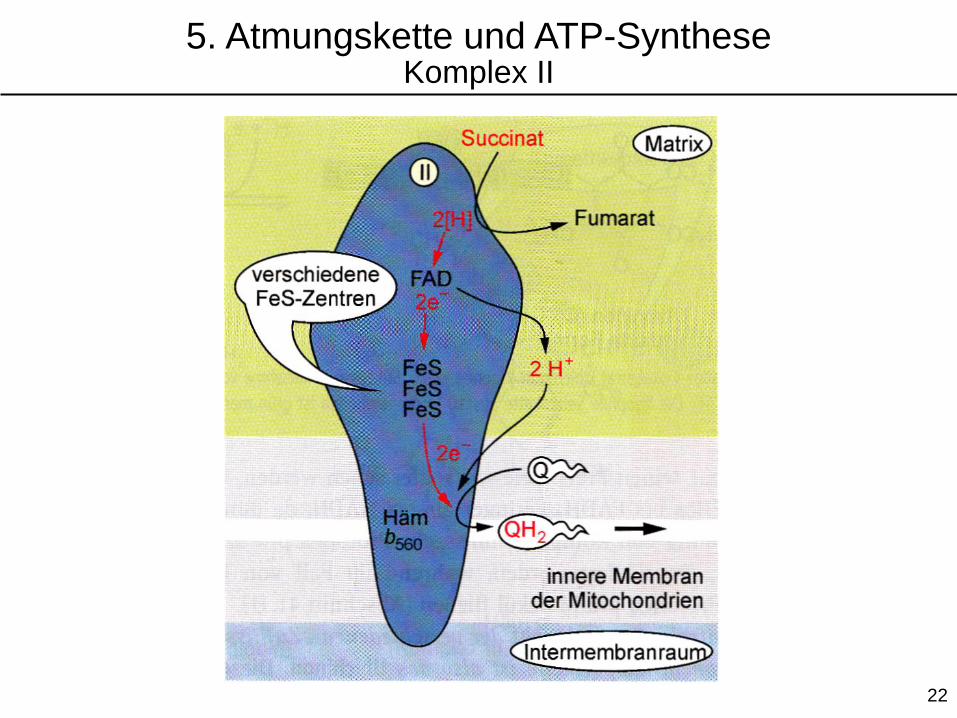
5. Atmungskette und ATP-SyntheseKomplex II
22
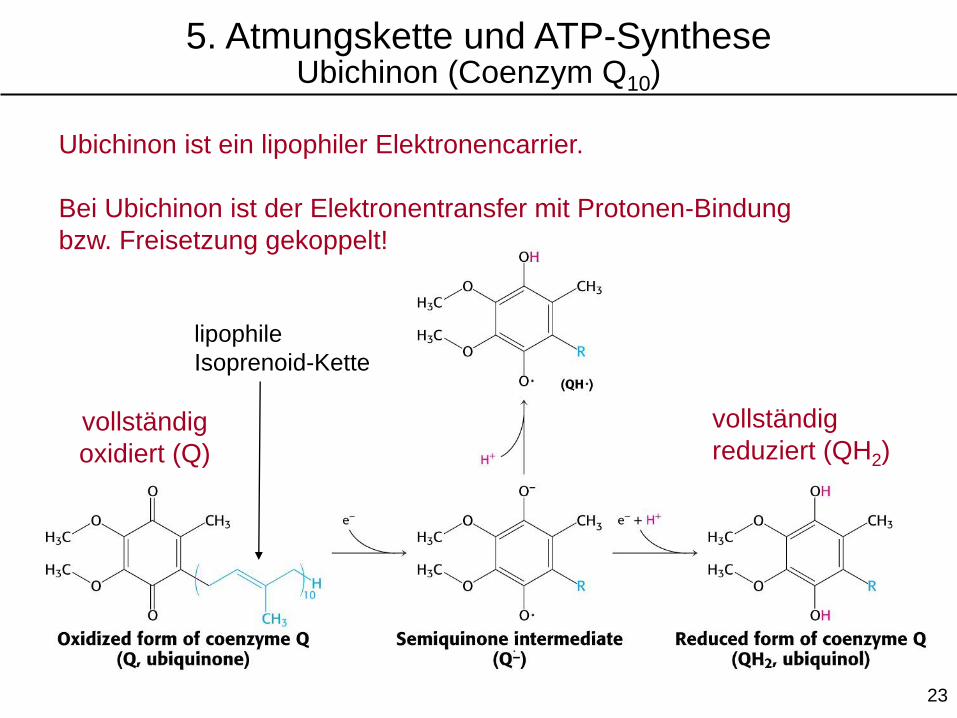
lipophile
Isoprenoid-Kette
vollständig
oxidiert (Q)
vollständig
reduziert (QH2)
Ubichinon ist ein lipophiler Elektronencarrier.
Bei Ubichinon ist der Elektronentransfer mit Protonen-Bindung
bzw. Freisetzung gekoppelt!
5. Atmungskette und ATP-SyntheseUbichinon (Coenzym Q10)
23
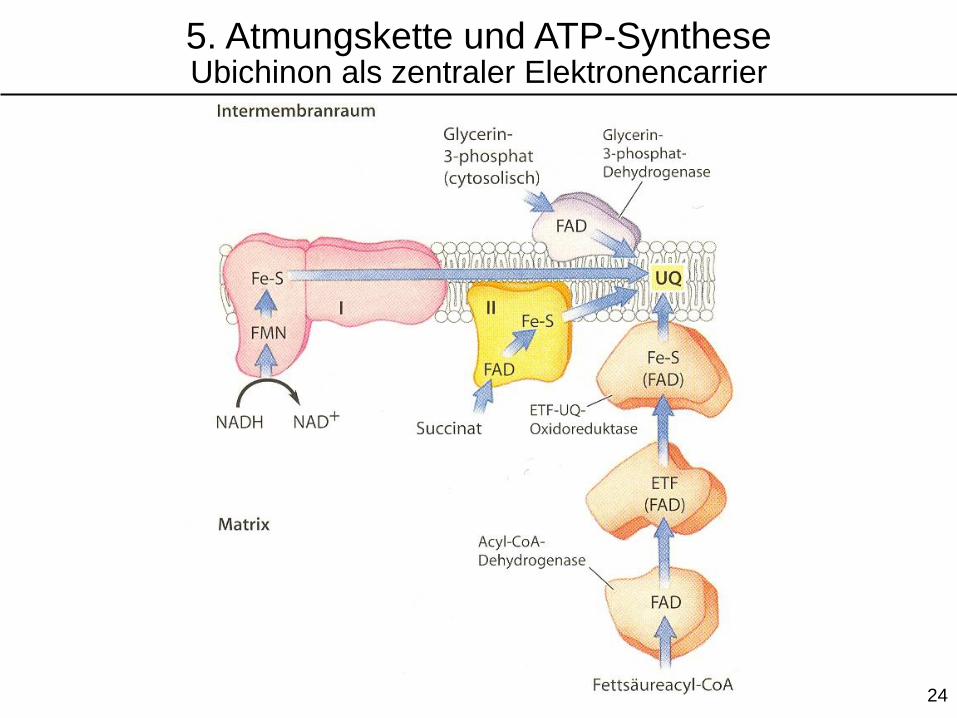
5. Atmungskette und ATP-SyntheseUbichinon als zentraler Elektronencarrier
24
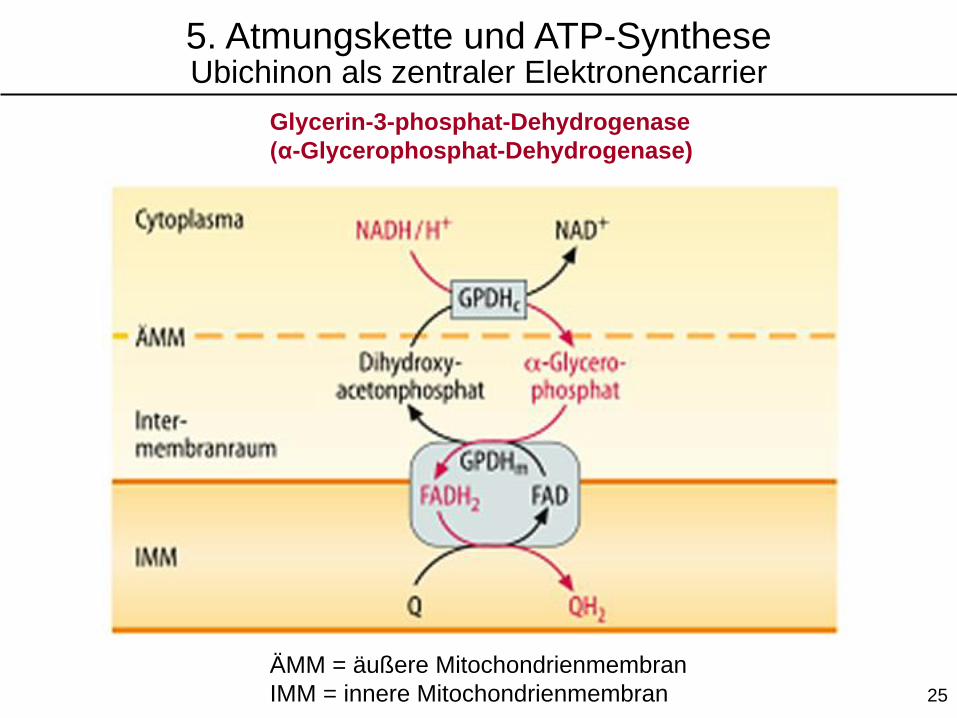
5. Atmungskette und ATP-SyntheseUbichinon als zentraler Elektronencarrier
Glycerin-3-phosphat-Dehydrogenase
(α-Glycerophosphat-Dehydrogenase)
ÄMM = äußere Mitochondrienmembran
IMM = innere Mitochondrienmembran 25
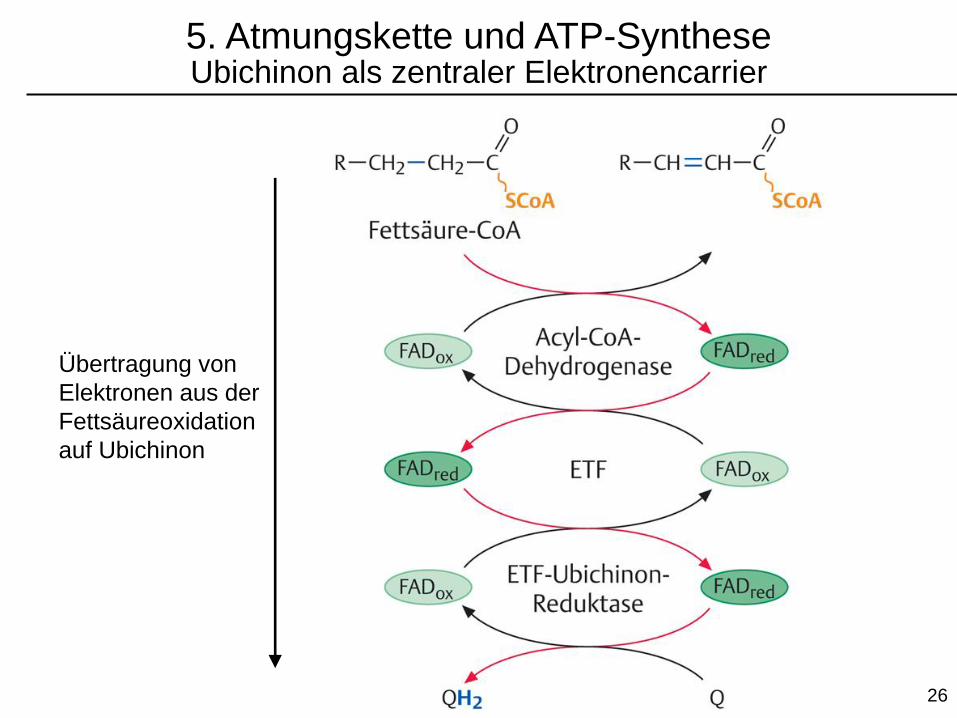
5. Atmungskette und ATP-SyntheseUbichinon als zentraler Elektronencarrier
Übertragung von
Elektronen aus der
Fettsäureoxidation
auf Ubichinon
26
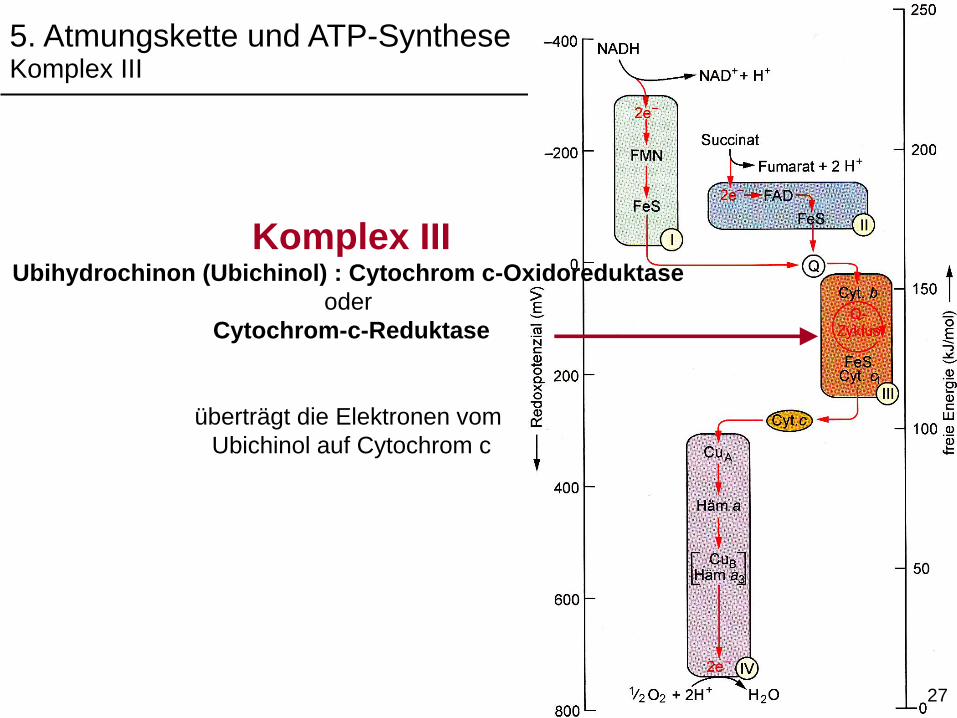
5. Atmungskette und ATP-SyntheseKomplex III
Komplex IIIUbihydrochinon (Ubichinol) : Cytochrom c-Oxidoreduktase
oder
Cytochrom-c-Reduktase
überträgt die Elektronen vom
Ubichinol auf Cytochrom c
27
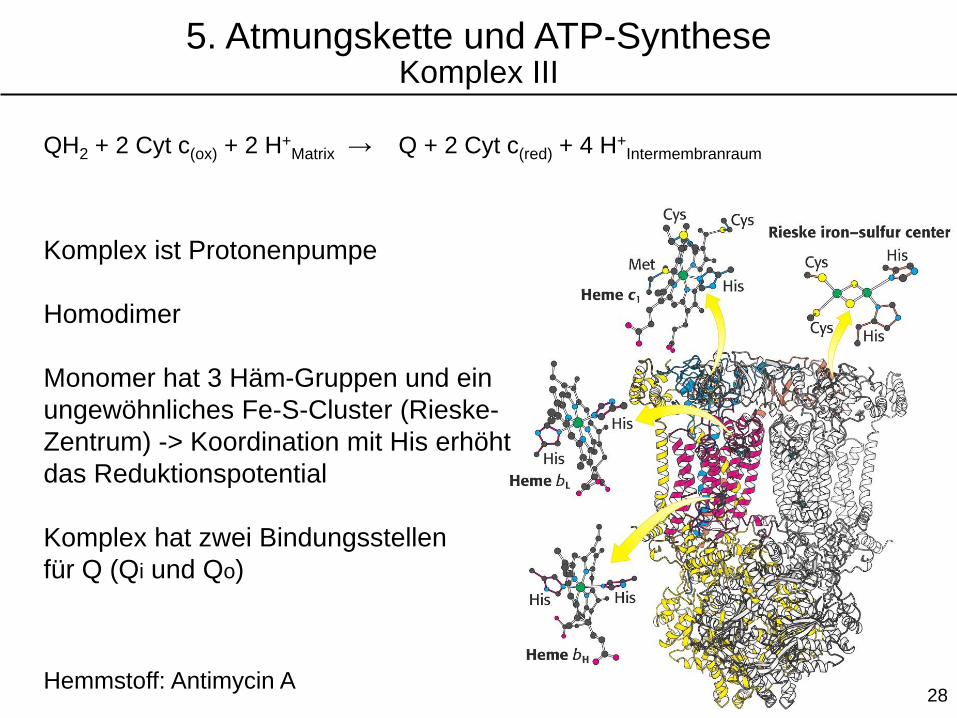
Komplex ist Protonenpumpe
Homodimer
Monomer hat 3 Häm-Gruppen und ein
ungewöhnliches Fe-S-Cluster (Rieske-
Zentrum) -> Koordination mit His erhöht
das Reduktionspotential
Komplex hat zwei Bindungsstellen
für Q (Qi und Qo)
Hemmstoff: Antimycin A
QH2 + 2 Cyt c(ox) + 2 H+Matrix → Q + 2 Cyt c(red) + 4 H+
Intermembranraum
5. Atmungskette und ATP-SyntheseKomplex III
28
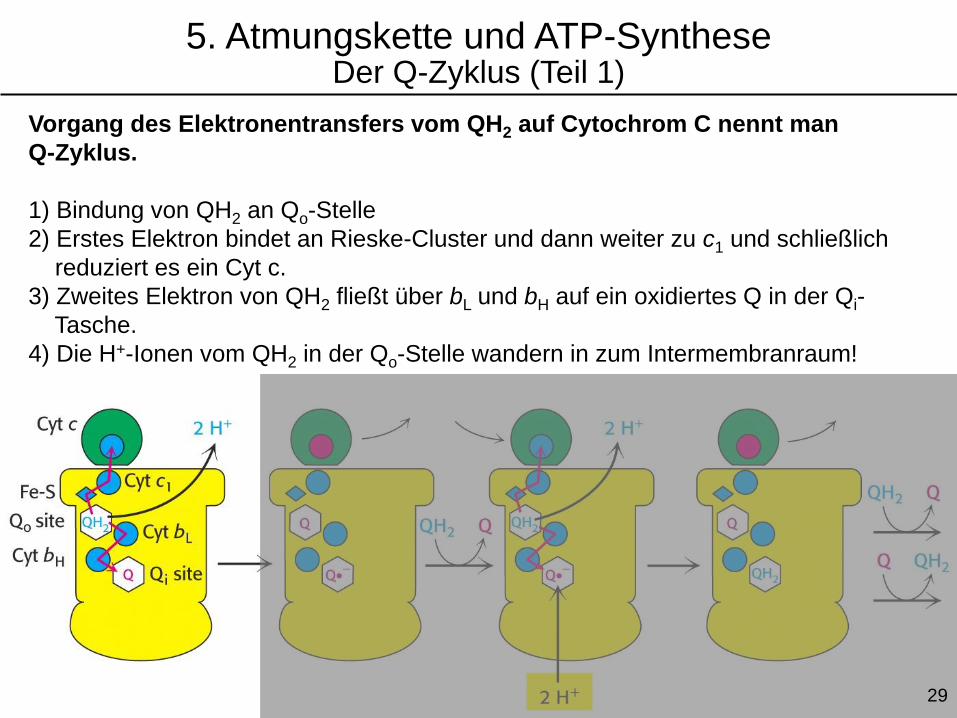
Vorgang des Elektronentransfers vom QH2 auf Cytochrom C nennt man
Q-Zyklus.
1) Bindung von QH2 an Qo-Stelle
2) Erstes Elektron bindet an Rieske-Cluster und dann weiter zu c1 und schließlich
reduziert es ein Cyt c.
3) Zweites Elektron von QH2 fließt über bL und bH auf ein oxidiertes Q in der Qi-
Tasche.
4) Die H+-Ionen vom QH2 in der Qo-Stelle wandern in zum Intermembranraum!
5. Atmungskette und ATP-SyntheseDer Q-Zyklus (Teil 1)
29
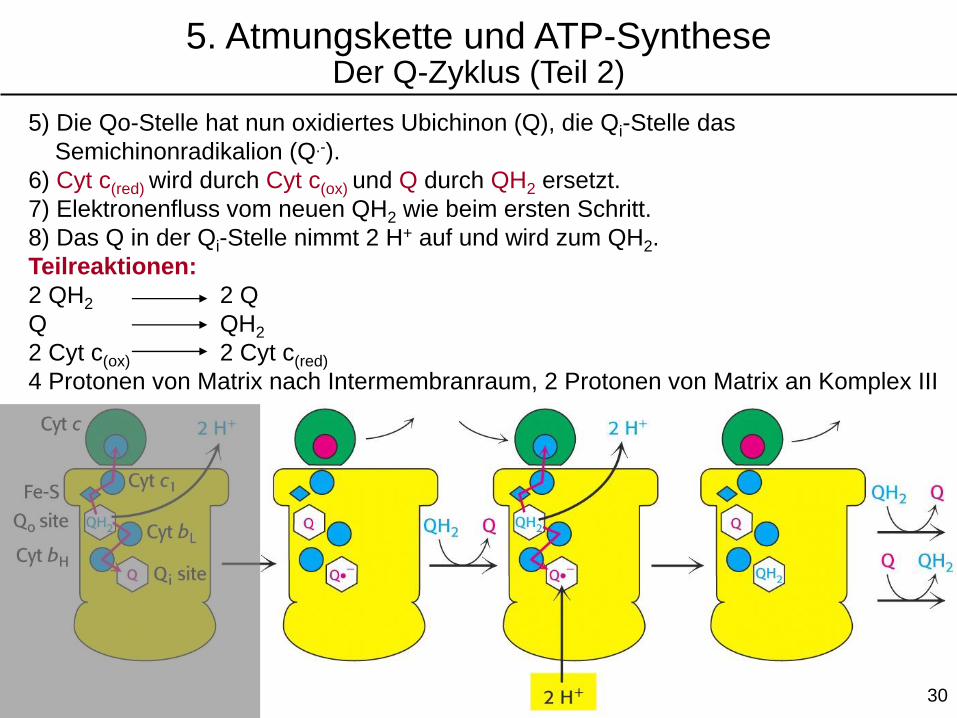
5) Die Qo-Stelle hat nun oxidiertes Ubichinon (Q), die Qi-Stelle das
Semichinonradikalion (Q.-).
6) Cyt c(red) wird durch Cyt c(ox) und Q durch QH2 ersetzt.
7) Elektronenfluss vom neuen QH2 wie beim ersten Schritt.
8) Das Q in der Qi-Stelle nimmt 2 H+ auf und wird zum QH2.
Teilreaktionen:
2 QH2 2 Q
Q QH2
2 Cyt c(ox) 2 Cyt c(red)
4 Protonen von Matrix nach Intermembranraum, 2 Protonen von Matrix an Komplex III
5. Atmungskette und ATP-SyntheseDer Q-Zyklus (Teil 2)
30
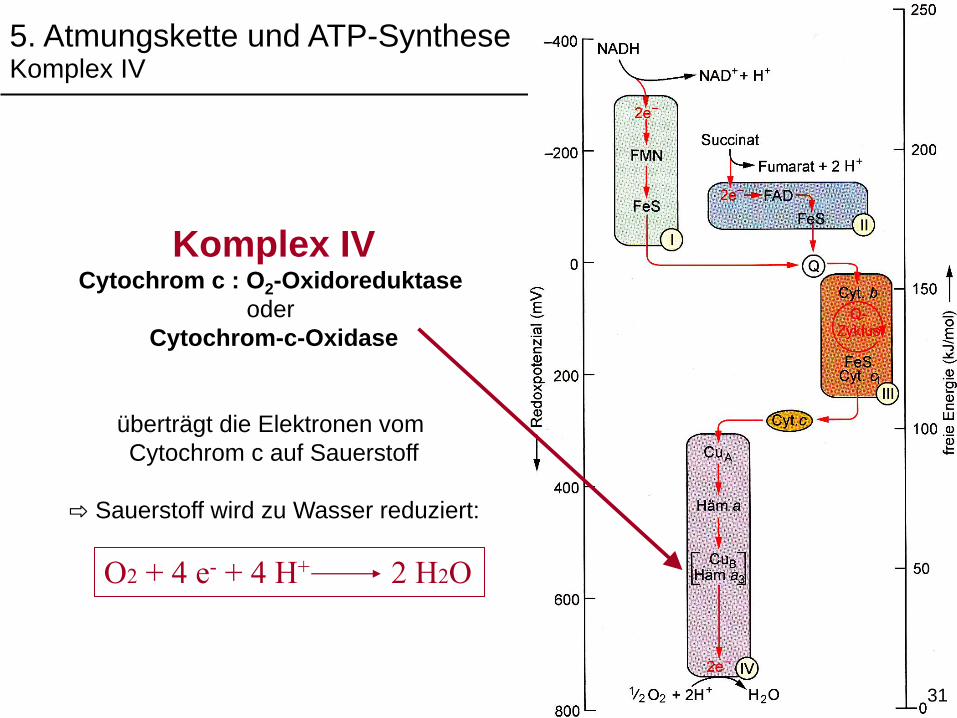
5. Atmungskette und ATP-SyntheseKomplex IV
Komplex IVCytochrom c : O2-Oxidoreduktase
oder
Cytochrom-c-Oxidase
überträgt die Elektronen vom
Cytochrom c auf Sauerstoff
⇨ Sauerstoff wird zu Wasser reduziert:
O2 + 4 e- + 4 H+ 2 H2O
31
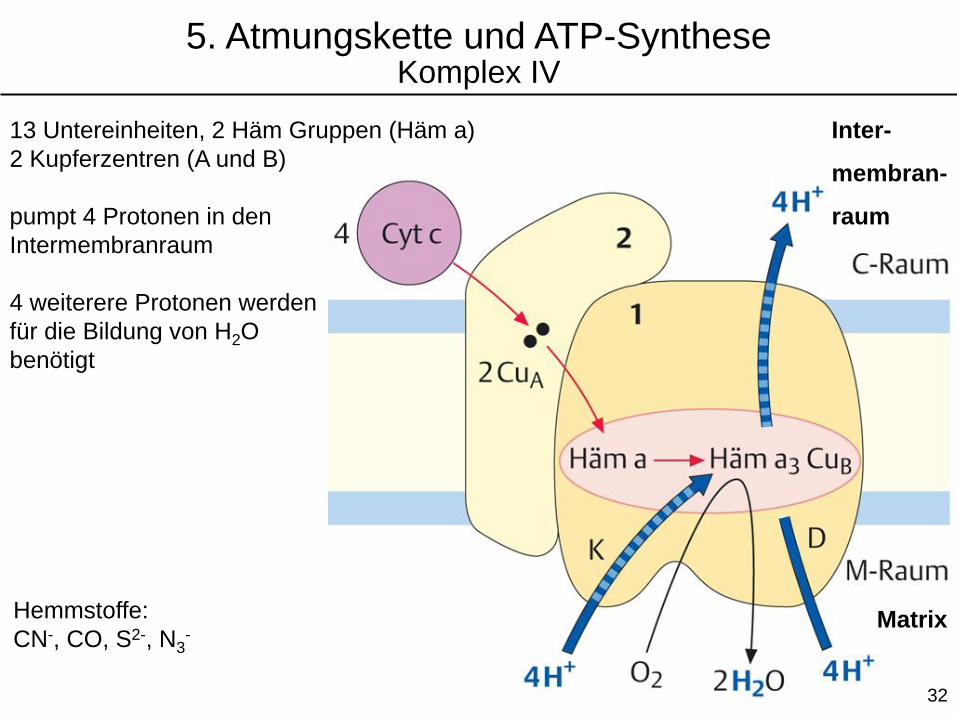
Hemmstoffe:
CN-, CO, S2-, N3-
13 Untereinheiten, 2 Häm Gruppen (Häm a)
2 Kupferzentren (A und B)
pumpt 4 Protonen in den
Intermembranraum
4 weiterere Protonen werden
für die Bildung von H2O
benötigt
5. Atmungskette und ATP-SyntheseKomplex IV
Inter-
membran-
raum
Matrix
32
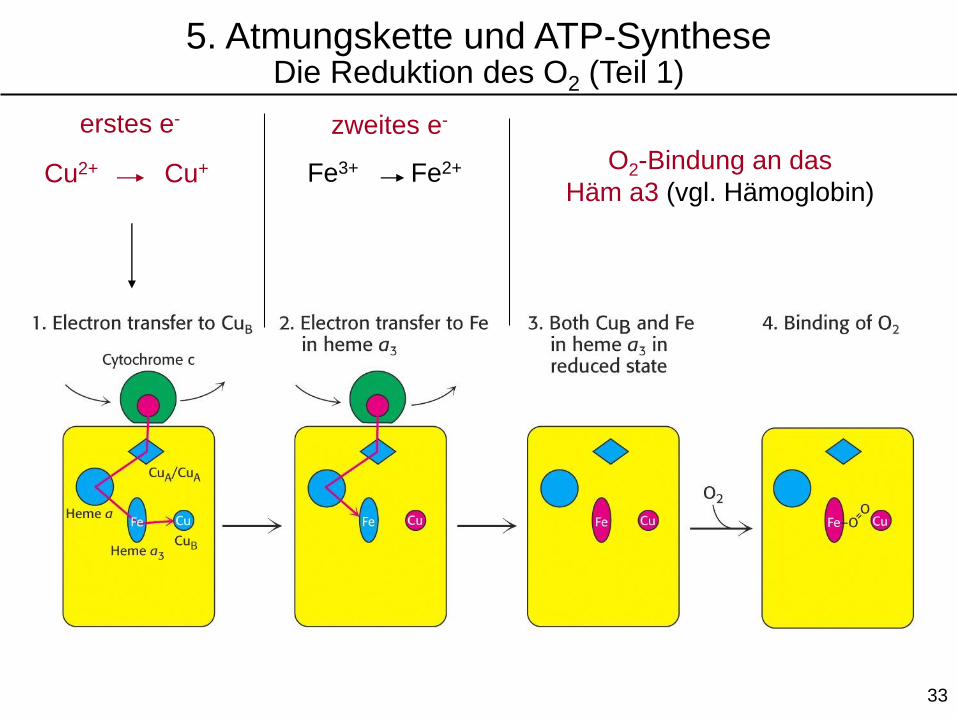
Cu2+ Cu+
erstes e- zweites e-
Fe3+ Fe2+ O2-Bindung an das
Häm a3 (vgl. Hämoglobin)
5. Atmungskette und ATP-SyntheseDie Reduktion des O2 (Teil 1)
33
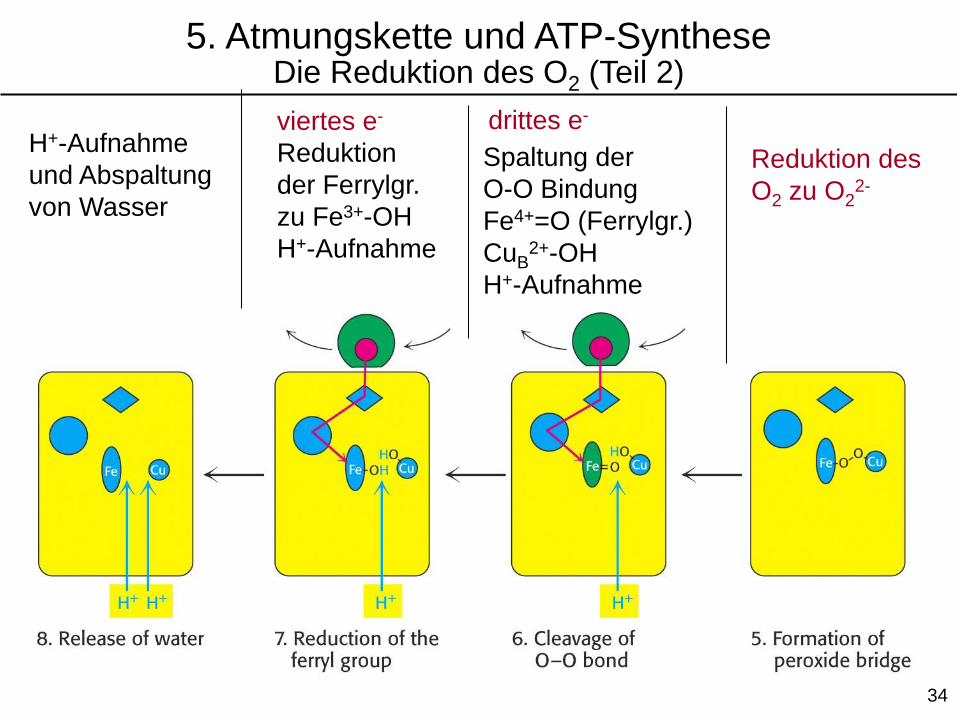
Reduktion des
O2 zu O22-
drittes e-
Spaltung der
O-O Bindung
Fe4+=O (Ferrylgr.)
CuB2+-OH
H+-Aufnahme
viertes e-
Reduktion
der Ferrylgr.
zu Fe3+-OH
H+-Aufnahme
H+-Aufnahme
und Abspaltung
von Wasser
5. Atmungskette und ATP-SyntheseDie Reduktion des O2 (Teil 2)
34
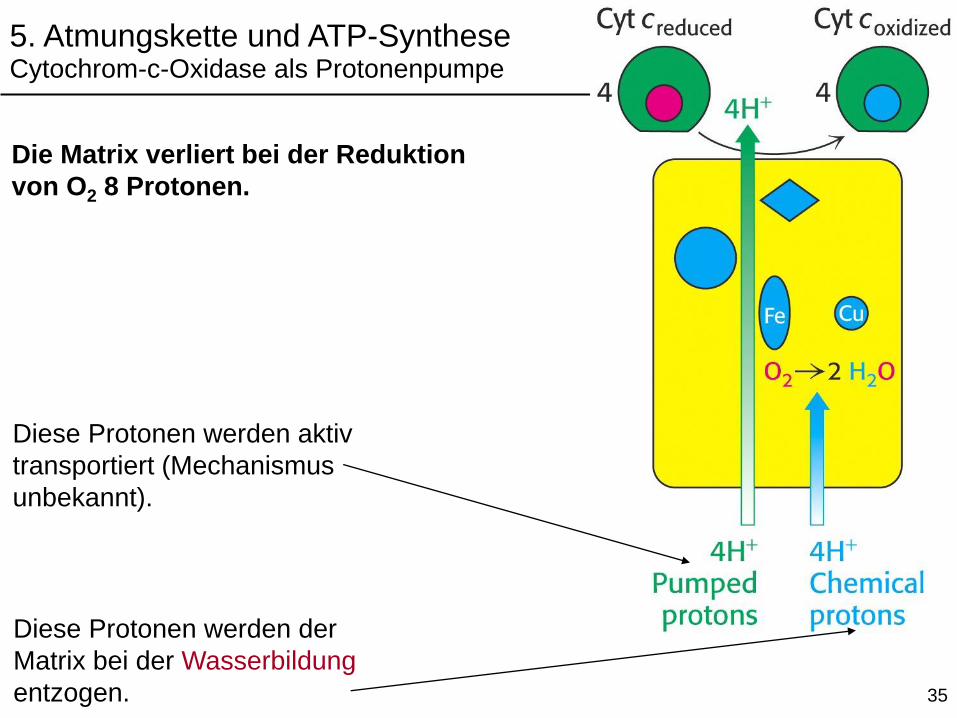
Diese Protonen werden der
Matrix bei der Wasserbildung
entzogen.
Diese Protonen werden aktiv
transportiert (Mechanismus
unbekannt).
Die Matrix verliert bei der Reduktion
von O2 8 Protonen.
5. Atmungskette und ATP-SyntheseCytochrom-c-Oxidase als Protonenpumpe
35
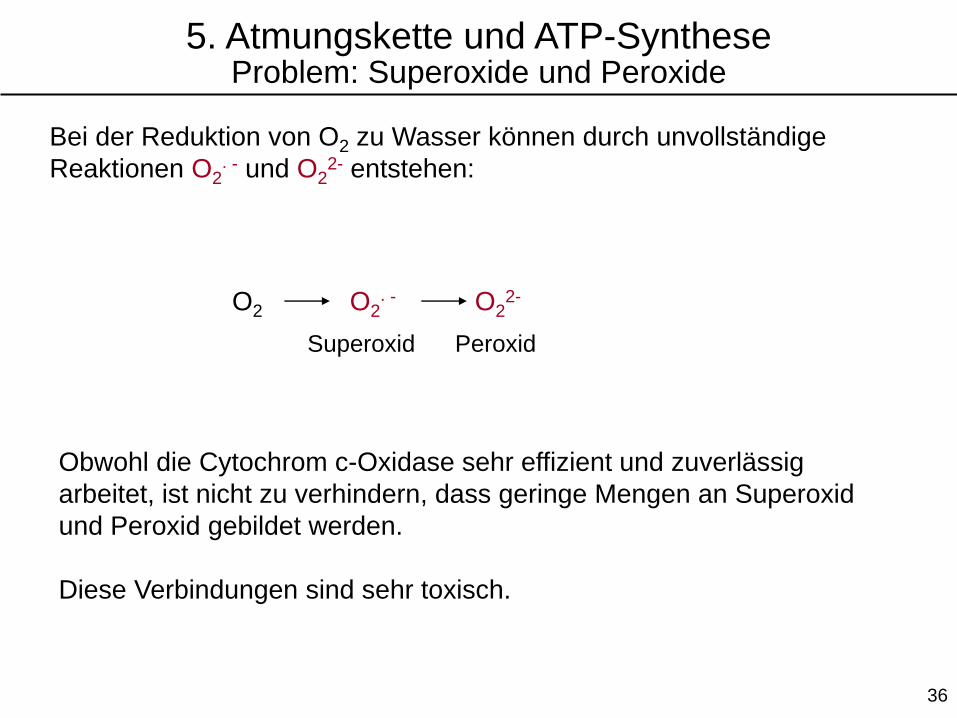
Bei der Reduktion von O2 zu Wasser können durch unvollständige
Reaktionen O2∙ - und O2
2- entstehen:
O2 O2∙ - O2
2-
Superoxid Peroxid
Obwohl die Cytochrom c-Oxidase sehr effizient und zuverlässig
arbeitet, ist nicht zu verhindern, dass geringe Mengen an Superoxid
und Peroxid gebildet werden.
Diese Verbindungen sind sehr toxisch.
5. Atmungskette und ATP-SyntheseProblem: Superoxide und Peroxide
36
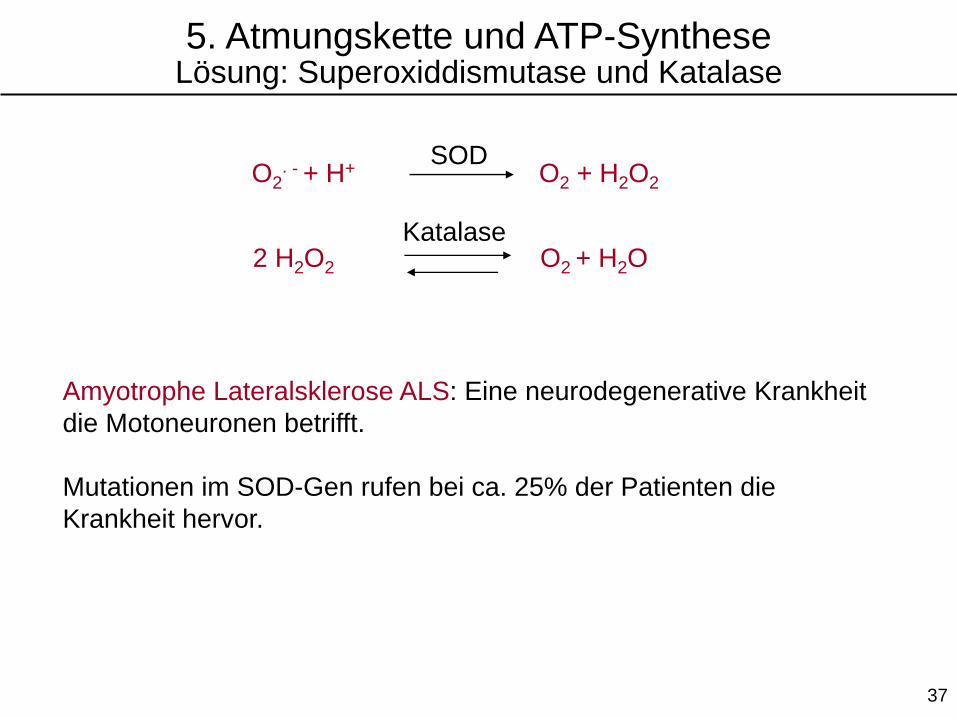
O2∙ - + H+ O2 + H2O2
SOD
2 H2O2 O2 + H2OKatalase
Amyotrophe Lateralsklerose ALS: Eine neurodegenerative Krankheit
die Motoneuronen betrifft.
Mutationen im SOD-Gen rufen bei ca. 25% der Patienten die
Krankheit hervor.
5. Atmungskette und ATP-SyntheseLösung: Superoxiddismutase und Katalase
37
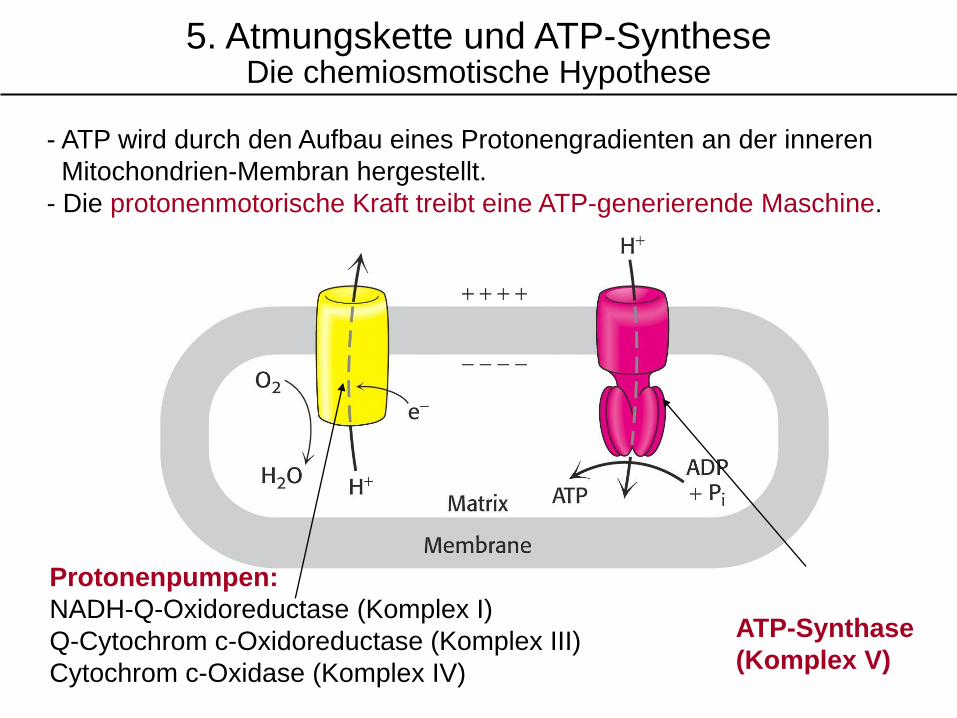
ATP-Synthase
(Komplex V)
Protonenpumpen:
NADH-Q-Oxidoreductase (Komplex I)
Q-Cytochrom c-Oxidoreductase (Komplex III)
Cytochrom c-Oxidase (Komplex IV)
- ATP wird durch den Aufbau eines Protonengradienten an der inneren
Mitochondrien-Membran hergestellt.
- Die protonenmotorische Kraft treibt eine ATP-generierende Maschine.
5. Atmungskette und ATP-SyntheseDie chemiosmotische Hypothese
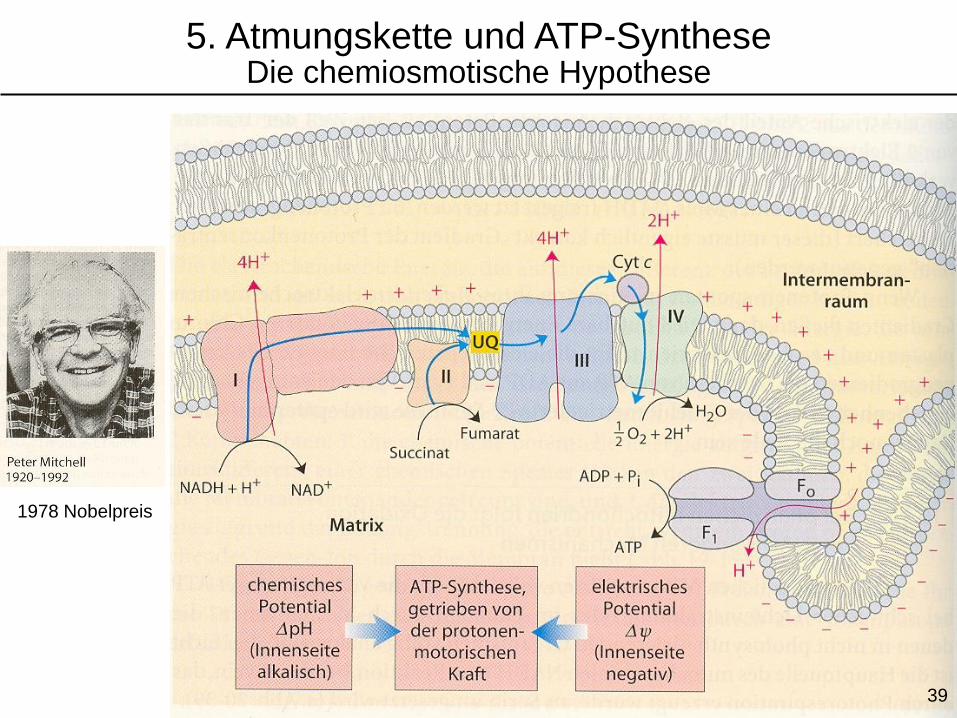
1978 Nobelpreis
5. Atmungskette und ATP-SyntheseDie chemiosmotische Hypothese
39
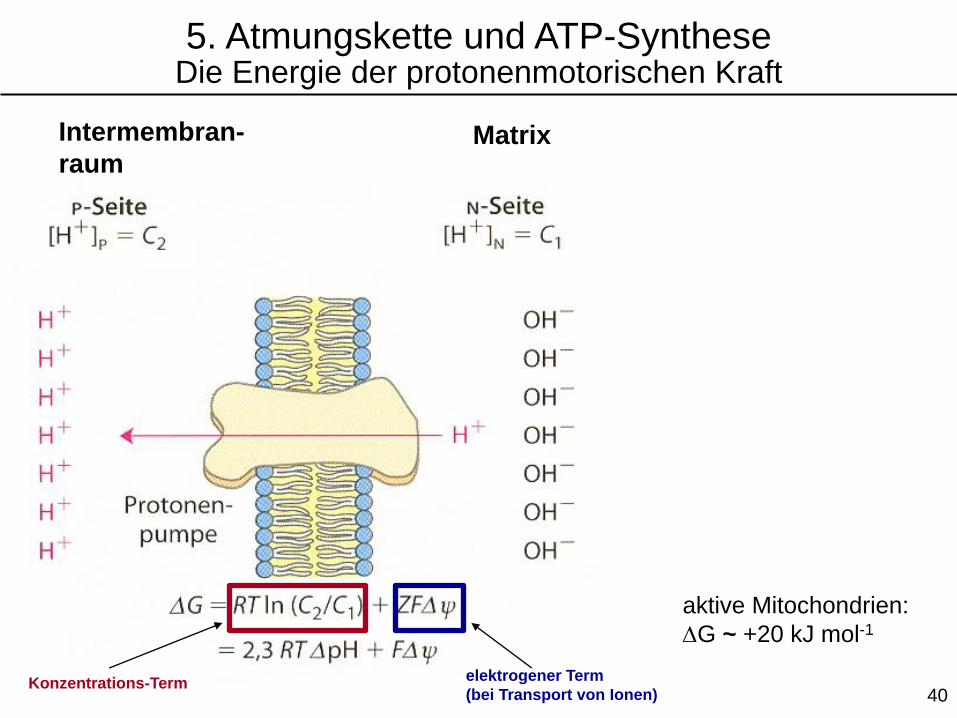
5. Atmungskette und ATP-SyntheseDie Energie der protonenmotorischen Kraft
Intermembran-
raumMatrix
Konzentrations-Termelektrogener Term
(bei Transport von Ionen)
aktive Mitochondrien:
DG ~ +20 kJ mol-1
40
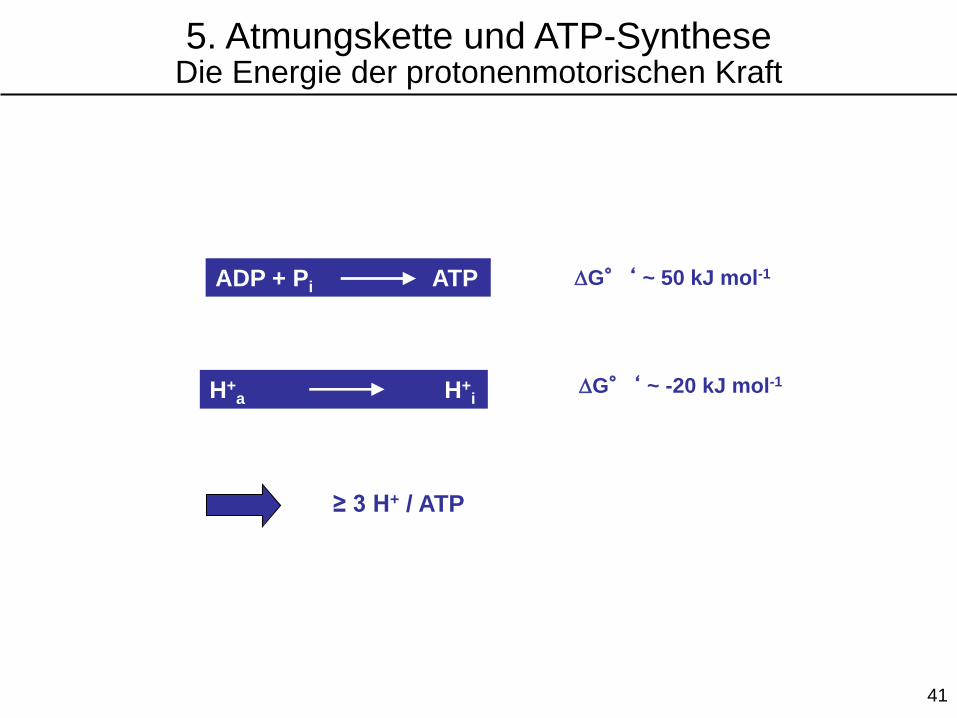
ADP + Pi ATP DG°‘ ~ 50 kJ mol-1
H+a H+
iDG°‘ ~ -20 kJ mol-1
≥ 3 H+ / ATP
5. Atmungskette und ATP-SyntheseDie Energie der protonenmotorischen Kraft
41
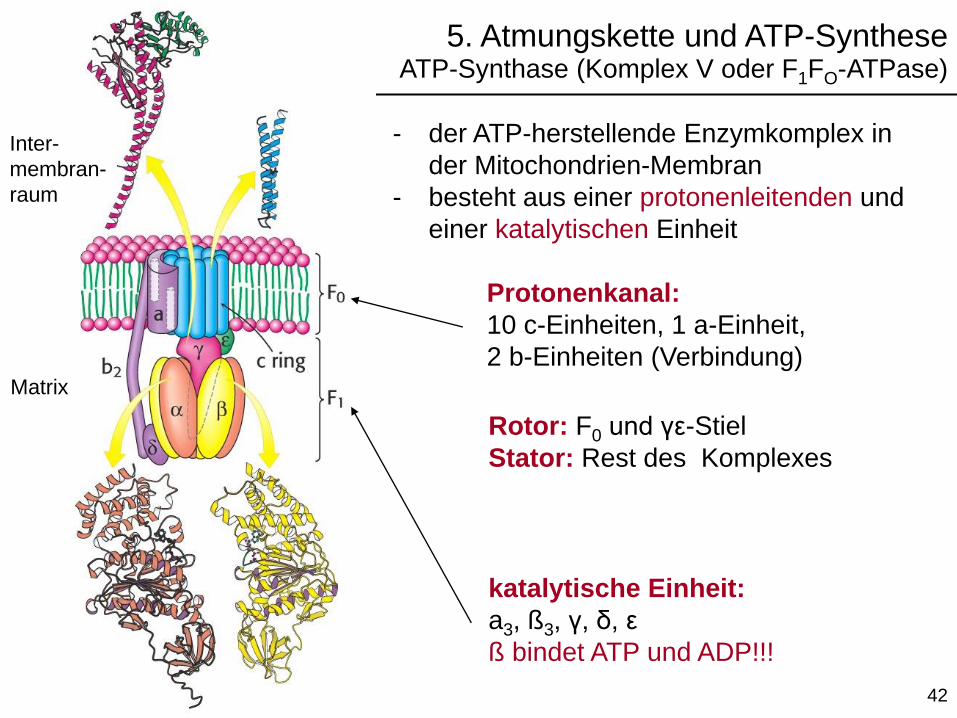
- der ATP-herstellende Enzymkomplex in
der Mitochondrien-Membran
- besteht aus einer protonenleitenden und
einer katalytischen Einheit
katalytische Einheit:
a3, ß3, γ, δ, ε
ß bindet ATP und ADP!!!
Inter-
membran-
raum
Protonenkanal:
10 c-Einheiten, 1 a-Einheit,
2 b-Einheiten (Verbindung)
Rotor: F0 und γε-Stiel
Stator: Rest des Komplexes
5. Atmungskette und ATP-SyntheseATP-Synthase (Komplex V oder F1FO-ATPase)
Matrix
42
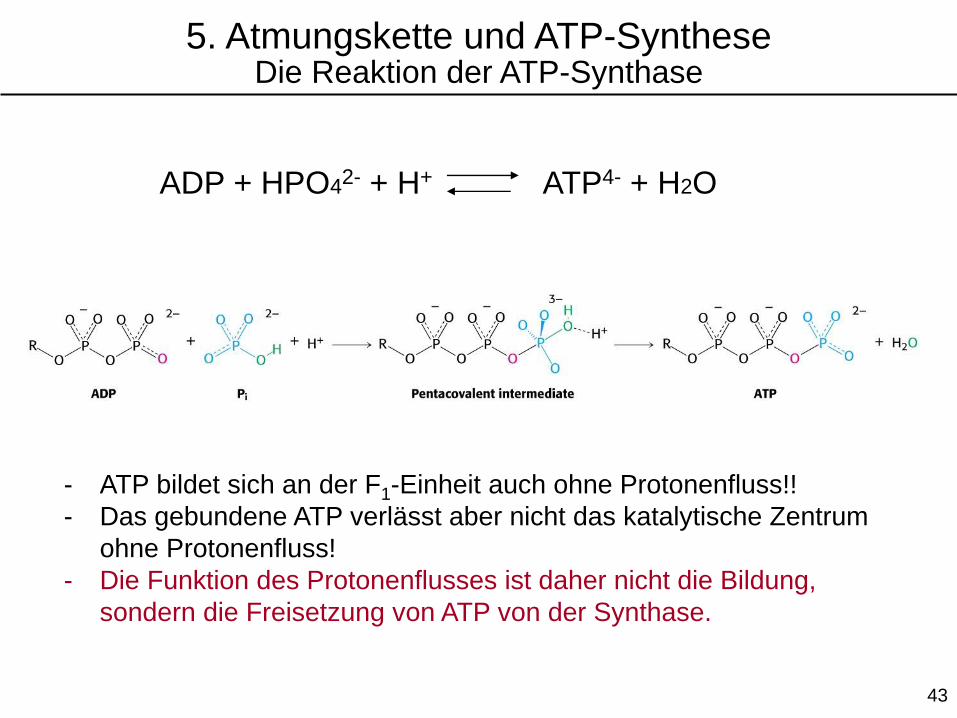
ADP + HPO42- + H+ ATP4- + H2O
- ATP bildet sich an der F1-Einheit auch ohne Protonenfluss!!
- Das gebundene ATP verlässt aber nicht das katalytische Zentrum
ohne Protonenfluss!
- Die Funktion des Protonenflusses ist daher nicht die Bildung,
sondern die Freisetzung von ATP von der Synthase.
5. Atmungskette und ATP-SyntheseDie Reaktion der ATP-Synthase
43
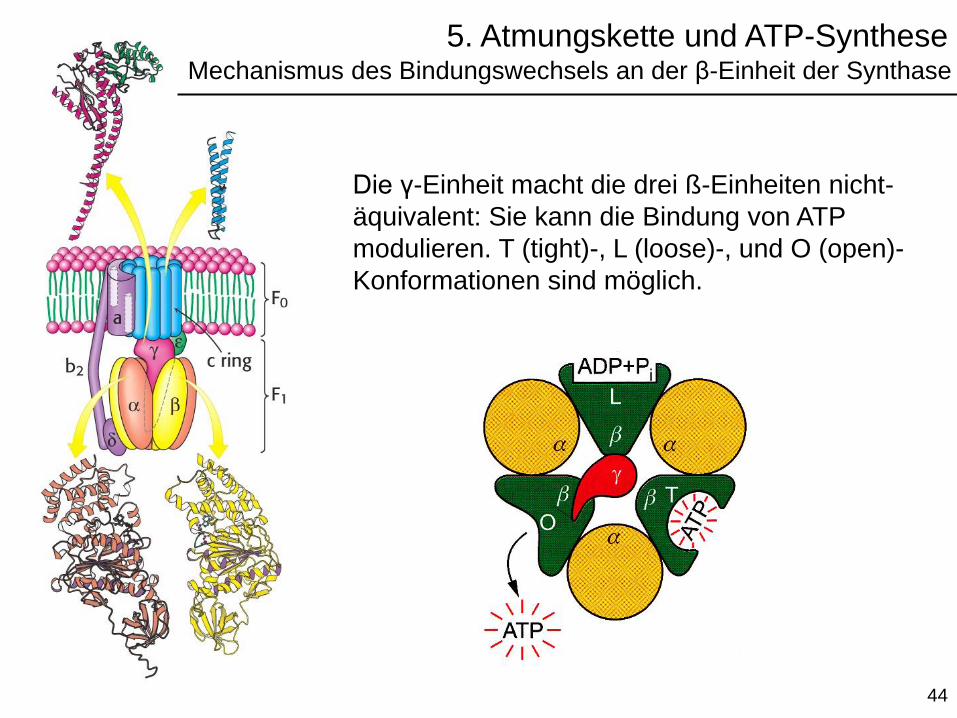
Die γ-Einheit macht die drei ß-Einheiten nicht-
äquivalent: Sie kann die Bindung von ATP
modulieren. T (tight)-, L (loose)-, und O (open)-
Konformationen sind möglich.
5. Atmungskette und ATP-SyntheseMechanismus des Bindungswechsels an der β-Einheit der Synthase
44
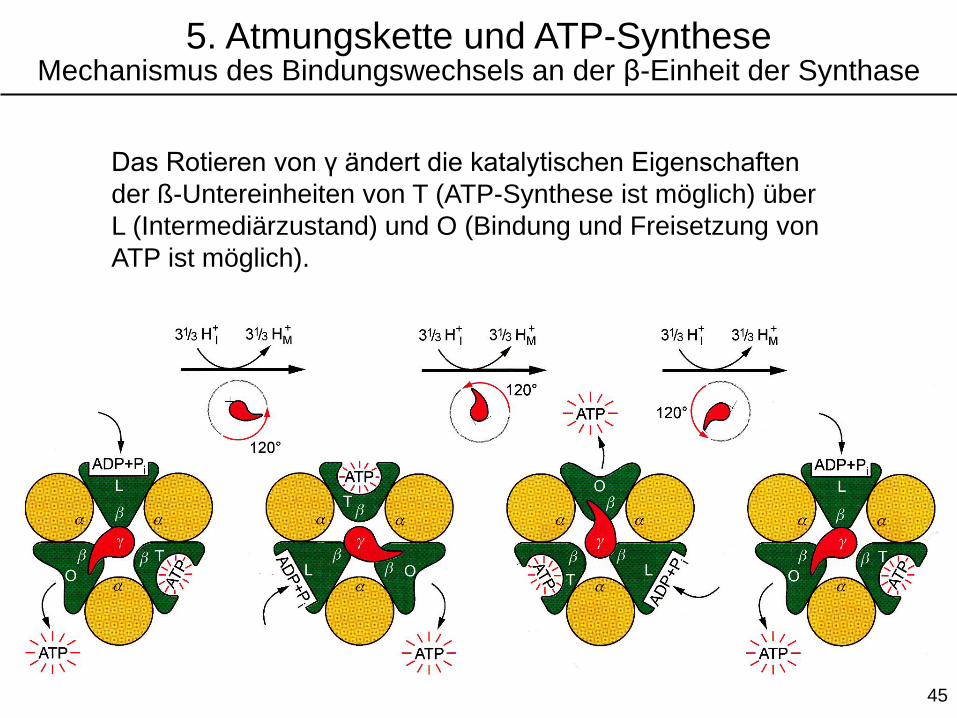
Das Rotieren von γ ändert die katalytischen Eigenschaften
der ß-Untereinheiten von T (ATP-Synthese ist möglich) über
L (Intermediärzustand) und O (Bindung und Freisetzung von
ATP ist möglich).
5. Atmungskette und ATP-SyntheseMechanismus des Bindungswechsels an der β-Einheit der Synthase
45
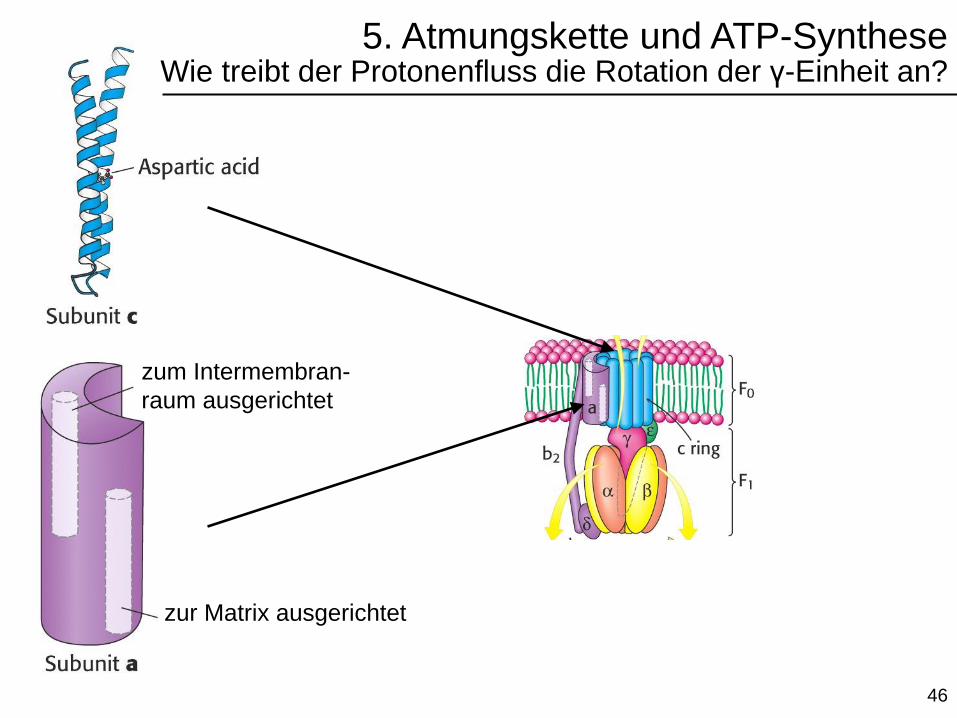
5. Atmungskette und ATP-SyntheseWie treibt der Protonenfluss die Rotation der γ-Einheit an?
zur Matrix ausgerichtet
zum Intermembran-
raum ausgerichtet
46
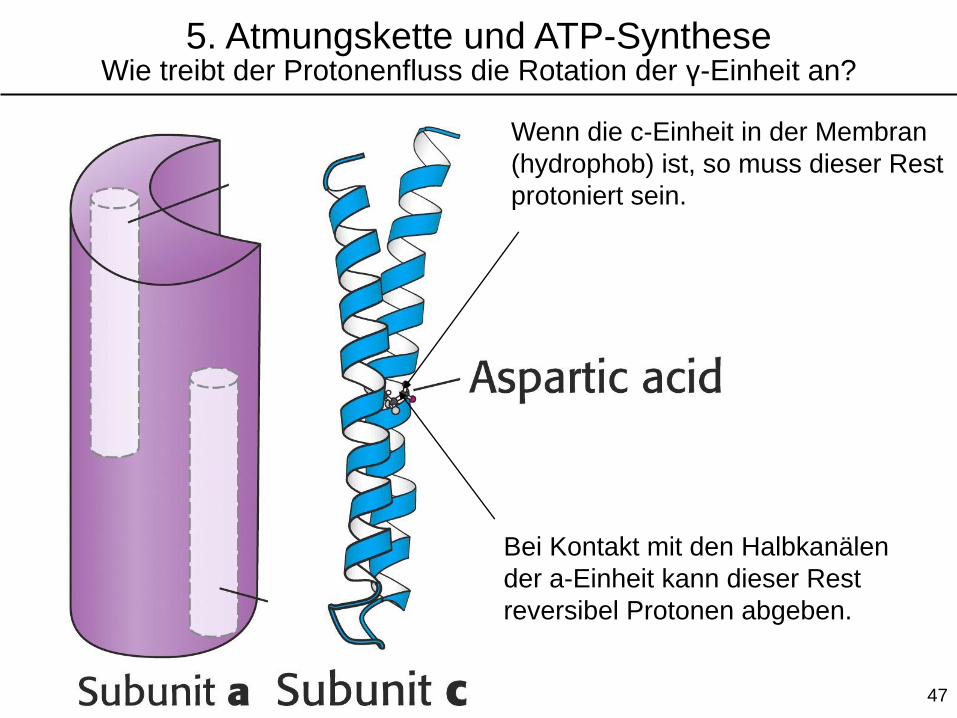
Wenn die c-Einheit in der Membran
(hydrophob) ist, so muss dieser Rest
protoniert sein.
Bei Kontakt mit den Halbkanälen
der a-Einheit kann dieser Rest
reversibel Protonen abgeben.
5. Atmungskette und ATP-SyntheseWie treibt der Protonenfluss die Rotation der γ-Einheit an?
47
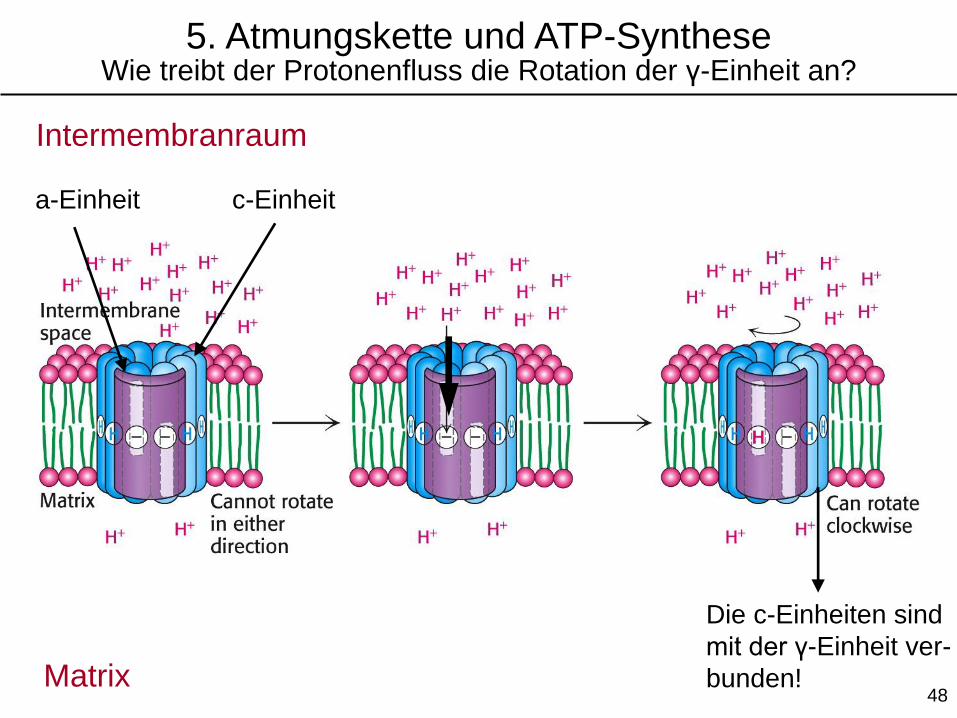
Die c-Einheiten sind
mit der γ-Einheit ver-
bunden!
5. Atmungskette und ATP-SyntheseWie treibt der Protonenfluss die Rotation der γ-Einheit an?
Intermembranraum
Matrix
c-Einheita-Einheit
48
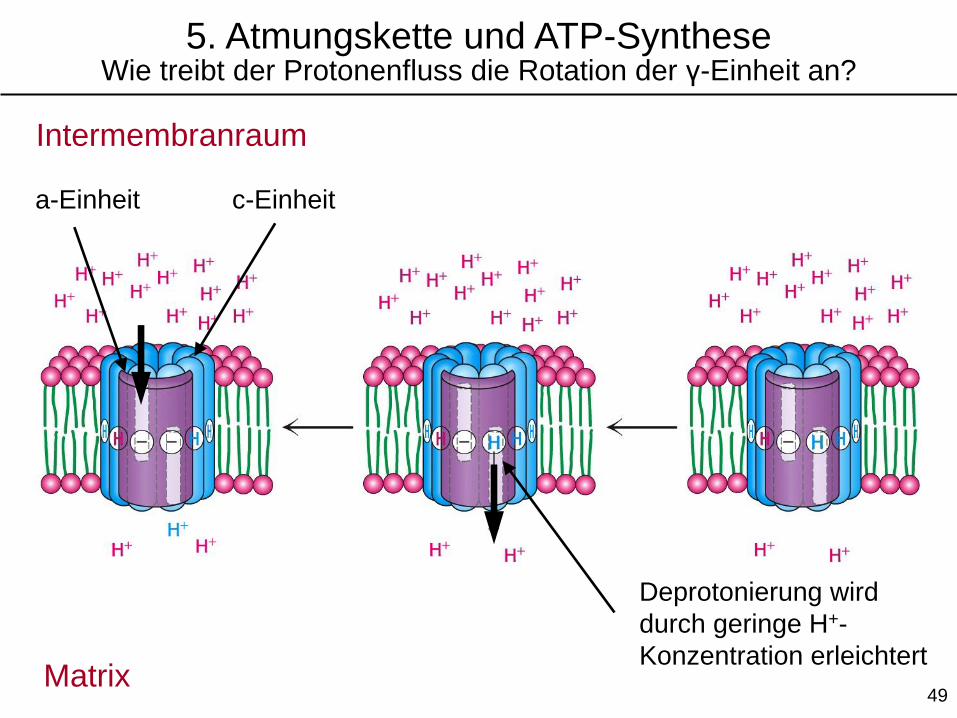
Matrix
Deprotonierung wird
durch geringe H+-
Konzentration erleichtert
c-Einheita-Einheit
5. Atmungskette und ATP-SyntheseWie treibt der Protonenfluss die Rotation der γ-Einheit an?
Intermembranraum
49
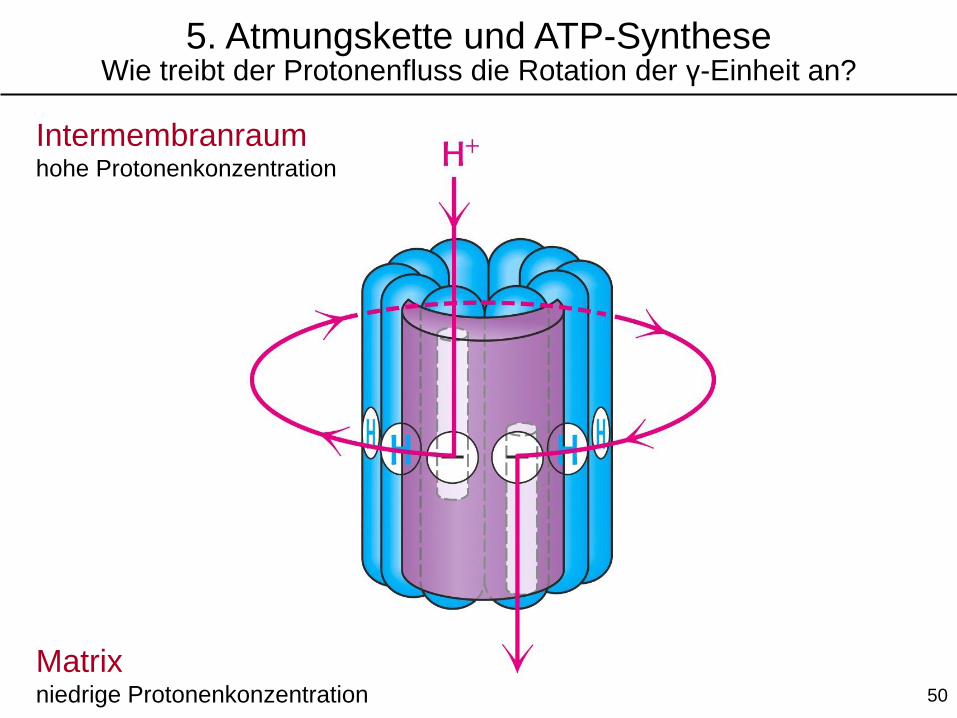
5. Atmungskette und ATP-SyntheseWie treibt der Protonenfluss die Rotation der γ-Einheit an?
Intermembranraumhohe Protonenkonzentration
Matrixniedrige Protonenkonzentration 50
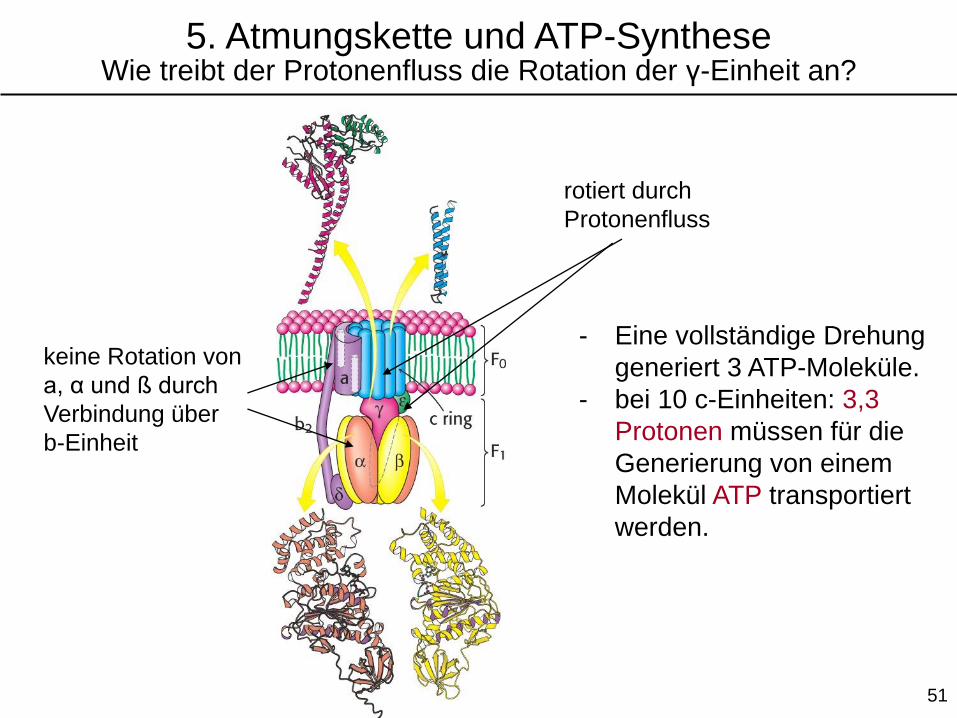
rotiert durch
Protonenfluss
keine Rotation von
a, α und ß durch
Verbindung über
b-Einheit
- Eine vollständige Drehung
generiert 3 ATP-Moleküle.
- bei 10 c-Einheiten: 3,3
Protonen müssen für die
Generierung von einem
Molekül ATP transportiert
werden.
5. Atmungskette und ATP-SyntheseWie treibt der Protonenfluss die Rotation der γ-Einheit an?
51
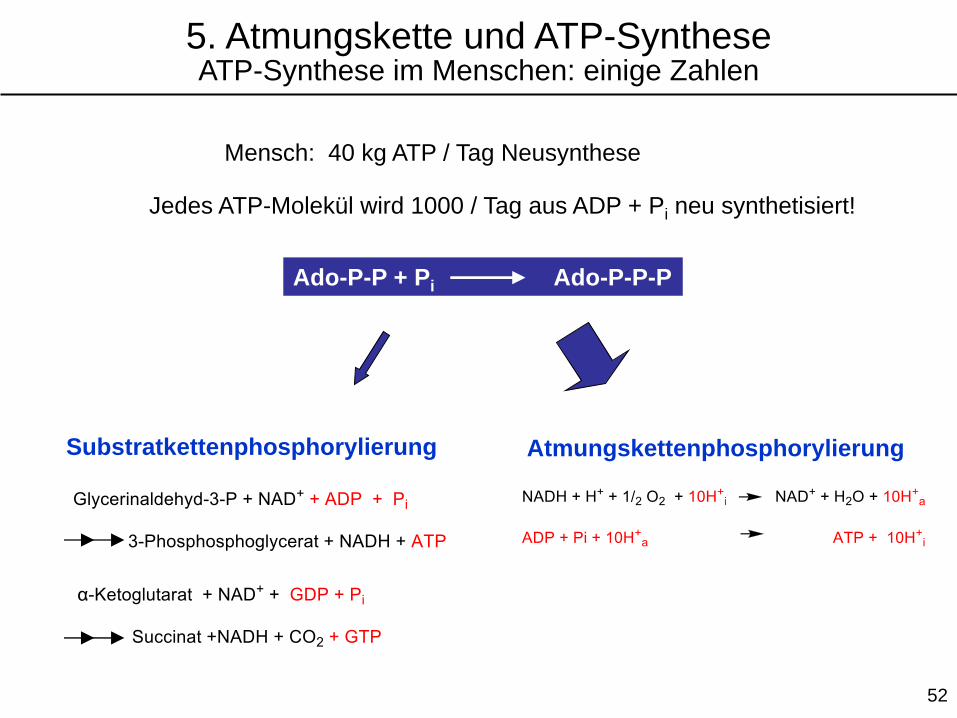
Mensch: 40 kg ATP / Tag Neusynthese
Jedes ATP-Molekül wird 1000 / Tag aus ADP + Pi neu synthetisiert!
Substratkettenphosphorylierung
Ado-P-P + Pi Ado-P-P-P
Atmungskettenphosphorylierung
5. Atmungskette und ATP-SyntheseATP-Synthese im Menschen: einige Zahlen
52
α
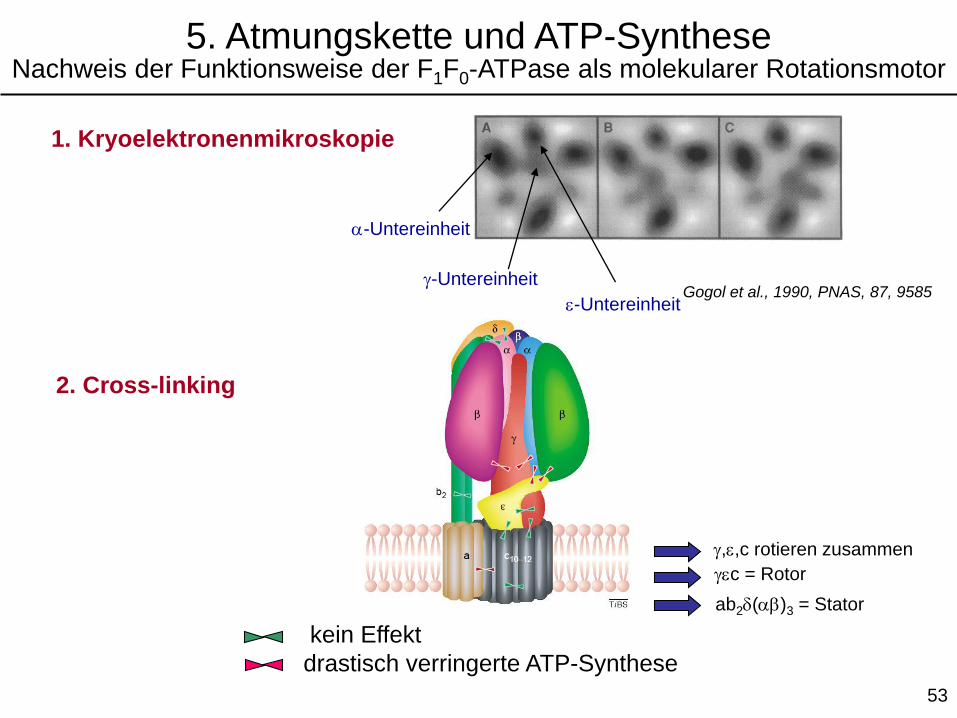
1. Kryoelektronenmikroskopie
a-Untereinheit
g-UntereinheitGogol et al., 1990, PNAS, 87, 9585
2. Cross-linking
kein Effekt
drastisch verringerte ATP-Synthese
g,e,c rotieren zusammen
gec = Rotor
ab2d(ab)3 = Stator
e-Untereinheit
5. Atmungskette und ATP-SyntheseNachweis der Funktionsweise der F1F0-ATPase als molekularer Rotationsmotor
53
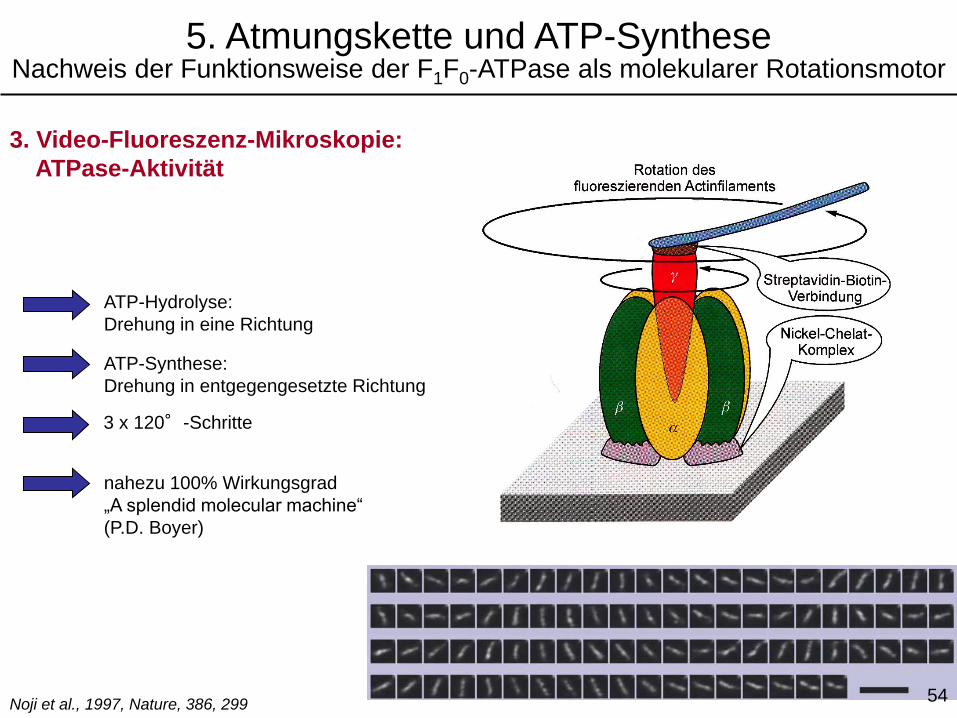
3. Video-Fluoreszenz-Mikroskopie:
ATPase-Aktivität
ATP-Hydrolyse:
Drehung in eine Richtung
ATP-Synthese:
Drehung in entgegengesetzte Richtung
nahezu 100% Wirkungsgrad
„A splendid molecular machine“
(P.D. Boyer)
Noji et al., 1997, Nature, 386, 299
3 x 120°-Schritte
5. Atmungskette und ATP-SyntheseNachweis der Funktionsweise der F1F0-ATPase als molekularer Rotationsmotor
54
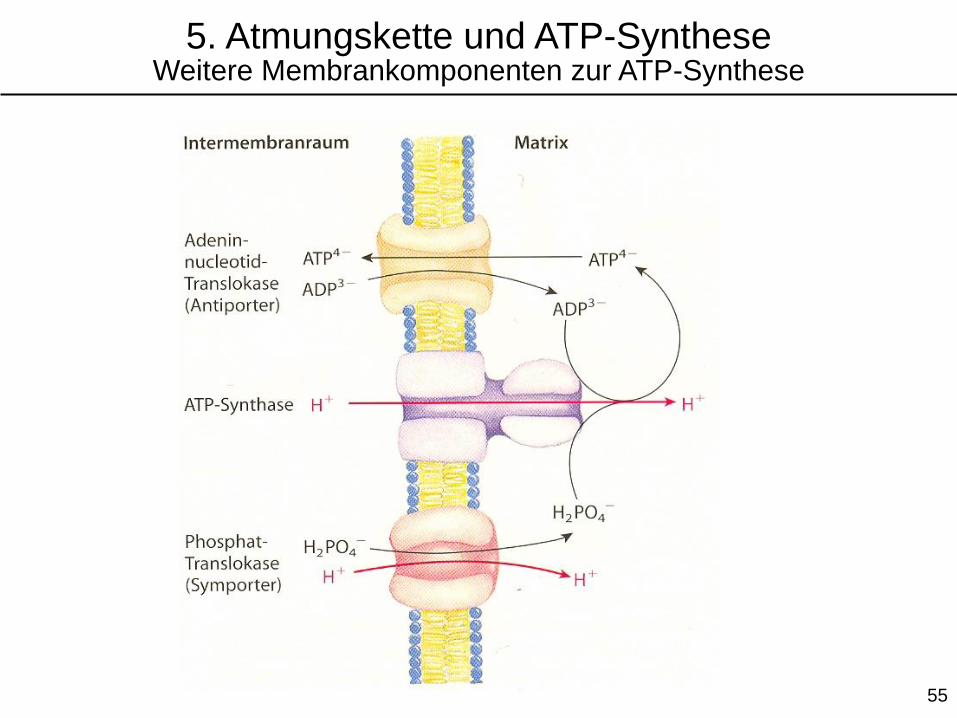
5. Atmungskette und ATP-SyntheseWeitere Membrankomponenten zur ATP-Synthese
55

Wieviel ATP entsteht ausgehend vom NADH+H+?
Geht man vom NADH+H+ aus, werden in der Atmungskette insgesamt 10 Protonen
aus dem Matrixraum in den Intermembranraum transportiert. Je 4 über Komplex I und
III sowie 2 über Komplex IV.
Nun wird ein Proton dafür benötigt, um ein Phosphat aus dem Zytoplasma ins
Mitochondrium zu transportieren und drei Protonen für die Phosphorylierung von
einem ADP.
Die Bildung eines ATPs kostet also 4 Protonen!
Aus der Oxidation von einem NADH+H+ entsteht in der Atmungskette 2,5 ATP
(weil: 10:4= 2,5).
5. Atmungskette und ATP-SyntheseATP-Ausbeute
56

Wie viel ATP entsteht ausgehend von FADH2?
Ausgehend vom FADH2 werden nicht 10 sondern nur 6 Protonen aus der Matrix
gepumpt.
Warum nur 6?
Weil der Einstieg in die Atmungskette erst bei Komplex I erfolgt, der ja selbst nicht
in der Lage ist, Protonen aus der Matrix zu pumpen.
Da die Bildung von ATP jedoch auch hier 4 Protonen kostet, entstehen deshalb
ausgehend vom FADH2 nur 1,5 ATP (weil: 6:4= 1,5).
5. Atmungskette und ATP-SyntheseATP-Ausbeute
57

Der Phosphat/Sauerstoff-Quotient (P/O-Quotient)
Mit dem P/O-Quotient kann man ausdrücken, wieviel Sauerstoff für die Bildung
einer bestimmten Menge an ATP benötigt wird.
Wegen der unterschiedlichen Energie der Elektronen von NADH+H+ und FADH2
ergibt sich für die beiden Reduktionsäquivalente auch ein unterschiedlicher P/O-
Quotient.
Beim NADH werden für einen Sauerstoff 2,5 Phosphate in ADP eingebaut, der
P/O-Quotient beträgt somit 2,5. Beim FADH2 sind es entsprechend nur 1,5.
5. Atmungskette und ATP-SyntheseATP-Ausbeute
58
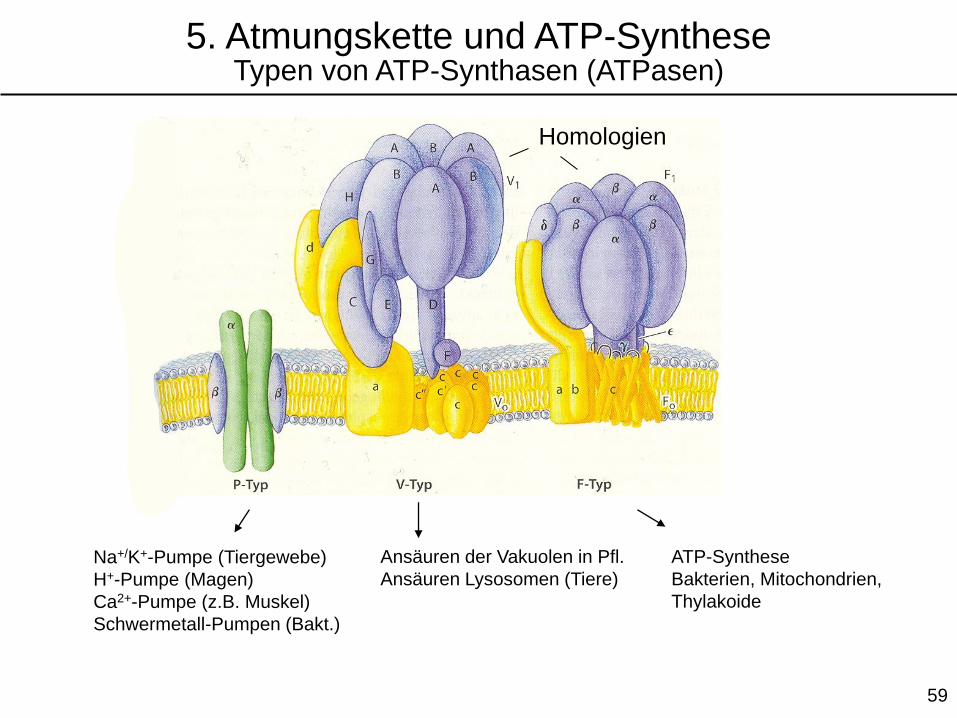
ATP-Synthese
Bakterien, Mitochondrien,
Thylakoide
Na+/K+-Pumpe (Tiergewebe)
H+-Pumpe (Magen)
Ca2+-Pumpe (z.B. Muskel)
Schwermetall-Pumpen (Bakt.)
Ansäuren der Vakuolen in Pfl.
Ansäuren Lysosomen (Tiere)
Homologien
5. Atmungskette und ATP-SyntheseTypen von ATP-Synthasen (ATPasen)
59
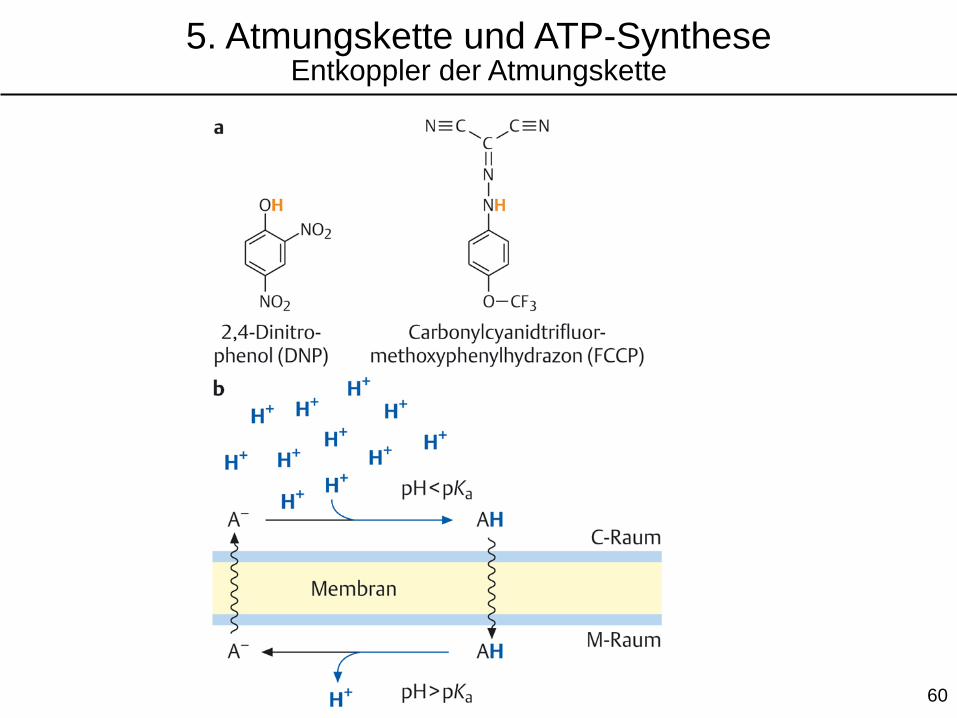
5. Atmungskette und ATP-SyntheseEntkoppler der Atmungskette
60
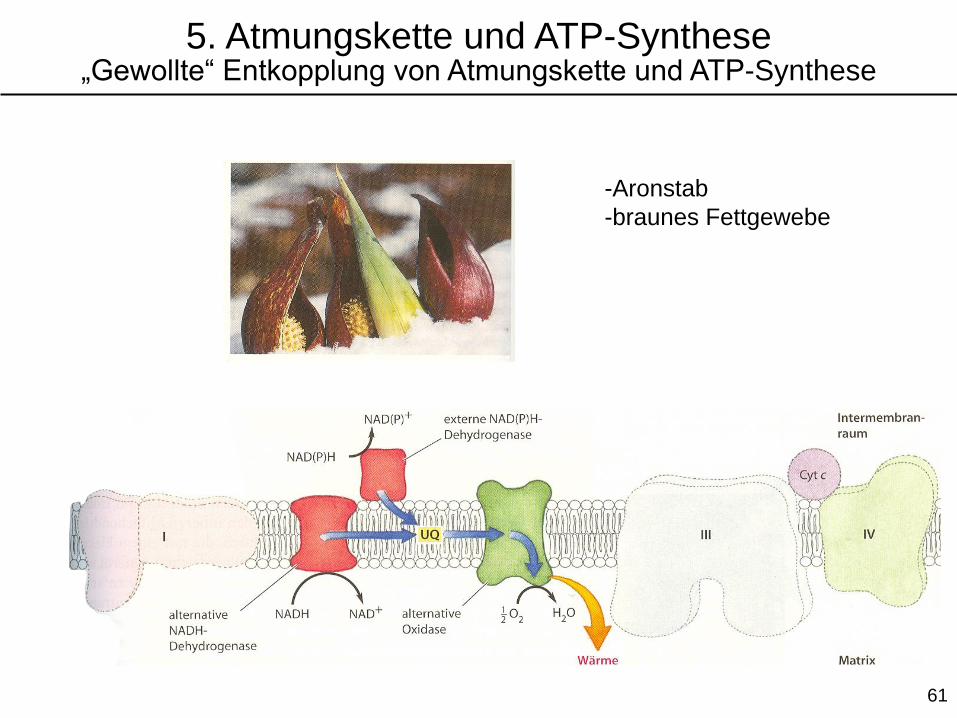
-Aronstab
-braunes Fettgewebe
5. Atmungskette und ATP-Synthese„Gewollte“ Entkopplung von Atmungskette und ATP-Synthese
61

5. Atmungskette und ATP-SyntheseZusammenfassung
62
In der ersten Phase der oxidativen Phosphorylierung werden die Elektronen der in den
katabolen Stoffwechselwegen gewonnenen Reduktionsäquivalente in der
Elektronentransportkette auf Sauerstoff übertragen.
Dies geschieht über mehrere große Membranproteinkomplexe, die Flavine, Eisen-Schwefel-
Zentren, Hämzentren und Kupferzentren als prosthetische Gruppen enthalten.
Beim Elektronenfluss wird in der ersten Phase ein Protonengradient über die innere
Mitochondrienmembran aufgebaut. In der zweiten Phase wird dieser Gradient von der ATP-
Synthase (Komplex V) zur ATP-Produktion genutzt.
Die Elektronen von NADH werden über die NADH-Dehydrogenase (Komplex I) in die
Atmungskette eingeschleust. Cytosolisch gebildetes NADH muss dazu indirekt über den Malat-
Aspartat-Shuttle in die mitochondriale Matrix eingeschleust werden.
Succinat-Dehydrogenase (Komplex II) des Citratzyklus und weitere FAD-abhängige
Dehydrogenasen wie α-Glycerophosphat-Dehydrogenase und ETF-Ubichinon-Oxidoreduktase
bilden ebenfalls Zuflüsse, indem Sie den mobilen Elektronenüberträger Ubichinon reduzieren.
Vom Ubichinon aus fließen die Elektronen über Cytochrom c-Reduktase (Komplex III),
Cytochrom c bzw. Cytochrom c-Oxidase (Komplex IV) zum Sauerstoff, der dabei zu Wasser
reduziert wird.

5. Atmungskette und ATP-SyntheseZusammenfassung
63
Die Elektronentranportkette pumpt zehn (ausgehend von Komplex I) bzw. sechs (ausgehend
von Komplex II) Protonen über die innere Mitochondrienmembran. Die dabei entstehende
protonenmotorische Kraft treibt die Rotation des membranständigen FO-Teils der ATP-
Synthase (Komplex V) an. Diese Drehbewegung wird auf den F1-Teil des Enzymkomplexes
übertragen und dort zur Synthese von ATP aus ADP und Pi genutzt.
Adeninnukleotid-Tranlokase und ein Phosphat-Carrier stellen unter Verbrauch von einem
gepumpten Proton sicher, dass ADP und Pi für die ATP-Synthese in die mitochondriale Matrix
hinein und neu synthetisiertes ATP aus den Mitochondrien hinaus transportiert werden.
Lipophile, schwache Säuren dienen als Entkoppler von Atmungskette und ATP-Synthese,
indem sie die innere mitochondriale Membran für Protonen durchlässig machen und so den
gebildeten H+-Gradienten gleich wieder zunichte machen. Dabei wird Energie in Form von
Wärme freigesetzt. Die „gewollte“ Entkopplung der Atmungskette von der ATP-Synthese wird
vom Aronstab und im brauen Fettgewebe zur Wärmeerzeugung genutzt.