Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL. CHEMISTRY Vol. 244, No. 9, Issue of May 10, pp. 22612268, 1969
P&ted in U.S.A.
Inhibition of Membrane-bound Adenosine Triphosphatase and of Cation Transport in Streptococcus faecalis by N,N’-Dicyclohexylcarbodiimide*
(Received for publication, December 23, 1968)
FRANKLIN M. HAROLD AND JAMES It. BAARDA
From the Division of Research, National Jewish Hospital, Denver, Colorado 80206, and the Department of Micro- biology, University of Colorado School of Medicine, Denver, Colorado 80220
CARL BARON AND ADOLPH ABRAM
From the Department of Biochemistry, University of Colorado School of Medicine, Denver, Colorado 80220
SUMMARY
N, N’-Dicyclohexylcarbodiimide (DCCD) was found to be a potent inhibitor of the membrane-bound ATPase of Sfrep- fococcus foe&is but did not inhibit the solubilized form of the enzyme. Inhibited membrane-bound ATPase was reactivated by releasing the enzyme from the membrane. Conversely, sensitivity to DCCD was restored by reconstitut- ing the ATPase-membrane complex from solubilized ATPase and depleted membranes. The results indicate that DCCD reacts covalently with a membrane component and inhibits the enzyme indirectly, perhaps by means of a transmitted effect on its conformation.
When added to intact cells, DCCD reduced the rate of glycolysis yet the ATP content of the cells was as high in the presence of the inhibitor as in its absence. DCCD strongly inhibited the degradation of ATP which ensued when glycol- ysis was stopped, either by removal of glucose or by addition of iodoacetate. The results suggest that DCCD inhibits ATPase in ho, just as it does in isolated membrane prepara- tions. The inhibition of over-all glycolysis may be secondary to the inhibition of ATP degradation.
DCCD also inhibited a number of energy-dependent trans- port processes, including the accumulation of Kf by exchange for H+ and Na+ and the uptake of phosphate and alanine. Inhibition was observed whether glucose or arginine served as energy source. The findings are consistent with the hypothesis that the membrane-bound ATPase of S. faecalis mediates energy transductions involved in membrane trans- port; inhibition of transport by DCCD would be a consequence of inhibition of the ATPase. Alternatively, DCCD may bind to sites on the membrane to inhibit indirectly both transport and ATP hydrolysis.
* This work was supported by Grants AI-03568 and GM- 05810 from the National Institutes of Health.
Adenosine triphosphatases are almost universally associated with biological membranes and have been implicated in both the generation and utilization of metabolic energy by membranous organelles. The ATPase of the inner mitochondrial membrane, for example, is thought to mediate the terminal step of oxidative phosphorylation (for reviews, see References 1 and 2). Plasma membranes of many animal cells contain ATPases which require Na+ and K+ for maximal activity, and these are almost certainly involved in the active transport of Na+ and K+ (3,4). ATPases are also commonly associated with membrane preparations from bact,eria (512), and it is presumed that they are concerned in energy transductions by microbial membranes.
Xtreptococcusfuecalisis of particular interest in this context as it lacks cytochromes and is thought to rely entirely (or at least, very largely) upon glycolysis for the generation of metabolic energy (13). A potent ATPase is associated with the plasma membranes of this organism (5) ; the enzyme is only weakly stim- ulated by Na+ and K+ (5, 14). Previous reports by Abrams (15), and his associates have described the release of the enzyme from the membranes, the partial characterization of the solu- bilized enzyme (16), and reattachment of the enzyme to empty binding sites on depleted membranes (17). The physiological function (or functions) of the ATPase has not been established, but indirect evidence implicated it in the transport of cations and of other metabolites (E-20).
Under the stimulus of the discovery (21-23) that the ATPase of the inner mitochondrial membrane is inhibited by N ,N’- dicyclohexylcarbodiimide, we have investigated the effect of this reagent on isolated plasma membranes and intact cells of S. fuecalis. We show here that DCCDr is a potent inhibitor of the membrane-bound ATPase, but has no effect on the solu- bilized enzyme. Reattachment of the enzyme to the membrane
1 The abbreviations used are: DCCD, N,N’-dicyclohexyl- carbodiimide: membrane-bound ATPase, two times washed plasma membrane preparation containing bound enzyme; re- leased ATPase, soluble ATPase obtained by release from mem- brane-bound ATPase.
2261
by guest on June 21, 2018http://w
ww
.jbc.org/D
ownloaded from
2262 Inhibition of ATPase and of Ion Transport in S. faecalis Vol. 244, No. 9
restores sensitivity to DCCD. Studies with intact cells were conducted in the expectation that physiological processes which depend upon the ATPase should therefore be inhibited by DCCD. DCCD does indeed inhibit the breakdown of ATP in the intact cell, and consequently reduces the rate of glycolysis. In addition, DCCD blocks energy-dependent net uptake of K+, phosphate, and alanine. The results are consistent with the hypothesis that the membrane-associated ATPase is involved in the act.ive transport of these metabolites.
METHODS AND MATERIALS
Preparation and Assay of Enzymes
Isolation of Native ATPase-Membrane Complex-The prepara- tion of membrane-bound ATPase from X. faecalis has been de- scribed (15,17). Briefly, cells were converted to protoplasts with lysozyme in high osmolar glycylglycine, followed by osmotic shock of the protoplasts in 1 mm MgSO1. The resulting mem- brane ghosts were then separated from the cytoplasmic fraction and washed twice with 1 mM MgS04. This twice-washed mem- brane preparation is referred to as (native) membrane-bound ATPase.
Preparation of Solubilized ATPase and Depleted Membranes- The procedure for solubilizing the ATPase has been described previously (15, 17). The membrane-bound ATPase prepara- tion described above was washed further, as follows: once with 2 M LiCl - 0.25 M Tris Cl, pH 7.5; once with 0.033 M Tris Cl, pH 7.5,l mMMgC12; and three times with 1 mM Tris Cl, pH 7.5. About 70 to 80% of the membrane-bound enzyme is released abruptly in a soluble form in these final three washes.
When the ATPase was to be used for reconstitution of the enzyme-membrane complex (17) it was concentrated by pre- cipitation with (NH&S04 between 20 and 80% saturation (16) and then redissolved in a small volume of Tris buffer.
The membrane residue remaining after release of ATPase as described above is referred to as depleted membranes. The de- pleted membranes still contain a small amount of ATPase.
Enzyme Assays-Routine assays of ATPase activity were per- formed by incubating the sample at 38” for 10 to 20 min in a volume of 1 .O ml containing 0.1 M Tris Cl (pH 7.5)-5 mM MgClz
ee _ mATPose
Li:: +I
c 0.04 - .- E
\ .- L -
sATPose
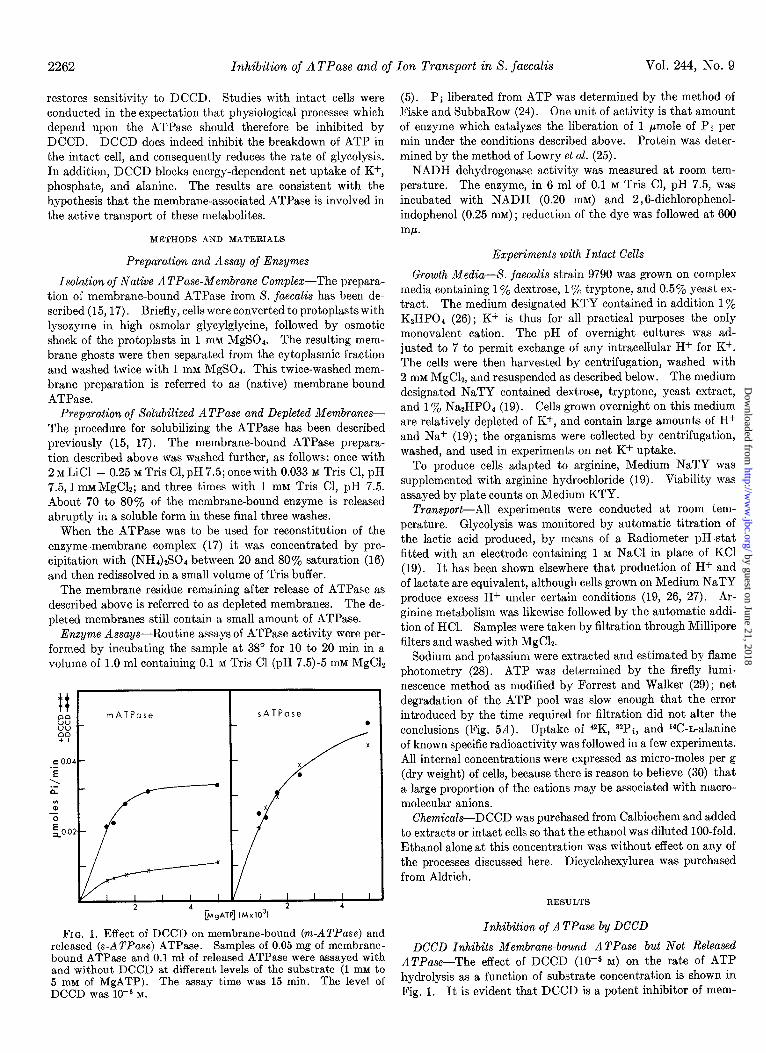
FIG. 1. Effect of DCCD on membrane-bound (m-ATPase) and released (s-ATPase) ATPase. Samples of 0.05 mg of membrane- bound ATPase and 0.1 ml of released ATPase were assayed with and without DCCD at different levels of the substrate (1 mM to 5 rnM of MgATP). The assay time was 15 min. The level of DCCD was 10eJ M.
(5). Pi liberated from ATP was determined by the method of Fiske and SubbaRow (24). One unit of activity is that amount of enzyme which catalyzes the liberation of 1 pmole of Pi per min under the conditions described above. Protein was deter- mined by the method of Lowry et al. (25).
NADH dehydrogenase activity was measured at room tem- perature. The enzyme, in 6 ml of 0.1 M Tris Cl, pH 7.5, was incubated with NADH (0.20 mM) and 2,6-dichlorophenol- indophenol (0.25 mM); reduction of the dye was followed at 600 w.
Experiments with Intact Cells
Growth Media---S. jaecalis strain 9790 was grown on complex media containing 1 To dextrose, 1 y0 tryptone, and 0.5% yeast ex- tract. The medium designated KTY contained in addition 1% KzHP04 (26) ; Kf is thus for all practical purposes the only monovalent cation. The pH of overnight cultures was ad- justed to 7 to permit exchange of any intracellular H+ for K+. The cells were then harvested by centrifugation, washed with 2 mM MgC12, and resuspended as described below. The medium designated NaTY contained dextrose, tryptone, yeast extract, and 1 To Na2HP04 (19). Cells grown overnight on this medium are relatively depleted of K+, and contain large amounts of H+ and Na+ (19); the organisms were collected by centrifugation, washed, and used in experiments on net K+ uptake.
To produce cells adapted to arginine, Medium NaTY was supplemented with arginine hydrochloride (19). Viability was assayed by plate counts on Medium KTY.
Transport-All experiments were conducted at room tem- perature. Glycolysis was monitored by automatic titration of the lactic acid produced, by means of a Radiometer pH-stat fitted with an electrode containing 1 M NaCl in place of KC1 (19). It has been shown elsewhere that production of H+ and of lactate are equivalent, although cells grown on Medium NaTY produce excess H+ under certain conditions (19, 26, 27). Ar- ginine metabolism was likewise followed by the automatic addi- tion of HCl. Samples were taken by filtration through Millipore filters and washed with MgCls.
Sodium and potassium were extracted and estimated by flame photometry (28). ATP was determined by the firefly lumi- nescence method as modified by Forrest and Walker (29) ; net degradation of the ATP pool was slow enough that the error introduced by the time required for filtration did not alter the conclusions (Fig. 58). Uptake of 42K, 32Pi, and i4C-n-alanine of known specific radioactivity was followed in a few experiments. All internal concentrations were expressed as micro-moles per g (dry weight) of cells, because there is reason to believe (30) that a large proportion of the cations may be associated with macro- molecular anions.
ChemicaZsDCCD was purchased from Calbiochem and added to extracts or intact cells so that the ethanol was diluted IOO-fold. Ethanol alone at this concentration was without effect on any of the processes discussed here. Dicyclohexylurea was purchased from Aldrich.
RESULTS
Inhibition of ATPase by DCCD
DCCD Inhibits Membrane-bound ATPase but Not Released ATPuse-The effect of DCCD (10m5 M) on the rate of ATP hydrolysis as a function of substrate concentration is shown in Fig. 1. It is evident that DCCD is a potent inhibitor of mem-
by guest on June 21, 2018http://w
ww
.jbc.org/D
ownloaded from
Issue of May 10, 1969 F. M. Harold, J. R. Baarda, C. Baron, and A. Abrams 2263
brane-bound ATPase but has no detectable effect on the re- leased ATPase. The released ATPase was not affected by DCCD even when the enzyme was treated for 1 hour with a lo- fold higher concentration of DCCD. Dicyclohexylurea, a possible product of reactions involving DCCD, inhibited neither membrane-bound ATPase nor released ATPase. It would thus appear that attachment of the enzyme to a membrane component is required for DCCD inhibition.
The soluble cytoplasmic fraction prepared as described under “Methods and Materials” contains some ATPase activity (5), and this, like the released ATPase obtained from the membranes, was also insensitive to DCCD. It is likely that the released ATPase derived from the protoplast membrane and the ATPase found in the cytoplasmic fraction are the same enzyme, as they have identical electrophoretic mobilities in polyacrylamide gel
(16).
101
7
6 c m z5 - -0 0
2
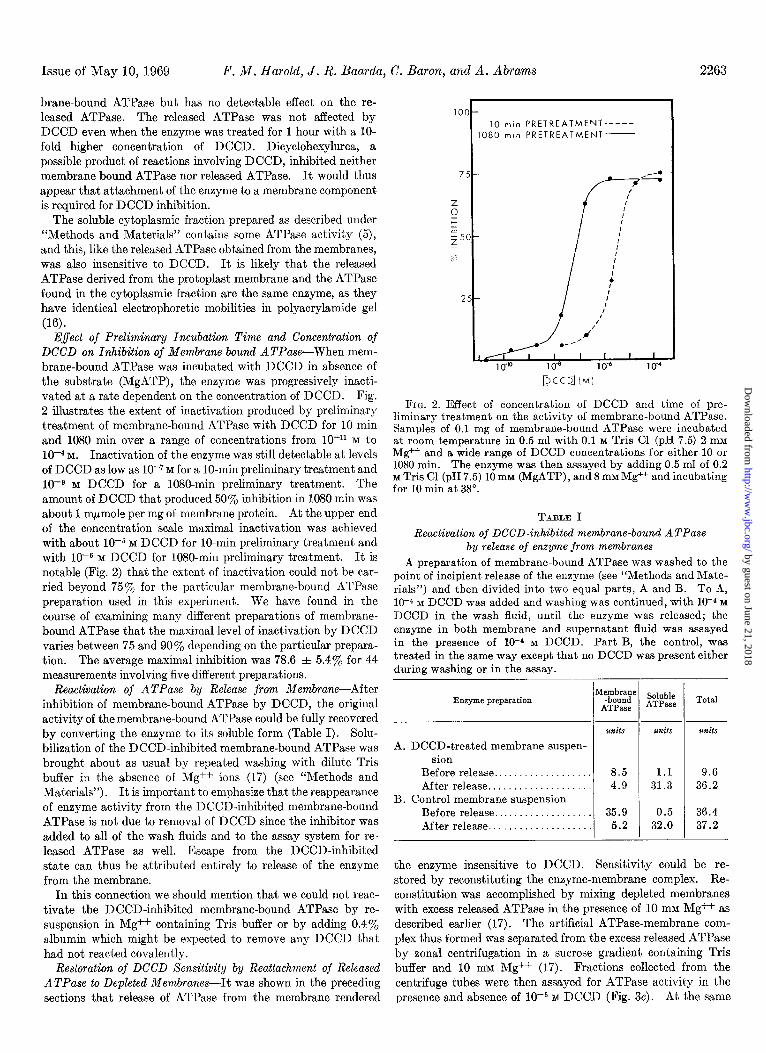
Effect of Preliminary Incubation Time and Concentration of DCCD on Inhibition of Membrane-bound ATPase-When mem- brane-bound ATPase was incubated with DCCD in absence of the substrate (MgATP), the enzyme was progressively inacti- vated at a rate dependent on the concentration of DCCD. Fig. 2 illustrates the extent of inactivation produced by preliminary treatment of membrane-bound ATPase with DCCD for 10 min and 1080 min over a range of concentrations from lo-” M to
10m4 M. Inactivation of the enzyme was still detectable at levels of DCCD as low as lo-‘M for a IO-min preliminary treatment and 10eg M DCCD for a lOSO-min preliminary treatment. The amount of DCCD that produced 50% inhibition in 1080 min was about 1 m/lmole per mg of membrane protein. At the upper end of the concentration scale maximal inactivation was achieved with about 10e5 M DCCD for lo-min preliminary treatment and with 10e6 M DCCD for 1080-min preliminary treatment. It is notable (Fig. 2) that the extent of inactivation could not be car- ried beyond 75% for the particular membrane-bound ATPase preparation used in this experiment. We have found in the course of examining many different preparations of membrane- bound ATPase that the maximal level of inactivation by DCCD varies between 75 and 90% depending on the particular prepara- tion. The average maximal inhibition was 78.6 f 5.4% for 44 measurements involving five different preparations.
FIG. 2. Effect of concentration of DCCD and time of pre- liminary treatment on the activity of membrane-bound ATPase. Samples of 0.1 mg of membrane-bound ATPase were incubated at room temperature in 0.5 ml with 0.1 M Tris Cl (pH 7.5) 2 mM Mg++ and a wide range of DCCD concentrations for either 10 or 1080 min. The enzyme was then assayed by adding 0.5 ml of 0.2 M Tris Cl (pH 7.5) 10 mM (MgATP), and 8 mM Mg++ and incubating for 10 min at 38”.
Reactivation of DCCD-inhibited membrane-bound ATPase by release of enzyme from membranes
A preparation of membrane-bound ATPase was washed to the point of incipient release of the enzyme (see “Methods and Mate- rials”) and then divided into two equal parts, A and B. To A, 10-J M DCCD was added and washing was continued, with 10-4 M
DCCD in the wash fluid, until the enzyme was released; the enzyme in both membrane and supernatant fluid was assayed in the presence of 10-d M DCCD. Part B, the control, was treated in the same way except that no DCCD was present either during washing or in the assay.
3-
1
5-
O-
5-
A
Reactivation of ATPase by Release from Membrane-After inhibition of membrane-bound ATPase by DCCD, the original activity of the membrane-bound ATPase could be fully recovered by converting the enzyme to its soluble form (Table I). Solu- bilieation of the DCCD-inhibited membrane-bound ATPase was brought about as usual by repeated washing with dilute Tris buffer in the absence of Mg++ ions (17) (see “Methods and Materials”). It is important to emphasize that the reappearance of enzyme activity from the DCCD-inhibited membrane-bound ATPase is not due to removal of DCCD since the inhibitor was added to all of the wash fluids and to the assay system for re- leased ATPase as well. Escape from the DCCD-inhibited state can thus be attributed entirely to release of the enzyme from the membrane.
In this connection we should mention that we could not reac- tivate the DCCD-inhibited membrane-bound ATPase by re- suspension in Mg++ containing Tris buffer or by adding 0.4% albumin which might be expected to remove any DCCD that had not reacted covalently.
Restoration of DCCD Sensitivity by Reattachment of Released ATPase to Depleted Membranes-It was shown in the preceding sections that release of ATPase from the membrane rendered
10 min PRETREATMENT-----
080 min PRETREATMENT-
DCCD] (Mi
TABLE I
Enzyme preparation
A. DCCD-treated membrane suspen- sion
Before release.................. After release...................
B. Control membrane suspension Before release.. . . . After release.. . .
units
8.5 1.1 9.6 4.9 31.3 36.2
35.9 0.5 36.4 5.2 32.0 37.2
Soluble ATPase
units
Total
1mits
the enzyme insensitive to DCCD. Sensitivity could be re- stored by reconstituting the enzyme-membrane complex. Re- constitution was accomplished by mixing depleted membranes with excess released ATPase in the presence of 10 mM Mg++ as described earlier (17). The artificial ATPase-membrane com- plex thus formed was separated from the excess released ATPase by zonal centrifugation in a sucrose gradient containing Tris buffer and 10 mM Mg++ (17). Fractions collected from the centrifuge tubes were then assayed for ATPase activity in the presence and absence of 10e5 M DCCD (Fig. 3~). At the same
by guest on June 21, 2018http://w
ww
.jbc.org/D
ownloaded from
2264 Inhibition of ATPase and of Ion Transport in S. faecalis Vol. 244, No. 9
ATPa;dpEyE;RANE
RECONSTITUTED c
b
DEPLETED
l-
a NATIVE
sATPose
1 -
15 TOP FRACTION NUMBER
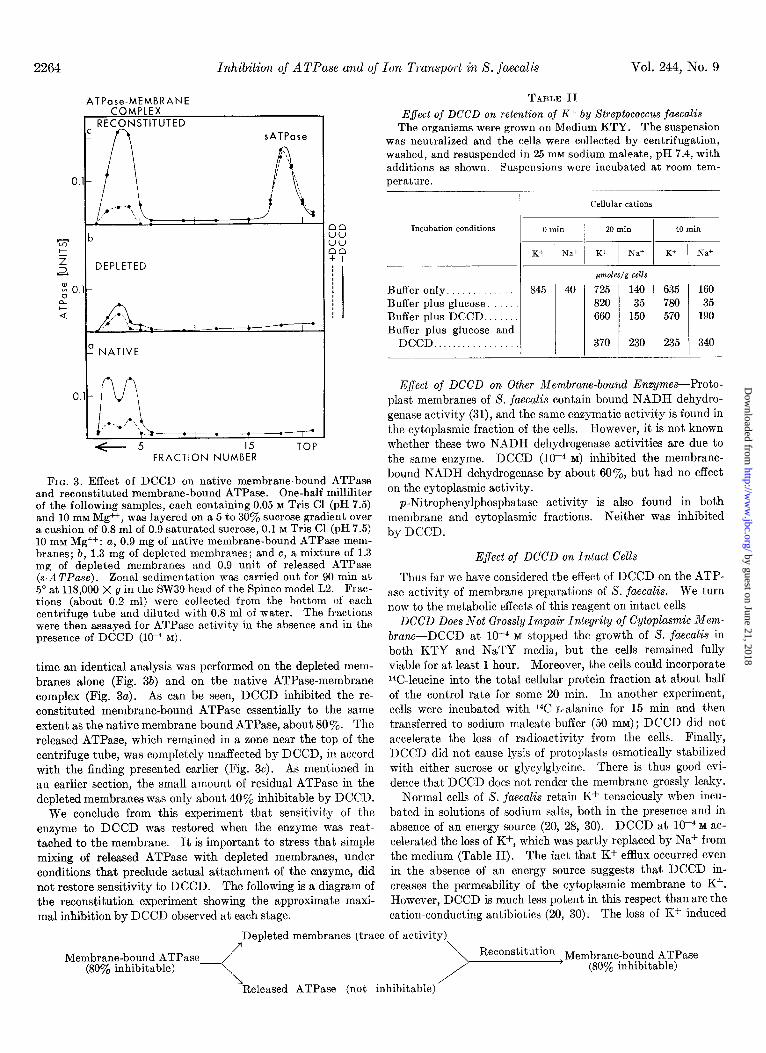
FIG. 3. Effect of DCCD on native membrane-bound ATPase and reconstituted membrane-bound ATPase. One-half milliliter of the following samples, each containing 0.05 M Tris Cl (pH 7.5) and 10 mM Mg++, was layered on a 5 to 30’y0 sucrose gradient over a cushion of 0.8 ml of 0.9 saturated sucrose, 0.1 M Tris Cl (pH 7.5) lo mM Mg++: a, 0.9 mg of native membrane-bound ATPase mem- branes; b, 1.3 mg of depleted membranes; and c, a mixture of 1.3 mg of depleted membranes and 0.9 unit of released ATPase (s-ATPnse). Zonal sedimentation was carried out for 90 min at 5” at 118,000 X g in the SW39 head of the Spinco model L2. Frac- tions (about 0.2 ml) were collected from the bottom of each centrifuge tube and diluted with 0.8 ml of water. The fractions were then assayed for ATPase activity in the absence and in the presence of DCCD (lo-* M).
time an identical analysis was performed on the depleted mem- branes alone (Fig. 3b) and on the native ATPase-membrane complex (Fig. 3~). As can be seen, DCCD inhibited the re- constituted membrane-bound ATPase essentially to the same extent as the native membrane-bound ATPase, about 80%. The released ATPase, which remained in a zone near the top of the centrifuge tube, was completely unaffected by DCCD, in accord with the finding presented earlier (Fig. 3~). As mentioned in an earlier section, the small amount of residual ATPase in the depleted membranes was only about 40% inhibitable by DCCD.
We conclude from this experiment that sensitivity of the enzyme to DCCD was restored when the enzyme was reat- tached to the membrane. It is important to stress that simple mixing of released ATPase with depleted membranes, under conditions that preclude actual attachment of the enzyme, did not restore sensitivity to DCCD. The following is a diagram of the reconstitution experiment showing the approximate maxi- mal inhibition by DCCD observed at each stage.
TABLE II Effect of DCCD on retention of K+ by Streptococcus faecalis
The organisms were grown on Medium KTY. The suspension was neutralized and the cells were collected by centrifugation, washed, and resuspended in 25 mM sodium maleate, pH 7.4, with additions as shown. Suspensions were incubated at room tem- perature.
Cellular cations
Incubation conditions 0 min 20 min 40 min
K+ Ill:1
Naf K+ Naf Kf Na+
~vLoles/g cells
Buffer only.. 845 40 725 140 635 160 Buffer plus glucose. 820 35 780 35 Buffer plus DCCD., 660 150 570 190
Buffer plus glucose and DCCD.. 370 230 235 340
Effect of DCCD on Other Membrane-bound Enzymes-Proto- plast membranes of S. faecalis contain bound NADH dehydro- genase activity (31), and the same enzymatic activity is found in the cytoplasmic fraction of the cells. However, it is not known whether these two NADH dehydrogenase activities are due to the same enzyme. DCCD (lop4 M) inhibited the membrane- bound NADH dehydrogenase by about SO%, but had no effect on the cytoplasmic activity.
p-Nitrophenylphosphatase activity is also found in both membrane and cytoplasmic fractions. Neither was inhibit,ed by DCCD.
EJect of DCCD on Intact Cells
Thus far we have considered the effect of DCCD on the ATP- ase activity of membrane preparations of S. faecalis. We turn now to the metabolic effects of this reagent on intact cells
DCCD Does Not Grossly Impair Integrity of Cytoplasmic Mem-
brune-DCCD at 10v4 M stopped the growth of 8. fuecalis in
both KTY and NaTY media, but the cells remained fully viable for at least 1 hour. Moreover, the cells could incorporate 14C-leucine into the total cellular protein fraction at about half of the control rate for some 20 min. In another experiment, cells were incubated with 14C-L-alanine for 15 min and then transferred to sodium maleate buffer (50 mM) ; DCCD did not accelerate the loss of radioactivity from the cells. Finally, DCCD did not cause lysis of protoplasts osmotically stabilized with either sucrose or glycylglycine. There is thus good evi- dence that DCCD does not render the membrane grossly leaky.
Normal cells of S. faecalis retain K+ tenaciously when incu- bated in solutions of sodium salts, both in the presence and in absence of an energy source (20, 28, 30). DCCD at lop4 M ac-
celerated the loss of K+, which was partly replaced by Na+ from the medium (Table II). The fact that K+ efflux occurred even in the absence of an energy source suggests that DCCD in- creases the permeability of the cytoplasmic membrane to K+. However, DCCD is much less potent in this respect than are the cation-conducting antibiot,ics (20, 30). The loss of K+ induced
Depleted membranes (trace of activity) 7 \
Membrane-bound ATPase , Reconstifution Membrane-bound ATPase (8OoJ, inhibitable)
L / (80’% inhibitable)
Released ATPase (not inhibitable)
by guest on June 21, 2018http://w
ww
.jbc.org/D
ownloaded from
Issue of May 10, 1969 F. M. Harold, J. R. Baa&a, C. Baron, and A. Abrams
by DCCD was slow, and was seen only in the presence of rela- tively high concentrations of Na+ (50 mM, but not 10 mM). The stimulation of K+ efflux in the presence of both DCCD and glu- cose is presumably related to the inhibition of net K+ accu- mulation by DCCD, and is discussed below.
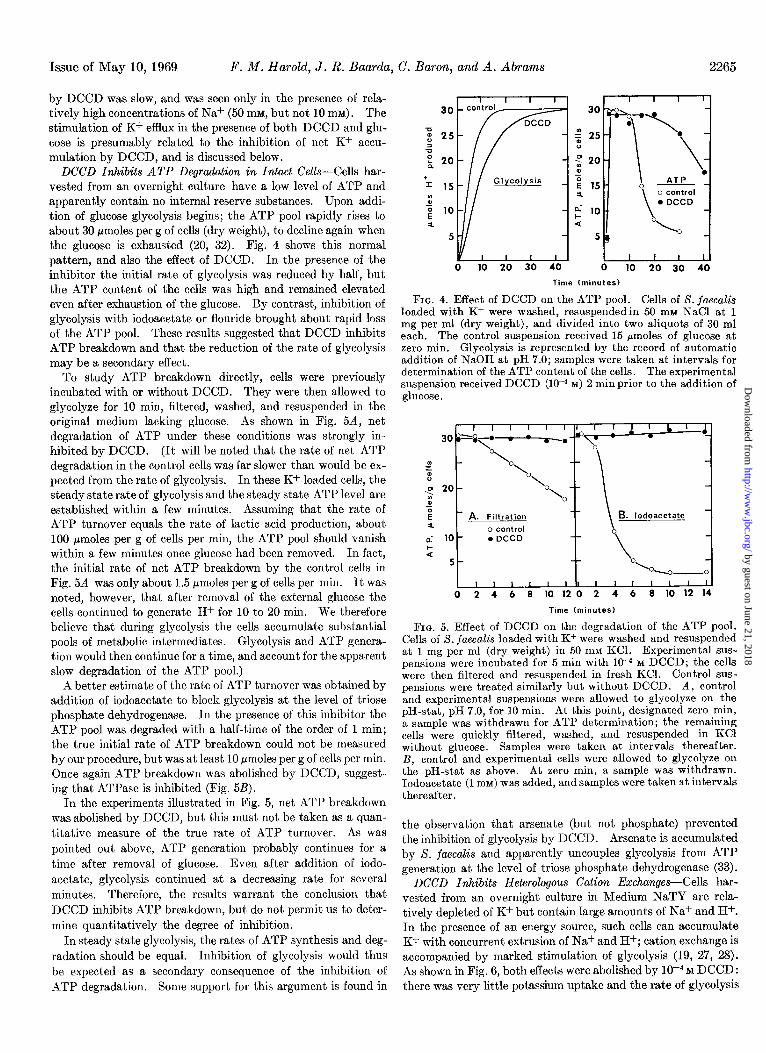
DCCD Inhibits ATP Degradation in Intact Cells-Cells har- vested from an overnight culture have a low level of ATP and apparently contain no internal reserve substances. Upon addi- tion of glucose glycolysis begins; the ATP pool rapidly rises to about 30 pmoles per g of cells (dry weight), to decline again when the glucose is exhausted (20, 32). Fig. 4 shows this normal pattern, and also the effect of DCCD. In the presence of the inhibitor the initial rate of glycolysis was reduced by half, but the ATP content of the cells was high and remained elevated even after exhaustion of the glucose. By contrast, inhibition of glycolysis with iodoacetate or flouride brought about rapid loss of the ATP pool. These results suggested that DCCD inhibits ATP breakdown and that the reduction of the rate of glycolysis may be a secondary effect.
Glycolysis _
p 20 0
= E 15 4
$ 10 a
5
Time (minutes)
To study ATP breakdown directly, cells were previously incubated with or without DCCD. They were then allowed to glycolyze for 10 min, filtered, washed, and resuspended in the original medium lacking glucose. As shown in Fig. 5A, net degradation of ATP under these conditions was strongly in- hibited by DCCD. (It will be noted that the rate of net ATP degradation in the control cells was far slower than would be ex- pected from the rate of glycolysis. In these K+-loaded cells, the steady state rate of glycolysis and the steady state ATP level are established within a few minutes. Assuming that the rate of ATP turnover equals the rate of lactic acid production, about 100 pmoles per g of cells per min, the ATP pool should vanish within a few minutes once glucose had been removed. In fact, the initial rate of net ATP breakdown by the control cells in Fig. 5A was only about 1.5 pmoles per g of cells per min. It was noted, however, that after removal of the external glucose the cells continued to generate H+ for 10 to 20 min. We therefore believe that during glycolysis the cells accumulate substantial pools of metabolic intermediates. Glycolysis and ATP genera- tion would then continue for a time, and account for the apparent slow degradation of the ATP pool.)
FIG. 4. Effect of DCCD on the ATP pool. Cells of S. juecaZis loaded with K+ were washed, resuspendedin 50 nnu NaCl at 1 mg per ml (dry weight), and divided into two aliquots of 30 ml each. The control suspension received 15 pmoles of glucose at zero min. Glycolysis is represented by the record of automatic addition of NaOH at pH 7.0; samples were taken at intervals for determination of the ATP content of the cells. The experimental suspension received DCCD (low4 M) 2 min prior to the addition of glucose.
I I I I I I 303~a~*--.
O\
I I I I I I Ill I I I I I I II 0 2 4 6 8 10 12 0 2 4 6 8 10 12 14
Time (minutes)
A better estimate of the rate of ATP turnover was obtained by addition of iodoacetate to block glycolysis at the level of triose phosphate dehydrogenase. In the presence of this inhibitor the ATP pool was degraded with a half-time of the order of 1 min; the true initial rate of ATP breakdown could not be measured by our procedure, but was at least 10 pmoles per g of cells per min. Once again ATP breakdown was abolished by DCCD, suggest- ing that ATPase is inhibited (Fig. 5B).
In the experiments illustrated in Fig. 5, net ATP breakdown was abolished by DCCD, but this must not be taken as a quan- titative measure of the true rate of ATP turnover. As was pointed out above, ATP generation probably continues for a time after removal of glucose. Even after addition of iodo- acetate, glycolysis continued at a decreasing rate for several minutes. Therefore, the results warrant the conclusion that DCCD inhibits ATP breakdown, but do not permit us to deter- mine quantitatively the degree of inhibition.
FIG. 5. Effect of DCCD on the degradation of the ATP pool. Cells of S. jaecalis loaded with K+ were washed and resuspended at 1 mg per ml (dry weight) in 50 mM KCl. Experimental sus- pensions were incubated for 5 min with 10-4 M DCCD; the cells were then filtered and resuspended in fresh KCl. Control sus- pensions were treated similarly but without DCCD. A, control and experimental suspensions were allowed to glycolyze on the pH-stat, pH 7.0, for 10 min. At this point, designated zero min, a sample was withdrawn for ATP determination; the remaining cells were ouickly filtered, washed, and resuspended in KC1 without glucose. - Samples were taken at intervals thereafter. B, control and experimental cells were allowed to glycolyze on the pH-stat as above. At zero min, a sample was withdrawn. Iodoacetate (1 mM) was added, and samples were taken at intervals thereafter.
the observation that arsenate (but not phosphate) prevented the inhibition of glycolysis by DCCD. Arsenate is accumulated by S. fuecalis and apparently uncouples glycolysis from ATP generation at the level of triose phosphate dehydrogenase (33).
In steady state glycolysis, the rates of ATP synthesis and deg- radation should be equal. Inhibition of glycolysis would thus be expected as a secondary consequence of the inhibition of
DCCD Inhibits Heterologous Cation Exchanges-Cells har- vested from an overnight culture in Medium NaTY are rela- tively depleted of K+ but contain large amounts of Na+ and H+. In the presence of an energy source, such cells can accumulate K+ with concurrent extrusion of Na+ and H+; cation exchange is accompanied by marked stimulation of glycolysis (19, 27, 28). As shown in Fig. 6, both effects were abolished by 1r4 M DCCD:
ATP degradation. Some support for this argument is found in there was very little potassium uptake and the rate of glycolysis
2265
by guest on June 21, 2018http://w
ww
.jbc.org/D
ownloaded from
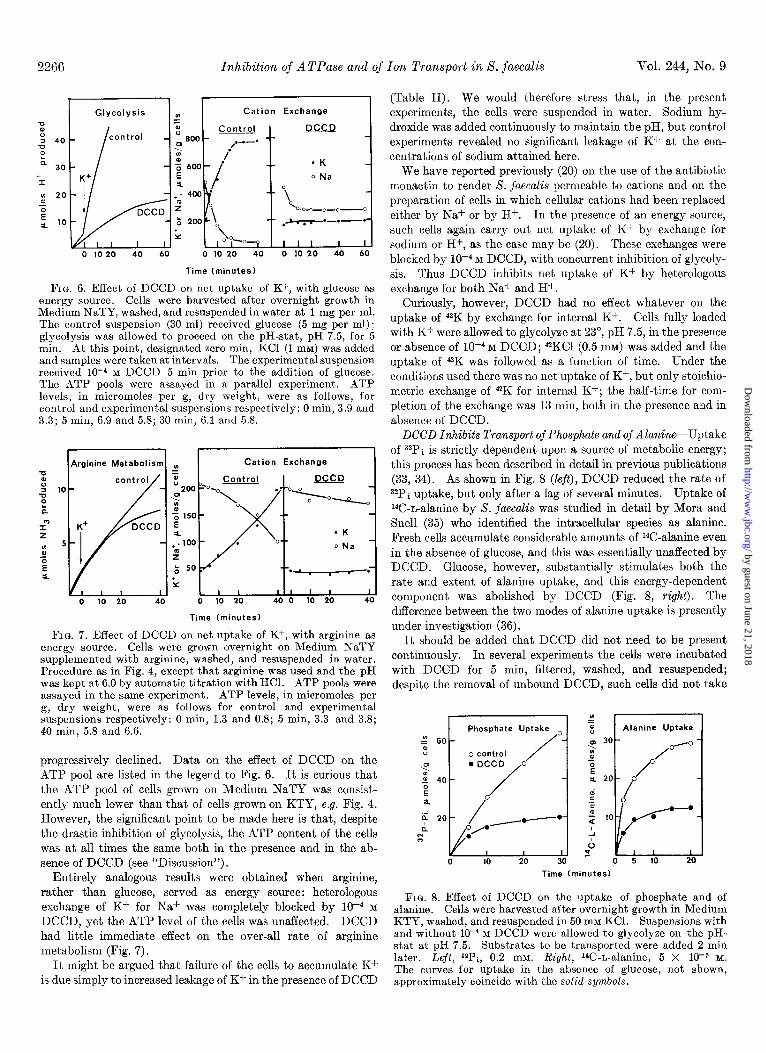
2266 Inhibition of A TPase and of Ton Transport in S. faecalis Vol. 244, No. 9
Cation Exchange
(Table II). We would therefore stress that, in the present experiments, the cells were suspended in water. Sodium hy- droxide was added continuously to maintain the pH, but control experiments revealed no significant leakage of K+ at the con- centrations of sodium attained here.
0 10 20 40 60 0 10 20 40 0 10 20 40 60
Time (minutes)
FIG. 6. Effect of DCCD on net uptake of K+, with glucose as energy source. Cells were harvested after overnight growth in Medium NaTY, washed, and resuspended in water at 1 mg per ml. The control suspension (30 ml) received glucose (5 mg per ml); glycolysis was allowed to proceed on the pH-stat, pH 7.5, for 5 min. At this point, designated zero min, KC1 (1 mM) was added and samples were taken at intervals. The experimental suspension received lo-4 M DCCD 5 min prior to the addition of glucose. The ATP pools were assayed in a parallel experiment. ATP levels, in micromoles per g, dry weight, were as follows, for control and experimental suspensions respectively: 0 min, 3.9 and 3.3; 5 min, 6.9 and 5.8; 30 min, 6.1 and 5.8.
We have reported previously (20) on the use of the antibiotic monactin to render 8. fuecalis permeable to cations and on the preparation of cells in which cellular cations had been replaced either by Na+ or by H+. In the presence of an energy source, such cells again carry out net uptake of K+ by exchange for sodium or H+, as the case may be (20). These exchanges were blocked by 1O-4 M DCCD, with concurrent inhibition of glycoly- sis. Thus DCCD inhibits net uptake of K+ by heterologous exchange for both Na+ and H+.
Curiously, however, DCCD had no effect whatever on the uptake of 42K by exchange for internal K+. Cells fully loaded with Kf were allowed to glycolyze at 23”, pH 7.5, in the presence or absence of lo-* M DCCD; 42KC1 (0.5 mM) was added and the uptake of 42K was followed as a function of time. Under the conditions used there was no net uptake of K+, but only stoichio- metric exchange of 42K for internal K+; the half-time for com- pletion of the exchange was 13 min, both in the presence and in absence of DCCD.
Cation Exchange -0 E
2 10
‘p
I” 2
: 5
s E a.
0 10 20 40 0 10 20 40 0 10 20 40
Time (minutes)
FIG. 7. Effect of DCCD on net uptake of K+, with arginine as energy source. Cells were grown overnight on Medium NaTY supplemented with arginine, washed, and resuspended in water. Procedure as in Fig. 4, except that arginine was used and the pH was kept at 6.0 by automatic titration with HCl. ATP pools were assayed in the same experiment. ATP levels, in micromoles per g, dry weight, were as follows for control and experimental suspensions respectively: 0 min, 1.3 and 0.8; 5 min, 3.3 and 3.8; 40 min, 5.8 and 6.6.
DCCD Inhibits Transport of Phosphate and of Alanine-Uptake of 32Pi is strictly dependent upon a source of metabolic energy; this process has been described in detail in previous publications (33, 34). As shown in Fig. 8 (left), DCCD reduced the rate of 32Pi uptake, but only after a lag of several minutes. Uptake of 14C-L-alanine by S. faecalis was studied in detail by Mora and Snell (35) who identified the intracellular species as alanine. Fresh cells accumulate considerable amounts of 14C-alanine even in the absence of glucose, and this was essentially unaffected by DCCD. Glucose, however, substantially stimulates both the rate and extent of alanine uptake, and this energy-dependent component was abolished by DCCD (Fig. 8, right). The difference between the two modes of alanine uptake is presently under investigation (36).
It should be added that DCCD did not need to be present continuously. In several experiments the cells were incubated wit,h DCCD for 5 min, filtered, washed, and resuspended; despite the removal of unbound DCCD, such cells did not take
60 60
progressively declined. Data on the effect of DCCD on the ATP pool are listed in the legend to Fig. 6. It is curious that the ATP pool of cells grown on Medium NaTY was consist- ently much lower than that of cells grown on KTY, e.g. Fig. 4. However, the significant point to be made here is that, despite the drastic inhibition of glycolysis, the ATP content of the cells was at all times the same both in the presence and in the ab- sence of DCCD (see “Discussion”).
Entirely analogous results were obtained when arginine, rather than glucose, served as energy source: heterologous exchange of K+ for Na+ was completely blocked by lo-* M
DCCD, yet the ATP level of the cells was unaffected. DCCD had little immediate effect on the over-all rate of arginine metabolism (Fig. 7).
Time (
0 5 10 20
Iminutes)
It might be argued that failure of the cells to accumulate K+ is due simply to increased leakage of K+ in the presence of DCCD
FIG. 8. Effect of DCCD on the uptake of phosphate and of alanine. Cells were harvested after overnight growth in Medium KTY, washed, and resuspended in 50 mM KCl. Suspensions with and without 10e4 M DCCD were allowed to glycolyze on the pH- stat at pH 7.5. Substrates to be transported were added 2 min later. Left, 32Pi, 0.2 mM. Right, W-L-alanine, 5 X low5 M.
The curves for uptake in the absence of glucose, not shown, approximately coincide with the solid symbols.
by guest on June 21, 2018http://w
ww
.jbc.org/D
ownloaded from

Issue of May 10, 1969 F. M. Harold, J. R. Baarda, C. Baron, and A. Abrams 2267
up K+, phosphate, or alanine. This is consistent with the finding (above) that removal of excess DCCD did not reactivate the membrane-bound ATPase.
DISCUSSION
The effects of DCCD on the ATPase of isolated protoplast membranes of X. juecalis are essentially similar to those obtained earlier by Holloway et al. (al), Beechey et al. (22), Roberton et al. (37), and Bulos and Racker (23) with mitochondria. DCCD inhibits the membrane-bound ATPase but not the solubilized enzyme (Fig. 1). The activity of DCCD-inhibited membrane-bound ATPase can be fully restored by releasing the enzyme from the membrane. Conversely, the sensitivity of the solubilized enzyme to DCCD is restored when the ATPase is reattached to the empty sites on membranes depleted of ATPase (Fig. 3). Finally, thereactionof DCCD with membrane- bound ATPase is dependent both on time and on DCCD con- centration (Fig. 2); a covalent reaction of DCCD with the membrane (22) is probable, although not established. In all, the results leave little doubt that the primary effect of DCCD is on a membrane component other than the ATPase protein itself, and that the inhibitory action is transmitted secondarily to the attached enzyme.
The characteristics of the ATPase (21-23, 37, 38) and of certain other enzymes (39) of mitochondria and chloroplasts are modified when they are associated with membranes; the term “allotopy” was coined by Racker (40) to designate this phenome- non. The present finding, that the sensitivity of S. jaecalis membrane-bound ATPase to inhibition by DCCD is lost when the enzyme is released from the membrane, warrants extension of the concept of allotopy to the bacterial plasma membrane. Let us note here that the ATPases of mitochondria and of S. jaecalis serve quite different functions-the former is thought to be involved in ATP synthesis, whereas the latter may consume ATP to energize membrane transport. It is therefore re- markable that ATPases from these diverse sources respond to DCCD in essentially the same manner. The fundamental unity of function suggested by this observation is an essential feature of Mitchell’s chemiosmotic hypothesis (2).
Let us now turn to the effects of DCCD on ATP metabolism in intact cells. Concentrations of DCCD comparable to those which inhibit the membrane-bound ATPase in vitro strongly inhibit ATP degradation in vivo, and also reduce the rate of glycolysis, but apparently do not interfere with the generation of ATP (Figs. 4 to 7). It seems reasonable to conclude that DCCD inhibits the membrane-bound ATPase in the intact cell, and that this ATPase catalyzes ATP degradation in vivo, at least in nongrowing cells which must turn over ATP in order to maintain glycolysis. This interpretation is consistent with the observation that the inhibition of glycolysis by DCCD was prevented by arsenate, which probably dissociates lactic acid production from the generation and turnover of ATP (33). Our observations are thus in accord with concepts originally formu- lated by Meyerhof (41) to explain the “Harden-Young effect,” i.e. the accumulation of hexose phosphates in cell-free extracts of yeast. Whether hexose phosphates accumulate in S. jaecalis inhibited by DCCD has not been determined.
The other essential finding is that a number of energy-de- pendent transport processes were severely inhibited by DCCD. Net uptake of K+ by heterologous exchange for Na+ and H+ was completely blocked (Figs. 6 and 7), whether glucose or
arginine served as energy source. We and others (19, 20) have previously argued that, since both glycolysis and arginine metabolism provide energy for cation transport, the product common to these pathways, ATP, is the most likely immediate energy donor. DCCD probably inhibits ATP hydrolysis by the membrane-bound ATPase, but does not interfere with ATP production. Taking all of the results at face value, the most economical interpretation is that the ATPase is directly or indirectly involved in the transport of cations. Indeed, the same argument applied to the uptake of phosphate and of alanine, which are also inhibited by DCCD (Fig. 8).
Cells treated with DCCD, while not grossly leaky, do tend to lose K+ by exchange for Na+ from the medium; efflux of K+ was actually stimulated by the simultaneous presence of both DCCD and glucose (Table II). Although we do not fully understand this phenomenon, it is likely that the stimulation of K+ efflux by glucose plus DCCD is another aspect of the inhibi- tion by DCCD of net K+ uptake. In a previous paper (28) we proposed that net uptake and retention of K+ are mediated by an energy-dependent transport system which selects K+ for translocation inward, Na+ and H+ for extrusion from the cells. The membrane-bound ATPase may well be part of such a transport system, and its inhibition by DCCD could result in impairment of the capacity to discriminate between Na+ and K+. Consequently, K+-loaded cells glycolyzing in the presence of excess Na+ would tend to lose K+ by exchange for Naf. This admittedly speculative model also suggests an explanation for the observation that DCCD inhibits K+ accumulation by heter- ologous exchange for Na+ and H”, but not autologous exchange of &K for K+, since autologous exchange does not require discrimination between K+ and Na+.
The logical weakness of the argument that the ATPase mediates cation transport resides in the assumption that DCCD blocks only the ATPase. In fact, it is quite clear that DCCD reacts not with the ATPase protein per se but with an unidentified constituent of the membrane, and that the inhibition of ATPase activity is secondary. Moreover, there is at least partial inhibition of membrane-bound NADH dehydrogenase activity. DCCD is a highly reactive substance and may, for all we know, react with diverse sites exposed at the cell surface (21). Thus we cannot rigorously exclude the possibility that there is no causal relationship whatever between the inhibition of ATPase and the inhibition of transport, but we do consider this improbable.
In view of these reservations, it is useful to consider the effects of DCCD in the light of findings made with other systems. DCCD is an inhibitor of oxidative phosphorylation and coupled respiration in mitochondria (22, 37), and thus belongs to a heterogeneous class of inhibitors of which oligomycin and related antibiotics have been most extensively studied. Oligomycin, like DCCD, inhibits the membrane-bound but not the solu- bilized ATPase of the inner mitochondrial membrane (37, 38). The antibiotic prevents mitochondrial ion transport energized by ATP (42). Most relevant to the present work, oligomycin inhibits both cation transport and the Na+-K+-stimulated ATP- ase of erythrocytes (43) ; it will be recalled that erythrocytes, like X. jaecalis, rely upon glycolysis for the generation of metabolic energy. Very recently, Harris et al. (44) and Green et al. (45) have shown that oligomycin and rutamycin prevent the con- version of mitochondrial membranes into an energized state thought to be involved in both oxidative phosphorylation and in osmotic work. These analogies suggest an alternative inter-
by guest on June 21, 2018http://w
ww
.jbc.org/D
ownloaded from

2268 Inhibition of ATPase and of Ion Transport in S. faecalis Vol. 244, No. 9
pretation of our results: DCCD may react with sites on the 22. plasma membranes such that the membrane can no longer 23. undergo configurational changes associated with transport. X
’ If these conformational changes necessarily accompany ATP hydrolysis, an inhibition of both transport and ATPase activity 25.
would be expected. 26.
1. PULLMAN, M. E., AND SCHATZ, G., Annu. Rev. Biochem., 36,
2. 3. 4. 5.
539 (1967). MITCHELL, P., Biol. Rev. (Cambridge), 41, 445 (1966). SKOU. J. C., Phusiol. Rev.. 45. 596 (1965). AL&S, R.‘W.,&w. Rek Biochenk, 36; 727 (1967). ARRAMS, A., MCNAMARA, P., AND JOHNSON, F. B., J. Biol.
Chem., 236, 3659 (1960).
28.
29.
30. 31.
6. DRAPEAU, G. R., AND MACLEOD, R. A., J. Bacterial., 86, 1413 (1963).
7. WEIBULL. C.. GREENAWALT. J. W.. AND L6w. H.. J. Biol. 32. 33.
8. Chem.,i37,‘847 (1962). ’ ’
I I
BOSE, S. K., AND GEST, I-I., Biochem. Biophys. Acta, 96, 159 (1965).
34.
9. 10.
35. MORA, J., AND SNELL, E. E., Biochemistry, 2, 136 (1963). 36. HAROLD, F. M., AND BAARDA, J. R., J. Bacterial., 96, 2026
11. 12.
ROTTEM, S., AND RAZIN, S., J. Bacterial., 92, 714 (1966). MUNOZ, ‘E.,‘FREER, J. fi., ELLAR, D. J., AND SALT&N, M. R. J.,
Biochim. Biovhus. Acta. 160. 531 (1968). GROSS, R., AND COLES, fi. W:, J. ikzctekiol., 96, 1322 (1968). HAFKENSCHEID, J. C. M., AND BONTING, S. L., Biochim. Bio-
phys. Acta, 161, 204 (1968).
37.
13. DEIBEL, R. L., Bacterial. Rev., 28, 330 (1964). 14. NEUJAHR, H. Y., HANSSON, E., AND FERM, R., Acta Chem.
&and., 21, 182 (1967). 15. 16. 17. 18. 19.
ABRAMS, A., J. Biol. Chem., 240, 3675 (1965). ABRAMS, A., AND BARON, C., Biochemistry, 6, 225 (1967). ABRAM& A.; AND BARON, C., Biochemistry, 7, 501 (1968). ABRAMS, A., J. Biol. Chem., 236, 1281 (1960). ZARLENGO, M., AND SCHULTZ, S. G., Biochim. Biophys. Acla,
38. 39. 40. 41. 42.
43.
126, 308 (1966). 44. 20. HAROLD, F. M., AND BAARDA, J. R., J. Bacterial., 96,816 (1968). 21. HOLLOWAY, C. T., ROBERTON, A. M., KNIGHT, I. G., AND 45.
BEECHEY, It. B., Biochem. J., 100, 79 P (1966).
REFERENCES 27.
BEECHEY, R. B., ROBERTON, A. M., HOLLOWAY, C. T., AND KNIGHT, I. G., Biochemistry, 6, 3867 (1967).
BULOS, B., AND RACKER, E., J. Biol. Chem., 243, 3891 (1968). FISKE, C. H., .~ND SUBBAROW, Y., J. Biol. Chem., 66, 375
(1925). LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L.. AND RANDALL,
R. J., J. Biol. Chem., 193, 265 (1951). ABRAMS, A., J. Biol. Chem., 234,383 (1959). ZARLENGO, M., AND ABRAMS, A., Biochim. Biophys. Acta, 71,
66 (1963). HAROLD, F. M., HAROLD, R. L., BAARDA, J. R., AND ABRAMS,
A., Biochemistry, 6, 1777 (1967). FORREST, W. W., AND WALKER, D. J., J. Bacterial., 89, 1448
(1965). HAROLD F. M., AND BAARDA, J. R., J. Bacterial., 94, 53 (1967). ABRAMS, A., Abstracts Fifth International Congress of Bio-
chemistry, Moscow, 1961, London, Pereamon Press, 1961, p. 273.
FORREST, W. W., J. Bacterial., 90, 1013 (1965). HAROLD, F. M., AND BAARDA, J. R., J. Bacterial., 91, 2257
(1966). HAROLD, F. M., HAROLD, R. L., AND ABRAMS, A., J. Biol.
Chem., 240, 3145 (1965).
(1968)‘. ROBERTON, A. M., HOLLOWAY, C. T., KNIGHT, I. G., AND
BEECHEY. R. B.. Biochem. J.. 108. 445 (1968). KAGAWA, Y., AND PACKER, E., i. Biol. Chem.,b41,2461 (1966). WOODWARD, D. O., Fed. P,roc., 27, 1167 (1968). RACKER, E., Fed. Proc., 26, 1335 (1967). MEYERHOF, O., J. Biol. Chem., 180, 575 (1949). BIELAWSKI, J., AND LEHNINGER, A. L., J. Biol. Chem., 241,
4316 (1966). WHITTAM, R., WHEELER, K. P., AND BLAKE, A., Nature, 203,
720 (1964). HARRIS, R. A., PENNISTON, J. T., ASAI, J., AND GREEN, D.
E., Proc. Nat. Acad. Sci. U. S. A., 69, 830 (1968). GREEN, D. E., ASAI, J., HARRIS, R. A., AND PENNISTON, J.
T., Arch. Biochem. Biophys., 126, 684 (1968).
by guest on June 21, 2018http://w
ww
.jbc.org/D
ownloaded from

Franklin M. Harold, James R. Baarda, Carl Baron and Adolph Abrams'-DicyclohexylcarbodiimideN,N by Streptococcus faecalisTransport in
Inhibition of Membrane-bound Adenosine Triphosphatase and of Cation
1969, 244:2261-2268.J. Biol. Chem.
http://www.jbc.org/content/244/9/2261Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/244/9/2261.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 21, 2018http://w
ww
.jbc.org/D
ownloaded from





























