Embed Size (px)
Citation preview

REVIEW
Inhibitors of Protein Tyrosine Phosphatase 1B from Marine NaturalProducts
Yue Zhou,a,b,1 Weirui Zhang,a,b,1 Xiaoyu Liu,a,b Haobing Yu,a,b Xiaoling Lu,*a,b and Binghua Jiao*a,b
aMarine Biopharmaceutical Institute, Second Military Medical University, Xiangyin Road 800, Shanghai 200433, P. R.China, e-mail: [email protected], [email protected]
bDepartment of Biochemistry and Molecular Biology, College of Basic Medical Sciences, Second Military MedicalUniversity, Shanghai 200433, P. R. China, e-mail: [email protected], [email protected]
The ocean is a capacious area with the most abundant biological resources on the earth. The particularity of themarine ecological environment (high pressure, high salt, and hypoxia) makes the marine species survival competitionfiercely, forcing many marine organisms in the process of life to produce a great deal of secondary metabolites withspecial structures and biological activities. In this article, 118 natural products which were isolated from four kinds ofmarine organisms, sponges, algae, soft corals and fungus, showing PTP1B inhibitory activity were summarized from2010 to 2016, which may become the leading compounds towards treating Diabetes mellitus (DM). What’s more, webriefly summarized the structure–activity relationship of PTP1B inhibitors.
Keywords: Natural products, Secondary metabolites, PTP1B, Inhibitory activity.
Contents1. Introduction2. Sponges3. Algae4. Soft Corals5. Fungi6. Conclusions
1. Introduction
The ocean is a huge treasure trove of natural prod-ucts, from where many drugs have been developedagainst various diseases.
Protein tyrosine phosphatase 1B (PTP1B) belongsto the protein tyrosine phosphatase family, whichspecifically hydrolyses the aromatic phosphate. Onone hand, in insulin signaling, it plays a key role as anegative regulator through dephosphorylating acti-vated insulin receptor (IR) and insulin receptor sub-strate (1IRS-1). Besides, under conditions of growthfactor receptor bound protein (Grb2), the
dephosphorylation of IRS-1 catalyzed by PTP1B wasaccelerated. The overexpression of PTP1B in the tissuecells will reduce protein tyrosine kinase (PTK) activity,the insulin receptor cannot be combined with insulin,thereby causing insulin resistance.[1 – 3] On the otherhand, in the JAK-STAT signaling pathway, which is themain axis of leptin receptor signaling transduction,inactive leptin receptor can couple with janus kinase(JAK) to get tyrosine kinases activity. The leptin recep-tor related kinase JAK2 and activator of transcription-3(STAT3) can be dephosphorylated by PTP1B and beinactive, leading leptin receptor does not respond toleptin and induce leptin resistance.[4 – 7]
These may all induce the occurrence and develop-ment of type-2 diabetes mellitus (T2DM) and obesity.Diabetes mellitus (DM) is a chronic endocrine disordermetabolic disease, which is characterized by hyper-glycemia. Among all the types, the overwhelmingmajority is T2DM, which is based on insufficient insu-lin secretion and insulin resistance. In recent years,PTP1B has become a new target as a negative regula-tory factor to treat T2DM and obesity.[8]
Based on these previous studies, PTP1B, as a newdiabetes target, sparked the interest of the scientiststo find new secondary metabolites as PTP1B inhibitor
1 Co-first authors who contributed equally in thismanuscript.
DOI: 10.1002/cbdv.201600462 Chem. Biodiversity 2017, 14, e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland
(1 of 12) e1600462
with corresponding PTP1B inhibitory activity. Accord-ing to the specifically structures and activities of themarine natural compounds, we summarized the newcompounds isolated from marine organisms in fouraspects, sponges, algae, soft corals and fungi, possess-ing PTP1B inhibitory activity from 2010 to 2016 in thisreview.
2. Sponges
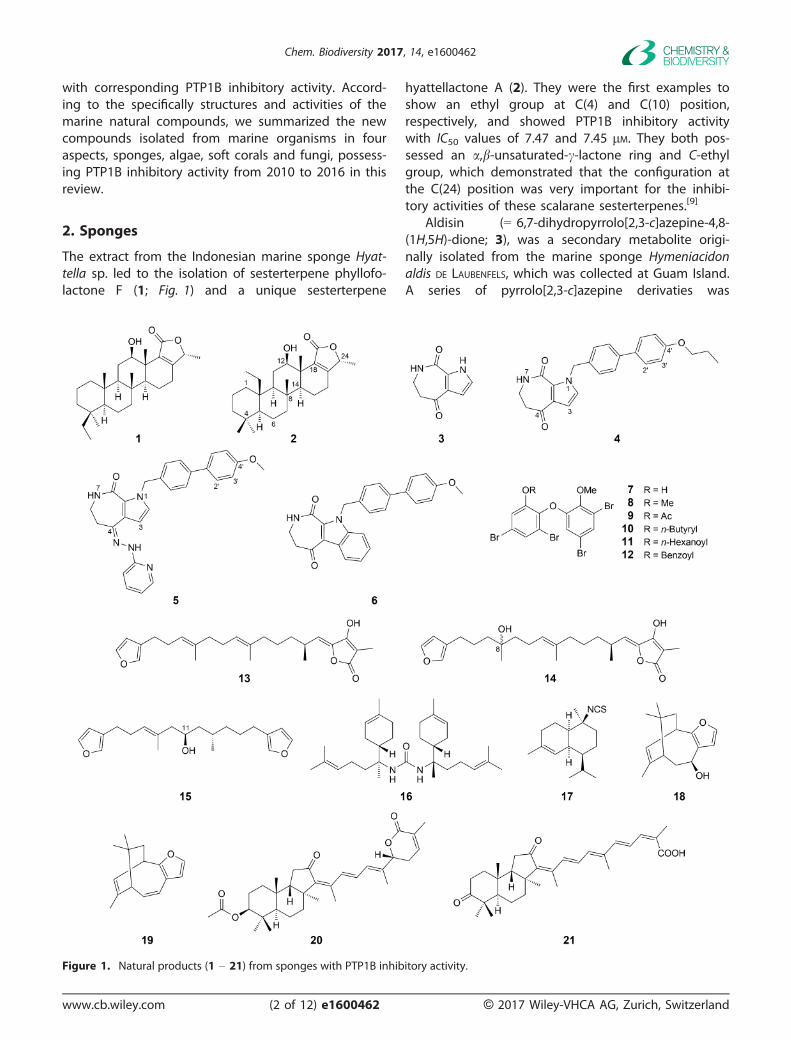
The extract from the Indonesian marine sponge Hyat-tella sp. led to the isolation of sesterterpene phyllofo-lactone F (1; Fig. 1) and a unique sesterterpene
hyattellactone A (2). They were the first examples toshow an ethyl group at C(4) and C(10) position,respectively, and showed PTP1B inhibitory activitywith IC50 values of 7.47 and 7.45 lM. They both pos-sessed an a,b-unsaturated-c-lactone ring and C-ethylgroup, which demonstrated that the configuration atthe C(24) position was very important for the inhibi-tory activities of these scalarane sesterterpenes.[9]
Aldisin (= 6,7-dihydropyrrolo[2,3-c]azepine-4,8-(1H,5H)-dione; 3), was a secondary metabolite origi-nally isolated from the marine sponge Hymeniacidonaldis DE LAUBENFELS, which was collected at Guam Island.A series of pyrrolo[2,3-c]azepine derivaties was
Figure 1. Natural products (1 – 21) from sponges with PTP1B inhibitory activity.
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (2 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland

synthesized, in which 4, 5, and 6 showed interestinginhibition against PTP1B with IC50 values of 16.36,14.93, 13.92 lM, respectively. Primary structure–activityrelationships (SARs) analysis showed that the biphenylmoiety at 1-positon and aromatic groups at 4-positoncontributed to the increase of inhibitory activity whilethe indoloazepine analogue 6 exhibiting comparableinhibitory potency implied another scaffold for designof PTP1B inhibitor.[10]
Bioassay-guided separation of the ethanol extractof an Indonesian marine sponge Lamellodysidea herba-cea led to the isolation of 2-(30,50-dibromo-20-methoxy-phenoxy)-3,5-dibromophenol (7). Furthermore, itsmethyl ether derivative 8 and four ester derivatives(acetyl (9), butyryl (10), hexanoyl (11), and benzoyl(12)) were prepared. All of them showed potent inhi-bitory activity with IC50 values of 0.85, 1.7, 0.62, 0.68,0.69, and 0.97 lM, respectively. Among them, com-pound 8 may be the most interesting one as a newleading compound possessing potent PTP1B inhibitoryactivity and no apparent cytotoxicity.[11]
Separation of the EtOH extracts of two Indonesiamarine sponges led to the isolation of two furanoses-terterpenoids (7E,12E,18S,20Z)-variabilin (13) and(12E,18S,20Z)-8-hydroxyvariabilin (14) from Ircinia sp.and a C21 furanoterpenoid furospongin-1 (15) fromSpongia sp., which inhibited PTP1B activity with IC50values of 1.5, 7.1, and 9.9 lM, respectively. The com-parison of compounds 13 and 14 indicating that theOH group at C(8) in 14 was unfavorable for the inhibi-tion of PTP1B activity.[12]
N,N’-Bis[(6R,7S)-7-amino-7,8-dihydro-a-bisabolen-7-yl]urea (16), (1R,6S,7S,10S)-10-isothiocyanato-4-amor-phene (17) were isolated from the EtOH extract of theOkinawan marine sponge Axinyssa sp. collected at Iri-omote Island. Compound 16 inhibited PTP1B activitywith IC50 value of 1.9 lM, while 17 inhibited PTP1Bactivity in a dose-dependent manner with IC50 valuesof 17 lM.[13]
The sesquiterpene euryspongin A (18) was isolatedfrom a marine sponge Euryspongia sp., collecting at Iri-omote Island, Okinawa, Japan. Though this compoundshowed novel unique sesquiterpenes structure, nopotent inhibition activity towards PTP1B was found.Dehydroeuryspongin A (19), the dehydro derivative of18, had PTP1B inhibitory activity with IC50 value of3.6 lM.[14]
The acetone extract of the Hainan sponge Stellettasp., belonging to the family Ancorinidae, which mayhave taxonomic relationships with genus Jaspis of anunidentified sponge, due to the isolation of the samecompound, 22,23-dihydrostellettin D (20), yielded astrong PTP1B inhibitor named stellettin G (21) with
IC50 value of 4.1 � 0.9 lM. Besides, stellettin G alsoshowed moderate cytotoxic activity against A549 andHL-60 cells.[15]
A new meroditerpene, 26-O-ethylstrongylophorine-14 (22; Fig. 2), with known 26-O-methylstrongylophor-ine-16 (23), strongylophorines-3 (24), -15 (25) and -17(26), were isolated from the Okinawan marine spongeStrongylophora strongilate. Compounds 22 – 26 inhib-ited the activity of PTP1B with IC50 values of 8.7, 8.5,9.0, 11.9, and 14.8 lM, respectively. The compoundswhich possessed an acetal moiety (22, 23, 25) showedstronger inhibitory activities than those of the lactonederivatives and diol derivative (26).[16]
New hippolides-related sesterterpenes, 27 – 29,were isolated from the crude fractions with PTP1Binhibitory activity of the South China Sea sponge Hip-pospongia lachne. Compounds 27 and 29 exhibitedmoderate PTP1B inhibitory activities with IC50 valuesof 5.2 and 8.7 lM, while compound 28 with a weakIC50 values of 14 lM, respectively.[17]
A new steroidal ketone, (22E,24S)-24-methylcho-lesta-4,7,22,25-tetraene-3-one (30), with an ergosta-22,25-diene side chain, exhibited significant inhibitoryactivity against PTP1B with an IC50 value of4.27 � 0.55 lM, was obtained from the South ChinaSea marine sponge Xestospongia testudinaria.[18]
A new polyacetylene, named isopetrosynol (31),together with three known compounds, petrosynol(32), adociacetylene D (33), (5R)-3,15,27-triaconta-triene-1,29-diyn-5-ol (34) were isolated from the EtOHextract of the marine sponge Halichondria cf. paniceacollected at Iriomote Island. Compound 31 inhibitedPTP1B activity with IC50 values of 8.2 � 0.3 lM, whileits stereoisomer 32 showed only 28.9 � 4.5% inhibi-tion at 21.6 lM, indicating that the configurations ofOH groups at C(14) and C(17) markedly affected thisactivity. Besides, compounds 33 and 34 inhibitedPTP1B activity with IC50 values of 7.8 � 0.5 and 12.2� 0.5 lM, respectively.[19]
Four new sesquiterpene quinines, dysidavaronesA – D, possessing the novel ‘dysidavarane’ carbonskeleton, were isolated from the sponge Dysideaavara, South China Sea. Among them, dysidavaronesA (35) and D (36) suppressed the activity of PTP1Bwith IC50 values of 9.98 and 21.6 lM.[20]
Melophlin C (37) was isolated from the EtOHextract of the marine sponge Petrosia sp., collected inthe coral reefs of North Sulawesi, Indonesia in 2013,showing PTP1B inhibitory activity with an IC50 value of14.6 lM.[21]
Three novel polyketides, woodylides A – C, wereobtained from the ethanol extract of the South ChinaSea sponge Plakortis simplex, which showed new
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (3 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland
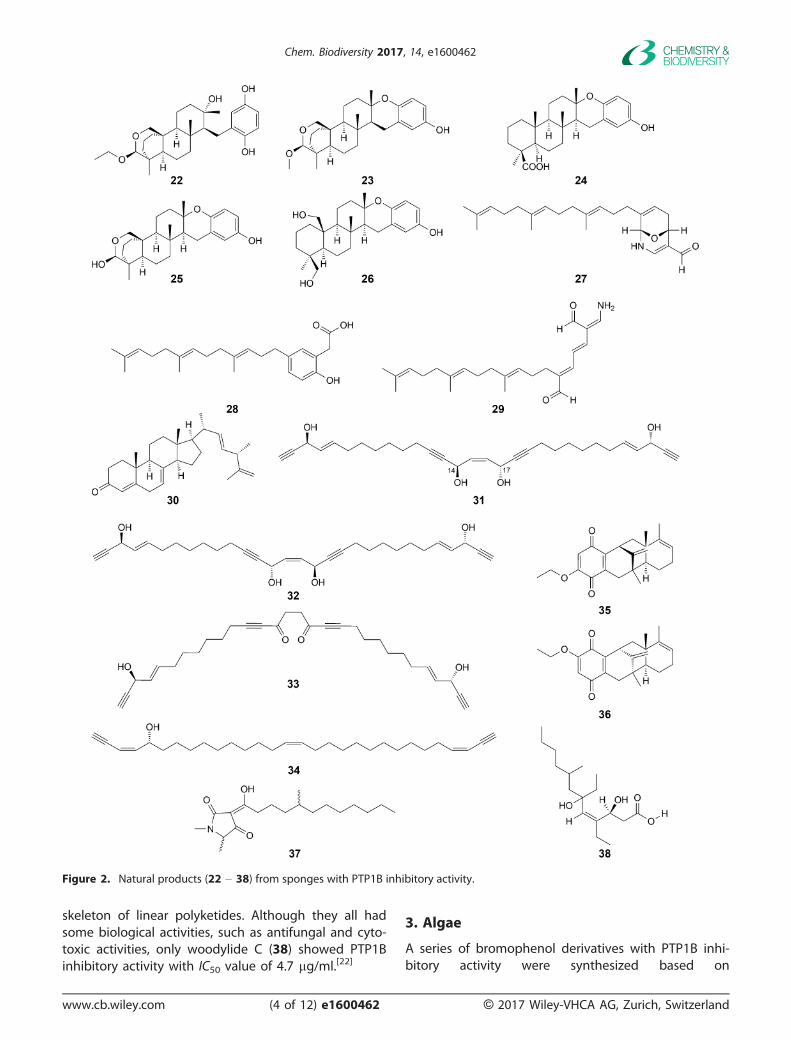
skeleton of linear polyketides. Although they all hadsome biological activities, such as antifungal and cyto-toxic activities, only woodylide C (38) showed PTP1Binhibitory activity with IC50 value of 4.7 lg/ml.[22]
3. Algae
A series of bromophenol derivatives with PTP1B inhi-bitory activity were synthesized based on
Figure 2. Natural products (22 – 38) from sponges with PTP1B inhibitory activity.
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (4 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland
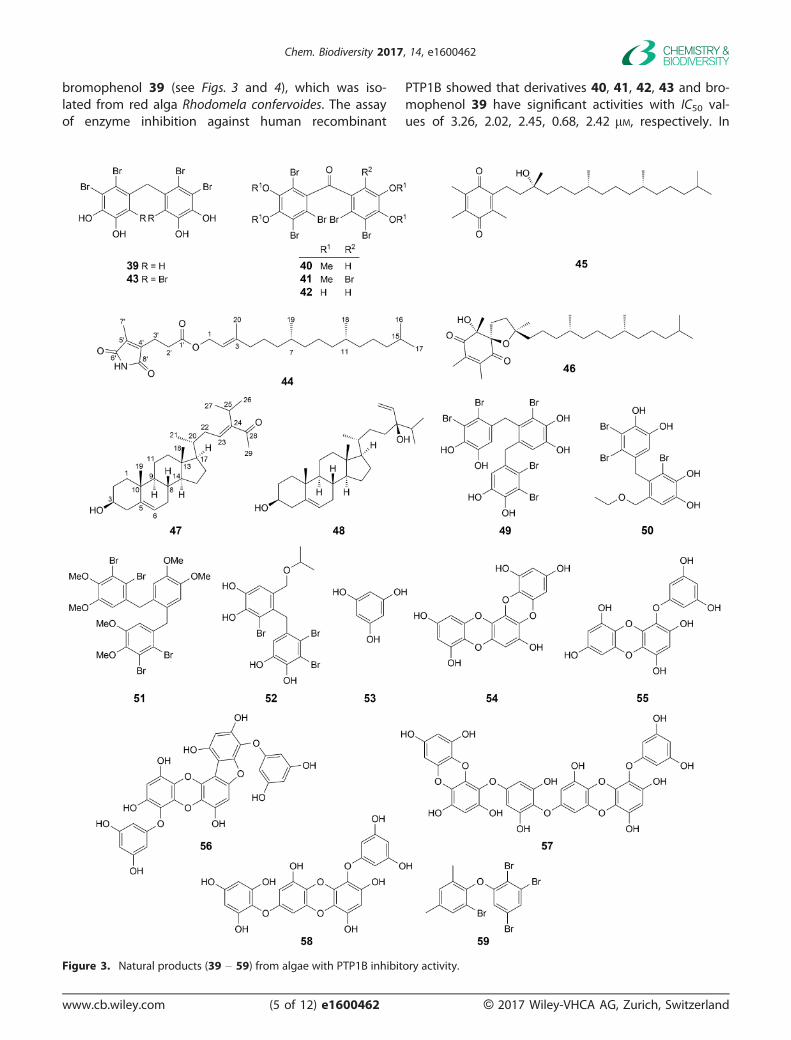
bromophenol 39 (see Figs. 3 and 4), which was iso-lated from red alga Rhodomela confervoides. The assayof enzyme inhibition against human recombinant
PTP1B showed that derivatives 40, 41, 42, 43 and bro-mophenol 39 have significant activities with IC50 val-ues of 3.26, 2.02, 2.45, 0.68, 2.42 lM, respectively. In
Figure 3. Natural products (39 – 59) from algae with PTP1B inhibitory activity.
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (5 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland
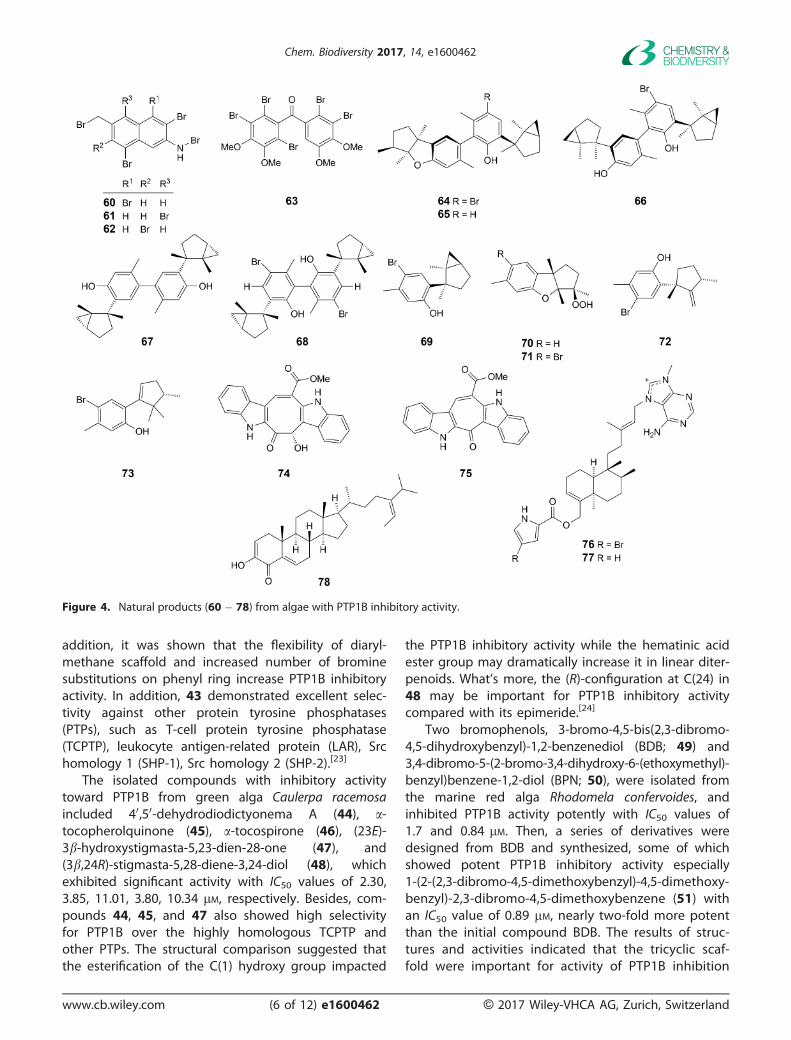
addition, it was shown that the flexibility of diaryl-methane scaffold and increased number of brominesubstitutions on phenyl ring increase PTP1B inhibitoryactivity. In addition, 43 demonstrated excellent selec-tivity against other protein tyrosine phosphatases(PTPs), such as T-cell protein tyrosine phosphatase(TCPTP), leukocyte antigen-related protein (LAR), Srchomology 1 (SHP-1), Src homology 2 (SHP-2).[23]
The isolated compounds with inhibitory activitytoward PTP1B from green alga Caulerpa racemosaincluded 40,50-dehydrodiodictyonema A (44), a-tocopherolquinone (45), a-tocospirone (46), (23E)-3b-hydroxystigmasta-5,23-dien-28-one (47), and(3b,24R)-stigmasta-5,28-diene-3,24-diol (48), whichexhibited significant activity with IC50 values of 2.30,3.85, 11.01, 3.80, 10.34 lM, respectively. Besides, com-pounds 44, 45, and 47 also showed high selectivityfor PTP1B over the highly homologous TCPTP andother PTPs. The structural comparison suggested thatthe esterification of the C(1) hydroxy group impacted
the PTP1B inhibitory activity while the hematinic acidester group may dramatically increase it in linear diter-penoids. What’s more, the (R)-configuration at C(24) in48 may be important for PTP1B inhibitory activitycompared with its epimeride.[24]
Two bromophenols, 3-bromo-4,5-bis(2,3-dibromo-4,5-dihydroxybenzyl)-1,2-benzenediol (BDB; 49) and3,4-dibromo-5-(2-bromo-3,4-dihydroxy-6-(ethoxymethyl)-benzyl)benzene-1,2-diol (BPN; 50), were isolated fromthe marine red alga Rhodomela confervoides, andinhibited PTP1B activity potently with IC50 values of1.7 and 0.84 lM. Then, a series of derivatives weredesigned from BDB and synthesized, some of whichshowed potent PTP1B inhibitory activity especially1-(2-(2,3-dibromo-4,5-dimethoxybenzyl)-4,5-dimethoxy-benzyl)-2,3-dibromo-4,5-dimethoxybenzene (51) withan IC50 value of 0.89 lM, nearly two-fold more potentthan the initial compound BDB. The results of struc-tures and activities indicated that the tricyclic scaf-fold were important for activity of PTP1B inhibition
Figure 4. Natural products (60 – 78) from algae with PTP1B inhibitory activity.
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (6 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland
and the multi-bromine atoms (4 – 5) attached to thearyl rings for PTP1B inhibition. In addition, 3,4-dibromo-5-(2-bromo-3,4-dihydroxy-6-(isopropoxymethyl)benzyl)benzene-1,2-diol (HPN; 52) was a synthetic ana-logue of BPN with enhanced inhibitory activity againstPTP1B with IC50 value at 0.63 lM and highly selectivityagainst other PTPs (TCPTP, LAR, SHP-1, SHP-2).[25][26]
Six phlorotannins, phloroglucinol (53), dioxinode-hydroeckol (54), eckol (55), phlorofurofucoeckol-A(56), dieckol (57), and 7-phloroeckol (58), were iso-lated from two perennial brown algae, Ecklonia
stolonifera and Eisenia bicyclis, belonging to the familyLaminareaceae in the mid-Pacific coast. Compounds55 – 58 showed stronger inhibitory activity againstPTP1B than the positive control with IC50 values of2.64 � 0.04, 0.56 � 0.10, 1.18 � 0.02, and 2.09 �0.09 lM, respectively, while the other two compoundsshowed moderate activity (Table 1).[27]
Five new highly brominated metabolites, 2-bromo-4,6-dimethylphenyl 2,3,5-tribromophenyl ether (59),1,2,5-tribromo-3-bromoamino-7-bromomethylnaphthalene(60), 2,5,8-tribromo-3-bromoamino-7-bromomethylna-
Table 1. Natural products from marine organisms with weak activity
Location Sources/Species Metabolite IC50 (lM) References
The South ChinaSea
Sponge Hippospongialachne
Hippolide A (96)Hippolide B (97)
96: 23.8197: 39.67
[41]
Arctic regions Sponge Stryphnus fortis Ianthelline (98) Weak, dose-dependent manner [42]Sponge Aplysina spp. 99 – 102 99, 100: Selective activity
101, 102: moderate activity[43]
Okinawan Sponge Strongylophorastrongilate
Strongylophorine-2 (103)Strongylophorine-8 (104)
103: > 24.4104: 21.2
[16]
The South ChinaSea
Sponge Hippospongialachne
Compound 105 33 [17]
Iriomote Island Sponge Axinyssa sp. Axinisothiocyanate J (106) 50% inhibition at 36 lM [13]The East ChinaSea
Red alga Laurenciaokamurai YAMADA
Compound 69 45.2 [29]
Hainan coastlinesof China
Red alga Laurencia similis 1,2,5-Tribromo-3-bromoamino-7-bromomethylnaphthalene (60)2,5,8-Tribromo-3-bromoamino-7-bromomethylnaphthalene (61)2,5,6-Tribromo-3-bromoamino-7-bromomethylnaphthalene (62)
60: 10261: 65.362: 69.8
[28]
Mid-Pacific coast Ecklonia stolonifera andEisenia bicyclis
Phloroglucinol (53)Dioxinodehydroeckol (54)
53: 55.48 � 1.8554: 29.97 � 4.52
[27]
Green alga Caulerparacemosa
trans-Phytol (107)trans-Phytyl acetate (108)(3b,24S)-Stigmasta-5,28-diene-3,24-diol (109)(22E)-3b-Hydroxycholesta-5,22-dien-24-one (110)
107: 32.6108: 50.02109: 41.70110: 49.97
[24]
South China Sea Soft coralSarcophytontrocheliophorumMarenzeller
Cembrene C (111)Ketoemblide (112)
111: 26.6112: 27.2
[34]
Yalong Bay,Hainan Province,China
The soft coralsS. flexibilis
7a-Hydroxycrassarosterol A (113) 33.05 [44]
Lingshui Bay,Hainan Province
The soft coral Sinulariadepressa TIXIER-DURIVAULT
Compound 114 22.7 [36]
Wan Island, Korea Penicillium sp. SF-5203Penicillium sp. SF-5295
Cyclopenol (115)Echinulin (116)Viridicatol (117)
115: 30116: 29.4117: 64
[40]
Daya Bay,Shenzhen city,GuangdongProvince, China
Endophytic fungusGuignardia sp.KcF8derived of a mangroveplant Kandelia candel
Guignardin C (118) 25.7 [45]
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (7 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland

phthalene (61), 2,5,6-tribromo-3-bromoamino-7-bro-momethylnaphthalene (62), and 20,50,60,5,6-penta-bromo-30,40,3,4-tetramethoxybenzophenone (63), wereisolated from the red alga Laurencia similis, collectingfrom Hainan coastlines of P. R. China. Compounds 59and 63 showed strong PTP1B inhibitory activities withIC50 values of 2.97 and 2.66 lM, and the other threenew compounds showed moderate inhibitory activityagainst PTP1B (Table 1).[28]
From the East China Sea red alga Laurencia okamu-rai YAMADA, three novel heterodimeric laurane-typesesquiterpenoids, laurokamurols A – C (64 � 66),were isolated along with eight known related mono-meric ones. The results for testing their activitiesshowed that all of the dimers, compounds 64 – 66,and two synthetic compounds 67 and 68 exhibitedsignificant inhibitory effect of PTP1B with IC50 valuesof 8.1, 12.5, 6.1, 10.0, and 5.6 lM, respectively, compa-rable with that of the positive control oleanolic acid(IC50 = 3.3 lM). All of the isolated monomers were notactive, except for 69, which showed weak inhibitoryactivity.[29]
The red alga Laurencia okamurai, collected off thecoast of Nanji Island, Zhejiang Province, China, inDecember 2013 led to the isolation of newsesquiterpenoid, debromo-3b-hydroperoxyaplysin(70), along with known compounds 3b-hydroperoxy-aplysin (71), isolaurinterol (72), and laurokamurene A(73) with PTP1B inhibitory activity with IC50 values of13.0, 14.9, 7.0, and 4.9 lg/ml, respectively. In a pre-liminary SAR study, both 70 and 71 with (R)-config-uration at C(3) position exhibited strong PTP1Binhibitory activities, while their epimers were inactive,
suggesting that the may be significant for the activ-ity.[30]
The investigation of the Chinese marine green algaCaulerpa racemosa resulted in the discovery of a newminor bisindole alkaloid, racemosin C (74), and oneknown related metabolite, caulersin (75), which werecharacterized by a naturally unprecedented 8-hydroxy-2,4,6-cyclooctatrienone ring fused with two indole sys-tems. They both exhibited significant PTP1B inhibitoryactivity with IC50 values of 5.86 � 0.57 and 7.14 �1.00 lM, respectively.[31]
Agelasine G (76), and ageline B (77) were isolatedfrom the Okinawan marine sponge Agelas nakamuraiHOSHINO. Compound 76 inhibited protein tyrosine phos-phatase 1B (PTP1B) activity with an IC50 value of 15 lM,but the debromo derivative (77) was not active indicat-ing that the Br atom seems necessary for its activity.[32]
In order to obtain bioactive second metabolites, anew sterol named thunberol (78) was isolated in thebrown alga Sargassum thunbergii, a kind of prolificseaweed which grew along the coast of East ChinaSea, exhibiting significant PTP1B inhibitory activitywith an IC50 value of 2.24 lg/ml.[33]
4. Soft Corals
There are some cembrane diterpenoids with anti-PTP1B activity were isolated from the South China Seasoft coral Sarcophyton trocheliophorum MARENZELLER. Theisolated compounds were sarcophytonolide N (79; seeFig. 5), sarcrassin E (80), and (4Z,12Z,14E)-sarcophyto-lide (81), which showed significant inhibitory activityagainst human PTP1B enzyme with IC50 values at 5.95,
Figure 5. The natural products (79 – 85) from soft corals with PTP1B inhibitory activity.
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (8 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland

6.33, and 15.4 lM, respectively. Later, a new cap-nosane diterpene sarsolilides B (82) and its knownanalogue sarsolilide A (83) were isolated from this softcoral, and exhibited PTP1B inhibitory activity with IC50values of 27.1 � 2.6 and 6.8 � 0.9 lM. Based on thepreliminary comparison between the structures ofthose compounds, it’s provided that methyl estergroup at C(18) significantly increases the enzyme inhi-bitory activity.[34][35]
(3b,4a,5a)-4-Methylergost-24(28)-ene-3-ol (84) andergosta-4,24(28)-dien-3-one (85) were isolated fromthe soft coral Sinularia depressa TIXIER-DURIVAULT off Ling-shui Bay, Hainan Province, exhibiting inhibitory activitywith IC50 values of 19.4, 15.3 lM. It was indicated thatsubstitution of 8-OH should be responsible for thedecrease of PTP1B inhibitory activity while acetylationon 3-OH may increase it.[36]
5. Fungi
A new styrylpyrone-type metabolite penstyrylpyrone(86; see Fig. 6) was isolated from the methylethylke-tone extract of marine-derived fungus Penicillium sp.JF-55 cultures, along with two known metabolites,anhydrofulvic acid (87) and citromycetin (88). Com-pounds 86 and 87 were suggested to bind to the active
site within PTP1B, exhibiting PTP1B inhibitory activity ina competitive manner with IC50 values of 5.28 and1.90 lM, while compound 88, which shared close struc-tural features to 87, did not show any inhibitory activityup to 25.8 lM, which manifested that the linear tricyclicsystem and the position of the carbonyl groups in com-pound 87 might be important for the binding withactive site of PTP1B.[37]
From the Indonesian ascidian-derived Penicilliumverruculosum strain TPU1311, a new merosesquiter-pene, verruculide A (89), and two known congeners,chrodrimanins A (90) and H (91), were isolated anddemonstrated to have PTP1B inhibitory activity. Thestudy showed that 89, 90, and 91 inhibited the activ-ity of PTP1B with IC50 values of 8.4, 8.5, and 14.9 lM,respectively. According to the comparison of theiractivities and structures with others revealed that theacetylation of the 40-OH group rather than the twoOH groups at the C(7) and C(70) positions should beresponsible for the significantly reducing of the PTP1Bactivity. It was the first time demonstrating com-pounds in the chrodrimanin family can exhibit inhibi-tory activity against PTP1B.[38]
Tanzawaic acids A (92) and B (93) were discoveredfrom a marine-derived fungus Penicillium sp. SF-6013and showed equivalent inhibitory effects against
Figure 6. The natural products (86 – 95) from fungi with PTP1B inhibitory activity.
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (9 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland
PTP1B activity with the IC50 value of 8.2 lM. They wereisolated from the sea urchin Brisaster latifrons, col-lected from the Sea of Okhotsk (N 53°22.6260 E
144°24.2000). The assay based on structural–activityrelationships between tanzawaic acids and its deriva-tives suggested that the presence of the hydroxy
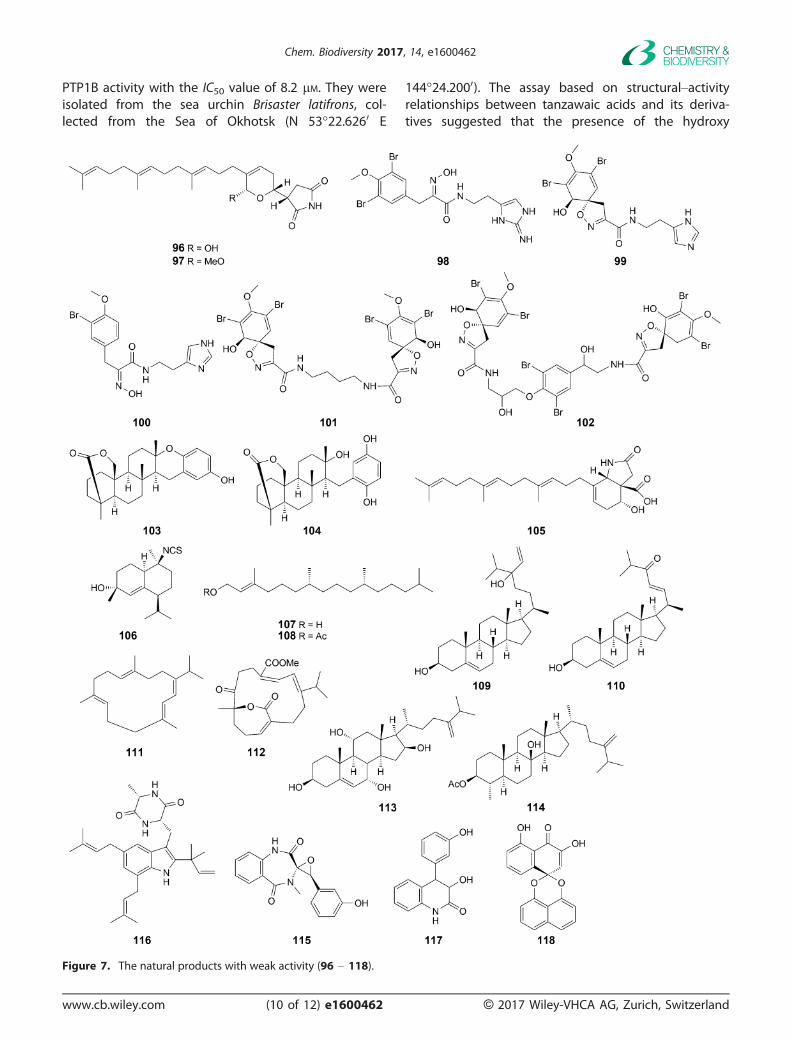
Figure 7. The natural products with weak activity (96 – 118).
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (10 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland

group at C(10) might be the important structure indiminishing PTP1B inhibitory.[39]
Fructigenine A (94) was isolated from the strainPenicillium sp. SF-5203, an intertidal sediment sample,collected from Wan Island, Korea, in January 2008. Fla-voglaucin (95) was extracted from the strain SF-5295,isolated from an unidentified sponge manually col-lected using scuba diving equipment off the shores ofJeju Island in February 2009. Compound 94 exhibitedthe PTP1B inhibitory activity in a noncompetitive man-ner with an IC50 value of 10.7 lM, while the IC50 valueof Compound 95 was determined as 13.4 lM in acompetitive manner.[40]
There are also some compounds (Fig. 7) from mar-ine organisms with weak activity up to 20 lM summa-rizing in the Table 1.
6. Conclusion
In this review, 118 marine natural products from2010 to 2016 with PTP1B inhibitory activity werereported. To base on the sources, compounds withPTP1B inhibitory activity can be mostly classified asfollowing: sponges, algae, soft corals and fungi. Fig. 8shows the proportion of the four general categories.The figure shows that the activated products weremainly found in sponges and algae, which producedabundant secondary metabolites due to their uniquechemical defense system formed in the deep seaenvironment and surrounding with large amounts ofsymbiotic bacteria. However, more than 1000 newcompounds of marine natural products with particu-lar types of bioactivity were discovered every yearand the main focus of attention was the anticanceractivity. So more awareness of the detection ofPTP1B inhibitory activity should be raised in novelnatural compounds.
In view of the remarkable value of drug targetingin PTP1B, there has been a few methods screening forthe inhibitor, such as colorimetric method usingp-nitrophenyl phosphate (PNPP) as substrate, fluores-cence method, scintillation proximity assay (SPA) andso on. Besides, a high-throughput method was estab-lished based on those to screen high activity PTP1Binhibitors more efficiently. However, at present stagethose technologies may lead to the emergence of thefalse positive results which will bring more difficultiesto drug follow-up studies. Moreover, the mechanismactions of those compounds have not yet fully clearand prevent the further related research development.
In a word, the immaturity of the detectionmethod and the unclear mechanism of action ofthose compounds were responsible for the unlistedeffective PTP1B inhibitor drugs in the clinic. What’smore, as a new target for treating DM or obesity, alot of work still need to be carried out such as study-ing the drug metabolism and the side effects in thebody. Till now, there are only two PTP1B inhibitors inclinical trials: the MSI-1436 in the USA,[46] JTT-551 inJapan.[47]
In order to further development, searching for newactive natural compounds and derivatives as usefultargeted drugs, finding appropriate methods to testPTP1B inhibitory activity, strengthening the relatedmechanism research and establishing medicine modelstill need our efforts for a period time in the future. Inthese aspects, searching for effective leading com-pounds was the most fundamental and most urgentaffair we need to do in recent times.
Acknowledgements
The work was funded by National Natural ScienceFoundation of China (NSFC) (41306197, 41606173),Guangdong Innovative Development of Marine EconomyRegional Demonstration Project (GD2012-D01-001).
Conflict of Interest
The authors declare no conflict of interest.
References
[1] H. T. Le, S. Ponniah, C. J. Pallen, Biochem. Biophys. Res.Commun. 2004, 314, 321.
[2] S. Rocchi, S. Tartare-Deckert, D. Sawka-Verhelle, A. Gamha,E. van Obberghen, Endocrinol. 1996, 137, 4944.
[3] T. Tenev, H. Keilhack, S. Tomic, B. Stoyanov, M. Stein-Ger-lach, R. Lammers, A. V. Krivtsov, A. Ullrich, F. D. B€ohmer, J.Biol. Chem. 1997, 272, 5966.
[4] M. G. Mohi, B. G. Neel, Curr. Opin. Genet Dev. 2007, 17, 23.
Figure 8. Compounds distribution of PTP1B inhibitory activitiesin the four types of organism.
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (11 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland

[5] K. E. You-Ten, E. S. Muise, A. Iti�e, E. Michaliszyn, J. Wagner,S. Jothy, W. S. Lapp, M. L. Tremblay, J. Exp. Med. 1997, 186,683.
[6] S. Galic, M. Klingler-Hoffmann, M. T. Fodero-Tavoletti, M. A.Puryer, T.-C. Meng, N. K. Tonks, T. Tiganis, Mol. Cell. Biol.2003, 23, 2096.
[7] N. K. Tonks, C. D. Diltz, E. H. Fischer, J. Biol. Chem. 1988,263, 6722.
[8] D. X. Luo, H. E. Yao, X. P. Gao, Y. Z. Zhang, Nat. Prod. Res.Dev. 2005, 17, 606.
[9] D. B. Abdjul, H. Yamazaki, O. Takahashi, R. Kirikoshi, R. E.Mangindaan, M. Namikoshi, Bioorg. Med. Chem. Lett. 2015,25, 904.
[10] J. Xie, J. Tian, L. Su, M. Huang, X. Zhu, F. Ye, Y. Wan, Bioorg.Med. Chem. Lett. 2011, 21, 4306.
[11] H. Yamazaki, D. A. Sumilat, S. Kanno, K. Ukai, H. Rotinsulu,D. S. Wewengkang, M. Ishikawa, R. E. P. Mangindaan, M.Namikoshi, J. Nat. Med. 2013, 67, 730.
[12] D. B. Abdjul, H. Yamazaki, S. Kanno, D. S. Wewengkang, H.Rotinsulu, D. A. Sumilat, K. Ukai, M. M. Kapojos, M. Nami-koshi, Bioorg. Med. Chem. Lett. 2017, 27, 1159.
[13] D. B. Abdjul, S. Kanno, H. Yamazaki, K. Ukai, M. Namikoshi,Bioorg. Med. Chem. Lett. 2016, 26, 315.
[14] H. Yamazaki, T. Nakazawa, D. A. Sumilat, O. Takahashi, K.Ukai, S. Takahashi, M. Namikoshi, Bioorg. Med. Chem. Lett.2013, 23, 2151.
[15] D.-Q. Xue, S.-C. Mao, X.-Q. Yu, Y.-W. Guo, Biochem. Syst.Ecol. 2013, 49, 101.
[16] J.-S. Lee, D. B. Abdjul, H. Yamazaki, O. Takahashi, R. Kiri-koshi, K. Ukai, M. Namikoshi, Bioorg. Med. Chem. Lett.2015, 25, 3900.
[17] S.-J. Piao, W.-H. Jiao, F. Yang, Y.-H. Yi, Y.-T. Di, B.-N. Han, H.-W. Lin, Mar. Drugs 2014, 12, 4096.
[18] W.-F. He, D.-Q. Xue, L.-G. Yao, J. Li, H.-L. Liu, Y.-W. Guo, J.Asian Nat. Prod. Res. 2016, 18, 195.
[19] D. B. Abdjul, H. Yamazaki, O. Takahashi, R. Kirikoshi, K.Ukai, M. Namikoshi, Chem. Pharm. Bull. 2016, 64, 733.
[20] W.-H. Jiao, X.-J. Huang, J.-S. Yang, F. Yang, S.-J. Piao, H.Gao, J. Li, W.-C. Ye, X.-S. Yao, W.-S. Chen, H.-W. Lin, Org.Lett. 2012, 14, 202.
[21] W. Maarisit, H. Yamazaki, S. Kanno, A. Tomizawa, H. Rotin-sulu, D. S. Wewengkang, D. A. Sumilat, K. Ukai, M. M. Kapo-jos, M. Namikoshi, Bioorg. Med. Chem. Lett. 2017, 27, 999.
[22] H.-B. Yu, X.-F. Liu, Y. Xu, J.-H. Gan, W.-H. Jiao, Y. Shen, H.-W. Lin, Mar. Drugs 2012, 10, 1027.
[23] D. Shi, J. Li, B. Jiang, S. Guo, H. Su, T. Wang, Bioorg. Med.Chem. Lett. 2012, 22, 2827.
[24] P. Yang, D.-Q. Liu, T.-J. Liang, J. Li, H.-Y. Zhang, A.-H. Liu,Y.-W. Guo, S.-C. Mao, Bioorg. Med. Chem. 2015, 23, 38.
[25] B. Jiang, D. Shi, Y. Cui, S. Guo, Arch. Pharm. 2012, 345, 444.[26] D. Shi, S. Guo, B. Jiang, C. Guo, T. Wang, L. Zhang, J. Li,
Mar. Drugs 2013, 11, 350.[27] H. E. Moon, N. Islam, B. R. Ahn, S. S. Chowdhury, H. S.
Sohn, H. A. Jung, J. S. Choi, Biosci. Biotechnol. Biochem.2011, 75, 1472.
[28] J. Qin, H. Su, Y. Zhang, J. Gao, L. Zhu, X. Wu, H. Pan, X. Li,Bioorg. Med. Chem. Lett. 2010, 20, 7152.
[29] X.-L. Li, T. Kurt�an, J.-C. Hu, A. M�andi, J. Li, X.-W. Li, Y.-W.Guo, J. Agric. Food Chem. 2017, 65, 1550.
[30] X.-L. Li, W.-F. He, J. Li, L.-F. Lan, X.-W. Li, Y.-W. Guo, J. AsianNat. Prod. Res. 2015, 17, 1146.
[31] H. Yang, D.-Q. Liu, T.-J. Liang, J. Li, A.-H. Liu, P. Yang, K. Lin,X.-Q. Yu, Y.-W. Guo, S.-C. Mao, B. Wang, J. Asian Nat. Prod.Res. 2014, 16, 1158.
[32] D. B. Abdjul, H. Yamazaki, S. Kanno, O. Takahashi, R. Kiri-koshi, K. Ukai, M. Namikoshi, J. Nat. Prod. 2015, 78, 1428.
[33] W.-F. He, L.-G. Yao, H.-L. Liu, Y.-W. Guo, J. Asian Nat. Prod.Res. 2014, 16, 685.
[34] L.-F. Liang, L.-X. Gao, J. Li, O. Taglialatela-Scafati, Y.-W. Guo,Bioorg. Med. Chem. 2013, 21, 5076.
[35] L.-F. Liang, T. Kurt�an, A. M�andi, L.-X. Gao, J. Li, W. Zhang,Y.-W. Guo, Eur. J. Org. Chem. 2014, 1841.
[36] L.-F. Liang, X.-J. Wang, H.-Y. Zhang, H.-L. Liu, J. Li, L.-F. Lan,W. Zhang, Y.-W. Guo, Bioorg. Med. Chem. Lett. 2013, 23,1334.
[37] D. S. Lee, J. H. Jang, W. Ko, K. S. Kim, J. H. Sohn, M. S.Kang, J. S. Ahn, Y. C. Kim, H. Oh, Mar. Drugs. 2013, 11,1409.
[38] H. Yamazaki, W. Nakayama, O. Takahashi, R. Kirikoshi, Y.Izumikawa, K. Iwasaki, K. Toraiwa, K. Ukai, H. Rotinsulu, D.S. Wewengkang, D. A. Sumilat, R. E. Mangindaan, M. Nami-koshi, Bioorg. Med. Chem. Lett. 2015, 25, 3087.
[39] T. H. Quang, N. T. T. Ngan, W. Ko, D.-C. Kim, C.-S. Yoon, J.H. Sohn, J. H. Yim, Y.-C. Kim, H. Oh, Bioorg. Med. Chem.Lett. 2014, 24, 5787.
[40] J. H. Sohn, Y.-R. Lee, D.-S. Lee, Y.-C. Kim, H. Oh, J. Microbiol.Biotechnol. 2013, 23, 1206.
[41] S.-J. Piao, H.-J. Zhang, H.-Y. Lu, F. Yang, W.-H. Jiao, Y.-H. Yi,W.-S. Chen, H.-W. Lin, J. Nat. Prod. 2011, 74, 1248.
[42] K. Ø. Hanssen, J. H. Andersen, T. Stiberg, R. A. Engh, J.Svenson, A.-M. Genevi�ere, E. Hansen, Anticancer Res. 2012,32, 4287.
[43] G. Carr, F. Berrue, S. Klaiklay, I. Pelletier, M. Landry, R. G.Kerr, Methods 2014, 65, 229.
[44] W.-T. Chen, H.-L. Liu, L.-G. Yao, Y.-W. Guo, Steroids 2014,92, 56.
[45] W. Ai, X. Wei, X. Lin, L. Sheng, Z. Wang, Z. Tu, X. Yang, X.Zhou, J. Li, Y. Liu, Tetrahedron 2014, 70, 5806.
[46] K. A. Lantz, S. G. E. Hart, S. L. Planey, M. F. Roitman, I. A.Ruiz-White, H. R. Wolfe, M. P. McLane, Obesity 2010, 18,1516.
[47] M. Ito, S. Fukuda, S. Sakata, H. Morinaga, T. Ohta, J. Dia-betes Res. 2014, 680348.
Received December 8, 2016Accepted February 27, 2017
Chem. Biodiversity 2017, 14, e1600462
www.cb.wiley.com (12 of 12) e1600462 © 2017 Wiley-VHCA AG, Zurich, Switzerland

本文献由“学霸图书馆-文献云下载”收集自网络,仅供学习交流使用。
学霸图书馆(www.xuebalib.com)是一个“整合众多图书馆数据库资源,
提供一站式文献检索和下载服务”的24 小时在线不限IP
图书馆。
图书馆致力于便利、促进学习与科研,提供最强文献下载服务。
图书馆导航:
图书馆首页 文献云下载 图书馆入口 外文数据库大全 疑难文献辅助工具





























