Embed Size (px)
Citation preview

of August 20, 2018.This information is current as
and Its Natural Deletion in C57BL/10 Miceof a Novel Goblet Cell Lectin, Intelectin-2,
: Inducible ExpressionTrichinella spiralisto Innate BALB/c Enteric Epithelial Responses
R. P. MillerHughWilliam H. Colledge, Jin-Kyu Lee, Michael Pierce and
Alan D. Pemberton, Pamela A. Knight, John Gamble,
http://www.jimmunol.org/content/173/3/1894doi: 10.4049/jimmunol.173.3.1894
2004; 173:1894-1901; ;J Immunol
Referenceshttp://www.jimmunol.org/content/173/3/1894.full#ref-list-1
, 6 of which you can access for free at: cites 26 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2004 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Innate BALB/c Enteric Epithelial Responses toTrichinellaspiralis: Inducible Expression of a Novel Goblet Cell Lectin,Intelectin-2, and Its Natural Deletion in C57BL/10 Mice1
Alan D. Pemberton,2* Pamela A. Knight,* John Gamble,† William H. Colledge,† Jin-Kyu Lee,‡
Michael Pierce,‡ and Hugh R. P. Miller*
Infection of mice with the nematode parasiteTrichinella spiralis induces changes in the proteome of the jejunal epithelium,including substantial up-regulation of a novel variant of interlectin. In this study we sequence this novel lectin, termed intelectin-2,and compare expression levels duringT. spiralis infection of resistant (BALB/c) with susceptible (C57BL/10) mouse strains.Intelectin-2 was cloned and sequenced from BALB/c mRNA extracted on day 14 of infection, and was found to have 91% aminoacid identity with intelectin (within our study termed intelectin-1). Intelectin-2 transcripts were up-regulated early (day 3) duringinfection with T. spiralis in BALB/c mice, suggesting an innate response, and levels remained high through to day 14 (time ofparasite rejection). Immunohistochemistry of jejunal sections with a rabbit polyclonal Ab to Xenopus laevis 35-kDa corticalgranule lectin (XL35; 68% identity with intelectin-2) followed a similar pattern, with intense labeling of goblet and Paneth cellsat day 14. However, intelectin-2 transcripts and protein were absent, and immunohistochemistry negative when C57BL/10 micewere infected with T. spiralis. Genomic PCR and Southern blotting confirmed that the intelectin-2 gene is absent from theC57BL/10 genome. The presence of intelectin-2 in resistant BALB/c mice, its absence from the susceptible C57BL/10 strain andthe kinetics of its up-regulation during T. spiralis infection suggest that this novel lectin may serve a protective role in the innateimmune response to parasite infection. The Journal of Immunology, 2004, 173: 1894–1901.
T he mucus layer that coats and protects the simple epitheliaof the gut and lung is composed of mucin glycoproteinsand additional components that help to protect the epithe-
lium against invasion by pathogens. In the gastrointestinal tract,such components include antibacterial defensins (1), lysozyme (2),trefoil factors (3), and members of the galactose-binding lectinfamily (galectins) (4). The mucus layer plays a significant role inthe response to intestinal nematodes in which the worms becomephysically entrapped in mucus and are expelled (reviewed in Ref.5). Mucus trapping is associated with changes in the physical prop-erties of mucus, and worm expulsion in primary infection is asso-ciated with goblet cell hyperplasia and augmented mucus synthesis(5). Certain lectins, such as the sheep galectin, ovgal11, also ap-pear to be up-regulated in response to parasite infection (6). Pro-posed roles for such lectins in the lumenal environment includeopsonization of bacteria and binding to mucins, to alter the vis-coelastic properties of mucus itself (4, 6).
We recently described (7) the appearance of a novel lectin inextracts of exfoliated jejunal epithelium from BALB/c mice in-fected with the nematode parasite Trichinella spiralis. This pro-
tein, which was absent in uninfected mice, appeared to be a closelyrelated variant of mouse intelectin, which is expressed by smallintestinal Paneth cells (8). Mouse intelectin belongs to a smallfamily of related lectins, with members also described in Xenopuslaevis (9–11) and in human (12, 13). Various roles have beenpostulated for the Xenopus lectins in fertility and development (10,11). Human intelectin (12) (also known as HL-1 (13) and lacto-ferrin receptor (14)) was expressed as a recombinant protein, andfound to bind oligosaccharides, including galactofuranose moetiesand to galactofuranose-containing bacterial cell wall preparations(12). Hence, a role in recognition of bacteria was proposed. Thislectin is expressed in the small intestine and colon, but is alsohighly expressed in heart tissue (12, 13), and has been immuno-localized to endothelium (13). The second human intelectin vari-ant, HL-2, was found to be expressed specifically in the smallintestine (13).
The aim of this study was to define the site of expression of thenovel mouse intelectin (7) and its relationship to the developmentof resistance against the intestinal nematode T. spiralis. We havesequenced this intelectin variant and compared its expression withthat of the previously described mouse intelectin (8). Our resultsshow that the expression of the intelectin variant is confined to theileum in normal BALB/c mice, but that during infection there isstrong jejunal expression in goblet and Paneth cells that peaks atthe time of worm expulsion. In contrast, the genome of theC57BL/10 strain, in which expulsion of T. spiralis is delayed (15),lacks this gene and there is no detectable expression in the gut.
Materials and MethodsMouse strains
For all experiments, with the exception of Southern blot analysis, BALB/cand C57BL/10 mice were supplied by BD Biosciences (Cowley, U.K.) and129/SvEv mice were purchased from B&K Universal (Hull, U.K.). ForSouthern blot analysis, BALB/c mice were supplied by Charles River
*Division of Veterinary Clinical Studies, University of Edinburgh, Easter Bush Vet-erinary Centre, Roslin, Midlothian, United Kingdom; †Department of Physiology,University of Cambridge, Cambridge, United Kingdom; and ‡Department of Bio-chemistry, University of Georgia, Athens, GA 30602-7229
Received for publication January 22, 2004. Accepted for publication May 13, 2004.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by a Grant from the Wellcome Trust (Award No. 060312)and from the Cystic Fibrosis Trust.2 Address correspondence and reprint requests to Dr. Alan Pemberton, Divisionof Veterinary Clinical Studies, Easter Bush Veterinary Centre, University ofEdinburgh, Roslin, EH25 9RG Midlothian, United Kingdom. E-mail address:[email protected]
The Journal of Immunology
Copyright © 2004 by The American Association of Immunologists, Inc. 0022-1767/04/$02.00
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Breeding Laboratories (Margate, U.K.); C57BL/6J and C57BL/10 mice byHarlan Olac (Bicester, U.K.); and 129S6/SvEv mice were a gift from R.Thresher (University of Cambridge, Cambridge, U.K.).
Parasite infections and tissue preparation
Maintenance, infection, and recovery of T. spiralis larvae was based onstandard methods (16). Sex-matched 8- to 15-wk-old BALB/c orC57BL/10 mice were infected by gavage with 200–300 muscle larvae permouse in 0.2 ml of PBS/0.1% agar. Larvae were freshly isolated frommuscle cysts from 30 to 90 day infected BALB/c mice. To check infectionswere successful, adult worms were isolated from groups of four to five ofthe mice at 6–7 days after infection as previously described (17). ForRT-PCR analysis, mice were killed on day 14 following infection (wormexpulsion phase) and epithelium exfoliated by EDTA perfusion as previ-ously described (18, 19). Samples were also prepared from age-matcheduninfected controls. Additional infections were set up in age- and sex-matched BALB/c mice, which were killed on days 1, 3, 7, 14, 28, and 56postinfection (p.i.)3 for epithelium isolation. Stripped epithelium was trans-ferred directly into TriReagent (Sigma-Aldrich, Poole, U.K.) for RNA iso-lation as previously described (19) or processed for protein extraction asdescribed below. Subsequent purification and removal of contaminatingDNA using DNA-free DNase (Ambion, Austin, TX) has been previouslydescribed (17). To check the purity of stripped epithelium preparations,samples were simultaneously fixed in 4% parafomaldehyde for embedding,sectioning, and H&E staining as previously described (19). Tissue samplesfor total RNA extraction were also collected from nonperfused uninfectedand infected female BALB/c mice day 14 following T. spiralis infection.Tissues collected were: stomach, duodenum, jejunum, ileum, caecum, co-lon, pancreas, kidney, spleen, liver, lung, brain, tongue, mesenteric lymphnode, heart, uterus, and skin (ear pinna). The samples were collected intoRNA-Later (Ambion) and total RNA isolated using TriReagent (Sigma-Aldrich) as previously described (17), followed by DNase treatment asearlier described.
For immunohistochemistry, age- and sex-matched BALB/c andC57BL/10 mice were infected with T. spiralis as we described and killed14 days after infection. Three-centimeter lengths of jejunum were removedstarting 10 cm from the pylorus, fixed in 4% paraformaldehyde, and pro-cessed and sectioned as previously described (20). In a second experiment,samples of jejunum were obtained from T. spiralis-infected age- and sex-matched BALB/c mice (bred at Easter Bush Veterinary Centre, Universityof Edinburgh, Edinburgh, U.K.) on days 7, 14, and 28, and also fromuninfected control mice. All mice were �15 wk of age.
Sequencing strategy
RNA from isolated epithelium from T. spiralis-infected BALB/c mice (day14) was reverse transcribed as previously described (21). Primer designwas informed by comparison of mouse intelectin-1 cDNA sequence withhuman and frog analogues, and by MALDI-TOF analysis of tryptic digestsof intelectin-2 separated by two-dimensional gel electrophoresis (2D-GE)(7), which gave five peptide fragments matching sections of the deducedamino acid sequence of mouse intelectin-1 (29–39, 51–59, 100–107, 240–245, and 246–255). cDNA was amplified using PCR primers ILNF1 andILNR2 (see following for all primer sequences). Amplifications were con-ducted for 40 s at 94°C, 40 s at 60°C, and 120 s at 72°C for 35 cycles. Theresultant single 654-bp PCR product was purified (High Pure PCR ProductPurification kit; Roche, Mannheim, Germany) and sequenced. To sequencethe 5� and 3� ends, a 5�/3� RACE kit (Boehringer Mannheim, Mannheim,U.K.) was used, following the manufacturer’s instructions. For 3� RACE,first-strand cDNA was amplified by PCR using primer ILN2P2 and the 3�dT-anchor primer, and the identity of PCR products was confirmed byhybridization with ILN2P1 as a probe. For the 5� end, RNA was reversetranscribed using ILN2P3 and avian myeloblastosis virus reverse transcrip-tase. The cDNA was purified and a 3� end homopolymeric tail was addedusing dATP and terminal transferase. The dA-tailed cDNA was amplifiedby two rounds of PCR using the 5�-dT-anchor primer and the gene-specificprimers ILN2P4 and ILN2P5. Amplifications were conducted for 15 s at94°C, 30 s at 55°C, and 60 s at 72°C for 30 cycles. PCR products werehybridized with ILNF1, and positive PCR products from both rounds ofPCR cloned and sequenced.
Resultant PCR products were visualized on ethidium bromide stained1.6% agarose gels and images recorded using Kodak Digital Science ImageStation 440CF and 1D Image Analysis software. To confirm the identity ofthe PCR products, gels were subjected to Southern blotting following stan-
dard protocols and hybridized using dioxygenin-labeled gene-specific probes,using methods previously described (20). Positive PCR products were clonedinto the TA Cloning vector pCR2.1 (Invitrogen, Leek, The Netherlands).Positive clones were identified by restriction enzyme digestion and Southernhybridizations. All nucleotide sequences were determined by dideoxy chain-termination cycle sequencing (DBS Genomics, School of Biological andBiomedical Sciences, University of Durham, Durham, U.K.). Sequence datawere analyzed with the help of web-based programs: Genestream ResourceCenter (http://www2.igh.cnrs.fr/bin/lalign-guess.cgi); Java Molecular BiologyDirectory (http://members.aol.com/_ht_a/lucatoldo/myhomepage/JaMBW);Restriction Mapper (http://www.restrictionmapper.org); National Centre forBiotechnology Information (http://www.ncbi.nlm.nih.gov); and The SequenceManipulation Site (http://www.ualberta.ca/�stothard/javascript/index.html).
Detection of transcripts by semiquantitative RT-PCR
Total RNA (1 �g) from isolated epithelium, or other tissues as alreadydescribed, was reverse transcribed using 2.5 �M (dT)15 as previously de-scribed (20). One-twentieth the volume was amplified by PCR using gene-specific primers for intelectin-1 (ILN1P6F/ILN1P7R) and intelectin-2(ILN2P6F/ILN2P7R), or for the housekeeping gene GAPDH as previouslydescribed (17) with equivalent quantities of nonreverse transcribed RNA asnegative controls. Reaction conditions were optimized to ensure the num-ber of cycles used correlated with the amplification stage of the PCR.Amplifications for intelectin-1 and intelectin-2 were conducted for 40 s at94°C, 40 s at 60°C, and 120 s at 72°C for 35 cycles (32 cycles for epithelialRNA preparations). Amplifications for GAPDH were conducted for 40 s at94°C, 40 s at 60°C, and 120s at 72°C for 35 cycles, in a final magnesiumconcentration of 1.5 mM, pH 8.3. To confirm specificity of the intelectin-1and intelectin-2 PCR products, restriction digests of 4 �l of each 50-�lPCR product were conducted with EcoR1, which only cuts the intelectin-2PCR product to give digests of 200 bp and 660 bp, and SacI, which onlycuts the intelectin-1 PCR product to give digests of 472 bp and 388 bp) (seeFig. 1). In addition, single PCR products from two day 14 epitheliumsamples amplified using ILN1P6F/ILN1P7R or ILN2P6F/ILN2P7R werepurified and sequenced as described to verify the specificity of the primers.PCR products were visualized as earlier described and hybridized with acommon probe for both intelectins, ILNR2, to detect any weak bands.
Long-template (LT)-PCR
To confirm the presence or absence of intelectin-1 and intelectin-2 se-quences in the genome of BALB/c, C57BL/10, and 129/SvEv strains ofmice, genomic DNA was prepared using a Qiagen DNeasy Tissue kit (Qia-gen, Hilden, Germany) according to the manufacturer’s instructions. LT-PCR was conducted using methods and PCR conditions previously de-scribed, but using an annealing temperature of 60°C (20). The primers usedwere ILNP6F/ILNR1 for intelectin-1, and ILN2P6F/ILN2P5 for intelec-tin-2. These primers were predicted from the intelectin-1 genomic se-quence to amplify LT-PCR products of 3185 and 3146 bp from the in-telectin-1 and intelectin-2 genes, respectively. PCR products werehybridized with ILNF1, a common probe for both genes.
Primers and probes
The sequences of the primers were the following: ILNF1 (sense) TCTCTATTTCCTGCGCACGAA; ILNR1 (antisense) GCTGACTGGACCATCGATCAC; ILNR2 (antisense) GGAAGAATCCTCCTCCACCGA; ILN2P1(sense) TACTCTCCCTCTGGCCGGAAT; ILN2P2 (sense) TGGTCTCTACCAGAAGTATCCGG; ILN2P3 (antisense) CCTTCTCCATATTTCACCGGATA; ILN2P4 (antisense) TCTGCCTGGATGTCGAAGTAG; ILN2P5(antisense) CAGGTTGTTCTCGTGCACACTA; ILN1P6F (sense) TTTATCATGGTGGCTACCAGAGG; ILN1P7R (antisense) GAGTTCCATATCCATCCCAATC; ILN2P6F (sense) TTTATCATGATTGCCACGAGAGT;ILN2P7R (antisense) GAGTTCCATATCCATTCGCATC; INTLF (sense)TGGGGCAATTCTTTCTTTTCCTCTC; INTLR (antisense) CTGCTGTACCCATTGTGAGTTCCAT. Primers and internal complementary probes weredesigned with the help of the “RawPrimer” program available onhttp://alces.med.umn.edu/rawprimer.html.
Southern blot analysis
Genomic DNA was prepared from mouse spleens, digested with HindIII,and transferred to Hybond-N� membranes (Amersham Biosciences, Pis-cataway, NJ) according to standard protocols. The blot was then hybridizedto an intelectin-1 probe, random-prime labeled (Amersham Biosciences)with [�-32P]dCTP. This probe consists of 812 bp of the intelectin-1 codingregion (NM_010584; bases 164–975 inclusive) and was generated by PCRusing the primers INTLF and INTLR. For hybridization, membranes were
3 Abbreviations used in this paper: p.i., postinfection; 2D-GE, two-dimensional gelelectrophoresis; LT-PCR, long-template PCR.
1895The Journal of Immunology
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

prehybridized at 65°C in a buffer containing 5� SSC, 5� Denhardt’s so-lution, 0.5% SDS, and 50 �g/ml sheared salmon sperm DNA (Stratagene,La Jolla, CA), labeled probe added, and hybridization continued overnight.The membranes were sequentially washed at 65°C in low (2� SSC, 0.1%SDS), medium (1� SSC, 0.1% SDS), and high (0.1� SSC, 0.1% SDS)stringency buffers as required.
Two-dimensional gel electrophoresis
Jejunal epithelium was obtained from BALB/c and C57BL/10 mice at day14 of T. spiralis infection, as previously described. To inactivate rapidlyany residual proteases, samples were extracted with 1 ml of 60 mM Tris-HCl, pH 6.8, 2% SDS, 10% glycerol, preheated to 95°C. The samples werethen heated on a boiling water bath for 5 min, followed by cooling andcentrifugation at 20,000 � g for 30 min at 15°C. Aliquots of extracts(typically 5–15 �l) were diluted in rehydration buffer (8 M urea, 2%CHAPS, 0.4% DTT, 0.2% Bio-Lytes 3-10 (Bio-Rad, Hercules, CA)) togive a final SDS concentration of �0.1% and CHAPS:SDS ratio of �20:1.Samples were then analyzed by 2D-GE as previously described (7).Briefly, first dimensional separation was with immobilized 7-cm or 17-cmpH gradient 3-10 strips (Bio-Rad) with focusing for 20,000 Vh or 45,000Vh, respectively, using a Bio-Rad Isoelectric Focusing IEF cell. Stripswere then equilibrated and run on 12% acrylamide SDS gels as previouslydescribed (7). Gels were then either stained with silver nitrate (PlusOnemethod; Amersham Biosciences) followed by imaging with a flatbed scan-ner (ImageScanner; Amersham Biosciences), or transferred to membrane(Immobilon P; Millipore, Bedford, MA) for Western blotting.
Western blotting
Following transblotting to Immobilon P membrane, blots were blockedwith 50 mM Tris-HCl, pH 7.5, 0.15M NaCl, 1% milk powder, 0.5% Tween80. Blots were then incubated for 1.5 h with either 1 �g/ml rabbit anti-XL35 Ig (13) or 1 �g/ml control rabbit IgG in blocking solution. Followingwashing, blots were then treated for 1 h with mouse monoclonal anti-rabbitIgG alkaline phosphatase conjugate (Sigma-Aldrich; 1/20,000), thenwashed and developed with 5-bromo-4-chloro-3-indolyl phosphate(BCIP)/NBT.
Immunostaining with anti-XL35 Ab
Samples of small intestine were collected from BALB/c, 129/SvEv, andC57BL/10 mice at days 0 and 14 of T. spiralis infection (see above), andfurther samples from BALB/c mice at day 7 and 28 of infection. Tissueswere fixed in 4% paraformaldehyde, processed into paraffin blocks andsections (4 �m) were made onto coated slides (Snowcoat X-tra; Surgipath,Winnipeg, Manitoba, Canada). Sections were dewaxed in xylene and en-dogenous peroxidase activity blocked by treatment with hydrogen perox-ide. After rehydration, sections were treated with 0.1% trypsin in 1 mMTris-HCl, pH 7.5, 0.1% CaCl2 at 37°C for 20 min. Slides were washed,blocked with PBS containing 0.5 M NaCl and 0.5% Tween 80, and incu-bated with anti-XL35 Ig (13)(1 �g/ml in blocking buffer) or control rabbitIgG (1 �g/ml) for 1 h at 21°C. After treatment with biotinylated donkeyanti-rabbit IgG (BA4001, 1/400; Vector Laboratories, Burlingame, CA),followed by avidin-HRP conjugate (ABC kit; Vector Laboratories) sec-tions were stained with diaminobenzidine, counterstained with hematoxy-lin, then dehydrated and mounted.
ResultsCloning and sequencing
Direct sequencing of the PCR product from primers ILNF1 andILNR2 fortuitously gave only the intelectin-2 sequence, rather thana mixture of intelectin-1 and intelectin-2 sequence as expected.This presumably reflects a higher concentration of intelectin-2mRNA in the source preparation (jejunum from day 14 T. spiralis-infected BALB/c mouse). In retrospect, the primer ILNF1 had onemismatch with respect to intelectin-2. Further sequence informa-tion was obtained by 5� and 3� RACE, as described in Materialsand Methods. The full cDNA sequence thus obtained for intelec-tin-2 is shown in Fig. 1, aligned with that previously obtained (8)for intelectin-1. The total sequence obtained for intelectin-2 was1136 nucleotides, with 941 bp of open reading frame (Fig. 1; Gen-Bank accession no. AY217760). Using primers ILN2P6F andILN2P7R, the sequence was confirmed in three other samples ofRNA from isolated epithelium (BALB/c, day 14 p.i.) and found to
be identical. This cDNA has 94% identity with that for intelec-tin-1. Analysis of the sequences shows a unique EcoR1 restrictionsite in intelectin-2, and a unique SacI site in intelectin-1 (Fig. 1).
The deduced amino acid sequence for intelectin-2 is shown inFig. 2, aligned with the sequences for mouse intelectin-1, the twohuman analogues HL-1 and HL-2, and the frog analogue XL35,with which it has 91%, 79%, 77%, and 68% amino acid identity,respectively. Regions of the intelectin-2 sequence are indicatedthat correspond to the tryptic mass fingerprinting of the corre-sponding 2D-GE spot.
Differential PCR for intelectin-1 and intelectin-2
To investigate the expression of intelectin-1 and intelectin-2 inuninfected and T. spiralis-infected jejunal epithelium, specificprimers were designed. Amplifications between forward primersILN1P6F and ILN2P6F, specific for intelectin-1 and intelectin-2,respectively, and common reverse primer ILNR2, gave products of860 bp (Fig. 3A). A single PCR product from both sets of primerswas partially sequenced to confirm their specificity for intelectin-1and intelectin-2. It was noted that although intelectin-1 was ex-pressed in both uninfected and infected epithelium, transcripts forintelectin-2 were only detected in infected samples.
To confirm the identity of all PCR products generated by theILN1P6F and ILN2P6F primers, restriction digests were con-ducted with SacI or EcoR1, which have unique cleavage sites inthe intelectin-1 and intelectin-2 sequences, respectively (Fig. 1).Restriction enzyme digests of the PCR products are shown in Fig.3A. SacI digestion produced restriction fragments of the expectedsize (472 bp and 388 bp) only from intelectin-1 PCR products,whereas EcoR1 produced the expected restriction fragments onlyfrom intelectin-2 PCR products.
Anatomical distribution of intelectins in the mouse
RNA samples from various anatomical sites were analyzed by RT-PCR for intelectin-1 and intelectin-2 (Fig. 3B). The distribution ofintelectin-1 transcripts appeared to be largely restricted to the in-testine, and in particular, no transcripts were detected in heart tis-sue, in contrast to the distribution of human intelectin (12). In-telectin-2 was even more restricted in its distribution, only beingdetected in the ileum of uninfected mice. The effect of infectionwas negligible on intelectin-1 distribution, apart from a possibleup-regulation in gastric tissues. However, intelectin-2 expressionwas markedly increased on infection, with a strong RT-PCR sig-nal being detected in small intestine and colon but not in stomach(Fig. 3B).
Intelectin-1 transcripts were abundant in uninfected jejunal ep-ithelium as detected by RT-PCR, and remained at high levelsthroughout infection with T. spiralis (Fig. 3C). Intelectin-2 tran-scripts were detected neither in uninfected jejunum nor at day 1 ofinfection, but were up-regulated by day 3 p.i. and strongly ex-pressed at days 7 and 14. Intelectin-2 transcripts were markedlyreduced at day 28, and were undetectable in samples collectedfrom day 56 p.i.
2D-GE and Western blotting
2D-GE resolved intelectin-2 in extracts of T. spiralis-infected (day14) BALB/c epithelium as a complex of 4 or 5 spots (Fig. 4A) aspreviously described (7). Western blotting of such separations withanti-XL35 Ig detected only the intelectin-2 spots (Fig. 4C), con-firming the specificity of the Ab. Western blotting with controlrabbit IgG was negative (data not shown).
1896 ENTERIC EXPRESSION OF INTELECTIN-2
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The response of C57BL/10 mice, which reject T. spiralis fromthe gut much slower than the BALB/c strain, was compared. Ex-tracts from day 14 of T. spiralis infection did not exhibit intelec-tin-2 spots by 2D-GE (Fig. 4B), and Western blotting with anti-XL35 Ig was negative (data not shown). This suggested thatintelectin-2 expression was much reduced or absent in theC57BL/10 strain on T. spiralis infection.
Immunostaining with anti-XL35 Abs
Jejunum from day 14 of T. spiralis infection in BALB/c, 129/SvEv, and C57BL/10 mice was immunostained with polyclonalrabbit anti-XL35 Ig. Immunostaining of goblet and Paneth cells
was observed in infected BALB/c and 129/SvEv mice (Figs. 5, Aand B, respectively), but not in C57BL/10 mice. Normal rabbit IgGcontrols were negative (data not shown). Anti-XL35 immunostain-ing of BALB/c jejunum from days 0, 7, 14, and 28 of T. spiralisinfection (Fig. 5, D, E, F, and G, respectively) showed that stainingwas absent in uninfected jejunum (Fig. 5D), despite the presenceof high levels of transcripts for intelectin-1. However, staining wasevident at days 7 and 14 of infection in goblet cells (Fig. 5, E andF, respectively) and Paneth cells (data not shown). Significantly,Fig. 5, E and F show immunoreactive material being released api-cally from goblet cells into the lumen. At day 28 (Fig. 5G), fol-lowing resolution of infection, there was little remaining evidence
FIGURE 1. cDNA sequence encoding mouse intelectin-2. The cDNA sequence obtained for mouse intelectin-2 (mITLN2) is aligned with the publishedsequence for mouse intelectin-1 (mITLN1) (8). Locations of sequencing primers (see Materials and Methods) are underlined. The SacI and EcoR1restriction sites are indicated, which selectively cut intelectin-1 and intelectin-2, respectively. Start and stop codons are shown in upper case.
1897The Journal of Immunology
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

of anti-XL35 immunoreactivity in goblet cells. Normal rabbit IgGcontrols from day 0 and day 14 of infection were also negative(Fig. 5, H and I, respectively). The staining pattern with anti-XL35is therefore consistent with the expression of intelectin-2, as de-tected by RT-PCR and 2D-GE, whereas intelectin-1 could not bedetected when using this Ab to immunostain sections that none-theless express high levels of intelectin-1 mRNA transcripts.
Comparison of intelectin-1 and intelectin-2 expression byBALB/c, C57BL/10, and 129 strains of mice
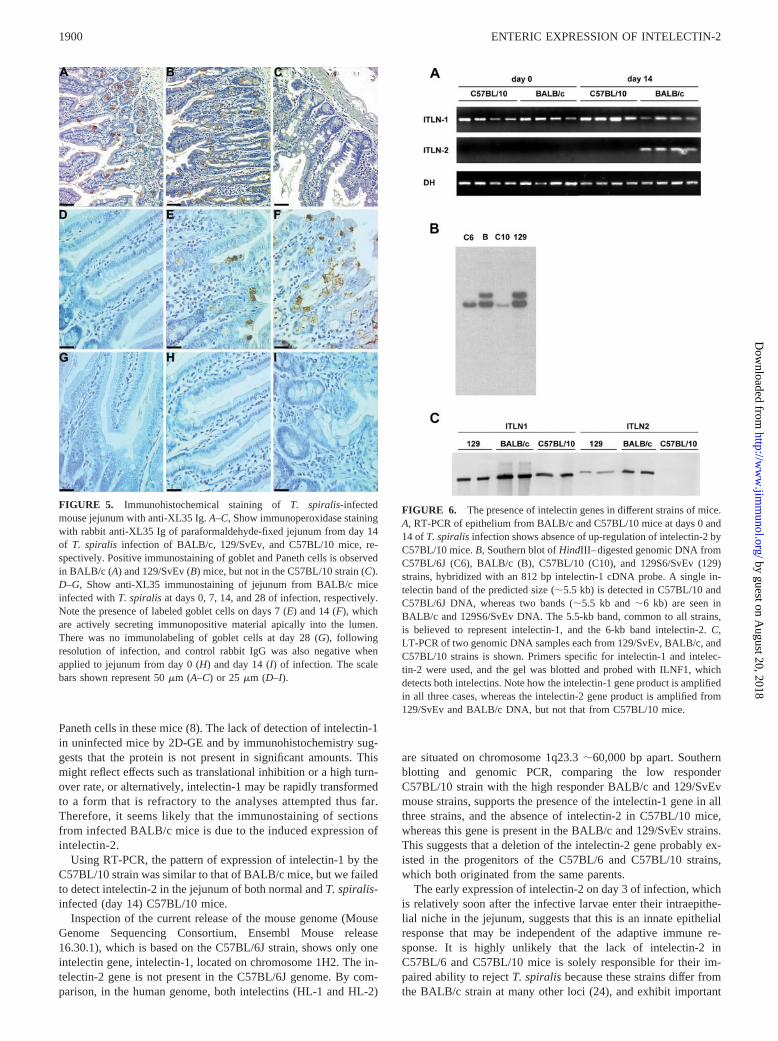
Using primers specific for intelectin-1 and intelectin-2 (see previ-ous), RT-PCR was conducted on RNA from the jejunal epitheliumof uninfected and T. spiralis-infected (day 14) C57BL/10 mice(Fig. 6A). As with BALB/c mice, intelectin-1 was strongly ex-pressed by C57BL/10 mice at both day 0 and day 14. However, nointelectin-2 transcripts were noted for C57BL/10 mice at any stageof infection, in agreement with the lack of intelectin-2 protein aspreviously shown.
The full genomic sequence of the closely related C57BL/6Jstrain (provided by the Mouse Genome Sequencing Consortium athttp://www.ensembl.org/Mus_musculus with version 16.30.01, re-lease date 06/05/03) indicated a complete absence of the intelec-tin-2 gene in this strain. The intelectin-1 gene was present, and 20kb of genomic DNA sequence encompassing the intelectin-1 gene
was downloaded for sequence analysis. This predicted that a singleband of �5.5 kb would be detected in a Southern blot of HindIII-digested DNA, using an 812-bp intelectin-1 cDNA probe.
Therefore the corresponding Southern blot was performed, com-paring the genomic organization of intelectin gene(s) in BALB/cand 129S6/SvEv strains, which respond rapidly to T. spiralis (22),with C57BL/6J and C57BL/10 strains, in which worm expulsion isdelayed (23).
Southern blot analysis indicated the presence of the expected5.5-kb restriction fragment in DNA from C57BL/10 and C57BL/6Jmice (Fig. 6B), confirming the presence of a single intelectin genein these strains. However, an additional band of �6 kb was detectedin DNA from BALB/c and 129S6/SvEv mice (Fig. 6B), suggestingthat a second intelectin gene is present in these mice. This observationis consistent with our 2D-GE (Fig. 4), immunostaining (Fig. 5), andPCR (Fig. 6A) data, which demonstrates that a second form of in-telectin is expressed in BALB/c mice but not in C57BL/10 mice. Asthe 5.5-kb band is common to all tracks in the figure, it is reasonableto suggest that it represents the intelectin-1 gene.
A further investigation using LT-PCR was conducted ongenomic DNA from C57BL/10, BALB/c, and 129/SvEv strains.The PCR primers were designed to span two exons, based on thegenomic sequence of intelectin-1. As expected, all three strainsgave the appropriate PCR product for intelectin-1 (Fig. 6C), but
FIGURE 2. Alignment of deduced amino acid sequences of intelectin analogues from mouse, man and frog. Mouse intelectin-1 and intelectin-2 arealigned with human intelectin analogues HL-1 and HL-2, and the Xenopus laevis lectin, XL35. Sections of the mouse intelectin-2 sequence confirmed bytryptic mass fingerprinting (7) are underlined.
1898 ENTERIC EXPRESSION OF INTELECTIN-2
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

the intelectin-2 product was only observed for BALB/c and 129/SvEv mice. Therefore, we conclude that the intelectin-2 gene isabsent from C57BL/10 mice, and that it is present in both BALB/cand 129/SvEv strains.
DiscussionWe describe in this report a novel epithelially derived lectin, in-telectin-2, that is exclusively expressed in the intestine in BALB/cand 129/SvEv mice. It is located in the goblet-Paneth cell popu-lation, is substantially up-regulated at the site of infection with T.spiralis, and levels of expression of intelectin-2 are maximal at thetime of worm expulsion. In our previous study (7), intelectin-2 wasfound to be the second most abundant protein, after �-actin, in2D-GE of exfoliated jejunal epithelium on day 14 or day 15 ofinfection. The fact that the protein is located in the two cell typesthat secrete apically into the gut lumen indicates that intelectin-2 islikely to be released into the gut lumen. The presence of luminalsecretion was confirmed in this study by immunohistochemistry,and considering the abundance of intelectin-2, it is likely that sub-
stantial quantities of intelectin-2 are secreted into the lumen bygoblet cells during the expulsion phase of T. spiralis infection.
Mass spectrometric analysis of tryptic digests of intelectin-2showed that it was similar to the previously described lectin, in-telectin (8). Using sequence similarities, and regions of consensussequence between other species variants of intelectin, we deter-mined the cDNA sequence for intelectin-2. This cDNA encodes a313 amino acid glycoprotein with 91% identity with mouseintelectin-1.
In uninfected BALB/c mice, mRNA for intelectin-1 was ex-pressed at high levels throughout the gut, but was not detected insignificant amounts in other tissues, including the heart, in whichhuman intelectin has been shown to be highly expressed (12).Nematode infection did not cause any apparent change in intelec-tin-1 expression in the gut. By contrast, intelectin-2 expression inuninfected mice was confined to the ileum, and was highly up-regulated during nematode infection, occurring throughout the gut.Increased message for intelectin-2 was noted in the jejunum asearly as day 3 of infection and was high at days 7 and 14, butreturned to undetectable levels at day 56, following resolution ofinfection.
The intelectin response to infection was also studied using anaffinity-purified rabbit antiserum to the frog intelectin analogue,XL35. Western blotting 2D-GE separations of exfoliated epithelialextracts of nematode-infected jejunum with the Abs detected agroup of spots at the identical location to intelectin-2. These Abswould be expected to detect both intelectin-1 and intelectin-2equally, so the fact that no further immunoreactive spots were seenin extracts from infected mice would suggest that intelectin-1 isnot present in the extracts.
The up-regulation of intelectin-2 expression as early as day 3 ofT. spiralis infection, as well as during the expulsion phase, sug-gests that this lectin may be a component of the early innate re-sponse to nematode infection. It was of interest therefore to com-pare the expression of intelectins in the low responder C57BL/10strain, which fail to expel T. spiralis efficiently (15). Comparisonof 2D-GE profiles of exfoliated jejunal epithelium from T. spiralis-infected BALB/c and C57BL/10 mice indicated an apparent lackof up-regulation of intelectin-2 in the latter strain. This was furtherconfirmed by the immunostaining of goblet cells and Paneth cellsin sections from T. spiralis-infected BALB/c but not C57BL/10mice. There was virtually no staining in uninfected mice, which isnoteworthy because mRNA for intelectin-1 is highly expressed by
FIGURE 3. Analysis of expression of intelectins-1 and intelectin-2 byRT-PCR. A, RT-PCR analysis of epithelial RNA from BALB/c mice atdays 0 and 14 of T. spiralis infection is shown. Primers were specific forintelectin-1 (ILN1P6F/ILNR2) and for intelectin-2 (ILN2P6F/ILNR2);both giving products of 860 bp. PCR products were digested with EcoR1and SacI. Intelectin-1 products were uncut by EcoR1, whereas intelectin-2products gave restriction fragments of 660 and 200bp as expected (see Fig.1). SacI restriction of intelectin-1 products gave fragments of 472 and 388bp as expected (see Fig. 1), whereas intelectin-2 products were uncut. Thisindicates the specificity of the primers. B, RT-PCR analysis with primersfor intelectin-1, intelectin-2, and GAPDH (DH) of RNA extracted fromBALB/c mice at days 0 and 14 of T. spiralis infection is shown. Tissuessamples were collected from stomach (S), duodenum (D), jejunum (J),ileum (I), caecum (Ca), colon (Co), kidney (K), spleen (Sp), liver (L), lung(Lu), brain (B), tongue (T), mesenteric lymph node (M), pancreas (P), ear(E), heart (H), and uterus (U). C, The comparative expression in BALB/cmice of intelectin-1 and intelectin-2 and GAPDH in the jejunal epitheliumby RT-PCR at days 0, 1, 3, 7, 14, 28, and 56 of T. spiralis infection.
FIGURE 4. Two-dimensional electrophoretic analysis of T. spiralis-in-fected BALB/c and C57BL/10 jejunal epithelium. Jejunal epithelium washarvested and extracted as described in Materials and Methods. Sampleswere also analyzed by 2D-GE, and the area containing intelectin-2 spots isshown. A, BALB/c epithelium from day 14 of T. spiralis infection is shownas silver stained. Note the presence of two major intelectin-2 forms, withone subsidiary form to the upper left and two further forms to the lowerright. The positions of �-actin (ACT) and �-enolase (ENOA) are alsoindicated. B, C57BL/10 epithelium from day 14 of T. spiralis infection isshown. Note the absence of intelectin-2 spots. C, Anti-XL35 Western blotof BALB/c epithelium extract from day 14 of T. spiralis infection. Notehow the pattern of anti-XL35 immunostaining corresponds to the intelec-tin-2 spots indicated in A. The presumed glycoforms of intelectin-2 span apI range from �5.2 to 5.7, and molecular mass range from �37 kDa to40 kDa.
1899The Journal of Immunology
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Paneth cells in these mice (8). The lack of detection of intelectin-1in uninfected mice by 2D-GE and by immunohistochemistry sug-gests that the protein is not present in significant amounts. Thismight reflect effects such as translational inhibition or a high turn-over rate, or alternatively, intelectin-1 may be rapidly transformedto a form that is refractory to the analyses attempted thus far.Therefore, it seems likely that the immunostaining of sectionsfrom infected BALB/c mice is due to the induced expression ofintelectin-2.
Using RT-PCR, the pattern of expression of intelectin-1 by theC57BL/10 strain was similar to that of BALB/c mice, but we failedto detect intelectin-2 in the jejunum of both normal and T. spiralis-infected (day 14) C57BL/10 mice.
Inspection of the current release of the mouse genome (MouseGenome Sequencing Consortium, Ensembl Mouse release16.30.1), which is based on the C57BL/6J strain, shows only oneintelectin gene, intelectin-1, located on chromosome 1H2. The in-telectin-2 gene is not present in the C57BL/6J genome. By com-parison, in the human genome, both intelectins (HL-1 and HL-2)
are situated on chromosome 1q23.3 �60,000 bp apart. Southernblotting and genomic PCR, comparing the low responderC57BL/10 strain with the high responder BALB/c and 129/SvEvmouse strains, supports the presence of the intelectin-1 gene in allthree strains, and the absence of intelectin-2 in C57BL/10 mice,whereas this gene is present in the BALB/c and 129/SvEv strains.This suggests that a deletion of the intelectin-2 gene probably ex-isted in the progenitors of the C57BL/6 and C57BL/10 strains,which both originated from the same parents.
The early expression of intelectin-2 on day 3 of infection, whichis relatively soon after the infective larvae enter their intraepithe-lial niche in the jejunum, suggests that this is an innate epithelialresponse that may be independent of the adaptive immune re-sponse. It is highly unlikely that the lack of intelectin-2 inC57BL/6 and C57BL/10 mice is solely responsible for their im-paired ability to reject T. spiralis because these strains differ fromthe BALB/c strain at many other loci (24), and exhibit important
FIGURE 5. Immunohistochemical staining of T. spiralis-infectedmouse jejunum with anti-XL35 Ig. A–C, Show immunoperoxidase stainingwith rabbit anti-XL35 Ig of paraformaldehyde-fixed jejunum from day 14of T. spiralis infection of BALB/c, 129/SvEv, and C57BL/10 mice, re-spectively. Positive immunostaining of goblet and Paneth cells is observedin BALB/c (A) and 129/SvEv (B) mice, but not in the C57BL/10 strain (C).D–G, Show anti-XL35 immunostaining of jejunum from BALB/c miceinfected with T. spiralis at days 0, 7, 14, and 28 of infection, respectively.Note the presence of labeled goblet cells on days 7 (E) and 14 (F), whichare actively secreting immunopositive material apically into the lumen.There was no immunolabeling of goblet cells at day 28 (G), followingresolution of infection, and control rabbit IgG was also negative whenapplied to jejunum from day 0 (H) and day 14 (I) of infection. The scalebars shown represent 50 �m (A–C) or 25 �m (D–I).
FIGURE 6. The presence of intelectin genes in different strains of mice.A, RT-PCR of epithelium from BALB/c and C57BL/10 mice at days 0 and14 of T. spiralis infection shows absence of up-regulation of intelectin-2 byC57BL/10 mice. B, Southern blot of HindIII–digested genomic DNA fromC57BL/6J (C6), BALB/c (B), C57BL/10 (C10), and 129S6/SvEv (129)strains, hybridized with an 812 bp intelectin-1 cDNA probe. A single in-telectin band of the predicted size (�5.5 kb) is detected in C57BL/10 andC57BL/6J DNA, whereas two bands (�5.5 kb and �6 kb) are seen inBALB/c and 129S6/SvEv DNA. The 5.5-kb band, common to all strains,is believed to represent intelectin-1, and the 6-kb band intelectin-2. C,LT-PCR of two genomic DNA samples each from 129/SvEv, BALB/c, andC57BL/10 strains is shown. Primers specific for intelectin-1 and intelec-tin-2 were used, and the gel was blotted and probed with ILNF1, whichdetects both intelectins. Note how the intelectin-1 gene product is amplifiedin all three cases, whereas the intelectin-2 gene product is amplified from129/SvEv and BALB/c DNA, but not that from C57BL/10 mice.
1900 ENTERIC EXPRESSION OF INTELECTIN-2
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

differences in their ability to produce regulatory cytokines, such asIL-10, in response to infections (25). However, the fact that in-telectin-2 is so markedly up-regulated in response to infection,becoming one of the most abundantly expressed proteins in in-fected epithelium (7), suggests that it may play an important rolein the innate immune response to parasite infection. The putativerole has yet to be determined, and it may indeed be multifunc-tional, including recognition of foreign carbohydrate moieties, asdescribed for human intelectin (12), and the alteration of mucinproperties, by similarity to the aggregation of the mucin-like eggjelly coat protein by the Xenopus intelectin analogue, XL35 (26).In the few immunohistochemical sections in which worms wereevident, there was no sign of anti-XL35 staining on the wormsurface (data not shown). In view of the widespread up-regulationof intelectin-2 throughout the gut, rather than just at the site ofinfection, a role for this lectin in preserving the mucosal barriermight be more likely.
Future studies should investigate the signals responsible for up-regulating intelectin-2 expression, and whether they can induceintelectin-2 expression at mucosal surfaces other than the intestinalepithelium. Importantly, the viscoelastic properties of purified mu-cins in the presence or absence of intelectin-2 should be explored.Furthermore, because there is a natural knockout of the gene inC57BL/10 mice, the effect of making these mice transgenic forgoblet cell-specific expression of intelectin-2, on the expulsion ki-netics of T. spiralis, would demonstrate its potential role in theinnate protective response against infection. It would also be im-portant to determine whether HL-1 or HL-2, the human analoguesto mouse intelectins, are regulated in a similar manner to theirmurine counterparts.
AcknowledgmentsWe thank Dr. Jeremy Brown for assistance in microscopy imaging andJudith Pate for technical assistance.
References1. Ouellette, A. J., and M. E. Selsted. 1996. Paneth cell defensins: endogenous
peptide components of intestinal host defense. FASEB J. 10:1280.2. Peeters, T., and G. Vantrappen. 1975. The Paneth cell: a source of intestinal
lysozyme. Gut 16:553.3. Kindon, H., C. Pothoulakis, L. Thim, K. Lynch-Devaney, and D. K. Podolsky.
1995. Trefoil peptide protection of intestinal epithelial barrier function: cooper-ative interaction with mucin glycoprotein. Gastroenterology 109:516.
4. Wasano, K., and Y. Hirakawa. 1997. Recombinant galectin-1 recognizes mucinand epithelial cell surface glycocalyces of gastrointestinal tract. J. Histochem.Cytochem. 45:275.
5. Miller, H. R. 1987. Gastrointestinal mucus, a medium for survival and for elim-ination of parasitic nematodes and protozoa. Parasitology 94(Suppl.):S77.
6. Dunphy, J. L., A. Balic, G. J. Barcham, A. J. Horvath, A. D. Nash, andE. N. Meeusen. 2000. Isolation and characterization of a novel inducible mam-malian galectin. J. Biol. Chem. 275:32106.
7. Pemberton, A. D., P. A. Knight, S. H. Wright, and H. R. P. Miller. 2004. Pro-teomic analysis of mouse jejunal epithelium and its response to infection with theintestinal nematode, Trichinella spiralis. Proteomics 4:1101.
8. Komiya, T., Y. Tanigawa, and S. Hirohashi. 1998. Cloning of the novel geneintelectin, which is expressed in intestinal Paneth cells in mice. Biochem. Bio-phys. Res. Commun. 251:759.
9. Roberson, M. M., and S. H. Barondes. 1982. Lectin from embryos and oocytesof Xenopus laevis: purification and properties. J. Biol. Chem. 257:7520.
10. Lee, J. K., P. Buckhaults, C. Wilkes, M. Teilhet, M. L. King, K. W. Moremen,and M. Pierce. 1997. Cloning and expression of a Xenopus laevis oocyte lectinand characterization of its mRNA levels during early development. Glycobiology7:367.
11. Nagata, S., M. Nakanishi, R. Nanba, and N. Fujita. 2003. Developmental expres-sion of XEEL, a novel molecule of the Xenopus oocyte cortical granule lectinfamily. Dev. Genes Evol. 213:368.
12. Tsuji, S., J. Uehori, M. Matsumoto, Y. Suzuki, A. Matsuhisa, K. Toyoshima, andT. Seya. 2001. Human intelectin is a novel soluble lectin that recognizes galacto-furanose in carbohydrate chains of bacterial cell wall. J. Biol. Chem. 276:23456.
13. Lee, J. K., J. Schnee, M. Pang, M. Wolfert, L. G. Baum, K. W. Moremen, andM. Pierce. 2001. Human homologs of the Xenopus oocyte cortical granule lectinXL35. Glycobiology 11:65.
14. Suzuki, Y. A., K. Shin, and B. Lonnerdal. 2001. Molecular cloning and functionalexpression of a human intestinal lactoferrin receptor. Biochemistry 40:15771.
15. Tuohy, M., D. A. Lammas, D. Wakelin, J. F. Huntley, G. F. Newlands, andH. R. Miller. 1990. Functional correlations between mucosal mast cell activityand immunity to Trichinella spiralis in high and low responder mice. ParasiteImmunol. 12:675.
16. Wakelin, D., and M. M. Wilson. 1977. Transfer of immunity to Trichinella spi-ralis in the mouse with mesenteric lymph node cells: time of appearance ofeffective cells in donors and expression of immunity in recipients. Parasitology74:215.
17. Knight, P. A., S. H. Wright, C. E. Lawrence, Y. Y. Paterson, and H. R. Miller.2000. Delayed expulsion of the nematode Trichinella spiralis in mice lacking themucosal mast cell-specific granule chymase, mouse mast cell protease-1. J. Exp.Med. 192:1849.
18. Bjerknes, M., and H. Cheng. 1981. Methods for the isolation of intact epitheliumfrom the mouse intestine. Anat. Rec. 199:565.
19. Rosbottom, A., P. A. Knight, G. McLachlan, E. M. Thornton, S. W. Wright,H. R. Miller, and C. L. Scudamore. 2002. Chemokine and cytokine expression inmurine intestinal epithelium following Nippostrongylus brasiliensis infection.Parasite Immunol. 24:67.
20. Wastling, J. M., P. Knight, J. Ure, S. Wright, E. M. Thornton, C. L. Scudamore,J. Mason, A. Smith, and H. R. Miller. 1998. Histochemical and ultrastructuralmodification of mucosal mast cell granules in parasitized mice lacking the �-chy-mase, mouse mast cell protease-1. Am. J. Pathol. 153:491.
21. Wastling, J. M., C. L. Scudamore, E. M. Thornton, G. F. Newlands, andH. R. Miller. 1997. Constitutive expression of mouse mast cell protease-1 innormal BALB/c mice and its up-regulation during intestinal nematode infection.Immunology 90:308.
22. Lawrence, C. E., J. C. Paterson, L. M. Higgins, T. T. MacDonald,M. W. Kennedy, and P. Garside. 1998. IL-4-regulated enteropathy in an intestinalnematode infection. Eur. J. Immunol. 28:2672.
23. Lammas, D. A., D. Wakelin, L. A. Mitchell, M. Tuohy, K. J. Else, andR. K. Grencis. 1992. Genetic influences upon eosinophilia and resistance in miceinfected with Trichinella spiralis. Parasitology 105(Pt. 1):117.
24. Schalkwyk, L. C., M. Jung, A. Daser, M. Weiher, J. Walter, H. Himmelbauer, andH. Lehrach. 1999. Panel of microsatellite markers for whole-genome scans andradiation hybrid mapping and a mouse family tree. Genome Res. 9:878.
25. Kaushik, R. S., J. E. Uzonna, Y. Zhang, J. R. Gordon, and H. Tabel. 2000. Innateresistance to experimental African trypanosomiasis: differences in cytokine(TNF-�, IL-6, IL-10 and IL-12) production by bone marrow-derived macro-phages from resistant and susceptible mice. Cytokine 12:1024.
26. Arranz-Plaza, E., A. S. Tracy, A. Siriwardena, J. M. Pierce, and G. J. Boons.2002. High-avidity, low-affinity multivalent interactions and the block topolyspermy in Xenopus laevis. J. Am. Chem. Soc. 124:13035.
1901The Journal of Immunology
by guest on August 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from





















![Slamf1 -/- [ BALB/c.129]](https://img.pdfslide.net/doc/110x75/56815051550346895dbe5296/slamf1-balbc129.jpg)







