Embed Size (px)
Citation preview

NeuroMolecular Medicine 179 Volume 4, 2003
Interactive Effects of Excitotoxic Injury and DietaryRestriction on Microgliosis and Neurogenesisin the Hippocampus of Adult Mice
Jaewon Lee,1 Wendy W. Auyeung,1 and Mark P. Mattson*,1,2
1Laboratory of Neurosciences, National Institute on Aging Gerontology Research Center, 5600 NathanShock Drive, Baltimore, MD 21224. 2Department of Neuroscience, Johns Hopkins University
School of Medicine, 725 N. Wolfe Street, Baltimore, MD 21205
Received June 2, 2003; Accepted July 3, 2003
Abstract
Responses to neuronal degeneration are complex, involving activation of microglia, astro-cytes, and synaptic remodeling. It has also been suggested that neuronal injury stimulates neu-rogenesis, the production of new neurons from neural stem cells. Because dietary restriction(DR) can increase hippocampal neurogenesis and promotes the survival of neurons followinginjury, we determined the effects of DR on the responses of neural stem cells, microglia, andastrocytes in the hippocampus to seizure-induced hippocampal damage. Mice on ad libitum orDR diets were given an intrahippocampal injection of kainate, administered the DNA precur-sor bromodeoxyuridine (BrdU) during a 5-d period, and euthanized 1 d or 3 wk later. Althoughkainate greatly increased the numbers of BrdU-labeled cells, it did not enhance neurogenesisand damaged neurons were not replaced. Instead, most BrdU-labeled cells were either prolif-erating microglia or neural progenitor cells that subsequently died. Microgliosis was transientand was strongly correlated with the amount of damage to CA3 neurons, whereas astrocytosiswas delayed and not correlated with neuronal loss. Surprisingly, neurogenesis was not increasedin response to seizure-induced damage, and although DR increased basal neurogenesis, it didnot promote neurogenesis following brain injury. DR significantly decreased seizure-inducedmicrogliosis, but did not affect astrocytosis. Our findings show that DR suppresses injury-induced microgliosis suggesting a contribution of a reduced microglial response to the neuro-protective effects of DR.
Index Entries: Astrocytes; apoptosis; microglia; epilepsy; seizures; stem cells; proliferation.
NeuroMolecular MedicineCopyright © 2003 Humana Press Inc.All rights of any nature whatsoever reserved.ISSN 1535-1084/03/04:179–195/$25.00
*Author to whom all correspondence and reprint requests should be addressed. E-mail: [email protected]
ORIGINAL ARTICLE

180 Lee, Auyeung, and Mattson
NeuroMolecular Medicine Volume 4, 2003
Introduction
In contrast to many other tissues that replace deadcells by nearby mitotic cells, the mammalian brainhas very little regenerative capacity, thus making itvulnerable to injury and disease. However, the braindoes contain populations of neural stem cells,located predominantly in the subventricular zoneand dentate gyrus of the hippocampus, capable ofself-renewal and of differentiation into neurons andglia (Kuhn and Svendsen, 1999). The potential ofneural stem cells to produce neurons that integrateinto functional circuits offers hope for the develop-ment of restorative therapies for ischemic, traumatic,and degenerative brain diseases (Rao and Mattson,2001; Rossi and Cattaneo, 2002). It has been reportedthat damage to the hippocampus can stimulate theproliferation of stem cells and the production of newgranule neurons from resident progenitors (Gouldand Tanapat, 1997). Seizures can induce neurogen-esis in the dentate subgranular zone (Bengzon et al.,1997), suggesting that the brain possesses a regen-erative ability. However, brain injury can also stim-ulate the proliferation of microglia and astrocytes(Bruce et al., 1996; Vezzani et al., 1999; Gebicke-Haerter, 2001), and previous studies have not clearlyestablished the phenotypes of cells proliferating inresponse to neuronal injury.
Several physiological stimuli can stimulate neu-rogenesis including environmental enrichment(Kempermann et al., 1997; Nilsson et al., 1999), phys-ical exercise (van Praag et al., 1999), and dietaryrestriction (Lee et al., 2000; Lee et al., 2002a; Matt-son et al., 2002). The signaling mechanisms thatmediate the effects of injury and physiological stim-uli on the proliferation, differentiation, and survivalof neural stem cells are beginning to be identifiedand include neurotrophic factors, such as bFGF,IGF-1, and brain-derived neurotrophic factor(BDNF) (Carro et al., 2000; Yoshimura et al., 2001;Lee et al., 2002a, 2002b; Cheng et al., 2003; Duan etal., 2003). Recent studies suggest that dietary fac-tors can have profound effects on the structure andfunction of the adult brain, and on its vulnerabilityto injury and disease. Dietary restriction (i.e.,reduced calorie intake or intermittent fasting) hasbeen known to increase the life span of many dif-ferent species, including rodents (Goodrick et al.,1983; Yu et al., 1985), and can increase the resistanceof neurons to dysfunction and death in animal
models of Alzheimer’s disease (AD) (Bruce-Kelleret al., 1999; Zhu et al., 1999), Parkinson’s disease(Duan and Mattson, 1999), and stroke (Yu and Matt-son, 1999). Studies have documented alterations inneurogenesis in animal models of AD (Haughey etal., 2002a, 2002b) and stroke (Jiang et al., 2001; Jinet al., 2001). We recently reported that DR inducesincreased hippocampal neurogenesis in adult ratsand mice (Lee et al., 2000; Lee et al., 2002). Similarto the effect of enriched environments (Kemper-mann et al., 1997; Nilsson et al., 1999), DR does notincrease the proliferation of neural stem cells, butdoes increase survival of their neuronal progeny(Lee et al., 2000; Lee et al., 2002a). It is not known ifDR affects the responses of neural stem cells and/orglial cells to brain injury. The findings described inthe present study document effects of DR on neu-rogenesis and microgliosis in a mouse model ofseizure-induced brain injury.
Materials and Methods
Mice, Diets, and Administration of Kainic Acid and BrdU
Four-week-old adult male C57BL/6 miceobtained from the National Cancer Institute weremaintained under temperature- and light-controlledconditions (20–23°C, 12 h light/12 h dark cycle).After 1 mo of adjustment time in the animal facil-ity, the mice were divided into two groups, an adlibitum (AL) group that had continual access to food,and a DR group that had access to food only everyother day. Previous studies have shown that ratsand mice maintained on such an intermittent fast-ing feeding schedule live longer and exhibitincreased resistance of neurons to injury comparedto animals fed AL(Goodrick et al., 1983; Bruce-Kelleret al., 1999; Anson et al., 2003). Three months afterdiet initiation, mice from each diet regimen werefasted overnight; anesthetized using an inhalationanesthetic (isofluorane) mixed with oxygen, andplaced in a stereotaxic head holder. The skull wasexposed along the midline and a convulsant doseof kainate (0.15 µg in 0.5 µL sterile saline, pH 7.2)was injected unilaterally into dorsal hippocampus(stereotaxic coordinates: 2.0 mm posterior tobregma, 2.4 mm lateral to bregma, and 1.4 mm belowthe surface of the brain). This method of adminis-tering kainate results in excitotoxic damage to CA3

Hippocampal Microgliosis and Neurogenesis 181
NeuroMolecular Medicine Volume 4, 2003
and CA1 neurons, but not to dentate granule neu-rons. All mice included in this study underwentseizures during the first 2 h after kainate adminis-tration. Seizure responses were rated as describedpreviously (Schauwecker, 2000). To evaluate neu-rogenesis and gliosis, mice were given a dailyintraperitoneal injection of bromodeoxyuridine(100 mg/kg body weight of BrdU) for 5 d beginning2 d after kainite injection. Half of the mice in eachdiet group were euthanized 1 d after the last BrdUinjection and the other half were euthanatized 3 wkafter the last BrdU injection. The time line for thesestudies is shown in Fig. 1A. All procedures com-plied with National Institutes of Health guidelinesand were approved by the Institutional Animal Careand Use Committee.
Immunohistochemistry
Mice were perfused transcardially with 4% para-formaldehyde and their brains were removed, post-fixed at 4°C overnight, and transferred to a 30%sucrose solution until they sank. Then brains werefrozen in isopentane and stored at –80°C. The brainswere sectioned serially at 40 µm in the coronal planeusing a freezing microtome; saving every sectioncontaining the hippocampal formation. The proto-col for immunostaining of brain sections with BrdUantibody was similar to that described previously(Lee et al., 2002a). Briefly, free-floating sections weretreated with 0.6% H2O2 in Tris-buffered saline, (TBS)pH 7.5, to block endogenous peroxidases, and DNAwas denatured by exposing sections sequentially toheat, acid, and base. The sections were incubated in0.1% TBS 5% Triton X-100 goat serum (TBS-TS) for30 min, and incubated with primary anti-BrdU anti-body (rat monoclonal; Accurate Chemicals, 1/400)in TBS-TS overnight at 4°C. Sections were furtherprocessed using a biotinylated secondary goat anti-rat IgG antibody (Vector Laboratories, 1/200),avidin-peroxidase complex, and diaminobenzidine.Stained sections were mounted onto slides andcounter-stained with cresyl violet to measure thevolume of the granule cell layer.
To determine the phenotype of BrdU-labeled cellswe performed double and triple labeling on adja-cent sections of brains as described (Lee et al., 2002).After DNA denaturation, sections were incubatedfor 1 h in a solution containing 5% normal goat
serum, and 0.1 % Triton X-100 in TBS. Primary anti-bodies were then added and the sections were incu-bated overnight at 4°C. The primary antibodiesincluded a rat monoclonal antibody against BrdU(Accurate Chemicals, 1/400 dilution), a rat mono-clonal antibody against CD11b antigen (Mac-1;Serotec, 1/1000 dilution), a rabbit polyclonal anti-body against GFAP(Sigma, 1/500 dilution), a mousemonoclonal antibody against the neuron-specificnuclear antigen NeuN (Chemicon, 1/500 dilution),a mouse monoclonal antibody against MAP2ab(Sigma, 1/500 dilution), and a mouse monoclonalantibody against BrdU (Becton Dickinson, 1/400dilution). Brain sections were then washed with TBSand incubated for 1 h in the presence of the combi-nation in anti-rat IgG labeled with AlexaFluor-488,anti-rabbit IgG labeled with AlexaFluor-633, anti-rat IgG labeled with AlexaFluor-633, anti-rabbit IgGlabeled with AlexaFluor-568, and anti-mouse IgGlabeled with AlexaFluor-568 (Molecular Probes;1/500 dilution). Confocal images were acquiredusing a Zeiss 510 CSLM microscope. Because of Mac-1 antigen sensitivity to the DNA denaturation pre-treatment, sections were incubated with Mac-1 priorto DNA denaturation, and then post-fixed with 4%paraformaldehyde for 30 min at room temperature.
Some brain sections were stained with BrdU anti-body in combination with either GFAP or Mac-1antibodies using different enzyme systems withdifferent substrates to label each antigen. Thesemethods allowed us to quantify gliogenesis usingunbiased stereological analysis. Briefly, free-floatingsections were treated with 0.6% H2O2 in TBS andthe sections were incubated in a TBS-TS (0.1% TBS5% Triton X-100 goat serum) blocking solution.The sections were incubated with antibodies againstMac-1 or GFAP in TBS-TS overnight at 4°C. Sectionswere further processed using a biotinylated sec-ondary antibody (Vector Laboratories, 1/200),avidin-alkaline phosphatase complex and substrate(VECTASTAIN® ABC-AP kit and Vector® Red Alka-line Phosphatase Substrate Kit I). Endogenous alka-line phosphatase activity was blocked by theaddition of levamisole (Sigma) to the buffer priorto the preparation of the working solution. Sectionsstained with Mac-1 or GFAP were pretreated withacid and base to denature DNA. The sections wereblocked with TBS-TS (0.1% TBS 5% Triton X-100horse serum) for 30 min, and incubated with pri-mary anti-BrdU antibody (mouse monoclonal;

182 Lee, Auyeung, and Mattson
NeuroMolecular Medicine Volume 4, 2003
Becton Dickinson, 1/400 dilution) in TBS-TSovernight at 4°C. Sections were further processedand developed using a biotinylated secondary horseanti-mouse IgG antibody (Vector Laboratories,1/200), avidin-peroxidase complex, and dia-minobenzidine. Stained sections were mounted ontoslides and cover-slipped with xylene-based mount-ing medium.
Cell Counting Methods
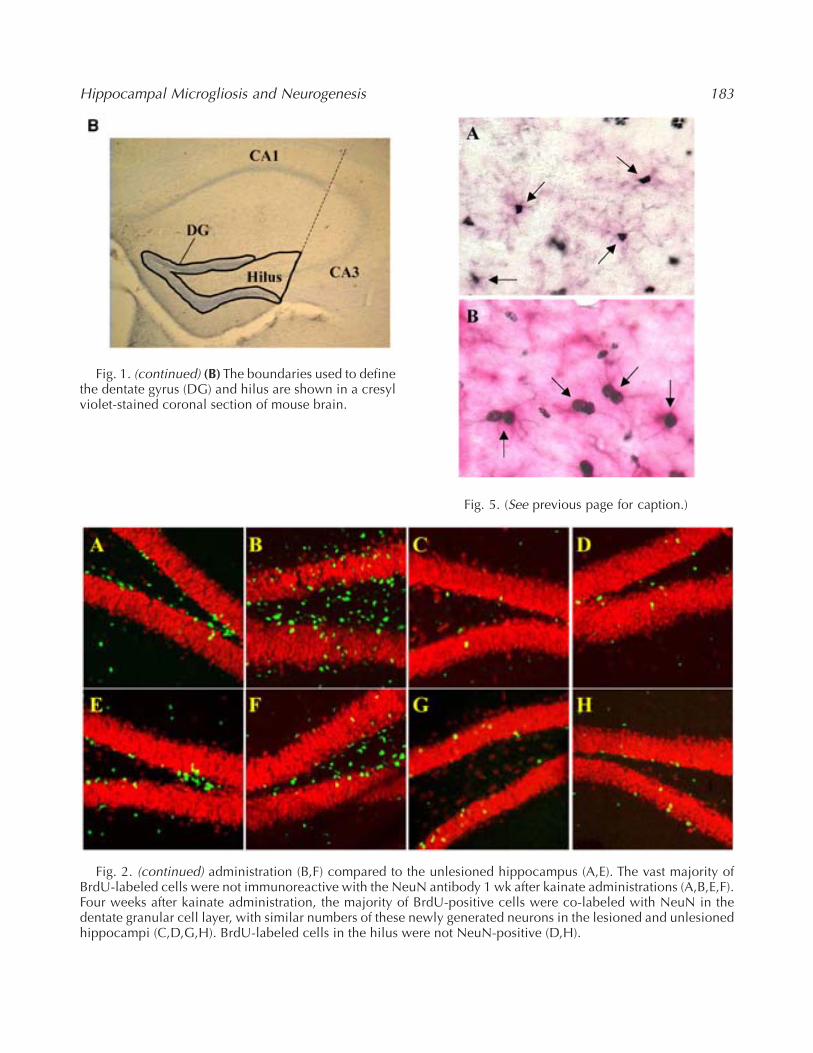
These methods are similar to those described pre-viously (Lee et al., 2002a, 2002b). The dentate gyrusand hilus were defined in accordance with themouse brain atlas of Franklin and Paxinos, (1997)as the granule cell layer excluding the molecularlayer, and the hilus was defined as the region withindentate granule cell layer extending to CA3 pyra-midal neurons (Fig. 1B). To quantify neurogenesis,BrdU and NeuN double-positive cells within thegranule cell layer of dentate gyrus were counted.
To quantify gliogenesis, BrdU-positive cells double-labeled with either GFAP or Mac-1 antibodies werecounted in the hilus. The total number of BrdU-positive cells in the dentate gyrus of each mouse wasestimated using the optical fractionator technique(West, 1993) assisted by a computer-based system,Stereologer(tm) (SPA, Alexandria, VA) using meth-ods similar to those described previously (Long etal., 1998). Estimates of region volume were assessedusing the Cavalieri method (Gundersen and Jensen,1987). Cells in every sixth section throughout theentire rostro-caudal extent of the hippocampus werecounted. For each section, the reference space wasdelineated by outlining at low power (×5 objective;on-screen magnification = ×138); identification ofBrdU-positive cells was accomplished at high power(×63 objective; on-screen magnification = ×1714).The dimension of the sampling frames was 49.2 µmin length by 49.2 µm in width and 14 µm in depth.The guard height for each section was 1 µm. Theoptical fractionator technique estimates the number
Fig. 1. Experimental design. (A) Time schedule for administration of kainate and BrdU, and sacrifice of mice.Mice were maintained on either AL or DR regimen for 3 mo, and all mice in each diet group were provided foodovernight before kainate administration (d 1). Kainate was injected unilaterally into the dorsal hippocampus (dayone) and seizure responses were monitored for 48 h (d 1,2). BrdU was administered once daily during a 5-d period.Mice were euthanized either 1 d (d 8 after kainate administration) or 3 wk (d 29 after kainate administration) afterthe last BrdU dose. (continued, opposite page, top left)
Fig. 2. (opposite page, bottom) Confocal microscopic images showing the phenotypes of newly generated cellsin the brains of mice that had been administered kainate. Brain sections from mice maintained on AL (A–D) andDR (E–H) were double-labeled with antibodies against BrdU (green) and NeuN (red: neuronal marker). The num-bers of BrdU-labeled cells in the dentate gyrus of the lesioned hippocampus were increased 1 wk after kainate
Fig. 5. (opposite page, top right) Microglia (A) and astrocytes (B) co-label with BrdU in the hilus of hippocam-pus. Enzymatic labeling methods were used to double-label cells using the BrdU antibody in combination witheither GFAP or Mac-1 antibodies. Arrows indicated BrdU-labeled microglia (A) and astrocytes (B).
Hippocampal Microgliosis and Neurogenesis 183
Fig. 1. (continued) (B) The boundaries used to definethe dentate gyrus (DG) and hilus are shown in a cresylviolet-stained coronal section of mouse brain.
Fig. 2. (continued) administration (B,F) compared to the unlesioned hippocampus (A,E). The vast majority ofBrdU-labeled cells were not immunoreactive with the NeuN antibody 1 wk after kainate administrations (A,B,E,F).Four weeks after kainate administration, the majority of BrdU-positive cells were co-labeled with NeuN in thedentate granular cell layer, with similar numbers of these newly generated neurons in the lesioned and unlesionedhippocampi (C,D,G,H). BrdU-labeled cells in the hilus were not NeuN-positive (D,H).
Fig. 5. (See previous page for caption.)

184 Lee, Auyeung, and Mattson
NeuroMolecular Medicine Volume 4, 2003
of cells by multiplying the sum of cells counted bythe reciprocal of the fraction of the region sampled.Volume densities were calculated by dividing thenumber of BrdU-positive cells counted by the totalvolume sampled of the reference space. The volumeof the sampled reference space was the number ofdissectors multiplied by the volume of one dissec-tor. Two investigators (J. L. and W. W. A.) collectedall stereological data for this study without know-ing the group identification of mice.
Assessment of Kainate-InducedHippocampal DamageThe damage in regions CA1, CA3, and hilus caused
by KA injection was determined by examination of8–10 cresyl violet-stained brain sections spacedevenly through the entire extent of the hippocam-pus. The relative amount of cell loss and damage toneurons was rated on a scale of 0 (no damage) to 5(complete cell loss). Analyses were performed bythree different blinded observers and averaged.
Statistical AnalysisCell count data were analyzed by one-way
ANOVA and post-hoc comparison of means werebased on Fisher’s protected least significant differ-ences (PLSD) procedure. Correlations among indi-vidual estimates of total BrdU-labeled cells indentate gyrus and the damage to hippocampal sub-fields were calculated by linear regression. p valuesless than 0.05 were considered statistically signifi-cant. Analyses were performed using StatView®
software.
ResultsKainate-Induced Injury Increases
Proliferation of Cells in the DentateGyrus, But Does Not IncreaseNeurogenesis: Modification by Dietary Restriction
Two-month-old mice were maintained on eitheran AL diet or an every other day feeding DR regi-men. Mice in the DR group were weighed on bothfeeding and nonfeeding days and the values plot-ted were on the feeding days for each mouse. Thebody weight and food consumption of each mousewas monitored every week until the mice were euth-
anized for analyses. The rate of increase in bodyweight of the DR mice was less than that of the ALmice. The body weights of the two groups were sig-nificantly different within 1 mo and continued todiverge until the termination of the experimentat which time their body weights differed by an aver-age of 15% (AL, 33.5 + 0.62 g; DR, 28.5 + 0.43 g;n = 12 mice per diet group; p < 0.01). Mice maintainedon the DR schedule consumed approximately 16%less food over time than did mice fed AL (AL,22.1 + 0.83 g/mouse/wk; DR, 18.9 + 0.36; n = 12 miceper diet group; p < 0.02), indicating that they weregorging on the days they did have access to food.
After three months on either ALor DR diets, micewere given a unilateral injection of kainate into thedorsal hippocampus at a dose that induced seizures.All mice exhibited seizures, and there were no sig-nificant differences in the intensity or duration ofthe seizure responses between groups AL and DR(data not shown). However, reduced mortality wasdetected in mice maintained on DR compared to ALfed mice (33% in DR group vs 47 % in AL group).Two days after kainate administration, mice weregiven five daily injections of bromodeoxyuridine(BrdU) to label newly generated cells. Mice wereeuthanized either one day or three weeks after thelast BrdU injection (corresponding to 1 wk or 4 wkafter kainate administration, respectively) andBrdU-immunoreactive cells in the dentate gyruswere quantified. At the 1-d post-BrdU time point,the number of BrdU positive cells in the dentategyrus was significantly increased in the kainate-injected (lesioned) side of hippocampus in mice inboth diet groups (Table 1). However, there was nosignificant effect of diet on the numbers of BrdU-labeled cells in either the lesioned or contralateralunlesioned hippocampus at the 1-d post-BrdU timepoint. Four weeks after kainate administration therewere significantly more BrdU-positive cells in thedentate gyrus of the unlesioned hippocampus inmice maintained on DR compared to mice fed AL(see Fig. 2 and Table 1). Although greater numbersof BrdU-labeled cells were present in the dentategyrus of the lesioned hippocampus, the survival ofthe labeled cells was significantly less than the sur-vival of labeled cells in the unlesioned hippocam-pus (Table 1). The volume of the dentate gyrus wasnot affected by kainate injection or diet (Table 1).
To determine the phenotypes of the newly gen-erated cells, we performed confocal immunohisto-

Hippocampal Microgliosis and Neurogenesis 185
NeuroMolecular Medicine Volume 4, 2003
chemical analyses of brains using an antibodyagainst NeuN, a nuclear protein expressed only inmature neurons in combination with the BrdU anti-body (Fig. 2). At the 1-d post-BrdU time point, thevast majority of BrdU-positive cells were confinedto the subgranular zone of the dentate gyrus in theunlesioned hippocampus (Fig. 2A,E). However,numerous BrdU-positive cells were presentthroughout the dentate gyrus of the lesioned hip-pocampus (Fig. 2B,F). One day after BrdU admin-istration BrdU-labeled cells in the dentate gyruswere not immunoreactive with the NeuN antibody(Fig. 2A,B,E,F). Three weeks after BrdU adminis-tration, many BrdU-positive cells were located inthe granule cell layer of dentate gyrus and wereNeuN positive, suggesting they had migrated fromthe subgranular region and differentiated into neu-rons (Fig. 2C,D,G,H). However, the numbers ofNeuN/BrdU double-positive cells were not differ-ent in the lesioned and unlesioned hippocampi, sug-gesting that, although proliferation of neural stemcells was increased in response to the kainate lesion,those newly generated cells do not differentiate intoneurons. The BrdU-labeled cells present one dayafter BrdU administration likely died during thesubsequent 3-wk period as it has been reported thatthe majority of BrdU-labeled cells in dentate gyrus
of the unlesioned hippocampus undergo apoptosisresulting in a progressive decrease in the numberof labeled cells over time (Lee et al., 2002a, 2002b).At 3 wk after BrdU administration, there were sig-nificantly more BrdU-labeled cells in the unlesioneddentate gyrus of DR mice compared to AL mice(Table 1). Although the majority of BrdU-labeledcells in the dentate granule cell layer were co-labeledwith the neuronal marker NeuN, some of the BrdU-labeled cells in the dentate gyrus did not expressNeuN; these cells were present at the highest levelsin kainate-lesioned hippocampi (Fig. 2D,H).
Kainate Induces Microgliosis and Astrocytosis in Damaged Regions of Hippocampus
To determine the effects of DR on glial cellresponses to seizure-induced neuronal injury, wefirst characterized the presence and relative num-bers of astrocytes and microglia in hippocampi ofAL-fed mice at 1- and 4-wk time points after kainateadministration. Triple-label staining of brain sec-tions was performed using antibodies against BrdU(white), MAP-2 to label neurons (green) and glial
Table 1Proliferation and Survival of Brdu-Labeled Cells in the Hippocampal Dentate Gyrus
of Mice on Ad Libitum and Dietary Restriction Feeding Regimens
AL DR
Unlesioned Lesioned Unlesioned Lesioned
Proliferation, 1 d 3351 ± 345.0 8192 ±1285.3c 2830 ± 297.2 8674 ± 1703.5c
Survival, 3 wk 1836 ± 73.9d 2695 ± 380.7c 2266 ± 131.9 b 3179 ± 588.6c
Survival (%), 3 wk 55 ± 2.2 33 ± 4.6 80 ± 4.7a 39 ± 7.2 Regional Volume1, 1d 0.202 ± 0.014 0.168 ± 0.015 0.174 ± 0.007 0.176 ± 0.014Regional Volume1, 3 wk 0.160 ± 0.016 0.153 ± 0.014 0.156 ± 0.010 0.166 ± 0.016
Note: Values for proliferation and survival are the total number of BrdU-positive cells in the dentate gyrus(see Materials and Methods section). Values for regional volume are mm3. Kainate (0.15 µg in 0.5 µL) was admin-istered into the dorsal hippocampus, then all mice received BrdU (100 mg per kg) for 5 d. Cell proliferation wasassessed on one day after the last BrdU injection, and survival of the BrdU-labeled cells was determined 3 wkafter last injection of BrdU (n = 5 to 8 mice per group). Values are the mean ± SEM.
a,bSignificantly different from AL group (a: p < 0.01; b: p < 0.05).cSignificantly different from the unlesioned side (p < 0.01).dSignificantly different from the 1-d time point (p < 0.01).

186 Lee, Auyeung, and Mattson
NeuroMolecular Medicine Volume 4, 2003
fibrillary acidic protein (GFAP) to label astrocytes(red) (Fig. 3A-H).
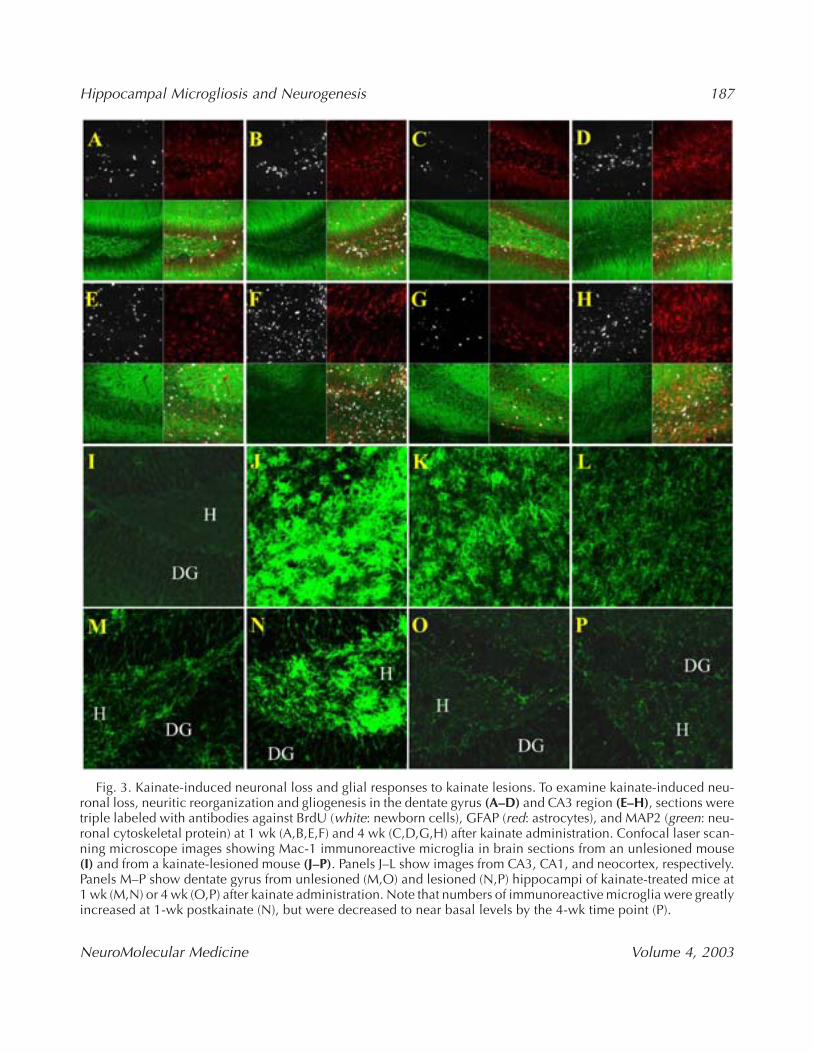
One week after kainate administration, MAP-2immunoreactivity was dramatically reduced in theCA3 region and the hilus of lesioned hippocampicompared to unlesioned hippocampi (compare panelsB and F with panels A and E in Fig. 3). Activation ofastrocytes occurred in both the lesioned and unle-sioned hippocampus (Fig. 3A,B,E,F). Four weeks afterkainate administration levels of GFAP and MAP-2immunoreactivities were increased in the damagedCA3 and hilar regions of the lesioned hippocampicompared to the 1-wk postkainate time point(Fig. 3D,H). At the 4-wk postkainate time point, themajority of BrdU-labeled cells in the damagedregions of the hippocampus were GFAP-positive(Fig. 3D,H). Although MAP-2 immunoreactivity wasincreased in the hilus and CA3 region at the 4-wkpostkainate time point, we did not detect any cellsdouble-labeled with BrdU and NeuN antibodies, sug-gesting that the increase in MAP-2 immunoreactivityresulted from growth/reorganization of dendrites ofadjacent surviving neurons.
To determine the responses of microglia tokainate-induced seizures and neuronal damage, weemployed an antibody (Mac-1) to complementreceptor-3 (CR3), that is expressed in microglia, butnot in other cell types in the brain (Jensen et al.,1997). Mac-1-positive cells were rare in the hip-pocampus of control mice that had not been injectedwith kainate (Fig. 3I), but were numerous in thelesioned, and to a lesser extent in the unlesioned,hippocampus of mice killed 1 wk after kainateadministration (Fig. 3J–P). Four weeks after kainateadministration the numbers of Mac-1-positive cellshad decreased dramatically in both the lesioned andunlesioned hippocampi compared to the 1-wk postkainate time point (Fig. 3O,P). Evaluation of themorphologies of the microglia, according to the clas-sification of Grassi-Zucconi et al. (2002) revealedthat, at 1 wk after kainate administration, hyper-trophic microglia and bushy cells (that are charac-terized by markedly enlarged soma and short, thickprocesses) were concentrated in the regions (CA3,hilus and CA1) that are the most sensitive to seizure-induced damage (Fig. 3I,K,N). Rod-shaped cells andamoeboid-phagocytic cells were more common inthe unlesioned neocortex and hippocampi (Fig.3L,M). Four weeks after kainate administration themajority of microglial cells exhibited a ramified rest-
ing morphology in both the lesioned and unlesionedhippocampi (Fig. 3O,P).
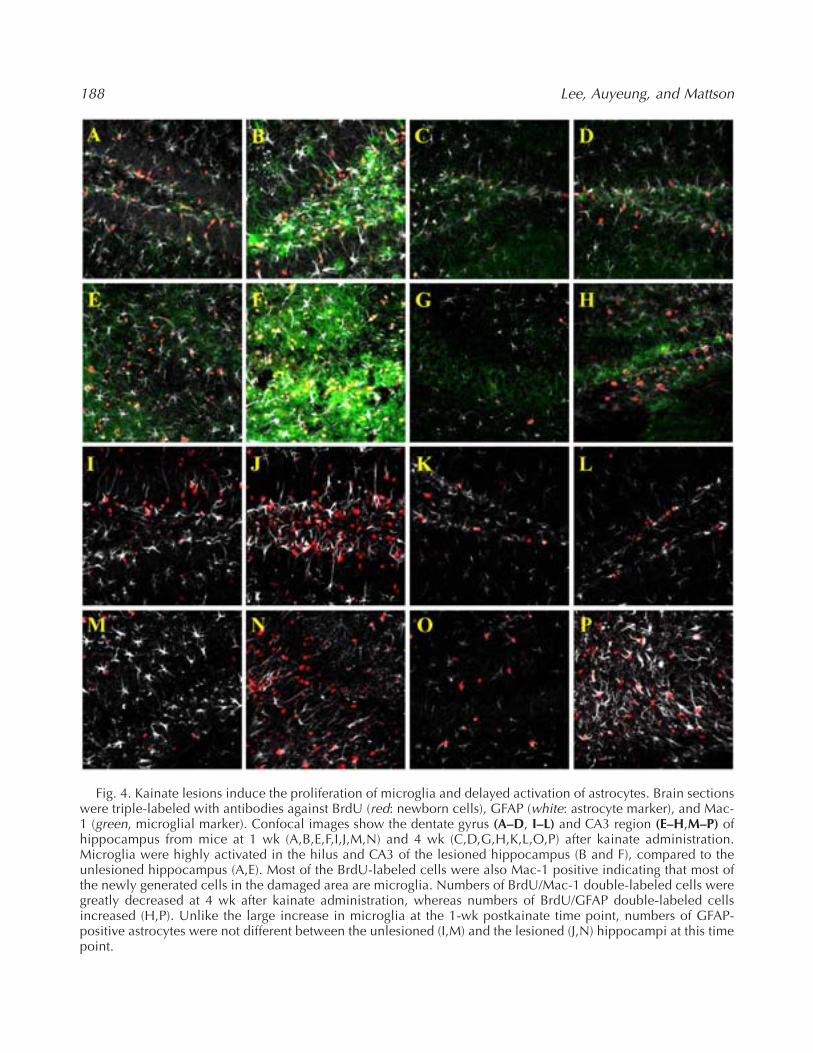
Because the numbers of microglia were increasedat the 1-wk time point and decreased at the 4-wktime point, which was similar to the numbers ofBrdU-labeled cells at these two time points, wetriple-labeled brain sections with BrdU, Mac-1, andGFAP antibodies (Fig. 4). Because the Mac1 antigenis vulnerable to the acid-base pretreatment requiredto denature DNA for BrdU immunohistochemistry,sections were immunostained with Mac-1 first. Atthe 1-wk postkainate time point, the majority ofBrdU-positive cells in the lesioned hippocampiwere Mac-1-positive (Fig. 4B,F). A few BrdU/Mac1double-positive cells were also present in the unle-sioned hippocampi at the 1-wk time point in bothlesioned and unlesioned hippocampi at the 4-wktime point (Fig. 4A,C–E,G,H).
GFAP-positive cells were present in boththe lesioned and unlesioned hippocampi at the1-wk postkainate time point; and were more numer-ous than in control mice not administered kainate(Fig. 4I,J,M,N). Four weeks after kainate adminis-tration, numbers of GFAP-positive cells remainedincreased in both the lesioned and unlesioned hip-pocampi; at this time point most of the BrdU-labeledcells in the injured regions of hippocampus werealso GFAP positive (Fig. 4P).
Dietary Restriction DecreasesKainate-Induced MicrogliosisBut Does Not Affect Astrocytosis
The numbers of newly generated microglia andastrocytes were quantified by counting BrdU-labeled cells that were also either Mac1- or GFAP-positive using unbiased stereological methods. Asanticipated, the nuclei of Mac1-positive microglia(Fig. 5A) were smaller than the nuclei of GFAP-positive astrocytes (Fig. 5B). One day after BrdUadministration (1 wk after kainate administration),there was a significant increase in the number ofBrdU positive microglial cells in the hilus of lesionedhippocampi compared to the unlesioned hip-pocampi in both AL and DR mice. However, thenumbers of BrdU-labeled microglia in the hilus ofthe lesioned hippocampi from mice maintained onDR was significantly less than that of mice on theAL diet (Table 2). Three weeks after BrdU injection
Hippocampal Microgliosis and Neurogenesis 187
NeuroMolecular Medicine Volume 4, 2003
Fig. 3. Kainate-induced neuronal loss and glial responses to kainate lesions. To examine kainate-induced neu-ronal loss, neuritic reorganization and gliogenesis in the dentate gyrus (A–D) and CA3 region (E–H), sections weretriple labeled with antibodies against BrdU (white: newborn cells), GFAP (red: astrocytes), and MAP2 (green: neu-ronal cytoskeletal protein) at 1 wk (A,B,E,F) and 4 wk (C,D,G,H) after kainate administration. Confocal laser scan-ning microscope images showing Mac-1 immunoreactive microglia in brain sections from an unlesioned mouse(I) and from a kainate-lesioned mouse (J–P). Panels J–L show images from CA3, CA1, and neocortex, respectively.Panels M–P show dentate gyrus from unlesioned (M,O) and lesioned (N,P) hippocampi of kainate-treated mice at1 wk (M,N) or 4 wk (O,P) after kainate administration. Note that numbers of immunoreactive microglia were greatlyincreased at 1-wk postkainate (N), but were decreased to near basal levels by the 4-wk time point (P).
188 Lee, Auyeung, and Mattson
Fig. 4. Kainate lesions induce the proliferation of microglia and delayed activation of astrocytes. Brain sectionswere triple-labeled with antibodies against BrdU (red: newborn cells), GFAP (white: astrocyte marker), and Mac-1 (green, microglial marker). Confocal images show the dentate gyrus (A–D, I–L) and CA3 region (E–H,M–P) ofhippocampus from mice at 1 wk (A,B,E,F,I,J,M,N) and 4 wk (C,D,G,H,K,L,O,P) after kainate administration.Microglia were highly activated in the hilus and CA3 of the lesioned hippocampus (B and F), compared to theunlesioned hippocampus (A,E). Most of the BrdU-labeled cells were also Mac-1 positive indicating that most ofthe newly generated cells in the damaged area are microglia. Numbers of BrdU/Mac-1 double-labeled cells weregreatly decreased at 4 wk after kainate administration, whereas numbers of BrdU/GFAP double-labeled cellsincreased (H,P). Unlike the large increase in microglia at the 1-wk postkainate time point, numbers of GFAP-positive astrocytes were not different between the unlesioned (I,M) and the lesioned (J,N) hippocampi at this timepoint.

Hippocampal Microgliosis and Neurogenesis 189
NeuroMolecular Medicine Volume 4, 2003
(4 wk after kainate administration), significantlyfewer BrdU-positive microglia were present in thehilus of both the lesioned an unlesioned hippocampicompared to the 1-d post-BrdU time point; this wastrue in both AL and DR mice. There was no signif-icant difference in the numbers of BrdU and Mac-1double-positive cells at the 3-wk post-BrdU timepoint in DR mice compared to AL mice (Table 2).
At the 1-d post-BrdU time point, there was not asignificant increase in the numbers of BrdU-positive astrocytes in the hilus of lesionedhippocampi compared to the contralateral hilus.However, the numbers of GFAP-positive astrocyteswere significantly increased in the hilus of thelesioned hippocampi from mice maintained oneither AL or DR diets at the 4-wk postkainate timepoint (Table 2). There was no significant effect ofdiet on the number of newly generated astrocytes(Table 2). The delayed appearance of BrdU-positiveastrocytes suggested that many of the BrdU-labeledcells present during the early postkainate timeperiod subsequently migrated into the damagedarea of hippocampus and differentiated into GFAP-positive astrocytes. In two separate experiments,the volume of the hilus was not significantly affected
by diet regimen, kainate administration, or time ofsacrifice (data not shown).
Regression Analysis of Newly Generated Cellsand Hippocampal Damage
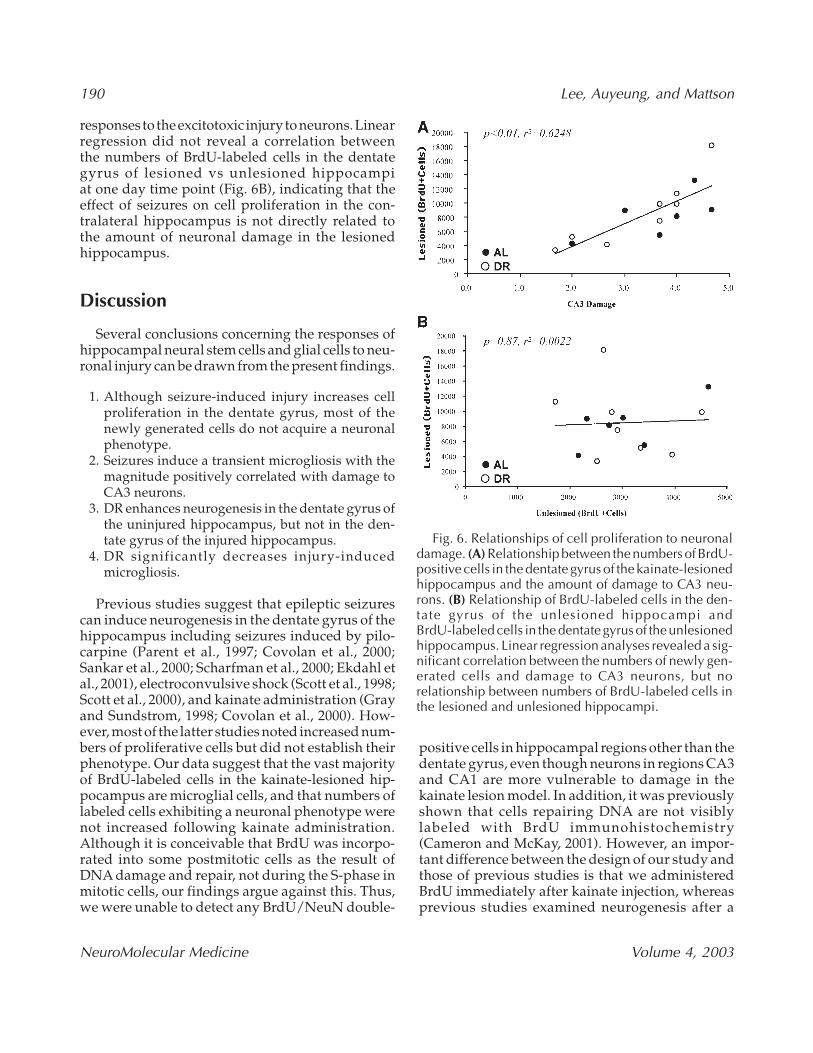
Linear regression analyses was used to determineif there was a correlation between the numbers ofBrdU-labeled cells in the dentate gyrus and theextent of hippocampal damage. There was a sig-nificant positive correlation between the numbersof newly generated cells in the dentate gyrus oflesioned hippocampi at 1 d after the last BrdUadministration and the relative amount of damageto CA3 neurons (Fig. 6A). Microglial cells are respon-sive to kainate-induced hippocampal damage, andtheir activation has been correlated with amount ofneuronal death (Andersson et al., 1991). Therefore,the positive correlation between the numbers ofBrdU-labeled cells in the dentate gyrus and thedamage to CA3 neurons suggests that increased pro-liferation of neural stem cells and microglia are
Table 2Effects of Kainate and Dietary Restriction on Numbers of Microglia
and Astrocytes in the Hilar Region of the Hippocampus
AL DR
Unlesioned Lesioned Unlesioned Lesioned
Microglia, 1 d 3074 ± 476.2 12357 ± 1495.8b 2217 ± 328.5 8595 ± 1655.6a,b
Microglia 3 wk 1232 ± 237.0c 3276 ± 816.8c 1035 ± 103.1d 2949 ± 902.4c
Astrocytes, 1d 908 ± 91.5 1257 ± 241.7 756 ± 100.8 961 ± 135.5Astrocytes, 3 wk 787 ± 94.0 1537 ± 212.9b 807 ± 201.1 1592 ± 187.1b,c
Note: Kainate (0.15 µg in 0.5µL) was injected into the dorsal hippocampus, and then all mice received BrdU(100 mg/kg) for 5 d. Kainate-induced gliogenesis in the hilus region was assessed on 1-d and 3 wk after lastBrdU injection. BrdU-positive cells that co-labeled with either Mac-1 (microglial marker) or GFAP (astrocytemarker) were quantified in the hilus regions of hippocampus (n = four to five mice per group). Values are themean ± SEM
aSignificantly different from the corresponding AL value (p < 0.01).bSignificantly different from the unlesioned side (p < 0.01).cSignificantly different from the 1-d time point (p < 0.01).dSignificantly different from the 1-d time point (p < 0.05).
190 Lee, Auyeung, and Mattson
NeuroMolecular Medicine Volume 4, 2003
responses to the excitotoxic injury to neurons. Linearregression did not reveal a correlation betweenthe numbers of BrdU-labeled cells in the dentategyrus of lesioned vs unlesioned hippocampiat one day time point (Fig. 6B), indicating that theeffect of seizures on cell proliferation in the con-tralateral hippocampus is not directly related tothe amount of neuronal damage in the lesionedhippocampus.
Discussion
Several conclusions concerning the responses ofhippocampal neural stem cells and glial cells to neu-ronal injury can be drawn from the present findings.
1. Although seizure-induced injury increases cellproliferation in the dentate gyrus, most of thenewly generated cells do not acquire a neuronalphenotype.
2. Seizures induce a transient microgliosis with themagnitude positively correlated with damage toCA3 neurons.
3. DR enhances neurogenesis in the dentate gyrus ofthe uninjured hippocampus, but not in the den-tate gyrus of the injured hippocampus.
4. DR significantly decreases injury-inducedmicrogliosis.
Previous studies suggest that epileptic seizurescan induce neurogenesis in the dentate gyrus of thehippocampus including seizures induced by pilo-carpine (Parent et al., 1997; Covolan et al., 2000;Sankar et al., 2000; Scharfman et al., 2000; Ekdahl etal., 2001), electroconvulsive shock (Scott et al., 1998;Scott et al., 2000), and kainate administration (Grayand Sundstrom, 1998; Covolan et al., 2000). How-ever, most of the latter studies noted increased num-bers of proliferative cells but did not establish theirphenotype. Our data suggest that the vast majorityof BrdU-labeled cells in the kainate-lesioned hip-pocampus are microglial cells, and that numbers oflabeled cells exhibiting a neuronal phenotype werenot increased following kainate administration.Although it is conceivable that BrdU was incorpo-rated into some postmitotic cells as the result ofDNA damage and repair, not during the S-phase inmitotic cells, our findings argue against this. Thus,we were unable to detect any BrdU/NeuN double-
positive cells in hippocampal regions other than thedentate gyrus, even though neurons in regions CA3and CA1 are more vulnerable to damage in thekainate lesion model. In addition, it was previouslyshown that cells repairing DNA are not visiblylabeled with BrdU immunohistochemistry(Cameron and McKay, 2001). However, an impor-tant difference between the design of our study andthose of previous studies is that we administeredBrdU immediately after kainate injection, whereasprevious studies examined neurogenesis after a
Fig. 6. Relationships of cell proliferation to neuronaldamage. (A) Relationship between the numbers of BrdU-positive cells in the dentate gyrus of the kainate-lesionedhippocampus and the amount of damage to CA3 neu-rons. (B) Relationship of BrdU-labeled cells in the den-tate gyrus of the unlesioned hippocampi andBrdU-labeled cells in the dentate gyrus of the unlesionedhippocampus. Linear regression analyses revealed a sig-nificant correlation between the numbers of newly gen-erated cells and damage to CA3 neurons, but norelationship between numbers of BrdU-labeled cells inthe lesioned and unlesioned hippocampi.

Hippocampal Microgliosis and Neurogenesis 191
NeuroMolecular Medicine Volume 4, 2003
delay of 4–5 d. It therefore remains to be determinedif neurogenesis increases at later time pointsafter kainate administration and if this is modifiedby DR.
It has been suggested that increased proliferationof neural stem cells in response to injury is an adap-tive response that may result in the replacement ofdamaged or dead neurons (Peterson, 2002). Ourresults confirm previous studies showing that theproliferation of cells in the dentate gyrus increasesin response to kainate-induced seizures (Gray andSundstrom, 1998; Covolan et al., 2000), but suggestthat very few if any of the newly generated cells dif-ferentiate into neurons. On the contrary, a compari-son of numbers of BrdU-labeled cells present in thedentate gyrus at 1 wk vs 4 wk after kainate admin-istration suggests that a higher percentage of thenewly generated cells died in the lesioned hip-pocampus compared to the nonlesioned hippocam-pus. The cells that did survive appeared to becomeprimarily astrocytes. There was no significant dif-ference in the numbers of NeuN/BrdU double-positive cells in the dentate gyrus of lesioned andunlesioned hippocampi of mice. Moreover, we wereunable to detect NeuN/BrdU double-positive cellsin other regions of the hippocampus, including thedamaged CA3 region. Therefore, the injury-inducedenhancement of neural stem cell proliferation doesnot result in replacement of damaged neurons, atleast not in this excitotoxic lesion model.
The enhanced death of newly generated cells inthe lesioned hippocampus suggests that the exci-totoxic insult not only promotes death of CA3 neu-rons, but may also enhance the death of thedifferentiating progeny of neural stem cells. How-ever, Parent et al. (1997) suggested that prolongedseizures stimulate dentate granule cell neurogene-sis, and that hippocampal network plasticity asso-ciated with epileptogenesis may arise from aberrantconnections formed by newly born dentate granulecells. In the latter study, they showed that prolifer-ative activity in the dentate gyrus was significantlyincreased at 3 d, remained elevated at 6 and 13 d,and returned to baseline levels by 27 d after pilo-carpine treatment. Therefore, there is still a possi-bility of neurogenesis that can replace the damagedneurons by new neurons generated at later timepoints than was examined in the present study. Weobserved a recovery of MAP2 immunoreactivity inthe absence of BrdU immunoreactivity in the dam-
aged hilus (Fig. 3D) and CA3 (Fig. 3H). However,this recovery of MAP2 immunoreactivity was notthe result of generation of new neurons in theseregions; instead, it was apparently the result ofgrowth or reorganization of dendrites of survivingneurons in these regions.
Although previous studies have reported theenhanced cell proliferation (BrdU-labeling) andincreased numbers of microglia, in response toseizure-induced brain injury, it has not been estab-lish whether the microglia were newly-generatedcells. Our analysis of brain sections double-labeledwith BrdU with Mac-1 antibodies revealed that themajority of BrdU-positive cells in lesioned hippo-campus during the first week after kainate admin-istration are microglia. The microglial proliferationwas most prevalent in the molecular layers of thelesioned regions of hippocampus (CA3 and hilus),but there were also BrdU-labeled microglia in thedentate gyrus. Kainate-induced microgliosis wastransient, and numbers of BrdU/Mac-1 double-positive cells were decreased at the 4 wk post-kainate time point. This decrease may be the resultof migration of microglia away from the injury sites,or death of the microglia. The positive correlationbetween numbers of newly generated cells in thedentate gyrus and the amount of damage to CA3pyramidal cells, suggests that the proliferation ofthese cells is triggered by a damage-related signalrather than by a direct effect of kainate on the pro-liferative cells. In contrast to microglia, we foundthat numbers of newly generated astrocytes (BrdU/GFAP double-positive) remained elevated in thelesioned hippocampus through the 4 wk post-kainate time point. Numbers of BrdU/GFAPdouble-positive cells in the hilus were increased betweenthe 1-wk and 4-wk postkainate time points sug-gesting some of these cells may have arisen fromneural stem cells that differentiated into astrocytesduring the 3-wk time interval. Interestingly, Songet al. (2002) recently suggested that adult astrocytesfrom hippocampus are capable of regulating neu-rogenesis by instructing the stem cells to adopt aneuronal fate. Additionally, astrocytes may induceand stabilize CNS synapses and integrate neuronalinputs and modulate synaptic activity (Haydon,2001; Ullian et al., 2001). Therefore, newly differen-tiated astrocytes might provide both supportivesynaptogenesis and active regenerative neurogen-esis in the damaged area of hippocampus. It was

192 Lee, Auyeung, and Mattson
NeuroMolecular Medicine Volume 4, 2003
previously suggested that DR enhances neurogen-esis by increasing BDNF production (Lee et al., 2000;Lee et al., 2002a, 2002b) and, indeed, BDNF has beenshown to promote the differentiation and survivalof newly generated neurons in the adult rodent brain(Benraiss et al., 2001). In addition to BDNF, kainateinduces the production of tumor necrosis factor-α,interleukins 1 and 6, transforming growth factor-β,and basic fibroblast growth factor (Morgan et al.,1993; Bruce et al., 1996; Vezzani et al., 1999;Yoshimura et al., 2001). One or more of these factorsmay contribute to the lack of effect of DR on neu-rogenesis in the lesioned hippocampus.
DR can increase the resistance of neurons to exci-totoxic, metabolic, and oxidative insults (Bruce-Keller et al., 1999; Duan and Mattson, 1999; Yu andMattson, 1999). Neuroprotection may be mediatedby increased production of brain-derived neu-rotrophic factor (Duan et al., 2001a; Duan et al.,2001b) and protein chaperones (Lee et al., 1999; Guoet al., 2000). However, data in the present study sug-gest that DR might also protect neurons by reduc-ing activation of microglia. Activation andproliferation of microglia in response to brain injuryprovides a mechanism for removing damaged neu-rons, but may also contribute to the neurodegener-ative process (Gebicke-Haerter, 2001). We found thatthe number of newly generated microglia was sig-nificantly decreased in the kainate-lesioned hip-pocampi of mice that had been maintained on a DRfeeding regimen. This result provides the first evi-dence that diet can affect microglial responses toinjury to the nervous system, a finding of potentialclinical relevance for various disorders that involvemicroglial activation (Hays, 1998; Lee et al., 2002c).It has been suggested that adult hippocampal neu-rogenesis may play an important role in learningand memory processes (Feng et al., 2001; Shors etal., 2001). However, neurogenesis may not repopu-late the large numbers of neurons lost as the resultof acute injuries or in age-related neurodegenera-tive disorders. Although DR can enhance hip-pocampal neurogenesis in adult rats and mice (Leeet al., 2000; Lee et al., 2002a, 2002b), we did notobserve any effect of DR on the proliferation or sur-vival of BrdU-labeled cells in the dentate gyrus ofthe kainate lesioned hippocampus. In the unle-sioned hippocampus, DR did not affect cell prolif-eration in the dentate gyrus but did cause asignificant increase in the number of surviving
BrdU-labeled cells and NeuN/BrdU double-labeledcells indicating that neurogenesis was increased.Previous studies showed that DR could reduce neu-ronal damage and improve function outcome inanimal models of stroke (Yu and Mattson, 1999),Alzheimer’s disease (Bruce-Keller et al., 1999; Zhuet al., 1999), and Parkinson’s disease (Duan andMattson, 1999). The ability of DR to enhance basalneurogenesis and reduce microgliosis, suggests thatDR may be a valuable approach for reducing dis-ease risk and improving functional outcome in var-ious neurodegenerative conditions.
References
Andersson P. B., Perry V. H., and Gordon S. (1991) TheCNS acute inflammatory response to excitotoxicneuronal cell death. Immunol. Lett. 30, 177–181.
Anson R. M., Guo Z., de Cabo R., et al. (2003) Inter-mittent fasting dissociates beneficial effects ofdietary restriction on glucose metabolism and neu-ronal resistance to injury from calorie intake. Proc.Natl. Acad. Sci. USA 100, 6216–6220.
Bengzon J., Kokaia Z., Elmer E., Nanobashvili A.,Kokaia M., and Lindvall O. (1997) Apoptosis andproliferation of dentate gyrus neurons after singleand intermittent limbic seizures. Proc. Natl. Acad.Sci. USA 94, 10432–10437.
Benraiss A., Chmielnicki E., Lerner K., Roh D., andGoldman S. A. (2001) Adenoviral brain-derivedneurotrophic factor induces both neostriatal andolfactory neuronal recruitment from endogenousprogenitor cells in the adult forebrain. J. Neurosci.21, 6718–6731.
Bruce A. J., Boling W., Kindy M. S., et al. (1996) Alteredneuronal and microglial responses to excitotoxicand ischemic brain injury in mice lacking TNFreceptors. Nat. Med. 2, 788–794.
Bruce-Keller A. J., Umberger G., McFall R., and Matt-son M. P. (1999) Food restriction reduces braindamage and improves behavioral outcome fol-lowing excitotoxic and metabolic insults. Ann.Neurol. 45, 8–15.
Cameron H. A. and McKay R. D. (2001) Adult neuro-genesis produces a large pool of new granule cellsin the dentate gyrus. J. Comp. Neurol. 435, 406–417.
Carro E., Nunez A., Busiguina S., and Torres-Aleman I.(2000) Circulating insulin-like growth factor Imediates effects of exercise on the brain. J. Neu-rosci. 20, 2926–2933.

Hippocampal Microgliosis and Neurogenesis 193
NeuroMolecular Medicine Volume 4, 2003
Cheng A., Wang S., Cai J., Rao M. S., and Mattson M. P.(2003) Nitric oxide acts in a positive feedback loopwith BDNF to regulate neural progenitor cell pro-liferation and differentiation in the mammalianbrain. Dev. Biol. 258, 319–333.
Covolan L., Ribeiro L. T., Longo B. M., and MelloL. E. (2000) Cell damage and neurogenesis inthe dentate granule cell layer of adult rats afterpilocarpine- or kainate-induced status epilepticus.Hippocampus 10, 169–180.
Duan W. and Mattson M. P. (1999) Dietary restrictionand 2-deoxyglucose administration improvebehavioral outcome and reduce degeneration ofdopaminergic neurons in models of Parkinson’sdisease. J. Neurosci. Res. 57, 195–206.
Duan W., Guo Z., and Mattson M. P. (2001a) Brain-derived neurotrophic factor mediates an excito-protective effect of dietary restriction in mice.J. Neurochem. 76, 619–626.
Duan W., Lee J., Guo Z., and Mattson M. P. (2001b)Dietary restriction stimulates BDNF productionin the brain and thereby protects neurons againstexcitotoxic injury. J. Mol. Neurosci. 16, 1–12.
Duan W., Guo Z., Jiang H., Ware M., Li X. J., and Matt-son M. P. (2003) Dietary restriction normalizesglucose metabolism and BDNF levels, slows diseaseprogression, and increases survival in huntingtinmutant mice. Proc. Natl. Acad. Sci. USA100, 2911–2916.
Ekdahl C. T., Mohapel P., Elmer E., and Lindvall O.(2001) Caspase inhibitors increase short-term sur-vival of progenitor-cell progeny in the adult ratdentate gyrus following status epilepticus. Eur.J. Neurosci.14, 937–945.
Feng R., Rampon C., Tang Y. P. et al. (2001) Deficientneurogenesis in forebrain-specific presenilin-1knockout mice is associated with reduced clear-ance of hippocampal memory traces. Neuron 32,911–926.
Gebicke-Haerter P. J. (2001) Microglia in neurodegen-eration: molecular aspects. Microsc. Res. Tech. 54,47–58.
Goodrick C. L., Ingram D. K., Reynolds M. A., Free-man J. R., and Cider N. L. (1983) Effects of inter-mittent feeding upon growth, activity, and lifespanin rats allowed voluntary exercise. Exp. Aging Res.9, 203–209.
Gould E. and Tanapat P. (1997) Lesion-induced pro-liferation of neuronal progenitors in the dentategyrus of the adult rat. Neuroscience 80, 427–436.
Grassi Zucconi G., Semprevivo M., Laurenzi M. A.,and Giuditta A. (2002) Sleep impairment bydiethyldithiocarbamate in rat. Protective effects of
pre-conditioning and antioxidants. Brain Res. 939,87–94.
Gray W. P. and Sundstrom L. E. (1998) Kainic acidincreases the proliferation of granule cell progen-itors in the dentate gyrus of the adult rat. BrainRes. 790, 52–59.
Gundersen H. J. and Jensen E. B. (1987) The efficiencyof systematic sampling in stereology and its pre-diction. J. Microsc. 147, 229–263.
Guo Z., Ersoz A., Butterfield D.A., and Mattson M. P.(2000) Beneficial effects of dietary restriction oncerebral cortical synaptic terminals: preservationof glucose and glutamate transport and mito-chondrial function after exposure to amyloidbeta-peptide, iron, and 3-nitropropionic acid.J. Neurochem. 75, 314–320.
Haughey N. J., Liu D., Nath A., Borchard A. C., andMattson M. P. (2002a) Disruption of neurogenesisin the subventricular zone of adult mice, and inhuman cortical neuronal precursor cells in culture,by amyloid beta-peptide: implications for thepathogenesis of Alzheimer’s disease. Neuromole-cular Med. 1, 125–135.
Haughey N. J., Nath A., Chan S. L., Borchard A. C.,Rao M. S., and Mattson M. P. (2002b) Disruptionof neurogenesis by amyloid beta-peptide, and per-turbed neural progenitor cell homeostasis, inmodels of Alzheimer’s disease. J. Neurochem. 83,1509–1524.
Haydon P. G. (2001) GLIA: listening and talking to thesynapse. Nat. Rev. Neurosci. 2, 185–193.
Hays S. J. (1998) Therapeutic approaches to the treat-ment of neuroinflammatory diseases. Curr. Pharm.Des. 4, 335–348.
Jensen M. B., Finsen B., and Zimmer J. (1997)Morphological and immunophenotypic micro-glial changes in the denervated fascia dentata ofadult rats: correlation with blood–brain barrierdamage and astroglial reactions. Exp. Neurol. 143,103–116.
Jiang W., Gu W., Brannstrom T., Rosqvist R., and WesterP. (2001) Cortical neurogenesis in adult rats aftertransient middle cerebral artery occlusion. Stroke32, 1201–1207.
Jin K., Minami M., Lan J. Q., et al. (2001) Neuro-genesis in dentate subgranular zone androstral subventricular zone after focal cerebralischemia in the rat. Proc. Natl. Acad. Sci. USA 98,4710–4715.
Kempermann G., Kuhn H. G., and Gage F. H. (1997)More hippocampal neurons in adult mice livingin an enriched environment. Nature 386, 493–495.

194 Lee, Auyeung, and Mattson
NeuroMolecular Medicine Volume 4, 2003
Kuhn H. G. and Svendsen C. N. (1999) Origins,functions, and potential of adult neural stem cells.Bioessays 21, 625–630.
Lee J., Duan W., Long J. M., Ingram D. K., and Matt-son M. P. (2000) Dietary restriction increases thenumber of newly generated neural cells, andinduces BDNF expression, in the dentate gyrus ofrats. J. Mol. Neurosci. 15, 99–108.
Lee J., Seroogy K. B., and Mattson M. P. (2002a) Dietaryrestriction enhances neurotrophin expression andneurogenesis in the hippocampus of adult mice.J. Neurochem. 80, 539–547.
Lee J., Duan W. and Mattson M. P. (2002b) Evidencethat brain-derived neurotrophic factor is requiredfor basal neurogenesis and mediates, in part, theenhancement of neurogenesis by dietary restric-tion in the hippocampus of adult mice. J. Neuro-chem. 82, 1367–1375.
Lee J., Chan S. L., and Mattson M. P. (2002c) Adverseeffect of a presenilin-1 mutation in microglia resultsin enhanced nitric oxide and inflammatory cyto-kine responses to immune challenge in the brain.Neuromolecular Med. 2, 29–45.
Long J. M., Kalehua A. N., Muth N. J., et al. (1998)Stereological estimation of total microglia numberin mouse hippocampus. J. Neurosci. Methods 84,101–108.
Mattson M. P., Chan S. L., and Duan W. (2002) Modi-fication of brain aging and neurodegenerative dis-orders by genes, diet, and behavior. Physiol. Rev.82, 637–672.
Morgan T. E., Nichols N. R., Pasinetti G. M., and FinchC. E. (1993) TGF-beta 1 mRNA increases inmacrophage/microglial cells of the hippocampus inresponse to deafferentation and kainic acid-inducedneurodegeneration. Exp. Neurol. 120, 291–301.
Nilsson M., Perfilieva E., Johansson U., Orwar O., andEriksson P. S. (1999) Enriched environmentincreases neurogenesis in the adult rat dentategyrus and improves spatial memory. J. Neurobiol.39, 569–578.
Parent J. M., Yu T. W., Leibowitz R. T., et al. (1997) Den-tate granule cell neurogenesis is increased byseizures and contributes to aberrant networkreorganization in the adult rat hippocampus.J. Neurosci. 17, 3727–3738.
Paxinos G. and Franklin K. B. J. (1997) The mouse brainin stereotaxic coordinates. New York: AcademicPress, p. 132.
Peterson D. A. (2002) Stem cells in brain plasticity andrepair. Curr. Opin. Pharmacol. 2, 34–42.
Rao M. S. and Mattson M. P. (2001) Stem cells andaging: expanding the possibilities. Mech. AgeingDev. 122, 713–734.
Rossi F. and Cattaneo E. (2002) Opinion: neural stemcell therapy for neurological diseases: dreams andreality. Nat. Rev. Neurosci. 3, 401–409.
Sankar R., Shin D., Liu H., Katsumori H., and Waster-lain C. G. (2000) Granule cell neurogenesis afterstatus epilepticus in the immature rat brain. Epilep-sia 41, S53–S56.
Scharfman H. E., Goodman J. H., and Sollas A. L. (2000)Granule-like neurons at the hilar/CA3 border afterstatus epilepticus and their synchrony with areaCA3 pyramidal cells: functional implications ofseizure-induced neurogenesis. J. Neurosci. 20,6144–6158.
Schauwecker P. E. (2000) Seizure-induced neuronaldeath is associated with induction of c-JunN-terminal kinase and is dependent on geneticbackground. Brain Res. 884, 116–128.
Scott B. W., Wang S., Burnham W. M., De Boni U., andWojtowicz J. M (1998) Kindling-induced neuro-genesis in the dentate gyrus of the rat. Neurosci.Lett. 248, 73–76.
Scott B. W., Wojtowicz J. M., and Burnham W. M. (2000)Neurogenesis in the dentate gyrus of the rat fol-lowing electroconvulsive shock seizures. Exp.Neurol. 165, 231–236.
Shors T. J., Miesegaes G., Beylin A., Zhao M., Rydel T.,and Gould, E. (2001) Neurogenesis in the adult isinvolved in the formation of trace memories.Nature 410, 372–376.
Song H., Stevens C. F., and Gage F. H. (2002a) Astrogliainduce neurogenesis from adult neural stem cells.Nature 417, 39–44.
Ullian E. M., Sapperstein S. K., Christopherson K. S.,and Barres B. A. (2001) Control of synapse numberby glia. Science 291, 657–661.
van Praag H., Kempermann G., and Gage, F. H. (1999)Running increases cell proliferation and neuro-genesis in the adult mouse dentate gyrus. Nat.Neurosci. 2, 266–270.
Vezzani A., Conti M., De Luigi A., et al. (1999)Interleukin-1beta immunoreactivity and microgliaare enhanced in the rat hippocampus by focal kainateapplication: functional evidence for enhancement ofelectrographic seizures. J. Neurosci. 19, 5054–5065.
West M. J. (1993) New stereological methods for count-ing neurons. Neurobiol. Aging 14, 275–285.
Yoshimura S., Takagi Y., Harada J., et al. (2001)FGF-2 regulation of neurogenesis in adult hip-

Hippocampal Microgliosis and Neurogenesis 195
NeuroMolecular Medicine Volume 4, 2003
pocampus after brain injury. Proc. Natl. Acad. Sci.USA 98, 5874–5679.
Yu B. P., Masoro E. J., and McMahan C. A. (1985) Nutri-tional influences on aging of Fischer 344 rats: I.Physical, metabolic, and longevity characteristics.J. Gerontol. 40, 657–670.
Yu Z. F. and Mattson M. P. (1999) Dietary restric-tion and 2-deoxyglucose administration reduce
focal ischemic brain damage and improvebehavioral outcome: evidence for a precon-ditioning mechanism. J. Neurosci. Res. 57,830–839.
Zhu H., Guo Q., and Mattson M. P. (1999) Dietaryrestriction protects hippocampal neurons againstthe death- promoting action of a presenilin-1 muta-tion. Brain Res. 842, 224–229.





























