Embed Size (px)
Citation preview

Nda
JYa
b
c
d
e
a
ARR2AA
KDANHLM
1
ncppAc
oH
w
h0
Int. J. Devl Neuroscience 49 (2016) 67–78
Contents lists available at ScienceDirect
International Journal of Developmental Neuroscience
j ourna l ho me page: www.elsev ier .com/ locate / i jdevneu
europrotective effects of docosahexaenoic acid on hippocampal celleath and learning and memory impairments in a valproiccid-induced rat autism model
ingquan Gaoa,b, Xuelai Wanga, Hongli Sunc, Yonggang Caoc, Shuang Lianga, Han Wanga,anming Wangd, Feng Yange, Fengyu Zhange,∗∗,1, Lijie Wua,∗,1
Department of Child and Adolescent Health, School of Public Health, Harbin Medical University, Harbin, Heilongjiang 150081, ChinaDepartment of Nursing, School of Public Health, Harbin Medical University, Harbin, Heilongjiang 150081, ChinaDepartment of Basic Medical Sciences, School of Public Health, Harbin Medical University, Harbin, Heilongjiang 150081, ChinaDepartment of Pharmacology, Harbin Medical University in Daqing, Daqing, Heilongjiang 163319, ChinaThe Global Clinical and Translational Research Institute, Bethesda, MD 20814, USA
r t i c l e i n f o
rticle history:eceived 10 July 2015eceived in revised form0 November 2015ccepted 20 November 2015vailable online 27 November 2015
eywords:ocosahexaenoic acid (DHA)utismeuroprotective
a b s t r a c t
Prenatal exposure to valproic acid (VPA) in rat offspring is capable of inducing experimental autism withneurobehavioral aberrations. This study investigated the effect of docosahexaenoic acid (DHA) on hip-pocampal cell death, learning and memory alteration in an experimental rat autism model. We found thatDHA supplementation (75, 150 or 300 mg/kg/day, 21 days) rescued the VPA (600 mg/kg) induced DHAreduction in plasma and hippocampus in a dose-dependent manner, increased the levels of hippocampalp-CaMKII and p-CREB without affecting total protein level, and altered BDNF-AKT-Bcl-2 signaling path-way, as well as inhibited the activity of caspase-3. DHA also influenced the content of malondialdehyde(MDA) and the activities of antioxidant enzymes in the VPA-treated offspring. Consistent with the previ-ous results, we also observed that 300 mg/kg DHA supplementation markedly increased the cell survival,decreased the cell apoptosis, and increased mature neuronal cell in the hippocampus in VPA-treated
ippocampal cellearning impairmentemory impairment
offspring. Utilizing the Morris water maze test, we found that DHA prevented cognitive impairment inoffspring of VPA-treated rats. The data suggested that DHA may play a neuroprotective role in hippocam-pal neuronal cell and ameliorates dysfunctions in learning and memory in this rat autism model. Thus,DHA could be used as treatment intervention for mitigating behavioral dysfunctions in autism spectrum
disorder (ASD).. Introduction
Autism Spectrum Disorders (ASD) are significant and complexeurodevelopmental disorders with persistent deficits in socialommunication and social interactions and restricted, repetitiveatterns of behavior, interests or activities, which are usuallyresent in the early developmental period (American Psychiatric
ssociation, 2013). The prevalence of ASD have increased dramati-ally in the past decade (Maenner and Durkin, 2010; Rutter, 2005),∗ Corresponding author at: Department of Child and Adolescent Health, Schoolf Public Health, Harbin Medical University, 157 Baojian Road, Nangang District,arbin 150081, China.
∗∗ Corresponding author.E-mail addresses: [email protected] (F. Zhang), [email protected],
[email protected] (L. Wu).1 Equally contributed to this paper.
ttp://dx.doi.org/10.1016/j.ijdevneu.2015.11.006736-5748/© 2016 Published by Elsevier Ltd. on behalf of ISDN.
© 2016 Published by Elsevier Ltd. on behalf of ISDN.
up to the current estimated prevalence of 1 out of 68 in youngchildren under age of 8 years (CDC, 2012).
ASD patients are usually accompanied by reducing global cog-nitive function score (Meador et al., 2009); about 75% of them havelearning and memory dysfunction (Polleux and Lauder, 2004). Thehippocampus is known to play a critical role in the processing ofspatial learning and memory, and is one of the major brain lociimpaired in animals with autism (Edgin and Pennington, 2005).Several studies have found that VPA treatment suppresses long-term potentiation (LTP) in hippocampal slices (Lee et al., 1996; Wuet al., 2010).
Docosahexaenoic acid (DHA, 22:6n-3), a component of thephospholipid structure of cellular membranes in the brain, isessential for normal neurological function (Fedorova and Salem,
2006; Strickland, 2014). Postnatal DHA status has been foundassociated with neurodevelopment (Agostoni et al., 1995). WhileDHA accumulates rapidly during brain development from prena-tal day 7 to postnatal day 16 in the whole rat brain (Green et al.,
6 euros
1bAas(dpsE
ednr2pOAs2D2pwpD
mm
2
2
uitTm
2
cdMA2ciwatmisfitrwioc
8 J. Gao et al. / Int. J. Devl N
999), and from the third trimester of gestation to 2 years afterirth in the human brain (Lauritzen et al., 2001; Martinez, 1992).dditionally, epidemiological and clinical studies have indicated
link between low level of plasma DHA and mental diseasesuch as depression (McNamara and Carlson, 2006); schizophreniaHorrocks and Farooqui, 2004) and attention-deficit/hyperactivityisorder (Bekaroglu et al., 1996; Fedorova and Salem, 2006). Sup-lementation with DHA has been shown to ameliorate clinicalymptoms of these disorders (Colangelo et al., 2009; Das, 2004;msley et al., 2003; Horrocks and Farooqui, 2004; Rees et al., 2009).
A number of studies have focused on DHA and ASD. Van Elstt al. (2014) stated that a change in the ratio of n-6–n-3, especiallyuring early life, may induce developmental changes in brain con-ectivity, synaptogenesis, cognition and behavior that are directlyelated to ASD. Several studies (Al-Farsi et al., 2013; Foley et al.,012; Wiest et al., 2009) have found that the level of DHA inlasma is lower in children with ASD than children without it.ur recent metabolomics study using two independent cohorts ofSD patients and normal controls suggest that altered DHA level inerum is one of major metabolites associated with ASD (Wang et al.,016). In addition, studies have reported that the dietary ratio ofHA and arachidonic acid (AA) affect characteristic of ASD (Malone,012; Yui, 2012; Yui et al., 2011). Some studies failed to provideositive findings in children with ASD using DHA supplementationith 400 mg/day (Johnson et al., 2010), possibly due to small sam-le size and limited outcome measures, but it is not clear whetherHA treatment has potential benefit to ASD.
We designed this study to evaluate the effect of DHA supple-ent on impaired learning and memory and to understand possibleechanisms of DHA in an autism rat animal model.
. Methods and materials
.1. Ethical approval
Great efforts were made to minimize the number of animalssed and their suffering. All animal experiments were conducted
n strict accordance with the criteria described in the National Insti-utes of Health Guide for the Care and Use of Laboratory Animals.his study was reviewed and approved by the Animal Care Com-ittee of Harbin Medical University.
.2. Animals
Experimentally naïve male and female Wistar rats were pur-hased from a commercial breeder (YISI: Changchun, China),elivered to the campus of Harbin Medical University in Daqing.ale and female fed separately and housed in groups of 3–4.
ll rats were maintained under standard laboratory conditions at2 ± 2 ◦C, with 50 ± 10% relative humidity and on a 12 h-light/darkycle, with food and water available ad libitum. Female rats weigh-ng 250–280 g (n = 30) and male rats weighing 280–350 g (n = 20)
ere used. Before any experimental procedure was carried out,nimals were allowed one week to acclimatize to the experimen-al rooms. Then, their fertility cycles were controlled, and they
ated overnight during protesters. Vaginal smears were examinedn the following morning, and placed on glass slides for analy-is. If spermatozoa were found in the morning examination, therst day of pregnancy (E0) was determined. Each pregnant rat washen housed separately. Female rats were divided into two groups:eceiving VPA treatment (n = 16) and controls (n = 4). VPA (Sigma)
as dissolved in 0.9% saline for a concentration of 250 mg/ml. Ratsn the treatment group received a single intraperitoneal injectionf 600 mg/kg VPA on E12.5 day after conception, and those in theontrol group were injected with the same amount of physiolog-
cience 49 (2016) 67–78
ical saline at the same time (Schneider and Przewłocki, 2005). Allfemale rats were housed individually and allowed to raise their ownlitters. All rat offspring exposed to VPA during gestation developeda characteristic ‘kink’ in the tail (Foley et al., 2012; Vorhees, 1987),which was easily distinguishable from age-matched controls. Theoffspring rats were weaned on postnatal day (PND) 21.
The following experiments were conducted on the male off-spring. On the 14th PND all VPA-induced male offspring rats (n = 60)were randomly divided into four groups by different marker withtrinitrophenol: one without DHA treatment (n = 16) and three withDHA treatment at different dosages of 75 mg/kg (n = 14), 150 mg/kg(n = 14) and 300 mg/kg (n = 16). DHA doses were based on recom-mended human doses and adjusted for a 10-fold higher metabolicrate of rats (75, 150 and 300 mg/kg for rats corresponding approx-imately to 500, 1000 and 2000 mg daily for humans, respectively).From the 14th (Yochum et al., 2008) to 35th PND, DHA was admin-istered by gavage (1 ml/day), and every group was given the samevolume of corn oil, and all the rats were executed on the 40th PND.
2.3. Total DHA content analysis of offspring plasma andhippocampus
Ultra-Performance Liquid Chromatography (UPLC) analysis wasperformed to test the DHA level of the hippocampus and plasmain rats by different groups. Wet tissue was weighed accurately(hippocampus about 0.03 g or plasma 200 �l), as well as 600 �lor 400 �l ethanol added respectively. Hippocampus tube histor-rhexis for 30 s and plasma tube vortex for 30 s. Internal standards(prostaglandin B1) PGB1 (400 �l, 100 pmol/ml ethanol solution),acetic acid (2 �l, 88% w/v), and BHT (1 �l, 0.5% w/v ethanol solutionas an antioxidant) were added to the sample, following high-speedshocks for 30 s. After mixing, the sample was left on ice for 1 h toextract the arachidonic acid metabolites followed by centrifugationat 15,000 rpm for 20 min at 4 ◦C. Supernatant was moved to anotherfresh tube and diluted with 3 ml distilled water. Then the solu-tion was applied to a Strata-X polymeric SPE column (200 mg/ml)that had been preconditioned with 100% ethanol 5 ml and 25%ethanol 5 ml. The column was washed with distilled water 10 ml,and 25% ethanol 5 ml which was allowed to run dry. Thereafter, theeicosanoids were eluted from the column with ethyl acetate con-taining 0.0002% BHT 5 ml, and then ethyl acetate layer was driedby N2 flushing on ice bath. After one hour, the dried sample wasreconstituted in 0.05% acetic acid (2000 �l, ethanol solution) astest solution. UPLC analysis was conducted on a Waters AcquityTM
UPLC system. A Waters AcquityTM UPLC BEH C18 column (1.7 �mparticle sizes, 2.1 × 50 mm column) was used, and the column wasmaintained at 35 ◦C. Sample injection volume was 5 �l, and sampleinjector was maintained at 4 ◦C throughout the analysis. Mobilephase B is pure water; phase A is acetonitrile and flow rate is0.20 ml/min. Mobile phase gradient ran from 35% A to 90% A over7 min, returning to 35% A over 1.5 min, and was held at 35% A for1.5 min for re-equilibration.
2.4. Morris water maze task
The Morris water maze (MWM) task is an established test ofspatial learning and reference memory. The MWM apparatus, awhite circular pool with a diameter of 120 cm and a height of 45 cm,was placed in an experimental room. The pool was geographicallydivided into four equal quadrants. Four different visual cues weremounted on the four directions of the walls. The pool was filledwith opaque water to a height of 25 cm; the water temperature
was 21–22 ◦C. A 10 cm diameter platform was located in the centerof a quadrant of the tank and submerged 1.5 cm below the watersurface. A video camera was mounted above the center of the tankto record all trials. MWM tests were done for five consecutive days,
euros
setfmpdwlt1ihbp
wtshttsptft1a
2
(i7iwa4nis
2
NiisSf
2
itwpwr1t3
J. Gao et al. / Int. J. Devl N
tarting on the 36th PND. Prior to the test, training was done twiceach day for two-and-a-half subsequent days, that is, a total of 5imes. At the initial training, all rats were allowed to swim freelyor 120 s in order to be familiar with the novel environment of the
aze and to locate and climb onto the escape platform. The pur-ose of this procedure was to decrease the level of stress in ratsuring MWM task. During following four training sessions, a ratas placed in the water facing the wall at one of four random start
ocations (north, south, east and west). Each rat was then allowedo find the submerged platform within 60 s and to rest on it for0 s. If rat failed to find a hidden platform within the assigned time,
t was placed on the platform for 10 s. The rat was returned to itseated cage for 5 min inter-trial intervals; and then the rat woulde placed in water at a different start location to repeat the samerocedure. Each rat was trained separately.
After training, the rats were tested for 5 times and the resultsere documented as official experimental results. From the first to
he fourth time, rats were placed in the water facing the wall, oppo-ite to the quadrant of the platform, as a starting location. The ratad to find the submerged platform, and the escape latency (timeo reach the platform) was used as an index of performance in theask. For rats who did not reach the platform, the latency time waset at 60 s. Throughout the training and official tests, the submergedlatform remained in the same position. At the fifth time, a proberial was performed for 60 s, in which the platform was removedrom the tank. A rat was placed in the water at the same start loca-ion. The time of rat passed through the circular area (diameter,0 cm) which formerly contained the submerged platform duringcquisition, were taken as measures of spatial memory.
.5. Tissue processing for histology
For histological examination, on PND 40, rats from each groupn = 6) were deeply anesthetized with chloral hydrate (300 mg/kg,.p.) and perfused with 0.1 M phosphate-buffered saline (PBS, PH.4), following that they were perfused by 4% paraformaldehyde
n 0.1 M phosphate buffer. After perfusion, whole brain of a ratas carefully dissected, then fixed in 4% paraformaldehyde for 48 h
nd cryo-protected by infiltration with 30% sucrose for 3 days at◦C. Frozen tissues containing the hippocampus were then cut at aominal thickness of 7 mm in the horizontal plane using a freez-
ng microtome (CM1900 UV, Leica, Germany). All sections wereubsequently stored in −80 ◦C refrigerator.
.6. Nissl staining
Neuronal cell survival was assessed by Nissl staining. Inissl staining, after rinsing with deionized water, sections were
mmersed in 0.1% cresyl violet at 37 ◦C for 18 min, and then rins-ng with deionized water again. Sections were dehydrated, coverlipped with neutral balsam and observed under a light microscope.ix brain sections were selected from each animal and processedor counting.
.7. NeuN immunohistochemistry
Mature neurons were assessed by NeuN immunohistochem-stry. After rinsing with PBS, tissue sections were sequentiallyreated with 0.3% hydrogen peroxide for 10 min and then rinsingith PBS. After that, sections were treated with 10% normal goatlasma for 15 min at room temperature and sequentially incubatedith mouse anti-NeuN (1:50, Chemicon, CA) overnight at 4 ◦C. After
insing, sections were exposed to biotinylated goat-mouse IgG for5 min at 37 ◦C, and after rinsing with PBS, they were exposedo streptavidin peroxidase (Vector, Burlingame, CA) for 15 min at7 ◦C. Next step, sections were soaked in 3.3-diaminobenzidine
cience 49 (2016) 67–78 69
(DAB) for about 5 min. Water was then added to stop DAB reac-tion. The stained sections were dehydrated using a graded series ofethanol solutions, cleared in xylene, cover slips with neutral balsamand examined with a light microscope. Control sections were incu-bated with 10% normal goat plasma instead of primary antibody.All subsequent incubations were performed as described above andno positive immunoreaction was observed.
2.8. TUNEL staining
TUNEL staining was carried out on sections by using the in situcell death detection kit, according to the manufacturer’s instruc-tions. Briefly, sections were immersed in 4% paraformaldehyde for30 min. And after rinsing with PBS, sections were treated with0.3% hydrogen peroxide for 30 min, treated with proteinase K(20 mg/ml) for 25 min at room temperature. Subsequently, the sec-tions were incubated with the TUNEL reaction mixture for 1 hat 37 ◦C. After washing with PBS, sections were incubated withconverter-POD for 30 min and then visualized with DAB. Sectionswere then counter stained with hematoxylin for 3 min and rinsedunder running water. After dehydration in graded ethanol seriesand transparent in xylene, the brain sections were mounted ontogelatin-coated slides. Apoptotic cell counting was performed in thehippocampus (CA1) in the hemisphere. Cells with yellow–browngranules in the nucleus were considered to be apoptotic cells.
2.9. Enzymatic activity measurement
Enzymatic activities of glutathione (GSH), glutathione peroxi-dase (GSH-PX), superoxide dismutase (SOD) and malondialdehyde(MDA) were measured according to manufacturer’s instructions indifferent commercial assay kits (Jian Cheng Bioengineering Insti-tute, Nanjing, China). Activities of GSH and GSH-PX were assayedby quantifying the oxidation rate of reduced glutathione to oxi-dized glutathione by Hydrogen peroxide. SOD activity was assessedin hippocampus homogenate by calculating rate inhibition ofnucleotide oxidation. The results were defined as U/mg protein. Theconcentration of MDA was determined at a wavelength of 532 nm,by measuring thiobarbituric acid (TBA) reacting substances. Levelsof MDA were expressed per mg protein nmol MDA. Protein contentwas determined by Coomassie brilliant blue method using bovineplasma albumin as standard (Li et al., 2008).
2.10. Western blot assays
The Western blot assay was performed on the hippocampussamples of rats from different groups. Hippocampus samples werewashed three times with PBS and were homogenized in an ice-cold RIPA lysis buffer (50 mM Tris–HCl, 150 mM NaCl, 10% glycerol,1% Nonidet P-40, 5 mM EDTA and 1 mM phenylmethylsulfonylfluoride) (Beyotime, China). Homogenates were centrifuged at13,000 rpm for 15 min at 4 ◦C. Supernatants were collected andtotal protein concentrations were determined by BCA methodusing bovine plasma albumin as the standard. Equal aliquots (60 g)of protein samples were applied to 10% or 12% SDS-PAGE gels,transferred to pure nitrocellulose membranes (PerkinElmer LifeSciences, Boston, MA), and blocked with 5% non-fat milk in Tris-buffered saline with 0.1% Tween-20 (TBS-T). Membranes werethen probed separately with following primary antibodies: anti-beta-actin (1:5000, Cell Signaling technology, USA), monoclonalmouse (1:2000, Cell Signaling technology, USA), anti-caspase-3(1:1000, Cell Signaling technology, USA), polyclonal rabbit (1:2000,
Cell Signaling technology, USA), anti-Bax (1:1000, Cell Signalingtechnology, USA), polyclonal rabbit (1:2000, Cell Signaling tech-nology, USA), anti-Bcl-2 (1:1000, Cell Signaling technology, USA),monoclonal rabbit (1:2000, Cell Signaling technology, USA), anti-
70 J. Gao et al. / Int. J. Devl Neuroscience 49 (2016) 67–78
Fig. 1. Reduced DHA concentration in the plasma and hippocampus of VPA-treated rats was rescued with administration of DHA by gavage feeding. VPA-treated rats weres rum (Bi ntrol;
C(SiUarGTs1esb
2
b(
ampled on the 40th PND. UPLC total ion chromatograms of hippocampus (A) and sen hippocampus in different groups (Mean ± S.E.M. n = 6). ##, P < 0.01, for VPA vs. co
REB (1:1000, Cell Signaling technology, USA), monoclonal rabbit1:2000, Cell Signaling technology, USA), anti-CaMKII (1:1000, Cellignaling technology, USA), monoclonal rabbit (1:2000, Cell Signal-ng technology, USA), anti-BDNF (1:200, Cell Signaling technology,SA), monoclonal rabbit (1:2000, Cell Signaling technology, USA),nti-Akt (1:1000, Cell Signaling technology, USA), monoclonalabbit (1:2000, Cell Signaling technology, USA), and mouse anti-APDH (1:2000, Santa Cruz Biotechnology, USA) at 4 ◦C overnight.hen membranes were incubated with anti-rabbit or anti-mouseecondary antibodies (Beyotime, China) at room temperature for
h. Chemiluminescent detection was performed by an ECL West-rn blotting detection kit. Results were analyzed by Quantity Oneoftware to obtain the optical density ratio of the target protein toeta- actin.
.11. Caspase-3 activity assay
Caspase-3 activity of hippocampus homogenates was assayedy cleavage of chromogenic caspase substrates, Ac-DEVD-pNAacetyl-Asp-Glu-Val-Asp p-nitroanilide) which is a caspase-3 sub-
) samples. C. Total DHA content in plasma in different groups; D. Total DHA content*, P < 0.05 and **, P < 0.01, for VPA vs. DHA treatment.
strate. The amount of caspase-3 was measured at a wave length of405 nm as directed by the kit (Beyotime Biotechnology). Hippocam-pal protein samples were prepared as described in Western blotanalysis. Approximately 60 �g of total protein was added to a reac-tion buffer containing Ac-DEVD-pNA (2 mM), incubated at 37 ◦C for4 h and absorbance of yellow pNA was measured by a spectrometerat a wave length of 405 nm. Specific caspase-3 activity which wasnormalized for total protein in hippocampus was then expressedas fold of baseline caspase-3 activity in the control group.
2.12. Statistical analysis
The means of the experimental data described above and associ-ated standard error were calculated and compared between groups.
One-way ANOVA was performed followed by Dunnett’s test forindividual comparisons between group means. All statistical analy-ses were carried out using SPSS 18.0 software; and the significancelevel was set at 0.05.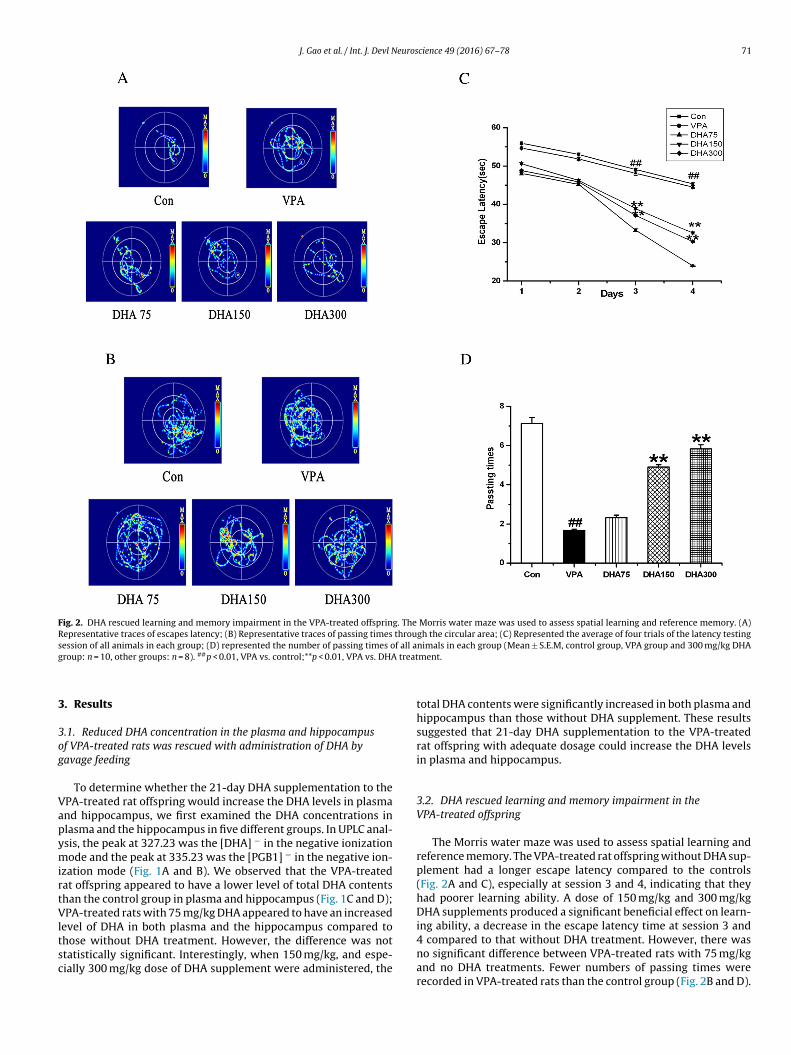
J. Gao et al. / Int. J. Devl Neuroscience 49 (2016) 67–78 71
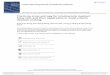
Fig. 2. DHA rescued learning and memory impairment in the VPA-treated offspring. The Morris water maze was used to assess spatial learning and reference memory. (A)Representative traces of escapes latency; (B) Representative traces of passing times through the circular area; (C) Represented the average of four trials of the latency testings f all ag treat
3
3og
VapymirtVltsc
ession of all animals in each group; (D) represented the number of passing times oroup: n = 10, other groups: n = 8). ##p < 0.01, VPA vs. control;**p < 0.01, VPA vs. DHA
. Results
.1. Reduced DHA concentration in the plasma and hippocampusf VPA-treated rats was rescued with administration of DHA byavage feeding
To determine whether the 21-day DHA supplementation to thePA-treated rat offspring would increase the DHA levels in plasmand hippocampus, we first examined the DHA concentrations inlasma and the hippocampus in five different groups. In UPLC anal-sis, the peak at 327.23 was the [DHA] − in the negative ionizationode and the peak at 335.23 was the [PGB1] − in the negative ion-
zation mode (Fig. 1A and B). We observed that the VPA-treatedat offspring appeared to have a lower level of total DHA contentshan the control group in plasma and hippocampus (Fig. 1C and D);PA-treated rats with 75 mg/kg DHA appeared to have an increased
evel of DHA in both plasma and the hippocampus compared to
hose without DHA treatment. However, the difference was nottatistically significant. Interestingly, when 150 mg/kg, and espe-ially 300 mg/kg dose of DHA supplement were administered, thenimals in each group (Mean ± S.E.M, control group, VPA group and 300 mg/kg DHAment.
total DHA contents were significantly increased in both plasma andhippocampus than those without DHA supplement. These resultssuggested that 21-day DHA supplementation to the VPA-treatedrat offspring with adequate dosage could increase the DHA levelsin plasma and hippocampus.
3.2. DHA rescued learning and memory impairment in theVPA-treated offspring
The Morris water maze was used to assess spatial learning andreference memory. The VPA-treated rat offspring without DHA sup-plement had a longer escape latency compared to the controls(Fig. 2A and C), especially at session 3 and 4, indicating that theyhad poorer learning ability. A dose of 150 mg/kg and 300 mg/kgDHA supplements produced a significant beneficial effect on learn-ing ability, a decrease in the escape latency time at session 3 and
4 compared to that without DHA treatment. However, there wasno significant difference between VPA-treated rats with 75 mg/kgand no DHA treatments. Fewer numbers of passing times wererecorded in VPA-treated rats than the control group (Fig. 2B and D).
72 J. Gao et al. / Int. J. Devl Neuroscience 49 (2016) 67–78
Fig. 3. DHA rescued neuronal loss and apoptosis in the offspring hippocampus of VPA-treated rats. A. Representative photomicrographs of offspring hippocampal CA1 andCA3 regions with Nissl staining (×20X and ×40X); B and C. Cell counts per visual field (×40X) found in the slides with Nissl staining in the regions of hippocampal CA1andCA3, respectively; D. Representative photomicrographs of offspring hippocampal CA1 and CA3 regions with NeuN immunohistochemistry (×20X and ×40X); E and F. Thenumber of positive cells (dark brown indicate NEUN positive cells) per visual field (×40X) found in the slides with NeuN immunohistochemistry in the regions of hippocampalCA1 and CA3, respectively; G. Representative photomicrographs of offspring hippocampal CA1 region with TUNEL staining (×20X and ×40X); H. The number of positive cells(dark brown indicate apoptotic cell) per visual field (×40X) found in the slides with TUNEL staining in the regions of hippocampal CA1. (Mean ± S.E.M, n = 6). ##p < 0.01, VPAvs. control; *p < 0.05, and **p < 0.01, VPA vs. DHA treatment.
J. Gao et al. / Int. J. Devl Neuroscience 49 (2016) 67–78 73
Fig. 4. Increased peroxidation in the offspring hippocampus of the VPA-treated rats was inhibited by enhanced activities of antioxidant enzymes induced by DHA supple-m n MDr 1, VPA
AwiDai
3t
httpslniegclw
entation. (A–D) showed the effects of DHA supplementation at different doses oespectively. (Mean ± S.E.M, n = 6). ##p < 0.01, VPA vs. control; *p < 0.05, and **p < 0.0
s shown in Fig. 2B and D, interestingly, VPA-treated rat offspringith 150 mg/kg and 300 mg/kg of DHA supplement have significant
ncrease in the number of passing times compared to those withoutHA supplement. Taken together, the data suggest that 150 mg/kgnd 300 mg/kg of DHA treatment may ameliorate spatial memorympairments in the VPA-treated rats.
.3. DHA rescued neuronal loss and increase mature neurons inhe hippocampus of the VPA-treated rat offspring
To confirm whether DHA has any neuroprotective effect on theippocampus in the VPA-treated rat offspring, we performed his-ological examination of Nissl staining to assess neuronal loss inhe hippocampus. We observed extensive neuronal changes in hip-ocampal CA1 and CA3 regions of rat brains (Fig. 3A). For Nissltaining, neurons that contained Nissl substance in the cytoplasm,oose chromatin and the nuclei appeared as pallid blue, prominentucleoli were considered to be normal, and damaged neurons were
dentified by dark blue, cavitations around the nucleus and the pres-nce of pyknotic homogeneous nuclei. In Nissl staining of control
roup, the neuronal cell outline was clear and the structure wasompact with abundant cytoplasm and cell body. Evident neuronaloss and neuronal degeneration were observed in the VPA groupith cells arranged sparsely with vague cell outlines. The number of
A, SOD, GSH and GSH-PX activities in hippocampus of VPA-treated rat offspring, vs. DHA treatment.
cells with eumorphism was significantly smaller in the VPA-treatedrats compared to normal control rats (Fig. 3B and C). DHA sup-plementation, especially at the dose of 150 mg/kg and 300 mg/kg,substantially increased the proportion of neurons that survived(Fig. 3B and C).
Similarly, the study in NeuN immunohistochemistry also indi-cated the decreasing of mature neurons in the pyramidal cells ofhippocampal CA1 and CA3 regions in the VPA-treated rats with-out DHA supplement (Fig. 3D). The number of positive cells (darkbrown indicates NeuN positive cells) was much lower in the VPA-treated group without DHA treatment compared to the controls inthe hippocampal CA1 and CA3 (Fig. 3E and F). DHA supplementa-tion attenuated the neuronal damage following VPA treatment onNeuN immunoreaction of hippocampal CA1 and CA3, although noeffects were found in 75 mg/kg DHA treatment to the offspring ofVPA-treated rats (Fig. 3E and F).
TUNEL staining of tissue sections was carried out to determine ifDHA supplementation could prevent apoptosis in the hippocampusin the VPA-treated rat offspring. Significantly more apoptotic cellswere observed in the VPA group compared to the control group
(Fig. 3G). DHA supplementation, especially at dose of 150 mg/kgand 300 mg/kg, substantially decreased the proportion of apop-totic neurons on the hippocampus in the VPA-treated rat offspring,
7 euroscience 49 (2016) 67–78
am
slt
3re
mwtipgtp
eohgbwopawfahVthbtsMo
3o
3A3cr34s
3m
lptea
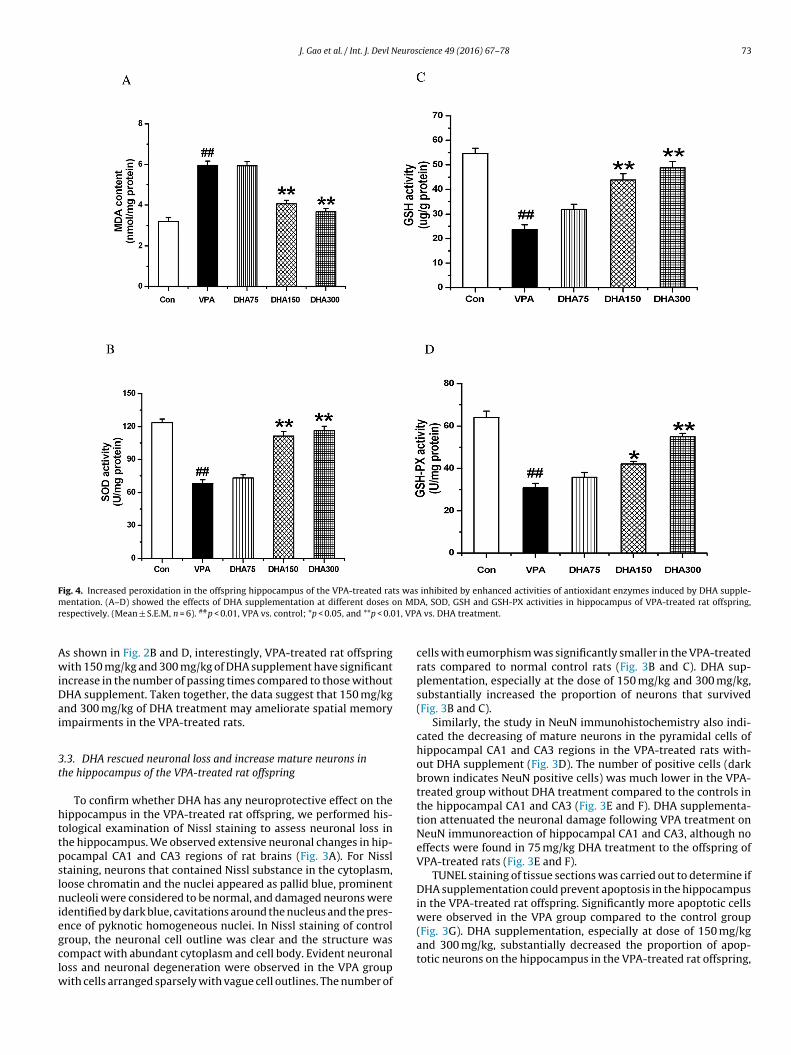
Fig. 5. DHA inhibited the increased activity of caspase-3 in the hippocampus of VPA-
4 J. Gao et al. / Int. J. Devl N
lthough no effects were found at a dose of 75 mg/kg DHA supple-entation (Fig. 3H).Taking together, evidences from three different histochemical
tudies support that DHA supplementation could rescue neuronaloss and increase mature neurons in the hippocampus of the VPA-reated rat offspring.
.4. Increased peroxidation in the hippocampus of VPA-treatedat offspring was inhibited by enhanced activities of antioxidantnzymes induced by DHA supplementation
To further investigate the neuroprotective role of DHA, we firsteasured MDA content and the activities of antioxidant enzymes,hich are believed to be involved in neuronal damage and protec-
ion (Meguid et al., 2011). As shown in Fig. 4A, the level of MDA, anndex of lipid peroxidation, significantly increased in the hippocam-us of VPA-treated rat offspring compared to the normal controlroup. After DHA supplementation (75, 150 and 300 mg/kg, respec-ively) to the offspring of VPA-treated rats, the VPA-mediated lipideroxidation significantly decreased with the increase of dosage.
The activities of antioxidants and anti-oxidative enzymes inach group are presented in Fig. 4B–D. Fig. 4B illustrated the activityf SOD in different groups. The activity of SOD in VPA -treated groupippocampus was significantly diminished compared to the controlroup. The offspring of VPA-treated rats supplemented with DHAy a dose of 75 mg/kg did not change the SOD activity. However,hen the dosage increased to 150 mg/kg or 300 mg/kg, VPA-treated
ffspring significantly increased in hippocampal SOD activity com-ared to those without DHA application. In this study, we alsossayed GSH and GSH-PX, which are an antioxidant and an enzymeith peroxidase activity to protect important cellular components
rom oxidative damage (Meguid et al., 2011). As shown in Fig. 4Cnd D, GSH concentration and GSH-PX activity had decreased in theippocampus of VPA-treated group compared to the control rats.PA-treated rat offspring with 75 mg/kg DHA administration tend
o have an increased GSH concentration and GSH-PX activity in theippocampus compared with those without DNA administration,ut the differences were not statistically significant. Whereas withhe increase in the dosage of DHA, GSH concentration increasedignificantly compared with those without DHA supplementation.eanwhile, the more dramatic elevation of GSH-PX activity was
bserved in the DHA supplementation at 150 and 300 mg/kg.
.5. DHA inhibited the activity of caspase-3 in the hippocampusf VPA-treated rats
To examine whether DHA supplementation suppresses caspase- activity, colorimetric analysis was carried out for a further study.s shown in Fig. 5, caspase-3 activity was markedly enhanced by65.02% in the VPA-treated rats without DHA supplementationompared with the normal control rat group. When VPA-treatedat offspring with DHA supplementation at dose of 75, 150 and00 mg/kg, all significantly reduced caspase-3 activity by 11.82%,0.26% and 59.44%, respectively, compared to those without DHAupplementation.
.6. DHA influenced BDNF-AKT-Bcl-2 signaling pathway toediate the enhanced expression of p-CaMKII and p-CREB
Previous studies demonstrated that BDNF-AKT-Bcl-2 intracel-ular signaling and caspase-3 pathway are involved in neuronal
rotection and apoptosis (Sheikh et al., 2010). To ascertain whetherhese signaling pathways could be affected by DHA treatment, wexamined the expressions of BDNF, p-Akt, Bcl-2, Bax and caspase-3fter exposure to different doses of DHA.treated rats. (Mean ± S.E.M, n = 6). ##p < 0.01, VPA vs. control; *p < 0.05, and **p < 0.01,VPA vs. DHA treatment.
We first observed a significant increase in BDNF expression inthe VPA-treated rat offspring compared to normal controls. Inter-estingly, the high doses of DHA treatment with 150 mg/kg and300 mg/kg further elevated the BDNF (Fig. 6A), but no effect wasfound at a dose of 75 mg/kg. We then assayed the expressionof p-Akt, one of downstream molecules in the BDNF-TrkB sig-naling pathway. The p-Akt/Akt expression ratio was significantlydecreased in the VPA-treated rat offspring compared to the con-trol group (Fig. 6B); and in parallel, the levels of p-Akt were alsoincreased at a dose of 150 and 300 mg/kg DHA supplementation.However, no changes in p-Akt were observed after DHA adminis-tration at a dose of 75 mg/kg in the VPA-treated rat offspring. Therewere no differences in total Akt expression found among all fivegroups.
Bcl-2 is specifically considered as an important anti-apoptoticprotein. We observed a significant decrease in the ratio of Bcl-2/GAPDH expression in the VPA-treated rat offspring compared tothe control group (Fig. 6C). Bcl-2 was rescued by DHA administra-tion in a dose-dependent manner.
Since Bax-dependent caspase-3 activation could be a key deter-minant in VPA-induced apoptosis in hippocampal neurons, wethus tested whether Bax and caspasin-3 expression changed inthis autism-like rats and DHA could rescue these alterations. Theexpression ratios of Bax/�-actin and caspase-3/GAPDH were sig-nificantly increased in the VPA-treated rat offspring compared tothe controls (Fig. 6D). The Bax/�-actin ratios were dramaticallydecreased after DHA supplementation compared to VPA-treatedrats without DHA treatment. Similarly, caspase-3/GAPDH expres-sions were decreased with DHA treatment in a dose-dependentmanner compared to the VPA-treated rats without DHA treatment(Fig. 6E).
CaMKII/CREB signaling proteins were regarded as the importantregulatory molecules in learning and memory formation. West-ern blot analysis was performed to evaluate the effects of DHAon the p-CaMKII and CaMKII protein expression in the hippocam-pus in different groups. The ratio of p-CaMKII/CaMKII expressionwas significantly decreased in the VPA-treated group compared tocontrols (Fig. 6F). The ratios were elevated in the VPA-treated ratswith different doses of DHA treatment compared to those withoutDHA supplementation except at dose of 75 mg/kg DHA treatment.
In contrast, total CaMKII expressions were not altered in all VPA-treated rats with and without DHA treatment. Further, we assessedthe effects of DHA on the expression of p-CREB and CREB, and found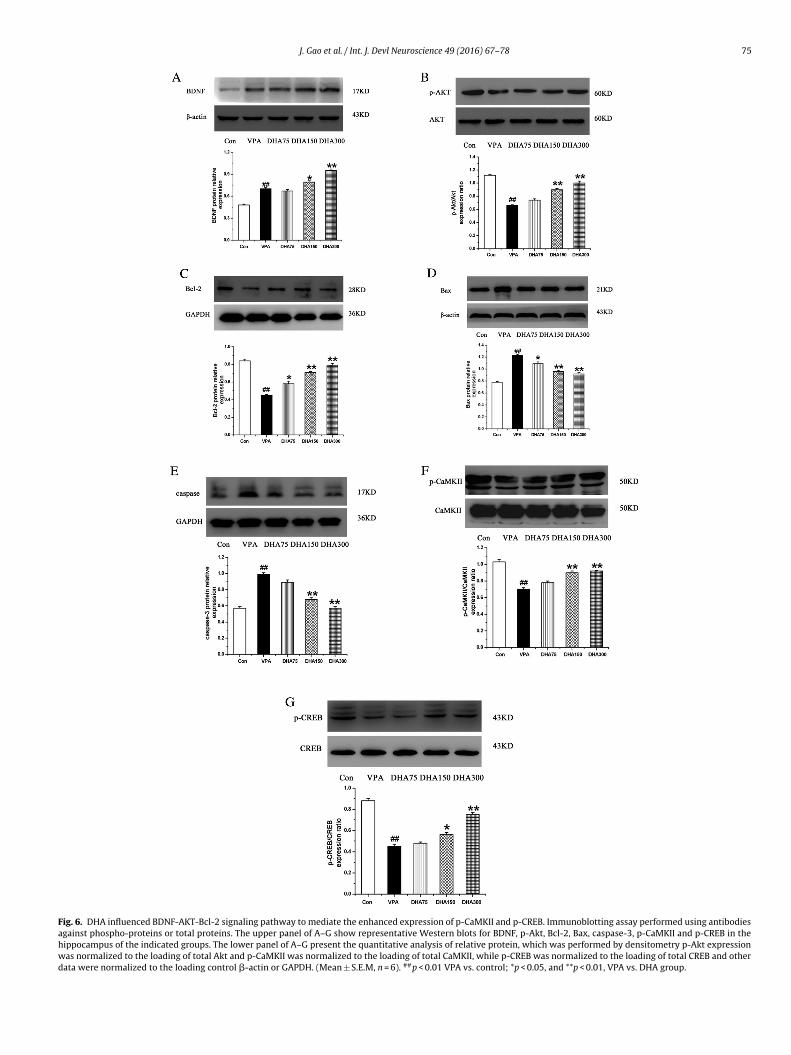
J. Gao et al. / Int. J. Devl Neuroscience 49 (2016) 67–78 75
Fig. 6. DHA influenced BDNF-AKT-Bcl-2 signaling pathway to mediate the enhanced expression of p-CaMKII and p-CREB. Immunoblotting assay performed using antibodiesagainst phospho-proteins or total proteins. The upper panel of A–G show representative Western blots for BDNF, p-Akt, Bcl-2, Bax, caspase-3, p-CaMKII and p-CREB in thehippocampus of the indicated groups. The lower panel of A–G present the quantitative analysis of relative protein, which was performed by densitometry p-Akt expressionwas normalized to the loading of total Akt and p-CaMKII was normalized to the loading of total CaMKII, while p-CREB was normalized to the loading of total CREB and otherdata were normalized to the loading control �-actin or GAPDH. (Mean ± S.E.M, n = 6). ##p < 0.01 VPA vs. control; *p < 0.05, and **p < 0.01, VPA vs. DHA group.

7 euros
two3wrt
4
Doitita
vswmAtabmeAatfcRsdiiAfooSpicl
rptgalep(DnEtmch
6 J. Gao et al. / Int. J. Devl N
hat the ratio of p-CREB/CREB proteins in the VPA-treated groupas reduced compared to the controls (Fig. 6G). VPA-treated rat
ffspring with DHA supplementation at a dose of 150 mg/kg and00 mg/kg increased the ratios of p-CREB/CREB compared to thoseithout DHA supplementation. Importantly, levels of total CREB
emained unchanged in all VPA-treated rats with and without DHAreatment.
. Discussion
In this study, we investigated the impacts of different doses ofHA supplementation in an experimental autism animal modeln biochemical alterations, and cognition. We found that DHAntervention reduced the expression of apoptosis protein and pro-ected against the neuronal loss in the hippocampus. DHA can alsoncrease the expression of phosphorylated CaMKII and CREB pro-eins and enhance cognitive ability, which could reverse abnormalutism-like behaviors.
It appears that DHA supplementation is an effective drug inter-ention on treating the VPA-induced rat model of autism. DHAerved as structural components of neuronal cell membranes,hich is specifically enriched in the brain and essential for nor-al neurological function (Fedorova and Salem, 2006; Su, 2010).ltered fluidity of the neuronal membrane phospholipids affects
he structure of membrane-bound receptors, which, in turn, hasn effect on their function and activity (Yehuda et al., 1999). It haseen reported that DHA plays a significant role in maintaining opti-al membrane integrity and fluidity (Fatemi et al., 2005; Lillycrop
t al., 2005). The decreased levels of DHA were observed in bothSD children (Foley et al., 2012; Wiest et al., 2009) and VPA-inducedutism rats, which are consistent with our results. In addition, reac-ive oxygen species (ROS) is believed as being a potential factoror an elevated metabolism rate of DHA. ASD was also reported toause a significant increase in lipid peroxidation. Basal amounts ofOS can be quickly scavenged by endogenous antioxidant enzymesuch as SOD, GSH and GSH-PX, which play a role in lipid signaling inefense against oxidative stress (Ghezzo et al., 2013). Further, MDA
s the product of cell membrane lipid peroxidation, and the changen MDA content is one of indication for the degree of cell oxidation.dequate supply of DHA is therefore supposed to be indispensable
or maintaining optimal tissue function. We observed an increasedxidative stress by increased MDA in the brain of VPA-induced ratffspring, and DHA efficiently blocked the alteration. In this study,OD, GSH and GSH-PX were significantly elevated after DHA sup-lementation in VPA-treated rat offspring. This indicates that an
ncrease in DHA modifies the physical properties of the neuronalell membranes underlying neuroprotection against VPA injury, ateast, related to its antioxidant property.
We showed that VPA-treated rats demonstrated significanteductions in the phosphorylation of CaMKII and CREB in the hip-ocampus. CaMKII, a Ca2+-activated enzyme highly abundant inhe hippocampus, is activated by Ca2+/CaM binding and under-oes a rapid autophosphorylation at Thr-286. Increase in CaMKIIutophosphorylation by DHA could be essential for the long-asting enhancement in synaptic efficacy following a LTP (Yamagatat al., 2009). We found VPA-induced CaMKII and CREB dephos-horylation can be partially recovered by DHA supplementation150 mg/kg and 300 mg/kg), implying that the therapeutic effect ofHA on cognitive dysfunction might be due to improvements inormal synaptic transmission and plasticity (Arsenault et al., 2011;bert and Greenberg, 2013; Takeuchi et al., 2013). With our VPA-
reated animal model, we identified a novel mechanism that DHAight ameliorate the learning and memory function to improveognitive function through CaMKII/CREB signaling pathway in theippocampus. In the Morris water maze task, there were significant
cience 49 (2016) 67–78
differences between groups with and without DHA supplementa-tion to the VPA-treated offspring, suggesting that both 150 and300 mg/kg DHA supplementation enhance learning and memoryof the VPA-treated rat offspring.
Apoptosis is one of the main types of programmed cell death andinvolves a series of biochemical events leading to a characteristiccell morphology and death. Previously, it was reported that autis-tic brains showed a loss of pyramidal neurons and granule cells inthe hippocampus (Bauman and Kemper, 2005; Duong et al., 1986;Fatemi et al., 2000). Several studies also found that the activity ofBDNF-Akt-Bcl-2 anti-apoptotic signaling pathway is compromisedin autistic subjects (Sheikh et al., 2010). Bcl-2 is a membrane-bound protein that strongly inhibits apoptosis and enhances cellsurvival (Reed, 1997; Sasaki et al., 2006), and Akt was reported to beinvolved in promoting neuronal survival (Dudek et al., 1997). Acti-vated Akt stimulated the changes in Bcl-2 expression and showedanti-apoptosis effects (Pugazhenthi et al., 2000). BDNF is one of theneurotrophic factors that regulate the activities of Akt, and playsan important role in supporting neuronal survival and maintainingseveral neuronal activities. To further investigate the mechanism ofDHA supplementation for that rescue phenotypes in VPA-treatedrat offspring, we examined caspase-3, Bax, Bcl-2, Akt and BDNFexpression in the hippocampus in different groups of rats. Caspase-3 is a crucial executioner of apoptosis since it is either partially ortotally responsible for the proteolytic cleavage of many key pro-teins (Erhardt et al., 2001). Our studies showed that caspase-3 andBax were significantly increased, whereas Bcl-2 and Akt were sig-nificantly decreased in the VPA-treated group. Conversely, Bcl-2and Akt were significantly increased while caspase-3 and Bax weresignificantly decreased after treatment with higher dose of DHAsupplementation. Increased BDNF expression in the VPA-treatedgroup is postulated as a result of the body’s negative feedbackin this study. BDNF expression was further increased significantlyafter DHA supplementation at the dose of 150 and 300 mg/kg, sug-gesting that DHA may also influence the BDNF expression. Thesedemonstrate that BDNF-Akt-Bcl2 signaling pathway could be oneof the key (direct or indirect) targets of DHA supplementation totreat with the VPA-treated offspring.
Impairments in social interaction and communication alongwith restrictive and repetitive behaviors are the core symptomsof ASD (American Psychiatric Association, 2013). Some evidencesuggested that ASD patients are usually accompanied with the spa-tial learning and memory deficits (Bennetto et al., 1996; Salowitzet al., 2013; Steele et al., 2007). Moreover, several reports indicatedthat visuospatial information processing deficits may contributeto functional motor coordination deficits in autism and impairtheir ability to imitate, which is a real-life behavior important forsocial interaction and the development of language skills (Stoneet al., 1997; Rogers et al., 2003; Salowitz et al., 2013). So the spa-tial learning and memory deficits are correlated with poor socialresponsivity and expressive language skills in children with ASD.In this study, we found DHA supplementation improved the spatiallearning and memory ability of the VPA-induced rat autism model.Therefore, we hypothesize that DHA supplementation may play arole in spatial memory ability improvements in children with ASD,and in turn, affect the communication and social interaction skillsof them.
In conclusion, this is the first demonstration that DHA plays aneuroprotective role in ameliorating learning and memory impair-ments in autism animal model. Given that DHA is a molecule with avery low toxicity profile at virtually any dose and long-term usage(Rees et al., 2009), our study may provide further evidence that DHA
could be used as therapeutic intervention to mitigate behavioraldysfunctions in patients with ASD, mediated in part by increased
euros
Cp
C
A
ohM
R
A
A
A
A
B
B
B
C
C
D
D
D
E
E
E
E
F
F
F
F
G
G
J. Gao et al. / Int. J. Devl N
aMKII and CREB phosphorylation via BDNF-Akt-Bcl2 signalingathway.
onflict of interest
The authors declare no competing financial interests.
cknowledgements
This work was supported by the Natural Science Foundationf China (81273094). We are most grateful to Dr. Peng Wang forer kind cooperation in Research Center, Daqing Campus of Harbinedical University.
eferences
gostoni, C., Riva, E., Trojan, S., Bellù, R., Giovannini, M., 1995. Docosahexaenoicacid status and developmental quotient of healthy term infants. Lancet 346,638.
l-Farsi, Y.M., Waly, M.I., Deth, R.C., Al-Sharbati, M.M., Al-Shafaee, M., Al-Farsi, O.,Al-Khaduri, M.M., Al-Adawi, S., Hodgson, N.W., Gupta, I., Ouhtit, A., 2013.Impact of nutrition on serum levels of docosahexaenoic acid among Omanichildren with autism. Nutrition 29, 1142–1146.
merican Psychiatric Association, 2013. Diagnostic and statistical manual ofmental disorders, 5th ed. American Psychiatric Association, Washington, DC,pp. 50–59.
rsenault, D., Julien, C., Tremblay, C., Calon, F., 2011. DHA improves cognition andprevents dysfunction of entorhinal cortex neurons in 3xTg-AD mice. PLoS One6, e17397.
auman, M.L., Kemper, T.L., 2005. Neuroanatomic observations of the brain inautism: a review and future directions. Int. J. Dev. Neurosci. 23, 183–187.
ekaroglu, M., Asian, Y., Gedik, Y., Deger, O., Mocan, H., Erduran, E., Karahan, C.,1996. Relationships between serum free fatty acids and zinc, and attentiondeficit hyperactivity disorder: a research note. J. Child Psychol. Psychiatry 37,225–227.
ennetto, L., Pennington, B.F., Rogers, S.J., 1996. Intact and impaired memoryfunctions in autism. Child Dev. 67, 1816–1835.
DC (2012) Prevalence of autism spectrum disorders–Autism and DevelopmentalDisabilities Monitoring Network, 14 sites, United States, 2008. MMWR SurveillSumm 61, 1–19.
olangelo, L.A., He, K., Whooley, M.A., Daviglus, M.L., Liu, K., 2009. Higher dietaryintake of long-chain �-3 polyunsaturated fatty acids is inversely associatedwith depressive symptoms in women. Nutrition 25, 1011–1019.
as, U.N., 2004. Can perinatal supplementation of long-chain polyunsaturated fattyacids prevents schizophrenia in adult life? Med. Sci. Monit. 10, Hy33–Hy37.
udek, H., Datta, S.R., Franke, T.F., Birnbaum, M.J., Yao, R., Cooper, G.M., Segal, R.A.,Kaplan, D.R., Greenberg, M.E., 1997. Regulation of neuronal survival by theserine-threonine protein kinase Akt. Science 275, 661–665.
uong, T., Robinson, H., BADG, Ritvo, A., 1986. Lower Purkinje cell counts in thecerebella of four autistic subjects: initial findings of the UCLA-NSAC AutopsyResearch Report. Am. J. Psychiatry 143, 862–866.
bert, D.H., Greenberg, M.E., 2013. Activity-dependent neuronal signalling andautism spectrum disorder. Nature 493, 327–337.
dgin, J.O., Pennington, B.F., 2005. Spatial cognition in autism spectrum disorders:superior, impaired, or just intact? J. Autism Dev. Disord. 35, 729–745.
msley, R., Oosthuizen, P., van Rensburg, S.J., 2003. Clinical potential of omega-3fatty acids in the treatment of schizophrenia. CNS Drugs 17, 1081–1091.
rhardt, J.A., Ohlstein, E.H., Toomey, J.R., Gabriel, M.A., Willette, R.N., Yue, T.-L.,Barone, F.C., Parsons, A.A., 2001. Activation of caspase-3/caspase-3-like activityin rat cardiomyocytes by an RGD peptide, but not the GPIIb/IIIa antagonistlotrafiban. Thromb. Res. 103, 143–148.
atemi, S., Earle, J., McMenomy, T., 2000. Hippocampal CA4 reelin-positiveneurons. Mol. Psychiatry 5, 571.
atemi, S.H., Snow, A.V., Stary, J.M., Araghi-Niknam, M., Reutiman, T.J., Lee, S.,Brooks, A.I., Pearce, D.A., 2005. Reelin signaling is impaired in autism. Biol.Psychiatry 57, 777–787.
edorova, I., Salem Jr., N., 2006. Omega-3 fatty acids and rodent behavior.Prostaglandins Leukotri. Essent. Fatty Acids 75, 271–289.
oley, A.G., Gannon, S., Rombach-Mullan, N., Prendergast, A., Barry, C., Cassidy,A.W., Regan, C.M., 2012. Class I histone deacetylase inhibition amelioratessocial cognition and cell adhesion molecule plasticity deficits in a rodentmodel of autism spectrum disorder. Neuropharmacology 63, 750–760.
hezzo, A., Visconti, P., Abruzzo, P.M., Bolotta, A., Ferreri, C., Gobbi, G., Malisardi, G.,Manfredini, S., Marini, M., Nanetti, L., Pipitone, E., Raffaelli, F., Resca, F., Vignini,
A., Mazzanti, L., 2013. Oxidative stress and erythrocyte membrane alterationsin children with autism: correlation with clinical features. PLoS One 8, e66418.reen, P., Glozman, S., Kamensky, B., Yavin, E., 1999. Developmental changes in ratbrain membrane lipids and fatty acids: the preferential prenatal accumulationof docosahexaenoic acid. J. Lipid Res. 40, 960–966.
cience 49 (2016) 67–78 77
Horrocks, L.A., Farooqui, A.A., 2004. Docosahexaenoic acid in the diet: itsimportance in maintenance and restoration of neural membrane function.Prostaglandins Leukotri. Essent. Fatty Acids 70, 361–372.
Johnson, C., Handen, B., Zimmer, M., Sacco, K., 2010. Polyunsaturated fatty acidsupplementation in young children with autism. J. Dev. Phys. Disabil. 22, 1–10.
Lauritzen, L., Hansen, H.S., Jørgensen, M.H., Michaelsen, K.F., 2001. The essentialityof long chain n-3 fatty acids in relation to development and function of thebrain and retina. Prog. Lipid Res. 40, 1–94.
Lee, G.Y.-P., Brown, L.M., Teyler, T.J., 1996. The effects of anticonvulsant drugs onlong-term potentiation (LTP) in the rat hippocampus. Brain Res. Bull. 39, 39–42.
Li, J., Liu, W., Ding, S., Xu, W., Guan, Y., Zhang, J.H., Sun, X., 2008. Hyperbaric oxygenpreconditioning induces tolerance against brain ischemia–reperfusion injuryby upregulation of antioxidant enzymes in rats. Brain Res. 1210, 223–229.
Lillycrop, K.A., Phillips, E.S., Jackson, A.A., Hanson, M.A., Burdge, G.C., 2005. Dietaryprotein restriction of pregnant rats induces and folic acid supplementationprevents epigenetic modification of hepatic gene expression in the offspring. J.Nutri. 135, 1382–1386.
Maenner, M.J., Durkin, M.S., 2010. Trends in the prevalence of autism on the basisof special education data. Pediatrics 126, e1018–e1025.
Malone, J.P., 2012. The systems theory of autistogenesis. SAGE Open 2, 25.Martinez, M., 1992. Tissue levels of polyunsaturated fatty acids during early
human development. J. Pediatr. 120, S129–S138.McNamara, R.K., Carlson, S.E., 2006. Role of omega-3 fatty acids in brain
development and function: potential implications for the pathogenesis andprevention of psychopathology. Prostaglandins Leukotri. Essent. Fatty Acids75, 329–349.
Meador, K.J., Baker, G.A., Browning, N., Clayton-Smith, J., Combs-Cantrell, D.T.,Cohen, M., Kalayjian, L.A., Kanner, A., Liporace, J.D., Pennell, P.B., 2009.Cognitive function at 3 years of age after fetal exposure to antiepileptic drugs.N. Engl. J. Med. 360, 1597–1605.
Meguid, N.A., Dardir, A.A., Abdel-Raouf, E.R., Hashish, A., 2011. Evaluation ofoxidative stress in autism: defective antioxidant enzymes and increased lipidperoxidation. Biol. Trace Elem. Res. 143, 58–65.
Polleux, F., Lauder, J.M., 2004. Toward a developmental neurobiology of autism.Ment. Retard. Dev. Disabil. Res. Rev. 10, 303–317.
Pugazhenthi, S., Nesterova, A., Sable, C., Heidenreich, K.A., Boxer, L.M., Heasley, L.E.,Reusch, J.E.-B., 2000. Akt/protein kinase B up-regulates Bcl-2 expressionthrough cAMP-response element-binding protein. J. Biol. Chem. 275,10761–10766.
Reed, J.C., 1997. Double identity for proteins of the Bcl-2 family. Nature 387,773–776.
Rees, A.-M., Austin, M.-P., Owen, C., Parker, G., 2009. Omega-3 deficiency associatedwith perinatal depression: case control study. Psychiatry Res. 166, 254–259.
Rogers, S.J., Hepburn, S.L., Stackhouse, T., Wehner, E., 2003. Imitation performancein toddlers with autism and those with other developmental disorders. J. ChildPsychol. Psychiatry 44, 763–781.
Rutter, M., 2005. Autism research: lessons from the past and prospects for thefuture. J. Autism Dev. Disord. 35, 241–257.
Salowitz, N.M., Eccarius, P., Karst, J., Carson, A., Schohl, K., Stevens, S., Van Hecke,A.V., Scheidt, R.A., 2013. Brief report: visuo-spatial guidance of movementduring gesture imitation and mirror drawing in children with autism spectrumdisorders. J. Autism Dev. Disord. 43, 985–995.
Sasaki, T., Kitagawa, K., Yagita, Y., Sugiura, S., Omura-Matsuoka, E., Tanaka, S.,Matsushita, K., Okano, H., Tsujimoto, Y., Hori, M., 2006. Bcl2 enhances survivalof newborn neurons in the normal and ischemic hippocampus. J. Neurosci. Res.84, 1187–1196.
Schneider, T., Przewłocki, R., 2005. Behavioral alterations in rats prenatallyexposed to valproic acid: animal model of autism. Neuropsychopharmacology30, 80–89.
Sheikh, A.M., Malik, M., Wen, G., Chauhan, A., Chauhan, V., Gong, C.X., Liu, F.,Brown, W.T., Li, X., 2010. BDNF-Akt-Bcl2 antiapoptotic signaling pathway iscompromised in the brain of autistic subjects. J. Neurosci. Res. 88, 2641–2647.
Steele, S.D., Minshew, N.J., Luna, B., Sweeney, J.A., 2007. Spatial working memorydeficits in autism. J. Autism Dev. Disord. 37, 605–612.
Stone, W.L., Ousley, O.Y., Littleford, C.D., 1997. Motor imitation in young childrenwith autism: what’s the object? J. Abnorm. Child Psychol. 25, 475–485.
Strickland, A.D., 2014. Prevention of cerebral palsy, autism spectrum disorder, andattention deficit–Hyperactivity disorder. Med. Hypotheses 82, 522–528.
Su, H.-M., 2010. Mechanisms of n-3 fatty acid-mediated development andmaintenance of learning memory performance. J. Nutri. Biochem. 21, 364–373.
Takeuchi, K., Gertner, M.J., Zhou, J., Parada, L.F., Bennett, M.V., Zukin, R.S., 2013.Dysregulation of synaptic plasticity precedes appearance of morphologicaldefects in a Pten conditional knockout mouse model of autism. Proc. Natl.Acad. Sci. U. S. A. 110, 4738–4743.
Van Elst, K., Bruining, H., Birtoli, B., Terreaux, C., Buitelaar, J.K., Kas, M.J., 2014. Foodfor thought: Dietary changes in essential fatty acid ratios and the increase inautism spectrum disorders. Neurosci. Biobehav. Rev. 45, 369–378.
Vorhees, C.V., 1987. Teratogenicity and developmental toxicity of valproic acid inrats. Teratology 35, 195–202.
Wang, Han, Liang, Shuang, Wang, Maoqing, et al., 2016. Potential serumbiomarkers from a metabolomics study of autism. J. Psychiatry Neurosci. 41
(January (1)), 27–37.Wiest, M., German, J., Harvey, D., Watkins, S., Hertz-Picciotto, I., 2009. Plasma fattyacid profiles in autism: a case-control study. Prostaglandins Leukotri. Essent.Fatty Acids 80, 221–227.

7 euros
W
Y
Y
autism spectrum disorders. Seishin Shinkeigaku Zasshi 114, 934–940.Yui, K., Koshiba, M., Nakamura, S., Onishi, M., 2011. Therapeutic effects of larger
8 J. Gao et al. / Int. J. Devl N
u, P., Jiang, L., Chen, H., 2010. Sodium valproate at the therapeutic concentrationinhibits the induction but not the maintenance phase of long-term potentiationin rat hippocampal CA1 area. Biochem. Biophys. Res. Commun. 391, 582–586.
amagata, Y., Kobayashi, S., Umeda, T., Inoue, A., Sakagami, H., Fukaya, M.,Watanabe, M., Hatanaka, N., Totsuka, M., Yagi, T., 2009. Kinase-dead knock-in
mouse reveals an essential role of kinase activity ofCa2+/calmodulin-dependent protein kinase II� in dendritic spine enlargement,long-term potentiation, and learning. J. Neurosci. 29, 7607–7618.ehuda, S., Rabinovitz, S., Mostofsky, D.I., 1999. Essential fatty acids are mediatorsof brain biochemistry and cognitive functions. J. Neurosci. Res. 56, 565–570.
cience 49 (2016) 67–78
Yochum, C.L., Dowling, P., Reuhl, K.R., Wagner, G.C., Ming, X., 2008. VPA-inducedapoptosis and behavioral deficits in neonatal mice. Brain Res. 1203, 126–132.
Yui, K., 2012. Useful pharmacologic treatment in impaired social interaction in
doses of arachidonic acid added to DHA on social impairment and its relationto alterations of polyunsaturated fatty acids in individuals with autismspectrum disorders. Nihon Shinkei Seishin Yakurigaku Zasshi 31, 117–124.




![Preparation of titania/hydroxyapatite (TiO2/HAp) composite ...or.nsfc.gov.cn/bitstream/00001903-5/77915/1/1000007277269.pdf · Pentachlorophenol (PCP) ... synthesis of TiO ... [17]](https://img.pdfslide.net/doc/110x75/5a7333487f8b9a9d538e6543/preparation-of-titaniahydroxyapatite-tio2hap-composite-ornsfcgovcnbitstream00001903-5779151.jpg)