Embed Size (px)
Citation preview


Inverse Folding and Sequence-StructureMaps of Ribonucleic Acids
Peter SchusterInstitut für Theoretische Chemie und Molekulare
Strukturbiologie der Universität Wien
Inverse Problem Workshop
IPAM, UCLA, 22.10.2003

Web-Page for further information:
http://www.tbi.univie.ac.at/~pks

1. The role of RNA in the cell and the notion of structure
2. RNA folding
3. Inverse folding of RNA
4. Sequence structure maps, neutral networks, and intersection
5. Reference to experimental data
6. Concluding remarks

1. The role of RNA in the cell and the notion of structure
2. RNA folding
3. Inverse folding of RNA
4. Sequence structure maps, neutral networks, and intersection
5. Reference to experimental data
6. Concluding remarks

RNA
RNA as scaffold for supramolecular complexes
ribosome? ? ? ? ?
RNA as adapter molecule
GA
C...
CU
G ..
.
leu
genetic code
RNA as transmitter of genetic information
DNA
...AGAGCGCCAGACUGAAGAUCUGGAGGUCCUGUGUUC...
messenger-RNA
protein
transcription
translation
RNA as of genetic informationworking copy
RNA as carrier of genetic information
RNA
RNA
viruses and retroviruses
as information carrier inevolution and
evolutionary biotechnologyin vitro
RNA as catalyst
ribozyme
The RNA
DNA protein
world as a precursor of the current + biology
RNA as regulator of gene expression
gene silencing by small interfering RNAs
RNA is modified by epigenetic control
RNA
RNA
editing
Alternative splicing of messenger
RNA is the catalytic subunit in supramolecular complexes
Functions of RNA molecules

OCH2
OHO
O
PO
O
O
N1
OCH2
OHO
PO
O
O
N2
OCH2
OHO
PO
O
O
N3
OCH2
OHO
PO
O
O
N4
N A U G Ck = , , ,
3' - end
5' - end
NaØ
NaØ
NaØ
NaØ
RNA
nd 3’-endGCGGAU AUUCGCUUA AGUUGGGA G CUGAAGA AGGUC UUCGAUC A ACCAGCUC GAGC CCAGA UCUGG CUGUG CACAG
3'-end
5’-end
70
60
50
4030
20
10
Definition of RNA structure
5'-e

RNA sequence
Empirical parameters
Biophysical chemistry: thermodynamics and
kinetics
RNA structure
Inverse folding of RNA:
Biotechnology,design of biomolecules
with predefined structures and functions
RNA folding:
Structural biology,spectroscopy of biomolecules, understanding
molecular function
Sequence, structure, and function

Definition and physical relevance of RNA secondary structures
RNA secondary structures are listings of Watson-Crick and GU wobble base pairs, which are free of knots and pseudokots.
„Secondary structures are folding intermediates in the formation of full three-dimensional structures.“
D.Thirumalai, N.Lee, S.A.Woodson, and D.K.Klimov. Annu.Rev.Phys.Chem. 52:751-762 (2001):

5'-End
5'-End 3'-End
3'-End
70
60
50
4030
20
10
GCGGAUUUAGCUCAGDDGGGAGAGCMCCAGACUGAAYAUCUGGAGMUCCUGUGTPCGAUCCACAGAAUUCGCACCASequence
Secondary structure
The RNA secondary structure lists the double helical stretches or stacks of a folded single strand molecule

The three-dimensional structure of a short double helical stack of B-DNA
James D. Watson, 1928- , and Francis Crick, 1916- ,Nobel Prize 1962
1953 – 2003 fifty years double helix

Canonical Watson-Crick base pairs:
cytosine – guanineuracil – adenine
W.Saenger, Principles of Nucleic Acid Structure, Springer, Berlin 1984

5'-End
5'-End 3'-End
3'-End
70
60
50
4030
20
10
GCGGAUUUAGCUCAGDDGGGAGAGCMCCAGACUGAAYAUCUGGAGMUCCUGUGTPCGAUCCACAGAAUUCGCACCASequence
Secondary structure

5'-End
5'-End
5'-End
3'-End
3'-End
3'-End
70
60
50
4030
20
10
GCGGAUUUAGCUCAGDDGGGAGAGCMCCAGACUGAAYAUCUGGAGMUCCUGUGTPCGAUCCACAGAAUUCGCACCASequence
Secondary structure
Symbolic notation
Í
A symbolic notation of RNA secondary structure that is equivalent to the conventional graphs

Tertiary elements in RNA structure
1. Different classes of pseudoknots
2. Different classes of non-Watson-Crick base pairs
3. Base triplets, G-quartets, A-platforms, etc.
4. End-on-end stacking of double helices
5. Divalent metal ion complexes, Mg2+, etc.
6. Other interactions involving phosphate, 2‘-OH, etc.

Tertiary elements in RNA structure
1. Different classes of pseudoknots
2. Different classes of non-Watson-Crick base pairs
3. Base triplets, G-quartets, A-platforms, etc.
4. End-on-end stacking of double helices
5. Divalent metal ion complexes, Mg2+, etc.
6. Other interactions involving phosphate, 2‘-OH, etc.

3'-end
"H-type pseudoknot"
5'-end
3'-end
pseudoknot
"Kissing loops"
5'-end
·· (((( ····· [[ ·)))) ···· ((((( · ]] ·····))))) ···
Two classes of pseudoknots in RNA structures

Tertiary elements in RNA structure
1. Different classes of pseudoknots
2. Different classes of non-Watson-Crick base pairs
3. Base triplets, G-quartets, A-platforms, etc.
4. End-on-end stacking of double helices
5. Divalent metal ion complexes, Mg2+, etc.
6. Other interactions involving phosphate, 2‘-OH, etc.

Twelve families of base pairsWatson-Crick / Hogsteen / Sugar
edgeCis / Transorientation
N.B. Leontis, E. Westhof, Geometric nomenclature and classification of RNA base pairs. RNA 7:499-512, 2001.

Tertiary elements in RNA structure
1. Different classes of pseudoknots
2. Different classes of non-Watson-Crick base pairs
3. Base triplets, G-quartets, A-platforms, etc.
4. End-on-end stacking of double helices
5. Divalent metal ion complexes, Mg2+, etc.
6. Other interactions involving phosphate, 2‘-OH, etc.

5'-End
3'-End7060
50
4030
20
10
5'-End
3'-End
70
60
50
4030
20
10
End-on-end stacking of double helical regions yields the L-shape of tRNAphe

1. The role of RNA in the cell and the notion of structure
2. RNA folding
3. Inverse folding of RNA
4. Sequence structure maps, neutral networks, and intersection
5. Reference to experimental data
6. Concluding remarks

How to compute RNA secondary structures
Efficient algorithms based on dynamic programming are available for computation of minimum free energy and many suboptimal secondary structures for given sequences.
M.Zuker and P.Stiegler. Nucleic Acids Res. 9:133-148 (1981)
M.Zuker, Science 244: 48-52 (1989)
Equilibrium partition function and base pairing probabilities in Boltzmann ensembles of suboptimal structures.J.S.McCaskill. Biopolymers 29:1105-1190 (1990)
The Vienna RNA Package provides in addition: inverse folding (computing sequences for given secondary structures), computation of melting profiles from partitionfunctions, all suboptimal structures within a given energy interval, barrier tress of suboptimal structures, kinetic folding of RNA sequences, RNA-hybridization and RNA/DNA-hybridization through cofolding of sequences, alignment, etc.. I.L.Hofacker, W. Fontana, P.F.Stadler, L.S.Bonhoeffer, M.Tacker, and P. Schuster. Mh.Chem.125:167-188 (1994)
S.Wuchty, W.Fontana, I.L.Hofacker, and P.Schuster. Biopolymers 49:145-165 (1999)
C.Flamm, W.Fontana, I.L.Hofacker, and P.Schuster. RNA 6:325-338 (1999)
Vienna RNA Package: http://www.tbi.univie.ac.at

G
GG
G
GG
G GG
G
GG
G
GG
G
U
U
U
U
UU
U
U
U
UU
A
A
AA
AA
AA
A
A
A
A
UC
C
CC
C
C
C
C
C
CC
C
5’-end 3’-end
Folding of RNA sequences into secondary structures of minimal free energy, DG0300

G
GG
G
GG
G GG
G
GG
G
GG
G
U
U
U
U
UU
U
U
U
UU
A
A
AA
AA
AA
A
A
A
A
UC
C
CC
C
C
C
C
C
CC
C
5’-end 3’-end
i j
k l
Edges: i·j,k·l á S .... base pairs
(i) i· i+1 á S .... backbone(ii) #base pairs per node = {0,1}(iii) if i·j and l·k á S, then
i<k<j ñ i<l<j ....pseudoknot exclusion
Folding of RNA sequences into secondary structures of minimal free energy, DG0300

G
GG
G
GG
G GG
G
GG
G
GG
G
U
U
U
U
UU
U
U
U
UU
A
A
AA
AA
AA
A
A
A
A
UC
C
CC
C
C
C
C
C
CC
C
5’-end 3’-end
free energy of stacking < 0
L∑∑∑∑ ++++=∆
loopsinternalbulges
loopshairpin
pairsbaseofstacks
,3000 )()()( iblklij ninbnhgG
Folding of RNA sequences into secondary structures of minimal free energy, DG0300

hairpinloop
hairpinloop
stack
stack
stack
hairpin loop
stack
free end
freeend
freeend
hairpin loop
hairpinloop
stack
stack
free end
freeend joint
hairpin loop
stackstack
stack
internal loop
bulgem
ultiloop
Elements of RNA secondary structures as used in free energy calculations

Maximum matching
An example of a dynamic programming computation of the maximum number of base pairs
Back tracking yields the structure(s).
i i+1 i+2 k
Xi,k-1
j-1 j
Xk+1,j
j+1
[ k+1,j ][i,k-1]
( ){ }1,,11,1,1, )1(max,max ++−−≤≤+ ++= jkjkkijkijiji XXXX ρ
Minimum free energy computations are based on empirical energies
GGCGCGCCCGGCGCC
GUAUCGAAAUACGUAGCGUAUGGGGAUGCUGGACGGUCCCAUCGGUACUCCA
UGGUUACGCGUUGGGGUAACGAAGAUUCCGAGAGGAGUUUAGUGACUAGAGGRNAStudio.lnk

Maximum matching j 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15i G G C G C G C C C G G C G C C1 G * * 1 1 1 1 2 3 3 3 4 4 5 6 62 G * * 0 1 1 2 2 2 3 3 4 4 5 63 C * * 0 1 1 1 2 3 3 3 4 5 54 G * * 0 1 1 2 2 2 3 4 5 55 C * * 0 1 1 2 2 3 4 4 46 G * * 1 1 1 2 3 3 3 47 C * * 0 1 2 2 2 2 38 C * * 1 1 1 2 2 29 C * * 1 1 2 2 2
10 G * * 1 1 1 211 G * * 0 1 112 C * * 0 113 G * * 114 C * *15 C *
An example of a dynamic programming computation of the maximum number of base pairs
Back tracking yields the structure(s).
i i+1 i+2 k
Xi,k-1
j-1 j
Xk+1,j
j+1
[ k+1,j ][i,k-1]
( ){ }1,,11,1,1, )1(max,max ++−−≤≤+ ++= jkjkkijkijiji XXXX ρ
Minimum free energy computations are based on empirical energies
GGCGCGCCCGGCGCC
GUAUCGAAAUACGUAGCGUAUGGGGAUGCUGGACGGUCCCAUCGGUACUCCA
UGGUUACGCGUUGGGGUAACGAAGAUUCCGAGAGGAGUUUAGUGACUAGAGGRNAStudio.lnk

S1(h)
S9(h)
Free
ene
rgy
GD
0
Minimum of free energy
Suboptimal conformations
S0(h)
S2(h)
S3(h)
S4(h)
S7(h)
S6(h)
S5(h)
S8(h)
G
GG
G
GG
G GG
G
GG
G
GG
G
U
U
U
U
UU
U
U
U
UU
A
A
AA
AA
AA
A
A
A
A
UC
C
CC
C
C
C
C
C
CC
C
5’-end 3’-end
The minimum free energy structures on a discrete space of conformations

Free
ene
rgy
GD
0
"Reaction coordinate"
Sk
S{Saddle point T {k
Free
ene
rgy
GD
0
Sk
S{
T {k
"Barrier tree"
Definition of a ‚barrier tree‘

5.10
2
8
1415
1817
23
19
2722
38
45
25
3633
3940
4341
3.30
7.40
5
3
7
4
109
6
1312
3.10
11
2120
16
2829
26
3032
424644
24
353437
49
31
4748
S0S1
Kinetic folding
S0
S1
S2
S3S4S5 S6
S7S8
S10S9
Suboptimal structures
lim t Á¸ finite folding time
5.90
A typical energy landscape of a sequence with two (meta)stable comformations

Kinetics RNA refolding between a long living metastable conformation and the minmum free energy structure

1. The role of RNA in the cell and the notion of structure
2. RNA folding
3. Inverse folding of RNA
4. Sequence structure maps, neutral networks, and intersection
5. Reference to experimental data
6. Concluding remarks

Minimum free energycriterion
Inverse folding of RNA secondary structures
The idea of inverse folding algorithm is to search for sequences that form a given RNA secondary structure under the minimum free energy criterion.

Structure

CUGGGAAAAAUCCCCAGACCGGGGGUUUCCCCGG
Compatible sequenceStructure
5’-end
3’-end

CUGGGAAAAAUCCCCAGACCGGGGGUUUCCCCGG
G
G
G
G
G
G
G
C
C
C
C G
G
G
G
C
C
C
C
C
C
C
U A
U
U
G
U
AA
A
A
U
Compatible sequenceStructure
5’-end
3’-end

CUGGGAAAAAUCCCCAGACCGGGGGUUUCCCCGG
G
G
G
G
G
G
G
C
C
C
C
U
U
G
G
G
G
G
C
C
C
C
C
C
C
U
U
AA
A
A
A
U
Compatible sequenceStructure
5’-end
3’-end
Single nucleotides: A U G C, , ,
Single bases pairs are varied independently

CUGGGAAAAAUCCCCAGACCGGGGGUUUCCCCGG
G
G
C
C
C
C G
G
G
G
C
C
G
G
G
G
G
C
C
C
C
C
U A
U
U
G
U
AA
A
A
U
Compatible sequenceStructure
5’-end
3’-end
Base pairs: AU , UAGC , CGGU , UG
Base pairs are varied in strict correlation

CUGGGAAAAAUCCCCAGACCGGGGGUUUCCC
GG
U
CUGGGAAAAAUCCCCAGACCGGGGGUUUCCCCGG
G G
G G
G G
G G
G G
G G
G G
C U
C C
C C
C C
U U
U U
G G
G G
G G
G G
G G
C C
C C
C C
C C
C C
C C
C C
U U
U U
A AA A
A A
A A
A A
U U
Compatible sequencesStructure
5’-end 5’-end
3’-end 3’-end

CUGGGAAAAAUCCCCAGACCGGGGGUUUCCGCGG
G
G
G
G
G
G
G
CG
C
C
U
U
G
G
GG
G
C
C
C
C
C
C
C
U
U
AA
A
A
A
U
Structure Incompatible sequence
5’-end
3’-end

.... GC UC ....CA
.... GC UC ....GU
.... GC UC ....GA .... GC UC ....CU
d =1H
d =1H
d =2H
City-block distance in sequence space 2D Sketch of sequence space
Single point mutations as moves in sequence space

0
421 8 16
10
19
9
14
6
13
5
11
3
7
12
21
17
22
18
25
20
26
24
28
272315 29 30
31
Binary sequences are encodedby their decimal equivalents:
= 0 and = 1, for example,
"0" 00000 =
"14" 01110 = ,
"29" 11101 = , etc.
≡
≡
≡
,
C
CCCCC
C C
C
G
GGG
GGG G
Mutant class
0
1
2
3
4
5
Hypercube of dimension n = 5 Decimal coding of binary sequences
Sequence space of binary sequences of chain lenght n = 5

CGTCGTTACAATTTA GTTATGTGCGAATTC CAAATT AAAA ACAAGAG.....
CGTCGTTACAATTTA GTTATGTGCGAATTC CAAATT AAAA ACAAGAG.....
G A G T
A C A C
Hamming distance d (I ,I ) = H 1 2 4
d (I ,I ) = 0H 1 1
d (I ,I ) = d (I ,I )H H1 2 2 1
d (I ,I ) d (I ,I ) + d (I ,I )H H H1 3 1 2 2 3¶
(i)
(ii)
(iii)
The Hamming distance between sequences induces a metric in sequence space

Target structure Sk
Initial trial sequences
Target sequence
Stop sequence of anunsuccessful trial
Intermediate compatible sequences
Approach to the target structure Sk in the inverse folding algorithm

Inverse folding algorithm
I0 Á I1 Á I2 Á I3 Á I4Á ... Á Ik Á Ik+1 Á ... Á It
S0Á S1Á S2Á S3Á S4Á ... Á SkÁ Sk+1Á ... Á St
Ik+1 = Mk(Ik) and DdS(Sk,Sk+1) = dS(Sk+1,St) - dS(Sk,St) < 0
M ... base or base pair mutation operator
dS (Si,Sj) ... distance between the two structures Si and Sj
‚Unsuccessful trial‘ ... termination after n steps

Minimum free energycriterion
Inverse folding of RNA secondary structures
1st2nd3rd trial4th5th
The inverse folding algorithm searches for sequences that form a given RNA secondary structure under the minimum free energy criterion.

1. The role of RNA in the cell and the notion of structure
2. RNA folding
3. Inverse folding of RNA
4. Sequence structure maps, neutral networks, and intersection
5. Reference to experimental data
6. Concluding remarks

Minimal hairpin loop size:
nlp Æ 3
Minimal stack length:nst Æ 2
Recursion formula for the number of acceptable RNA secondary structures

Computed numbers of minimum free energy structures over different nucleotide alphabets
P. Schuster, Molecular insights into evolution of phenotypes. In: J. Crutchfield & P.Schuster, Evolutionary Dynamics. Oxford University Press, New York 2003, pp.163-215.

Hamming distance d (S ,S ) = H 1 2 4
d (S ,S ) = 0H 1 1
d (S ,S ) = d (S ,S )H H1 2 2 1
d (S ,S ) d (S ,S ) + d (S ,S )H H H1 3 1 2 2 3¶
(i)
(ii)
(iii)
The Hamming distance between structures in parentheses notation forms a metric in structure space

RNA sequences as well as RNA secondary structures can be visualized as objects in metric spaces. At constant chain length the sequence space is a (generalized) hypercube.
The mapping from RNA sequences into RNA secondary structures is many-to-one. Hence, it is redundant and not invertible.
RNA sequences, which are mapped onto the same RNA secondary structure, are neutral with respect to structure. The pre-images of structures in sequence space are neutral networks. They can be represented by graphs where the edgesconnect sequences of Hamming distance dH = 1.


Sk I. = ( )ψfk f Sk = ( )
Sequence space Structure space Real numbers
Mapping from sequence space into structure space and into function

Sk I. = ( )ψfk f Sk = ( )
Sequence space Structure space Real numbers

Sk I. = ( )ψfk f Sk = ( )
Sequence space Structure space Real numbers
The pre-image of the structure Sk in sequence space is the neutral network Gk

Neutral networks are sets of sequences forming the same structure. Gk is the pre-image of the structure Sk in sequence space:
Gk = y-1(Sk) π {yj | y(Ij) = Sk}
The set is converted into a graph by connecting all sequences of Hamming distance one.
Neutral networks of small RNA molecules can be computed by exhaustive folding of complete sequence spaces, i.e. all RNA sequences of a given chain length. This number, N=4n , becomes very large with increasing length, and is prohibitive for numerical computations.
Neutral networks can be modelled by random graphs in sequence space. In this approach, nodes are inserted randomly into sequence space until the size of the pre-image, i.e. the number of neutral sequences, matches the neutral network to be studied.

Random graph approach to neutral networks
Sketch of sequence spaceStep 00

Random graph approach to neutral networks
Sketch of sequence spaceStep 01

Random graph approach to neutral networks
Sketch of sequence spaceStep 02

Random graph approach to neutral networks
Sketch of sequence spaceStep 03

Random graph approach to neutral networks
Sketch of sequence spaceStep 04

Random graph approach to neutral networks
Sketch of sequence spaceStep 05

Random graph approach to neutral networks
Sketch of sequence spaceStep 10

Random graph approach to neutral networks
Sketch of sequence spaceStep 15

Random graph approach to neutral networks
Sketch of sequence spaceStep 25

Random graph approach to neutral networks
Sketch of sequence spaceStep 50

Random graph approach to neutral networks
Sketch of sequence spaceStep 75

Random graph approach to neutral networks
Sketch of sequence spaceStep 100

λj = 27 = 0.444 ,/12 λk = ø l (k)j
| |Gk
λ κ cr = 1 - -1 ( 1)/ κ-
λ λk cr . . . .>
λ λk cr . . . .<
network is connectedGk
network is connectednotGk
Connectivity threshold:
Alphabet size : = 4k ñ kAUGC
G S Sk k k= ( ) | ( ) = y y-1 U { }I Ij j
k lcr
2 0.5
3 0.423
4 0.370
GC,AU
GUC,AUG
AUGC
Mean degree of neutrality and connectivity of neutral networks

A connected neutral network

Giant Component
A multi-component neutral network

Reference for postulation and in silico verification of neutral networks

GkNeutral Network
Structure S k
Gk Cà k
Compatible Set Ck
The compatible set Ck of a structure Sk consists of all sequences which form Sk as its minimum free energy structure (the neutral network Gk) or one of itssuboptimal structures.
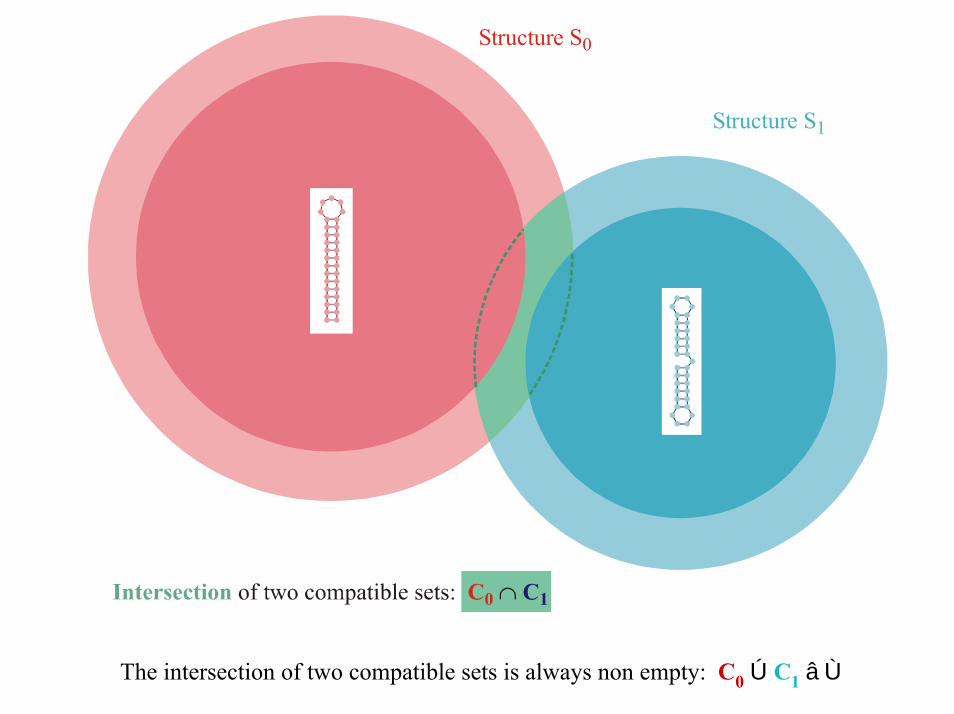
Structure S 0
Structure S 1
The intersection of two compatible sets is always non empty: C0 Ú C1 â Ù

Reference for the definition of the intersection and the proof of the intersection theorem
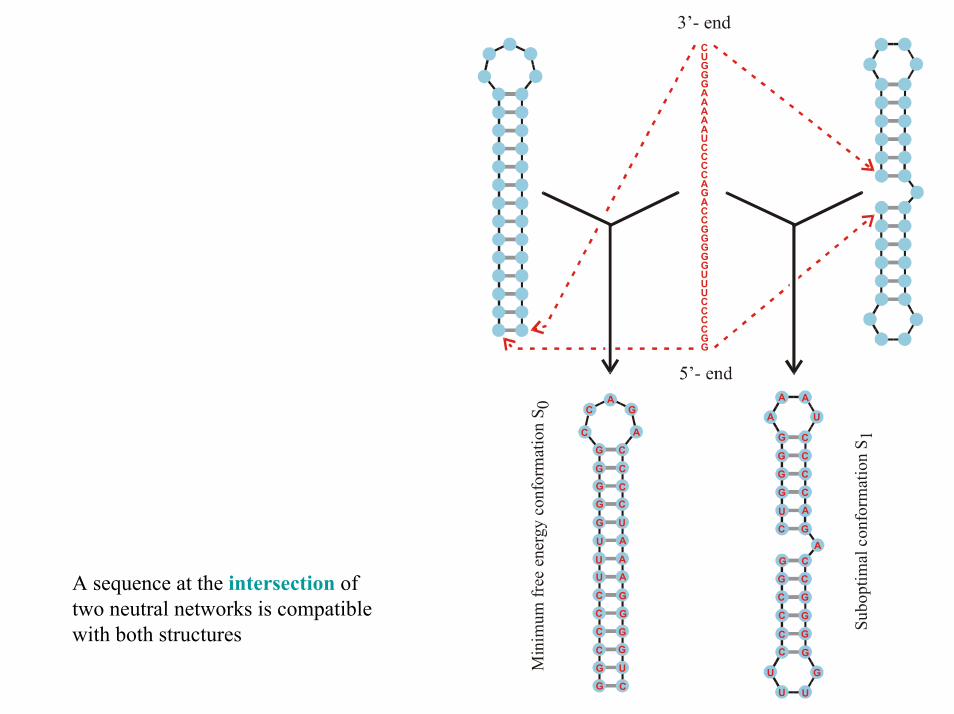
CUGGGAAAAAUCCCCAGACCGGGGGUUUCCCCGG
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
C
C
C
C
C
C
C
C
UU
UU
U
U
G
G
G
G
G
C
C
C
C
C
C
C
CC
C
C
C
CU
UU
AA
A
AA
A
A
A
A
A
U
3’- end
Min
imum
free
ene
rgy
conf
orm
atio
n S 0
Subo
ptim
al c
onfo
rmat
ion
S 1
C
G
A sequence at the intersection of two neutral networks is compatible with both structures

5.10
5.902
8
1415
1817
23
19
2722
38
45
25
3633
3940
4341
3.30
7.40
5
3
7
4
109
6
1312
3.10
11
2120
16
2829
26
3032
424644
24
353437
49
31
4748
S0S1
basin '1'
long livingmetastable structure
basin '0'
minimum free energy structure
Barrier tree for two long living structures


1. The role of RNA in the cell and the notion of structure
2. RNA folding
3. Inverse folding of RNA
4. Sequence structure maps, neutral networks, and intersection
5. Reference to experimental data
6. Concluding remarks

A ribozyme switch
E.A.Schultes, D.B.Bartel, Science 289 (2000), 448-452

Two ribozymes of chain lengths n = 88 nucleotides: An artificial ligase (A) and a natural cleavage ribozyme of hepatitis-d-virus (B)

The sequence at the intersection:
An RNA molecules which is 88 nucleotides long and can form both structures

Two neutral walks through sequence space with conservation of structure and catalytic activity

Nature , 323-325, 1999402
Catalytic activity in the AUG alphabet

Nature , 841-844, 2002420
Catalytic activity in theDU alphabet
5'-End
3'-End
70
60
50
4030
20
10
RNA clover-leaf secondary structures of sequences with chain length n=76
tRNAphe

Target structure Sk
Initial trial sequences
Target sequence
Stop sequence of anunsuccessful trial
Intermediate compatible sequences
Approach to the target structure Sk in the inverse folding algorithm
5'-End5'-End 5'-End5'-End
3'-End3'-End 3'-End3'-End
7070 7070
60606060
5050 5050
4040 4040 3030 3030
2020 20
20
1010 1010
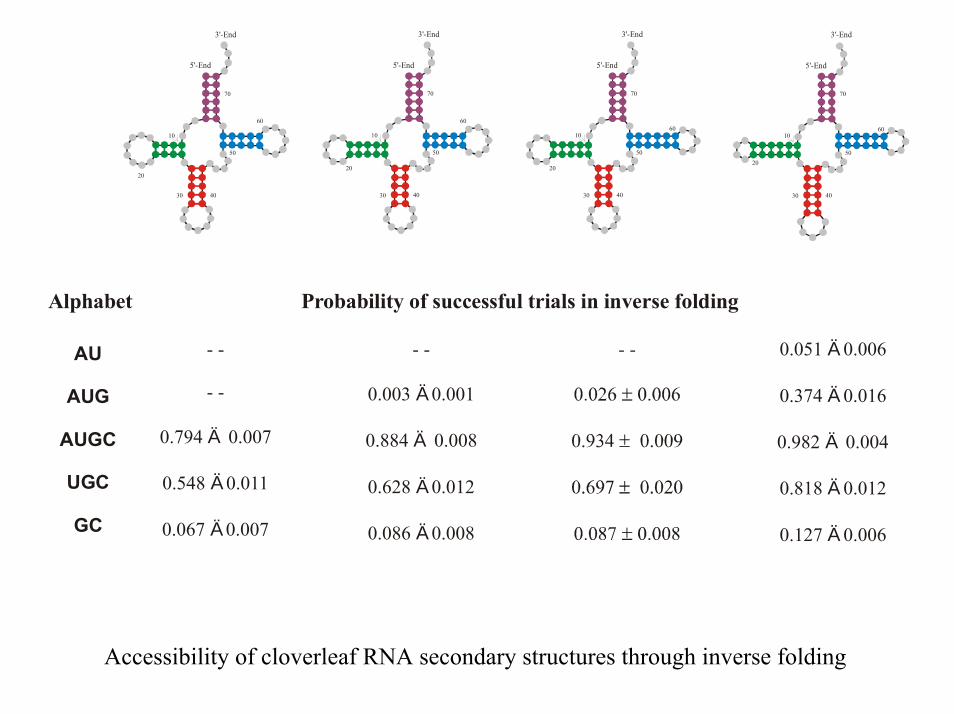
Alphabet Probability of successful trials in inverse folding
AU
AUG
AUGC
UGC
GC
- -
- -
0.794 0.007
0.548 0.011
0.067 0.007
Ä
Ä
Ä
- -
0.003 0.001
0.884 0.008
0.628 0.012
Ä
Ä
Ä
0.086 0.008Ä
0.051 0.006
0.374 0.016
0.982 0.004
0.818 0.012
0.127 0.006
Ä
Ä
Ä
Ä
Ä
Accessibility of cloverleaf RNA secondary structures through inverse folding

5'-End5'-End 5'-End5'-End
3'-End3'-End 3'-End3'-End
7070 7070
60606060
5050 5050
4040 4040 3030 3030
2020 20
20
1010 1010
Alphabet Degree of neutrality l
AU
AUG
AUGC
UGC
GC
- -
- -
0.275 0.064
0.263 0.071
0.052 0.033
Ä
Ä
Ä
- -
0.217 0.051
0.279 0.063
0.257 0.070
Ä
Ä
Ä
0.057 0.034Ä
0.073 0.032
0.201 0.056
0.313 0.058
0.250 0.064
0.068 0.034
Ä
Ä
Ä
Ä
Ä
`
Degree of neutrality of cloverleaf RNA secondary structures over different alphabets

1. The role of RNA in the cell and the notion of structure
2. RNA folding
3. Inverse folding of RNA
4. Sequence structure maps, neutral networks, and intersection
5. Reference to experimental data
6. Concluding remarks

Concluding remarks
1. At constant chain lengths the number of RNA sequences exceeds the number of secondary structures by orders of magnitude.
2. The pre-images of common structures in sequence space are extended and connected neutral networks.
3. The intersection of the sets of compatible sequences of two structures is always non-empty.
4. Inverse folding allows for the design of RNA molecules with predefined structures and properties.

Acknowledgement of support
Fonds zur Förderung der wissenschaftlichen Forschung (FWF)
Projects No. 09942, 10578, 11065, 1309313887, and 14898
Jubiläumsfonds der Österreichischen Nationalbank
Project No. Nat-7813
European Commission: Project No. EU-980189
Siemens AG, Austria
The Santa Fe Institute and the Universität Wien
The software for producing RNA movies was developed by Robert Giegerich and coworkers at the Universität Bielefeld
Universität Wien

Coworkers
Universität WienWalter Fontana, Santa Fe Institute, NM
Christian Reidys, Christian Forst, Los Alamos National Laboratory, NM
Peter Stadler, Bärbel Stadler, Universität Leipzig, GE
Ivo L.Hofacker, Christoph Flamm, Universität Wien, AT
Andreas Wernitznig, Michael Kospach, Universität Wien, ATUlrike Langhammer, Ulrike Mückstein, Stefanie Widder
Jan Cupal, Kurt Grünberger, Andreas Svrček-Seiler, Stefan WuchtyAndreas DeStefano
Ulrike Göbel, Institut für Molekulare Biotechnologie, Jena, GEWalter Grüner, Stefan Kopp, Jaqueline Weber

Web-Page for further information:
http://www.tbi.univie.ac.at/~pks





























