Embed Size (px)
Citation preview

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 244, 745–750 (1998)ARTICLE NO. RC988328
Involvement of Mac-1-Mediated Adherenceand Sphingosine 1-Phosphate in Survivalof Phorbol Ester-Treated U937 Cells
Koichi Hamada, Hitoshi Nakamura, Tsukasa Oda,* Tetsuo Hirano,Noriaki Shimizu, and Hiroyasu Utiyama1
The Life Science Group, Faculty of Integrated Arts and Sciences, Hiroshima University, Kagamiyama 1-7-1, Higashi,Hiroshima 739-8521, Japan; and *Helix Research Institute, Yana 1532-3, Kisarazu 292-0812, Japan
Received February 17, 1998
has recently been advanced that phorbol esters viaPhorbol esters exert a dual function in human leuke- protein kinase C (PKC) and phosphoinositides via
mia cells, induction of differentiation and activation phosphoinositide 3-kinase are key mediators of theof integrin-mediated functions. Here we have shown activated state (10). b2 integrins, LFA-1 (CD11a/that the plastic adherence of phorbol ester-treated CD18), Mac-1 (CD11b/CD18), and p150,95 (CD11c/U937 cells is mediated by expression of integrin Mac-
CD18) are all critical to monocyte and granulocyte1 (CD11b/CD18) on the cell surface and that these ad-adhesion-dependent functions as deduced from stud-herent cells exhibit anoikis (apoptosis when adherenties in patients with leucocyte adhesion deficiency (11,cells are detached or adherence is inhibited). We used12). Mac-1 is essential for binding to C3bi and otherU937-derived clones overexpressing either antisenseligands, spreading, phagocytosis, and extravasationRNAs antisense to CD11b and CD18 mRNAs or mRNAof phorbol ester-activated monocytes and granulo-from a truncated mutant CD11b gene. We have alsocytes (13-17). Whereas the inflammatory cytokinesshown that apoptosis in non-adherent cells or anoikissuch as lipopolysaccharide, Interleukin-1, and tumorwas mediated by sphingosine and that survival of ad-
herent cells was achieved by a shift of the dynamic necrosis factor-a (TNF-a) delay the spontaneousbalance between sphingosine and sphingosine 1-phos- apoptosis in monocytes and neutrophils (18-21) byphate toward the latter by adherence-activated sphin- inducing adherence to extracellular matrix with up-gosine 1-kinase. q 1998 Academic Press regulated CD11b/CD18 and CD11c/CD18 (21), in-
flammatory monocytes (22), granulocytes (23), andphorbol ester-activated tissue macrophages (24) areprogrammed to undergo apoptosis by detachment
Many types of mammalian cells are programmed (anoikis in a wider sense). Monocytes and neutro-to undergo apoptosis unless survival signals are pro- phils which have phagocytosed bacteria also die byvided by integrin-mediated cell adhesion to the extra- apoptosis (25). Removal of apoptotic cells by macro-cellular matrix, a process called apoptosis by default phage phagocytosis protects surrounding tissues(1, 2). Apoptotic cell death that occurs when normal from inflammatory cell contents (23). Pre-apoptoticcells are detached from substrate or prevented from senescent neutrophils show reduced adhesion byattaching is called anoikis (3, 4). Integrin-dependent functional uncoupling of integrin receptors from cyto-signal transduction pathways have been described in plasmic factors, without alteration of the surface ex-general (5-7), or more specifically, relating to survival pression of CD11b/CD18 and CD11c/CD18 (26).and anoikis in epithelial, endothelial, and fibroblas- Phorbol esters induce monocyte/macrophage-dif-tic cells (8, 9). ferentiation of the human promonocytic cell line
Regulation of adhesion by integrin activation path- U937 (27) as typically evidenced by newly expressedways is also central to phagocytosis of opsonized tar- Mac-1 and p150,95 on the cell surface (28). Phorbolgets by granulocytes and macrophages, and a notion esters also activate the expressed integrins (29-31),
and induce both phagocytosis of latex beads (32) andadherence to plastic substrate (33). Phorbol ester-1 Corresponding author. Fax: 81-824-24-0759. E-mail: utiyama@
ipc.hiroshima-u.ac.jp. treated U937 and human myeloblastic leukemia HL-
0006-291X/98 $25.00Copyright q 1998 by Academic PressAll rights of reproduction in any form reserved.
745
AID BBRC 8328 / 694d$$$201 03-11-98 11:17:25 bbrcgs AP: BBRC

Vol. 244, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ered saline (PBS(0)) and fixed for 10 min at room temperature. After60 cells are programmed to undergo apoptosis (34),blocking for 20 min with normal goat serum, cells were washed andbut adherent cells exhibit prolonged survival (35).suspended in 50 ml wash buffer (PBS/BSA) containing 0.25 mg/mlHere we suggest that plastic, although never in- dextran sulfate, 0.2% sodium azide, and 0.1% BSA in PBS(0). Cells
gested, may be the target of phagocytosis by activated were incubated with primary antibody OKM-1 to CD11b at 4 7C for30 min, washed with PBS/BSA, and resuspended in 50 ml FITC-goatU937 cells. In this case, plastic-adherent U937 cellsanti-mouse IgG (TAGO, USA). Cells were washed with PBS/BSA,are similar to monocytes engaged in phagocytosis andresuspended, and analyzed on a flow cytometer EPICS-PROFILE IIthus are protected from apoptosis by survival signals(Coulter, USA). Cells which were not treated with primary antibody
provided by integrin-mediated cell adhesion. There- were used as negative controls.fore, we chose to study phorbol ester-treated U937cells to elucidate the signaling pathway for integrin- RT-PCRmediated survival of phagocytosing cells, which re-
Steady-state levels of ectopic mRNA expression in transfectedmains to be well characterized. cells were quantified by reverse transcription-polymerase chainreaction (RT-PCR) by modification of a method described pre-viously (44). Total RNA was prepared in a single step by acidMATERIALS AND METHODSguanidinium thiocyanate-phenol-chloroform extraction (45), anddigested with RNase-free DNase I (RQ-1, Promega, USA). Reverse
Cell Culture and Drug Treatment transcription by MMLV reverse transcriptase (United States Bio-chemicals, USA) was done with RNA samples and appropriateHuman U937 promonocytic cells obtained from ATCC were grown3 *-primers 20 nucleotides in length. cDNA sequences from thein plastic dishes at 377C in RPMI 1640 medium (Nissui, Japan) sup-exogenous, transfected genes were selectively amplified by PCRplemented with 10% fetal calf serum (Irvine Scientific, USA) andby using the same 3 *-primer and a 5*-primer hybridizing to a100 mg/ml kanamycin. Cells were passaged every 3 days and seededsequence between the TATA box and the multi-cloning site in theat 1.5 1 105 cells/ml. The doubling time was 24 h. The cell viability
determined by trypan blue exclusion was higher than 90%. Phorbol12,13-dibutyrate (PDB) (Sigma, USA) was added to a cell culture of4 1 105 cells/ml at 100 nM (36). Counting of adherent cells under aphase contrast microscope was based on adherence to dishes follow-ing light tapping. Cells were treated with sphingolipids in serum-free RPMI 1640 supplemented with 5 mg/ml insulin (Sigma, USA)and 5 mg/ml transferrin (Gibco BRL, USA) after preculture for 24 h inthe same medium (37, 38). N,N-dimethylsphingosine (DMS) (Biomol,USA) was added from a stock solution of 25 mg/ml in ethanol andsphingosine 1-phosphate (SPP) (Biomol, USA) was delivered as acomplex with bovine serum albumin (BSA) in 4 mg/ml BSA (fattyacid-free, Sigma, USA) (37).
Transfection and Clonal Selection
Stably transfected U937 cell lines, As11bN and As11bC overex-pressing CD11b antisense RNA, As18N overexpressing CD18 anti-sense RNA, and GFFKR overexpressing a mutant CD11b gene encod-ing a protein with a truncated cytoplasmic tail, were prepared usingthe pH3M expression vector (39) and the cDNA plasmids for CD11b(40) and CD18 genes (41). The antisense sequence against CD11bcorresponds to the first 1276 base pairs from the translation startsite to the EcoR V restriction site in As11bN and the Pst I/Hind IIIrestriction fragment of 1556 base pairs of the C-terminal domain inAs11bC. The antisense sequence against CD18 in As18N correspondsto the first 1597 base pairs from the translation start site to theSca I restriction site. The truncated CD11b gene in GFFKR clone,prepared by oligonucleotide-directed mutagenesis (42), encodes for amutant protein which lacks the C-terminal cytoplasmic domain ex-cept 7 amino acids, KLGFFKR. The integrity of the construct in theexpression plasmid was confirmed by DNA sequencing. Expressionplasmids were co-transfected with one tenth amount of pSV2neo (43)by electroporation at 300 mFarad, 250 kV in Gene Pulser (BIO-RAD, FIG. 1. Apoptosis in non-adherent cells and survival in adherentUSA), and stable transfectants were selected using growth medium cells in PDB-treated U937 cells. (A) U937 cells (4 1 105 cells/ml)containing 0.4 mg/ml G418 (Gibco BRL, USA). Stably transfected were treated with 100 nM PDB, adherent (closed bars) and non-clones were obtained by limiting dilution and maintained in selection adherent cells (hatched bars) were separately fixed, stained withmedium. DAPI, and cells with fragmented nuclei were counted. (B) U937 cells
were incubated in the presence of the indicated amount of EDTAwith (hatched bars) or without (open bars) PDB for 24 h, wholeFlow Cytometrycells were recovered, and cells with fragmented nuclei were counted.Triplicate counting was performed with a minimum of 200 cellsA cell culture of 0.4 ml (5 1 105 cells/ml) was added to an equal
volume of 4% paraformaldehyde in Mg2/, Ca2/-free phosphate-buff- counted.
746
AID BBRC 8328 / 694d$$$202 03-11-98 11:17:25 bbrcgs AP: BBRC

Vol. 244, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 2. Flow cytometric analysis with monoclonal antibody against CD11b, and RNA expression from exogenous genes determinedby RT-PCR in clones of U937-derived stable transfectants. As11bN [(1) and 1] and As11bC [(2) and 2] overexpress antisense RNAagainst CD11b mRNA, and As18N [(3) and 3] overexpresses antisense RNA against CD18 mRNA. GFFKR [(5) and 5] overexpressesmutant CD11b mRNA encoding a protein with a truncated cytoplasmic tail, KLGFFKR. Untreated U937 cells [(7) and 7] and vectortransfectants [(4), (6), 4, and 6] are controls. Results of negative control staining obtained without using primary antibody are shownby dotted curves.
pH3M vector. PCR reactions were performed for 25 cycles with RESULTS AND DISCUSSIONdenaturation at 92 7C for 1.8 min, annealing at 43 7C for 2 min,and polymerization at 72 7C for 2.2 min. b-actin mRNA expression
PDB treatment (100 nM) of U937 cells induced 60%was measured as a control by RT-PCR with appropriate primers18 nucleotides in length. The PCR product was quantified by elec- cell adherence to plastic culture dishes by 3 h, and atrophoretic separation in an agarose gel, staining with ethidium saturated value of 90% was reached by 12 h (databromide and analysis in an image analyzer (Fas-II, TOYOBO, not shown). The adherent cells survived for 48 h, butJapan). apoptosis of non-adherent cells increased with time
and reached 60% after 48 h (Fig. 1A). When the inte-Assay of Apoptosis. grin-mediated adhesion was inhibited with EDTA,
the percentage of apoptotic cells after 24 h of PDB(a) Nuclear fragmentation. 105 cells were fixed with an equal
treatment increased dose-dependently (Fig. 1B).volume of 5% paraformaldehyde, neutralized by one tenth volume ofThese results clearly show anoikis in PDB-treated1 M Tris-HCl (pH 7.2) and centrifuged onto a glass slide using a
cytospin apparatus. Dried plates were stained with 2 mg/ml 4,6-dia- U937 cells.midino-2-phenylindole (DAPI) in SLOW FADE (Molecular Probes; Next we demonstrate that the plastic adherence ofUSA). Cells with fragmented nuclei were counted under an Olympus PDB-treated U937 cells is predominantly mediatedOMT2 inverted fluorescence microscope equipped with the appro-
by b2 integrin Mac-1, though both Mac-1 and p150,95priate epifluorescence filters at a final magnification of 2001.have been shown to induce adhesion (31). Mac-1 ex-
(b) DNA fragmentation. A previously described procedure (46) pression on the cell surface was inhibited in U937-was used with a slight modification. Briefly, 5 1 105 cells were lysedderived clones which overexpress antisense RNAs,in a lysis buffer containing 0.5% N-lauroylsarcosine in 10 mM EDTA,As11bN and As11bC (against different portions of the50 mM Tris-HCl (pH 8), and the lysate was digested with RNase A
(Sigma, USA), followed by proteinase K. Nucleic acids were precipi- CD11b mRNA) and As18N (against part of the CD18tated in 2 volumes of ethanol, dissolved in 20 ml TE buffer (10 mM mRNA). Since heterodimer formation in the cyto-Tris-HCl (pH 8), 1 mM EDTA), and analyzed by electrophoresis in plasm is a prerequisite for the cell surface-expressiona 2% agarose gel and staining with ethidium bromide. The DNA
of integrin receptors (47-49), the antisense RNAmigration was assessed in an image analyzer (FAS-II, TOYOBO,Japan). against CD11b mRNA specifically blocks Mac-1 ex-
747
AID BBRC 8328 / 694d$$$202 03-11-98 11:17:25 bbrcgs AP: BBRC
Vol. 244, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
expression showed more resistance to apoptosis (datanot shown).
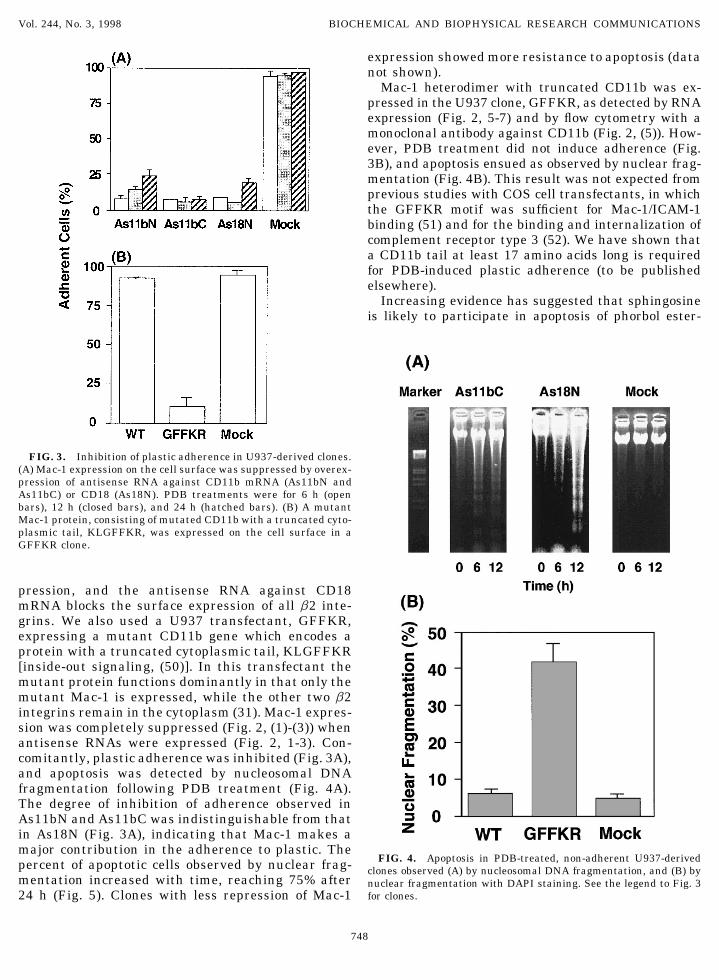
Mac-1 heterodimer with truncated CD11b was ex-pressed in the U937 clone, GFFKR, as detected by RNAexpression (Fig. 2, 5-7) and by flow cytometry with amonoclonal antibody against CD11b (Fig. 2, (5)). How-ever, PDB treatment did not induce adherence (Fig.3B), and apoptosis ensued as observed by nuclear frag-mentation (Fig. 4B). This result was not expected fromprevious studies with COS cell transfectants, in whichthe GFFKR motif was sufficient for Mac-1/ICAM-1binding (51) and for the binding and internalization ofcomplement receptor type 3 (52). We have shown thata CD11b tail at least 17 amino acids long is requiredfor PDB-induced plastic adherence (to be publishedelsewhere).
Increasing evidence has suggested that sphingosineis likely to participate in apoptosis of phorbol ester-
FIG. 3. Inhibition of plastic adherence in U937-derived clones.(A) Mac-1 expression on the cell surface was suppressed by overex-pression of antisense RNA against CD11b mRNA (As11bN andAs11bC) or CD18 (As18N). PDB treatments were for 6 h (openbars), 12 h (closed bars), and 24 h (hatched bars). (B) A mutantMac-1 protein, consisting of mutated CD11b with a truncated cyto-plasmic tail, KLGFFKR, was expressed on the cell surface in aGFFKR clone.
pression, and the antisense RNA against CD18mRNA blocks the surface expression of all b2 inte-grins. We also used a U937 transfectant, GFFKR,expressing a mutant CD11b gene which encodes aprotein with a truncated cytoplasmic tail, KLGFFKR[inside-out signaling, (50)]. In this transfectant themutant protein functions dominantly in that only themutant Mac-1 is expressed, while the other two b2integrins remain in the cytoplasm (31). Mac-1 expres-sion was completely suppressed (Fig. 2, (1)-(3)) whenantisense RNAs were expressed (Fig. 2, 1-3). Con-comitantly, plastic adherence was inhibited (Fig. 3A),and apoptosis was detected by nucleosomal DNAfragmentation following PDB treatment (Fig. 4A).The degree of inhibition of adherence observed inAs11bN and As11bC was indistinguishable from thatin As18N (Fig. 3A), indicating that Mac-1 makes amajor contribution in the adherence to plastic. The
FIG. 4. Apoptosis in PDB-treated, non-adherent U937-derivedpercent of apoptotic cells observed by nuclear frag- clones observed (A) by nucleosomal DNA fragmentation, and (B) bymentation increased with time, reaching 75% after nuclear fragmentation with DAPI staining. See the legend to Fig. 3
for clones.24 h (Fig. 5). Clones with less repression of Mac-1
748
AID BBRC 8328 / 694d$$$202 03-11-98 11:17:25 bbrcgs AP: BBRC

Vol. 244, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
treated cells (53, 54). Macrophagic differentiation ofHL-60 cells by exposure to phorbol esters is blocked bysphingosine and PKC inhibitor (55, 56). Sphingosineand sphinganine, in turn, induce apoptosis in U937 andHL-60 cells, which is enhanced by PKC inhibitor andsuppressed by PKC activator (57). It has recently beendemonstrated that TNF-a-induced apoptosis (58) isblock by activation of sphingosine 1-kinase, whichshifts the dynamic balance between sphingosine andsphingosine 1-phosphate toward the latter (38, 59).Here we show that this mechanism also regulates theapoptosis of non-adherent cells and survival of adher-ent cells following phorbol ester-treatment of U937cells. DMS, also a strong inhibitor of PKC, inducedapoptosis in serum-free medium (Fig. 6A), which wascounteracted by exogenous SPP (Fig. 6B). In addition,we found that phorbol ester-induced apoptosis of adher-ence-defective, U937-derived GFFKR cells was sup-pressed by SPP in all clones studied (33, 21, and 52)(Fig. 6C). SPP has also been reported to block apoptosisby serum withdrawal in rat pheochromocytoma PC12cells (60). Finally, we observed the activation of sphin-gosine 1-kinase in plastic-adherent HL-60 cells inducedby PDB treatment (submitted).
FIG. 6. (A) Induction of apoptosis in U937 cells with DMS, (B)inhibition of DMS-induced apoptosis (5 mM) by simultaneous addi-tion of SPP. (C) Induction of apoptosis in 100 nM PDB-treated (12h), non-adherent GFFKR clones, 33, 21, and 52 (hatched bars) andits suppression with 10 mM SPP (closed bars).
ACKNOWLEDGMENTS
We thank Drs. T. A. Springer for CD11b and CD18 cDNA plasmids,H. Matsuda for use of the flow cytometer in his laboratory, and K.Akahori, S. Ikegami, and T. Suda for stimulating discussions. Thiswork was supported in part by a grant from the Ministry of Educa-tion, Science, and Culture of Japan.
REFERENCES
1. Raff, M. C. (1992) Nature 356, 397–400.2. Ishizaki, Y., Cheng, L., Mudge, A. W., and Raff, M. C. (1995)
Mol. Biol. Cell 6, 1443–1458.3. Frisch, S. M., and Francis, H. (1994) J. Cell Biol. 124, 619–626.FIG. 5. Apoptosis progression with time in PDB-treated, non-4. Ruoslahti, E., and Reed, J. C. (1994) Cell 77, 477–478.adherent U937-derived clones, observed by nuclear fragmentation
after DAPI staining. See the legend to Fig. 3 for clones. 5. Meredith, Jr, J. E., Winitz, S., Lewis, J. M., Hess, S., Ren, X. D.,
749
AID BBRC 8328 / 694d$$$203 03-11-98 11:17:25 bbrcgs AP: BBRC

Vol. 244, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Renshaw, M. W., and Schwartz, M. A. (1996) Endocr. Rev. 17, 33. Hass, R., Bartels, H., Topley, N., Hadam, M., Kohler, L., Goppelt-Strube, M., and Resch, K. (1989) Eur. J. Cell Biol. 48, 282–293.207–220.
34. Gunji, H., Hass, R., and Kufe, D. (1992) J. Clin. Invest. 89, 954–6. Hynes, R. O. (1992) Cell 69, 11–25.960.7. Clark, E. A., and Brugge, J. S. (1995) Science 268, 233–239.
35. Solary, E., Bertrand, R., and Pommier, Y. (1994) Leukemia 8,8. Meredith, J. E., and Schwartz, M. A. (1997) Trends Cell Biol. 7,792–797.146–150.
36. Oda, T., Kitajima, K., Hirano, T., Shimizu, N., and Utiyama, H.9. Bates, R. C., Lincz, L. F., and Burns, G. F. (1995) Cancer Metas- (1993) Int. J. Hematol. 58, 125–128.
tasis Rev. 14, 191–203.37. Zhang, H., Desai, N. N., Olivera, A., Seki, T., Brooker, G., and
10. Kolanus, W., and Seed, B. (1997) Curr. Opin. Cell Biol. 9, 725– Spiegel, S. (1991) J. Cell Biol. 114, 155–167.731. 38. Cuvillier, O., Pirianov, G., Kleuser, B., Vanek, P. G., Coso, O. A.,
11. Arnaout, M. A. (1990) Blood 75, 1037–1050. Gutkind, S., and Spiegel, S. (1996) Nature 381, 800–803.12. Anderson, D. C., and Springer, T. A. (1987) Annu. Rev. Med. 38, 39. Aruffo, A., and Seed, B. (1987) Proc. Natl. Acad. Sci. USA 84,
175–194. 8573–8577.13. Anderson, D. C., Miller, L. J., Schmalstieg, F. C., Rothlein, R., 40. Corbi, A. L., Kishimoto, T. K., Miller, L. J., and Springer, T. A.
and Springer, T. A. (1986) J. Immunol. 137, 15–27. (1988) J. Biol. Chem. 263, 12403–12411.41. Kishimoto, T. K., O’Connor, K., Lee, A., Roberts, T. M., and14. Wright, S. D., and Meyer, B. C. (1986) J. Immunol. 136, 1759–
Springer, T. A. (1987) Cell 48, 681–690.1764.42. Cormack, B. (1993) in Current Protocols in Molecular Biology,15. Wright, S. D., and Silverstein, S. C. (1982) J. Exp. Med. 156,
pp. 8.5.1–8.5.9, Greene and Wiley-Interscience, New York.1149–1164.43. Muller, R., and Wagner, E. F. (1984) Nature 311, 438–442.16. Wright, S. D., Licht, M. R., Craigmyle, L. S., and Silverstein,44. Kinoshita, T., Imamura, J., Nagai, H., and Shimotohno, K. (1992)S. C. (1984) J. Cell. Biol. 99, 336–339.
Anal. Biochem. 206, 231–235.17. O’Shea, J. J., Brown, E. J., Seligmann, B. E., Metcalf, J. A.,45. Chomczynski, P., and Sacchi, N. (1987) Anal. Biochem. 162, 156–Frank, M. M., and Gallin, J. I. (1985) J. Immunol. 134, 2580–
159.2587.46. Martin, S. J., Lennon, S. V., Bonham, A. M., and Cotter, T. G.18. Savill, J. S., Wyllie, A. H., Henson, J. E., Walport, M. J., Henson,
(1990) J. Immunol. 145, 1859–1867.P. M., and Haslett, C. (1989) J. Clin. Invest. 83, 865–875.47. Sastre, L., Kishimoto, T. K., Gee, C., Roberts, T., and Springer,19. Liles, W. C. (1997) Curr. Opin. Infec. Dis. 10, 165–170. T. A. (1986) J. Immunol. 137, 1060–1065.
20. Mangan, D. F., Welch, G. R., and Wahl, S. M. (1991) J. Immunol. 48. Springer, T. A., Thompson, W. S., Miller, L. J., Schmalstieg,146, 1541–1546. F. C., and Anderson, D. C. (1984) J. Exp. Med. 160, 1901–1918.
21. Watson, R. W., Rotstein, O. D., Nathens, A. B., Parodo, J., and 49. Ho, M. K., and Springer, T. A. (1983) J. Biol. Chem. 258, 2766–Marshall, J. C. (1997) J. Immunol. 158, 945–953. 2769.
22. Mangan, D. F., and Wahl, S. M. (1991) J. Immunol. 147, 3408– 50. Williams, M. J., Hughes, P. E., O’Toole, T. E., and Ginsberg,3412. M. H. (1994) Trends Cell Biol. 4, 109–112.
23. Savill, J. (1997) J. Leukoc. Biol. 61, 375–380. 51. Hibbs, M. L., Xu, H., Stacker, S. A., and Springer, T. A. (1991)Science 251, 1611–1613.24. Munn, D. H., Beall, A. C., Song, D., Wrenn, R. W., and Throck-
52. Rabb, H., Michishita, M., Sharma, C. P., Brown, D., and Arnaout,morton, D. C. (1995) J. Exp. Med. 181, 127–136.M. A. (1993) J. Immunol. 151, 990–1002.25. Baran, J., Guzik, K., Hryniewicz, W., Ernst, M., Flad, H. D., and
53. Ohta, H., Sweeney, E. A., Masamune, A., Yatomi, Y., Hakomori,Pryjma, J. (1996) Infect. Immun. 64, 4242–4248.S., and Igarashi, Y. (1995) Cancer Res. 55, 691–697.26. Dransfield, I., Stocks, S. C., and Haslett, C. (1995) Blood 85,
54. Sakakura, C., Sweeney, E. A., Shirahama, T., Hakomori, S., and3264–3273.Igarashi, Y. (1996) Febs Lett. 379, 177–180.
27. Sundstrom, C., and Nilsson, K. (1976) Int. J. Cancer 17, 565–55. Merrill, Jr, A. H., Sereni, A. M., Stevens, V. L., Hannun, Y. A.,577.
Bell, R. M., and Kinkade, Jr, J. M. (1986) J. Biol. Chem. 261,28. Miller, L. J., Schwarting, R., and Springer, T. A. (1986) J. Immu- 12610–12615.
nol. 137, 2891–2900. 56. Kolesnick, R. N. (1989) J. Biol. Chem. 264, 7617–7623.29. Keizer, G. D., Te, V. A., Schwarting, R., Figdor, C. G., and De 57. Jarvis, W. D., Fornari, F. A., Traylor, R. S., Martin, H. A.,
Vries, J. E. (1987) Eur. J. Immunol. 17, 1317–1322. Kramer, L. B., Erukulla, R. K., Bittman, R., and Grant, S. (1996)30. Skoglund, G., Patarroyo, M., Forsbeck, K., Nilsson, K., and Ingel- J. Biol. Chem. 271, 8275–8284.
man-Sundberg, M. (1988) Cancer Res. 48, 3168–3172. 58. Obeid, L. M., Linardic, C. M., Karolak, L. A., and Hannun, Y. A.(1993) Science 259, 1769–1771.31. Nueda, A., Lopez-Rodriguez, C., Rubio, M. A., Sotillos, M., Post-
igo, A., del Pozo, M. A., Vega, M. A., and Corbi, A. L. (1995) Cell 59. Spiegel, S., and Milstien, S. (1995) J. Membr. Biol. 146, 225–Immunol. 164, 163–169. 237.
60. Edsall, L. C., Pirianov, G. G., and Spiegel, S. (1997) J. Neurosci.32. Harris, P. E., Ralph, P., Litcofsky, P., and Moore, M. A. (1985)Cancer Res. 45, 9–13. 17, 6952–6960.
750
AID BBRC 8328 / 694d$$$203 03-11-98 11:17:25 bbrcgs AP: BBRC





























