Embed Size (px)
Citation preview

1
INTERACTION OF SYNTENIN-1 AND THE NG2 PROTEOGLYCAN IN MIGRATORY
OLIGODENDROCYTE PRECURSOR CELLS
Nivedita Chatterjee1*, Judith Stegmüller
1,2*, Philipp Schätzle
1, Khalad Karram
1, Michael Koroll
3,
Hauke B. Werner4, Klaus-Armin Nave
4, and Jacqueline Trotter
1
Molecular Cell Biology, Dept. of Biology, Johannes Gutenberg University of Mainz, Germany1, Present
address: Max-Planck-Institute of Experimental Medicine, Göttingen Germany 2, Max-Delbrück-Center for
Molecular Medicine, Berlin Germany3, Max-Planck-Institute of Experimental Medicine Department of
Neurogenetics, Göttingen Germany4
Running Title: Syntenin-1 binds NG2 in oligodendroglial progenitors
*These two authors contributed equally
Address correspondence to: J. Trotter, Molecular Cell Biology, Dept. of Biology, Bentzelweg 3, Johannes
Gutenberg University of Mainz 55128 Mainz, Germany
Tel (49) 6131 3920263; Fax (49) 6131 3923840; Email: [email protected]
Migration of oligodendrocyte precursors
along axons is a necessary prerequisite for
myelination, but little is known about
underlying mechanisms. NG2 is a large
membrane proteoglycan implicated in
oligodendrocyte migration. Here we show
that a PDZ domain protein, termed syntenin-
1, interacts with NG2 and that syntenin-1 is
necessary for normal rates of migration. The
association of syntenin-1 with NG2, identified
in a yeast-2-hybrid screen, was confirmed by
co-localization of both proteins within
processes of oligodendroglial precursor cells
and by co-immunoprecipitation from cell
extracts. Syntenin-1 also colocalizes with NG2
in 'co-capping' assays, demonstrating a
lateral association of both proteins in live
oligodendrocytes. RNAi-mediated
downregulation of syntenin-1 in glial cells
results in a significant reduction of migration
in vitro, as does the presence of polyclonal
antibody against NG2. Thus Syntenin plays a
role in the migration of oligodendroglial
precursors, and we suggest that NG2-
syntenin-1 interactions contribute to this.
The NG2 proteoglycan is a type I
membrane protein that is expressed by a variety
of immature cells of several embryonic tissue
origins including glia, muscle progenitor cells,
and pericytes (1). In the central nervous system,
expression of NG2 was originally thought to
specify oligodendroglial progenitor cells (OPC),
but more recent data suggests that NG2-
expressing cells encompass a wider range of
immature glial cells in white and grey matter.
These include glia making synaptic-like contacts
to neurons in the hippocampus and cerebellum
(2) and glial cells specifically associated with
the nodes of Ranvier (3). Interestingly, many
NG2+ cells are both proliferative and motile or
exhibit local process motility (4,5). Antibodies
to the NG2 extracellular domain inhibit
migration of OPC and immature Schwann cells
in in vitro migration assays (5,6) and NG2 also
plays a role in cell spreading in melanoma
tumours which express the proteoglycan MCSP,
the human ortholog of NG2 (7). Identifying the
intracellular NG2 interacting proteins should aid
in elucidating the function of this multi-domain
protein in migratory cells of the oligodendrocyte
lineage.
The intracellular domain of NG2
consists of the C-terminal 76 amino acids and
has the PSD-95, Dlg and ZO-1 (PDZ)-binding
motif (QYWV) which can interact with PDZ-
domain containing proteins (8). In order to
define relevant intracellular partners of the
glycoprotein, we have carried out a yeast two
hybrid screen, using the complete intracellular
domain of NG2 as bait. One of the proteins that
we identified is syntenin-1 (also termed mda-9).
Syntenin-1 is a widely expressed PDZ protein
that is often overexpressed in highly migratory
metastatic tumors including melanoma (9).
Here we demonstrate that syntenin is
expressed by primary oligodendrocytes and
show functional studies using the
oligodendroglial precursor cell line Oli-neu. We
provide biochemical, morphological and
functional data demonstrating that NG2 and
syntenin-1 form a complex, which we suggest is
one component regulating oligodendroglial
precursor migration.
http://www.jbc.org/cgi/doi/10.1074/jbc.M706074200The latest version is at JBC Papers in Press. Published on January 24, 2008 as Manuscript M706074200
Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
EXPERIMENTAL PROCEDURES
Animals- NMRI mice were obtained
from the Central Animal Facility of the
University of Mainz.
Antibodies- The following primary
antibodies were used: polyclonal (pc) against rat
syntenin-1 (10), against syntenin-1 generated
using a synthetic peptide corresponding to 286-
298 aa or 250-C-terminus in mouse (Synaptic
Systems, Göttingen, Germany / Abcam,
Cambridge, UK), against EGFP (BD
Biosciences ), against AN2 which recognize
mouse NG2 (5); monoclonal (mc) AN2
antibody(5), anti-GFAP (Glial Fibrillary Acidic
Protein, Boehringer, Mannheim, Germany), anti-
SMI-31 against neurofilament (Sternberger
Monoclonals Incorporated, Lutherville, USA),
anti-aa3 against PLP (M.B. Lees, Waltham,
MA), anti-8-18-C5 against MOG (C. Linington,
Aberdeen, Scotland), anti-SDL.3D10 against !-
tubulin isotype III (Sigma-Aldrich, Munich,
Germany) anti-11-5B against CNPase (Sigma-
Aldrich, Munich, Germany)
Cell Culture and Transfection-
HEK293T cells were cultured in DMEM with
10% Fetal Calf Serum and 2mM glutamine.
Primary oligodendrocyte cultures were prepared
from embryonic day 14-16 mice as described
previously (11). Cells were grown on poly-L-
lysine coated coverslips in modified Sato
medium (12) supplemented with B27, 10 ng/ml
platelet-derived growth factor, 5 ng/ml basic
fibroblast growth factor and 1% horse serum.
Proliferation of astrocytes was prevented by
treatment with Mitomycin C at 10"g/ml for 2.5h
(Sigma-Aldrich), when primary cells were used
for Western blot analysis. The Oli-neu cell line
was cultured according to Trotter and colleagues
(13) on poly-L-Lysine-coated coverslips in
SATO medium containing 1% Horse Serum.
Expression vectors were transfected by
conventional electroporation (20"g/300"l, 4x106
cells/ml). 2mM sodium-butyrate was added to
enhance expression of constructs with a CMV
promoter. Syntenin-1 directed synthetic siRNAs
(target sequences TACGTCAGCATAGTACA-
TTTA and CAGATTGCAGATATACTGTCA,
80 pmol each) and non silencing control siRNA
(target sequence AATTCTCCGAACGTGTCA-
CGT, 160 pmol) were purchased from Qiagen
and nucleofected into 106 Oli-neu cells using the
“AMAXA Basic Nucleofection Protocol for
Primary Mammalian Neural Cells”.
Coimmunoprecipitation- HEK293T
cells were transfected with EGFP-syntenin-1
(10) and the NG2del construct which contains
one-fourth of the extracellular domain and the
complete transmembrane and cytoplasmic tail of
NG2 (8). 24 hours after transfection cells were
washed with phosphate-buffered saline (PBS),
incubated for 1 h in methionine/cysteine-free
medium and were then metabolically labeled
with 100"Ci/ml [35S] Met/Cys for 4 h. Cells
were washed twice with HBSS fortified with
cold Met/Cys and then lysed on ice in 1% Triton
X-100, 50 mM Tris, pH 7.4, 150 mM NaCl and
a protease inhibitor cocktail of iodoacetamide
(18mg/ml in H2O), PMSF (100mM in
isopropanol), pepstatin (5mg/ml in DMSO),
antipain (1mg/ml in DMSO), aprotinin (1mg/ml
in water), benzamidine-HCl (26mg/ml in H2O,
leupeptin (5mg/ml in DMSO). The lysates were
chilled for 30 min and centrifuged at 300g for 5
min to remove nuclei. For immunoprecipitation
the following antibodies were used: rabbit pc
AN2, rabbit pc EGFP, rabbit pc syntenin-1
(Synaptic Systems). Lysates were pre-absorbed
with Protein A-Sepharose (Amersham
Biosciences) for 1 h at 4°C (preclear), followed
by incubation with primary antibodies overnight
at 4°C. Immunoprecipitations were obtained by
addition of Protein A-Sepharose beads and 1h
incubation on a head-over-tail rotator.
Precipitates were washed five times with radio-
immune precipitation assay buffer (RIPA, 0.1%
SDS, 1% Nonidet P-40, 1% sodium
deoxycholate, 150 mM NaCl, 50 mM Tris, pH
7) before adding sample buffer and resolving the
proteins by SDS-PAGE. Gels were dried,
exposed to screens, and evaluated with a
PhosphorImager (Raytest).
Coimmunoprecipitation of endogenous
NG2 with transfected EGFP-syntenin-1 and
endogenous syntenin-1 from Oli-neu cells was
carried out following an identical protocol.
Immunofluorescence Staining- Cells
were washed with PBS, fixed for 10 min with
4% paraformaldehyde (PFA), washed with PBS,
permeabilized for 5 min with 0.05% Triton X-
100, washed with PBS, and blocked with BME
and 10% Horse serum. Primary antibodies
(against syntenin (10)) were diluted in blocking
buffer and incubated for 1 h or overnight in a
humidified chamber. Cells were washed three
times with blocking buffer and incubated for 30
min with appropriate secondary Cy2- and Cy3-
conjugated secondary antibodies (Dianova,
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
Hamburg, Germany), diluted in blocking buffer.
In case of double labeling, primary antibodies
and their corresponding secondary antibodies
were separately added. Coverslips were washed
in distilled water and then mounted in Moviol
for analysis by confocal microscopy (Leica,
Germany).
Capping of Surface Molecules - Primary
oligodendrocytes (2 div = days in vitro) on
coverslips were incubated with mc AN2
antibody (1:10) for 10 min at 4°C. After washing
with PBS, cells were incubated with goat-anti-
rat-Cy2 antibody for 20 min at 37°C, or 4°C: the
latter for control cells. Subsequently, cells were
fixed for 5 min with 4% PFA, permeabilized
with 0.1% TritionX-100 (5 min), blocked with
BME/10%HS and incubated overnight with pc
syntenin-1 antibody. The secondary goat-anti-
rabbit-Cy3 antibody was added the next day,
before mounting the cells in moviol.
Western Blot Analysis- SDS-PAGE was
performed using 4-12% NuPage gels
(Invitrogen, Karlsruhe, Germany). Proteins were
blotted onto polyvinylidene difluoride
membranes (Hybond P, Amersham
Biosciences), blocked with 4% milk powder in
PBS and 0.1% Tween 20 (PBST). Proteins were
detected by sequential incubation with primary
antibodies for 2 hours at room temperature (RT)
and secondary horseradish peroxidase-
conjugated anti-species antibodies (Dianova) for
1 hour at RT. Blots were developed with
enhanced chemiluminescence (Pierce).
Isolation of syntenin-1 with Yeast Two
Hybrid Screen- The entire 76 amino acid
COOH-terminal region of mouse NG2 (NH2-
RKRNKT ... NGQYWV-COOH, GenBank
accession number AF352400) was fused to the
GAL4 binding domain by cloning it into the
pGBT9 vector (Clontech) with XbaI/HindIII.
The resulting bait construct was designated
pGBT9cyto. Using the lithium acetate method,
the yeast strain CG1945 was transformed
sequentially with pGBT9cyto and a 9-12 week
old postnatal mouse brain MATCHMAKER
cDNA library in pACT2 (Clontech). 33 x 106
transformants were screened. Transformants
were grown on SD medium-Leu-Trp-His plates;
5 mM 3-amino-1,2,4-triazole was added to the
medium to suppress leaky HIS3 reporter gene
expression. Positive clones were tested for beta-
galactosidase gene activity: yeast colonies were
grown on SD-Leu-Trp-His, transferred onto
reinforced nitrocellulose membrane, submerged
in liquid nitrogen, and placed on a Z-buffer/X-
gal-soaked Whatman (Z-buffer: 16.1 g/liter
Na2HPO4*7H2O, 5.5 g/liter NaH2PO4*H2O,
0.75 g/liter KCl, 0.246 g/liter MgSO4*7H2O,
pH 7; Z-buffer/X-gal solution: 100 ml of Z-
buffer, 0.27 ml of beta-mercaptoethanol, 1.67 ml
of 20 mg/ml X-gal stock solution). Blue color
was allowed to develop for 30 min-3 h. The
specificity of the NG2-syntenin-1 interaction
was confirmed by beta-galactosidase assay and
growth selection of cotransformed yeast cells
with pGBT9cyto and isolated library plasmids.
To map the PDZ binding motif at the C-terminus
of NG2, individual mutations of the 0, -1, -2,
and -3 positions of the C-terminal peptide
QYWV* were introduced by PCR, cloned into
pGBT9, and designated as NG2 0G (Val
mutated to Gly), NG2 1G (Trp to Gly), NG2 2G
(Tyr to Gly), NG2 2F (Tyr to Phe), NG2 3G
(Gln to Gly). Mutant NG2 constructs were
cotransformed with syntenin-1 f.l (full length)
and syntenin-1 PDZ1-2. Yeast cells were grown
on double dropout medium and assayed for beta-
galactosidase gene activity and additionally
selected for growth on triple dropout medium.
Scratch Migration Assay- Transfected
cells were plated on poly-L-lysine-coated
gridded coverslips (15 mm, Bellco Glass). Cells
were cultured until they reached 80-90%
confluence but were kept at least 16 h prior to
experimental manipulation. Cell-free areas were
generated by gently scratching the cell
monolayer with a sterile blue Gilson-pipette tip.
Coverslips were subsequently washed with PBS
and placed in 3 cm dishes containing
preconditioned Sato medium. Phase images (40x
magnifications) were captured approximately
every 6 hours over a period of 1.5 days. The
“magnetic pen tool” of Photoshop CS2 was used
to manually define the area covered by cells.
The marked (covered) areas were exported and
quantified using Image-Pro Plus software
(MediaCybernetics, MD). The software
calculated the marked area as a percentage of the
whole picture. The results at each time point
were normalized by subtracting the value of the
starting area covered by the cells. In this study
we have equated the percental increase of
marked areas with the migration of the cells.
RESULTS
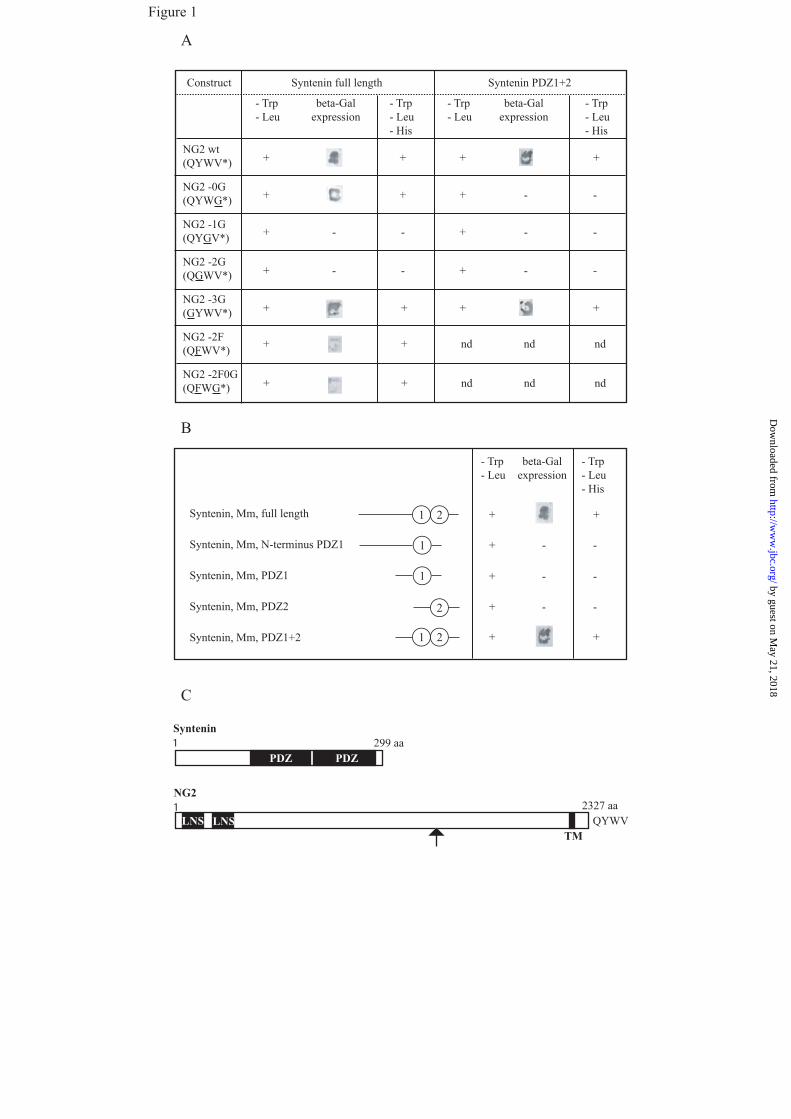
Identification of syntenin-1 as an
intracellular partner of NG2 via yeast two
hybrid screening- To elucidate the mechanism
of NG2 function in migration, we sought to
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
identify novel NG2-interacting proteins using a
Yeast-Two-Hybrid analysis. The complete
mouse NG2 cytoplasmic region consisting of
76 amino acids (RKRN ... QYWV*,
* = translation stop codon) was used as a bait to
screen a postnatal mouse brain cDNA library
(Clontech). Screening of 33 x 106 transformants
identified several potential binding partners
including GRIP (8), and four independent
different library plasmids encoding the PDZ
protein syntenin-1 (14). Each plasmid contained
the entire syntenin-1 cDNA, which corresponds
to the published mouse sequence (14). The
interaction was verified by co-transforming
yeast cells with NG2 and syntenin-1. The
transformed yeast cells were viable on double
and triple dropout media and in addition were
tested positive in the ß-galactosidase assay
(Fig.1 A, row1). This finding suggests that
syntenin-1 is a novel interacting partner of NG2.
The C-terminus of NG2 displays a canonical
PDZ binding motif (QYWV). To validate the
requirement of the PDZ motif for the interaction
of NG2 with syntenin-1, we carried out point
mutation analyses and found that the terminal
three amino acid residues in the PDZ-
recognition site of NG2 are important for
binding to the PDZ domains of syntenin-1 (Fig1,
rows 2-6). These data demonstrate that NG2’s
PDZ binding motif QYWV is critical for the
binding to syntenin-1.
Syntenin-1 harbors a tandem repeat of
two PDZ domains. To determine which PDZ
domain is required for the interaction with NG2,
we carried out deletion analyses. Syntenin-1
deletion mutants were generated by PCR using
full length mouse Syntenin-1 as a template,
cloned into EcoRI/BamHI sites of the pACT2
vector and verified by sequencing. The
following deletion mutants were tested for
interaction with NG2: Syntenin-1-N-
terminus/PDZ1 (MSLYP-HKDSS), Syntenin-1-
PDZ1 (RAEIK-HKDSS), Syntenin-1-PDZ2
(FERTV-TIPEV*), Syntenin-1-PDZ1/2
(RAEIK-TIPEV*). Both PDZ domains were
required for binding to NG2 (Fig. 1B).
Syntenin-1 is expressed by immature
NG2-positive and by more mature
oligodendrocytes in vitro- We determined the
expression of syntenin-1 in primary
oligodendrocytes and the cell line Oli-neu (Fig.
2). Lysates of primary oligodendrocytes (2 div),
Oli-neu cells (Fig. 2 A, lane 2 and 3) and total
brain were subjected to Western blot analysis
using the syntenin-1 antibody. We found that
syntenin-1 is expressed in both primary
oligodendrocytes and Oli-neu cells (Fig. 2 A,
lanes 2 and 3). Total brain lysate served as a
positive control to validate the 36kD syntenin-1
band (lane 1). Interestingly, the highly migratory
Oli-neu cells reveal a high expression of
syntenin-1. These results show that syntenin-1 is
expressed in oligodendroglial cells.
Next, we characterized the
developmental profile of syntenin-1 in primary
oligodendrocytes during differentiation in vitro
(Fig. 2B). The expression of syntenin-1
increases with time in culture. The decrease of
NG2, which is expressed by immature
oligodendrocytes and the increase of PLP, a
marker for more mature oligodendrocytes,
suggests that the increase in syntenin-1
expression parallels oligodendrocyte maturation.
The possibility that the syntenin-1 signals arise
from neuronal and/or astrocytic contamination
was excluded by controlling for expression of
the neuronal protein neurofilament and the
asctrocyte-specific protein GFAP. We did not
observe detectable amounts of neurofilament,
and the low amount of GFAP expression did not
correlate with the increase of syntenin-1 over
time. These results suggest an increase in
syntenin-1 expression in cultures of maturing
oligodendrocytes.
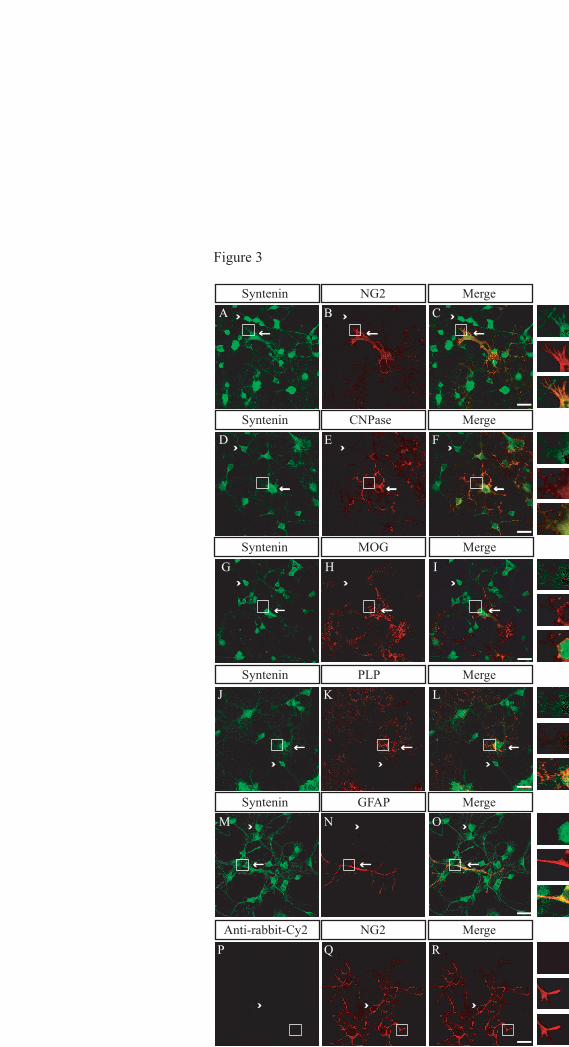
To corroborate this finding, we
subjected cultured oligodendrocytes to
immunocytochemistry using stage-specific
markers for oligodendrocytes. We found
syntenin-1 expression at different stages of
oligodendrocyte differentiation (Fig 3). All
NG2-positive cells at 2 div expressed syntenin-
1. In addition, we found that many more mature
oligodendrocytes in the culture, identified by
CNP, MOG and PLP expression, also express
syntenin-1. These data demonstrate that
syntenin-1 is expressed by immature as well as
by differentiated oligodendrocytes. The
extensive syntenin-positive process outgrowth in
older cells may contribute to the increase in
syntenin-1 with time in culture.
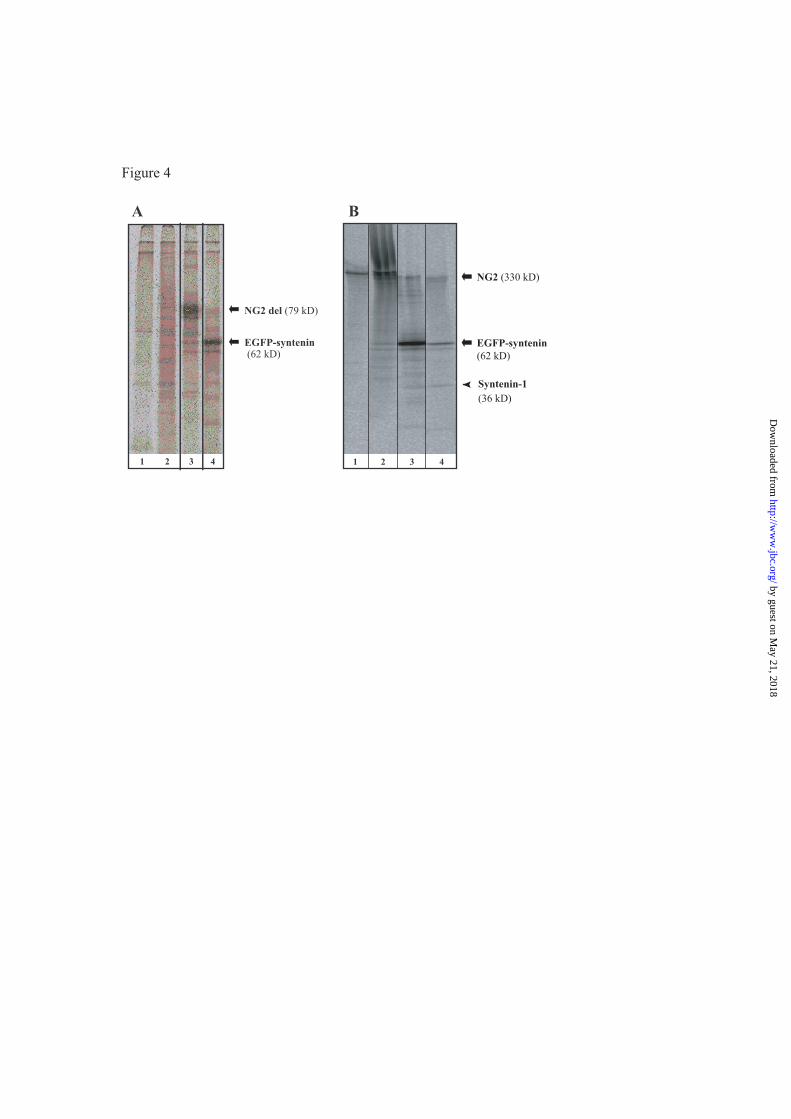
Co-immunoprecipitation of NG2 and
syntenin-1- To validate the interaction of
syntenin-1 and NG2 biochemically, HEK293T
cells were cotransfected with a plasmid
encoding EGFP-syntenin-1 (Enhanced Green
Fluorescent Protein-syntenin-1; (10) and a
plasmid encoding NG2del (encompassing one-
fourth of the extracellular domain and the
complete transmembrane and cytoplasmic tail of
NG2, Fig.1C) (8). A complex of EGFP-
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
syntenin-1 and NG2 was isolated from the
transfected cells after metabolic radiolabelling
by precipitating with polyclonal AN2 antibodies
against NG2 which recognize NG2del (Fig. 4A
lane 3) or polyclonal antibodies against EGFP,
which target EGFP-syntenin-1 (lane 4). This
finding suggests that NG2 and EGFP-syntenin-1
co-associate. Next, we examined the association
of syntenin-1 and NG2 in Oli-neu cells. All Oli-
neu cells are NG2-positive. In cells, which had
been transfected with EGFP-syntenin-1,
precipitation with polyclonal AN2 antibodies
precipitated endogenous NG2 as well as the
62kD EGFP-syntenin-1 (Fig. 4 B lane 2).
Precipitation using polyclonal antibodies against
EGFP precipitated EGFP-syntenin-1 and
associated NG2 (lane 3). Polyclonal antibodies
against syntenin-1 pulled down EGFP-syntenin-
1, endogenous NG2 and gave a weak signal for
endogenous syntenin-1 (arrowhead, Lane 4).
Co-immunoprecipitation of endogenous
NG2 and endogenous syntenin-1 from native
Oli-neu did not yield a syntenin-1 signal: this
may be due to the difficulty of extracting NG2
and syntenin-1 from cytoskeletal-associated
components Syntenin-1 has been reported to be
difficult to extract from membrane fractions
with detergents, high salt, carbonate and freeze-
thawing (15) compatible with a cytoskeleton
association. Furthermore, it is likely that only a
small fraction of each protein is associated in a
complex.
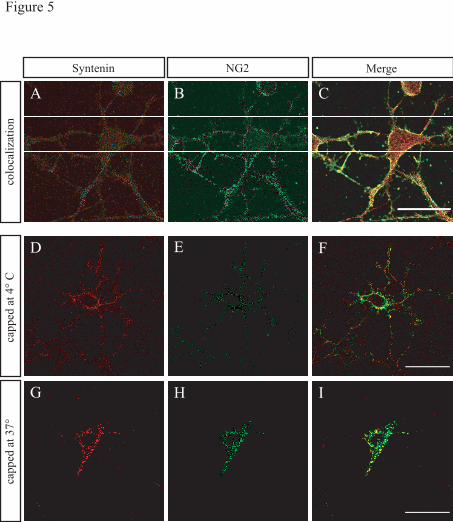
NG2 colocalizes with syntenin-1 in
NG2-expressing primary glial cells in vitro-
Primary oligodendrocytes were double-labeled
with AN2 monoclonal antibody which
recognizes NG2, and polyclonal antibody
against syntenin-1 (Fig. 5 A-C). While NG2 was
predominantly localized at the cell surface, on
the cell body and processes, syntenin-1 is
expressed throughout the whole cell in a
punctate manner. Strikingly, we found that NG2
and syntenin-1 colocalize at the surface of the
cell as indicated by the yellow punctate
fluorescence in the overlay (Fig. 5A-C). The
colocalization is particularly intense at distinct
varicosities in the processes: these regions may
indicate points were branching has just started.
Capping of NG2 on glial cells causes a
parallel redistribution of syntenin-1- To
corroborate that NG2 and syntenin-1 form a
complex in primary oligodendrocytes, we
performed a co-capping assay (Fig. 5 D-I).
Clustering and parallel movement of NG2 and
syntenin-1 was explored by incubating live
oligodendrocytes (2 div) with AN2 monoclonal
antibody for 10 minutes at 4° C. This was
followed by addition of a fluorescent secondary
antibody and incubation at 37° C. A patching
and capping of NG2 was induced. A similar
redistribution of a population of the endogenous
syntenin-1 was observed resulting in clusters of
NG2 and syntenin-1 at or near the cell surface,
bolstering our finding that NG2 and syntenin-1
associate in oligodendrocytes.
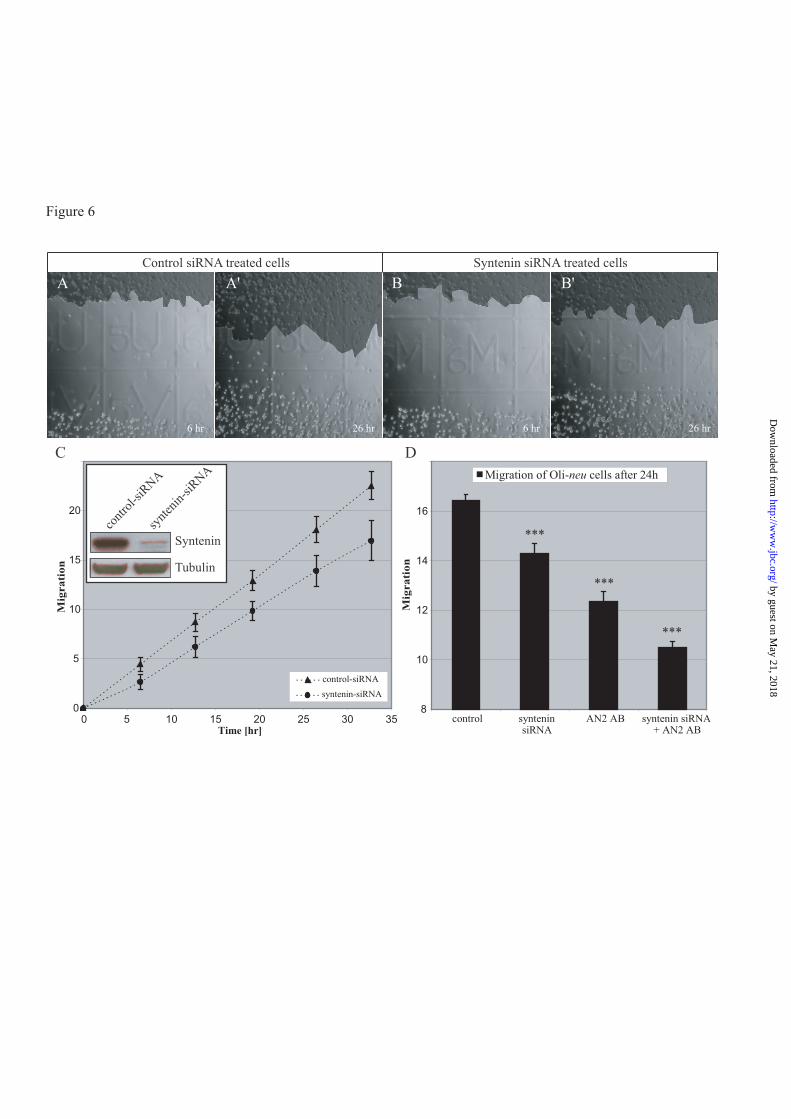
Syntenin-1 promotes migration of Oli-
neu cells- NG2 has been implicated in spreading
and migration (4-6,16,17). If syntenin-1-NG2
binding were functionally important, loss of
syntenin-1 should reduce the migratory
behaviour of Oli-neu cells and primary NG2-
positive oligodendrocytes. We took a siRNA
approach to acutely knockdown syntenin-1 in
Oli-neu cells. Oli-neu cells were transfected with
either control siRNA or syntenin-1 siRNA. By
Western blot analysis, we found an efficient
knockdown of syntenin-1 in Oli-neu cells 16
hours after transfection as compared to control
siRNA-transfected cells (Fig.6, insert). Western
blot analysis at different times after transfection
demonstrated that the syntenin knockdown was
stable up to at least 54 hours after transfection
(data not shown), and thus maintained during the
whole time course of the migration assays.
Control and syntenin-1 siRNA-transfected Oli-
neu cells were then analyzed in a scratch assay:
knockdown of syntenin-1 significantly reduced
the migration of Oli-neu cells as compared to
control cells (Fig. 6C). Inclusion of polyclonal
AN2 antibody directed against the whole NG2
molecule which recognizes primarily the protein
core (5) reduced migration to a greater extent
than syntenin-1 siRNA alone (Fig. 6D). When
both these approaches were combined in the
assay, a stronger inhibition of migration was
found than with either syntenin-1 siRNA or AN2
antibodies alone, suggesting that the signaling
pathways in the migration promoting effects of
NG2 and syntenin are not completely
overlapping. These data demonstrate that
syntenin-1, like NG2, promotes migration of
Oli-neu cells.
DISCUSSION
PDZ proteins as partners of NG2- The
NG2 proteoglycan binds to several PDZ-domain
proteins with its C-terminal QYWV motif. This
has been shown for MUPPI 1 and GRIP (8,18).
PDZ domain proteins act as intracellular
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
scaffolds coordinating and compartmentalizing
molecules involved in signal transduction. The
majority of the PDZ-domain-containing proteins
are associated with plasma membrane proteins
and they are generally restricted to specific
subcellular domains such as synapses or cell-cell
contact points (19). Here, we characterize
syntenin-1 as an additional intracellular ligand
of NG2.
Expression of syntenin-1- Syntenin-1
was first identified as an adaptor protein binding
syndecan in focal adhesion complexes (14), and
later observed to be associated with a number of
membrane proteins as part of multi-protein
assemblies, notably at the pre- and post-synaptic
terminal (20-22). Syntenin-1 shows a
widespread tissue expression at both fetal and
adult stages (14) which is particularly high in
fetal kidney, liver, lung and brain (15). In the
nervous system, syntenin-1 has been shown to
bind the cell adhesion molecule neurofascin
(10). In vivo subpopulations of neurons have
been reported to express syntenin-1: cell bodies
and dendrites of pyramidal cells show a diffuse
expression of syntenin-1 (23).
Cloning of mouse and rat syntenin-1s
(also known as melanoma differentiation
associated gene-9 or mda-9) revealed that the
molecules are highly homologous (9): the rodent
and human mda-9/syntenin-1 is nearly identical
at the level of PDZ-1 and the COOH-terminal
domains. The N-terminal portion is the most
divergent: in humans this domain is 81% and
77% identical to the mouse and rat domains,
respectively. A second family member,
syntenin-2 also exists (10).
Cell lysates from cultured primary
oligodendrocytes, the cell line Oli-neu and
whole mouse brain homogenate all show
expression of syntenin-1 by Western
blotting and immunofluorescence. Both a
membrane-associated and a cytosolic
distribution of the protein were observed,
similar to observations in lymphocytes (24).
In both Oli-neu and primary cells the
intracellular staining at high resolution often
appeared punctated and may represent
vesicular association. Colocalization of NG2 and syntenin-1-
In primary oligodendrocyte cultures an intense
colocalization of NG2 and syntenin-1 at
varicosities in OPC processes was seen. Recent
work reported that in neurons overexpression of
syntenin-1 increased the number of dendritic
protrusions (25).
Subpopulations of syntenin-1 and NG2
co-cap. Syntenin-1 is a component of early
secretory pathways (26) and has been found to
be expressed by early apical recycling
endosomes (27) and to bind to the GTPase Rab 5
(28).
Only a sub-population of syntenin-1
colocalizes with NG2 further reiterating that
syntenin-1 (like NG2) has several binding
partners and only a subpopulation of syntenin-1
interacts with NG2 at any given time point.
Namely, it has been reported that NG2 can
interact with MUPP1 (18) or GRIP1(8). At
present, no experimental evidence exists that
MUPP1 or GRIP1 are involved in the migration
or process motility of NG2-positive glia or
oligodendrocyte progenitors. Our results suggest
that a subset of NG2 molecules cooperate with
syntenin-1 in a molecular complex required for
cellular motility, while the interaction with
MUPP1 or GRIP1 is important for other cellular
functions (discussed in(8,18). Our results are
similar to observations in T and B lymphocytes
where a subpopulation of syntenin-1 colocalizes
with the binding partner CD6 after capping (24).
CD6 like NG2 is a type I membrane
glycoprotein expressed by thymocytes, mature T
and B lymphocytes and in some neuronal
subpopulations in basal ganglia and cerebellar
cortex (29).
Molecular characteristics of the NG2
syntenin-1 binding- The interaction of syntenin-
1 with NG2 requires both PDZ domains; it has
not been ascertained whether the binding is
cooperative. Syntenin-1 has the capacity to
multimerize (10): this may explain the
observation that an endogenous syntenin-1 band
was detected in immunoprecipitates of Oli-neu
expressing EGFP-syntenin-1. PDZ domains
have the propensity to multimerize as homo-
multimers or hetero-multimers (30) and can
multimerize via PDZ-independent mechanisms
(31). Moreover, not all interactions with
syntenin-1 require both domains (32). A
syntenin-1-NG2 interaction does not thus
exclude simultaneous binding of other ligands.
The ability to form multimers further increases
the possibility of the simultaneous interaction
with several ligands.
Functional implications of the NG2-
syntenin-1 interaction- Actin-binding domains
have not been identified in the cytoplasmic tail
of NG2: however NG2 plays a role in migration,
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
metastasis and adhesion complexes. The
association of NG2 with downstream
cytoskeletal machinery must therefore be
mediated by linker proteins. The high expression
levels of both NG2 and syntenin-1 in immature
glia correlates with the motility of these cells as
well as localized motility of their processes.
Knockdown of syntenin-1 reduces migration of
Oli-neu cells. Syntenin-1 may be an adaptor
molecule which connects NG2 to downstream
components and may play a role in mediating
cytoskeletal changes during movement of NG2-
expressing cells and / or their processes.
However, in many cell types NG2 interacts in
cis with integrins, which may promote migration
independent of syntenin (33). Similarly,
syntenin-1 may additionally promote migration
by mechanisms independent of its interaction
with NG2. Syntenin can mediate cytoskeletal
dynamics at the plasma membrane: syntenin-1
overexpression induces the formation of long,
branching plasma membrane extensions (14). In
astrocytomas NG2 has been compartmentalized
with membrane cytoskeletal linkers such as
ERM proteins which interact with actin (16).
The expression of ERM proteins by
oligodendrocytes has not been reported.
Interestingly, immature Schwann cells express
NG2 and also the ERM protein
Merlin/Schwannomin which is a reported
binding partner of syntenin-1 (34).
The association of syntenin-1 with
endocytic compartments suggest that it may
regulate endocytosis and trafficking of NG2 as
has been shown for syndecan via an interaction
of syntenin-1 with phosphatidylinositol 4,5
bisphosphate (PIP2) (35,36).
In more mature oligodendrocytes, which
we show also express syntenin-1 but are NG2-
negative and non-migratory, syntenin-1 is likely
to have different binding partners and could
conceivably influence the massive membrane
remodeling involved in myelin sheet formation
(37).
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
REFERENCES
1. Nishiyama, A. (2007) Neuroscientist 13(1), 62-76
2. Bergles, D. E., Roberts, J. D., Somogyi, P., and Jahr, C. E. (2000) Nature 405(6783), 187-
191
3. Butt, A. M., Kiff, J., Hubbard, P., and Berry, M. (2002) J Neurocytol 31(6-7), 551-565
4. Eisenmann, K. M., McCarthy, J. B., Simpson, M. A., Keely, P. J., Guan, J. L., Tachibana,
K., Lim, L., Manser, E., Furcht, L. T., and Iida, J. (1999) Nat Cell Biol 1(8), 507-513
5. Niehaus, A., Stegmuller, J., Diers-Fenger, M., and Trotter, J. (1999) J Neurosci 19(12),
4948-4961
6. Schneider, S., Bosse, F., D'Urso, D., Muller, H., Sereda, M. W., Nave, K., Niehaus, A.,
Kempf, T., Schnolzer, M., and Trotter, J. (2001) J Neurosci 21(3), 920-933
7. Pluschke, G., Vanek, M., Evans, A., Dittmar, T., Schmid, P., Itin, P., Filardo, E. J., and
Reisfeld, R. A. (1996) Proc Natl Acad Sci U S A 93(18), 9710-9715
8. Stegmuller, J., Werner, H., Nave, K. A., and Trotter, J. (2003) The Journal of biological
chemistry 278(6), 3590-3598
9. Sarkar, D., Boukerche, H., Su, Z. Z., and Fisher, P. B. (2004) Pharmacol Ther 104(2),
101-115
10. Koroll, M., Rathjen, F. G., and Volkmer, H. (2001) The Journal of biological chemistry
276(14), 10646-10654
11. Kramer, E. M., Koch, T., Niehaus, A., and Trotter, J. (1997) The Journal of biological
chemistry 272(14), 8937-8945
12. Trotter, J., Bitter-Suermann, D., and Schachner, M. (1989) Journal of neuroscience
research 22(4), 369-383
13. Jung, M., Kramer, E., Grzenkowski, M., Tang, K., Blakemore, W., Aguzzi, A., Khazaie,
K., Chlichlia, K., von Blankenfeld, G., Kettenmann, H., and et al. (1995) Eur J Neurosci
7(6), 1245-1265
14. Grootjans, J. J., Zimmermann, P., Reekmans, G., Smets, A., Degeest, G., Durr, J., and
David, G. (1997) Proc Natl Acad Sci U S A 94(25), 13683-13688
15. Zimmermann, P., Tomatis, D., Rosas, M., Grootjans, J., Leenaerts, I., Degeest, G.,
Reekmans, G., Coomans, C., and David, G. (2001) Mol Biol Cell 12(2), 339-350
16. Makagiansar, I. T., Williams, S., Dahlin-Huppe, K., Fukushi, J., Mustelin, T., and
Stallcup, W. B. (2004) The Journal of biological chemistry 279(53), 55262-55270
17. Majumdar, M., Vuori, K., and Stallcup, W. B. (2003) Cell Signal 15(1), 79-84
18. Barritt, D. S., Pearn, M. T., Zisch, A. H., Lee, S. S., Javier, R. T., Pasquale, E. B., and
Stallcup, W. B. (2000) J Cell Biochem 79(2), 213-224
19. Sheng, M., and Sala, C. (2001) Annu Rev Neurosci 24, 1-29
20. Torres, R., Firestein, B. L., Dong, H., Staudinger, J., Olson, E. N., Huganir, R. L., Bredt,
D. S., Gale, N. W., and Yancopoulos, G. D. (1998) Neuron 21(6), 1453-1463
21. Lin, D., Gish, G. D., Songyang, Z., and Pawson, T. (1999) The Journal of biological
chemistry 274(6), 3726-3733
22. Hirbec, H., Perestenko, O., Nishimune, A., Meyer, G., Nakanishi, S., Henley, J. M., and
Dev, K. K. (2002) The Journal of biological chemistry 277(18), 15221-15224
23. Ohno, K., Koroll, M., El Far, O., Scholze, P., Gomeza, J., and Betz, H. (2004) Mol Cell
Neurosci 26(4), 518-529
24. Gimferrer, I., Ibanez, A., Farnos, M., Sarrias, M. R., Fenutria, R., Rosello, S.,
Zimmermann, P., David, G., Vives, J., Serra-Pages, C., and Lozano, F. (2005) J Immunol
175(3), 1406-1414
25. Hirbec, H., Martin, S., and Henley, J. M. (2005) Mol Cell Neurosci 28(4), 737-746
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
26. Fernandez-Larrea, J., Merlos-Suarez, A., Urena, J. M., Baselga, J., and Arribas, J. (1999)
Mol Cell 3(4), 423-433
27. Fialka, I., Steinlein, P., Ahorn, H., Bock, G., Burbelo, P. D., Haberfellner, M., Lottspeich,
F., Paiha, K., Pasquali, C., and Huber, L. A. (1999) The Journal of biological chemistry
274(37), 26233-26239
28. Tomoda, T., Kim, J. H., Zhan, C., and Hatten, M. E. (2004) Genes Dev 18(5), 541-558
29. Mayer, B., Funke, I., Seed, B., Riethmuller, G., and Weiss, E. (1990) J Neuroimmunol
29(1-3), 193-202
30. Srivastava, S., Osten, P., Vilim, F. S., Khatri, L., Inman, G., States, B., Daly, C., DeSouza,
S., Abagyan, R., Valtschanoff, J. G., Weinberg, R. J., and Ziff, E. B. (1998) Neuron 21(3),
581-591
31. Hsueh, Y. P., Kim, E., and Sheng, M. (1997) Neuron 18(5), 803-814
32. Xu, X. Z., Choudhury, A., Li, X., and Montell, C. (1998) J Cell Biol 142(2), 545-555
33. Karram, K., Chatterjee, N., and Trotter, J. (2005) J Anat 207(6), 735-744
34. Jannatipour, M., Dion, P., Khan, S., Jindal, H., Fan, X., Laganiere, J., Chishti, A. H., and
Rouleau, G. A. (2001) The Journal of biological chemistry 276(35), 33093-33100
35. Zimmermann, P., Meerschaert, K., Reekmans, G., Leenaerts, I., Small, J. V.,
Vandekerckhove, J., David, G., and Gettemans, J. (2002) Mol Cell 9(6), 1215-1225
36. Zimmermann, P., Zhang, Z., Degeest, G., Mortier, E., Leenaerts, I., Coomans, C., Schulz,
J., N'Kuli, F., Courtoy, P. J., and David, G. (2005) Dev Cell 9(3), 377-388
37. Simons, M., and Trotter, J. (2007) Curr Opin Neurobiol
Acknowledgements
We thank Lilia Niedens and Ulrike Stapf for excellent technical assistance, Reinhard Windoffer for advice
with the use of Image Pro Plus and the Gemeinnützige Hertie Stiftung, the Deutsche
Forschungsgemeinschaft (SPPs Cell Polarity and Glia-Synapse) and the European Union FP6 (Signalling
and Traffic) for financial support.
Figure 1. Syntenin-1 binds to NG2 via both PDZ domains
A, the amino acids tryptophan, tyrosine and valine in the PDZ-binding motif of NG2 are essential for
binding to syntenin-1. Yeast cells were transformed with full length syntenin-1 or a syntenin-1 deletion
mutant containing PDZ1-2 together with different PDZ binding motif mutants of NG2. These were then
tested for ß-galactosidase activity in triple amino acid deficient medium. nd = not determined
B, both PDZ domains of syntenin-1 are required for binding to NG2. Yeast were transformed with the
cytoplasmic tail of NG2 together with different syntenin-1 deletion mutants and tested for ß-galactosidase
activity and growth in triple-amino acid deficient medium.
C, Schematic diagram of Syntenin and NG2. PDZ indicates two PSD95/Discslarge/Zona Occludens-1
domains, LNS indicates two LamininG/Neurexin/Sex hormone-binding globulin domains, TM indicates
the transmembrane domain and the arrow indicates the position of the deletion (aa478-aa2164) in NG2del.
The amino acids QYWV represent the PDZ binding motif.
Figure 2. Syntenin-1 is expressed by oligodendrocytes in culture
A, Lysates from whole brain lysate (lane 1), cultured (2 div) primary oligodendrocytes (lane 2), and Oli-
neu (lane 3) were immunoblotted using the polyclonal syntenin-1 antibody.
B, Lysates from primary oligodendrocyte cultures were analyzed for expression of syntenin-1 at different
stages of maturity by western blotting. NG2 represents an early marker in the lineage, while PLP is
expressed by more matured cells. Probing for neurofilament and GFAP confirmed that the increase in
syntenin-1 expression is not due to neuronal or astrocytic proliferation.
Figure 3. Syntenin-1 expression in cultured glial cells
Oligodendrocyte cultures (2 div) were immunolabelled for syntenin (green, A, D, G, J, M) and either NG2
(red, B, Q), CNP (red, E), MOG (red, H), or PLP (red, K) identifying different stages of oligodendrocyte
development. Astrocyte contamination was visualized by staining for GFAP (red, N). Overlays show the
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
expression of syntenin in oligodendrocytes (C, F, I, L) and astrocytes (O). Arrow heads show single-
labeled cells and arrows show double-labeled cells. White boxes show enlarged areas of double-labeled
cells. The secondary antibody control for the Syntenin staining is shown in P (green). Scale bar equals 20
"m.
Figure 4. Syntenin-1 and NG2 form a complex in transfected HEK293 cells and in the OPC line Oli-
neu.
A, HEK293T cells were transfected with a plasmid encoding EGFP-tagged syntenin-1 together with the
NG2del plasmid, a deletion construct of NG2 missing most of the extracellular region but retaining the two
LamininG-Neurexin-Sex hormone-binding globulin (LNS) domains and the complete cytoplasmic tail (8).
The cells were metabolically labeled and lysates were subjected to immunoprecipitation with pc AN2
recognising NG2del and pc EGFP antibodies.
Lane 1: Pre-clear; Lane 2: Non-transfected cells, immunoprecipitation with pc AN2; Lane 3: Transfected
cells, immunoprecipitation with pc AN2; Lane 4: Transfected cells, immunoprecipitation with pc EGFP,
B, Oli-neu cells were transfected with the EGFP-syntenin-1 plasmid and lysates were subjected to
immunprecipitation. Lane 1: Untransfected cells, immunoprecipitation with pc AN2; Lane 2: Transfected
cells, immunoprecipitation with pc AN2; Lane 3: Transfected cells, immunoprecipitation with pc EGFP;
Lane 4: Transfected cells, immunoprecipitation with pc syntenin-1 antibodies.
Figure 5. Syntenin-1 and NG2 colocalize and co-cap in immature oligodendrocytes in culture
Cultured oligodendrocytes (2 div) were stained for syntenin-1 (A) and NG2 (B). An Overlay (C) shows
intense colocalization at the plasma membrane.
To demonstrate co-capping: Primary oligodendrocytes were incubated with the mc AN2 antibody at 4°C,
followed by incubation with fluorescent secondary anti-rat antibody at 4°C (D-F) or 37°C (G-I). The cells
were then fixed and permeabilized. Subsequently, the cells were subjected to immunocytochemistry with
the syntenin-1 antibody at room temperature. Single section confocal images show co-capping of NG2 and
syntenin-1 at 37°C in presence of mc AN2.
Scale bar equals 20"m.
Figure 6. RNAi knockdown of syntenin-1 affects migration of Oli-neu cells
Oli-neu cells were transfected with syntenin-1 siRNA and subsequently incubated for 16h. A scratch was
performed using a pipette tip. Images were taken at regular intervals over the course of 34 h. A, A’ and B,
B’ show an example for control as well as syntenin siRNA treated cells after 6h or 26h, respectively. Areas
which were covered by cells were manually marked and quantified at each time point. C, Oli-neu cells
with reduced syntenin-1 expression migrate less than the control siRNA treated cells. Error bars represent
the standard deviation of the mean (n=12). C insert, Western blot analysis of cells from the same
transfections used in the migration assays at 16 hours after transfection, verified the successful knockdown
of syntenin-1 protein. D, polyclonal AN2 antibody against NG2 reduces migration of Oli-neu cells.
Antibody and syntenin-1 siRNA together cause a stronger reduction in migration than either treatment
alone. Results are expressed as the mean of 20 different migration fronts with standard deviation (*** P
value <0,0001 by comparison with the control).
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from
A
B
beta-Galexpression
NG2 wt(QYWV*)
NG2 -0G(QYWG*)
NG2 -1G(QYGV*)
NG2 -3G(GYWV*)
NG2 -2F(QFWV*)
NG2 -2F0G(QFWG*)
NG2 -2G(QGWV*)
+
Construct
- Trp- Leu- His
- Trp- Leu- His
- Trp- Leu
- Trp- Leu
beta-Galexpression
Syntenin PDZ1+2Syntenin full length
+ + +
+ + + --
+ - + ---
+ - + ---
+ + + +
+ + nd ndnd
+ + nd ndnd
1
1 2
1
2
1 2
Syntenin, Mm, full length
Syntenin, Mm, N-terminus PDZ1
Syntenin, Mm, PDZ1
Syntenin, Mm, PDZ2
Syntenin, Mm, PDZ1+2
beta-Galexpression
- Trp- Leu
- Trp- Leu- His
+
+ -
+ -
+ -
+
-
-
-
+
+
1 299 aa
1 2327 aa
PDZ PDZ
LNS LNS TM
QYWV
NG2
Syntenin
C
Figure 1 by guest on M
ay 21, 2018http://w
ww
.jbc.org/D
ownloaded from

) 0 1 P ( n i a r B 2 4 7 11
Primary oligo- dendrocytes (div)
Syntenin (36 kDa)
NG2 (330 kDa)
PLP (25 kDa)
Neurofilament (200 kDa) GFAP (50 kDa)
Tubulin (50 kDa)
) 0 1 P ( n i a r B
v i d 2 . d o g i l o y r a m i r P
u e n - i l O
Syntenin (36 kDa)
Tubulin (50 kDa)
A B
1 2 3
Figure 2
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from
A B CMergeNG2Syntenin
MergeCNPaseD E F
MergeNG2Anti-rabbit-Cy2P Q R
Syntenin
MergeMOGG H I
Syntenin
MergePLPJ K L
Syntenin
M N OMergeGFAPSyntenin
Figure 3
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from
A
NG2 (330 kD)
EGFP-syntenin (62 kD)
B
EGFP-syntenin (62 kD)
NG2 del (79 kD)
3 4 2 1 3 4 2 1
Syntenin-1 (36 kD)
Figure 4
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from
0
5
1 0
1 5
2 0
0 5 1 0 1 5 2 0 2 5 3 0 3 5 T i m e [ h r ]
M i g
r a t i o
n
c o n t r o l - s i R N A s y n t e n i n - s i R N A
Syntenin
Tubulin
A N
R i s - l o r t n o c
A N
R i s - n i n e t n y s
C
Control siRNA treated cells Syntenin siRNA treated cells
6 hr 26 hr
A B A' B'
6 hr 26 hr
D
c o n t r o l s y n t e n i n A N 2 A B s y n t e n i n s i R N A + A N 2 A B
M i g r a t i o n o f O l i - n e u c e l l s a f t e r 2 4 h
s i R N A
M i g
r a t i o
n
***
***
***
8
1 0
1 2
1 4
1 6
Figure 6
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koroll, Hauke B. Werner, Klaus-Armin Nave and Jacqueline TrotterNivedita Chatterjee, Judith Stegmüller, Philipp Schätzle, Khalad Karram, Michael
precursor cellsInteraction of syntenin-1 and the NG2 proteoglycan in migratory oligodendrocyte
published online January 24, 2008J. Biol. Chem.
10.1074/jbc.M706074200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from