Embed Size (px)
Citation preview

lable at ScienceDirect
Journal of Human Evolution 74 (2014) 96e113
Contents lists avai
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
The late Early Pleistocene human dental remains from Uadi Aalad andMulhuli-Amo (Buia), Eritrean Danakil: Macromorphology andmicrostructure
Clément Zanolli a,*, Luca Bondioli b, Alfredo Coppa c, Christopher M. Dean d,Priscilla Bayle e, Francesca Candilio c, Silvia Capuani f, Diego Dreossi g, Ivana Fiore c,David W. Frayer h, Yosief Libsekal i, Lucia Mancini g, Lorenzo Rook j, Tsegai Medin Tekle i,k,Claudio Tuniz a,c, l, Roberto Macchiarelli m,n
aMultidisciplinary Laboratory, The ‘Abdus Salam’ International Centre for Theoretical Physics, Trieste, ItalybMuseo Nazionale Preistorico Etnografico ‘Luigi Pigorini’, Rome, ItalycDipartimento di Biologia Ambientale, Università di Roma ‘La Sapienza’, Rome, ItalydDepartment of Cell and Developmental Biology, University College London, UKeUMR 5199 PACEA, Université Bordeaux 1, FrancefCNR-IPCF, Dipartimento di Fisica, Università di Roma ‘La Sapienza’, Rome, Italyg Elettra-Sincrotrone Trieste S.C.p.A., SYRMEP Group, Basovizza, ItalyhDepartment of Anthropology, University of Kansas, Lawrence, USAiNational Museum of Eritrea, Asmara, EritreajDipartimento di Scienze della Terra, Università di Firenze, Italyk Institut Catala de Paleoecologia Humana i Evolució Social, Universitat Rovira i Virgili, Tarragona, SpainlCentre for Archaeological Science, University of Wollongong, AustraliamDépartement de Préhistoire, UMR 7194, MNHN, Paris, FrancenDépartement Géosciences, Université de Poitiers, France
a r t i c l e i n f o
Article history:Received 10 November 2013Accepted 22 April 2014Available online 19 May 2014
Keywords:BuiaHomo erectus/ergasterEast AfricaTeethExternal morphologyInternal structure
* Corresponding author.E-mail address: [email protected] (C. Za
http://dx.doi.org/10.1016/j.jhevol.2014.04.0050047-2484/� 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
Fieldwork performed during the last 15 years in various Early Pleistocene East African sites has signifi-cantly enlarged the fossil record of Homo erectus sensu lato (s.l.). Additional evidence comes from theDanakil Depression of Eritrea, where over 200 late Early to early Middle Pleistocene sites have beenidentified within a w1000 m-thick sedimentary succession outcropping in the Dandiero Rift Basin, nearBuia. Along with an adult cranium (UA 31), which displays a blend of H. erectus-like and derived morpho-architectural features and three pelvic remains, two isolated permanent incisors (UA 222 and UA 369)have also been recovered from the 1 Ma (millions of years ago) Homo-bearing outcrop of Uadi Aalad.Since 2010, our surveys have expanded to the nearby (4.7 km) site of Mulhuli-Amo (MA). This is afossiliferous area that has been preliminarily surveyed because of its exceptional concentration ofAcheulean stone tools. So far, the site has yielded 10 human remains, including the unworn crown of alower permanent molar (MA 93). Using diverse analytical tools (including high resolution mCT and mMRI),we analysed the external and internal macromorphology and microstructure of the three specimens, andwhenever possible compared the results with similar evidence from early Homo, H. erectus s.l.,H. antecessor, H. heidelbergensis (from North Africa), Neanderthals and modern humans. We also assessedthe UA 369 lower incisor from Uadi Aalad for root completion timing and showed that it compares wellwith data for root apex closure in modern human populations.
� 2014 Elsevier Ltd. All rights reserved.
nolli).
Introduction
Geological, paleontological and paleoanthropological fieldresearch has been carried out in the Dandiero Rift Basin, northernDanakil Depression of Eritrea since 1994. This research has led to

C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113 97
the discovery of over 200 late Early to early Middle Pleistocene siteswith widespread evidence of vertebrate faunal remains and lithicartefacts within the nearly 1000 m-thick fluvio-deltaic-lacustrinesedimentary succession outcropping 100 km south of Massawa(Abbate et al., 1998, 2004; Ghinassi et al., 2009; review in; Rooket al., 2012). Thus far, in an area covering about 40 km2, cranial,dental and postcranial human remains have been discovered in twolocalities near the Buia village: Uadi (Wadi) Aalad andMulhuli-Amo(Coppa et al., 2012).
The continental basin fill (the so-called Maebele Synthem)consists, from bottom to top, of six lithostratigraphic units: fluvialBukra sand and gravel, fluvio-deltaic Alat Formation, fluvial Warasand and gravel, lacustrine Goreya Formation; fluvio-deltaic Arosand; and alluvial fan Addai fanglomerate (Abbate et al., 2004;Papini et al., 2014). The Homo-bearing deposits belong to the up-per part of the 70e100 m-thick fluvio-deltaic Alat Formation(Ghinassi et al., 2009).
The upper part of the Alat Formation hosts the top of the Jar-amillo subchron (0.99 Ma), which occurs about 10 m below thetransition to the overlying Wara sand and gravel. The Matuyama-Brunhes boundary (0.78 Ma) is located about 5 m above the baseof the Aro sand unit (Albianelli and Napoleone, 2004). Confirmationof the magnetostratigraphically-based chronological setting comesfrom fission track dating (Bigazzi et al., 2004) and mammal bio-chronology (Ferretti et al., 2003; Martínez-Navarro et al., 2004,2010; Rook et al., 2010, 2012).
All human remains discovered so far occur in the upper part ofthe Alat Formation, where the transition from a deltaic to alluvialsetting is characterized by high frequency, potentially millennial-scale, lake-level oscillations (units DL5, FL2a and FL2b in Ghinassiet al., 2009). Specifically, the concentration of fossil bones at thebase of the FL2bwas associatedwith an increase in fluvial dischargethat caused winnowing of the immediately underlying fossil-bearing fluvio-deltaic deposits. As a whole, both the sedimentaryrecord documenting the evolution of fluvio-deltaic and lacustrinesystems (Ghinassi et al., 2009; Rook et al., 2012), and the vertebratefaunal assemblages predominated by taxa known for their waterdependence (Delfino et al., 2004; Martínez-Navarro et al., 2004),clearly indicate a relatively open paleoenvironmental scenariocharacterized by the presence of moist grassed habitats adjacent topersistent water (Rook et al., 2012).
First identified in the Uadi Aalad (UA) site, the 5e6 m-thickHomo-bearing layer produced a virtually complete adult craniumpreserving the face (UA 31), two permanent incisors (UA 222 andUA 369), and three pelvic portions (UA 173, UA 405, UA 466)(Abbate et al., 1998; Macchiarelli et al., 2004a; Bondioli et al., 2006).Compared with the Indonesian and Chinese Homo erectus sensustricto (s.s.) sample (review in Antón, 2003), as well as with Africanspecimens such as OH 9 and, to a lesser extent, KNM-ER 3733 (Baab,2008; Rightmire, 2013) and to the chronogeographically closecalvaria from Daka, Middle Awash (Gilbert and Asfaw, 2008), UA 31displays a blend of H. erectus/ergaster-like and derived morpho-architectural features more commonly found in Middle Pleisto-cene specimens. These features include a high positioning of themaximum parietal breadth, weak parietal keeling along themidline, slight parasagittal flattening, and from sub-vertical toslightly downwards converging parietal walls, documentingextensive variation in late Early Pleistocene East African Homo(Macchiarelli et al., 2004a).
Since 2010, the systematic survey of a fossiliferous area near tothe exceptionally preserved A006 site (the so-called ‘handaxesesplanade’) previously reported for its extensive concentration ofOldowan and Acheulean lithic tools (Martini et al., 2004), has led tothe discovery of new fossil human remains at theMulhuli-Amo (MA)locality, 4.7 km south of the UA Homo site (Coppa et al., 2012, 2014).
The nearly 15 m-thick sedimentary succession of Mulhuli-Amoconsists of deltaic and fluvial sediments. The lower interval (ca. 5m) consists of a sandy Gilbert-type delta deposit. The middle in-terval (5 m) is made of pedogenized muddy deposits with isolatedsandy fluvial channels responsible for the transport and accumu-lation of bone remains and stone tools in their basal parts. Finally,the 5 m-thick upper interval of the succession consists of fluvialchannelized gravelly sand capped with pedogenized mud. Even ifmost fossil remains from this site are spread on the eroded surface,most of the bones and Acheulean artefacts occur in situ at the baseof the third interval.
The human fossil-bearing levels of Uadi Aalad andMulhuli-Amoare remarkably similar in sedimentary facies and depositionalhistory (cf. Ghinassi et al., 2009; Rook et al., 2012), and areconsidered to sample the same stratigraphic horizon belonging tothe Alat. Besides the presence of reptile remains including croco-diles (Crocodylus niloticus), turtles (Pelusios sinuatus), and monitorlizards (Varanus niloticus), the vertebrate assemblage at Mulhuli-Amo consists of the same mammal taxa represented at UadiAalad (i.e., Elephas, Hippopotamus, Kolpochoerus, Kobus, Pelorovis).
A total of 10 cranial and dental human remains, likely from twoadults and one immature individual, have been collected so far atMulhuli-Amo. They consist of an isolated frontal fragment bearing athick right torus (MA 14) and eight parietal fragments (MA 64 andMA 88aef) associated with a temporal bone fragment (MA 89), allfrom a single adult cranium showing structural (thickness distri-bution) and architectural features (proportions, curvature) closelyfitting the morphology represented by UA 31. Finally, the assem-blage includes an isolated permanent lower molar crown (MA 93)(Coppa et al., 2014).
Compared with the Southeast Asian and Indonesian hypodigm(Indriati, 2004; Kaifu et al., 2005), the human dental assemblagefrom chronologically well-constrained Early Pleistocene Africansites (e.g., McDougall et al., 2012) is qualitatively and quantitativelyrelatively rich until about 1.4 Ma (millions of years ago) (cf. Wood,1991), while evidence from the later Early Pleistocene is scanty(Schwartz and Tattersall, 2003; Suwa et al., 2007; Brink et al., 2012).The material we examine here thus enlarges this sample and con-tributes to partially fill the gap still existing between the dentalrecords available so far for African H. erectus sensu lato (s.l.) on onehand, and Homo heidelbergensis on the other hand.
Building on the announcement (Abbate et al., 1998) and pre-liminary general description of the two incisors from Uadi Aalad,collected in 1995 and 1997, respectively (Macchiarelli et al., 2004a),and the molar crown fromMulhuli Amo, collected in 2011, here weprovide details of their external and internal morphology. Morespecifically, we compare the structure of the three Eritrean speci-mens sampling H. erectus/ergaster with some earlier and later hu-man taxa, particularly with the evidence from the early MiddlePleistocene North African sample of Tighenif, using a variety ofinvestigative approaches and analytical tools granting high reso-lution access to their microstructure (Zanolli and Mazurier, 2013).In doing so, we reveal for the first time the primitive versus derivednature of some tooth structural features in African H. erectus s.l.near the end of the Early Pleistocene and investigate their evolu-tionary polarity at a macroregional scale.
Materials and methods
The three fossil teeth from the Buia area represent an upper leftlateral permanent incisor (UA 222) and a lower left central per-manent incisor (UA 369) from the Homo site of Uadi Aalad (Abbateet al., 1998; Macchiarelli et al., 2004a), and a lower left M1/M2crown (MA 93) fromMulhuli-Amo (Coppa et al., 2012, 2014; Zanolliet al., 2013). The incisors preserve the crown and root, while the

C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e11398
molar is represented only by the crown. This material is perma-nently stored at the Geo-Paleontological Laboratory of the NationalMuseum of Eritrea.
Outer morphology
The degree of occlusal wear was assessed according to Smith(1984), with description of the nonmetric crown topographyfollowing Scott and Turner (1997), based on Arizona State Univer-sity Dental Anthropology System (ASUDAS) scores (Turner et al.,1991). Maximum mesiodistal (MD) and buccolingual (BL) crowndiameters (in mm) were measured on the original specimens withMitutoyo Digimatic calipers (to the nearest 0.1 mm) and then onthe unsmoothed virtual surfaces generated after segmentation ofthe microtomographic record of each specimen (see below). Sincethe MD dimension of both incisors is affected by interproximalwear, we limited the comparative analysis to the BL diameter. Forlinear measurements, the Eritrean specimens were variablycompared with a number of fossil and recent samples (Table 1).
Crown microwear
Microwear texture analysis was attempted on the three speci-mens following the standard procedures described by Mahoney(2006). The teeth were first cleaned with cotton soaked inalcohol andmolds of their crown surfaces were obtained using highresolution silicone impression material (Zhermack Elite HD). A cast
Table 1Fossil and recent comparative samples used for crown linear measurements.
Samples Tooth References
H. habilis-rudolfensisfrom East Africa (HHR)
UI2 Wood, 1991LI1 Stynder et al., 2001LMs Wood, 1991; Leakey et al., 2012
African H. erectus/ergaster fromKenya and Ethiopia (HEA)
UI2 Wood, 1991LI1 Stynder et al., 2001LMs Wood, 1991; Suwa et al., 2007
H. erectus from Georgia (HEG) UI2 Martinón-Torres et al., 2008LI1 Martinón-Torres et al., 2008LMs Martinón-Torres et al., 2008;
Lordkipanidze et al., 2013H. erectus from Java (HEJ) UI2 Grine and Franzen, 1994;
Zanolli, 2013LI1 Stynder et al., 2001LMs Wood, 1991; Widianto, 1993;
Kaifu et al., 2005; Zanolli, 2013H. erectus from China (HEC) UI2 Wood, 1991
LI1 Wood, 1991LMs Wood, 1991
H. antecessor from AtapuercaGran Dolina (HA)
UI2 Bermúdez de Castro et al., 1999LI1 Bermúdez de Castro et al., 1999LMs Bermúdez de Castro et al., 1999
North African H. heidelbergensis fromTighenif and Rabat (HHNA)
UI2 Martinón-Torres et al., 2008LI1 Stynder et al., 2001LMs Wood, 1991
European H. heidelbergensis fromAtapuerca Sima delos Huesos (HHE)
UI2 Martinón-Torres et al., 2012LI1 Martinón-Torres et al., 2012LMs Martinón-Torres et al., 2012
Neanderthals (NEA) UI2 Voisin et al., 2012LI1 Voisin et al., 2012LMs Voisin et al., 2012
Near Eastern early H. sapiensfrom Qafzeh (NEEHS)
UI2 Vandermeersch, 1981LI1 Vandermeersch, 1981LMs Vandermeersch, 1981
European early Upper Paleolithichumans (EUPH)
UI2 Frayer, 1977LI1 Hillson et al., 2010LMs Frayer, 1977
Recent (Holocene) humans (RH) UI2 Frayer, 1977LI1 Frayer, 1977LMs Frayer, 1977
of each specimen was produced using transparent epoxy resin(Hardrock 554, Phase Inc.). Positive transparent araldite replicaswere analysed using a stereomicroscope and the patterns ofmicrostriations evaluated by scanning electron microscopy (SEM)(EVO 60 Leitz GmbH). For the latter analysis, the replicas weresputter coated with gold by means of an Emitech K550X system.However, in contrast with the two incisors, MA 93 did not provideany observable details.
High resolution magnetic resonance micro-imaging and root growthassessment
To assess non-destructively the growth-related dentine micro-structures (daily von Ebner and long-period Andresen lines) insubfossil and fossil teeth (Bondioli et al., 2013), we developed a newmagnetic resonance microimaging (mMRI) analytical approach,based on previous successful high resolution MRI imaging ofmodern human teeth (Weiger et al., 2012). Indeed, techniques inmagnetic resonance imaging (MRI) have advanced rapidly in thepast two decades (Haack et al., 1999), and can now be applied inplace of more destructive methods. Magnetic resonance imagingutilizes the magnetic moment exhibited by many naturally occur-ring nuclei, with the proton in the hydrogen nucleus being ofparticular importance. Protons are highly abundant in all tissuesand have high magnetic resonance sensitivity. Since the composi-tion of most biological tissue is dominated by water, the differencesin the water environment of adjacent tissue structures can bemanipulated by MRI to effectively visualize tissue structure. Morerecently, magnetic resonance microimaging (mMRI) methods havebeen developed and refined (Callaghan, 1995) to provide imagescharacterized by a spatial resolution of approximately 100 mm orsmaller. They have been particularly useful for investigating poroussystems (Strange et al., 1993; Allen et al., 1997) and bones (De Santiset al., 2010; Wehrli, 2013). Specifically, mMRI techniques are able toretrieve high resolution information about the density, the meandiameter, and the distribution of microfeatures in porous systems.Indeed, when the pores of a porous system, like the dentine, arefilled with a liquid containing protons (usually water), it is possibleto obtain indirect information about the microstructural charac-teristics of this material from the signal of the liquid entrapped inthe microstructures. Currently, MRI is rarely used in dental diag-nosis and research mainly because of the very short transverserelaxation times of the mineralized tooth tissues.
We performed a preliminary validation test on two perfectlypreserved Neolithic permanent teeth (an upper incisor and acanine), after comparatively assessing their root microstructure(dentine tubules and Andresen lines) by means of mCT, mMRI andhistology (Bondioli et al., 2013). We applied this technique tovisualize the dentine microanatomy and tentatively assess thegrowth pattern in UA 369. To do so, this well preserved lower leftcentral permanent incisor was immersed in a thin glass walled tubeused to contain samples in nuclear magnetic resonance spectros-copy, which had been filled with distilled water at the temperatureof 18 �C. Images of UA 369 were obtained by using Multi Slice MultiEcho (MSME) imaging sequences with repetition times (TR) equalto 380.0 ms, echo times (TE) equal to 3.5 ms, field of view (FOV)equal to 14 � 14 mm2, matrix 256 � 256, and number of scansaveraged (ns) equal to 1024. A total of 11 slices were selected of slicethickness (STH) equal to 200 mm to investigate the central zone ofUA 369 dentine. The in-plane resolution in each image slice was55 � 55 mm2.
All measurements were performed on a Bruker 9.4T Avance-400system located at the Physics Department of the ‘La Sapienza’University of Rome, operating with a micro-imaging probe (12 mminternal diameter bore) and equipped with a gradient unit

Table 2Linear, surface, and volumetric variables used for assessing internal structuralvariation and molar crown tissue proportions.
Variables Definitions
c The enamel area assessed on a buccolingual virtualsection (mm2)
b The dentine area under the enamel surface on abuccolingual virtual section (mm2)
e The enamel-dentine junction (EDJ) length on abuccolingual virtual section (mm)
AET (¼c/e) The bi-dimensional average enamel thickness (mm)RET (¼100*
AET/(b1/2))The scale-free, bi-dimensional relative enamelthickness (Martin, 1985)
Ve The volume of the enamel cap (mm3)Vcd The volume of the coronal dentine (mm3)Vcp The volume of the coronal pulp (mm3)Vcdp The volume of the coronal dentine, including the
coronal aspect of the pulp chamber (mm3)Vc The total crown volume, including enamel, dentine, and
pulp (mm3)SEDJ The EDJ surface (mm2)Vcdp/Vc
(¼100*Vcdp/Vc)The percent of coronal volume that is dentine and pulp(%)
3D AET (¼Ve/SEDJ) The three-dimensional average enamel thickness (mm)3D RET (¼100*3D
AET/(Vcdp1/3))The scale-free three-dimensional relative enamelthickness (see Kono, 2004; Olejniczak et al., 2008a)
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113 99
characterized by a maximum gradient strength of 1200 mT/m, anda rise time of 100 ms.
Root extension rate was calculated from the mMRI images ac-cording to the method detailed in Macchiarelli et al. (2006), Dean(2009), and Dean and Cole (2013), which can be applied to bothfossil and extant human teeth. Continuous data were collected onthe lingual and labial aspects along almost the entire root length ofUA 369. After outlining themain visible growth incremental lines onthe image of this specimen, dentine tubules were traced from theroot surface just beneath the last formed enamel at the cervix (pointA) to a second point 200 mm along the dentine tubules into rootdentine (point B). This thickness of dentine forms at a rate of 2.23e2.50 mm/day and takes ca. 80e90 days to form (Dean, 1998, 1999,2012; Dean and Cole, 2013). A line parallel with the orientation ofthe growthmarkerswas drawn from the end (point B) of the 200 mmsegment back to the root surface (point C). The distance betweenpoint A andpoint C represents the length that the root had extendedin ca. 80e90 days. An extension rate was calculated in mm/day forthis distance along the root. This procedure was then repeated suchthatpoint CbecamepointA in each calculation along the root length.
In H. erectus from Java (S7-37), direct measurements of dailydentine increments in an upper M1 equalled 1.43 mm per dayover the first 50 mm, 2.46 mm per day between 50 and 100 mm,and 2.71 mm per day between 100 and 200 mm from the cement-dentine junction (CDJ) (Dean et al., 2001; Dean, 2009). The overallmean daily rate over 200 mm of dentine formation from the CDJ(point A to point B) was 2.4 mm per day in this upper M1, but noequivalent data exist for anterior teeth in African H. erectus.Previously, a rate of 2.5 mm per day has been used to calculate 80days formation for a 200 mm thickness of newly formed rootdentine in modern human teeth. However, Smith et al. (2007)have argued that in some teeth of Pan troglodytes dentine for-mation rates average closer to 2 mm per day at the cervix. Dean(2009) and Dean and Cole (2013) also observed rates ofw2.0 mm per day in some modern human anterior teeth. Becauseit remains unclear what the exact range of daily dentine forma-tion rates is likely to have been in UA 369, we have adopted acautious approach and have used both 80 and 90 day formationtimes (equivalent to 2.2e2.5 mm per day) to provide what we feelis the most likely range of estimates for the time taken to formconsecutive lengths of root along the CDJ.
Microtomographic-based inner structure
The three specimens were detailed using X-ray micro-tomography (mCT) at the SYRMEP beamline (crown portion of thetooth) and at the Tomolab station (whole tooth) of the ElettraSincrotrone Trieste laboratory (Basovizza, Italy). Themeasurementsperformed on the whole tooth were carried out according to thefollowing parameters: 100 kV voltage, 80 mA current, 4.0 s expo-sition time per projection, and a projection each 0.25�, for the in-cisors; 130 kV, 61 mA, 4.5 s, and 0.15� for MA 93. Their final volumeswere reconstructed with an isotropic voxel size of 10.0 and 8.3 mm,respectively. Using Amira v.5.3 (Visualization Sciences Group Inc.)and ImageJ (Schneider et al., 2012), a semi-automatic, threshold-based segmentation was carried out following the half-maximumheight method (HMH; Spoor et al., 1993) and the region of inter-est thresholding protocol (ROI-Tb; Fajardo et al., 2002) takingrepeated measurements on different slices of the virtual stack(Coleman and Colbert, 2007).
Besides the pulp chamber volume of the incisors, 14 linear,surface, and volumetric variables describing internal structuralvariation and tissue proportions were digitally measured (orderived) on MA 93 (Table 2). Intra- and inter-observer tests formeasurement accuracy run by two observers revealed differencesless than 4%, which is similar to previous analyses (e.g., Macchiarelliet al., 2008; Bondioli et al., 2010; Zanolli et al., 2012).
Observations on the pulp cavity of the two incisors from UadiAalad were compared with similar evidence fromH. heidelbergensisfrom Tighenif (isolated lower I1; Zanolli, 2011; Zanolli andMazurier, 2013), Neanderthals (Ehringsdorf G3 [upper I2] andRegourdou 1 [lower I1]; Macchiarelli et al., 2013; NESPOS Database,2013); and recent humans (original data).
To assess site-specific variation in their radicular dentinethickness, ad hoc imaging techniques were used to virtually un-roll the roots of the two incisors (15e85% portion of the total rootlength) and to generate their morphometric maps (Bayle, 2008;Bondioli et al., 2010; Bayle et al., 2011; Macchiarelli et al.,2013). For comparative purposes, we also considered thefollowing specimens: the upper I2 Javanese H. erectus specimenMI92.2 (HEJ: Zanolli, 2011); the North African H. heidelbergensisisolated lower I1 from Tighenif (HHNA: Zanolli, 2011; Zanolli andMazurier, 2013); the two Neanderthal specimens Ehringsdorf G3(upper I2) and Regourdou 1 (lower I1) (NEA: Macchiarelli et al.,2013; NESPOS Database, 2013); and two recent human incisors(EH: original data).
In all three Eritrean specimens, enamel thickness topographicdistribution was rendered through a 3D map generated using achromatic scale where thickness increases from thin (dark blue) tothick (red) (Macchiarelli et al., 2008, 2013; Bayle et al., 2011). Forcomparative purposes, similar cartographies have been generatedfor a selected number of specimens representing H. erectus fromSangiran (MI92.2 [upper I2] and NG0802.3 [lower M2]; Zanolli,2011, 2013); North African H. heidelbergensis (isolated lower I1and Tighenif 2 [lower M1 and M2]; Zanolli, 2011; Zanolli andMazurier, 2013); Neanderthals (Krapina D122 [upper I2], Regour-dou 1 [lower I1], La Chaise de Vouthon S5 [lower M1], Krapina D10[lower M2]; Macchiarelli et al., 2013; NESPOS Database, 2013); andrecent humans of European origin (original data).
The results of the 2e3D virtual analyses performed on MA 93were variably compared with a number of fossil and recent samples(Table 3).
Nonmetric features revealed by virtual imaging at the EDJ of MA93 were scored according to Skinner et al. (2008a) for the accessorycusps, Bailey et al. (2011) and Martínez de Pinillos et al. (2014) forthe trigonid crest pattern (see also Macchiarelli et al., 2006), andMartinón-Torres et al. (2014) for the talonid crest expression.

Table 3Fossil and recent comparative samples used for assessing 2e3D molar crown tissue proportions.
Specimens/samples References
H. habilis-rudolfensis from East Africa (HHR) Smith et al., 2012South African early Homo (SAEH) Smith et al., 2012Javanese H. erectus (HEJ) Zanolli, 2011North African H. heidelbergensis from Tighenif (HHNA) Zanolli, 2011; Zanolli and Mazurier, 2013Neanderthals (NEA) Olejniczak et al., 2008a; Kupczik and Hublin, 2010;
Smith et al., 2012; Macchiarelli et al., 2013; NESPOS Database, 2013North African Aterians (AFMH) Kupczik and Hublin, 2010African-Near Eastern anatomically modern fossil humans
(ANEFMH) from Morocco (Dar es-Soltane, Grotte desContrebandiers, Jebel Irhoud), South Africa (Die KeldersCave, Equus Cave) and Israel (Qafzeh)
Smith et al., 2012
The European early Upper Paleolithic specimenfrom Lagar Velho (EUPH)
Bayle et al., 2010
Recent humans (RH) Olejniczak et al., 2008a; Smith et al., 2012; original data
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113100
Geometric morphometric analyses
Comparative geometric morphometric analyses of the MA 93EDJ were performed on the unsmoothed reconstructed virtualsurfaces by placing a total of seven landmarks on the apex of theprotoconid, metaconid, entoconid and hypoconid, and at eachintermediate lowest point between two dentine horns along thedentine marginal ridge, except between the two distal horns (seeZanolli et al., 2012). Because of the variable presence of thehypoconulid, this latter cusp and the distal marginal ridge werenot considered (Zanolli and Mazurier, 2013). By using the packageMorpho 0.24.1 (Schlager, 2013) for R v.3.0.1 (R Development CoreTeam, 2013), we performed a generalized Procrustes analysis(GPA) and a between-group principal component analysis(bgPCA) based on the Procrustes shape coordinates (Mitteroeckerand Bookstein, 2011). For the specific purposes of this analysis,MA 93 was compared with a number of fossil and recent samples(Table 4).
Results
Description of the specimens
UA 222 (Uadi Aalad) This upper left lateral permanent incisor(ULI2) from an adult individual is complete from crown to root(Fig. 1A). Macroscopically, there is some evidence of weathering,root etching and sediment scratching. From the mCT record, it alsoappears that the pulp cavity is filled with homogeneous fine-grained sediment. The tooth shows advanced occlusal wear (stage4), with functional enamel loss and extensive emerging dentinecreating a slightly labiolingual to mesiodistal oriented bevel.Interproximal wear is more marked distally, where thefacet almost reaches the CEJ. While the occlusal surface issmooth, with rare pits and few microstriations, SEM analysis of
Table 4Fossil and recent comparative samples used for in the geometric morphometric analyse
Samples Sites Lower
Javanese H. erectusa SangiranNorth African H. heidelbergensisa TighenifNeanderthals Krapina KRD77, KRD7
KRD81, KRD1La Chaise-de-Vouthon S5, S14e7, S4RegourdouEhringsdorf Ehringsdorf I
Recent humans 14
a Added a posteriori in the bgPCA analysis.
its labial and, to a minor extent, lingual crown aspects revealed anumber of fine striations covering the entire surfaces. Thescratches, which can be interpreted as microwear traces (cf.Ungar et al., 2008; Krueger and Ungar, 2010), are mostly verticalor slightly oblique (85e90� range), sub-parallel and very thin (2e3 mm) (Fig. 2A). However, some larger furrows show thecharacteristics of trampling damage, with broad entry,rectangular section, and narrow end (Fiore et al., 2004).
The uncorrected maximum MD crown diameter is 6.7 mm,while the BL diameter is 7.2 mm (contra Macchiarelli et al., 2004a;see Table 5). The root, which is complete with a closed apex dis-playing a thin cementum deposit on virtual sections, exhibits lon-gitudinal grooves, the distal one being deeper. From the lowestenamel cervical point to its apex, root length is 18.8 mm.
The enamel shows no evidence of linear enamel hypoplasia,hypocalcifications or macro-defects, and no calculus or patholog-ical lesions are noticeable. While part of its crown has beenremoved by extensive wear, based on the preserved enamel UA 222lacks shovelling (grade 0e1), an interruption groove and a distincttuberculum dentale, but does show labial convexity (Fig. 1A) andalso possesses a longitudinal lingual groove that is slightly deviatedmesially (cf. Martinón-Torres et al., 2012), reaching the wavy rim ofthe preserved occlusal-lingual margin.
Microtomographic-based virtual sections reveal relatively thinenamel (maximum 0.66 mm). The EDJ perfectly mirrors the outermorphology. Despite a slight collar constriction from its occlusalaspect to mid-root level, the pulp cavity shows a slightly labiolin-gually compressed ovoid outline. Conversely, its apical portion ismesiodistally flattened, with marked labiolingual expansion(Fig. 3A). Its total volume is 30.7 mm3. The standardized morpho-metric map of its virtually unrolled root reveals an area of abso-lutely thicker dentine (maximum 3.3 mm) on the labial aspect nearthe cervix, while thinner dentine is deposited on two vertical stripsrunning along the mesial and distal aspects (Fig. 4A).
s of the molar EDJ.
M1 Lower M2 References
NG0802.3 Zanolli, 2011Tighenif 2 Zanolli and Mazurier, 2013
9, KRD80,05
KRD1, KRD6, KRD10,KRD86, KRD104, KRD107
NESPOS Database, 2013
9, BDJ4C9Regourdou 1
17 Original data

Figure 1. The upper left I2 UA 222 (A) and the lower left I1 UA 369 (B) from Uadi Aalad, and the lower left M1/M2 MA 93 from Mulhuli-Amo (C). For each specimen, the imagesshow: the original (in lingual and mesial views for UA 222 and UA 369, in occlusal and buccal views for MA 93); a mesiodistal (left) and a labiolingual (right) virtual section for UA222 and UA 369, and two virtual sections for MA 93 respectively passing through the mesial (upper) and the buccal cusps (lower); the microtomographic-based reconstruction of(from the left): the outer surface (the enamel in red), the dentine (yellow), the occlusal view (for the two incisors) and, uniquely for MA 93, the pulp cavity (cyan) (same views as theoriginals). b, buccal; l, lingual; m, mesial; o, occlusal. Scale bars, 1 cm. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version ofthis article.)
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113 101
UA 369 (Uadi Aalad) This specimen represents a complete lowerleft central incisor (LLI1) from an adult. Serial virtual slices reveal apulp chamber filled almost completely by a matrix, which occa-sionally causes contrast attenuation with the dentine (Fig. 1B). UA369 also exhibits advanced occlusal wear (stage 4; Smith, 1984),with a major mesiodistal to labiolingual orientation resulting in aweakly concave occlusal surface whose enamel rim is thickerlabially (0.9 mm versus 0.4 mm lingually). Interproximal wear ismore marked mesially. Under the stereomicroscope, the occlusalsurface is smooth, with rare pits and few microstriations. TheSEM imaging shows a microwear pattern similar to that seen inUA 222 (Fig. 2B). Indeed, mastication-related pits and scratches,as well as sediment-induced striae and furrows, are present onboth labial and lingual crown aspects, the former havingundergone some etching-like process that obscures part of thefeatures, particularly near the occlusal border. The majority of thevertical, sub-parallel and thin striae (2e3 mm) correspond tomicrowear traces (Teaford, 2007; Ungar et al., 2008; Hlusko et al.,2013), while few longer (1e2 mm), slightly thicker (3e10 mm)and randomly oblique scratches more likely result from theabrasive action of variably sized sediment particles. Labially, closeto the CEJ, there are four long and relatively deep furrows oftaphonomic origin, with smooth walls and rounded bottom.Although the general microfeature pattern does not differ fromUA 222, some of the scratches occurring on both faces indicatethat UA 369 may have experienced slightly different taphonomicmicro-dynamics.
The uncorrected maximum mesiodistal diameter correspondsto 5.2 mm and the labiolingual diameter to 6.7 mm (Table 5).Similar to UA 222, no linear enamel hypoplasia, hypocalcifications,macro-defects, calculus deposits or pathological lesions arenoticeable on this specimen, and the preserved portion of thecrown shows no developed relief on its lingual and labial aspects, orshovelling.
Measured on virtual sections, maximum enamel radial thick-ness reaches 0.8 mm. Internally, the dentine crown surface caststhe outer morphology. Also in this lower incisor the upper portionof the pulp cavity, whose total volume corresponds to 15.9 mm3,shows a labiolingually compressed ovoid outline, followed by amarked mid-root labiolingual expansion and a mesiodistally flat-tened apical root morphology (Fig. 3B).
Well-developed longitudinal grooves run along both the mesialand distal radicular aspects, whose maximum length reaches16.3 mm. As revealed by its profile (Fig. 4B), thicker root dentine inthis lower incisor appears on the cervical half of the labial andlingual aspects (maximum 2.9 mm), while the thinnest values arefound along the apical third of the mesial and distal faces.
For magnetic resonance microimaging-based growth assess-ment, a total of 20 consecutive measurements were possible onboth lingual and buccal aspects of the root in UA 369 (Fig. 5A), butadditional measurements became too difficult close to the rootapex beyond 12.5 mm on the buccal aspect, and 13.5 mm on thelingual aspect of the root. The overall mean buccal extension rate(n ¼ 20 estimates) was 8.0 mm per day (range 3.0e10.8 mm per day)

Figure 2. SEM micrographs of the labial aspect of UA 222 (A) and UA 369 (B). Mostlyvertically-oriented thin microfeatures are visible on the central portion of the UA 222crown (A), where the two larger and deeper furrows crossing near the centre of theimage result from sediment abrasion. Numerous thin microfeatures are also present onthe centre-mesial area spot sampling the UA 369 crown (B), together with few obliquesediment traces and an etching damage on the upper right corner of the image. Scalebars, 100 mm.
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113102
and themean lingual extension ratewas 7.8 mmper day (range 4.9e10.0 mm per day). In both roots there was an early peak in rootextension rate (10.0 and 10.8 mm per day, respectively) between1.32 and 1.48 years post enamel completion.
These results (Fig. 5B) are presented first as cumulative plots ofincreasing lingual and buccal root length against root formationtime compared with data for a sample of 44 modern human incisortooth roots taken fromDean and Cole (2013). A second plot (Fig. 5C)puts root growth into context with crown growth for illustrativepurposes. Root formation in UA 369 was compared with data forcrown growth in the same modern human sample (Dean and Cole,2013). It was assumed that the unworn crown height in UA 369would have been close to that of other unworn lower incisorsattributed to H. erectus s.l., a good example of which is the lowercentral incisor of KNM-WT 15000 that measures 9.75 mm. Usingthis tooth to provide an estimate of the crown formation time in UA369 suggests that it would have taken close to 2.83 years to com-plete crown formation based on a nine day periodicity betweenadjacent long-period striae of Retzius in the enamel of the Eritrean
specimen (Dean et al., 2001: Table 1). These estimates of crownheight and crown formation time allow the plots for UA 369 to bevisualized alongside the modern human sample and show thatthere is little effect (compared with Fig 5B) on how rates of rootformation fall within the modern sample. Both plots (Fig. 5B and C)show that root formation in UA 369 could be reconstructed overapproximately five years.MA 93 (Mulhuli-Amo) This is a nearly unworn lower left first orsecond permanent molar crown (LLM1/M2) of a juvenile individual(Fig. 1C). The buccal and lingual surfaces are slightly etched fromroots and sediment, resulting in a coarse enamel texture. On thedistolingual surface, a shiny area of better preserved enamelmimics an interproximal facet, but its lingually displaced positionand its broad extension (larger than the mesial interproximalfacet) support a taphonomic, non-functional origin. While someshallow furrows are visible on the lingual aspect, the presence oflinear enamel hypoplasia cannot be confirmed here. Conversely,some hints of widely spaced perikymata are still visible in thecentre of the distal face, where the enamel remains least worn. Inthis position, there is indeed a weak expression of enamelhypoplasia, supporting a similar conclusion for the lingual aspect.The hypoplasia would have occurred during crown formation.
The rather squat crown reaches 6.1 mm in height at the meta-conid and 6.2 mm at the protoconid. It shows a rectangular outlinewith a length (MD) of 13.9 mm and a breadth (BL) of 11.9 mm(Table 5). A slightly polished wear facet on the protoconid and twosmaller facets on the entoconid and on the hypoconulid indicatethat the tooth had begun functional occlusion, but in the earlieststage (stage 1). A small, flat and ovoid interproximal mesial contactfacet (2.9 � 2.3 mm) is present, but there is no distal facet. Themesial wall converges cervically, while the opposite is true buccally,the lingual and distal walls being relatively vertical. The sub-horizontal cervico-enamel line is almost completely preservedand the fracture plane in the inferior part appears to follow theincremental mineralizing front, which suggests that the tooth wasat the crown complete stage of formation.
There are five main cusps, including a medium-large hypo-conulid (C5; stage 4), plus a small tuberculum intermedium (C7;stage 2), but no tuberculum sextum (C6). Relative cusp size isprotoconid > metaconid > hypoconid > entoconid > C5 > C7. Theocclusal grooves connect in a þ fissure pattern. Compared with theincised distal marginal ridge, the mesial margin is not crossed bythe central groove and is proportionally thicker and higher, forminga deep elongated mesial fovea (stage 3). Immediately behind thefovea, an uninterrupted sharp mid-trigonid crest (score 1A) linksthe two mesial cusps. Two distal crests depart from the protoconidand the metaconid, respectively. They converge in the trigonidbasin, but are interrupted in the centre by themain groove. Becausethey do not connect, the latter can be interpreted as an incompletedistal crest. Between the mid-trigonid crest and the distal trigonid-like feature, a short crest originating from the metaconid anglestowards the occlusal basin. No expression of a deflecting wrinkle isfound. The protostylid is absent and the intercuspal groove isrelatively shallow.
At the EDJ level, MA 93 exhibits a modestly elevated topography,with relatively low but sharp dentine horns systematically under-lying the blunter outer cusps, even at the level of the metaconulid-type C7 (Skinner et al., 2008a), which is located on the distalshoulder of the metaconid dentine horn and is slightly betterexpressed than at the outer surface. While on the occlusal surfacethe mid-trigonid crest is complete but slightly dipping at thesagittal sulcus level, its EDJ expression is continuous and of regularheight (grade 3; Bailey et al., 2011), originating slightly mesial fromthe metaconid dentine horn and joining the protoconid dentinemiddle lobe segment (middle-mesial configuration). Together with
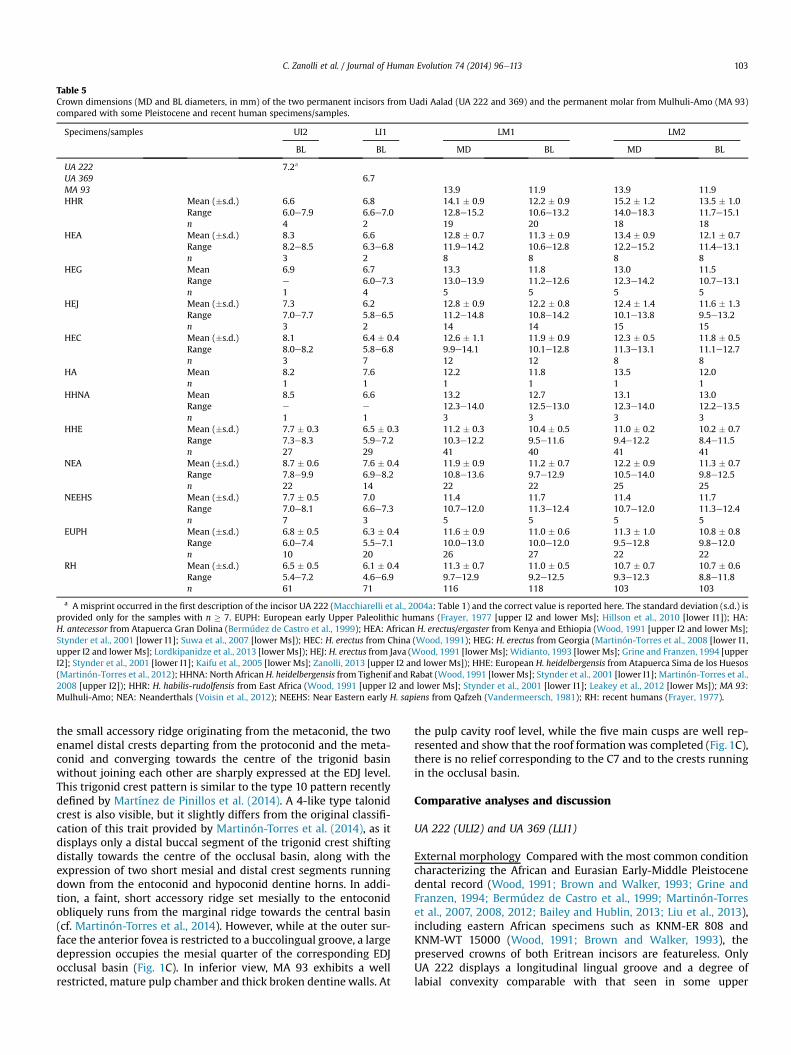
Table 5Crown dimensions (MD and BL diameters, in mm) of the two permanent incisors from Uadi Aalad (UA 222 and 369) and the permanent molar from Mulhuli-Amo (MA 93)compared with some Pleistocene and recent human specimens/samples.
Specimens/samples UI2 LI1 LM1 LM2
BL BL MD BL MD BL
UA 222 7.2a
UA 369 6.7MA 93 13.9 11.9 13.9 11.9HHR Mean (�s.d.) 6.6 6.8 14.1 � 0.9 12.2 � 0.9 15.2 � 1.2 13.5 � 1.0
Range 6.0e7.9 6.6e7.0 12.8e15.2 10.6e13.2 14.0e18.3 11.7e15.1n 4 2 19 20 18 18
HEA Mean (�s.d.) 8.3 6.6 12.8 � 0.7 11.3 � 0.9 13.4 � 0.9 12.1 � 0.7Range 8.2e8.5 6.3e6.8 11.9e14.2 10.6e12.8 12.2e15.2 11.4e13.1n 3 2 8 8 8 8
HEG Mean 6.9 6.7 13.3 11.8 13.0 11.5Range e 6.0e7.3 13.0e13.9 11.2e12.6 12.3e14.2 10.7e13.1n 1 4 5 5 5 5
HEJ Mean (�s.d.) 7.3 6.2 12.8 � 0.9 12.2 � 0.8 12.4 � 1.4 11.6 � 1.3Range 7.0e7.7 5.8e6.5 11.2e14.8 10.8e14.2 10.1e13.8 9.5e13.2n 3 2 14 14 15 15
HEC Mean (�s.d.) 8.1 6.4 � 0.4 12.6 � 1.1 11.9 � 0.9 12.3 � 0.5 11.8 � 0.5Range 8.0e8.2 5.8e6.8 9.9e14.1 10.1e12.8 11.3e13.1 11.1e12.7n 3 7 12 12 8 8
HA Mean 8.2 7.6 12.2 11.8 13.5 12.0n 1 1 1 1 1 1
HHNA Mean 8.5 6.6 13.2 12.7 13.1 13.0Range e e 12.3e14.0 12.5e13.0 12.3e14.0 12.2e13.5n 1 1 3 3 3 3
HHE Mean (�s.d.) 7.7 � 0.3 6.5 � 0.3 11.2 � 0.3 10.4 � 0.5 11.0 � 0.2 10.2 � 0.7Range 7.3e8.3 5.9e7.2 10.3e12.2 9.5e11.6 9.4e12.2 8.4e11.5n 27 29 41 40 41 41
NEA Mean (�s.d.) 8.7 � 0.6 7.6 � 0.4 11.9 � 0.9 11.2 � 0.7 12.2 � 0.9 11.3 � 0.7Range 7.8e9.9 6.9e8.2 10.8e13.6 9.7e12.9 10.5e14.0 9.8e12.5n 22 14 22 22 25 25
NEEHS Mean (�s.d.) 7.7 � 0.5 7.0 11.4 11.7 11.4 11.7Range 7.0e8.1 6.6e7.3 10.7e12.0 11.3e12.4 10.7e12.0 11.3e12.4n 7 3 5 5 5 5
EUPH Mean (�s.d.) 6.8 � 0.5 6.3 � 0.4 11.6 � 0.9 11.0 � 0.6 11.3 � 1.0 10.8 � 0.8Range 6.0e7.4 5.5e7.1 10.0e13.0 10.0e12.0 9.5e12.8 9.8e12.0n 10 20 26 27 22 22
RH Mean (�s.d.) 6.5 � 0.5 6.1 � 0.4 11.3 � 0.7 11.0 � 0.5 10.7 � 0.7 10.7 � 0.6Range 5.4e7.2 4.6e6.9 9.7e12.9 9.2e12.5 9.3e12.3 8.8e11.8n 61 71 116 118 103 103
a A misprint occurred in the first description of the incisor UA 222 (Macchiarelli et al., 2004a: Table 1) and the correct value is reported here. The standard deviation (s.d.) isprovided only for the samples with n � 7. EUPH: European early Upper Paleolithic humans (Frayer, 1977 [upper I2 and lower Ms]; Hillson et al., 2010 [lower I1]); HA:H. antecessor from Atapuerca Gran Dolina (Bermúdez de Castro et al., 1999); HEA: African H. erectus/ergaster from Kenya and Ethiopia (Wood, 1991 [upper I2 and lower Ms];Stynder et al., 2001 [lower I1]; Suwa et al., 2007 [lower Ms]); HEC: H. erectus from China (Wood, 1991); HEG: H. erectus from Georgia (Martinón-Torres et al., 2008 [lower I1,upper I2 and lowerMs]; Lordkipanidze et al., 2013 [lower Ms]); HEJ: H. erectus from Java (Wood, 1991 [lower Ms];Widianto, 1993 [lower Ms]; Grine and Franzen, 1994 [upperI2]; Stynder et al., 2001 [lower I1]; Kaifu et al., 2005 [lower Ms]; Zanolli, 2013 [upper I2 and lower Ms]); HHE: European H. heidelbergensis from Atapuerca Sima de los Huesos(Martinón-Torres et al., 2012); HHNA: North African H. heidelbergensis from Tighenif and Rabat (Wood, 1991 [lower Ms]; Stynder et al., 2001 [lower I1]; Martinón-Torres et al.,2008 [upper I2]); HHR: H. habilis-rudolfensis from East Africa (Wood, 1991 [upper I2 and lower Ms]; Stynder et al., 2001 [lower I1]; Leakey et al., 2012 [lower Ms]); MA 93:Mulhuli-Amo; NEA: Neanderthals (Voisin et al., 2012); NEEHS: Near Eastern early H. sapiens from Qafzeh (Vandermeersch, 1981); RH: recent humans (Frayer, 1977).
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113 103
the small accessory ridge originating from the metaconid, the twoenamel distal crests departing from the protoconid and the meta-conid and converging towards the centre of the trigonid basinwithout joining each other are sharply expressed at the EDJ level.This trigonid crest pattern is similar to the type 10 pattern recentlydefined by Martínez de Pinillos et al. (2014). A 4-like type talonidcrest is also visible, but it slightly differs from the original classifi-cation of this trait provided by Martinón-Torres et al. (2014), as itdisplays only a distal buccal segment of the trigonid crest shiftingdistally towards the centre of the occlusal basin, along with theexpression of two short mesial and distal crest segments runningdown from the entoconid and hypoconid dentine horns. In addi-tion, a faint, short accessory ridge set mesially to the entoconidobliquely runs from the marginal ridge towards the central basin(cf. Martinón-Torres et al., 2014). However, while at the outer sur-face the anterior fovea is restricted to a buccolingual groove, a largedepression occupies the mesial quarter of the corresponding EDJocclusal basin (Fig. 1C). In inferior view, MA 93 exhibits a wellrestricted, mature pulp chamber and thick broken dentine walls. At
the pulp cavity roof level, while the five main cusps are well rep-resented and show that the roof formationwas completed (Fig. 1C),there is no relief corresponding to the C7 and to the crests runningin the occlusal basin.
Comparative analyses and discussion
UA 222 (ULI2) and UA 369 (LLI1)
External morphology Compared with the most common conditioncharacterizing the African and Eurasian Early-Middle Pleistocenedental record (Wood, 1991; Brown and Walker, 1993; Grine andFranzen, 1994; Bermúdez de Castro et al., 1999; Martinón-Torreset al., 2007, 2008, 2012; Bailey and Hublin, 2013; Liu et al., 2013),including eastern African specimens such as KNM-ER 808 andKNM-WT 15000 (Wood, 1991; Brown and Walker, 1993), thepreserved crowns of both Eritrean incisors are featureless. OnlyUA 222 displays a longitudinal lingual groove and a degree oflabial convexity comparable with that seen in some upper
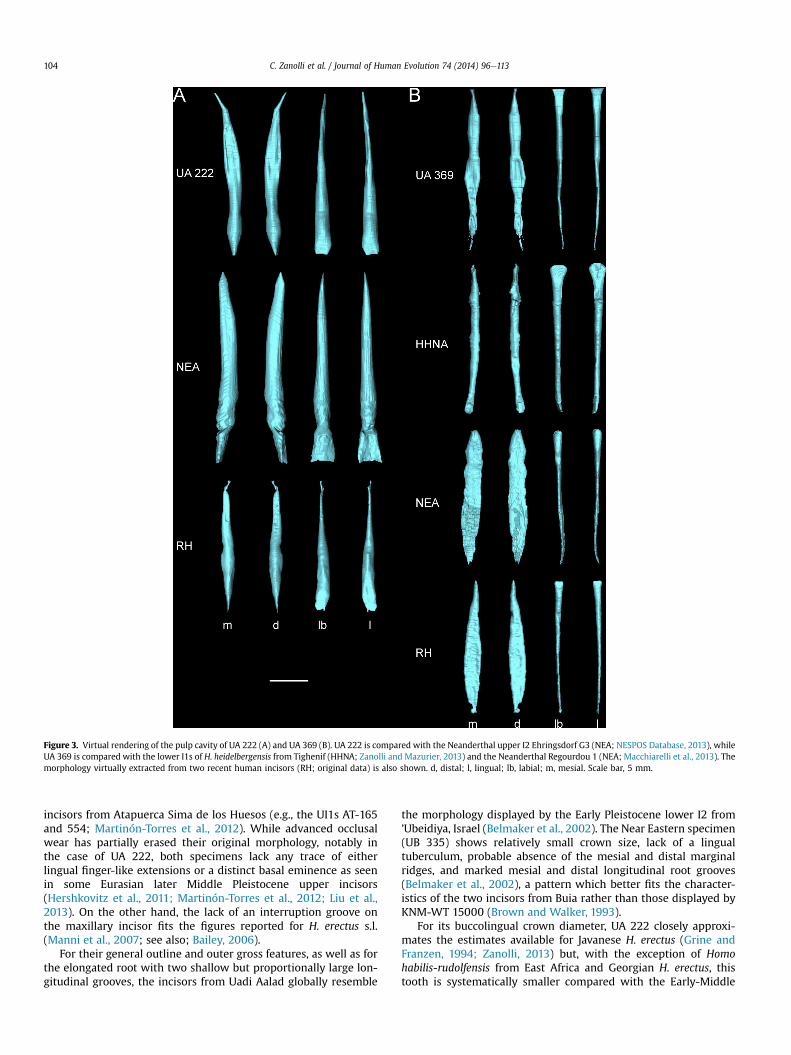
Figure 3. Virtual rendering of the pulp cavity of UA 222 (A) and UA 369 (B). UA 222 is compared with the Neanderthal upper I2 Ehringsdorf G3 (NEA; NESPOS Database, 2013), whileUA 369 is compared with the lower I1s of H. heidelbergensis from Tighenif (HHNA; Zanolli and Mazurier, 2013) and the Neanderthal Regourdou 1 (NEA; Macchiarelli et al., 2013). Themorphology virtually extracted from two recent human incisors (RH; original data) is also shown. d, distal; l, lingual; lb, labial; m, mesial. Scale bar, 5 mm.
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113104
incisors from Atapuerca Sima de los Huesos (e.g., the UI1s AT-165and 554; Martinón-Torres et al., 2012). While advanced occlusalwear has partially erased their original morphology, notably inthe case of UA 222, both specimens lack any trace of eitherlingual finger-like extensions or a distinct basal eminence as seenin some Eurasian later Middle Pleistocene upper incisors(Hershkovitz et al., 2011; Martinón-Torres et al., 2012; Liu et al.,2013). On the other hand, the lack of an interruption groove onthe maxillary incisor fits the figures reported for H. erectus s.l.(Manni et al., 2007; see also; Bailey, 2006).
For their general outline and outer gross features, as well as forthe elongated root with two shallow but proportionally large lon-gitudinal grooves, the incisors from Uadi Aalad globally resemble
the morphology displayed by the Early Pleistocene lower I2 from’Ubeidiya, Israel (Belmaker et al., 2002). The Near Eastern specimen(UB 335) shows relatively small crown size, lack of a lingualtuberculum, probable absence of the mesial and distal marginalridges, and marked mesial and distal longitudinal root grooves(Belmaker et al., 2002), a pattern which better fits the character-istics of the two incisors from Buia rather than those displayed byKNM-WT 15000 (Brown and Walker, 1993).
For its buccolingual crown diameter, UA 222 closely approxi-mates the estimates available for Javanese H. erectus (Grine andFranzen, 1994; Zanolli, 2013) but, with the exception of Homohabilis-rudolfensis from East Africa and Georgian H. erectus, thistooth is systematically smaller compared with the Early-Middle
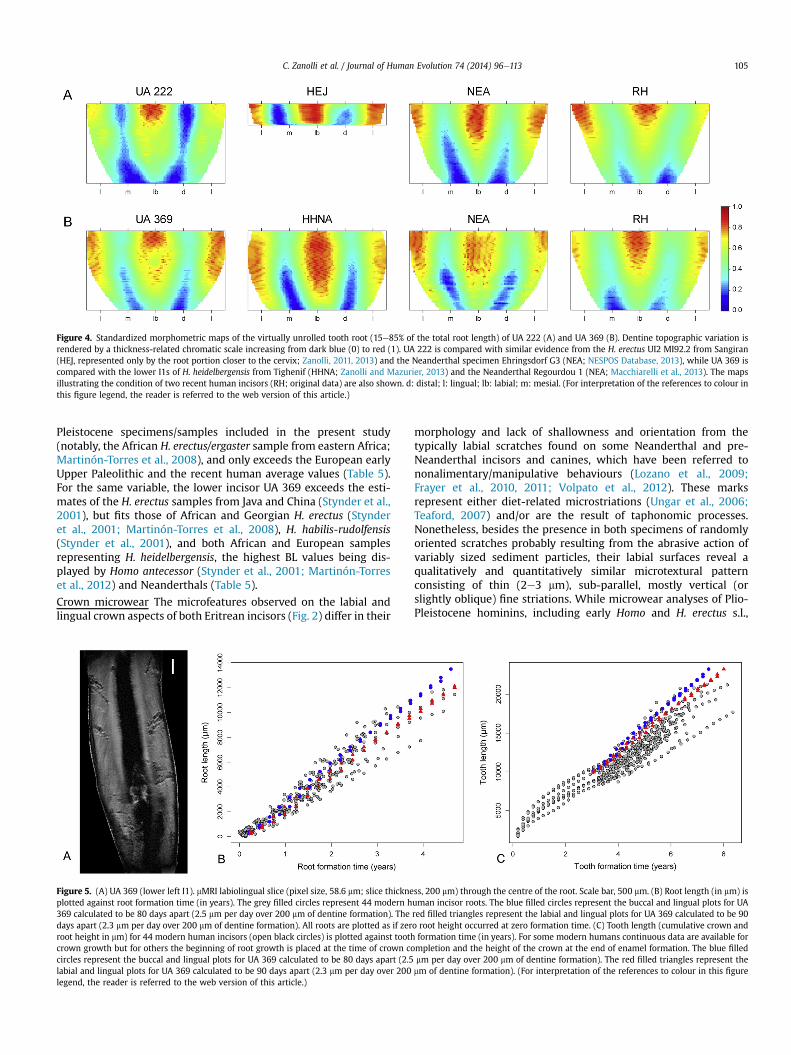
Figure 4. Standardized morphometric maps of the virtually unrolled tooth root (15e85% of the total root length) of UA 222 (A) and UA 369 (B). Dentine topographic variation isrendered by a thickness-related chromatic scale increasing from dark blue (0) to red (1). UA 222 is compared with similar evidence from the H. erectus UI2 MI92.2 from Sangiran(HEJ, represented only by the root portion closer to the cervix; Zanolli, 2011, 2013) and the Neanderthal specimen Ehringsdorf G3 (NEA; NESPOS Database, 2013), while UA 369 iscompared with the lower I1s of H. heidelbergensis from Tighenif (HHNA; Zanolli and Mazurier, 2013) and the Neanderthal Regourdou 1 (NEA; Macchiarelli et al., 2013). The mapsillustrating the condition of two recent human incisors (RH; original data) are also shown. d: distal; l: lingual; lb: labial; m: mesial. (For interpretation of the references to colour inthis figure legend, the reader is referred to the web version of this article.)
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113 105
Pleistocene specimens/samples included in the present study(notably, the African H. erectus/ergaster sample from eastern Africa;Martinón-Torres et al., 2008), and only exceeds the European earlyUpper Paleolithic and the recent human average values (Table 5).For the same variable, the lower incisor UA 369 exceeds the esti-mates of the H. erectus samples from Java and China (Stynder et al.,2001), but fits those of African and Georgian H. erectus (Stynderet al., 2001; Martinón-Torres et al., 2008), H. habilis-rudolfensis(Stynder et al., 2001), and both African and European samplesrepresenting H. heidelbergensis, the highest BL values being dis-played by Homo antecessor (Stynder et al., 2001; Martinón-Torreset al., 2012) and Neanderthals (Table 5).
Crown microwear The microfeatures observed on the labial andlingual crown aspects of both Eritrean incisors (Fig. 2) differ in their
Figure 5. (A) UA 369 (lower left I1). mMRI labiolingual slice (pixel size, 58.6 mm; slice thickneplotted against root formation time (in years). The grey filled circles represent 44 modern h369 calculated to be 80 days apart (2.5 mm per day over 200 mm of dentine formation). Thedays apart (2.3 mm per day over 200 mm of dentine formation). All roots are plotted as if zerroot height in mm) for 44 modern human incisors (open black circles) is plotted against toothcrown growth but for others the beginning of root growth is placed at the time of crown cocircles represent the buccal and lingual plots for UA 369 calculated to be 80 days apart (2.5labial and lingual plots for UA 369 calculated to be 90 days apart (2.3 mm per day over 200legend, the reader is referred to the web version of this article.)
morphology and lack of shallowness and orientation from thetypically labial scratches found on some Neanderthal and pre-Neanderthal incisors and canines, which have been referred tononalimentary/manipulative behaviours (Lozano et al., 2009;Frayer et al., 2010, 2011; Volpato et al., 2012). These marksrepresent either diet-related microstriations (Ungar et al., 2006;Teaford, 2007) and/or are the result of taphonomic processes.Nonetheless, besides the presence in both specimens of randomlyoriented scratches probably resulting from the abrasive action ofvariably sized sediment particles, their labial surfaces reveal aqualitatively and quantitatively similar microtextural patternconsisting of thin (2e3 mm), sub-parallel, mostly vertical (orslightly oblique) fine striations. While microwear analyses of Plio-Pleistocene hominins, including early Homo and H. erectus s.l.,
ss, 200 mm) through the centre of the root. Scale bar, 500 mm. (B) Root length (in mm) isuman incisor roots. The blue filled circles represent the buccal and lingual plots for UAred filled triangles represent the labial and lingual plots for UA 369 calculated to be 90o root height occurred at zero formation time. (C) Tooth length (cumulative crown andformation time (in years). For some modern humans continuous data are available formpletion and the height of the crown at the end of enamel formation. The blue filledmm per day over 200 mm of dentine formation). The red filled triangles represent themm of dentine formation). (For interpretation of the references to colour in this figure
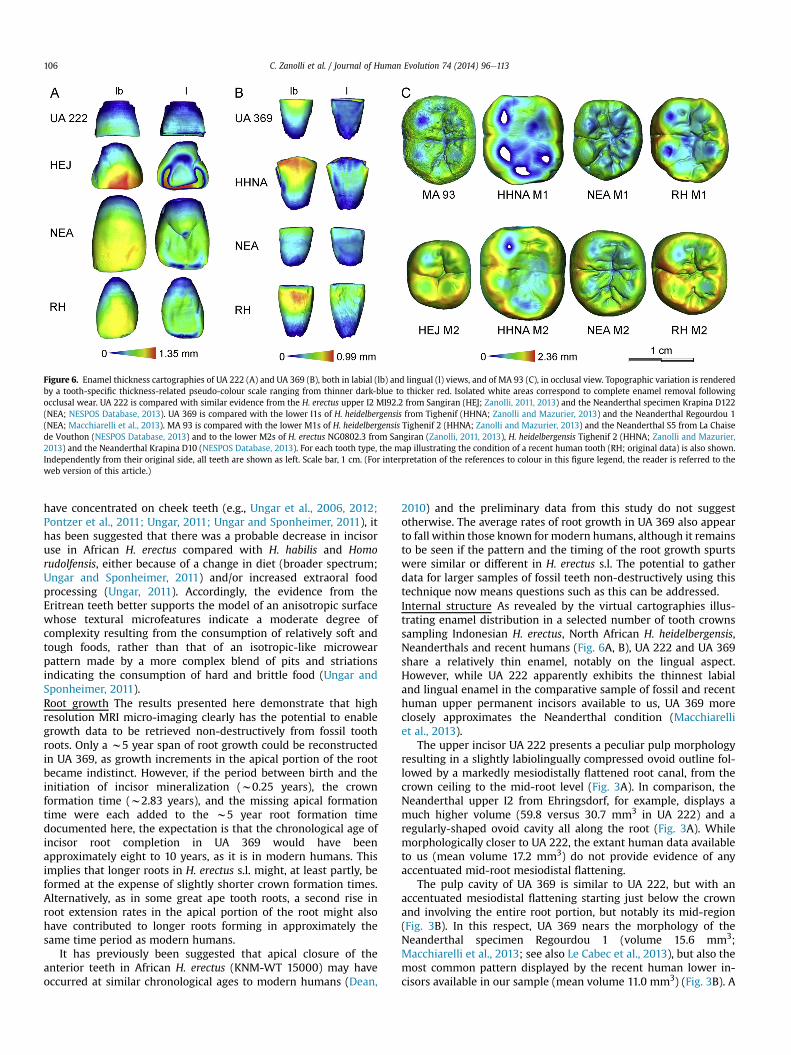
Figure 6. Enamel thickness cartographies of UA 222 (A) and UA 369 (B), both in labial (lb) and lingual (l) views, and of MA 93 (C), in occlusal view. Topographic variation is renderedby a tooth-specific thickness-related pseudo-colour scale ranging from thinner dark-blue to thicker red. Isolated white areas correspond to complete enamel removal followingocclusal wear. UA 222 is compared with similar evidence from the H. erectus upper I2 MI92.2 from Sangiran (HEJ; Zanolli, 2011, 2013) and the Neanderthal specimen Krapina D122(NEA; NESPOS Database, 2013). UA 369 is compared with the lower I1s of H. heidelbergensis from Tighenif (HHNA; Zanolli and Mazurier, 2013) and the Neanderthal Regourdou 1(NEA; Macchiarelli et al., 2013). MA 93 is compared with the lower M1s of H. heidelbergensis Tighenif 2 (HHNA; Zanolli and Mazurier, 2013) and the Neanderthal S5 from La Chaisede Vouthon (NESPOS Database, 2013) and to the lower M2s of H. erectus NG0802.3 from Sangiran (Zanolli, 2011, 2013), H. heidelbergensis Tighenif 2 (HHNA; Zanolli and Mazurier,2013) and the Neanderthal Krapina D10 (NESPOS Database, 2013). For each tooth type, the map illustrating the condition of a recent human tooth (RH; original data) is also shown.Independently from their original side, all teeth are shown as left. Scale bar, 1 cm. (For interpretation of the references to colour in this figure legend, the reader is referred to theweb version of this article.)
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113106
have concentrated on cheek teeth (e.g., Ungar et al., 2006, 2012;Pontzer et al., 2011; Ungar, 2011; Ungar and Sponheimer, 2011), ithas been suggested that there was a probable decrease in incisoruse in African H. erectus compared with H. habilis and Homorudolfensis, either because of a change in diet (broader spectrum;Ungar and Sponheimer, 2011) and/or increased extraoral foodprocessing (Ungar, 2011). Accordingly, the evidence from theEritrean teeth better supports the model of an anisotropic surfacewhose textural microfeatures indicate a moderate degree ofcomplexity resulting from the consumption of relatively soft andtough foods, rather than that of an isotropic-like microwearpattern made by a more complex blend of pits and striationsindicating the consumption of hard and brittle food (Ungar andSponheimer, 2011).Root growth The results presented here demonstrate that highresolution MRI micro-imaging clearly has the potential to enablegrowth data to be retrieved non-destructively from fossil toothroots. Only a w5 year span of root growth could be reconstructedin UA 369, as growth increments in the apical portion of the rootbecame indistinct. However, if the period between birth and theinitiation of incisor mineralization (w0.25 years), the crownformation time (w2.83 years), and the missing apical formationtime were each added to the w5 year root formation timedocumented here, the expectation is that the chronological age ofincisor root completion in UA 369 would have beenapproximately eight to 10 years, as it is in modern humans. Thisimplies that longer roots in H. erectus s.l. might, at least partly, beformed at the expense of slightly shorter crown formation times.Alternatively, as in some great ape tooth roots, a second rise inroot extension rates in the apical portion of the root might alsohave contributed to longer roots forming in approximately thesame time period as modern humans.
It has previously been suggested that apical closure of theanterior teeth in African H. erectus (KNM-WT 15000) may haveoccurred at similar chronological ages to modern humans (Dean,
2010) and the preliminary data from this study do not suggestotherwise. The average rates of root growth in UA 369 also appearto fall within those known for modern humans, although it remainsto be seen if the pattern and the timing of the root growth spurtswere similar or different in H. erectus s.l. The potential to gatherdata for larger samples of fossil teeth non-destructively using thistechnique now means questions such as this can be addressed.Internal structure As revealed by the virtual cartographies illus-trating enamel distribution in a selected number of tooth crownssampling Indonesian H. erectus, North African H. heidelbergensis,Neanderthals and recent humans (Fig. 6A, B), UA 222 and UA 369share a relatively thin enamel, notably on the lingual aspect.However, while UA 222 apparently exhibits the thinnest labialand lingual enamel in the comparative sample of fossil and recenthuman upper permanent incisors available to us, UA 369 moreclosely approximates the Neanderthal condition (Macchiarelliet al., 2013).
The upper incisor UA 222 presents a peculiar pulp morphologyresulting in a slightly labiolingually compressed ovoid outline fol-lowed by a markedly mesiodistally flattened root canal, from thecrown ceiling to the mid-root level (Fig. 3A). In comparison, theNeanderthal upper I2 from Ehringsdorf, for example, displays amuch higher volume (59.8 versus 30.7 mm3 in UA 222) and aregularly-shaped ovoid cavity all along the root (Fig. 3A). Whilemorphologically closer to UA 222, the extant human data availableto us (mean volume 17.2 mm3) do not provide evidence of anyaccentuated mid-root mesiodistal flattening.
The pulp cavity of UA 369 is similar to UA 222, but with anaccentuated mesiodistal flattening starting just below the crownand involving the entire root portion, but notably its mid-region(Fig. 3B). In this respect, UA 369 nears the morphology of theNeanderthal specimen Regourdou 1 (volume 15.6 mm3;Macchiarelli et al., 2013; see also Le Cabec et al., 2013), but also themost common pattern displayed by the recent human lower in-cisors available in our sample (mean volume 11.0 mm3) (Fig. 3B). A

C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113 107
mesiodistally flattened radicular canal compatible with the outlineof the Eritrean lower incisor also characterizes the lower I2 spec-imen UB 355 from ’Ubeidiya (Belmaker et al., 2002) and therecently reported Early Pleistocene lower I2 from Atapuerca Simadel Elefante, Spain (Prado-Simón et al., 2012). Conversely, the earlyMiddle Pleistocene isolated lower I1 from Tighenif (volume15.8 mm3) exhibits a spatula-like shape at crown level turning intoa regularly-shaped canal with an ovoid cross-section along theentire radicular portion (Zanolli and Mazurier, 2013) (Fig. 3B).
Topographic variation in root dentine thickness of both Eritreanincisors has been rendered through a morphometric map gener-ated by virtually unzipping and then unrolling 15e85% of the totalroot length (Fig. 4). Even if the significance (genetic versus func-tional) of the variation in root dentine thickness is not yet fullyunderstood (Bayle et al., 2012) and few specimens have beendetailed so far (Bayle, 2008; Bondioli et al., 2010; Bayle et al., 2011;Macchiarelli et al., 2013), we note some differences among thecartographies of the fossil and recent specimens considered in thisstudy. More specifically, while in UA 222 a unique thicker area isfound on the labial aspect near the CEJ, in the maps representingIndonesian H. erectus, Neanderthals and recent humans, verticallyextended thicker dentine is present both labially and lingually.Furthermore, at least in the Neanderthal and modern human roots,the regions where absolutely and relatively thinner dentine isfound aremuch less spread and confined to the lower portion of themap (Fig. 4A). Conversely, UA 369 root topography globally overlapsthe pattern of dentine thickness distribution expressed by theNorth African H. heidelbergensis and Neanderthal representativesused in this study, while the modern human cartography locallyshows relatively thinner dentine (Fig. 4B).
MA 93 (LLM1/M2)
External morphology The comparison between the occlusalmorphology of MA 93 and earlier East African lower molars sam-pling H. erectus/ergaster (e.g., KNM-ER 730, 820, 992, KNM-WT15000), as well as with the geographically closer specimens fromKonso (Suwa et al., 2007) and Melka Kunture (Condemi, 2007;Morgan et al., 2012) in Ethiopia, reveals a common generalpattern. However, while the presence of five main cuspsseparated by a Y-shaped groove is found almost ubiquitously inEarly Pleistocene African lower M1s (Wood, 1991; Bermúdez deCastro et al., 1999; Martinón-Torres et al., 2007, 2008), MA 93shows a cusp organization resulting in a þ fissure pattern morecommonly found in the Early Pleistocene lower M2s (Gómez-Robles et al., 2014). While not very common in the African record,the cruciform groove pattern is relatively frequent in IndonesianH. erectus (Zanolli, 2013) and in Middle Pleistocene Eurasianlower M2s and M3s (Bermúdez de Castro et al., 1999; Martinón-Torres et al., 2007, 2012; Bailey and Hublin, 2013).
Cusp size sequence in MA 93, with a larger protoconid than themetaconid and a medium-large hypoconulid, fits the averagepattern found in Early Pleistocene Afro-European lower M1s andM2s (e.g., KNM-ER 820 and 992 (Wood, 1991), ATD6-5 (Bermúdezde Castro et al., 1999)). In addition, MA 93 exhibits a well-developed and complete mid-trigonid crest, which even if foundin Georgian H. erectus and frequent in H. erectus s.s., is generallyabsent or interrupted by the central groove in African H. erectus/ergaster specimens like KNM-ER 820, 992 and KNM-WT 15000(Wood,1991; Brown andWalker,1993;Martinón-Torres et al., 2007,2008; Bailey and Hublin, 2013; Zanolli, 2013). Unlike most Afro-European H. erectus lower molars, like D211 (Martinón-Torreset al., 2008), KGA10-1 (Suwa et al., 2007), KNM-ER 820 and KNM-WT 15000 (Wood, 1991; Brown and Walker, 1993), the Eritreanmolar lacks any expression of a deflecting wrinkle, while the
presence of a C7, better expressed at the EDJ than at the outerenamel surface, is compatible with the high frequency of this traitobserved in H. erectus (Manni et al., 2007; Bailey and Hublin, 2013).Interestingly, in addition to its specific features, MA 93 closely re-sembles the LM1 KGA10-1 from Konso (Suwa et al., 2007), the twosharing a rectangular occlusal outline, the presence of a C7, the lackof protostylid (which is, conversely, quite a common feature inH. erectus s.l.; Manni et al., 2007), and low and nearly parallel lateralcrownwalls. To a lesser extent, the same is true for the lower M2 ofthe same specimen, even if the latter shows a slight buccolingualtapering of the talonid resulting into a more rounded occlusaloutline (Suwa et al., 2007).
Regardless of its serial position, MA 93 exhibits an absolutelymesiodistally extended crown (Table 5). If MA 93 is an M2, its MDdimension nears the averages of African H. erectus s.l. (Wood, 1991;Suwa et al., 2007) and of the Dmanisi sample (Martinón-Torreset al., 2008; Lordkipanidze et al., 2013), as well as the single valueavailable so far for H. antecessor (Bermúdez de Castro et al., 1999),but exceeds those of all remaining Early to Middle Pleistocenecomparative samples except H. habilis-rudolfensis, the latter againdisplaying the greatest anteroposterior crown diameter (Leakeyet al., 2012). Under this scenario, while the breadth of MA 93does not deviate from the estimates of most among the samplesconsidered in this analysis (notably, from African, Georgian, Chi-nese, and Javanese H. erectus; Wood, 1991; Suwa et al., 2007;Martinón-Torres et al., 2008; Lordkipanidze et al., 2013; Zanolli,2013), it is lower than measured in H. habilis-rudolfensis andNorth African H. heidelbergensis. The Eritrean molar also exceedsestimates from European H. heidelbergensis (Table 5).
Internal structure Comparative tooth crown tissue proportions inMA 93 and some fossil and recent human specimens/samples aregiven in Table 6 (2D estimates) and Table 7 (3D). The comparativerecord available for these kind of structural variables remainsquantitatively scanty in paleoanthropology and the extent ofintra-taxic/population variation is still virtually unknown, butregardless of whether assigned to M1 or M2, the Eritreanspecimen exhibits rather low 2D values, notably for the enamelarea (c). However, for dentine area (b) and enamel-dentinejunction length (e), the specimen fits the estimates from twoSouth African early Homo M1s (Smith et al., 2012), and is alsonear those from an isolated Javanese H. erectus M2 crown for thevariables c, e, and AET (Zanolli, 2011) (Table 6).
The number of comparative observations reported so far for 3Dinner tooth structural organization is even more limited, notablyfor the LM1s (Table 7), the most investigated fossil humans beingthe Neanderthals (Macchiarelli et al., 2006, 2007, 2008, 2013;Olejniczak et al., 2008a; Kupczik and Hublin, 2010). However,while for the enamel (Ve), dentine (Vcd), and pulp (Vcp) volumesthe Eritrean crown falls within the range of variation known forNeanderthals and recent humans, as an M2 it differs from the fig-ures expressed by the single representatives available to us ofIndonesian H. erectus (Zanolli, 2011) and H. heidelbergensis fromTighenif (Zanolli and Mazurier, 2013), as well as from the laterNorth African Aterians (Kupczik and Hublin, 2010).
A different picture is provided by the comparison for the percentratio Vcdp/Vc, which represents the coronal portion that is dentineand pulp, a directly comparable, size-independent parameterrather finely describing internal crown structural organization. Inthis case and specific comparative context, the structurally mostdistant signature with respect to MA 93 is represented by thevirtually unworn H. erectus lower M2 from Java used in this study(specimen NG0802.3; Zanolli, 2013). This specimen shows thelowest ratio of any Pleistocene specimen reported so far and con-trasts with the higher figures provided by our specimen sampling
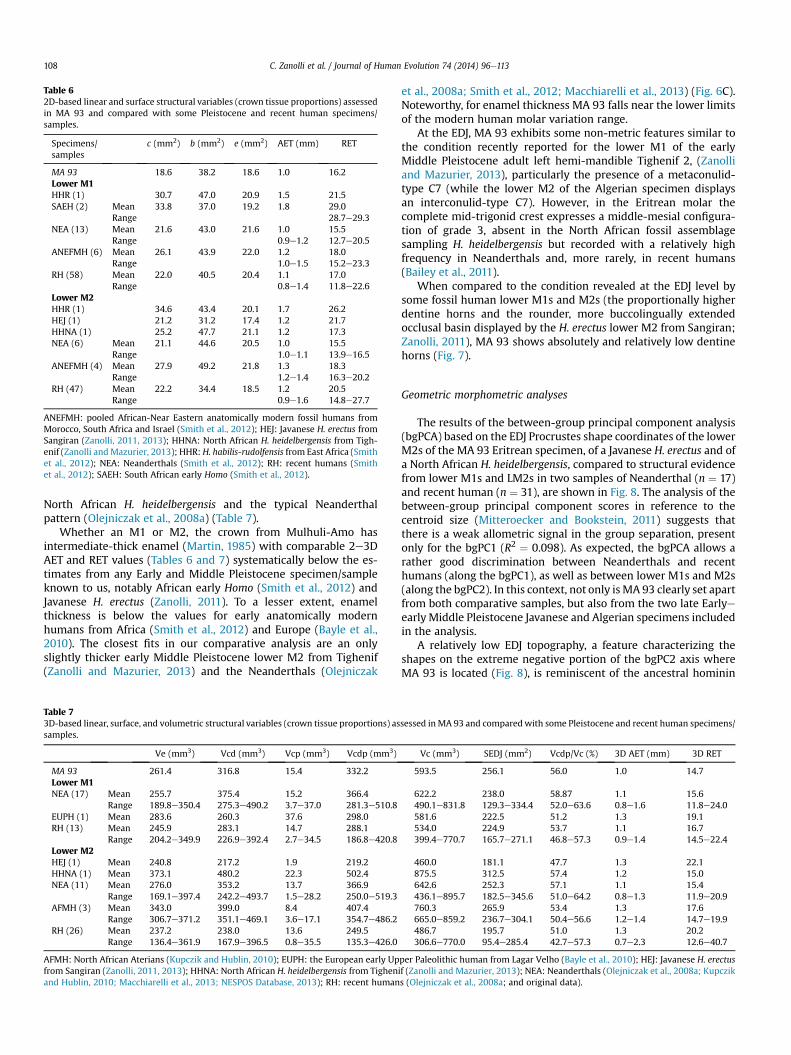
Table 62D-based linear and surface structural variables (crown tissue proportions) assessedin MA 93 and compared with some Pleistocene and recent human specimens/samples.
Specimens/samples
c (mm2) b (mm2) e (mm2) AET (mm) RET
MA 93 18.6 38.2 18.6 1.0 16.2Lower M1HHR (1) 30.7 47.0 20.9 1.5 21.5SAEH (2) Mean 33.8 37.0 19.2 1.8 29.0
Range 28.7e29.3NEA (13) Mean 21.6 43.0 21.6 1.0 15.5
Range 0.9e1.2 12.7e20.5ANEFMH (6) Mean 26.1 43.9 22.0 1.2 18.0
Range 1.0e1.5 15.2e23.3RH (58) Mean 22.0 40.5 20.4 1.1 17.0
Range 0.8e1.4 11.8e22.6Lower M2HHR (1) 34.6 43.4 20.1 1.7 26.2HEJ (1) 21.2 31.2 17.4 1.2 21.7HHNA (1) 25.2 47.7 21.1 1.2 17.3NEA (6) Mean 21.1 44.6 20.5 1.0 15.5
Range 1.0e1.1 13.9e16.5ANEFMH (4) Mean 27.9 49.2 21.8 1.3 18.3
Range 1.2e1.4 16.3e20.2RH (47) Mean 22.2 34.4 18.5 1.2 20.5
Range 0.9e1.6 14.8e27.7
ANEFMH: pooled African-Near Eastern anatomically modern fossil humans fromMorocco, South Africa and Israel (Smith et al., 2012); HEJ: Javanese H. erectus fromSangiran (Zanolli, 2011, 2013); HHNA: North African H. heidelbergensis from Tigh-enif (Zanolli andMazurier, 2013); HHR: H. habilis-rudolfensis from East Africa (Smithet al., 2012); NEA: Neanderthals (Smith et al., 2012); RH: recent humans (Smithet al., 2012); SAEH: South African early Homo (Smith et al., 2012).
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113108
North African H. heidelbergensis and the typical Neanderthalpattern (Olejniczak et al., 2008a) (Table 7).
Whether an M1 or M2, the crown from Mulhuli-Amo hasintermediate-thick enamel (Martin, 1985) with comparable 2e3DAET and RET values (Tables 6 and 7) systematically below the es-timates from any Early and Middle Pleistocene specimen/sampleknown to us, notably African early Homo (Smith et al., 2012) andJavanese H. erectus (Zanolli, 2011). To a lesser extent, enamelthickness is below the values for early anatomically modernhumans from Africa (Smith et al., 2012) and Europe (Bayle et al.,2010). The closest fits in our comparative analysis are an onlyslightly thicker early Middle Pleistocene lower M2 from Tighenif(Zanolli and Mazurier, 2013) and the Neanderthals (Olejniczak
Table 73D-based linear, surface, and volumetric structural variables (crown tissue proportions) assamples.
Ve (mm3) Vcd (mm3) Vcp (mm3) Vcdp (mm3)
MA 93 261.4 316.8 15.4 332.2Lower M1NEA (17) Mean 255.7 375.4 15.2 366.4
Range 189.8e350.4 275.3e490.2 3.7e37.0 281.3e510.8EUPH (1) Mean 283.6 260.3 37.6 298.0RH (13) Mean 245.9 283.1 14.7 288.1
Range 204.2e349.9 226.9e392.4 2.7e34.5 186.8e420.8Lower M2HEJ (1) Mean 240.8 217.2 1.9 219.2HHNA (1) Mean 373.1 480.2 22.3 502.4NEA (11) Mean 276.0 353.2 13.7 366.9
Range 169.1e397.4 242.2e493.7 1.5e28.2 250.0e519.3AFMH (3) Mean 343.0 399.0 8.4 407.4
Range 306.7e371.2 351.1e469.1 3.6e17.1 354.7e486.2RH (26) Mean 237.2 238.0 13.6 249.5
Range 136.4e361.9 167.9e396.5 0.8e35.5 135.3e426.0
AFMH: North African Aterians (Kupczik and Hublin, 2010); EUPH: the European early Upfrom Sangiran (Zanolli, 2011, 2013); HHNA: North African H. heidelbergensis from Tighenand Hublin, 2010; Macchiarelli et al., 2013; NESPOS Database, 2013); RH: recent human
et al., 2008a; Smith et al., 2012; Macchiarelli et al., 2013) (Fig. 6C).Noteworthy, for enamel thickness MA 93 falls near the lower limitsof the modern human molar variation range.
At the EDJ, MA 93 exhibits some non-metric features similar tothe condition recently reported for the lower M1 of the earlyMiddle Pleistocene adult left hemi-mandible Tighenif 2, (Zanolliand Mazurier, 2013), particularly the presence of a metaconulid-type C7 (while the lower M2 of the Algerian specimen displaysan interconulid-type C7). However, in the Eritrean molar thecomplete mid-trigonid crest expresses a middle-mesial configura-tion of grade 3, absent in the North African fossil assemblagesampling H. heidelbergensis but recorded with a relatively highfrequency in Neanderthals and, more rarely, in recent humans(Bailey et al., 2011).
When compared to the condition revealed at the EDJ level bysome fossil human lower M1s and M2s (the proportionally higherdentine horns and the rounder, more buccolingually extendedocclusal basin displayed by the H. erectus lower M2 from Sangiran;Zanolli, 2011), MA 93 shows absolutely and relatively low dentinehorns (Fig. 7).
Geometric morphometric analyses
The results of the between-group principal component analysis(bgPCA) based on the EDJ Procrustes shape coordinates of the lowerM2s of the MA 93 Eritrean specimen, of a Javanese H. erectus and ofa North African H. heidelbergensis, compared to structural evidencefrom lower M1s and LM2s in two samples of Neanderthal (n ¼ 17)and recent human (n ¼ 31), are shown in Fig. 8. The analysis of thebetween-group principal component scores in reference to thecentroid size (Mitteroecker and Bookstein, 2011) suggests thatthere is a weak allometric signal in the group separation, presentonly for the bgPC1 (R2 ¼ 0.098). As expected, the bgPCA allows arather good discrimination between Neanderthals and recenthumans (along the bgPC1), as well as between lower M1s and M2s(along the bgPC2). In this context, not only is MA 93 clearly set apartfrom both comparative samples, but also from the two late EarlyeearlyMiddle Pleistocene Javanese and Algerian specimens includedin the analysis.
A relatively low EDJ topography, a feature characterizing theshapes on the extreme negative portion of the bgPC2 axis whereMA 93 is located (Fig. 8), is reminiscent of the ancestral hominin
sessed in MA 93 and comparedwith some Pleistocene and recent human specimens/
Vc (mm3) SEDJ (mm2) Vcdp/Vc (%) 3D AET (mm) 3D RET
593.5 256.1 56.0 1.0 14.7
622.2 238.0 58.87 1.1 15.6490.1e831.8 129.3e334.4 52.0e63.6 0.8e1.6 11.8e24.0581.6 222.5 51.2 1.3 19.1534.0 224.9 53.7 1.1 16.7399.4e770.7 165.7e271.1 46.8e57.3 0.9e1.4 14.5e22.4
460.0 181.1 47.7 1.3 22.1875.5 312.5 57.4 1.2 15.0642.6 252.3 57.1 1.1 15.4436.1e895.7 182.5e345.6 51.0e64.2 0.8e1.3 11.9e20.9760.3 265.9 53.4 1.3 17.6665.0e859.2 236.7e304.1 50.4e56.6 1.2e1.4 14.7e19.9486.7 195.7 51.0 1.3 20.2306.6e770.0 95.4e285.4 42.7e57.3 0.7e2.3 12.6e40.7
per Paleolithic human from Lagar Velho (Bayle et al., 2010); HEJ: Javanese H. erectusif (Zanolli and Mazurier, 2013); NEA: Neanderthals (Olejniczak et al., 2008a; Kupcziks (Olejniczak et al., 2008a; and original data).
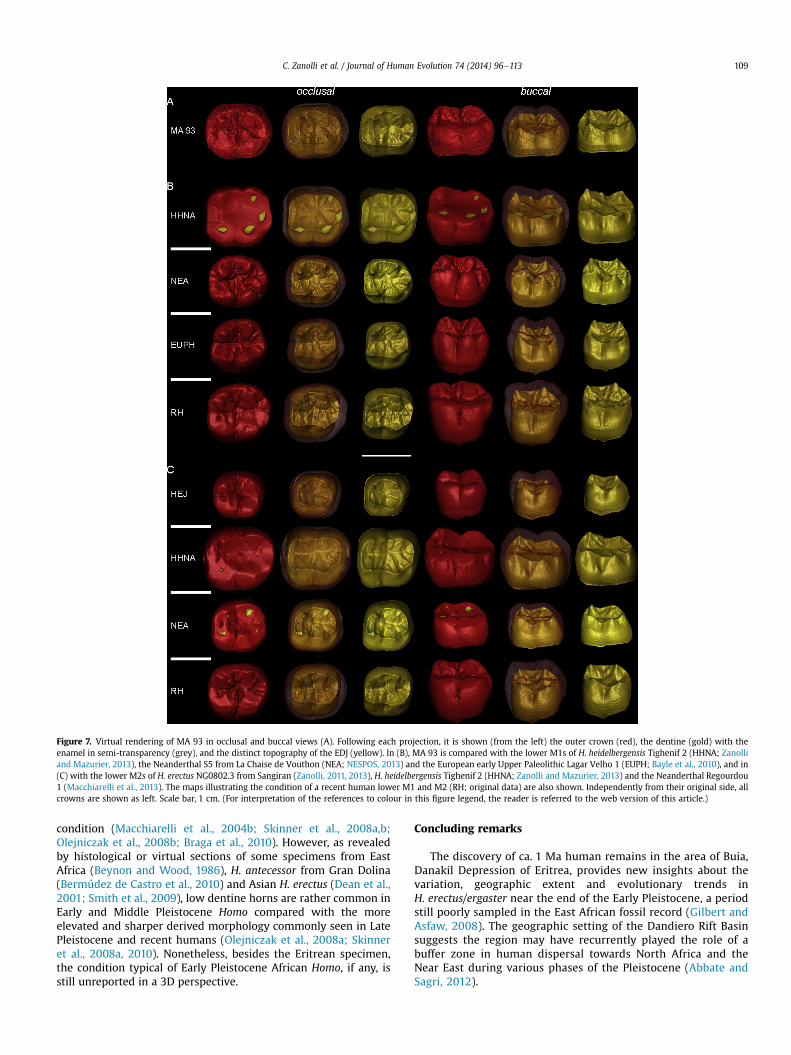
Figure 7. Virtual rendering of MA 93 in occlusal and buccal views (A). Following each projection, it is shown (from the left) the outer crown (red), the dentine (gold) with theenamel in semi-transparency (grey), and the distinct topography of the EDJ (yellow). In (B), MA 93 is compared with the lower M1s of H. heidelbergensis Tighenif 2 (HHNA; Zanolliand Mazurier, 2013), the Neanderthal S5 from La Chaise de Vouthon (NEA; NESPOS, 2013) and the European early Upper Paleolithic Lagar Velho 1 (EUPH; Bayle et al., 2010), and in(C) with the lower M2s of H. erectus NG0802.3 from Sangiran (Zanolli, 2011, 2013), H. heidelbergensis Tighenif 2 (HHNA; Zanolli and Mazurier, 2013) and the Neanderthal Regourdou1 (Macchiarelli et al., 2013). The maps illustrating the condition of a recent human lower M1 and M2 (RH; original data) are also shown. Independently from their original side, allcrowns are shown as left. Scale bar, 1 cm. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113 109
condition (Macchiarelli et al., 2004b; Skinner et al., 2008a,b;Olejniczak et al., 2008b; Braga et al., 2010). However, as revealedby histological or virtual sections of some specimens from EastAfrica (Beynon and Wood, 1986), H. antecessor from Gran Dolina(Bermúdez de Castro et al., 2010) and Asian H. erectus (Dean et al.,2001; Smith et al., 2009), low dentine horns are rather common inEarly and Middle Pleistocene Homo compared with the moreelevated and sharper derived morphology commonly seen in LatePleistocene and recent humans (Olejniczak et al., 2008a; Skinneret al., 2008a, 2010). Nonetheless, besides the Eritrean specimen,the condition typical of Early Pleistocene African Homo, if any, isstill unreported in a 3D perspective.
Concluding remarks
The discovery of ca. 1 Ma human remains in the area of Buia,Danakil Depression of Eritrea, provides new insights about thevariation, geographic extent and evolutionary trends inH. erectus/ergaster near the end of the Early Pleistocene, a periodstill poorly sampled in the East African fossil record (Gilbert andAsfaw, 2008). The geographic setting of the Dandiero Rift Basinsuggests the region may have recurrently played the role of abuffer zone in human dispersal towards North Africa and theNear East during various phases of the Pleistocene (Abbate andSagri, 2012).
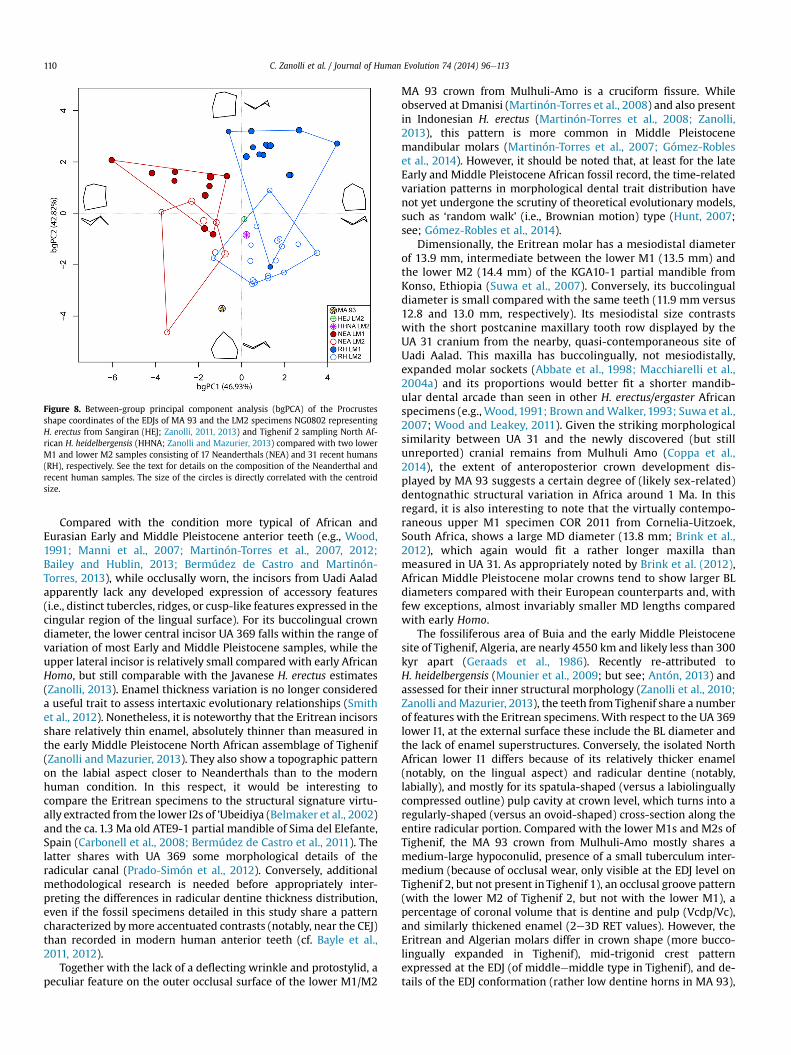
Figure 8. Between-group principal component analysis (bgPCA) of the Procrustesshape coordinates of the EDJs of MA 93 and the LM2 specimens NG0802 representingH. erectus from Sangiran (HEJ; Zanolli, 2011, 2013) and Tighenif 2 sampling North Af-rican H. heidelbergensis (HHNA; Zanolli and Mazurier, 2013) compared with two lowerM1 and lower M2 samples consisting of 17 Neanderthals (NEA) and 31 recent humans(RH), respectively. See the text for details on the composition of the Neanderthal andrecent human samples. The size of the circles is directly correlated with the centroidsize.
C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113110
Compared with the condition more typical of African andEurasian Early and Middle Pleistocene anterior teeth (e.g., Wood,1991; Manni et al., 2007; Martinón-Torres et al., 2007, 2012;Bailey and Hublin, 2013; Bermúdez de Castro and Martinón-Torres, 2013), while occlusally worn, the incisors from Uadi Aaladapparently lack any developed expression of accessory features(i.e., distinct tubercles, ridges, or cusp-like features expressed in thecingular region of the lingual surface). For its buccolingual crowndiameter, the lower central incisor UA 369 falls within the range ofvariation of most Early and Middle Pleistocene samples, while theupper lateral incisor is relatively small compared with early AfricanHomo, but still comparable with the Javanese H. erectus estimates(Zanolli, 2013). Enamel thickness variation is no longer considereda useful trait to assess intertaxic evolutionary relationships (Smithet al., 2012). Nonetheless, it is noteworthy that the Eritrean incisorsshare relatively thin enamel, absolutely thinner than measured inthe early Middle Pleistocene North African assemblage of Tighenif(Zanolli and Mazurier, 2013). They also show a topographic patternon the labial aspect closer to Neanderthals than to the modernhuman condition. In this respect, it would be interesting tocompare the Eritrean specimens to the structural signature virtu-ally extracted from the lower I2s of ’Ubeidiya (Belmaker et al., 2002)and the ca. 1.3 Ma old ATE9-1 partial mandible of Sima del Elefante,Spain (Carbonell et al., 2008; Bermúdez de Castro et al., 2011). Thelatter shares with UA 369 some morphological details of theradicular canal (Prado-Simón et al., 2012). Conversely, additionalmethodological research is needed before appropriately inter-preting the differences in radicular dentine thickness distribution,even if the fossil specimens detailed in this study share a patterncharacterized bymore accentuated contrasts (notably, near the CEJ)than recorded in modern human anterior teeth (cf. Bayle et al.,2011, 2012).
Together with the lack of a deflecting wrinkle and protostylid, apeculiar feature on the outer occlusal surface of the lower M1/M2
MA 93 crown from Mulhuli-Amo is a cruciform fissure. Whileobserved at Dmanisi (Martinón-Torres et al., 2008) and also presentin Indonesian H. erectus (Martinón-Torres et al., 2008; Zanolli,2013), this pattern is more common in Middle Pleistocenemandibular molars (Martinón-Torres et al., 2007; Gómez-Robleset al., 2014). However, it should be noted that, at least for the lateEarly and Middle Pleistocene African fossil record, the time-relatedvariation patterns in morphological dental trait distribution havenot yet undergone the scrutiny of theoretical evolutionary models,such as ‘random walk’ (i.e., Brownian motion) type (Hunt, 2007;see; Gómez-Robles et al., 2014).
Dimensionally, the Eritrean molar has a mesiodistal diameterof 13.9 mm, intermediate between the lower M1 (13.5 mm) andthe lower M2 (14.4 mm) of the KGA10-1 partial mandible fromKonso, Ethiopia (Suwa et al., 2007). Conversely, its buccolingualdiameter is small compared with the same teeth (11.9 mm versus12.8 and 13.0 mm, respectively). Its mesiodistal size contrastswith the short postcanine maxillary tooth row displayed by theUA 31 cranium from the nearby, quasi-contemporaneous site ofUadi Aalad. This maxilla has buccolingually, not mesiodistally,expanded molar sockets (Abbate et al., 1998; Macchiarelli et al.,2004a) and its proportions would better fit a shorter mandib-ular dental arcade than seen in other H. erectus/ergaster Africanspecimens (e.g., Wood,1991; Brown andWalker, 1993; Suwa et al.,2007; Wood and Leakey, 2011). Given the striking morphologicalsimilarity between UA 31 and the newly discovered (but stillunreported) cranial remains from Mulhuli Amo (Coppa et al.,2014), the extent of anteroposterior crown development dis-played by MA 93 suggests a certain degree of (likely sex-related)dentognathic structural variation in Africa around 1 Ma. In thisregard, it is also interesting to note that the virtually contempo-raneous upper M1 specimen COR 2011 from Cornelia-Uitzoek,South Africa, shows a large MD diameter (13.8 mm; Brink et al.,2012), which again would fit a rather longer maxilla thanmeasured in UA 31. As appropriately noted by Brink et al. (2012),African Middle Pleistocene molar crowns tend to show larger BLdiameters compared with their European counterparts and, withfew exceptions, almost invariably smaller MD lengths comparedwith early Homo.
The fossiliferous area of Buia and the early Middle Pleistocenesite of Tighenif, Algeria, are nearly 4550 km and likely less than 300kyr apart (Geraads et al., 1986). Recently re-attributed toH. heidelbergensis (Mounier et al., 2009; but see; Antón, 2013) andassessed for their inner structural morphology (Zanolli et al., 2010;Zanolli andMazurier, 2013), the teeth fromTighenif share a numberof features with the Eritrean specimens. With respect to the UA 369lower I1, at the external surface these include the BL diameter andthe lack of enamel superstructures. Conversely, the isolated NorthAfrican lower I1 differs because of its relatively thicker enamel(notably, on the lingual aspect) and radicular dentine (notably,labially), and mostly for its spatula-shaped (versus a labiolinguallycompressed outline) pulp cavity at crown level, which turns into aregularly-shaped (versus an ovoid-shaped) cross-section along theentire radicular portion. Compared with the lower M1s and M2s ofTighenif, the MA 93 crown from Mulhuli-Amo mostly shares amedium-large hypoconulid, presence of a small tuberculum inter-medium (because of occlusal wear, only visible at the EDJ level onTighenif 2, but not present in Tighenif 1), an occlusal groove pattern(with the lower M2 of Tighenif 2, but not with the lower M1), apercentage of coronal volume that is dentine and pulp (Vcdp/Vc),and similarly thickened enamel (2e3D RET values). However, theEritrean and Algerian molars differ in crown shape (more bucco-lingually expanded in Tighenif), mid-trigonid crest patternexpressed at the EDJ (of middleemiddle type in Tighenif), and de-tails of the EDJ conformation (rather low dentine horns in MA 93),

C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113 111
with Tighenif’s lower M2s more closely approaching the modernhuman condition (Zanolli and Mazurier, 2013).
As a whole, while quantitatively limited, comparative evidencefrom Buia/Mulhuli-Amo and Tighenif indicates a blend of primitive(mostly found in the earlier Danakil specimens), derived (mostlyfound in the later Algerian assemblage), and unique featurespointing to a significant degree of tooth variation at a macrore-gional scale and to a likely nonlinear pattern of dental morphos-tructural evolution across the Early to Middle Pleistocene. In thisrespect, while 2e3D virtual tooth imaging potentially disclosesnew promising perspectives in inter-taxic comparisons (e.g., Smithet al., 2012; Liu et al., 2013; Macchiarelli et al., 2013), we admit thatthe current paucity of similar evidence from more representativeEarly and Middle Pleistocene African assemblages still limits anyreliable conclusion about the variation patterns and evolutionarytrends in H. erectus/ergaster tooth endostructural morphology. Thisin turn reduces the chances of providing conclusive, dentally-basedsupport to the suggested speciation event occurring in Africa dur-ing the earlyMiddle Pleistocene (Rightmire, 2013). This emphasisesthe importance of continually developing methods, especiallythose that are non-desctructive, to extract more information out ofthe existing fossil record (Wood and Leakey, 2011).
Acknowledgements
The Buia Project (Eritrean-Italian Danakil Expedition: Anthropo-Archaeological and Geo-Paleontological Research Project), anongoing international project developed in agreement and collabo-rationwiththeEritreanNationalMuseumofAsmaraandtheNorthernRed Sea Regional Museum of Massawa, is granted by the ItalianMinistry for Education and Research (COFIN and PRIN national pro-jects), the Italian Ministry for Foreign Affairs (DGPCC-V; grants ARC-000387/2011, ARC-000504/2012, ARC-000853/2013), the University‘La Sapienza’ of Rome (‘Grandi Scavi Archeologici’ project; grantB88C13003900005 and B88C13000440005). During the last years,our field work has also benefited from the support provided by theItalian National Research Council, the University of Florence, theLeakey Foundation, the National Geographic Society, the SpanishMinistryof ScienceandEducation, theNationalMuseumofPrehistoryand Ethnography of Rome, the University of Poitiers, the French CNRS(MNHN Paris). For scientific collaboration, we acknowledge E. Abbate(Florence), M.G. Belcastro (Bologna), P. Bruni (Florence), G. Carnevale(Turin), M. Delfino (Turin), M. Ghinassi (Pavia), B. Martínez-Navarro(Tarragona), A. Mazurier (Poitiers), P. O’Higgins (York), O. Oms (Bar-celona),M. Papini (Florence),M. Pavia (Turin),M. Sagri (Florence).Weare also grateful to F. Bernardini (Trieste), M. Chech (Paris), F. Genchi(Bologna), A. Keset (Massawa), N. Kiflemariam (Massawa), F. Romag-noli (Florence), D. Tesfay (Massawa), A. Todero (Bologna), P. Ungar(Fayetteville), V. Vannata (Rome), and A. Zerai (Asmara). Very usefuland competent comments were provided by one referee when com-menting on the comparative evidence from Atapuerca. For detailedstratigraphic information about the Mulhuli-Amo site, we especiallythankM.Ghinassi andM.Papini. The specimenMA93wascollected in2011 by the Saho guide Hussein Umar. Finally, we recognize thatwithout the generous help and warm hospitality of the people fromthe Buia village, this work would simply have not been possible.
References
Abbate, E., Sagri, M., 2012. Early to Middle Pleistocene Homo dispersals from Africato Eurasia: Geological, climatic and environmental constraints. Quatern. Int.267, 3e19.
Abbate, E., Albianelli, A., Azzaroli, A., Benvenuti, M., Tesfamariam, B., Bruni, P.,Cipriani, N., Clarke, R.J., Ficcarelli, G., Macchiarelli, R., Napoleone, G., Papini, M.,Rook, L., Sagri, M., Tecle, T.M., Torre, D., Villa, I., 1998. A one-million-year-old
Homo cranium from the Danakil (Afar) Depression of Eritrea. Nature 393, 458e460.
Abbate, E., Beraky, W., Bruni, P., Falorni, P., Papini, M., Sagri, M., Simret, G.,Tewolde, M.T., 2004. Geology of the Homo-bearing Pleistocene Dandiero Basin(Buia Region, Eritrean Danakil Depression). Riv. Ital. Paleontol. Strat. 110, 5e34.
Albianelli, A., Napoleone, G., 2004. Magnetostratigraphy of the Homo-bearingPleistocene Dandiero Basin (Danakil Depression, Eritrea). Riv. Ital. Paleontol.Strat. 110, 35e44.
Allen, S.G., Stephenson, P.C.L., Strange, J.H., 1997. Morphology of porous mediastudied by nuclear magnetic resonance. J. Chem. Phys. 106, 7802e7811.
Antón, S.C., 2003. Natural history of Homo erectus. Yearb. Phys. Anthropol. 46, 126e170.
Antón, S.C., 2013. Homo erectus and related taxa. In: Begun, D.R. (Ed.), A Companionto Paleoanthropology. Blackwell Publishing Ltd., Pondicherry, pp. 497e516.
Baab, K.L., 2008. The taxonomic implications of cranial shape variation in Homoerectus. J. Hum. Evol. 54, 827e847.
Bailey, S.E., 2006. Beyond shovel-shaped incisors: Neandertal dental morphology ina comparative context. Period. Biol. 108, 253e267.
Bailey, S.E., Hublin, J.J., 2013. What does it mean to be dentally “modern”? In:Scott, G.R., Irish, J.D. (Eds.), Anthropological Perspectives on Tooth Morphology,Genetics, Evolution, Variation. Cambridge University Press, Cambridge,pp. 222e249.
Bailey, S.E., Skinner, M.M., Hublin, J.J., 2011. What lies beneath? An evaluation oflower molar trigonid crest patterns based on both dentine and enamelexpression. Am. J. Phys. Anthropol. 145, 505e518.
Bayle, P., 2008. Analyses quantitatives par imagerie à haute résolution des sé-quences de maturation dentaire et des proportions des tissus des dentsdéciduales chez les Néanderthaliens et les Hommes modernes. Ph.D. Disser-tation, Université Toulouse III Paul Sabatier.
Bayle, P., Macchiarelli, R., Trinkaus, E., Duarte, C., Mazurier, A., Zilhão, J., 2010. Dentalmaturational pattern and dental tissue proportions in the early Upper Paleolithicchild from Abrigo do Lagar Velho, Portugal. Proc. Natl. Acad. Sci. 107, 1338e1342.
Bayle, P., Bondioli, L., Macchiarelli, R., Mazurier, A., Puymerail, L., Volpato, V.,Zanolli, C., 2011. Three-dimensional imaging and quantitative characterizationof human fossil remains. Examples from the NESPOS database. In:Macchiarelli, R., Weniger, G.-C. (Eds.), Pleistocene Databases. Acquisition, Stor-ing, Sharing. Wissenschaftliche Schriften des Neanderthal Museums 4, Mett-mann, pp. 29e46.
Bayle, P., Bondioli, L., Dean, C., Macchiarelli, R., 2012. Unrolled roots: topographicvariation of the dentine thickness in Neandertal and modern human anteriorteeth. Proc. Eur. Soc. Study Hum. Evol. 1, 36 (abstract).
Belmaker, M., Tchernov, E., Condemi, S., Bar-Yosef, O., 2002. New evidence forhominid presence in the Lower Pleistocene of the Southern Levant. J. Hum. Evol.43, 43e56.
Bermúdez de Castro, J.M., Martinón-Torres, M., 2013. A newmodel for the evolutionof the human Pleistocene populations of Europe. Quatern. Int. 295, 102e112.
Bermúdez de Castro, J.M., Rosas, A., Nicolás, M.E., 1999. Dental remains fromAtapuerca-TD6 (Gran Dolina site, Burgos, Spain). J. Hum. Evol. 37, 523e566.
Bermúdez de Castro, J.M., Martinón-Torres, M., Prado, L., Gómez-Robles, A., Rosell, J.,López-Polín, L., Arsuaga, J.L., Carbonell, E., 2010. New immature hominin fossilfrom European Lower Pleistocene shows the earliest evidence of a modernhuman dental development pattern. Proc. Natl. Acad. Sci. 107, 11739e11744.
Bermúdez de Castro, J.M., Martinón-Torres, M., Gómez-Robles, A., Prado-Simón, L.,Martín-Francés, L., Lapresa, M., Olejniczak, A.J., Carbonell, E., 2011. Early Pleis-tocene human mandible from Sima del Elefante (TE) cave site in Sierra deAtapuerca (Spain): a comparative morphological study. J. Hum. Evol. 61, 12e25.
Beynon, A.D., Wood, B.A., 1986. Variations in enamel thickness and structure in EastAfrican hominids. Am. J. Phys. Anthropol. 70, 177e193.
Bigazzi, G., Balestrieri, M.L., Norelli, P., Oddone, M., 2004. Fission-track dating of atephra layer in the Alat formation of the Dandero Group (Danakil Depression,Eritrea). Riv. Ital. Paleontol. Strat. 110, 45e49.
Bondioli, L., Coppa, A., Frayer, D.W., Libsekal, Y., Rook, L., Macchiarelli, R., 2006.A one million year old human pubic symphysis. J. Hum. Evol. 50, 479e483.
Bondioli, L., Bayle, P., Dean, C., Mazurier, A., Puymerail, L., Ruff, C., Stock, J.T.,Volpato, V., Zanolli, C., Macchiarelli, R., 2010. Morphometric maps of long boneshafts and dental roots for imaging topographic thickness variation. Am. J. Phys.Anthropol. 142, 328e334.
Bondioli, L., Capuani, S., Coppa, A., Dean, C.M., Macchiarelli, R., Mancini, L.,Zanolli, C., 2013. Dentine growth patterns in human fossil teeth assessed byhigh resolution magnetic resonance micro-imaging. Proc. Europ. Soc. Stud.Hum. Evol. 2, 50 (abstract).
Braga, J., Thackeray, J.F., Subsol, G., Kahn, J.L., Maret, D., Treil, J., Beck, A., 2010. Theenamel-dentine junction in the postcanine dentition of Australopithecus afri-canus: intra-individual metameric and antimeric variation. J. Anat. 216, 62e79.
Brink, J.S., Herries, A.I.R., Moggi-Cecchi, J., Gowlett, J.A.J., Bousman, C.B., Hancox, J.P.,Grün, R., Eisenmann, V., Adams, J.W., Rossouw, L., 2012. First hominine remainsfrom a w1.0 million year old bone bed at Cornelia-Uitzoek, Free State Province,South Africa. J. Hum. Evol. 63, 527e535.
Brown, B., Walker, A., 1993. The dentition. In: Walker, A., Leakey, R.E. (Eds.), TheNariokotome Homo erectus Skeleton. Harvard University Press, Cambridge,pp. 161e192.
Callaghan, P.T., 1995. Principles of Nuclear Magnetic Resonance Microscopy. OxfordScience Publications, Oxford.
Carbonell, E., Bermúdez de Castro, J.M., Parés, J.M., Pérez-González, A., Ollé, A.,Mosquera, M., Cuenca-Bescós, G., García, N., Granger, D.E., Huguet, R., van der

C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113112
Made, J., Martinón-Torres, M., Rodríguez, X.P., Rosas, A., Sala, R., Stock, G.M.,Vallverdú, J., Vergés, J.M., Allué, E., Benito, A., Burjachs, F., Cáceres, I., Canals, A.,Díez, J.C., Lozano, M., Mateos, A., Navazo, M., Rodríguez, J., Rosell, J., Arsuaga, J.L.,2008. The first hominin of Europe. Nature 452, 465e469.
Coleman, M.N., Colbert, M.W., 2007. Technical note: CT thresholding protocols fortaking measurements on three-dimensional models. Am. J. Phys. Anthropol.133, 723e725.
Condemi, S., 2007. The Garba IV E mandible. In: Chavaillon, J., Piperno, M. (Eds.),Studies on the Early Paleolithic Site of Melka Kunture, Ethiopia. IIPP, Florence,pp. 687e701.
Coppa, A., Abbate, E., Bondioli, L., Bruni, P., Candilio, F., Delfino, M., Genchi, F.,Ghinassi, M., Libsekal, Y., Macchiarelli, R., Martínez-Navarro, B., Medin, T.,Papini, M., Pavia, M., Rook, L., Sagri, M., Zanolli, C., 2012. Mulhuli-Amo, a newlate Early Pleistocene paleoanthropological site in the northern DanakilDepression, Eritrea. Proc. Eur. Soc. Stud. Hum. Evol. 1, 58e59 (abstract).
Coppa, A., Bondioli, L., Candilio, F., Frayer, D.W., Libsekal, Y., Medin, T., Tesfay, D.,Zanolli, C., Macchiarelli, R., 2014. New 1 Ma old human cranial remains fromMulhuli-Amo, near Uadi Aalad, Danakil (Afar) depression of Eritrea. Am. J. Phys.Anthropol. 153 (S58), 97.
Dean, M.C., 1998. Comparative observations on the spacing of short-period (vonEbner’s) lines in dentine. Arch. Oral. Biol. 43, 1009e1021.
Dean, M.C., 1999. Hominoid tooth growth. Using incremental lines in dentine asmarkers of growth in modern human and fossil primate teeth. In: Hoppa, R.,FitzGerald, C. (Eds.), Human Growth in the Past: Studies from Bones and Teeth.Cambridge University Press, Cambridge, pp. 11e127.
Dean, M.C., 2009. Extension rates and growth in tooth height of modern human andfossil hominin canines and molars. In: Koppe, T., Meyer, G., Alt, K.W. (Eds.),Frontiers of Oral Biology: Interdisciplinary Dental Morphology, vol. 13. Karger,Basel, pp. 68e73.
Dean, M.C., 2010. Retrieving chronological age from dental remains of early fossilhominins to reconstruct human growth in the past. Phil. Trans. R. Soc. B 365,3397e3410.
Dean, M.C., 2012. Daily rates of dentine formation and root extension rates inParanthropus boisei, KNM-ER 1817, from Koobi Fora, Kenya. In: Reynolds, S.C.,Gallagher, A. (Eds.), African Genesis. Perspectives on Hominin Evolution. Cam-bridge University Press, Cambridge, pp. 268e279.
Dean, M.C., Cole, T.J., 2013. Human life history evolution explains dissociation be-tween the timing of tooth eruption and peak rates of root growth. PLoS One 8,e54534.
Dean, M.C., Leakey, M.G., Reid, D., Schrenk, F., Schwartz, G.T., Stringer, C., Walker, A.,2001. Growth processes in teeth distinguish modern humans from Homoerectus and earlier hominins. Nature 414, 628e631.
Delfino, M., Segid, A., Yosief, D., Shoshani, J., Rook, L., Libsekal, Y., 2004. Fossilreptiles from the Pleistocene Homo-bearing locality of Buia (Eritrea, NorthernDanakil Depression). Riv. Ital. Paleontol. Strat. 110, 51e60.
De Santis, S., Rebuzzi, M., Di Pietro, G., Fasano, F., Maraviglia, B., Capuani, S., 2010.In vitro and in vivo MR evaluation of internal gradient to assess trabecular bonedensity. Phys. Med. Biol. 55, 5767e5785.
Fajardo, R.J., Ryan, T.M., Kappelman, J., 2002. Assessing the accuracy of high reso-lution X-ray computed tomography of primate trabecular bone by comparisonswith histological sections. Am. J. Phys. Anthropol. 118, 1e10.
Ferretti, M.P., Ficcarelli, G., Libsekal, Y., Tecle, T.M., Rook, L., 2003. Fossil ele-phants from Buia (northern Afar Depression, Eritrea) with remarks on thesystematics of Elephas recki (Proboscidea, Elephantidae). J. Vert. Paleontol.23, 244e257.
Fiore, I., Bondioli, L., Coppa, A., Macchiarelli, R., Russom, R., Kashay, H.,Solomon, T., Rook, L., Libsekal, Y., 2004. Taphonomic analysis of the late EarlyPleistocene bone remains from Buia (Dandero Basin, Danakil Depression,Eritrea): evidence for large mammal and reptile butchering. Riv. Ital. Pale-ontol. Strat. 110, 89e97.
Frayer, D.W., 1977. Metric dental change in the European Upper Paleolithic andMesolithic. Am. J. Phys. Anthropol. 46, 109e120.
Frayer, D.W., Lalueza-Fox, C., Fiore, I., Radov�ci�c, J., Bondioli, L., 2010. Right handedNeandertals: Vindija and beyond. J. Anthropol. Sci. 88, 113e127.
Frayer, D.W., Lozano, M., Bermúdez de Castro, J.M., Carbonell, E., Arsuaga, J.L.,Radov�ci�c, J., Fiore, I., Bondioli, L., 2011. More than 500,000 years of right-handedness in Europe. Laterality 17, 51e69.
Geraads, D., Hublin, J.J., Jaeger, J.J., 1986. The Pleistocene hominid site of TernifineAlgeria: new results on the environment, age, and human industries. Quatern.Res. 25, 380e386.
Ghinassi, M., Papini, M., Yosieph, L., Rook, L., 2009. Palaeoenvironments of the BuiaHomo site: high-resolution facies analysis and non-marine sequence stratig-raphy in the Alat formation (Pleistocene Dandiero Basin, Danakil depression,Eritrea). Palaeogeogr. Palaeoclimatol. Palaeoecol. 280, 415e431.
Gilbert, W.H., Asfaw, B., 2008. Homo erectus. Pleistocene Evidence from the MiddleAwash, Ethiopia. University of California Press, London.
Gómez-Robles, A., Bermúdez de Castro, J.M., Arsuaga, J.L., Carbonell, E., Polly, P.D.,2014. No known hominin species matches the expected dental morphology ofthe last common ancestor of Neanderthals and modern humans. Proc. Natl.Acad. Sci. http://dx.doi.org/10.1073/pnas.1302653110.
Grine, F.E., Franzen, J.L., 1994. Fossil hominid teeth from the Sangiran Dome (Java,Indonesia). Cour. Forsch. -Inst. Senckenberg 171, 75e103.
Haack, E.M., Brown, R.W., Thompson, M.R., Venkatesan, R., 1999. Magnetic Reso-nance Imaging: Physical Principles and Sequence Design. J. Wiley and Sons, Inc.,New York.
Hershkovitz, I., Smith, P., Sarig, R., Rolf, Q., Rodríguez, L., García, R., Arsuaga, J.,Barkai, R., Gopher, A., 2011. Middle Pleistocene dental remains from Qesem Cave(Israel). Am. J. Phys. Anthropol. 144, 575e592.
Hillson, S.W., Parfitt, S.A., Bello, S.M., Roberts, M.B., Stringer, C.B., 2010. Two homininincisor teeth from the middle Pleistocene site of Boxgrove, Sussex, England.J. Hum. Evol. 59, 493e503.
Hlusko, L.J., Carlson, J.P., Guatelli-Steinberg, D., Krueger, K.L., Mersey, B., Ungar, P.S.,Defleur, A., 2013. Neanderthal teeth from Moula-Guercy, Ardèche, France. Am. J.Phys. Anthropol. 151, 477e491.
Hunt, G., 2007. The relative importance of directional change, random walks, andstasis in the evolution of fossil lineages. Proc. Natl. Acad. Sci. 104, 18404e18408.
Indriati, E., 2004. Indonesian fossil hominid discoveries from 1889e2003: catalogueand problems. In: Akiyama, S., Miyawaki, R., Kubodera, T., Higuchi, M. (Eds.),Proceedings of the 5th and 6th symposia on collection building and naturalhistory studies in Asia and the Pacific Rim, Nat. Sci. Mus. Monographs, vol. 24,pp. 163e177.
Kaifu, Y., Aziz, F., Baba, H., 2005. Hominid mandibular remains from Sangiran:1952e1986 collection. Am. J. Phys. Anthropol. 128, 497e519.
Kono, R., 2004. Molar enamel thickness and distribution patterns in extant greatapes and humans: New insights based on a 3-dimensional whole crownperspective. Anthropol. Sci. 112, 121e146.
Krueger, K.L., Ungar, P.S., 2010. Incisor microwear textures of five bioarcheologicalgroups. Int. J. Osteoarchaeol. 20, 549e560.
Kupczik, K., Hublin, J.J., 2010. Mandibular molar root morphology in Neanderthalsand Late Pleistocene and recent Homo sapiens. J. Hum. Evol. 59, 525e541.
Leakey, M.G., Spoor, F., Dean, M.C., Feibel, C.S., Antón, S.C., Kiarie, C., Leakey, L.N.,2012. New fossils from Koobi Fora in northern Kenya confirm taxonomic di-versity in early Homo. Nature 488, 201e204.
Le Cabec, A., Gunz, P., Kupczik, K., Braga, J., Hublin, J.J., 2013. Anterior tooth rootmorphology and size in Neanderthals: taxonomic and functional implications.J. Hum. Evol. 64, 169e193.
Liu, W., Schepartz, L.A., Xing, S., Miller-Antonio, S., Wu, X., Trinkaus, E., Martinón-Torres, M., 2013. Late Middle Pleistocene hominin teeth from Panxian Dadong,South China. J. Hum. Evol. 64, 337e355.
Lordkipanidze, D., Ponce de León, M.S., Margvelashvili, A., Rak, Y., Rightmire, G.P.,Vekua, A., Zollikofer, C.P.E., 2013. A complete skull from Dmanisi, Georgia, andthe evolutionary biology of early Homo. Science 342, 326e331.
Lozano, M., Mosquera, M., Bermúdez de Castro, J.M., Arsuaga, J., Carbonell, E., 2009.Right handedness of Homo heidelbergensis from Sima de los Huesos Atapuerca,Spain 500,000 years ago. Evol. Hum. Behav. 30, 369e376.
Macchiarelli, R., Bondioli, L., Chech, M., Coppa, A., Fiore, I., Russom, R., Vecchi, F.,Libsekal, Y., Rook, L., 2004a. The late Early Pleistocene human remains fromBuia, Danakil Depression, Eritrea. Riv. Ital. Paleontol. Strat. 110, 133e144.
Macchiarelli, R., Bondioli, L., Falk, D., Faupl, P., Illerhaus, B., Kullmer, O., Richter, W.,Said, H., Sandrock, O., Schäfer, K., Urbanek, C., Viola, B.T., Weber, G.W.,Seidler, H., 2004b. Early Pliocene hominid tooth from Galili, Somali Region,Ethiopia. Coll. Antropol. 28, 65e76.
Macchiarelli, R., Bondioli, L., Debénath, A., Mazurier, A., Tournepiche, J.-F., Birch, W.,Dean, C., 2006. How Neanderthal molar teeth grew. Nature 444, 748e751.
Macchiarelli, R., Mazurier, A., Volpato, V., 2007. L’apport des nouvelles technologiesà l’étude des Néandertaliens. In: Vandermeersch, B., Maureille, B. (Eds.), LesNéandertaliens. Biologie et Cultures. C.T.H.S., Paris, pp. 169e179.
Macchiarelli, R., Bondioli, L., Mazurier, A., 2008. Virtual dentitions: touchingthe hidden evidence. In: Irish, J.D., Nelson, G.C. (Eds.), Technique andApplication in Dental Anthropology. Cambridge University Press, Cam-bridge, pp. 426e448.
Macchiarelli, R., Bayle, P., Bondioli, L., Mazurier, A., Zanolli, C., 2013. From outer toinner structural morphology in dental anthropology. The integration of thethird dimension in the visualization and quantitative analysis of fossil remains.In: Scott, R.G., Irish, J.D. (Eds.), Anthropological Perspectives on ToothMorphology: Genetics, Evolution, Variation. Cambridge University Press, Cam-bridge, pp. 250e277.
Mahoney, P., 2006. Dental microwear from Natufian hunter-gatherers and earlyNeolithic farmers: comparisons between and within samples. Am. J. Phys.Anthropol. 130, 308e319.
Manni, F., Vargiu, R., Coppa, A., 2007. Neural network analysis by using the Self-Organizing Maps (SOMs) applied to human fossil dental morphology: a newmethodology. In: Bailey, S.E., Hublin, J.-J. (Eds.), Dental Perspectives on HumanEvolution. State-of-the-Art Research in Dental Paleoanthropology. Springer,Dordrecht, pp. 81e101.
Martin, L.B., 1985. Significance of enamel thickness in hominoid evolution. Nature314, 260e263.
Martínez de Pinillos, M., Martinón-Torres, M., Skinner, M.M., Arsuaga, J.L., Gracia-Téllez, A., Martínez, I., Martín-Francés, L., Bermúdez de Castro, J.M., 2014.Trigonid crests expression in Atapuerca-Sima de los Huesos lower molars: in-ternal and external morphological expression and evolutionary inferences. C.R.Palevol 13, 205e221.
Martínez-Navarro, B., Rook, L., Segid, A., Yosieph, D., Ferretti, M.P., Shoshani, J.,Tecle, T.M., Libsekal, Y., 2004. The large fossil mammals from Buia (Eritrea):systematics, biochronology and paleoenvironments. Riv. Ital. Paleontol. Strat.110, 61e88.
Martínez-Navarro, B., Rook, L., Papini, M., Libsekal, Y., 2010. A new species of bullfrom the Early Pleistocene paleoanthropological site of Buia (Eritrea): paral-lelism on the dispersal of the genus Bos and the Acheulian culture. Quatern. Int.212, 169e175.

C. Zanolli et al. / Journal of Human Evolution 74 (2014) 96e113 113
Martini, F., Libsekal, Y., Filippi, O., Ghebréher, A., Kashai, H., Kiros, A., Martino, G.,Okubatsion, D., Segid, A., Solomon, T., Teka, Z., Yosief, D., Yamane, S., 2004.Characterization of lithic complexes from Buia (Dandiero basin, Danakildepression, Eritrea). Riv. Ital. Paleontol. Strat. 110, 99e132.
Martinón-Torres, M., Bermúdez de Castro, J.M., Gómez-Robles, A., Arsuaga, J.L.,Carbonell, E., Lordkipanidze, D., Manzi, G., Margvelashvili, A., 2007. Dental ev-idence on the hominin dispersals during the Pleistocene. Proc. Natl. Acad. Sci.104, 13279e13282.
Martinón-Torres, M., Bermúdez de Castro, J.M., Gómez-Robles, A., Margvelshvill, A.,Prado, L., Lordkipanidze, D., Vekua, A., 2008. Dental remains from Dmanisi(Republic of Georgia): morphological analysis and comparative study. J. Hum.Evol. 55, 249e273.
Martinón-Torres, M., Bermúdez de Castro, J.M., Gómez-Robles, A., Prado-Simón, L.,Arsuaga, J.L., 2012. Morphological description and comparison of the dentalremains from Atapuerca-Sima de los Huesos site (Spain). J. Hum. Evol. 62, 7e58.
Martinón-Torres, M., Martínez de Pinillos, M., Skinner, M.M., Martín-Francés, L.,Gracia-Téllez, A., Martínez, I., Arsuaga, J.L., Bermúdez de Castro, J.M., 2014.Talonid crests expression at the enamel-dentine junction of hominin lowerpermanent and deciduous molars. C.R. Palevol 13, 223e234.
McDougall, I., Brown, F.H., Vasconcelos, P.M., Cohen, B.E., Thiede, D.S.,Buchanan, M.J., 2012. New single crystal 40Ar/39Ar ages improve time scale fordeposition of the Omo Group, Omo-Turkana Basin, East Africa. J. Geol. Soc.London 169, 213e226.
Mitteroecker, P., Bookstein, F.L., 2011. Linear discrimination, ordination, and thevisualization of selection gradients in modern morphometrics. Evol. Biol. 38,100e114.
Morgan, L.E., Renne, P.R., Kieffer, G., Piperno, M., Gallotti, R., Raynal, J.-P., 2012.A chronological framework for a long and persistent archaeological record:Melka Kunture, Ethiopia. J. Hum. Evol. 62, 104e115.
Mounier, A., Marchal, F., Condemi, S., 2009. Is Homo heidelbergensis a distinct spe-cies? New insight on the Mauer mandible. J. Hum. Evol. 56, 219e246.
NESPOS Database, 2013. Neanderthal Studies Professional Online Service. http://www.nespos.org.
Olejniczak, A.J., Smith, T.M., Feeney, R.N.M., Macchiarelli, R., Mazurier, A.,Bondioli, L., Rosas, A., Fortea, J., de la Rasilla, M., Garcia-Tabernero, A.,Radov�ci�c, J., Skinner, M.M., Toussaint, M., Hublin, J.-J., 2008a. Dental tissueproportions and enamel thickness in Neandertal and modern human molars.J. Hum. Evol. 55, 12e23.
Olejniczak, A.J., Smith, T.M., Skinner, M.M., Grine, F.E., Feeney, R.N.M., Thackeray, J.F.,Hublin, J.-J., 2008b. Three-dimensional molar enamel distribution and thicknessin Australopithecus and Paranthropus. Biol. Lett. 4, 406e410.
Papini, M., Ghinassi, M., Libsekal, Y., Rook, L., 2014. Geological map of the faciesassociations of the northern Dandiero Basin (Danakil depression, Eritrea,including the Pleistocene Buya Homo site). J. Maps 10, 126e135.
Pontzer, H., Scott, J.R., Lordkipanidze, D., Ungar, P.S., 2011. Dental microwear textureanalysis and diet in the Dmanisi hominins. J. Hum. Evol. 61, 683e687.
Prado-Simón, L., Martinón-Torres, M., Baca, P., Gómez-Roblez, A., Lapresa, M.,Carbonell, E., Bermúdez de Castro, J.M., 2012. A morphological study of thetooth roots of the Sima del Elefante mandible (Atapuerca, Spain): a new clas-sification of the teeth. Biological and methodological considerations. Anthropol.Sci. 120, 61e72.
R Development Core Team, 2013. R: a language and environment for statisticalcomputing. http://www.R-project.org.
Rightmire, G.P., 2013. Homo erectus and Middle Pleistocene hominins: brain size,skull form, and species recognition. J. Hum. Evol. 65, 223e252.
Rook, L., Ghinassi, M., Libsekal, Y., Martínez-Navarro, B., Medin, T., Papini, M., 2010.Stratigraphic context and taxonomic assessment of the large cercopithecoid(Primates, Mammalia) from the late Early Pleistocene palaeoanthropologicalsite of Buia (Eritrea). J. Hum. Evol. 59, 692e697.
Rook, L., Ghinassi, M., Carnevale, G., Delfino, M., Pavia, M., Bondioli, L., Candilio, F.,Coppa, A., Martínez-Navarro, B., Medin, T., Papini, M., Zanolli, C., Libsekal, Y.,2012. Stratigraphic context and paleoenvironmental significance of minor taxa(Pisces, Reptilia, Aves, Rodentia) from the late Early Pleistocene palae-oanthropological site of Buia (Eritrea). J. Hum. Evol. 64, 83e92.
Schlager, S., 2013. Morpho: calculations and visualizations related to geometricmorphometrics. R package version 0.24.1. http://sourceforge.net/projects/morpho-rpackage/?source¼directory.
Schneider, C.A., Rasband, W.S., Eliceiri, K.W., 2012. NIH Image to ImageJ: 25 years ofimage analysis. Nat. Methods 9, 671e675.
Schwartz, J.H., Tattersall, I., 2003. The Human Fossil Record. In: CraniodentalMorphology of Genus Homo (Africa and Asia), vol. 2. Wiley-Liss, Hoboken.
Scott, G.R., Turner II, C.G., 1997. The Anthropology of Modern Human Teeth. DentalMorphology and its Variation in Recent Human Populations. Cambridge Uni-versity Press, Cambridge.
Skinner, M.M., Wood, B.A., Boesch, C., Olejniczak, A.J., Rosas, A., Smith, T.M.,Hublin, J.-J., 2008a. Dental trait expression at the enamel-dentine junction oflower molars in extant and fossil hominoids. J. Hum. Evol. 54, 173e186.
Skinner, M.M., Gunz, P., Wood, B.A., Hublin, J.-J., 2008b. Enamel-dentine junction(EDJ) morphology distinguishes the lower molars of Australopithecus africanusand Paranthropus robustus. J. Hum. Evol. 55, 979e988.
Skinner, M.M., Evans, A., Smith, T.M., Jernvall, J., Tafforeau, P., Kupczik, K.,Olejniczak, A.J., Rosas, A., Radov�ci�c, J., Thackeray, J.F., Toussaint, M.,Hublin, J.-J., 2010. Contributions of enamel-dentine junction shape andenamel deposition to primate molar crown complexity. Am. J. Phys.Anthropol. 142, 157e163.
Smith, H.B., 1984. Patterns of molar wear in hunter-gatherers and agriculturalists.Am. J. Phys. Anthropol. 63, 39e56.
Smith, T.M., Reid, D.J., Dean, M.C., Olejniczak, A.J., Martin, L.B., 2007. Molar devel-opment in common chimpanzees (Pan troglodytes). J. Hum. Evol. 52, 201e216.
Smith, T.M., Olejniczak, A.J., Kupczik, K., Lazzari, V., de Vos, J., Kullmer, O.,Schrenk, F., Hublin, J.-J., Jacob, T., Tafforeau, P., 2009. Taxonomic assessment ofthe Trinil molars using non-destructive 3D structural and development anal-ysis. PaleoAnthropol., 117e129.
Smith, T.M., Olejniczak, A.J., Zermeno, J.P., Tafforeau, P., Skinner, M.M., Hoffmann, A.,Radov�ci�c, J., Toussaint, M., Kruszynski, R., Menter, C., Moggi-Cecchi, J.,Glasmacher, U.A., Kullmer, O., Schrenk, F., Stringer, C., Hublin, J.-J., 2012. Vari-ation in enamel thickness within the genus Homo. J. Hum. Evol. 62, 395e411.
Spoor, F., Zonneveld, F., Macho, G.A., 1993. Linear measurements of cortical boneand dental enamel by computed tomography: applications and problems. Am. J.Phys. Anthropol. 91, 469e484.
Strange, J.H., Rahman, M., Smith, E.G., 1993. Characterization of porous solids byNMR. Phys. Rev. Lett. 71, 3589e3591.
Stynder, D.D., Moggi-Cecchi, J., Berger, L.R., Parkington, J.E., 2001. Human mandib-ular incisors from the late Middle Pleistocene locality of Hoedjiespunt 1, SouthAfrica. J. Hum. Evol. 41, 369e383.
Suwa, G., Asfaw, B., Haile-Selassie, Y., White, T.D., Shigehiro Katoh, K.,WoldeGabriel, G., William, K.H., Nakaya, H., Beyene, Y., 2007. Early PleistoceneHomo erectus fossils fromKonso, Southern Ethiopia. Anthropol. Sci. 115,133e151.
Teaford, M.F., 2007. Dental microwear and paleoanthropology: cautions and pos-sibilities. In: Bailey, S.E., Hublin, J.-J. (Eds.), Dental Perspectives on HumanEvolution. Springer, Dordrecht, pp. 345e368.
Turner II, C.G., Nichol, C.R., Scott, G.R., 1991. Scoring procedures for key morpho-logical traits of the permanent dentition: the Arizona State University DentalAnthropology System. In: Kelley, M.A., Larsen, C.S. (Eds.), Advances in DentalAnthropology. Wiley-Liss, New York, pp. 13e31.
Ungar, P.S., 2011. Dental evidence for the diets of Plio-Pleistocene hominins. Yearb.Phys. Anthropol. 54, 47e62.
Ungar, P.S., Sponheimer, M., 2011. The diets of early hominins. Science 334, 190e193.Ungar, P.S., Grine, F.E., Teaford, M.F., El Zaatari, S.E., 2006. Dental microwear and
diets of African early Homo. J. Hum. Evol. 50, 78e95.Ungar, P.S., Scott, R.S., Scott, J.R., Teaford, M., 2008. Dental microwear analysis:
historical perspectives and new approaches. In: Irish, J.D., Nelson, G.C. (Eds.),Technique and Application in Dental Anthropology. Cambridge University Press,Cambridge, pp. 389e425.
Ungar, P.S., Krueger, K.L., Blumenschine, R.J., Njau, J., Scott, R.S., 2012. Dentalmicrowear texture analysis of hominins recovered by the Olduvai LandscapePaleoanthropology Project, 1995e2007. J. Hum. Evol. 63, 429e437.
Vandermeersch, B., 1981. Les Hommes Fossiles de Qafzeh (Israël). Cahiers de Palé-ontologie (Paléoanthropologie). CNRS, Paris.
Voisin, J.L., Condemi, S., Wolpoff, M.H., Frayer, D.W., 2012. A new online database(http://anthropologicaldata.free.fr) and a short reflection about the productiveuse of compiling Internet data. PaleoAnthropology, 241e244.
Volpato, V., Macchiarelli, R., Guatelli-Steinberg, D., Fiore, I., Bondioli, L., Frayer, D.W.,2012. Hand to mouth in a Neandertal: right handedness in Regourdou 1. PLoSOne 7 (1e7), e43949.
Wehrli, F.W., 2013. Magnetic resonance of calcified tissues. J. Magn. Reson 229, 35e48.
Weiger, M., Pruessmann, K.P., Bracher, A.-K., Köhler, S., Lehmann, V., Wolfram, U.,Hennel, F., Rasche, V., 2012. High-resolution ZTE imaging of human teeth. NMRBiomed 25, 1144e1151.
Widianto, H., 1993. Unité et diversité des hominidés fossiles de Java: Présentationde restes humains inédits. Ph.D. Dissertation, Muséum national d’Histoirenaturelle, Paris.
Wood, B.A., 1991. Koobi Fora Research Project. In: Hominid Cranial Remains fromKoobi Fora, vol. 4. Clarendon Press, Oxford.
Wood, B.A., Leakey, M., 2011. The Omo-Turkana basin fossil hominins and theircontribution to our understanding of human evolution in Africa. Evol.Anthropol. 20, 264e292.
Zanolli, C., 2011. L’organisation endostructurale de restes dentaires humains duPléistocène inférieur final-moyen initial d’Indonésie et d’Afrique, avec uneattention particulière à Homo erectus s.s. caractérisation comparative à hauterésolution et problématiques taxinomiques. Ph.D. Dissertation, Muséum Na-tional d’Histoire Naturelle, Paris.
Zanolli, C., 2013. Additional evidence for morpho-dimensional tooth crown varia-tion in a new Indonesian H. erectus sample from the Sangiran Dome (CentralJava). PLoS One 8 (1e15), e67233.
Zanolli, C., Mazurier, A., 2013. Endostructural characterization of theH. heidelbergensis dental remains from the early Middle Pleistocene site ofTighenif, Algeria. C.R. Palevol 12, 293e304.
Zanolli, C., Bayle, P., Macchiarelli, R., 2010. Tissue proportions and enamel thicknessdistribution in the early Middle Pleistocene human deciduous molars fromTighenif (Ternifine), Algeria. C.R. Palevol 9, 341e348.
Zanolli, C., Bondioli, L., Mancini, L., Mazurier, A., Widianto, H., Macchiarelli, R., 2012.Two human fossil deciduous molars from the Sangiran Dome (Java, Indonesia):outer and inner morphology. Am. J. Phys. Anthropol. 147, 472e481.
Zanolli, C., Bondioli, L., Candilio, F., Coppa, A., Dreossi, D., Frayer, D.W., Libsekal, Y.,Mancini, L., Rook, L., Medin, T., Tuniz, C., Macchiarelli, R., 2013. Endostructuralmorphology of the late Early Pleistocene human dental remains from UadiAalad and Mulhuli-Amo, Danakil (Afar) depression of Eritrea. Am. J. Phys.Anthropol. 150 (S56), 298 (abstract).




















![CYBERPSICOLOGIA [ CYBGIA ] - lumsa.it · - Il test UADI e altri strumenti psicodiagnostici. - La questione dell’aiuto psicologico online: aspetti tecno-strutturali ed etici delle](https://img.pdfslide.net/doc/110x75/5c69400109d3f263648cdfa4/cyberpsicologia-cybgia-lumsait-il-test-uadi-e-altri-strumenti-psicodiagnostici.jpg)








