Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Dec. 1974, p. 1361-1368Copyright 0 1974 American Society for Microbiology
Vol. 14, No. 6Printed in U.SA.
Kinetics of Inhibition of Influenza Virus Hemagglutination byHomologous Antihemagglutinin and AntineuraminidaseAntibodies as Measured by Means of the Photometric
Antibody Concentration Unit MethodJOACHIM DRESCHER, ULRICH DESSELBERGER, AND DIETRICH LUBACH
Institute of Virology, Medizinische Hochschule Hannover, D 3 Hannover-Kleefeld, Germany
Received for publication 11 September 1974
The kinetics of the reactions of antihemagglutinin (AH) and antineuramini-dase (AN) antibodies with homologous influenza virus were examined by use of aphotometric hemagglutination inhibition test (antibody concentration unit[ACU] test). The following results were obtained. (i) The isotherms describingthe reaction of AN antibodies with homologous virus were found to have muchsteeper slopes than did the corresponding AH antibody isotherm. This findingindicates that the photometric ACU method can discriminate objectivelybetween AH and AN antibodies. (ii) The reaction of mixtures of AH and ANantibodies with homologous virus was examined and found that AH antibodiescombine with virus independently from the presence of AN antibodies, whereasAH antibodies were found to reduce greatly the measurable degree of hemag-glutination inhibition by AN antibodies. (iii) A formula was developed andexperimentally confirmed by means of which the binding of virus by mixtures ofAH and AN antibodies can be predicted. Thereby, the influence of the relativeconcentration of AH and AN antibodies on the establishing of isotherms and ontheir subsequent use for antibody titration by means of the photometric ACUmethod was investigated. It was found that the procedure routinely employed forestablishing isotherms yielded data reflecting the reaction of AH antibodies only.As a consequence, the use of these isotherms will identify AH antibodies.
In a preceding publication (7), a highly accu-rate photometric method for the determinationof hemagglutination inhibiting influenza virusantibody has been described. This so calledphotometric ACU (antibody concentrationunit) method is based on the finding that, whenan excess of virus is reacted with graded dilu-tions of homologous antiserum, the ratio ofbound and free virus to the concentration ofantibody present can be described in terms ofFreundlich's adsorption isotherm (10).As applied, log CN/C.b = log A + 1/N log C',
where CN is the concentration of bound and C'of free virus, and Cab the concentration ofantibody present.CN is determined in operational terms as
difference between hemagglutinin concentra-tion units (HCUs) added (C) and HCUs recov-ered (C'). A and 1/N are two constants whichare experimentally determined for the strain ofvirus employed. Log A equals the intercept and1/N the slope of the straight line obtained whenplotting the values of log CN/Ca, against thevalues of log C'.
With heterologous reactants, the reactioncannot be described in terms of Freundlich'sisotherm, since a different type of proportional-ity pertains (2, 7). As a consequence, testingwhether or not the kinetics of the reaction of anunknown antiserum with a given strain of viruscan be described in terms of the isothermestablished for the strain of virus employedenables the reactions of virus with homologousand heterologous antibody to be objectivelydiscriminated (2, 7). This technique has beenutilized to test the specificity of antibody pat-terns recorded by conventional hemagglutina-tion inhibition (HI) tests in human sera (2, 3, 4).When these experiments were done, it was
not known' that under certain circumstancesantineuraminidase (AN) antibodies can causehemagglutination inhibition (11, 16). Therefore,it was of interest to test whether or not ANantibody can influence recognition of specificantihemagglutinin (AH) antibody by means ofthe photometric ACU method. For this purpose,the reaction between the virus strains A/Bel/42(HO N1), A/Sing/1/57 (H2 N2), and the recom-
1361
on July 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

DRESCHER, DESSELBERGER, AND LUBACH
binant A/Bel (HO)-A/Sing (N2) carrying thehemagglutinin of the strain A/Bel and theneuraminidase of the strain A/Sing/1/57 wasexamined. Since the strains A/Bel and A/Singwere found not to cross-react measurably, thereactions of A/Sing antibodies with A/Bel-A/Sing virus and of A/Bel-A/Sing antibodies withA/Sing virus represent AN antibody-mediatedhemagglutination inhibition.The experiments described in this paper were
designed to investigate the mutual influence ofAH and AN antibodies on the establishing ofadsorption isotherms and on their use for anti-body titration by means of the ACU method.For this purpose, the kinetics of reacting 'sepa-rately AH and AN antibodies with homologousvirus was investigated.
Since recognition of specific antibody bymeans of the ACU method is based on thekinetics of virus antibody interaction, the find-ing of different kinetics for AN and AH anti-body-virus interaction would indicate that bothtypes of antibodies can be discriminated bymeans of the ACU method.Furthermore, a formula was developed and
experimentally confirmed by means of whichthe binding of virus by mixtures of AH and ANantibodies can be predicted. By use of thisformula, the levels of AN antibodies influencingmeasurably the reaction of virus with AH anti-bodies can be determined.
In addition, it was felt that the knowledge ofthe kinetics of the reaction of AH and ANantibodies with homologous virus as obtained bythe experiments described in this paper couldcontribute to a better understanding of the roleof each type of antibody in immunity to influ-enza virus infection.
MATERIALS AND METHODSVirus. The strains of egg-adapted influenza virus
employed were: A/Sing/1/57 (H2 N2), A/Bel/42 (HON1), and the recombinant A/Bel/42 (HO)-A/Sing/I/57 (N2) carrying A/Bel/42 hemagglutinin and A/Sing/1/57 neuraminidase. (The virus suspensions used werefound to have the following ratios of neuraminidaseactivity [mU determined using fetuin purchased fromK and K Laboratories, Plainville, N.Y., as substrate]to hemagglutinin (HA) activity: A/Bel-0.021 mU/Ha,A/Bel-A/Sing-0.074 mU/HA, and A/Sing-0.012mU/HA.) In addition, the strains A/England/1/61(H2 N2), A/AA/1/65 (H2 N2), and A/Aichi/2/68 (H3N2) were used. Virus was purified by adsorption onand elution from BASO4 (8) and was suspended inbuffered saline (0.15 M NaCi), buffered at pH 7.0with 0.01 M phosphate.
Sera. Antisera were prepared by vaccination ofchickens and rabbits with the virus strains listedabove. The inoculation schedule was 10 intraperito-
neal injections given at 24-h intervals. Each contained100 HCUs of virus and was suspended in 20 ml ofbuffered saline (5). Blood samples were drawn prior tovaccination and 15, 30, 45, and 60 days thereafter.Sera were pretreated with M/90 KIO,. Prior to vacci-nation, none of the animals showed detectable anti-body against the virus strains employed.
Antibody titrations. (i) Hemagglutination inhibit-ing antibody were determined by means of the photo-metric ACU method (7). Titers were expressed interms of ACU units (7). (One ACU unit is defined asthat concentration of antibody which is just sufficientto give the value 1.0 for the ratio CN/AC'1/N whenreacted with homologous virus.) In addition, thereaction of sera with virus was characterized bydetermining the reciprocal highest serum dilution(d5*) yielding binding of 50% of 90 to 110 HCUs ofvirus by means of a photometric hemagglutinationinhibition test (5).
(ii) For establishing adsorption isotherms, the tech-nique used was that previously described (7). Eachisotherm was based on 80 to 130 measurements. The"standard serum" (7) was assigned a convenientnumerical value in order to have ACU titer valuescomputed by use of the isotherm in the same numeri-cal range of magnitude as HI titers determined byconventional HI tests. Hereby, the HI titers of A/Singantiserum against A/Bel-A/Sing virus and of A/Bel-A/Sing antiserum against A/Sing virus were usedwhen establishing the corresponding isotherms.The isotherms were tested for linearity as described
by Fazekas (9) as follows: for each isotherm, thevalues of 1/N and log A were calculated separately forvalues where C' (C' = concentration of free virus)ranged from 10 to 50 and for values where C' exceeded50. The values of 1/N and log A were tested forsignificant differences by means of a t test (P < 0.05).When the 1/N and log A values of these partialisotherms failed to differ significantly, the isothermwas considered linear.
(iii) HI pattern tests were carried out using fouragglutinating doses of virus. Titers were expressed interms of the reciprocal highest serum dilution yieldingcomplete hemagglutination inhibition.
(iv) Neuraminidase activity assays were carried outaccording to the method of Warren (15), using muco-protein from human urine prepared according toTamm and Horsfall (14) as substrate.
Sera were tested repeatedly for neuraminidaseinhibition as described by Seto and Rott (13), usingKIO4 pretreated sera.
Neuraminidase inhibition titers were expressed interms of the geometric means of the reciprocal highestserum dilutions yielding 50% inhibition, using neura-minidase starting concentrations yielding optical den-sity readings of 0.05 at a wave length of 549 nm in amodel DB Beckman spectrophotometer (light path of1 cm).
RESULTSPattern of reactions of virus strains A/Sing
(H2 N2), A/Bel (HO Ni), and the recombinantA/Bel (HO)-A/Sing (N2). The reactions of the
1362 J. VIROL.
on July 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

INHIBITION OF INFLUENZA VIRUS BY ACU METHOD
virus strains A/Bel, A/Sing, and the recombi-nant A/Bel-A/Sing with the corresponding anti-sera were examined by use of the photometrichemagglutination inhibition test (d,0), HI pat-tern test, and neuraminidase inhibition test.Representative examples of the results are givenin Table 1. Note that the A/Bel (HO)-A/Sing(N2) antiserum yielded AN antibody-mediatedhemagglutination inhibition of A/Sing virusand A/Sing antiserum of A/Bel (HO)-A/Sing(N2) virus. This conclusion is based on thefinding that the A/Bel antiserum failed to reactmeasurably with A/Sing virus and the A/Singantiserum with A/Bel virus.
Establishing of isotherms. The constants Aand 1/N of the isotherms describing the reac-
tion of the virus strains A/Sing, A/Bel, andA/Bel-A/Sing with the corresponding antiserawere determined where possible. Table 2 givesa survey on the adsorption isotherm constantsobtained. (The isotherms presented were foundto be linear.)The values of the constants A and 1/N were
found not to differ significantly (P < 0.5) for thereactions of A/Bel and A/Bel-A/Sing antiserawith A/Bel and A/Bel-A/Sing virus, indicatingthat these data were not measurably influencedby the presence-or absence of homologous ANantibodies.
Not shown for convenience in presentation isthat the ACU titers of A/Bel and of A/Bel-A/Sing antisera obtained by using A/Bel viruswere found not to differ significantly from theACU titers recorded when reacting these anti-sera with A/Bel-A/Sing virus. In contrast, theisotherms pertaining to AN antibody-mediatedhemagglutination inhibition (reaction of A/Singantisera with A/Bel-A/Sing virus and of A/Bel-A/Sing antisera with A/Sing virus) hadsignificantly (P < 0.05) higher values of theconstants 1/N and lower values of the constantA than the isotherms describing the reactions ofthese virus strains with AH antibodies (reactionof A/Bel antibody with A/Bel-A/Sing virus andof A/Sing antibody with A/Sing virus).This finding warrants the conclusion that AH
and AN antibodies reacted separately withhomologous virus will be identified correctly byuse of the ACU method.Reaction of mixtures of AH and AN anti-
bodies with homologous virus. To analyze thereaction of mixtures of AH and AN antibodieswith homologous virus, it was necessary toobtain formulas by means of which the virusconcentration bound by AH and AN antibodiesseparately can be precisely predicted.Binding of virus by AH antibodies. It was
found that when reacting C.b,H ACUs of AH
TABLE 1. Pattern of cross-reactions of strains A/Bel (HO Nl), A/Sing (H2 N2), and the recombinant A/Bel(HO)-A/Sing (N2)a
Antisera reacted with strain of virusAntisera obtained by
vaccination with A/Bel (HO N1) A/Bel (HO)-A/Sing (N2) A/Sing (H2 N2)strain of virus
dso HI NIT d5. HI NIT d5o HI NIT
A/Bel (HO N1) 3,936 1,120 90 3,950 1,120 < 10 <62 <28 < 10A/Bel (HO)-A/Sing 8,700 3,584 <10 13,987 3,584 57 700 560 1,527
A/Sing H2 N2) <62 < 28 < 10 3,800 112 245 15,090 j 8,960 2,740
a d0, Reciprocal highest serum dilution yielding binding of 50% of 90 to 110 HCUs; HI, reciprocal highestserum dilution yielding complete inhibition of four agglutinating doses of virus; NIT (neuraminidase inhibitiontest), reciprocal highest serum dilution inhibiting 50% of neuraminidase standard dose.
TABLE 2. Constants A and 1/N of the isotherms describing the reaction of strains A/Bel (HO Nl), A/Bel(HO)-A/Sing (N2), and A/Sing (H2 N2) with antibodies
Sera reacted with virus straina
Antisera oriented A/Bel (HO N1) A/Bel (HO)-A/Sing (N2) A/Sing (H2 N2)to strain of virus
A 1/N A 1/N A 1/N
A/Bel (HON 1) 151.2 0.3489 146.9 0.3403 - -
A/Bel (HO)-A/Sing (N2) 131.06 0.3512 135.7 0.3388 0.6567 1.413A/Sing (H2 N2) - - 2.268 1.0737 60.26 0.226
a -, No isotherm obtained, since sera failed to react with virus.
1363VOL. 14, 1974
on July 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

DRESCHER, DESSELBERGER, AND LUBACH
antibodies with C HCUs of homologous virusthe concentration of bound (CNNH) and freevirus (C 'H) can be precisely predicted by meansof equation 1, the derivation of which has beenpreviously (6) described:
CN,H = C - C'H (1)
where C'H equals ClyCab H, y equalsJOAHC(1/N,H - 1)/2.303 and AN and 1/N,H are theconstants of the isotherm describing the reac-tion of AH antibodies with homologous virus.Binding of virus by AN antibodies. It was
empirically found that equation 1 may not beapplied to data where 1/N exceeds 1.0. There-fore, it cannot be used for describing the reac-
tion of AN antibody with homologous virus.However, it was empirically found that thisreaction can be described in terms of equation2:
P = R log (TNQld) (2)
where P is the fraction of virus bound, TN is theconcentration ofAN antibodies in the undilutedserum in terms of ACU units, and d is thereciprocal serum dilution tested.R and Q are two constants which are experi-
mentally determined for the strain of virusemployed (Table 3). The determination of theseconstants and the derivation of equation 2 arepresented in the Appendix to this paper. Repre-sentative examples of the results obtained whentesting the validity of equation 2 are given inTable 4.An antiserum oriented to A/Sing virus was
reacted with A/Bel-A/Sing virus and the con-centration of bound virus observed was com-
pared with the binding of virus predicted by useof equation 2. Note that the values of theconcentration of bound virus predictedagreed well with the experimentally ob-served values.Binding of virus by mixtures ofAH and AN
antibodies. The binding of homologous virus bygraded dilutions of mixtures of AH antisera andAN antisera was photometrically determined,reacting mixtures of A/Sing and A/Bel antiserawith A/Bel-A/Sing virus and mixtures of A/Singand A/Bel-A/Sing antisera with A/Sing virus. Itwas found that the binding of virus by AH andAN antibody mixtures could be described bymeans of an equation derived by making thefollowing assumptions: it was assumed that AHantibody react independently from the presenceof AN antibody. Then, the concentration ofvirus bound (CNNH) when reacting a concentra-tion CG.,H of AH antibody with C HCUs ofhomologous virus is obtained by means ofequation 1.
It was further assumed that the binding ofvirus by AN antibodies equals the binding ofvirus expected when the concentration of virusfree after the reaction with AH antibodies (C'H)reacts with AN antibodies. Then, the virusstarting concentration operative against the ANantibodies equals C H = A/yCab,H and the con-
centration of virus bound by AN antibody(CN N) is obtained by use of equation 3:
CN.N = C R log (TN. Q/d)/yCab, H (3)
As a consequence, the total concentration ofbound virus (CN) equals the sum of the virusconcentrations bound by AH and by AN anti-bodies:
CN = C - C/yC ab. H+C.R log (TN. Q/d)/yCab. H (4)
TABLE 3. Constants R and Q of the formulaadescribing the reaction of antineuraminidase
antibody with homologous virus
Antiserum Reacted with R Qoriented to: virus strain
A/Sing (H2 N2) A/Bel-A/Sing 0.4713 29.644(HO H2)
A/Bel-A/Sing A/Sing (H2 N2) 0.576 12.12(HO N2)
aPercent bound virus = R log (TN. Q/d), where TNequals ACU titer of AN antibody and d is thereciprocal serum dilution tested.
TABLE 4. Examples of the reaction of AIBel-AlSing(HO N2) virus with A/Sing (H2 N2) antibodya
Reciprocalserum Cab.N C CNObs. CN.exp.dilution
560 1.786 161 131.3 130.81,120 0.893 161 115.6 108.02,240 0.446 161 96.9 85.14,480 0.223 161 58.4 62.28,960 0.112 161 40.9 39.61,120 0.893 76 53.3 51.02,240 0.446 76 44.3 40.24,480 0.223 76 29.7 29.48,960 0.112 76 17.2 18.72,240 0.446 38.7 22.1 20.44,480 0.223 38.7 13.8 15.08,960 0.112 38.7 10.4 9.52,240 0.446 23.7 12.7 12.54,480 0.223 23.7 8.5 9.2
a CaO,N, Concentration of AN antibody reacted; C,virus starting concentration; CN.Oft., concentration ofbound virus experimentally observed; CN.exp., concen-tration of bound virus calculated according to equa-tion 2.
1364 J. VIROL.
on July 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
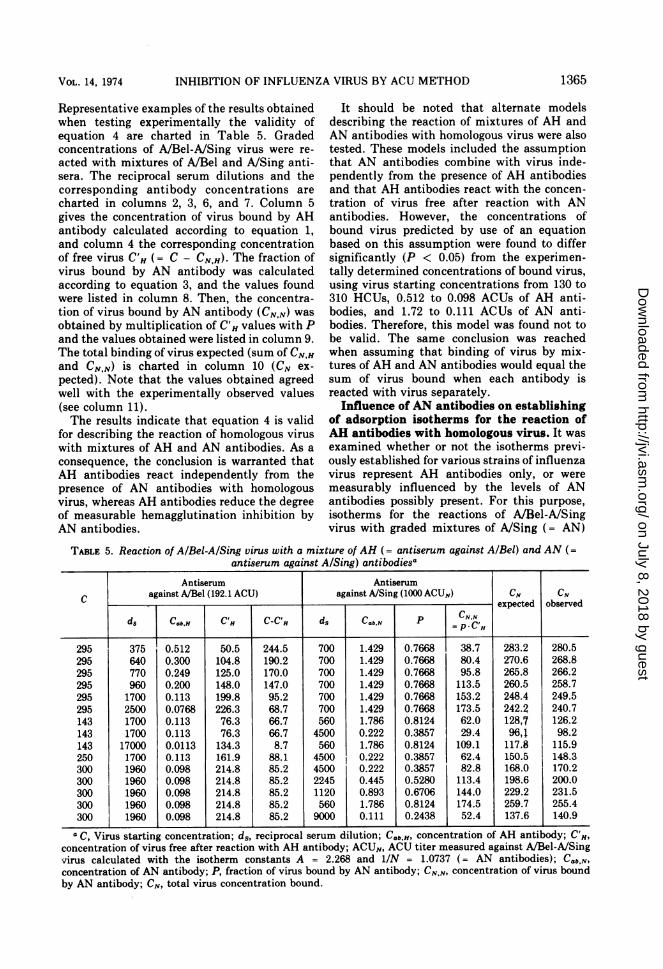
INHIBITION OF INFLUENZA VIRUS BY ACU METHOD
Representative examples of the results obtainedwhen testing experimentally the validity ofequation 4 are charted in Table 5. Gradedconcentrations of A/Bel-A/Sing virus were re-acted with mixtures of A/Bel and A/Sing anti-sera. The reciprocal serum dilutions and thecorresponding antibody concentrations arecharted in columns 2, 3, 6, and 7. Column 5gives the concentration of virus bound by AHantibody calculated according to equation 1,and column 4 the corresponding concentrationof free virus C'H (= C - CN.H). The fraction ofvirus bound by AN antibody was calculatedaccording to equation 3, and the values foundwere listed in column 8. Then, the concentra-tion of virus bound by AN antibody (CNN) wasobtained by multiplication of C'H values with Pand the values obtained were listed in column 9.The total binding of virus expected (sum of CN.Hand CN.N) is charted in column 10 (CN ex-pected). Note that the values obtained agreedwell with the experimentally observed values(see column 11).The results indicate that equation 4 is valid
for describing the reaction of homologous viruswith mixtures of AH and AN antibodies. As aconsequence, the conclusion is warranted thatAH antibodies react independently from thepresence of AN antibodies with homologousvirus, whereas AH antibodies reduce the degreeof measurable hemagglutination inhibition byAN antibodies.
It should be noted that alternate modelsdescribing the reaction of mixtures of AH andAN antibodies with homologous virus were alsotested. These models included the assumptionthat AN antibodies combine with virus inde-pendently from the presence of AH antibodiesand that AH antibodies react with the concen-tration of virus free after reaction with ANantibodies. However, the concentrations ofbound virus predicted by use of an equationbased on this assumption were found to differsignificantly (P < 0.05) from the experimen-tally determined concentrations of bound virus,using virus starting concentrations from 130 to310 HCUs, 0.512 to 0.098 ACUs of AH anti-bodies, and 1.72 to 0.111 ACUs of AN anti-bodies. Therefore, this model was found not tobe valid. The same conclusion was reachedwhen assuming that binding of virus by mix-tures of AH and AN antibodies would equal thesum of virus bound when each antibody isreacted with virus separately.
Influence of AN antibodies on establishingof adsorption isotherms for the reaction ofAH antibodies with homologous virus. It wasexamined whether or not the isotherms previ-ously established for various strains of influenzavirus represent AH antibodies only, or weremeasurably influenced by the levels of ANantibodies possibly present. For this purpose,isotherms for the reactions of A/Bel-A/Singvirus with graded mixtures of A/Sing (= AN)
TABLE 5. Reaction of AIBeI-AlSing virus with a mixture of AH (= antiserum against A/Bel) and AN (=antiserum against A/Sing) antibodiesa
Antiserum Antiserum
c against A/Bel (192.1 ACU) against A/Sing (1000 ACUN) CN CN. expected observed
ds Cab,H C'H C-C'H ds Cab.N P CN
lIC'CH
295 375 0.512 50.5 244.5 700 1.429 0.7668 38.7 283.2 280.5295 640 0.300 104.8 190.2 700 1.429 0.7668 80.4 270.6 268.8295 770 0.249 125.0 170.0 700 1.429 0.7668 95.8 265.8 266.2295 960 0.200 148.0 147.0 700 1.429 0.7668 113.5 260.5 258.7295 1700 0.113 199.8 95.2 700 1.429 0.7668 153.2 248.4 249.5295 2500 0.0768 226.3 68.7 700 1.429 0.7668 173.5 242.2 240.7143 1700 0.113 76.3 66.7 560 1.786 0.8124 62.0 128,7 126.2143 1700 0.113 76.3 66.7 4500 0.222 0.3857 29.4 96,4 98.2143 17000 0.0113 134.3 8.7 560 1.786 0.8124 109.1 117.8 115.9250 1700 0.113 161.9 88.1 4500 0.222 0.3857 62.4 150.5 148.3300 1960 0.098 214.8 85.2 4500 0.222 0.3857 82.8 168.0 170.2300 1960 0.098 214.8 85.2 2245 0.445 0.5280 113.4 198.6 200.0300 1960 0.098 214.8 85.2 1120 0.893 0.6706 144.0 229.2 231.5300 1960 0.098 214.8 85.2 560 1.786 0.8124 174.5 259.7 255.4300 1960 0.098 214.8 85.2 9000 0.111 0.2438 52.4 137.6 140.9n- X-r_ -,I _ _z_z _. _:_A. A. lA-; - ALU f"hA,. ?a u, Virus starting concentration; ds, reclprocal serum dilutuon; -ab,H, concentration 01 Ati antioOCy; L. H,
concentration of virus free after reaction with AH antibody; ACUN, ACU titer measured against A/Bel-A/Singvirus calculated with the isotherm constants A = 2.268 and 1/N = 1.0737 (= AN antibodies); Cb.tN,concentration of AN antibody; P, fraction of virus bound by AN antibody; CN,N, concentration of virus boundby AN antibody; CN, total virus concentration bound.
1365VOL. 14, 1974
on July 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

DRESCHER, DESSELBERGER, AND LUBACH
and A/Bel (= AH) antisera were established.The results obtained are given in Table 6. Theratio of AH (= A/Bel) to AN (= A/Sing)antibodies expressed in terms of d,0 values isgiven in column 2. Note that A/Bel antiserumwithout addition of A/Sing antiserum was usedin experiment no. 1. The number of dataobtained in each experiment is given in column3. Columns 4 and 6 list the isotherm constantsfound. By use of a t test (P < 0.05), it was
examined whether the 1/N and log A values ofexperiments 2 to 7 differed significantly fromthe corresponding value of experiment 1 (seecolumns 5 and 7). The standard errors of the1/N values were found not to exceed 0.005 andthose of the log A values not to exceed 0.025.The results obtained indicate that AN anti-bodies failed to influence measurably the ad-sorption isotherms, when the serum mixturescontained at least three times higher levels ofAH antibodies than AN antibodies in terms ofd,0 values (see experiments 2 to 5). In contrast,the presence of higher relative concentrations ofAN antibodies (see experiments 6 and 7) re-
sulted in a significant increase of 1/N and log Avalues.When the relative concentration of AN anti-
bodies was further increased (see experiments 8to 9), no isotherms were obtained since no linearrelationship between values of log CN/CGb andlog C' was recorded (see column 8). In theseexperiments, the partial isotherms calculatedfor data where C' ranged from 10 to 50 showed
significantly lower 1/N and higher log A valuesthan the partial isotherms calculated for datawhere C' exceeded 50. It should be noted thatthe influence of the relative concentration of AHand AN antibodies on the establishing of ad-sorption isotherms can be predicted by intro-ducing into equation 4 assumed values of TH,TN, and d, and by calculating isotherms usingthe values of free and bound virus obtained.The isotherms calculated by this procedure
for the reactions of A/Bel and A/Sing antibodymixtures with A/Bel-A/Sing virus agreed wellwith the isotherms experimentally determined.For the sake of brevity, no examples are given.In addition, antisera oriented to the virusstrains A/Sing/1/57 (H2 N2), A/England/1/61(H2 N2), A/AA/1/65 (H2 N2), and A/Aichi/2/68 (H3 N2), which had been previouslyused for establishing adsorption isotherms, were
tested in like manner with homologous virusand, in addition, with A/Bel-A/Sing virus.The ratios of AH to AN antibodies as mea-
sured in terms of d,0 values ranged from 9.7 to31.1. This finding suggests that the immuniza-tion procedure employed for preparing antiserafor establishing adsorption isotherms yields re-
latively low levels of AN antibodies and, as a
consequence, the isotherms established mea-
sure AH antibodies only.
DISCUSSIONWhen interpreting the results obtained, it
should be emphasized that all conclusions
TABLE 6. Determination of the constants A and 1/N of the isotherms describing the reactions ofA/Bel-A/Singvirus with graded mixtures of A/Bel and A/Sing antisera
Ratio of A/Bel Significance of Significance ofExpt to A/Sin iod/es N. o difference between difference betweenExpt to A/Silng antibodies NO. Of 1/N 1/N and 1/N value of Log A log A and log A value Linearitybno. in terms of d,5 values values expt no. 1 ofexpt.no.1
(dso,H/daJO.N)a(P < 0.05) (P < 0.05)
1 125 0.3403 2.16702 Yes2 11.9 87 0.3418 No 2.18752 No Yes3 9.1 115 0.3291 No 2.13987 No Yes4 6.0 76 0.3407 No 2.19865 No Yes5 3.0 98 0.3505 No 2.21484 No Yes6 2.0 112 0.4047 Yes 2.27085 Yes Yes7 0.6 87 0.5574 Yes 2.27875 Yes Yes8 0.24 104 No9 0.12 113 No
a do,,H, Reciprocal highest dilution of A/Bel serum yielding binding of 50% of 90 to 110 HCUs of A/Bel-A/Singvirus; dIO,N, reciprocal highest dilution of A/Sing serum yielding binding of 50% of 90 to 110 HCUs ofA/Bel-A/Sing virus.
b The linearity of each adsorption isotherm was tested by dividing the data into two groups, one comprisingvalues where the concentration of free virus ranged from 10 to 50 and a second where the concentration of freevirus ranged from 51 to 110. For each group, the values of log A and 1/N were calculated separately and testedfor significant differences by means of a t test (P < 0.05). When no significant difference between log A and 1/Nvalues was recorded, the isotherm was considered linear (9).
1366 J. VIROL.
on July 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

INHIBITION OF INFLUENZA VIRUS BY ACU METHOD
reached pertain to the strains of virus em-ployed, and that at present, no statement canbe made whether or not such conclusions arevalid for further virus strains or their recombi-nants.The data presented in this paper warrant the
conclusion that AH and AN antibodies havedrastically different kinetics of their reactionwith homologous virus as measured by means ofthe photometric ACU method. This findingindicates that the photometric ACU methodcan discriminate between AH and AN anti-bodies. When using an AH antibody isotherm,ACU titers will reflect AH antibodies, and viceversa.Using graded mixtures of AH and AN anti-
bodies, the influence of the relative concentra-tion of AH and AN antibodies on establishingadsorption isotherms was investigated, and itwas found that AN antibodies failed to influ-ence measurably the isotherm, if the ratio ofAHto AN antibodies exceeded 3.0. Since it wasfound that the vaccination schedule employedfor preparation of antisera used for establishingisotherms yields ratios of AH to AN antibodiesexceeding 3.0, this finding suggests that previ-ously established isotherms measure AH anti-body only. This conclusion is consistent with aprevious report (11), where AN antibody wasalso found not to influence measurement ofACU titers.A modification of the ACU method by means
of which both AH and AN antibodies can bedetermined will be described in a subsequentpublication. It might be questioned whether thefound difference between AH and AN antibodyisotherms is due to the fact that AN antibodieswere measured by means of a hemagglutinationinhibition test. Therefore, it should be men-tioned that when isotherms were established forthe reaction of AN antibodies with N2 neura-minidases of different strains by. use of enzymeinhibition test, an average value of the constant1/N of 1.09 was obtained (1), which is inexcellent agreement with the data presented inthis paper.The finding that AN antibody isotherms have
much steeper slopes than AH antibody iso-therms can be interpreted as follows: undercomparable conditions, the amount of virusbound per ACU unit of AN antibody willincrease much more rapidly with increasingvirus starting concentration than does the virusconcentration bound per ACU unit of AH anti-body. As an example, when reacting A/Bel-A/Sing virus with 0.1 ACUs of A/Sing antibody,an increase of the virus starting concentrationfrom 10 to 100 HCUs will result in 10-fold
increase of the virus concentration bound perACU unit, whereas with A/Bel antibody thisratio will increase 4.8-fold only. Vice versa, thebinding of virus by AN antibodies decreasesmuch more progressively with decreasing virusstarting concentration than does binding ofvirus by AH antibodies.The relatively low binding capacity of AN
antibody against low concentrations of viruscould be at least partially responsible for thefact that AN antibody do not prevent virusinfection where a low virus concentration isneeded but interferes with the release of virusfrom infected cells where a much higher virusconcentration is operative (16). When examin-ing the reaction of mixtures of AH and ANantibodies with homologous virus, it was foundthat binding of virus by AH antibodies was notinfluenced by the presence of AN antibodies. Incontrast, the presence of AH antibodies didgreatly reduce the measurable degree of hemag-glutination inhibition by AN antibodies, sincethe virus concentration bound by AN antibodieswas found to equal that virus concentrationbound if only the virus concentration free afterthe reaction with AH antibodies is reacted withAN antibodies. This finding could be explainedby the following hypothetical model: considerthat a virus concentration C is once reactedseparately with AH antibodies, yielding bindingof a- C HCUs, and once with AN antibodies,resulting in binding of b-C HCUs. Since AHand AN antibodies combine with different anti-genic sites, it is reasonable to assume that bothantibodies combine with virus independentlyfrom each other.Thus, when reacting a mixture of AH and AN
antibodies with C HCUs of virus, the concentra-tion of virus bound by AH antibodies will againequal a C, whereas the virus concentrationa *b * C is bound both by AH and AN antibodies.As a consequence, the binding of virus by ANantibodies observed equals b (C - a- C), whereC - aC is the virus concentration free afterreaction with AH antibodies. Thus, the de-crease in measurable binding of virus by ANantibodies in presence of AH antibodies is dueto the fact that virus inhibited by AH anti-bodies combines with AN antibodies withoutadditional inhibition of hemagglutinating activ-ity.
APPENDIXDetermination of the constants R and Q for
describing the reaction of AN antibodies withhomologous virus. Graded dilutions (lid) of ANantisera containing TN ACUs of AN antibodies werereacted with graded concentrations of homologous
1367VOL. 14, 1974
on July 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

DRESCHER, DESSELBERGER, AND LUBACH
virus, and the fraction of bound virus (P) was deter-mined under the conditions of the photometric ACUmethod.The slope (R) and the intercept (E) of the regres-
sion line relating values of P to values of log (TN/d)were determined by means of the method of leastsquares yielding:
P = R log (TN/d) + E (1)
This can be rearranged to give
10 = (TNI/dR) * 10E (2)
10P = (TN,.10E/1R/d)R (3)
Calling the value of 1OE/R Q gives:
P = R log (TN. Q/d) (4)
By means of equation 4, the fraction of virus boundwhen reacting a known concentration of AN anti-bodies (TN/d) with homologous virus can be calcu-lated. (Values of P larger than 1.0 are scored as 1.0.)
ACKNOWLEDGMENTSWe acknowledge with much appreciation the help of G. C.
Schild, World Influenza Centre, Mill-Hill, London, whogenerously supplied the virus strains A/Bel, A/Sing, andA/Bel-A/Sing. The mucoprotein samples employed were akind gift of E. Hertzberger, N. V. Philips Duphar, Weesp,The Netherlands. The excellent technical assistance of G.Andrae, B. Brunler, D. Friedrich, M. Stanschus, K. Stubben-dorff, and D. Weidauer is acknowledged.
LITERATURE CITED1. Baars, A. J., H. Frankena, and N. Masurel. 1971.
Antigenic relationship of influenza-virus neuramini-dase from Asian, Hong Kong and Equi-2 strains.Antonie van Leeuwenhoek. J. Microbiol. Serol. 37:209-218
2. Davenport, F. M., A. V. Hennessy, J. Drescher, J.Mulder, and T. Francis, Jr. 1964. Further observationson the relevance of serologic recapitulation of humaninfection with influenza viruses. J. Exp. Med.120:1087-1097.
3. Davenport, F. M., A. V. Hennessy, and E. Minuse. 1967.Further observations on the significance of A/
equine-2/63 antibodies in man. J. Exp. Med.126:1049-1061.
4. Davenport, F. M., A. V. Hennessy, and E. Minuse. 1968.The age distribution in humans of hemagglutinating-inhibiting antibodies reacting with avian strains ofinfluenza A virus. J. Immunol. 100:581-585.
5. Drescher, J. 1972. Patterns of cross-reaction of selectedinfluenza virus A2 strains isolated from 1957 to 1968 asdetermined by use of a photometric hemagglutinationinhibition test. Amer. J. Epidemiol. 95:549-556.
6. Drescher, J. 1973. Photometric method for determinationof doubly specific influenza antibody. Zentralbl. Bak-teriol. Hyg. Abt. I. Orig. 225:233-244.
7. Drescher, J., F. M. Davenport, and A. V. Hennessy.1962. Photometric methods for the measurement ofhemagglutinating viruses and antibody. II. Further ex-perience with antibody determinations and descriptionof a technique for analysis of virus mixtures. J. Immu-nol. 89:805-814.
8. Drescher, J., A. V. Hennessy, and F. M. Davenport. 1962.Photometric methods for the measurement of hemag-glutinating viruses and antibody. I. Further experiencewith a novel photometric method for measuring hemag-glutinins. J. Immunol. 89:794-804.
9. Frazekas de St. Groth, S. 1961. Methods in immuno-chemistry of viruses. 2. Evaluation of parameters fromequilibrium measurements. Aust. J. Exp. Biol.39:563-582.
10. Freundlich, H. 1928. Colloid and capillary chemistry, p.994. Dutton, New York.
11. Hennessy, A. V., and F. M. Davenport. 1972. Effect ofneuraminidase on antibody combining unit (ACU)titers of human sera determined by Drescher's photo-metric procedure. Proc. Soc. Exp. Biol. Med. 141:6f59-660.
12. Kilbourne, E. D. 1968. Recombination of influenza Aviruses ofhuman and animal origin. Science 160:74-76.
13. Seto, J. T., and R. Rott. 1966. Functional significance ofsialidase during influenza virus multiplication. Virol-ogy 30:731-737.
14. Tamm, I., and F. L. Horsfall, Jr. 1952. Mucoproteinderived from human urine which reacts with influenza,mumps and Newcastle disease virus. J. Exp. Med.95:71-97.
15. Warren, L. 1959. The thiobarbituric acid assay of sialicacids. J. Biol. Chem. 234:1971-1975.
16. Webster, R. G., and W. G. Laver. 1971. Antigenicvariation in influenza virus. Biology and chemistry.Progr. Med. Virol. 13:271-338.
1368 J. VIROL.
on July 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from















![BACTERIAL HEMAGGLUTINATION AND HEMOLYSIS · 1956] BACTERIALHEMAGGLUTINATIONANDHEMOLYSIS 167 Bacteria, too, cause agglutination of erythro- ofbacterial hemagglutination. (3) Specifichemag-](https://img.pdfslide.net/doc/110x75/5e099b8a8848c3026b1ffb45/bacterial-hemagglutination-and-hemolysis-1956-bacterialhemagglutinationandhemolysis.jpg)













