Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Dec. 1972, p. 1321-1334Copyright 0 1972 American Society for Microbiology
Vol. 112, No. 3Printed in U.S.A.
Mechanism of Inhibition of DeoxyribonucleicAcid Synthesis in Escherichia coli by
HydroxyureaNAVIN K. SINHA' AND D. P. SNUSTAD
Department of Genetics and Cell Biology, University of Minnesota, St. Paul, Minnesota 55101
Received for publication 14 September 1972
The effects of hydroxyurea on Escherichia coli B/5 physiology (increases in cellmass, number of viable cells, and deoxyribonucleic acid [DNA], RNA, andprotein concentrations) were studied in an attempt to find a concentration thatcompletely inhibits DNA synthesis and increase in number of viable cells but haslittle or no effect on other metabolic processes. These conditions were the mostclosely approached at an hydroxyurea concentration of 0.026 to 0.033 M. Aconcentration of 0.026 or 0.033 M was used in subsequent experiments to studythe site(s) of inhibition of DNA synthesis in E. coli B/5 by hydroxyurea.Hydroxyurea at a concentration of 10-2 M was found to inhibit ribonucleosidediphosphate reductase activity completely in crude extracts of E. coli. Thesynthesis of deoxyribonucleotides was greatly reduced when E. coli cells were
grown in the presence of 0.033 M hydroxyurea. Studies on the acid-soluble DNAprecursor pools showed that hydroxyurea causes a decrease in the concentrationof deoxyribonucleoside diphosphates and deoxyribonucleoside triphosphates andan increase in the total concentration of ribonucleotides. Sucrose densitygradient sedimentation of DNA from cells treated with 0.026 M hydroxyurea for30 min indicated that at this concentration hydroxyurea induces no detectablesingle- or double-strand breaks. In addition, both replicative and repairsyntheses of DNA were found to occur normally in toluene-treated cells in thepresence of relatively high concentrations of hydroxyurea. Pulse-chase studiesshowed that deoxyribonucleotides synthesized prior to the addition of hydrox-yurea to cells are utilized normally for DNA synthesis in the presence ofhydroxyurea. On the basis of these observations, we have concluded that theprimary, if not the only, site of inhibition of DNA synthesis in E. coli B/5 by lowconcentrations of hydroxyurea is the inhibition of the enzyme ribonucleosidediphosphate reductase.
Hydroxyurea (HU) has been shown to inhibitspecifically deoxyribonucleic acid (DNA) syn-thesis in bacteria (14, 40). This compound alsoinhibits DNA synthesis in mammalian cellsand possesses antineoplastic properties. De-spite intensive study of the effects of HU onbacterial metabolism over the last few years,considerable disagreement as to the exactmechanism of inhibition of DNA synthesis bythis chemical still exists.
It has been suggested by several workers thatHU inhibits DNA synthesis by interfering withthe formation of deoxyribonucleotide precur-sors of DNA by the enzyme ribonucleoside
I Present address: Department of Biology, MassachusettsInstitute of Technology, Cambridge, Mass. 02139.
diphosphate reductase (7, 12, 21, 25, 31, 45).Elford (12) has shown that HU drasticallyinhibits the ribonucleoside diphosphate reduc-tase activity in crude extracts of Escherichiacoli, but Rosenkranz and Jacobs (41) havepresented results contradicting this claim.HU has also been reported to inhibit theactivity of purified ribonucleotide reductasefrom E. coli (21). Of the four components of thisenzyme system (thioredoxin, thioredoxin re-ductase, and proteins Bi and B2), only proteinB2 is sensitive to this inhibitor. This proteinpossesses a sharp absorption peak at 410 nmthat originates from an iron-containing compo-nent of the enzyme (7). Upon incubation withHU, the inactivation of the protein B2 of the E.coli ribonucleotide reductase system is accom-
1321
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

SINHA AND SNUSTAD
panied by the loss of the 410-nm absorptionpeak. These observations led to the suggestionthat HU causes an inhibition of DNA synthesisby conversion of the iron-containing componentof protein B2 to an inactive form (7, 36).However, addition of iron to purified ribonu-cleotide reductase does not reverse the inhibi-tory effects of HU (25). Furthermore, hydrox-ylamine and hydroxyurethane, neither of whichis a specific inhibitor of DNA synthesis, alsoinhibit purified ribonucleotide reductase activi-ty. In the case of hydroxylamine, it has beenshown that the protein B2 is the site of inhibi-tory action, and the loss of reductase activity isaccompanied by a loss of the absorption peak at410 nm. The effects of HU on DNA synthesis inE. coli are completely reversible for at least 2hr, and this reversal can be accomplished in theabsence of protein synthesis (37), whereas theinhibition of purified ribonucleotide reductaseis not reversed by removal of the inhibitor (21).
Neuhard (31) and Neuhard and Thomassen(33) found the addition of HU to bacteria to re-sult in a decrease in the size of the acid-solubledeoxyribonucleoside triphosphate pools. Acid-soluble pool studies of Rosenkranz and Jacobs(41), on the other hand, led them to the conclu-sion that HU does not inhibit the synthesis ofdeoxyribonucleotides but rather the conversionof deoxyribonucleoside diphosphates to theirtriphosphate derivatives. To evaluate theseconflicting interpretations properly, the effectsof HU on the total ribonucleotide and de-oxyribonucleotide pools need to be studied.Warner and Hobbs (45) found that in phageT4-infected E. coli HU produces an increase inthe total ribonucleotide pools and a pronounceddecrease in the total deoxyribonucleotide pools.
If ribonucleotide reductase is the only site ofaction of this inhibitor, then at least a partialreversal might be expected after addition ofdeoxyribonucleosides to HU-inhibited cells.Such reversals have indeed been observed insome types of mammalian tissue cultures (2,3, 13, 24, 48), but none has been obtained inbacteria (38). -This failure to obtain such areversal by adding deoxyribonucleosides to bac-teria is expected since bacteria, unlike mam-malian cells, apparently lack nucleoside kinaseactivity specific for deoxyadenosine, deoxycyti-dine, and deoxyguanosine (18).An alternative hypothesis as to the mecha-
nism of inhibition of DNA synthesis by HU hasbeen proposed by Rosenkranz et al. (38). Theysuggest that HU inhibits DNA synthesis bycausing a disturbance in the structure of theDNA template, thus making it incapable of di-recting normal DNA replication.
In this paper, we report the results of studies
on the effects of HU on acid-soluble DNA pre-cursor pools and ribonucleotide reductase ac-tivity in E. coli, performed in an attempt to es-tablish whether HU inhibits the reduction ofribonucleotides in uninfected E. coli. In addi-tion, we have investigated the effects of HU onDNA replication and repair in toluene-treatedcells and examined the integrity of DNA fromHU-treated cells to evaluate the suggestionthat HU might be inhibiting DNA synthesis bya mechanism distinct from any effects on ribo-nucleotide reduction. Also, pulse-chase experi-ments were done in an attempt to detect anyHU-induced metabolic blocks in the pathway ofDNA synthesis by intact cells subsequent to theribonucleoside diphosphate reduction step.
MATERIALS AND METHODSMedia and bacterial strains. H broth (nutrient
broth, 8 g [Difco]; peptone, 5 g [Difco]; sodiumchloride, 5 g; glucose, 1 g; distilled water, 1 liter), Tbroth (tryptone, 10 g [Difco]; sodium chloride, 5 g;distilled water, 1 liter), EHA bottom layer agar (agar,10 g [Difco]; tryptone, 13 g [Difco]; sodium chloride,8 g; sodium citrate [dihydratel, 2 g; glucose, 1.3 g;distilled water, 1 liter), and EHA top-layer agar (agar,6.5 g Pifco]; tryptone, 13 g Pifco]; sodium chloride,8 g; sodium citrate [dihydrate], 2 g; glucose, 3 g;distilled water, 1 liter) (11) were used for growth ofovernight bacterial cultures, dilutions, and plating,respectively. M9CA medium (M9 medium of Adams[1 ] supplemented with 5 jg of thymidine per ml, 0.2%glycerol, and 0.3% Casamino Acids [Difco] which wasadded as a charcoal-filtered solution) was used forgrowth of bacteria throughout this study. E. coli B/5,a prototroph, was used in all experiments.
Radioisotopes and scintillation counting. Thy-midine-2-14C (55.7 mCi/mmole) and uracil-2- 4C (18.6mCi/mmole) were obtained from InternationalChemical and Nuclear Corp., Irvine, Calif. Thymi-dine-methyl-3H (11.3 Ci/mmole), [5- 3H]cytidine-5-diphosphate (13.5 Ci/mmole) and deoxyadenosine-5'-triphosphate-8-_ H-tetralithium (5.54 Ci/mmole)were obtained from Schwarz Bioresearch Inc.,Orangeburg, N.Y.
2, 5-Diphenyloxazole (PPO) and 1, 4-bis-2-(4-methyl-5-phenyloxazolyl)-benzene (dimethyl POPOP)were obtained from Packard Instrument Co., Down-er's Grove, Ill.Two scintillation fluids were used. For nonaqueous
materials, a solution containing 4 g of PPO and 0.25 gof dimethyl POPOP per liter of toluene was used. Foraqueous solutions, Bray solution (6) was used. Theradioactivity was counted in a Packard Tri-Carbliquid scintillation spectrometer.
Chemicals. HU was purchased from CalbiochemInc., Wood Dale, Ill. Polyethyleneimine-cellulose-coated plastic sheets for thin-layer chromatography(TLC) were obtained from Brinkman Instruments,Westbury, N.Y. Pancreatic deoxyribonuclease andribonuclease were purchased from Worthington Bio-chemical Corp., Freehold, N.J.Measurement of the effects of HU on cellular
metabolism. Cell mass was measured with a Beck-
1322 J. BACTERIOL.
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

VOL. 112, 1972 MECHANISM OF INHIBITI
man DB spectrophotometer at 650 nm. The numberof viable cells was determined by plating an appro-priate dilution on EHA bottom-layer agar plates. Fordetermination of DNA, ribonucleic acid (RNA), andprotein concentrations, cells were withdrawn at vari-ous times after addition of HU to log-phase culturesof E. coli B/5, and macromolecules were precipitatedby adding trichloroacetic acid to a final concentrationof 5%. The trichloroacetic acid precipitates wereallowed to sit in an ice bath for 30 min, after whichthey were centrifuged and washed three times with5-ml quantities of cold 5% trichloroacetic acid solu-tion before being dissolved in 5 ml of 0.1 M NaOHsolution. DNA concentration was determined by theindole assay (44); RNA was determined by the orcinolassay (10). Protein content was measured by themethod of Lowry et al. (22). Salmon sperm DNA,yeast RNA, and trypsin were used as standards in theabove assays.
Ribonucleotide reductase assays. Three liters ofexponentially growing cells (approximately 3 x 101cells/ml in M9CA medium) were chilled by addingcrushed ice, harvested by centrifugation, and eitherused immediately or frozen for up to 1 week for futureuse. The cell extracts were prepared, enzyme assaysperformed, and Dowex 5OW column chromatographywas done as described by Yeh et al. (47). The fractionsfrom the column were analyzed for radioactivity byadding 2 ml of each fraction to 10 ml of Bray solutionand counting in a scintillation spectrometer as de-scribed earlier.
Acid-soluble pool studies. Acid-soluble pyrimi-dine pools were labeled by growing log-phase cells ofE. coli B/5 in the presence of 0.1 Mmole of "4C-uracil(3.2 x 107 counts per min per Mmole; 18.6 mCi/mmole) per ml for 15 min. At the desired intervals,1-ml samples were withdrawn, and the cells werecollected by membrane filtration (Millipore Corp.).The cells were washed on the filters with three 2-mlportions of cold M9CA medium, and the membranefilters (Millipore Corp.) quickly immersed in 5 ml ofcold 5% trichloroacetic acid. The filters were vigor-ously shaken in the trichloroacetic acid solution for 15seconds and then allowed to stand in an ice bath for30 min or longer. The trichloroacetic acid precipitateswere centrifuged, and the supernatant fluid wasextracted 10 times with equal volumes of cold etherbefore being lyophilized to dryness. The residues weredissolved in 0.1 ml of water, and 10-Mliter sampleswere placed on PEI-cellulose plates with referencenucleotides (25 nmole of each) for TLC. To estimatethe amount of "4C-uracil incorporated into DNA, thetrichloroacetic acid precipitates were dissolved in 0.5ml of 0.3 M KOH and incubated for 18 hr at 30 C.Five ml of 10% trichloroacetic acid was added to eachsample, and the precipitates were allowed to sit in anice bath for 30 min. The precipitates were thencentrifuged and washed three times with 5-ml vo-lumes of cold 5% trichloroacetic acid before beingdissolved in 2-ml volumes of 0.1 M NaOH. Samples(1 ml) were then dried in scintillation vials andcounted.
PEI-cellulose plates were prepared for TLC as
described by Randerath and Randerath (34). Beforecarrying out two-dimensional chromatography to sep-
[ON OF DNA SYNTHESIS 1323
arate nucleotides, the TLC plates were washed withtris(hydroxymethyl)aminomethane (Tris)-methanoland methanol-water solutions as described by Neu-hard et al. (32) to remove salts and other interferingsubstances.
Monophosphates were separated by the method ofRanderath and Randerath (35). Di- and triphosphateswere separated by the method of Neuhard (30). Afterchromatographic separation, the plates were dried;the nucleotide-containing (as indicated by ultravioletabsorption) spots were cut out; and the radioactivityin each was counted.
Preparation of labeled DNA. Two identicalcultures of E. coli B/5 were grown for 6 to 8generations in M9CA medium supplemented with 0.5mg of uridine per ml. One culture also contained 0.5,sCi of '4C-thymidine per ml, and the second con-tained 25 MCi of 3H-thymidine per ml. When the celldensity reached about 4 x 101 cells/ml, 2-ml sampleswere withdrawn, filtered on membrane filters (Mil-lipore Corp.), washed with 10 ml of cold M9CAmedium, and resuspended in 2 ml of prewarmedM9CA medium. The "4C-labeled cells were added to amedium that also contained 26 mm HU. Both cul-tures were incubated another 30 min at 30 C withvigorous aeration. The two cultures were then pooled,and the DNA was extracted by using a gentleextraction procedure developed by T. Lash (Ph.D.thesis, Yale University, 1971). The DNA moleculesextracted by this procedure have a molecular weightof 1.5 x 108 to 2.5 x 108.
This extraction procedure involves freezing andthawing cells in the presence of an equal volume of30% sucrose in 0.1 M Tris-0.1 M ethylenediaminetet-raacetic acid (EDTA), pH 8.5, with a small amount(0.1 ml) of 0.5 M NaN,. This is followed by lysozymetreatment (final concentration of the enzyme, 4 x 10'units/ml) at 0 C for 15 min. The spheroplasts arelysed by addition of Sarkosyl to a final concentrationof 0.1%. RNA is digested with ribonuclease (finalconcentration, 40 gg/ml) at 37 C for 30 min. ThenPronase (a freshly prepared solution in 0.1 M Tris-chloride, pH 8.5) is added as an 80 mg/ml solution toachieve a final concentration of 2 mg/ml. Pronasetreatment is continued for 15 to 18 hr at 37 C. Beforeuse, both ribonuclease and Pronase were heated to 80C for 10 min and then quickly chilled in an ice bath toinactivate any contaminating deoxyribonucleases.After the Pronase treatment, the DNA solution isdialyzed against a buffer containing 0.01 M Tris,0.005 M EDTA, and 0.02 M sodium chloride. (One mlof 95% ethanol is added to each liter of dialysis bufferjust before use. The dialysis bags are prepared for useby boiling twice in 0.5% sodium carbonate-0.05%sodium citrate buffer, pH 8.6, and then once indistilled water.) The DNA solution is dialyzed againstthree consecutive 1-liter volumes of buffer over 24 hr(8 hr in each) at room temperature.
Sucrose density gradient centrifugation. Su-crose was dissolved in 2 M sodium chloride solution(containing 10-8 M EDTA) at pH 8.5 (0.01 M Tris) forneutral gradients and at pH 12.2 (0.02 M sodiumphosphate) for alkaline gradients. Gradients werealways prepared with freshly dissolved solutions ofsucrose. Linear sucrose gradients of 2.4 ml each of 5%
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

SINHA AND SNUSTAD
(w/v) and 20% (w/v) sucrose solutions were preparedas described by Martin and Ames (23) in 5-mlnitrocellulose tubes that had been pretreated with asilicone preparation (Siliclad) to prevent DNA fromsliding to the bottom of the tube (8). All gradientswere stored in the cold for at least 60 min before use.The DNA preparations (0.3 ml) were gently layeredon the gradients.
Centrifugations were carried out with an SW50.1Beckman rotor in a Beckman L2-65B centrifuge. Allcentrifugations were done at 18 C for 15 to 20 hr at20,000 rev/min. Ten-drop fractions were collected bypuncturing a hole in the bottom of the centrifugetube. The fractions were collected directly on What-man GF/A glass-fiber discs in scintillation vials anddried at 60 C, and the radioactivity was counted.DNA synthesis in toluene-treated cells. E. coli
B/5 cells were grown to a cell density of 5 x 108 to 6 x108 cells/ml, harvested by centrifugation, and treatedwith toluene as described by Moses and Richardson(27). DNA-synthesizing capacity was determined asdescribed by Moses and Richardson (28). Assaymixtures containing all the necessary componentswere prepared in a total volume of 0.18 ml to whichwas added 0.02 ml of a freshly prepared aqueoussolution of HU to obtain the desired final concentra-tion of the inhibitor in each assay tube. The tubeswere preincubated at 30 C for 15 min before adding108 to 2 x 108 cells in 0.1 ml of 0.05 M potassiumphosphate buffer. DNA synthesis was allowed toproceed for 15 min at 30 C.
Pulse-chase studies. A 16-ml culture was grownin M9CA medium at 30 C with vigorous aeration to acell density of approximately 4 x 108 cells per ml.I4C-uracil was added to a final concentration of 2Ci/ml (approximately 0.1 gmole/ml), and the cellswere incubated for another 25 min. A 1-ml samplewas then withdrawn, and the cells were collected on amembrane filter (Millipore Corp.) and washed threetimes with 2 ml of cold M9CA medium. Thereafter,the filter and the cells were shaken for 15 sec in 0.5 mlof ice-cold 1 M perchloric acid. Immediately afterremoval of the above sample, two 7-ml samples werewithdrawn and washed free from radioactive uracil bymembrane filtration (Millipore Corp.). The cells fromone of the 7-ml samples were suspended in 7 ml ofprewarmed M9CA medium supplemented with 2.5gmole of cold uracil per ml. The cells from the othersample were suspended in 7 ml of M9CA mediumthat also contained HU (0.033 M) in addition to colduracil (2.5 gmole/ml). Both cultures were grown at 30C with aeration. Samples (1 ml) were withdrawn atthe desired intervals, and the cells were collected onmembrane filters (Millipore Corp.) and washed withcold M9CA. The filters were then quickly immersedin 0.5 ml of 1 M perchloric acid and shaken toprecipitate macromolecules. Bovine serum albumin(1 mg) was added to each tube, and the precipitateswere allowed to sit in an ice-bath for 60 min. Theprecipitates were collected by centrifugation, RNAwas hydrolyzed, and the radioactivity in DNA wasestimated as previously described.The perchloric acid supernatant fluids were placed
in a boiling-water bath for 10 min to hydrolyze
nucleoside polyphosphates to nucleoside monophos-phates. The samples were brought to room tempera-ture and neutralized by using 8 M KOH solutioncontaining a small quantity of phenol red as a pHindicator. The precipitates were then centrifuged,and the supernatant fluids were used for the estima-tion of labeled ribo- and deoxyribonucleotides byTLC. A 0.1-ml sample of this supematant fluid waswithdrawn, and the radioactivity was counted afterthe addition of 10 ml of Bray solution to estimate thetotal acid-soluble radioactivity.PEI cellulose-coated plates were prepared for TLC
as described earlier. Ten-microliter amounts of thesamples were placed 1.5 cm apart along with 25nmoles of each reference nucleotide on a PEI-cel-lulose plate. Subsequent to application of the sam-ples, the TLC plates were washed with Tris-methanoland anhydrous methanol as described by Neuhard etal. (32). Ribonucleotides were separated from deox-yribonucleotides by one-dimensional chromatogra-phy in a solvent composed of 6 g of sodium tetrabo-rate, 3 g of boric acid, 25 g of ethylene glycol, and 70ml of water. The solvent front was allowed to ascendapproximately 15 cm above the origin. In this system,uracil, uridine, cytidine, deoxyuridine, deoxycyti-dine, and thymidine migrate with a RF between 0.7and 0.8. Pyrimidine-containing deoxyribonucleotides(deoxyuridine monophosphate, thymidine monophos-phate, and deoxycytosine monophosphate) move withan RF of about 0.5, and pyrimidine-containing ribonu-cleotides (cytosine monophosphate and uridine mon-ophosphate) possess a RF of approximately 0.28. Afterchromatography, the TLC plates were dried, and theradioactivity in nucleotides was estimated as de-scribed earlier.
RESULTSEffects of HU on bacterial metabolism.
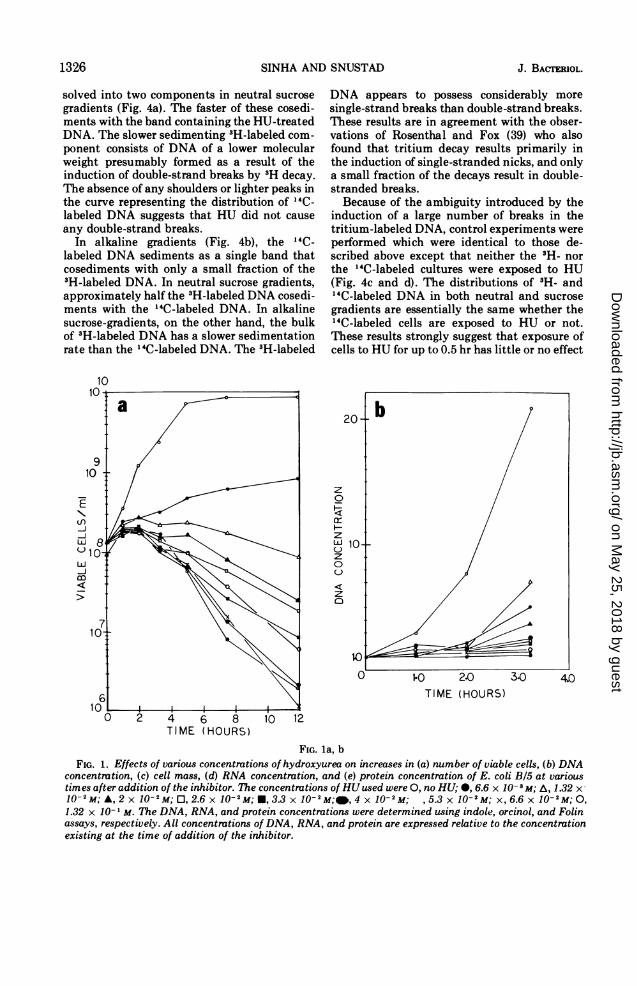
Preliminary studies indicated that the concen-tration of HU (0.1-0.2 M) commonly used byother workers (31, 37, 41, 42) in studies with E.coli resulted not only in an inhibition of DNAsynthesis but also in a pronounced inhibition ofRNA and protein synthesis. Therefore, theeffects of various concentrations of HU on E.coli B/5 metabolism, i.e., number of viablecells, cell mass, and DNA, RNA, and proteinconcentrations, were examined in an attempt tofind a concentration that completely inhibitsDNA synthesis and increase in number ofviable cells but has little or no effect on othermetabolic processes.
Increase in the number of viable cells (Fig. la)and DNA synthesis (Fig. lb) are the processesmost sensitive to HU. Number of viable cellsceases to show a prolonged increase at anyconcentration of HU higher than 0.02 M. Theeffect of HU on number of viable cells isreversible for about 2 hr, after which a slowdevitalization begins. The rate of cell death isdependent upon the concentration of HU pres-
1324 J. BACTERIOL.
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

MECHANISM OF INHIBITION OF DNA SYNTHESIS
ent (Fig. la). DNA synthesis is completely(greater than 95%) inhibited by an HU concen-tration of 0.026 M (Fig. lb). At none of the HUconcentrations used was there a net decrease inthe amount of DNA present, indicating thatthis compound does not induce a significantamount of degradation of DNA to acid-solublematerial. A study of the effects of HU onincreases in cell mass (Fig. lc), and RNA (Fig.ld) and protein (Fig. le) concentrations re-vealed that these processes are much less sensi-tive to the effects of this inhibitor. Nevertheless,there is a small, but significant, decrease in therates of increase in cell mass and RNA andprotein synthesis with each increase in HUconcentration. Therefore, in all further experi-ments, the lowest HU concentration giving acomplete inhibition of DNA synthesis(0.026-0.033 M) was used to minimize the othereffects of the inhibitor. Also, treatment wasalways for less than 2 hr to avoid irreversibledamage to the cells.When the effect of HU on the incorporation of
radioactive thymidine into DNA of E. coli B/5was investigated, a significant amount of labelwas found to be incorporated into trichloroa-cetic acid-insoluble material in the presence of0.026 M and even higher concentrations of HU(N. K. Sinha, unpublished results). This incor-poration probably represents repair synthesis.It may be mentioned however that no differencein the rates of thymidine incorporation wasobserved in a polA+ strain and its polA mutantderivative.CDP reductase activities. Since con-
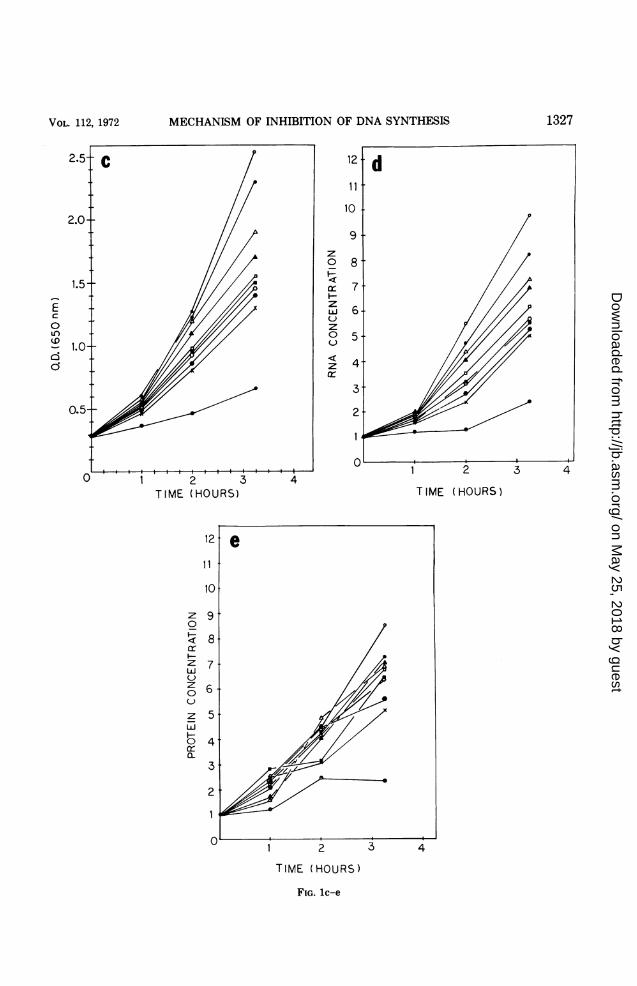
tradicting results have been reported by Elford(12) and Rosenkranz and Jacobs (41) regardingthe inhibition of ribonucleotide reductase incrude extracts of E. coli by HU, these experi-ments were repeated in this study (Fig. 2). AtHU concentrations of 10-3 M and higher, thecytidine diphosphate (CDP) reductase activitypresent in crude extracts of E. coli B/5 wasmarkedly reduced. The degree of inhibitionobtained with 0.01 M HU varied from 80 to100%. Our results are thus in agreement withthose of Elford (12) and contradict those ofRosenkranz and Jacobs (41).
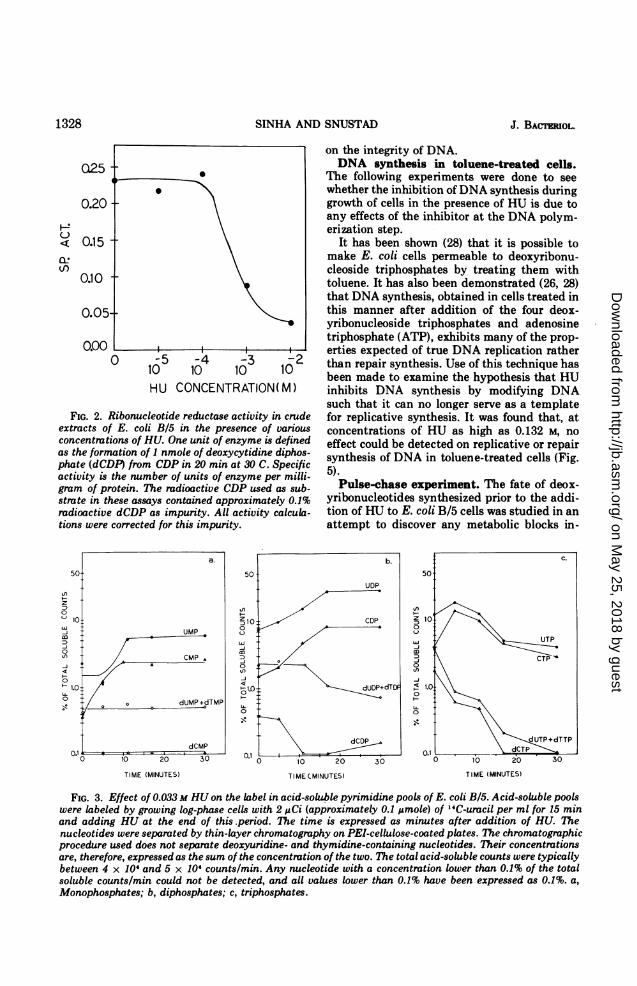
Effects of HU on ribonucleotide reduc-tion in vivo. The effect(s) of HU on the relativesizes of the ribo- and deoxv *ibonucleotide poolswas examined to see whether HU caused aninhibition of ribonucleotide reduction in vivo.The acid-soluble pyrimidine pool was labeled,HU was added, and the amount of radioactivitypresent in ribo- and deoxyribonucleosidemono-, di-, and triphosphates was examined avarious times (Fig. 3a, b, and c; Table 1). After
the addition of HU, the amount of label inribonucleoside mono- and diphosphates in-creases, whereas the labeled ribonucleoside tri-phosphates show an initial increase followed bya decrease in concentration. The deoxyribonu-cleoside monophosphates are present in quitelow amounts, and no change in their levels isseen, but addition of HU results in a decrease inthe concentration of deoxyribonucleoside di-and triphosphates. The decrease is especiallypronounced in the case of the triphosphates.Table 1 shows the total amounts of labeleduracil- and cytosine-containing nucleotides andtheir distribution in ribo- and deoxyribonucleo-tides. It is obvious from these results that bothuracil- and cytosine-containing ribonucleotidesshow an increase in their total amounts andthat the deoxyribonucleotides show a decreasein their total concentrations after the additionof HU to bacteria. In addition, the proportion oflabel present in nucleotides increases in thepresence of HU. The decrease in the amount ofdeoxyribonucleotides would be even more pro-nounced than those shown in Table 1 if thevalues expressed were corrected for the ob-served expansion of the nucleotide pools.Effects of HU on DNA integrity. A
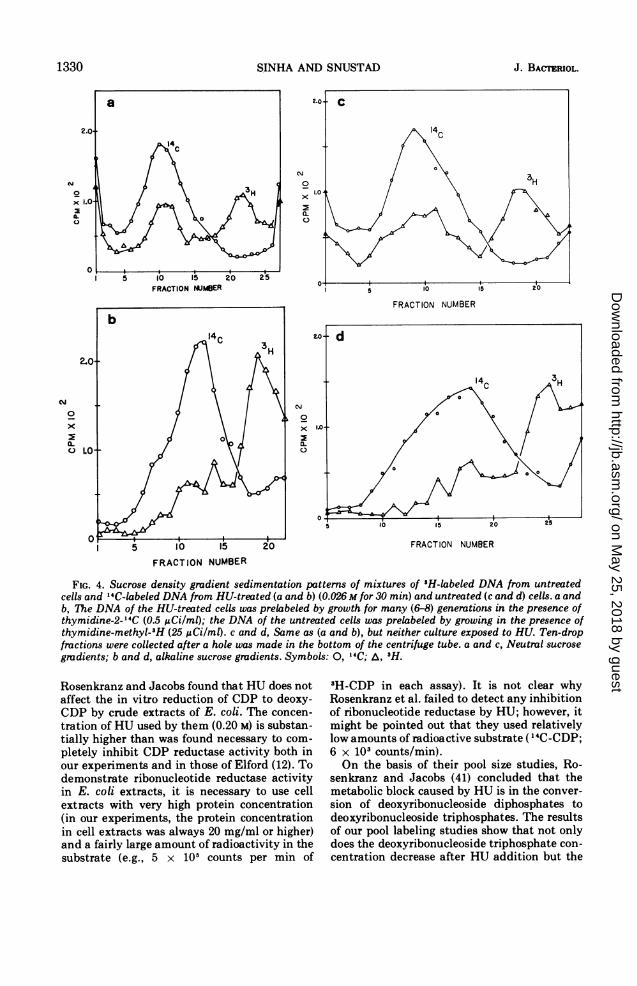
culture of E. coli B/5 was divided into twosubcultures; one subculture was labeled with"4C-thymidine and the other with 3H-thymi-dine. The radioactive precursors were subse-quently removed by membrane filtration (Mil-lipore Corp.). The cells with "4C-labeled DNAwere grown in M9CA medium supplementedwith HU (final concentration 0.026 M), andthose with 3H-labeled DNA were grown inunsupplemented medium, both for an addi-tional 30 min at 30 C. The treated and un-treated cells were then pooled, and their DNAwas extracted by a gentle extraction procedure.Mixing the cells before extraction insured thatany differences in the DNA of the two subcul-tures would be due to the effect(s) of HUtreatment rather than to any differences in thehandling procedure. Induction of any breaks inthe DNA of the treated cells due to HUtreatment will result in this DNA having alower, average molecular weight than the DNAfrom untreated cells. The cells with "4C-labeledDNA, rather than the 3H-labeled ones, weretreated with HU because tritium decay isknown to cause single- and double-strandbreaks in DNA (39). The labeled DNA mixturewas analyzed for HU-induced breaks in thestructure by sedimentation through neutral andalkaline gradients. The results of sucrose densi-ty gradient sedimentation are shown in Fig. 4aand b. The 3H-labeled (control) DNA is re-
VOL. 112, 1972 1325
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
SINHA AND SNUSTAD
solved into two components in neutral sucrosegradients (Fig. 4a). The faster of these cosedi-ments with the band containing the HU-treatedDNA. The slower sedimenting 'H-labeled com-ponent consists of DNA of a lower molecularweight presumably formed as a result of theinduction of double-strand breaks by 3H decay.The absence of any shoulders or lighter peaks inthe curve representing the distribution of "4C-labeled DNA suggests that HU did not causeany double-strand breaks.
In alkaline gradients (Fig. 4b), the "4C-labeled DNA sediments as a single band thatcosediments with only a small fraction of the3H-labeled DNA. In neutral sucrose gradients,approximately half the 3H-labeled DNA cosedi-ments with the "4C-labeled DNA. In alkalinesucrose-gradients, on the other hand, the bulkof 3H-labeled DNA has a slower sedimentationrate than the "4C-labeled DNA. The 3H-labeled
10
N..
-J-LJO 1(uLJ>
DNA appears to possess considerably moresingle-strand breaks than double-strand breaks.These results are in agreement with the obser-vations of Rosenthal and Fox (39) who alsofound that tritium decay results primarily inthe induction of single-stranded nicks, and onlya small fraction of the decays result in double-stranded breaks.Because of the ambiguity introduced by the
induction of a large number of breaks in thetritium-labeled DNA, control experiments wereperformed which were identical to those de-scribed above except that neither the 'H- northe "4C-labeled cultures were exposed to HU(Fig. 4c and d). The distributions of 'H- and"4C-labeled DNA in both neutral and sucrosegradients are essentially the same whether the"4C-labeled cells are exposed to HU or not.These results strongly suggest that exposure ofcells to HU for up to 0.5 hr has little or no effect
4.0TIME (HOURS)
12
FIG. la, bFIG. 1. Effects of various concentrations of hydroxyurea on increases in (a) number of viable cells, (b) DNA
concentration, (c) cell mass, (d) RNA concentration, and (e) protein concentration of E. coli B/5 at varioustimes afteraddition of the inhibitor. The concentrations of HUused were 0, no HU; 0, 6.6 x 10-3M; A, 1.32 x-10'-M; A, 2 x 10-2M; O, 2.6 x 10-2M; *, 3.3 x 10-2M;@,4xj10-2M; ,5.3x10-2M; x,6.6x10-2M;O,1.32 x 10- ' M. The DNA, RNA, and protein concentrations were determined using indole, orcinol, and Folinassays, respectively. All concentrations of DNA, RNA, and protein are expressed relative to the concentrationexisting at the time of addition of the inhibitor.
1326 J. BACTERIOL.
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
MECHANISM OF INHIBITION OF DNA SYNTHESIS
z0
a::iHzzLUJC)z0
z
TIME (HOURS)
1 2
TIME (HOURS)
FIG. lc-e
2
T IME (HOURS)
3 4
Ec
o-)
11
z0I-
zLiJC)z0C-)
z
HL05
1327VoL. 112, 1972
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
SINHA AND SNUSTAD
on the integrity of DNA.DNA synthesis in toluene-treated cells.
44* The following experiments were done to see* \ whether the inhibition of DNA synthesis during
growth of cells in the presence of HU is due toany effects of the inhibitor at the DNA polym-erization step.
It has been shown (28) that it is possible tomake E. coli cells permeable to deoxyribonu-cleoside triphosphates by treating them withtoluene. It has also been demonstrated (26, 28)that DNA synthesis, obtained in cells treated inthis manner after addition of the four deox-yribonucleoside triphosphates and adenosinetriphosphate (ATP), exhibits many of the prop-
>5 4 3
I2
erties expected of true DNA replication rather0 10 10 10 10 2 than repair synthesis. Use of this technique has
been made to examine the hypothesis that HUHU CONCENTRATION( M) inhibits DNA synthesis by modifying DNA
such that it can no longer serve as a templateRibonucleotide reductase activity in crude for replicative synthesis. It was found that, atf E. coli B/5 in the presence of various concentrations of HU as high as 0.132 M, notions of HU. One unit of enzyme is defined effect could be detected on replicative or repairnation of 1 nmole of deoxycytidine diphos- synthesis of DNA in toluene-treated cells (Fig.DP) from CDP in 20 min at 30 C. Specific 5 oc!thD n4ImhUr nf 1iIte nf anJy&mo nor miMiL )-acctutty Ms cne numuer of unuls of enzyme per mitus-
gram of protein. The radioactive CDP used as sub-strate in these assays contained approximately 0.1%radioactive dCDP as impurity. All activity calcula-tions were corrected for this impurity.
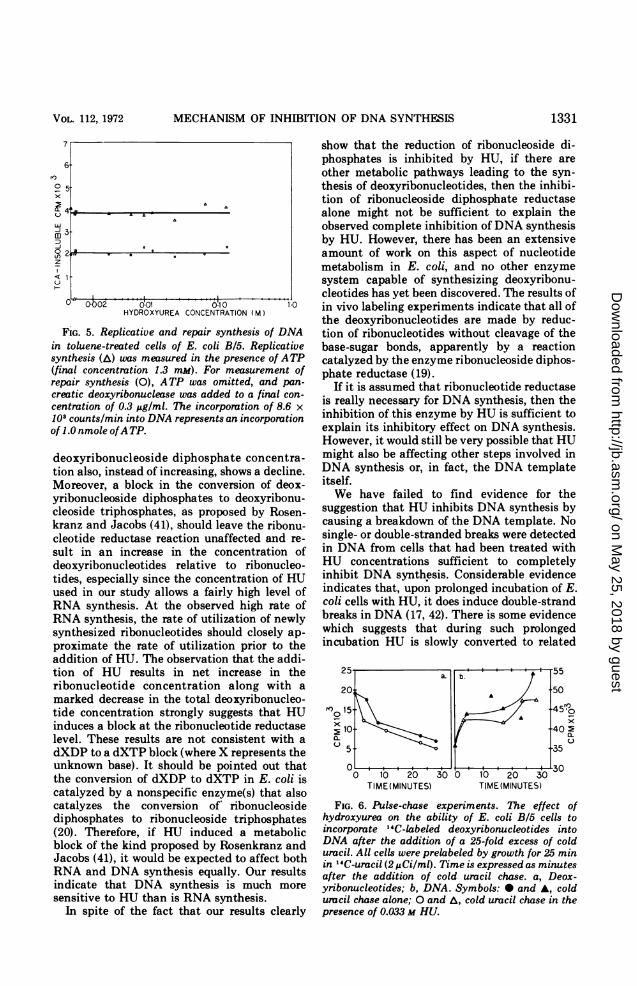
Pulse-chase experiment. The fate of deox-yribonucleotides synthesized prior to the addi-tion of HU to E. coli B/5 cells was studied in anattempt to discover any metabolic blocks in-
50
Z 1000
J
coJ
0< 1.0.0
0.10 10 20 30 , 10 20 30 °0 10 2 30
TIME (MINUTES) TIME (MINUTES) TIME (MINUTES)
FIG. 3. Effect of 0.033 M HU on the label in acid-soluble pyrimidine pools of E. coli B/5. Acid-soluble poolswere labeled by growing log-phase cells with 2 1ACi (approximately 0.1 gmole) of '4C-uracil per ml for 15 minand adding HU at the end of this.period. The time is expressed as minutes after addition of HU. Thenucleotides were separated by thin-layer chromatography on PEI-cellulose-coated plates. The chromatographicprocedure used does not separate deoxyuridine- and thymidine-containing nucleotides. Their concentrationsare, therefore, expressed as the sum of the concentration of the two. The total acid-soluble counts were typicallybetween 4 x 104 and 5 x 104 counts/min. Any nucleotide with a concentration lower than 0.1% of the totalsoluble counts/min could not be detected, and all values lower than 0.1% have been expressed as 0.1%. a,Monophosphates; b, diphosphates; c, triphosphates.
UC-)
FIG. 2.extracts oconcentraias the forrphate (dC.ni-t;llitlv Fe
C.
UTP
\UTP+dTTP\______ dCTP >
1328 J.' BACTOL.
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
MECHANISM OF INHIBITION OF DNA SYNTHESIS
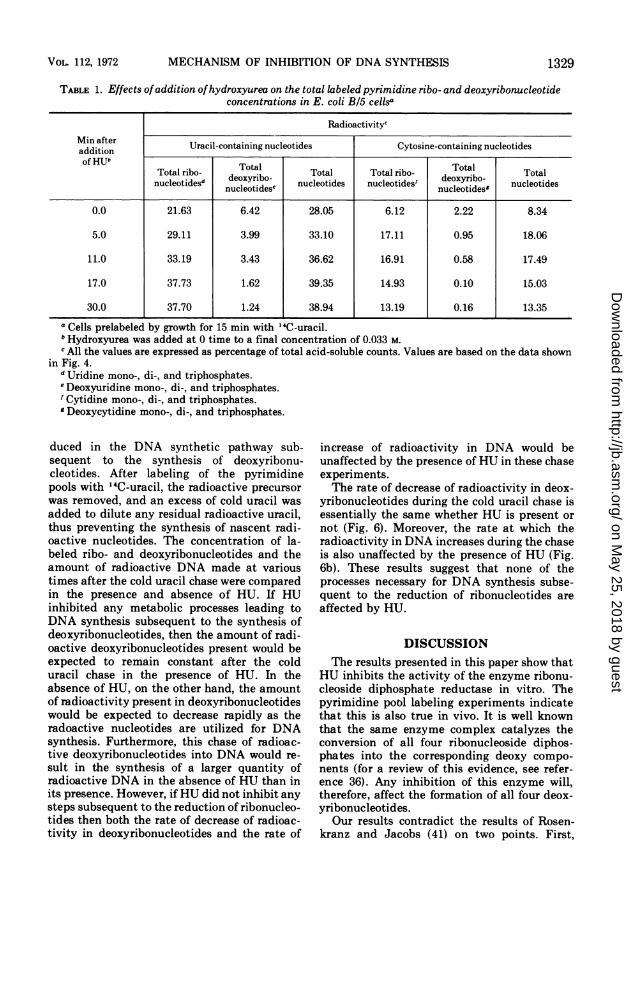
TABLE 1. Effects ofaddition ofhydroxyurea on the total labeledpyrimidine ribo- and deoxyribonucleotideconcentrations in E. coli B/5 cellsa
RadioactivitycMin after Uracil-containing nucleotides Cytosine-containing nucleotidesadditionof HU"
oa oaTotal ribo- Total Total Total ribo- Total TotalnucleotideSd nucleotideSe nucleotides nucleotidest nucleotidesg nucleotides
0.0 21.63 6.42 28.05 6.12 2.22 8.34
5.0 29.11 3.99 33.10 17.11 0.95 18.06
11.0 33.19 3.43 36.62 16.91 0.58 17.49
17.0 37.73 1.62 39.35 14.93 0.10 15.03
30.0 37.70 1.24 38.94 13.19 0.16 13.35
a Cells prelabeled by growth for 15 min with 14C-uracil.° Hydroxyurea was added at 0 time to a final concentration of 0.033 M.c All the values are expressed as percentage of total acid-soluble counts. Values are based on the data shown
in Fig. 4.d Uridine mono-, di-, and triphosphates.e Deoxyuridine mono-, di-, and triphosphates.' Cytidine mono-, di-, and triphosphates.' Deoxycytidine mono-, di-, and triphosphates.
duced in the DNA synthetic pathway sub-sequent to the synthesis of deoxyribonu-cleotides. After labeling of the pyrimidinepools with '4C-uracil, the radioactive precursorwas removed, and an excess of cold uracil wasadded to dilute any residual radioactive uracil,thus preventing the synthesis of nascent radi-oactive nucleotides. The concentration of la-beled ribo- and deoxyribonucleotides and theamount of radioactive DNA made at varioustimes after the cold uracil chase were comparedin the presence and absence of HU. If HUinhibited any metabolic processes leading toDNA synthesis subsequent to the synthesis ofdeoxyribonucleotides, then the amount of radi-oactive deoxyribonucleotides present would beexpected to remain constant after the colduracil chase in the presence of HU. In theabsence of HU, on the other hand, the amountof radioactivity present in deoxyribonucleotideswould be expected to decrease rapidly as theradoactive nucleotides are utilized for DNAsynthesis. Furthermore, this chase of radioac-tive deoxyribonucleotides into DNA would re-sult in the synthesis of a larger quantity ofradioactive DNA in the absence of HU than inits presence. However, ifHU did not inhibit anysteps subsequent to the reduction of ribonucleo-tides then both the rate of decrease of radioac-tivity in deoxyribonucleotides and the rate of
increase of radioactivity in DNA would beunaffected by the presence of HU in these chaseexperiments.The rate of decrease of radioactivity in deox-
yribonucleotides during the cold uracil chase isessentially the same whether HU is present ornot (Fig. 6). Moreover, the rate at which theradioactivity in DNA increases during the chaseis also unaffected by the presence of HU (Fig.6b). These results suggest that none of theprocesses necessary for DNA synthesis subse-quent to the reduction of ribonucleotides areaffected by HU.
DISCUSSIONThe results presented in this paper show that
HU inhibits the activity of the enzyme ribonu-cleoside diphosphate reductase in vitro. Thepyrimidine pool labeling experiments indicatethat this is also true in vivo. It is well knownthat the same enzyme complex catalyzes theconversion of all four ribonucleoside diphos-phates into the corresponding deoxy compo-nents (for a review of this evidence, see refer-ence 36). Any inhibition of this enzyme will,therefore, affect the formation of all four deox-yribonucleotides.Our results contradict the results of Rosen-
kranz and Jacobs (41) on two points. First,
VOL. 112, 1972 1329
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
SINHA AND SNUSTAD
i0 15
FRACTION NUMBER
2.0
0
0x0
C)
x
5 10 Is 20
FRACTION NUMBER
FRACTION NUMBER
FIG. 4. Sucrose density gradient sedimentation patterns of mixtures of 3H-labeled DNA from untreatedcells and 14C-labeled DNA from HU-treated (a and b) (0.026 M for 30 min) and untreated (c and d) cells. a andb, The DNA of the HU-treated cells was prelabeled by growth for many (6-8) generations in the presence ofthymidine-2-"4C (0.5 uCi/ml); the DNA of the untreated cells was prelabeled by growing in the presence ofthymidine-methyl-9H (25 ,gCi/ml). c and d, Same as (a and b), but neither culture exposed to HU. Ten-dropfractions were collected after a hole was made in the bottom of the centrifuge tube. a and c, Neutral sucrose
gmdients; b and d, alkaline sucrose gradients. Symbols: 0, 14C; A, 3H.
Rosenkranz and Jacobs found that HU does notaffect the in vitro reduction of CDP to deoxy-CDP by crude extracts of E. coli. The concen-
tration of HU used by them (0.20 M) is substan-tially higher than was found necessary to com-
pletely inhibit CDP reductase activity both inour experiments and in those of Elford (12). Todemonstrate ribonucleotide reductase activityin E. coli extracts, it is necessary to use cellextracts with very high protein concentration(in our experiments, the protein concentrationin cell extracts was always 20 mg/ml or higher)and a fairly large amount of radioactivity in thesubstrate (e.g., 5 x 105 counts per min of
3H-CDP in each assay). It is not clear whyRosenkranz et al. failed to detect any inhibitionof ribonucleotide reductase by HU; however, itmight be pointed out that they used relativelylow amounts of radioactive substrate ('4C-CDP;6 x 103 counts/min).On the basis of their pool size studies, Ro-
senkranz and Jacobs (41) concluded that themetabolic block caused by HU is in the conver-sion of deoxyribonucleoside diphosphates todeoxyribonucleoside triphosphates. The resultsof our pool labeling studies show that not onlydoes the deoxyribonucleoside triphosphate con-
centration decrease after HU addition but the
1330 J. BACTERIOL.
3H0
X I
C.)
N
0
x
C)
1.0
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
MECHANISM OF INHIBITION OF DNA SYNTHESIS
6rl)
° 5
m3
11
o t02 0*'o1ool,oHYDROXYUREA CONCENTRATION (M)
FIG. 5. Replicative and repair synthesis of DNAin toluene-treated cells of E. coli B/5. Replicativesynthesis (A) was measured in the presence of ATP(final concentration 1.3 mM). For measurement ofrepair synthesis (0), ATP was omitted, and pan-creatic deoxyribonuclease was added to a final con-centration of 0.3,ug/ml. The incorporation of 8.6 x103 counts/min into DNA represents an incorporationof 1.0 nmole ofATP.
deoxyribonucleoside diphosphate concentra-tion also, instead of increasing, shows a decline.Moreover, a block in the conversion of deox-yribonucleoside diphosphates to deoxyribonu-cleoside triphosphates, as proposed by Rosen-kranz and Jacobs (41), should leave the ribonu-cleotide reductase reaction unaffected and re-sult in an increase in the concentration ofdeoxyribonucleotides relative to ribonucleo-tides, especially since the concentration of HUused in our study allows a fairly high level ofRNA synthesis. At the observed high rate ofRNA synthesis, the rate of utilization of newlysynthesized ribonucleotides should closely ap-proximate the rate of utilization prior to theaddition of HU. The observation that the addi-tion of HU results in net increase in theribonucleotide concentration along with amarked decrease in the total deoxyribonucleo-tide concentration strongly suggests that HUinduces a block at the ribonucleotide reductaselevel. These results are not consistent with adXDP to a dXTP block (where X represents theunknown base). It should be pointed out thatthe conversion of dXDP to dXTP in E. coli iscatalyzed by a nonspecific enzyme(s) that alsocatalyzes the conversion of ribonucleosidediphosphates to ribonucleoside triphosphates(20). Therefore, if HU induced a metabolicblock of the kind proposed by Rosenkranz andJacobs (41), it would be expected to affect bothRNA and DNA synthesis equally. Our resultsindicate that DNA synthesis is much moresensitive to HU than is RNA synthesis.
In spite of the fact that our results clearly
show that the reduction of ribonucleoside di-phosphates is inhibited by HU, if there areother metabolic pathways leading to the syn-thesis of deoxyribonucleotides, then the inhibi-tion of ribonucleoside diphosphate reductasealone might not be sufficient to explain theobserved complete inhibition of DNA synthesisby HU. However, there has been an extensiveamount of work on this aspect of nucleotidemetabolism in E. coli, and no other enzymesystem capable of synthesizing deoxyribonu-cleotides has yet been discovered. The results ofin vivo labeling experiments indicate that all ofthe deoxyribonucleotides are made by reduc-tion of ribonucleotides without cleavage of thebase-sugar bonds, apparently by a reactioncatalyzed by the enzyme ribonucleoside diphos-phate reductase (19).
If it is assumed that ribonucleotide reductaseis really necessary for DNA synthesis, then theinhibition of this enzyme by HU is sufficient toexplain its inhibitory effect on DNA synthesis.However, it would still be very possible that HUmight also be affecting other steps involved inDNA synthesis or, in fact, the DNA templateitself.We have failed to find evidence for the
suggestion that HU inhibits DNA synthesis bycausing a breakdown of the DNA template. Nosingle- or double-stranded breaks were detectedin DNA from cells that had been treated withHU concentrations sufficient to completelyinhibit DNA synthesis. Considerable evidenceindicates that, upon prolonged incubation of E.coli cells with HU, it does induce double-strandbreaks in DNA (17, 42). There is some evidencewhich suggests that during such prolongedincubation HU is slowly converted to related
20O
re) 15-
E 10a-0 5.
a. b.
A
O ,,,,, ]SO, ,,...,0 10 20 300 1o20 30
TIME(MINUTES) TIME(MINUTES)
50
45Qpo
40 E
35
*30
FIG. 6. Pulse-chase experiments. The effect ofhydroxyurea on the ability of E. coli B/S cells toincorporate 14C-labeled deoxyribonucleotides intoDNA after the addition of a 25-fold excess of colduracil. All cells were prelabeled by growth for 25 minin ' 4C-uracil (2 MCi/ml). Time is expressed as minutesafter the addition of cold uracil chase. a, Deox-yribonucleotides; b, DNA. Symbols: 0 and A, colduracil chase alone; 0 and A, cold uracil chase in thepresence of 0.033 M HU.
!
1331VOL. 112, 1972
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

SINHA AND SNUSTAD
compounds such as N-hydroxyurethan and car-bamoyloxyurea, and that these are the realcausative agents of double-strand breaks inDNA. Purified carbamoyloxyurea causes exten-sive breakdown of DNA (43). N-hydroxyure-than, which is lethal to E. coli, also causesextensive degradation of the DNA templateand devitalization of E. coli cells (29). Both ofthese effects are evident during prolonged expo-sure of cells to HU. It seems possible thatdevitalization of cells and DNA breakdown incells exposed to HU for prolonged periods oftime is not actually due to HU but rather to itsderivatives such as carbamoyloxyurea and hy-droxyurethan. The inhibition ofDNA synthesisby HU, in contrast to the induction of breaks inDNA, is evident within a few minutes after theaddition of fairly low concentrations of inhibi-tor to cells.DNA synthesis in toluene-treated cells has
been shown to result in a semiconservativelyreplicated product made at a rate of synthesisclose to the in vivo rate of replication (28). Moreimportantly, DNA synthesis in toluene-treatedcells of E. coli mutants with temperature-sensi-tive blocks in DNA replication is also tempera-ture sensitive (26). In addition, DNA synthesisoccurs normally in toluene-treated cells of apolA mutant of E. coli which lacks the predomi-nant polymerase present in wild-type E. colicells (28). The above observations support theproposal that the DNA synthesis occurring intoluene-treated cells in the presence of ATP isreally true DNA replication. The observationthat HU concentrations as high as 0.13 M haveno detectable effect on ATP-dependent DNAsynthesis in toluene-treated cells indicatesthat HU has no effect(s) on DNA synthesis atthe DNA polymerization level, either by alter-ing the DNA template or by interfering withany of the other factors needed for true replica-tive polymerization.The results of the soluble-pool size studies
show that HU causes a drastic reduction in theconcentrations of deoxyribonucleotides synthe-sized. Moreover, the pulse-chase studies showthat deoxyribonucleotides synthesized prior tothe addition of HU can be utilized for DNAsynthesis at the normal rate in the presence ofHU. These results lead us to the conclusion thatall steps essential for DNA synthesis subse-quent to the reduction of ribonucleotides areunaffected by HU. The observations that DNAsynthesis can occur normally in toluene-treatedcells in the presence of fairly high concentra-tions of HU and that in vivo all steps necessaryfor DNA synthesis beyond the reduction ofribonucleotides proceed normally in the pre-
sence of HU, along with the evidence discussedbelow, suggest that HU inhibits DNA synthesisin E. coli solely by inhibiting ribonucleotidereductase.
In T4-infected E. coli, even though de novosynthesis of deoxyribonucleotides is completelyinhibited by HU, the deoxyribonucleotidesgenerated by the breakdown of the host chro-mosome are quite efficiently reutilized to syn-thesize progeny phage DNA in the presence ofhigh concentrations of HU (45). The averageyield is 14 phage particles per infected cell inthe presence of HU. This synthesis of phageDNA and viral particles cannot be attributedto a limited amount of escape synthesis or tothe presence of some other less efficient mech-anism of deoxyribonucleotide synthesis becauseno phage DNA or viral particles are made in thepresence of HU if bacteriophage T4 mutantsunable to carry out breakdown of the hostgenophore are used (46).
Lactobacillus leichmanii possesses a ribonu-cleotide reductase system quite different fromthe one found in E. coli. The reduction ofribonucleotides takes place at the triphosphatelevel and it is mediated not by an iron-contain-ing component but by a cobalt-containing coen-zyme (4, 5, 15). A similar enzyme systempresumably exists in Rhizobium (9). TheLactobacillus ribonucleotide reductase is notsensitive to HU (12), and these organisms areresistant to HU.A recent study of the metal-binding proper-
ties and antitumor activity of several 1-sub-stituted-3-hydroxyureas by Harmon et al. (16)revealed that only those compounds capable offorming complexes with Fe3+ possessed antitu-mor activity. The correlation between the abil-ity to bind iron and antimicrobial and antitu-mor properties along with the findings of Brownet al. (7), indicating that HU inhibits purifiedribonucleotide reductase of E. coli by interact-ing with the iron-containing component of theprotein B2, also provide indirect evidence sug-gesting that the primary, if not the only, effectof low concentrations of HU on DNA synthesisin E. coli (at least for approximately the first2 hr of exposure) is the result of its inhibition ofribonucleoside-diphosphate reductase.The observation that the addition of deox-
yribonucleosides does not reverse the inhibitioncaused by HU (38) does not exclude the possi-bility that ribonucleotide reductase is the ma-jor, if not the only, site of inhibition of DNAsynthesis in E. coli. The externally addeddeoxyribonucleosides have to be phos-phorylated before they can be used in DNAsynthesis. Even though E. coli cells do possess
1332 J. BACTERIOL.
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

MECHANISM OF INHIBITION OF DNA SYNTHESIS
thymidine kinase activity, it has been foundthat they lack deoxyadenosine, deoxycytidine,and deoxyguanosine kinase activities (18).Some mammalian systems may possess thesekinases, thus explaining the reversal of theeffects of HU by the addition of deoxyribonu-cleosides observed in mammalian tissue cul-tures (2, 3, 14, 24, 48). The fact that addition ofiron to HU-treated purified ribonucleotide re-ductase in vitro does not restore the activity ofthe enzyme (25) may simply mean that theproper conditions for the activation of theenzyme in vitro have not been achieved.
ACKNOWLEDGMENTSThis investigation was supported by Public Health Service
research grant AI-07946 from the National Institute ofAllergy and Infectious Diseases.
LITERATURE CITED1. Adams, M. H. 1959. Bacteriophages, p. 592. Interscience
Publishers, New York.2. Adams, R. L. P., and J. G. Lindsay. 1967. Hydrox) rea.
Reversal of inhibition and use as a cell-synchronizingagent. J. Biol. Chem. 242:1314-1317.
3. Baccheti, S., and G. F. Whitmore. 1969. The action ofhydroxyurea on mouse L-cells. Cell Tissue Kinet.2:193-211.
4. Batterham, T. J., R. K. Ghambeer, R. L. Blakley, and C.Brownson. 1967. Cobamides and ribonucleotide reduc-tion. IV. Stereochemistry of hydrogen transfer to thedeoxyribonucleotide. Biochemistry 6:1203-1208.
5. Blakley, R. L., R. K. Ghambeer, T. J. Batterham, and C.Brownson. 1966. Studies with hydrogen isotopes on themechanism of action of cobamide-dependent ribonu-cleotide reductase. Biochem. Biophys. Res. Commun.24:418-426.
6. Bray, G. A. 1960. A simple efficient liquid scintillator forcounting aqueous solutions in a liquid scintillationcounter. Anal. Biochem. 1:279-285.
7. Brown, N. C., R. Eliasson, P. Reichard, and L. Theland-er. 1969. Spectrum and iron content of protein B2 fromribonucleoside diphosphate reductase. Eur. J. Bio-chem. 9:512-518.
8. Burgi, E., and A. D. Hershey. 1963. Sedimentation rate asa measure of molecular weight of DNA. Biophys. J.3:309-321.
9. Cowles, J. R., H. J. Evans, and S. A. Russell. 1969. B,,coenzyme-dependent ribonucleotide reductase inRhizobium species and the effects of cobalt deficiencyon the activity of the enzyme. J. Bacteriol. 97:1460-1465.
10. Dische, F. 1955. Color reactions of the nucleic acidcomponents, p. 285. In E. Chargaff and J. N. Davidson(ed.), The nucleic acids, vol. 1, Academic Press Inc.,New York.
11. Edgar, R. S. 1963. Some technical considerations con-
cerning experiments on phage recombination, p. 19-36.In W. J. Burdette (ed.), Methodology in basic genetics.Holden-Day, San Francisco.
12. Elford, H. L. 1968. Effect of hydroxyurea on ribonucleo-tide reductase. Biochem. Biophys. Res. Commun.33:129-135.
13. Gale, G. R. 1968. Antagonism by deoxyribosides of theinhibitory action of certain hydroxamic acids on DNAsynthesis. Experientia 24:57-58.
14. Gale, G. R., S. M. Kendall, H. H. McLain, and S.
DuBois. 1964. Effect of hydroxyurea on Pseudomonasaeruginosa. Cancer Res. 24:1012-1016.
15. Gottesman, M. M., and W. S. Beck. 1966. Transfer ofhydrogen in the cobamide-dependent ribonucleotidereductase reaction. Biochem. Biophys. Res. Commun.24:353-359.
16. Harmon, R. E., J. C. Dabrowiak, D. J. Brown, S. K.Gupta, M. Herbert, and D. Chitharanjan. 1970. Metal-complexes of 1-substituted 3-hydroxyureas. J. Med.Chem. 13:577-579.
17. Jacobs, S. J., and H. S. Rosenkranz. 1970. Detection of areactive intermediate in the reaction between DNAand hydroxyurea. Cancer Res. 30:1084-1094.
18. Karlstrom, H. 0. 1970. Inability of Escherichia coli B toincorporate added deoxycytidine, deoxyadenosine, anddeoxyguanosine into DNA. Eur. J. Biochem. 17:68-71.
19. Karlstrom, O., and A. Larsson. 1967. Significance ofribonucleotide reduction in the biosynthesis of deox-yribonucleotides in Escherichia coli. Eur. J. Biochem.3:164-170.
20. Koerner, J. F. 1970. Enzymes of nucleic acid metabolism.Annu. Rev. Biochem. 39:291-322.
21. Krakoff, I. H., N. C. Brown, and P. Reichard. 1968.Inhibition of ribonucleoside diphosphate reductase byhydroxyurea. Cancer Res. 28:1559-1565.
22. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with a Folinphenol reagent. J. Biol. Chem. 193:265.
23. Martin, R. G., and B. N. Ames. 1961. A method fordetermining the sedimentation behavior of enzymes:application to protein mixtures. J. Biol. Chem.236:1372-1379.
24. Mohler, W. C. 1964. Cytotoxicity of hydroxyurea(NSC-32065) reversible by pyrimidine deoxyribosidesin a mammalian cell line grown in vitro. CancerChemother. Rep. 34:1-6.
25. Moore, E. C. 1969. The effects of ferrous ion anddithioerythritol on inhibition by hydroxyurea of ribo-nucleotide reductase. Cancer Res. 29:291-295.
26. Mordoh, J., Y. Hirota, and F. Jacob. 1970. On the processof cellular division in Escherichia coli. V. Incorporationof deoxyribonucleoside triphosphates by DNA-ther-mosensitive mutants of Escherichia coli also lackingDNA polymerase activity. Proc. Nat. Acad. Sci. U.S.A.67:773-778.
27. Moses, R. E., and C. C. Richardson. 1970. A new DNApolymerase activity of Escherichia coli. I. Purificationand properties of the activities present in E. coli polAl.Biochem. Biophys. Res. Commun. 41:1557-156-i.
28. Moses, R. E., and C. C. Richardson. 1970. Replicationand repair of DNA in cells of Escherichia coli treatedwith toluene. 1970. Proc. Nat. Acad. Sci. U.S.A.67:674-681.
29. Mullinix, K. P., and H. S. Rosenkranz. 1971. Effect ofN-hydroxyurethan on viability and metabolism ofEscherichia coli. J. Bacteriol. 105:556-564.
30. Neuhard, J. 1966. Studies on the acid-soluble nucleotidepool in thymine-requiring mutants of Escherichia coliduring thymine starvation. III. On the regulation of thedeoxyadenosine triphosphate and deoxycytidine tri-phosphate pools of Escherichia coli. Biochim. Biophys.Acta 129:104-115.
31. Neuhard, J. 1967. Studies on the acid-soluble nucleotidepool in Escherichia coli. IV. Effects of hydroxyurea.Biochim. Biophys. Acta 145:1-6.
32. Neuhard, J., E. Randerath, and K. Randerath. 1965.Ion-exchange thin layer chromatography. XIII. Resolu-tion of complex nucleoside triphosphate mixtures.Anal. Biochem. 13:211-222.
33. Neuhard, J., and E. Thomassen. 1971. Turnover of thedeoxynucleoside triphosphates in Escherichia coli 15Tduring thymine starvation. Eur. J. Biochem. 20:36-43.
VOL. 112, 1972 1333
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

SINHA AND SNUSTAD
34. Randerath, E., and K. Randerath. 1964. Resolution ofcomplex nucleotide mixtures by two-dimensional an-
ion exchange thin-layer chromatography. J. Chro-matogr. 16:126-129.
35. Randerath, K., and E. Randerath. 1965. Ion-exchangethin-layer chromatography. XIV. Separation of nucleo-tide sugars and nucleoside monophosphates on PEI-cellulose. Anal. Biochem. 13:575-579.
36. Reichard, P. 1968. The biosynthesis of deoxyribose, p. 77.John Wiley & Sons, New York.
37. Rosenkranz, H. S., and H. S. Carr. 1970. Hydroxyureaand Escherichia coli nucleoside diphosphate reductase.Cancer Res. 30:1926-1927.
38. Rosenkranz, H. S., H. S. Carr, and R. D. Pollak. 1967.Studies with hydroxyurea. VI. Effects of hydroxyureaon the metabolism of sensitive and resistant strains ofEscherichia coli. Biochim. Biophys. Acta 149:228-245.
39. Rosenthal, P. N., and M. S. Fox. 1970. Effects ofdisintegration of incorporated 3H and 32p on thephysical and biological properties of DNA. J. Mol.Biol. 54:441-463.
40. Rosenkranz, H. S., A. J. Garro, J. A. Levy, and H. S.Carr. 1966. Studies with hydroxyurea. I. The reversibleinhibition of bacterial DNA synthesis and the effect ofhydroxyurea on the bactericidal action of strep-tomycin. Biochim. Biophys. Acta 114:501-515.
41. Rosenkranz, H. S., and S. J. Jacobs. 1968. Inhibition ofDNA synthesis by hydroxyurea. Biol. Biochem. Eval.
Malignancy Exp. Hepatomas, Proc. U.S.-Jap. Conf.6:15-41.
42. Rosenkranz, H. S., S. J. Jacobs, and H. S. Carr. 1968.Studies with hydroxyurea. VIII. The deoxyribonucleicacid of hydroxyurea-treated cells. Biochim. Biophys.Acta 161:428-441.
43. Rosenkranz, H. S., and S. Rosenkranz. 1969. Degradationof DNA by carbamoyloxyurea-an oxidation productof hydroxyurea. Biochim. Biophys. Acta 195:266-267.
44. Short, E. C., H. R. Wamer, and J. F. Koemer. 1968. Theeffect of cupric ions on the indole reaction for thedetermination of deoxyribonucleic acid. J. Biol. Chem.243:3342-3344.
45. Wamer, H. R., and M. D. Hobbs. 1969. Effect ofhydroxyurea on replication of bacteriophage T4 inEscherichia coli. J. Virol. 3:331-336.
46. Warner, H. R., D. P. Snustad, S. E. Jorgensen, and J. F.Koemer. 1970. Isolation of bacteriophage T4 mutantsdefective in the ability to degrade host deoxyribonu-cleic acid. J. Virol. 5:700-708.
47. Yeh, Y. C., E. J. Dubovi, and I. Tessman. 1969. Control ofpyrimidine biosynthesis by phage T4: mutants unableto catalyze the reduction of cytidine diphosphate.Virology 37:615-623.
48. Young, C. W., G. Schochetman, S. Hodas, and M. E.Balis. 1967. Inhibition of DNA synthesis by hydrox-yurea: structure activity relationships. Cancer Res.27:535-540.
1334 J. BACTERIOL.
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from






![Furine and Pyrimidine Ribonucleoside Monophosphate ...cancerres.aacrjournals.org/content/canres/42/4/1326.full.pdf · [CANCER RESEARCH 42, 1326-1330, April1982] 0008-5472/82/0042-0000$02.00](https://img.pdfslide.net/doc/110x75/5a7e72ec7f8b9ae9398e75ca/furine-and-pyrimidine-ribonucleoside-monophosphate-cancer-research-42-1326-1330.jpg)






















