Embed Size (px)
Citation preview

Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520
Leukotriene C4 synthase: a pivotal enzyme incellular biosynthesis of the cysteinyl leukotrienes
Bing K. Lam∗, K. Frank AustenDepartment of Medicine, Division of Rheumatology, Immunology and Allergy, Brigham and Women’s Hospital,
Harvard Medical School, Boston, MA 02115, USA
Abstract
Leukotriene C4 synthase (LTC4S) conjugates LTA4 with glutathione (GSH) to form LTC4, theparent compound of the cysteinyl LTs. LTC4S is an 18 kDa membrane protein and functions as anoncovalent homodimer. The enzyme activity of LTC4S is augmented by Mg2+ and inhibited byCo2+ and the function of 5-lipoxygenase (LO) activating protein (FLAP) inhibitor MK-886. TheKm andVmax values are 3.6�M and 1.3�mol/mg/min for LTA4 and 1.6 mM and 2.7�mol/mg/minfor GSH, respectively. The deduced amino acid sequence and the predicted secondary structureof LTC4S shares significant homology to FLAP, mGST-2 and mGST-3 which are all membersof MAPEG protein superfamily. LTC4S and FLAP exhibited identical genomic organization offive exons and four introns. Site-directed mutagenesis suggests that Arg-51 is involved in openingthe epoxide ring of LTA4 and Tyr-93 in GSH thiolate anion formation during catalytic conjuga-tion. LTC4S is a TATA-less gene whose transcription assessed in a reporter construct involvedboth cell-specific and nonspecific regulatory elements. LTC4S−/− mice grow normally, and areattenuated for innate and adaptive immune inflammatory permeability responses.© 2002 Elsevier Science Inc. All rights reserved.
Keywords: Leukotriene C4; Conjugation; Mutagenesis; Inflammation
1. Introduction
Bronchial asthma is characterized by the presence of episodic bronchoconstriction, per-sistent hyper-responsiveness, and inflammation of the conducting airways[1,2]. The in-flammatory responses in asthma include luminal plugging with mucus; denudation of theepithelium[3] with subepithelial collagen deposition[4]; infiltration of the epithelium andsubmucosa with eosinophils and T helper 2 cells[5,6]; and degranulation of resident mastcells and infiltrating eosinophils[5]. Mast cells and eosinophils release both preformed
∗ Corresponding author. Tel.:+1-617-525-1270; fax:+1-617-525-1310.E-mail address: [email protected] (B.K. Lam).
0090-6980/02/$ – see front matter © 2002 Elsevier Science Inc. All rights reserved.PII: S0090-6980(02)00052-7

512 B.K. Lam, K. Frank Austen / Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520
granule proteins such as mast cell tryptase and eosinophil major basic protein, respectively,and newly generated lipid mediators such as cysteinyl leukotrienes (LTs) which contributeto the pathologic changes in asthma[1].
The specific involvement of the cysteinyl LTs in asthma is suggested by their potentbiological activities, their presence in the bronchial alveolar lavage fluid of asthmatic pa-tients and the clinical efficacy of 5-lipoxygenase (5-LO) inhibitors and cysteinyl LT receptorantagonists in the management of patients with bronchial asthma. In addition to compro-mising airway function when administered by inhalation to normal individuals, LTC4 andits metabolites, LTD4 and LTE4, are each even more potent in reducing the lung functionof the asthmatic patients[7]. These lipid mediators also stimulate mucus secretion frombronchial epithelial cells and increase pulmonary vascular permeability via endothelial cellcontraction at the post-capillary venules[8] and have been recovered from the airways ofasthmatic individuals at rest[9] and in increased amounts after allergen challenge[10],isocapnic hyperventilation[11], or aspirin challenge of susceptible patients[12]. Clinicalstudies with leukotriene biosynthesis inhibitors or with cysteinyl LT receptor antagonists,agents which are devoid of intrinsic bronchodilatory activity, leads to rather immediatebronchodilatation in patients with asthma[13]. These results suggest that chronic over-production of the cysteinyl leukotrienes may contribute to the abnormal resting airwaytone in some patients with asthma. This view is supported by the capacity of these agentsto inhibit the acute asthmatic responses to exercise[14], cold–dry air[15], allergen[16],and aspirin[17] and by their efficacy in the management of patients with chronic asthma[18].
2. Biosynthesis of cysteinyl leukotrienes
Cellular biosynthesis of cysteinyl leukotrienes in mast cells, basophils, monocyte/macrophages, and eosinophils is initiated by an increase in intracellular Ca2+ and acti-vation of phospholipase A2 (PLA2) in response to agonist stimulation. Cytosolic PLA2translocates to the perinuclear membrane and releases arachidonic acid from the mem-brane phospholipids[19]. 5-LO, which translocates from either cytoplasm or nucleoplasm[20] to the perinuclear membrane and is functionally modulated by tyrosine kinase[21],converts the released arachidonic acid to 5-HPETE and subsequently to LTA4 [22]. The in-tracellular activity of 5-LO on released arachidonic acid requires the associated function of5-LO activating protein (FLAP), an integral perinuclear membrane protein. LTC4 synthaseconjugates LTA4 with glutathione (GSH) to form LTC4 [23]. LTC4 synthase is the onlyenzyme in hematopoietic cells with the ability to conjugate GSH with LTA4 to form LTC4.Following carrier-mediated export[24], sequential cleavage of glutamic acid and glycinefrom the GSH moiety of LTC4 yields LTD4 and LTE4, respectively[25]. LTC4, LTD4 andLTE4 subsequently bind to the G-protein coupled CysLT1 or CysLT2 receptors to exert theirbiologic effects[26,27]. Whereas the CysLT1 receptor has higher affinity for LTD4 than forLTC4 or LTE4, the CysLT2 receptor has similar high affinity toward LTC4 and LTD4 andlower affinity for LTE4. In addition to cysteinyl leukotrienes, recombinant murine CysLT1receptor has also been shown to respond to UDP but not to other pyrimidinergic or puriner-gic nucleotides[28].

B.K. Lam, K. Frank Austen / Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520 513
3. LTC4 synthase protein
LTC4 synthase differs from conventional GSH S-transferases (GST), by its selectivityfor LTA4 and its analogs[23], microsomal localization, and failure to conjugate xenobi-otics. LTC4 synthase also exhibits differential susceptibility to inhibitors[29] and lacksimmunoreactivity to antibodies for known GSTs[30]. The enzyme is expressed in a limitednumber of cell types such as eosinophils, mast cells, basophils, monocyte/macrophages andin leukemic cell lines such as KG-1 cells and THP-1 cells; it is also present in platelets,which lack 5-LO.
LTC4 synthase was purified to homogeneity as an 18 kDa protein from THP-1[31] andKG-1 cells. The 18 kDa protein was proposed to function as a homodimer based on thesize of the active fraction by gel filtration column chromatography of both the native andthe recombinant protein[31,32]. LTC4 synthase activity is augmented by Mg2+ ions, andis inhibited by Co2+ ion [30] and by the FLAP inhibitor MK-886[33]. N-ethylmaleimide,a sulfhydryl modifying reagent that activates microsomal (m)GST-I, inhibits recombinantLTC4 synthase and also LTC4 synthase from guinea pig lung or from U-937 cells[23,30].
Purified human recombinant LTC4 synthase displayedKm values for LTA4 and GSH of3.6 and 1.6 mM, respectively, andVmax values of 1.3 and 2.7�mol/min/mg, respectively[34]. These values are similar to the values obtained with the LTC4 synthase purified fromDMSO-differentiated U-937 cells and to the purified recombinant mouse protein[30,34].
4. LTC4 synthase cDNA
Human LTC4 synthase cDNA encodes for a 150 amino acid residue polypeptide[33].The deduced amino acid sequence of LTC4 synthase reveals two potential protein kinase C(PKC) phosphorylation sites and one potentialN-glycosylation site. The deduced amino acidsequence reveals 31% overall amino acid identity with FLAP. The identity at the N-terminal2/3 of these proteins is 44% and includes the putative FLAP-like inhibitor binding domain.Furthermore, secondary structure analysis of the deduced LTC4 synthase indicated that it isnearly identical to FLAP, as their three respective hydrophobic domains overlap each otherand their two hydrophilic loops are similar in sizes. These features of LTC4 synthase areconsistent with reports of down regulation of this enzyme by PKC activation and the abilityof the FLAP inhibitor, MK-886, to inhibit LTC4 synthase activity[33–35]. The degree ofamino acid identity and the similarity of the predicted secondary structure between LTC4synthase and FLAP are also observed in mGST-2 and mGST-3[36]. The utilization of eitherarachidonic acid and/or GSH as their enzymatic substrates together with the similarity inthe predicted secondary structure for these integral membrane proteins have establishedthem as members of a new protein family, the membrane associated protein involved ineicosanoid and GSH metabolism or MEPEG[36].
Mouse LTC4 synthase also encodes for a 150 amino acid residue polypeptide and thededuced amino acid sequence displays an 88% amino acid identity to human LTC4 synthase.The putative FLAP-like inhibitor binding domain, the potentialN-glycosylation site, and thetwo potential PKC phosphorylation sites are all conserved in mouse LTC4 synthase. Thereare 18 different amino acid residues in mouse LTC4 synthase, nine of which are located at

514 B.K. Lam, K. Frank Austen / Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520
the carboxy-terminus of the protein. These differences in amino acid residues do not appearto alter mouse LTC4 synthase function as evidenced by the near identical enzyme kineticsand susceptibility to inhibition by MK-886 as compared to the human enzyme[33,34].
5. LTC4 synthase gene
The human LTC4 synthase gene comprises of five small exons and four introns and spans2.5 kbp[37]. The human gene for FLAP is >31 kbp in size. Nonetheless, the second, third,and fourth exons of LTC4 synthase are identical in size to those of the FLAP gene, while thefirst and the fifth exons differ minimally by the number of nucleotides in the 5′ and the 3′untranslated regions. Hence, the intron/exon junctions and gene organization are the samefor FLAP and LTC4 synthase. In addition, the exons of LTC4 synthase and FLAP alignidentically with regard to the amino acids that they encode.
The 5′ extension analysis of KG-1 mRNA revealed three putative transcription initiationsites in the human LTC4 synthase gene, located 66, 69, and 96 nucleotides upstream of theATG translation start site. The 5′ flanking region of human LTC4 synthase gene containsthe typical features of genes that have been identified with multiple transcription initiationsites: a high G/C content and at least one consensus sequence for Sp1 binding 18 nucleotidesupstream of the first transcription start site.
The 2.0 kb gene for mouse LTC4 synthase has an intron/exon organization identicalto the human LTC4 synthase and FLAP genes. The 5′ flanking sequence of the mouseLTC4 synthase gene, however, contains only one transcription initiation site 64 bp upstreamof the ATG start site by primer extension with lung RNA, and consensus transcriptionfactor binding motifs for AP-2, CEBP, PEA-3[38] and STAT-6 (Lam et al., unpublishedobservation).
Fluorescent in situ hybridization with the P1 plasmid containing the gene for LTC4 syn-thase localized this gene to the long arm of human chromosome 5, an area that correspondsto band 5q35[37]. The LTC4 synthase gene is located distal to the gene cluster for cytokinescentral to the Th2 cell phenotype and to genes implicated in allergic inflammation[37,39];this chromosomal organization is mimicked in mouse by a syntenic location of the LTC4synthase gene on mouse chromosome 11[38].
6. Transcriptional regulation of human LTC4 synthase
Promoter analysis of the 2.1 kb 5′ flanking sequence of the human LTC4 synthase generevealed the ability of the proximal 225-bp genomic fragment (nucleotides−228 to−3upstream of the translation start site) to induce the expression of a reporter gene product inLTC4S expressing THP-1 cells[40] but not in the nonexpressing K-562 cells. Deletionaland mutational analysis of the 225 bp fragment showed the presence of a nonspecific basalpromoter region between nucleotides−122 and−56 upstream of the ATG translation startsite and a cell specific region further upstream. The nonspecific region contains a Sp1 bindingmotif (GC box,−120 to−115) and an initiator element (Inr, CAGAC−66 to−62). ThisSp1 site binds Sp1 and Sp3 and is critical for transcription of the reporter gene. Mutation

B.K. Lam, K. Frank Austen / Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520 515
of a tandem CACCC motif (nucleotides−149 to−145 and−139 to−135) of the cellspecific region reduced the expression of the reporter gene. Mutation of the CACCC motifalso reduced the ability of both Sp1 and Kruppel-like factor Zf9/CPBP totrans-activatethe reporter gene implying that these transcription factors are involved in the cell-specificregulation of LTC4 synthase gene[40]. An A–C polymorphism at nucleotide−444 wasreported to be higher in Polish patients with aspirin-intolerant asthma but was not observedto be associated with aspirin-intolerant asthmatics in the USA[41,42].
Although not analyzed by reporter gene assay, consensus PKC-responsive elements forAP-1 and AP-2 at 807 and 877 nucleotides 5′ of the first transcription initiation site mayalso be involved in the transcription regulation of LTC4 synthase gene as the induction ofits enzyme activity has been reported in human erythroleukemia cells after treatment withphorbol 12-myristate 13-actate (PMA)[43]. In addition, upregulation of LTC4 synthasehas been reported in THP-1 cells treated with TGF� [44] and in in vitro derived humaneosinophils developed with IL-3 and IL-5[45]. It is possible that the regulation of LTC4synthase by IL-3 involves PI-3 kinase and the activation of NFAT transcription factor asthere is a putative NFAT binding motif located 648 nucleotides upstream of the translationstart site (Lam et al., unpublished observation).
Human mast cells derived in culture from cord blood mononuclear cells with stem cellfactor (SCF), IL-6 and IL-10, washed, and sustained with cytoprotective SCF reveal aremarkable induction of LTC4 synthase by IL-4. The increased expression of LTC4 synthaseis apparent for the transcript by Northern analysis, the protein by Western analysis, and forsubcellular function by conjugation of LTA4-methyl ester and GSH to form LTC4-methylester[46]. The integrated response to cross linking of the membrane receptor for IgE,FcεR1, is also increased by more than 20-fold compared to cells maintained in SCF alone.A signal transducer activator of transcription (STAT)-6 binding motif has been identified1.9 kb upstream of the ATG start site.
7. Catalytic mechanism of LTC4 synthase
Site-directed mutagenesis was carried out to examine the substrate binding sites and thecatalytic mechanisms for conjugation of LTA4 and GSH by human LTC4 synthase[32].Mutations of V49F, A52S, N55A and Y59F of the first hydrophilic loop, and mutations ofY97F and Y93F of the second hydrophilic loop, increased theKm of the recombinant enzymefor GSH, suggesting that these residues are involved in GSH binding. Point mutation ofthe Arg-51 of the first hydrophilic loop to Thr or Ile abolished function of the recombinantmutant protein, whereas mutations of Arg-51 to His or Lys provided fully active mutantprotein. The mutation of Y93F reduced the enzyme activity of the mutant protein to lessthan 1% of the wild type enzyme. In addition, there is a shift in pH optimum of the residualactivity of the mutant protein to that of spontaneous conjugation. Mutations of C56S andC82V, each alone or together, did not alter enzyme function.N-Ethylmaleimide treatmentinhibits C82V but not C56S enzyme (Lam et al., unpublished observation). These resultssuggest that LTC4 synthase is a noncovalent homodimer and thatN-ethylmaleimide inhibitswild type enzyme by modifying the free sulfhydryl group of Cys-56. Direct linkage of twoLTC4 synthase monomers by a 12 amino acid bridge gave rise to an active covalent-linked
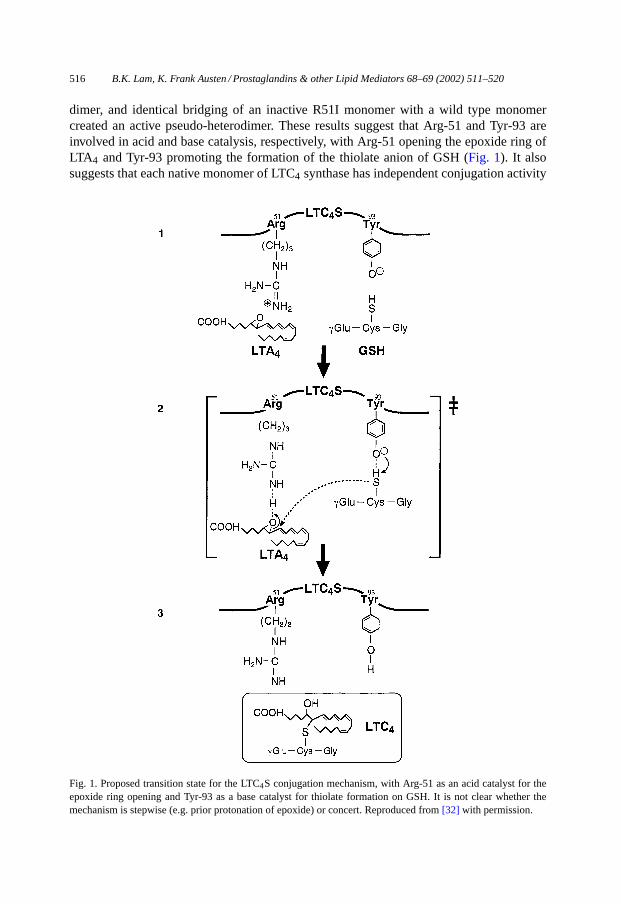
516 B.K. Lam, K. Frank Austen / Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520
dimer, and identical bridging of an inactive R51I monomer with a wild type monomercreated an active pseudo-heterodimer. These results suggest that Arg-51 and Tyr-93 areinvolved in acid and base catalysis, respectively, with Arg-51 opening the epoxide ring ofLTA4 and Tyr-93 promoting the formation of the thiolate anion of GSH (Fig. 1). It alsosuggests that each native monomer of LTC4 synthase has independent conjugation activity
Fig. 1. Proposed transition state for the LTC4S conjugation mechanism, with Arg-51 as an acid catalyst for theepoxide ring opening and Tyr-93 as a base catalyst for thiolate formation on GSH. It is not clear whether themechanism is stepwise (e.g. prior protonation of epoxide) or concert. Reproduced from[32] with permission.

B.K. Lam, K. Frank Austen / Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520 517
and that noncovalent dimerization of LTC4 synthase maintains the proper protein folding.It is interesting that the amino acid residue Tyr-93 is present in mGST-2, mGST-3, andFLAP. In contrast, Arg-51 is present in mGST-2 and mGST-3, which have the ability toconjugate both LTA4 and xenobiotics with GSH, but is not present in FLAP, which has noknown GSH-conjugating activity (32).
8. LTC4 synthase gene disrupted mice
The murine LTC4 synthase gene was disrupted by gene targeting technique such thatexons II–IV were deleted and replaced by a neomycin gene. Exons II–IV encode aminoacid residues 21–104 of the polypeptide. This region includes Arg-51 and Tyr-93 and othersubstrate binding residues that are critical for enzyme function. The targeted gene disrup-tion did not affect the growth and the fertility of the animals[47]. Disruption of the murineLTC4 synthase gene abrogated the ability of the strain-derived mast cells, obtained by bonemarrow culture with IL-3, to express LTC4 synthase transcript, protein and function. Dis-ruption of the LTC4 synthase gene greatly reduced or eliminated the capacity of varioustissues to conjugate LTA4-methyl ester with GSH to form LTC4-methyl ester establishingLTC4 synthase as the dominant enzyme in this proinflammatory pathway. Only the testisrevealed normal LTA4-methyl ester conjugating activity, likely reflecting the activities ofmGST-2, mGST-3 and/or other GSTs. Bone marrow-derived mast cells from LTC4S−/−animals did not synthesize LTC4 in response to IgE-dependent activation but generated nor-mal amount of PGD2 and LTB4 and exhibited normal degranulation responses (Table 1).There was an increase in 6-trans-LTB4 diastereoisomers but not 5-hydroxyeicosatetraenoicacid indicating decay of unused LTA4 rather than shunting to the LTA4 hydrolase pathway.There was a 50% reduction in IgE mediated passive cutaneous anaphylaxis of the ear in theLTC4S−/− animals revealing a prominent role of the cysteinyl leukotrienes in augmentingmast cell dependent permeability[47]. There was also a 50% suppression in the incrementin intraperitoneal permeability elicited by the injection of zymosan through its interactionwith the monocyte/macrophage�-glucan receptor. These observations indicate that LTC4synthase, not mGST-2 or mGST-3, is the major LTC4-producing enzyme in tissues whosefunction includes mediation of the increase in vascular permeability in innate or adaptiveimmune responses.
Table 1Release of�-HEX and production of eicosanoids by BMMC from LTC4S−/− and LTC4S+/+ mice BMMC weresensitized with 2�g/ml of mouse monoclonal IgE and stimulated with 10�g/ml of goat anti-mouse Ig
Genotype Production (pmol/106 cells) �-HEX (%)
LTC4 6-trans-LTB4a LTB4 5-HETE PGD2
LTC4S−/− <1.0 108.7± 8.3b 29.5± 3.2 87.3± 8.0 11.1± 1.7 33.3± 4.2LTC4S+/+ 100.2± 14.3 54.3± 5.9 28.1± 4.5 94.9± 9.9 8.3± 1.9 25.0± 4.6
The supernatants were analyzed by RP-HPLC for leukotrienes and by EIA for PGD2. Values are means± S.E.
(n = 3). Modified from[47].a Includes its diastereoisomers.b P < 0.01 as compared to LTC4S+/+.

518 B.K. Lam, K. Frank Austen / Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520
Acknowledgements
This work was supported in part by National Institutes of Health Grants AI22531,AI31599, AR36308, ES06105, HL03208 and HL36110.
References
[1] Busse WW, Calhun WF, Sedgwick JD. Mechanism of airway inflammation in asthma. Am Rev Respir1993;Dis.147:S20–4.
[2] Dunnill MS. The pathology of asthma, with special reference to changes in the bronchial mucosa. J Clin Path1960;13:27–33.
[3] Laitinen LA, Heino M, Laitinen A, Kava T, Haahtela T. Damage of the airway epithelium and bronchialreactivity in patients with asthma. Am Rev Respir Dis 1985;131:599–606.
[4] Roche WR, Beasley R, Williams JH, Holgate ST. Subepithelial fibrosis in the bronchi of asthmatics. Lancet1989;1:520–4.
[5] Beasley R, Roche WR, Roberts JA, Holgate ST. Cellular events in the bronchi in mild asthma and afterbronchial provocation. Am Rev Respir Dis 1989;139:806–17.
[6] Robinson DS, Hamid Q, Ying S, Tsicopoulos A, Barkan J, Bentley AM, et al. Predominant Th2-likebrochoalveolar T-lymphocyte population in atopic asthma. N Engl J Med 1992;326:298–304.
[7] Arm JP, O’Hickey SP, Hawksworth RJ, Fong CY, Crea AEG, Spur B, et al. Asthmatic airways have adisproportionate hyperresponsiveness to LTE4, as compared with normal airways, but not to LTC4, LTD4,methacholine, and histamine. Am Rev Respir Dis 1990;142:1112–8.
[8] Drazen JM, Austen KF. Leukotrienes and airway responses. Am Rev Respir Dis 1987;136:985–98.[9] Lam S, Chan H, LeRiche JC, Chan-Yeung M, Salari H. Release of leukotrienes in patients with bronchial
asthma. J Allergy Clin Immunol 1988;81:711–7.[10] Wenzel SE, Larsen GL, Johnston K, Voelkel NF, Westcott JY. Elevated levels of leukotriene C4 in
bronchoalveolar lavage fluid from atopic asthmatics after endobronchial allergen challenge. Am Rev RespirDis 1990;142:112–9.
[11] Pliss LB, Ingenito EP, Ingram RH, Pichurko B. Assessment of broncho-alveolar cell and mediator responseto isocapnic hyperpnea in asthma. Am Rev Respir Dis 1990;142:73–8.
[12] Christie PE, Tagari P, Ford-Hutchinson W, Charlesson S, Chee P, Arm JP, et al. Urinary leukotriene E4
concentrations increase after aspirin challenge in aspirin-sensitive asthmatic subjects. Am Rev Respir Dis1991;143:1025–9.
[13] Gaddy J, Margolskee D, Bush RK, Williams VC, Busse W. Broncho-dilatation with a potent and selectiveleukotriene D4 (LTD4) receptor antagonist (MK-571) in a patient with asthma. Am Rev Respir Dis1992;146:358–63.
[14] Manning PJ, Watson RM, Margolskee DJ, Williams VC, Schwartz JI, O’Byrne PM. Inhibition ofexercise-induced bronchoconstriction by MK-571, a patent leukotriene D4-receptor agonist. N Engl J Med1990;323:1736–9.
[15] Israel E, Dermarkarian R, Rosenberg M, Sperling R, Taylor G, Rubin P, et al. The effects of 5-lipoxygenaseinhibitor. N Engl J Med 1990;323:1740–4.
[16] Taylor IK, O’Shaughnessy KM, Fuller RW, Dollery CT. Effect of cysteinyl-leukotriene receptor antagonistICI 204,219 on allergen induced bronchoconstriction and airway hyperreactivity in atopic subjects. Lancet1991;337:690–4.
[17] Israel E, Fischer AR, Rosenberg MA, Lilly CM, Callery JC, Shapiro J, et al. The pivotal role of 5-lipoxygenaseproducts in the reaction of aspirin-induced asthma. Am Rev Respir Dis 1993;148:1447–51.
[18] Israel E, Rubin P, Kemp JP, Grossman J, Pierson W, Siegal SC, et al. The effect of inhibition of 5-lipoxygenaseby zileuton in mild-to-moderate asthma. Ann Int Med 1993;119:1059–66.
[19] Clark JD, Milona N, Knopf JL. Purification of a 110-kDa cytosolic phospholipase A2 from human monocyticcell line U937. Proc Natl Acad Sci USA 1990;87:7708–12.
[20] Brock TG, McNish RW, Bailie MB, Peters-Golden M. Rapid import of cytosolic 5-lipoxygenase into thenucleus of neutrophils after in vivo recruitment and in vitro adherence. J Biol Chem 1997;272:8276–80.

B.K. Lam, K. Frank Austen / Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520 519
[21] Lepley RA, Muskardin DT, Fitzpatrick FA. Tyrosine kinase activity modulates catalysis and translocation ofcellular 5-lipoxygenase. J Biol Chem 1996;271:6179–84.
[22] Rouzer CA, Matsumoto T, Samuelsson B. Single protein from human leukocytes possesses 5-lipoxygenaseand leukotriene A4 synthase activities. Proc Natl Acad Sci USA 1986;83:857–61.
[23] Yoshimoto T, Soberman RJ, Spur B, Austen KF. Properties of highly purified leukotriene C4 synthase ofguinea pig lung. J Clin Invest 1988;81:866–71.
[24] Lam BK, Owen Jr WF, Austen KF, Soberman RJ. The identification of a distinct export step following thebiosynthsis of leukotriene C4 by human eosinophils. J Biol Chem 1989;264:12885–9.
[25] Orning L, Kaijsen L, Hammarstrom S. In vivo metabolism of leukotriene C4 in man: urinary excretion ofleukotriene E4. Biochem Biophys Res Commun 1985;130:214–20.
[26] Lynch KR, O’neal GP, Liu Q, Im DS, Sawyer N, Metters KM, et al. Characterization of the human cysteinylleukotriene CysLT1 receptor. Nature 1999;399:789–93.
[27] Heise CE, O’Dowd BF, Figueroa DJ, Sawyer N, Nguyen T, Im DS, et al. Characterization of the humancysteinyl leukotriene 2 receptor. J Biol Chem 2000;275:30531–6.
[28] Mellor EA, Maekawa A, Austen KF, Boyce JA. Cysteinyl leukotriene receptor 1 is also a pyrimidinergicreceptor and is expressed by human mast cells. Proc Natl Acad Sci USA 2001;98:7964–9.
[29] Bach MK, Brashler JR, Peck RE, Morton Jr DR. Leukotriene C synthetase, a special glutathione S-transferase:properties of the enzyme and inhibitor studies with special reference to the mode of action of U-60257, aselective inhibitor of leukotriene synthesis. J Allergy Clin Immunol 1984;74:353–7.
[30] Nicholson DW, Klemba MW, Rasper DM, Metters KM, Zamboni RJ, Ford-Hutchinson AW. Purificationof human leukotriene C4 synthase from dimethylsulfoxide-differentiated U-937 cells. Eur J Biochem1992;209:725–34.
[31] Nicholson DW, Ali A, Vallancourt JP, Calaycay J, Mumford R, Zamboni R, et al. Purification to homogeneityand the N-terminal sequence of human leukotriene C4 synthase: a glutathione S-transferase composed of18-kDa subunits. Proc Natl Acad Sci USA 1993;90:2015–9.
[32] Lam BK, Penrose JF, Xu K, Baldasaro MH, Austen KF. Site-directed mutagenesis of human leukotriene C4
synthase. J Biol Chem 1997;272:13923–8.[33] Lam BK, Penrose JF, Freeman GJ, Austen KF. Expression cloning of a cDNA for human leukotriene C4
synthase, an integral membrane protein conjugating reduced glutathione to leukotriene A4. Proc Natl AcadSci USA 1994;91:7663–7.
[34] Lam BK, Penrose JF, Rokach J, Xu K, Baldasaro M, Austen KF. Molecular cloning, expression, andcharacterization of mouse leukotriene C4 synthase. Eur J Biochem 1996;238:606–12.
[35] Sjolinder M, Tornhamre S, Werga P, Edenius C, Lindgren JA. Phorbol ester-induced suppression of leukotrieneC4 synthase activity in human granulocytes. FEBS Lett 1995;377:87–91.
[36] Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson AW, Persson B. Common structural featuresof MAPEG: a widespread superfamily of membrane associated proteins with highly divergent functions ineicosanoid . Science 1999;8:689–92.
[37] Penrose JF, Spector J, Baldasaro M, Xu K, Boyce J, Arm JP. Molecular cloning of the gene for humanleukotriene C4 synthase: organization, nucleotide sequence, and chromosomal localization to 5q35. J BiolChem 1996;271:11356–61.
[38] Penrose JF, Baldasaro MH, Webster M, Xu K, Austen KF, Lam BK. Molecular cloning of the gene for mouseleukotriene C4 synthase. Eur J Biochem 1997;248:807–13.
[39] van Leeuwen BH, Martinson ME, Webb GC, Young IG. Molecular organization of the cytokine gene cluster,involving the human IL-3, IL-4, IL-5, and GM-CSF genes, on human chromosome 5. Blood 1989;73:1142–8.
[40] Zhao JL, Austen KF, Lam BK. Cell-specific transcription of leukotriene C4 synthase involves a Kruppel-liketranscription factor and Sp1. J Biol Chem 2000;275:8903–10.
[41] Sanak M, Simon HU, Szczeklik A. Leukotriene C4 synthase polymorphism and risk of aspirin-inducedasthma. Lancet 1997;350:1599–600.
[42] Van Sambeek R, Stevenson DD, Baldarsao M, Lam BK, Zhao JL, Yoshida A, et al. 5′ Flanking regionpolymorphism of the gene encoding leukotriene C4 synthase does not correlate with the aspirin-intolerantasthma phenotype in the United States. J Allergy Clin Immunol 2000;106:72–6.
[43] Soderstrom M, Bolling A, Hammarstrom S. Induction of leukotriene C4 synthase activity in differentiatinghuman erythroleukemia cells. Biochem Biophys Res Commun 1992;189:1043–9.

520 B.K. Lam, K. Frank Austen / Prostaglandins & other Lipid Mediators 68–69 (2002) 511–520
[44] Riddick CA, Serio KJ, Hodulik CR, Ring WL, Regan MS, Bigby TD. TGF-� increase leukotriene C4 synthaseexpression in the monocyte-like cell line, THP-1. J Immunol 1999;162:1101–7.
[45] Boyce JA, Lam BK, Penrose JF, Friend DS, Parsons S, Owen WF, et al. Expression of LTC4 synthase duringthe development of eosinophils in vitro from cord blood progenitors. Blood 1996;88:4338–47.
[46] Hsieh FH, Lam BK, Penrose JF, Austen KF, Boyce JA. T helper cell type 2 cytokines coordinately regulateimmunoglobulin E-dependent cysteinyl leukotriene production by human cord blood-derived mast cells:profound induction of leukotriene C4 synthase expression by interleukin 4. J Exp Med 2001;193:123–33.
[47] Kanaoka Y, Maekawa A, Penrose JP, Austen KF, Lam BK. Attenuated zymosan-induced peritoneal vascularpermeability and IgE-dependent passive cutaneous anaphylaxis in mice lacking leukotriene C4 synthase. JBiol Chem 2001;276:22608–13.










![Stimulation by Leukotriene D4of Increases Cytosolic … · LTC4,[3H]LTD4, 14,15(N)[3H]leukotrienes C4, D4;NIF,nifedipine. methionyl-leucyl-phenylalanine (fMLP)(1, 2)andleukotriene](https://img.pdfslide.net/doc/110x75/5e6b548066b3967a27736aa8/stimulation-by-leukotriene-d4of-increases-cytosolic-ltc43hltd4-1415n3hleukotrienes.jpg)


















