Embed Size (px)
Citation preview

LIANA CLIMBING STRATEGIES AND RELATIONSHIP WITH HOST TREES IN MO SINGTO FOREST DYNAMICS PLOT,
KHAO YAI NATIONAL PARK, THAILAND
NATTHIDA KHIEWBANYANG
A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR
THE DEGREE OF MASTER OF SCIENCE (ENVIRONMENTAL BIOLOGY)
FACULTY OF GRADUATE STUDIES MAHIDOL UNIVERSITY
2015
COPYRIGHT OF MAHIDOL UNIVERSITY

Thesis entitled
LIANA CLIMBING STRATEGIES AND RELATIONSHIP WITH HOST TREES IN MO SINGTO FOREST DYNAMICS PLOT,
KHAO YAI NATIONAL PARK, THAILAND
……………….………….…..………….. Miss Natthida Khiewbanyang Candidate
……………………….….…..………… Asst. Prof. Jenjit Khudamrongsawat, Ph.D. (Biology) Major advisor
……………………….….…..………….. Lect. Chanpen Saralamba, Ph.D. (Biology) Co-advisor
……………………….….…..………. Prof. Patcharee Lertrit, M.D., Ph.D. (Biochemistry) Dean Faculty of Graduate Studies Mahidol University
……………….….…..………………….. Assoc. Prof. Prayad Pokethitiyook, Ph.D. (Chemical Engineering) Program Director Master of Science Program in Environmental Biology Faculty of Science Mahidol University

Thesis entitled
LIANA CLIMBING STRATEGIES AND RELATIONSHIP WITH HOST TREES IN MO SINGTO FOREST DYNAMICS PLOT,
KHAO YAI NATIONAL PARK, THAILAND
was submitted to the Faculty of Graduate Studies, Mahidol University for the degree of Master of Science (Environmental Biology)
on March 9, 2015
……………….………….…..………… Miss Natthida Khiewbanyang Candidate
………………….…..…………………. Asst. Prof. Wirong Chanthorn, Ph.D. (Biology) Chair
……………………….….…..………… Prof. Patcharee Lertrit, M.D., Ph.D. (Biochemistry) Dean Faculty of Graduate Studies Mahidol University
……………………….….…..……….. Asst. Prof. Jenjit Khudamrongsawat, Ph.D. (Biology) Member ……………………….….…..………… Lect. Chanpen Saralamba, Ph.D. (Biology) Member ……………….….…..………………… Asst. Prof. Somsak Dangtip, Ph.D. (Applied Nuclear Physics) Acting Dean Faculty of Science Mahidol University

iii
ACKNOWLEDGEMENTS
I would like to express my gratitude to my advisor, Assistant Professor Dr.
Jenjit Khudamrongsawat for the valuable guidance, advice, and motivation. This
master thesis would not be possible without her continuous support and patience.
I am grateful to my committee members Dr. Chanpen Saralamba for
introducing me to the topic as well for the support, encouragements and valuable
advices through the learning process of this master thesis. I would also like to thank
Assistant Professor Dr. Wirong Chanthorn for being the external examiner of my
thesis defense including useful suggestions for improvement. My grateful thanks are
also extended to Miss. Anuttara Natalang who gave the permission to use all required
information, the necessary material to complete the research work and including useful
comments, remarks and engagements. I am using this opportunity to express my
gratitude to Professor Warren Y. Brockelman who gave me an opportunity to
participate and learn in Mo Singto Forest Dynamics Plot.
This thesis work is supported by Thailand Graduate Institute of Science
and Technology (TGIST), TGIST-01-55-020 under National Science and Technology
Development Agency (NSTDA).
I gratefully thank Department of National Parks, Wildlife and Plant
Conservation for permission to collect information in Khao Yai National Park.
Also, I would like to thank the assistants in my fieldwork, who have
willingly shared their precious time during the process of fieldwork. I thank my fellow
graduate students and friends for their various helps and supports.
Finally, I would like to thank my parents for their kindness, wonderful
support and understanding throughout my study.
Natthida Khiewbanyang

iii

iii

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 62
BIOGRAPHY
NAME Natthida Khiewbanyang
DATE OF BIRTH 28 October 1987
PLACE OF BIRTH Bangkok, Thailand
INSTITUTIONS ATTENDED Mahidol University, 2006-2009
Bachelor of Science
(Conservation Biology)
Mahidol University, 2010-2014
Master of Science
(Environmental Biology)
SCHOLARSHIP RECEIVED Scholarship for Young Scientists,
2010-2011
Thailand Graduate Institute of Science and
Technology, 2012-2013
HOME ADDRESS 105 Soi Suan Phak 29, Suan Phak Rd.,
Taling Chan, Bangkok, THAILAND 10170
E-mail: [email protected]

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 55
APPENDIX
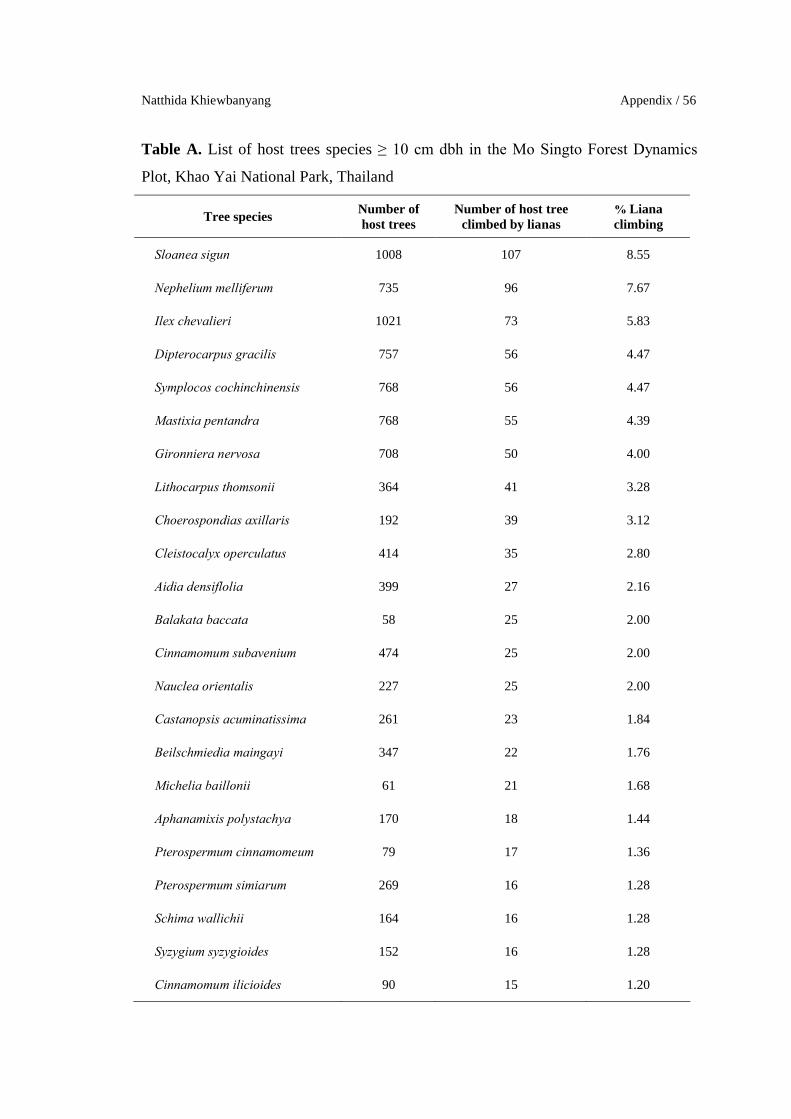
Natthida Khiewbanyang Appendix / 56
Table A. List of host trees species ≥ 10 cm dbh in the Mo Singto Forest Dynamics
Plot, Khao Yai National Park, Thailand
Tree speciesNumber of host trees
Number of host treeclimbed by lianas
% Liana climbing
Sloanea sigun 1008 107 8.55
Nephelium melliferum 735 96 7.67
Ilex chevalieri 1021 73 5.83
Dipterocarpus gracilis 757 56 4.47
Symplocos cochinchinensis 768 56 4.47
Mastixia pentandra 768 55 4.39
Gironniera nervosa 708 50 4.00
Lithocarpus thomsonii 364 41 3.28
Choerospondias axillaris 192 39 3.12
Cleistocalyx operculatus 414 35 2.80
Aidia densiflolia 399 27 2.16
Balakata baccata 58 25 2.00
Cinnamomum subavenium 474 25 2.00
Nauclea orientalis 227 25 2.00
Castanopsis acuminatissima 261 23 1.84
Beilschmiedia maingayi 347 22 1.76
Michelia baillonii 61 21 1.68
Aphanamixis polystachya 170 18 1.44
Pterospermum cinnamomeum 79 17 1.36
Pterospermum simiarum 269 16 1.28
Schima wallichii 164 16 1.28
Syzygium syzygioides 152 16 1.28
Cinnamomum ilicioides 90 15 1.20
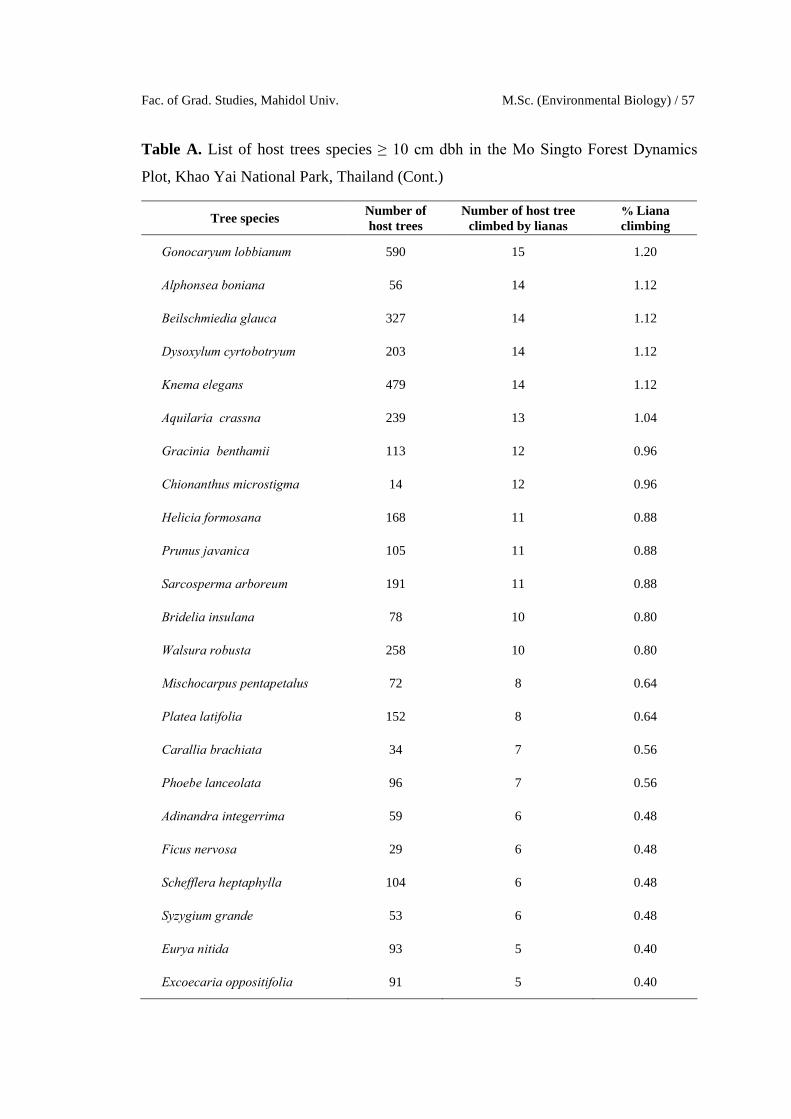
Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 57
Table A. List of host trees species ≥ 10 cm dbh in the Mo Singto Forest Dynamics
Plot, Khao Yai National Park, Thailand (Cont.)
Tree speciesNumber of host trees
Number of host treeclimbed by lianas
% Liana climbing
Gonocaryum lobbianum 590 15 1.20
Alphonsea boniana 56 14 1.12
Beilschmiedia glauca 327 14 1.12
Dysoxylum cyrtobotryum 203 14 1.12
Knema elegans 479 14 1.12
Aquilaria crassna 239 13 1.04
Gracinia benthamii 113 12 0.96
Chionanthus microstigma 14 12 0.96
Helicia formosana 168 11 0.88
Prunus javanica 105 11 0.88
Sarcosperma arboreum 191 11 0.88
Bridelia insulana 78 10 0.80
Walsura robusta 258 10 0.80
Mischocarpus pentapetalus 72 8 0.64
Platea latifolia 152 8 0.64
Carallia brachiata 34 7 0.56
Phoebe lanceolata 96 7 0.56
Adinandra integerrima 59 6 0.48
Ficus nervosa 29 6 0.48
Schefflera heptaphylla 104 6 0.48
Syzygium grande 53 6 0.48
Eurya nitida 93 5 0.40
Excoecaria oppositifolia 91 5 0.40
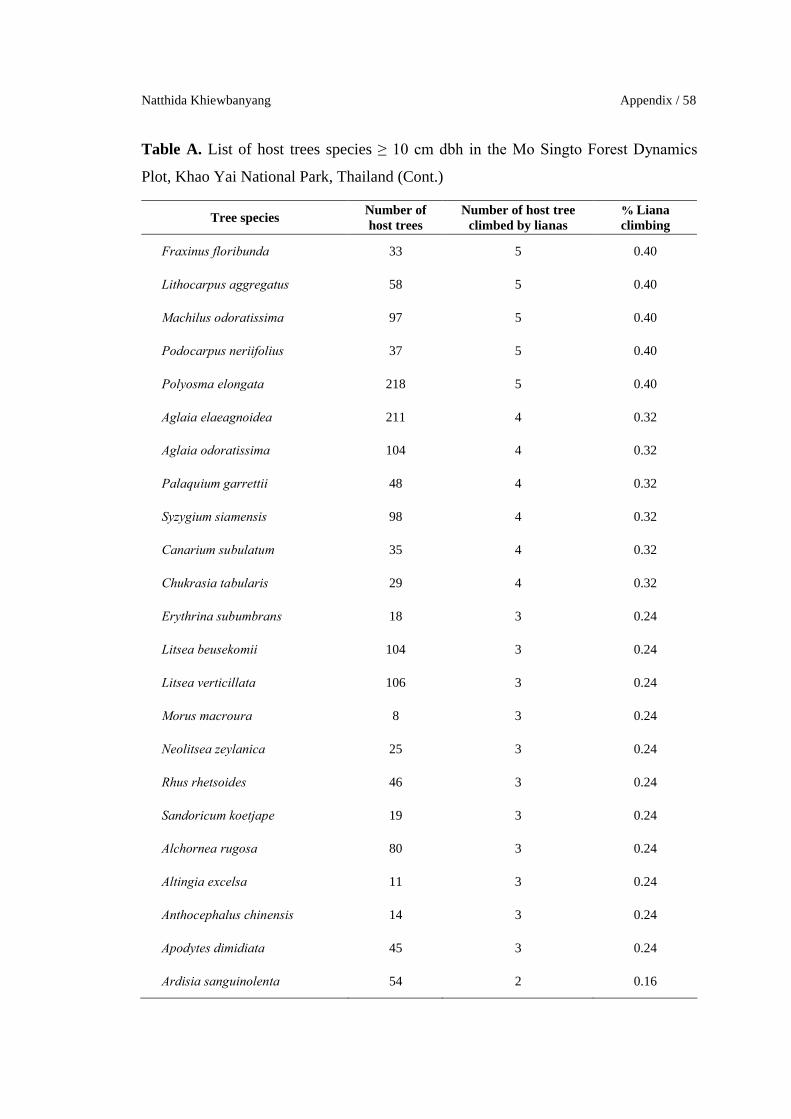
Natthida Khiewbanyang Appendix / 58
Table A. List of host trees species ≥ 10 cm dbh in the Mo Singto Forest Dynamics
Plot, Khao Yai National Park, Thailand (Cont.)
Tree speciesNumber of host trees
Number of host treeclimbed by lianas
% Liana climbing
Fraxinus floribunda 33 5 0.40
Lithocarpus aggregatus 58 5 0.40
Machilus odoratissima 97 5 0.40
Podocarpus neriifolius 37 5 0.40
Polyosma elongata 218 5 0.40
Aglaia elaeagnoidea 211 4 0.32
Aglaia odoratissima 104 4 0.32
Palaquium garrettii 48 4 0.32
Syzygium siamensis 98 4 0.32
Canarium subulatum 35 4 0.32
Chukrasia tabularis 29 4 0.32
Erythrina subumbrans 18 3 0.24
Litsea beusekomii 104 3 0.24
Litsea verticillata 106 3 0.24
Morus macroura 8 3 0.24
Neolitsea zeylanica 25 3 0.24
Rhus rhetsoides 46 3 0.24
Sandoricum koetjape 19 3 0.24
Alchornea rugosa 80 3 0.24
Altingia excelsa 11 3 0.24
Anthocephalus chinensis 14 3 0.24
Apodytes dimidiata 45 3 0.24
Ardisia sanguinolenta 54 2 0.16
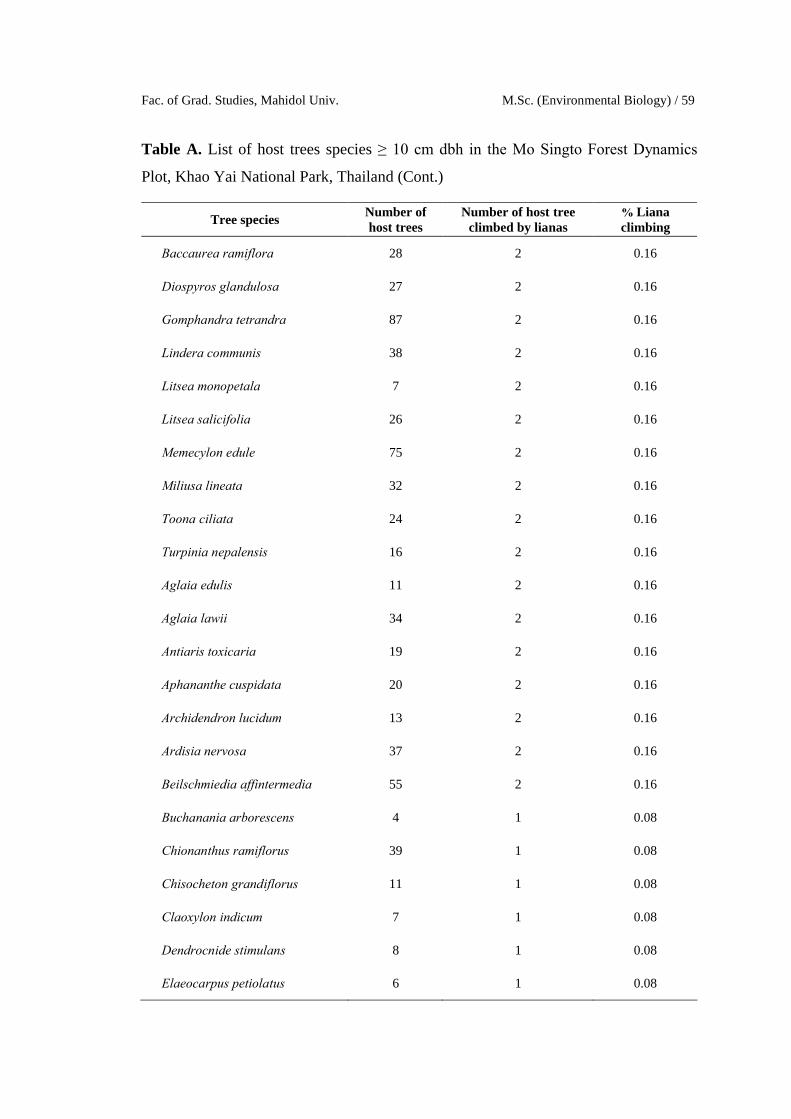
Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 59
Table A. List of host trees species ≥ 10 cm dbh in the Mo Singto Forest Dynamics
Plot, Khao Yai National Park, Thailand (Cont.)
Tree speciesNumber of host trees
Number of host treeclimbed by lianas
% Liana climbing
Baccaurea ramiflora 28 2 0.16
Diospyros glandulosa 27 2 0.16
Gomphandra tetrandra 87 2 0.16
Lindera communis 38 2 0.16
Litsea monopetala 7 2 0.16
Litsea salicifolia 26 2 0.16
Memecylon edule 75 2 0.16
Miliusa lineata 32 2 0.16
Toona ciliata 24 2 0.16
Turpinia nepalensis 16 2 0.16
Aglaia edulis 11 2 0.16
Aglaia lawii 34 2 0.16
Antiaris toxicaria 19 2 0.16
Aphananthe cuspidata 20 2 0.16
Archidendron lucidum 13 2 0.16
Ardisia nervosa 37 2 0.16
Beilschmiedia affintermedia 55 2 0.16
Buchanania arborescens 4 1 0.08
Chionanthus ramiflorus 39 1 0.08
Chisocheton grandiflorus 11 1 0.08
Claoxylon indicum 7 1 0.08
Dendrocnide stimulans 8 1 0.08
Elaeocarpus petiolatus 6 1 0.08
Natthida Khiewbanyang Appendix / 60
Table A. List of host trees species ≥ 10 cm dbh in the Mo Singto Forest Dynamics
Plot, Khao Yai National Park, Thailand (Cont.)
Tree speciesNumber of host trees
Number of host treeclimbed by lianas
% Liana climbing
Eugenia grata 16 1 0.08
Ficus glaberrima 10 1 0.08
Ficus stricta 5 1 0.08
Ficus vasculosa 17 1 0.08
Hopea odorata 1 1 0.08
Hydnocarpus ilicifolius 2 1 0.08
Ilex affchapaensis 3 1 0.08
Lithocarpus pattaniensis 25 1 0.08
Macaranga denticulata 1 1 0.08
Macaranga siamensis 18 1 0.08
Machilus affsalicina 8 1 0.08
Mallotus paniculatus 1 1 0.08
Mangifera longipetiolata 25 1 0.08
Markhamia stipulata 4 1 0.08
Nothapodytes montana 37 1 0.08
Ormosia sumatrana 13 1 0.08
Phoebe cathia 12 1 0.08
Platymitra macrocarpa 22 1 0.08
Podocarpus imbricatus 3 1 0.08
Pouteria stellibacca 3 1 0.08
Prunus arborea 28 1 0.08
Saurauia roxburghii 57 1 0.08
Shorea henryana 2 1 0.08
Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 61
Table A. List of host trees species ≥ 10 cm dbh in the Mo Singto Forest Dynamics
Plot, Khao Yai National Park, Thailand (Cont.)
Tree speciesNumber of host trees
Number of host treeclimbed by lianas
% Liana climbing
Sterculia balanghas 5 1 0.08
Tetrameles nudiflora 18 1 0.08
Triadica cochinchinensis 4 1 0.08
Viburnum cylindricum 50 1 0.08
Xanthophyllum virens 18 1 0.08
Total 15620 1251

Fac. of Grad. Studies, MahidolUniv. Thesis / iv
LIANA CLIMBING STRATEGIES AND RELATIONSHIP WITH HOST TREES IN MO
SINGTO FOREST DYNAMICS PLOT, KHAO YAI NATIONAL PARK, THAILAND
NATTHIDA KHIEWBANYANG 5337755SCEB/M
M.Sc. (ENVIRONMENTAL BIOLOGY)
THESIS ADVISORY COMMITTEE: WIRONG CHANTHORN, Ph.D.
JENJIT KHUDAMRONGSAWAT, Ph.D., CHANPEN SARALAMBA, Ph.D.
ABSTRACT
Lianas are woody climbing plants exhibiting a unique habit that requires host
trees as physical supports. High diversity of lianas in tropical forests involves its various and
distinct climbing strategies to ascend to the forest canopies in addition to species richness. The
diverse tactics of its climbing strategies are dependent on variations in the host tree
characteristics, particularly, the sizes and bark textures of host trees. The perennial plant
characteristics of trees on 30.5 hectares of the Mo Singto Forest Dynamics Plot, which is a
seasonal evergreen forest located in the Khao Yai National Park in Thailand, were examined.
Thirty ramets per liana species with diameter at breast height ≥ 3 cm of 1,560 liana ramets
were examined, and six climbing strategies were classified in this study. The dominant
climbing strategy was twiners, followed by the combination of twiners and scramblers,
scramblers, tendril, hook and adventitious root climbers, respectively. Liana ramet size was
positively correlated with the host tree size. However, this relationship varied with the type of
climbing strategies. Most twiners, hooks and tendril climbers tended to use small host trees.
The scrambler and the combination of twiner and scrambler ramets appeared to be
independent of host tree sizes. Adventitious root climbers tended to use large host trees. Most
ramets used host trees with slightly rough bark texture more often than other types of bark
texture with the exception of the adventitious root climbers that were often found on trees with
rough bark texture. In conclusion, the presence and abundance of ramets with different
climbing strategies could be associated with sizes and bark textures of host trees.
KEY WORDS: LIANAS / CLIMBING STRATEGY / SEASONAL EVERGREEN
FOREST / KHAO YAI NATIONAL PARK / HOST TREES
62 pages

Fac. of Grad. Studies, MahidolUniv. Thesis / v
กลวิธีการเลี้อยของเถาวัลยและความสัมพันธกับพืชใหอาศัยในแปลงสํารวจถาวรมอสิงโต อุทยานแหงชาติเขาใหญLIANA CLIMBING STRATEGIES AND RELATIONSHIP WITH HOST TREES IN MO SINGTO FOREST DYNAMICS PLOT, KHAO YAI NATIONAL PARK, THAILAND
ณัฐธิดา เขียวบานยาง 5337755 SCEB/M
วท.ม. (ชีววิทยาสภาวะแวดลอม)
คณะกรรมการที่ปรึกษาวิทยานิพนธ: วิรงค จันทร, Ph.D., เจนจิต คูดํารงสวัสดิ์, Ph.D., จันทรเพ็ญ ศรลัมพ, Ph.D.
บทคัดยอเถาวัลยหรือไมเถาเนื้อแข็งเปนพืชที่มีวิสัยเฉพาะตัวโดยมีโครงสรางของลําตนทอดเลื้อยหรือปน
ปายตนไมอื่นเพื่อยึดลําตนใหทรงตัวอยูได ความหลากชนิดของเถาวัลยในปาเขตรอนยังเกี่ยวของกับความหลากหลายของกลวิธีการเลื้อยที่เถาวัลยใชสําหรับเคลื่อนที่ไปสูเรือนยอด ความหลากหลายของกลวิธีการเลื้อยดังกลาวอาจขึ้นอยูกับลักษณะของพืชใหอาศัยโดยเฉพาะขนาดและลักษณะเปลือก จึงทําการศึกษาความสัมพันธดังกลาวบริเวณปาดิบชื้นแบบมีฤดูกาลในแปลงสํารวจถาวรขนาดพื้นที่ 30.5 เฮกตาร โดยเลือกศึกษาจากเถาวัลย 52 ชนิด ชนิดละ 30 ตน ขนาดเสนผาศูนยกลางลําตนมากกวาหรือเทากับ 3 เซนติเมตร จํานวนทั้งสิ้น 1,560 ตนและจําแนกกลวิธีการเลื้อย จากการศึกษาสามารถจําแนกกลวิธีการเลื้อยไดหกวิธี คือ การใชลําตนหรือกิ่งเลื้อยพัน (Twiners) การใชลําตนหรือกิ่งเลื้อยพันและพาดพิง (The combination of twiners and scramblers) การเลื้อยพันแบบพาดพิง (Scramblers) การใชมือพัน (Tendrils) การใชขอเกี่ยว (Hooks) และการใชรากพิเศษ (Adventitious root climbers) นอกจากนี้ยังพบวาขนาดของเถาวัลยมีความสัมพันธกับขนาดของตนไม แตอยางไรก็ดีความสัมพันธนี้ยังขึ้นอยูกับกลวิธีการเลื้อยที่เถาวัลยเลือกใชดวย โดยเถาวัลยที่ใชลําตนหรือกิ่งเลื้อยพัน ใชขอเกี่ยวและใชมือพันมักจะปรากฏบนตนไมที่มีขนาดเล็ก เถาวัลยที่ใชลําตนหรือกิ่งเลื้อยพันและพาดพิงและการเลื้อยพันแบบพาดพิงมักจะไมขึ้นอยูกับขนาดของตนไม และเถาวัลยที่ใชรากพิเศษมักจะปรากฎบนตนไมที่มีขนาดใหญ เถาวัลยสวนใหญจะปรากฎบนตนไมที่มีเปลือกคอนขางขรุขระยกเวนเถาวัลยที่ใชรากพิเศษมักจะปรากฎอยูบนตนไมที่มีเปลือกขรุขระมาก ในการศึกษาครั้งนี้แสดงใหเห็นวาการปรากฎและจํานวนของเถาวัลยที่ใชกลวิธีการเลื้อยตางๆกันมีความสัมพันธกับขนาดและลักษณะของเปลือกของพืชใหอาศัย
62 หนา

vi
CONTENTS
Page
ACKNOWLEDGEMENTS iii
ABSTRACT (ENGLISH) iv
ABSTRACT (THAI) v
LIST OF TABLES viii
LIST OF FIGURES ix
CHAPTER I INTRODUCTION 1
CHAPTER II LITERATURE REVIEWS 4
2.1 Lianas 4
2.2 Liana abundance and diversity 5
2.3 The roles and functions of lianas 6
2.4 Liana climbing strategies 6
2.5 Liana climbing stratrgies and their limiting factors 7
CHAPTER III MATERIALS AND METHODS 10
3.1 The Mo Singto Forest Dynamics Plot 10
3.2 Liana database and sampling 13
3.3 Liana climbing strategies, liana and host tree sizes, 14
and host tree bark textures
3.3.1 Climbing strategy classification 14
3.3.2 Liana and host tree size classification 19
3.3.3 Bark texture classification 20
3.4 Data anlyses 24
3.4.1 Liana climbing strategies 24
3.4.2 Relationships between liana size and host tree size, 24
liana climbing strategies and host tree size,
and liana climbing strategies and bark texture

vii
CONTENTS (cont.)
Page
CHAPTER IV RESULTS 26
4.1 Liana and host tree abundance 26
4.2 Liana climbing strategies 29
4.3 Liana and host tree relationship 31
4.3.1 Relationship between liana ramet sizes 31
and host tree sizes
4.3.2 Relationship between liana climbing strategies 33
and host tree sizes
4.3.3 Relationship between liana climbing strategies 40
and different host tree bark textures
CHAPTER V DISCUSSION 41
5.1 Lianas and host tree abundance 41
5.2 Lianas climbing strategies 41
5.3 Liana and host tree relationships 44
5.3.1 Relationship between liana ramet sizes 44
and host tree sizes
5.3.2 Relationship between liana climbing strategies 44
and host tree sizes
5.3.3 Relationship between liana climbing strategies 46
and host tree bark textures
REFERENCES 49
APPENDIX 55
BIOGRAPHY 62

Natthida Khiewbanyang Materials and Methods / 12
Figure 3.2 Map of the terrain on the Mo Singto Forest Dynamics Plot showing the
sampling area of 30.5 ha in four different habitats (hilltop, valley, slope and lowland)
and 20-meter survey points, the elevation ranging from 725-815 m above sea level
with 5-meter contour intervals, and the scale displaying distances on the axes in meters
(Brockelman et al., 2011).
m

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 13
3.2 Liana database and sampling
The data in this study obtained from liana and tree censuses managed by
the Ecology Laboratory of the National Center for Genetic Engineering and
Biotechnology (BIOTEC), under the National Science and Technology Development
Agency (NSTDA). Inventories of lianas and trees were kept in the form of computer
database using Microsoft Access with the following information: all living lianas and
tree stem diameter at breast height (dbh), species identification, and their locations that
were plotted into the x and y coordinate system. For this study, the 2011 recensus data
were used for liana ramet sampling.
A liana genet was defined as a rooted stems arising from a vegetative stem
of a plant. It could climb on a host trees but sometimes fell back to the ground. A
ramet (distinct genetic individual) ≥ 3 cm dbh was defined as an independent
individual that was not connected to any other aboveground or belowground stems
(Gerwing, 2004).
Trees were considered to be hosts when liana ramets climbed the trunk or
branch of the trees and were classified as primary host trees. They were the initial
support (Nabe-Nielsen, 2001).
All liana ramets with their host trees were assigned specific tagging
numbers. Monocotyledons, specifically rattans, that were the only monocot species
found in the study site and some hemiepiphytes were excluded because they were
represented by very few species and low in abundance. Lianas that could not
confidently be identified were also excluded from analyses. All selected liana ramets
were verified their living status to confirm the record in the database.
This study focused on the common species of liana ramets. All living
species were divided into common and rare species based on their densities (number
of ramets per hectare). Common species were species found at least one individual per
hectare while rare species were those with less than one individual per hectare. Thirty
ramets for each common species of lianas whose diameter at breast height (dbh) were
greater than or equal to 3 cm and found to climb host trees were randomly selected
from their tagging numbers by using a computer program (RAND function in
Microsoft office 2007). Thus, a total of 1,560 ramets of common liana species were
examined.

Natthida Khiewbanyang Materials and Methods / 14
3.3 Liana climbing strategies, liana and host tree sizes, and host tree
bark textures
3.3.1 Climbing strategy classification
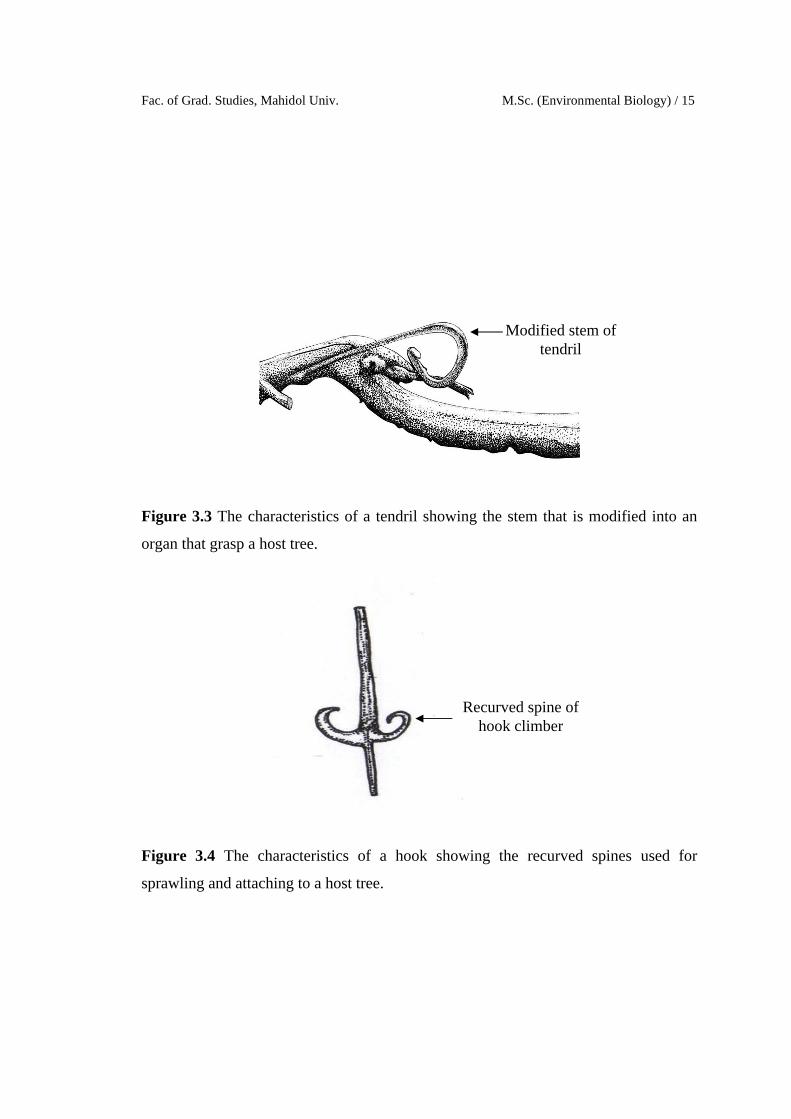
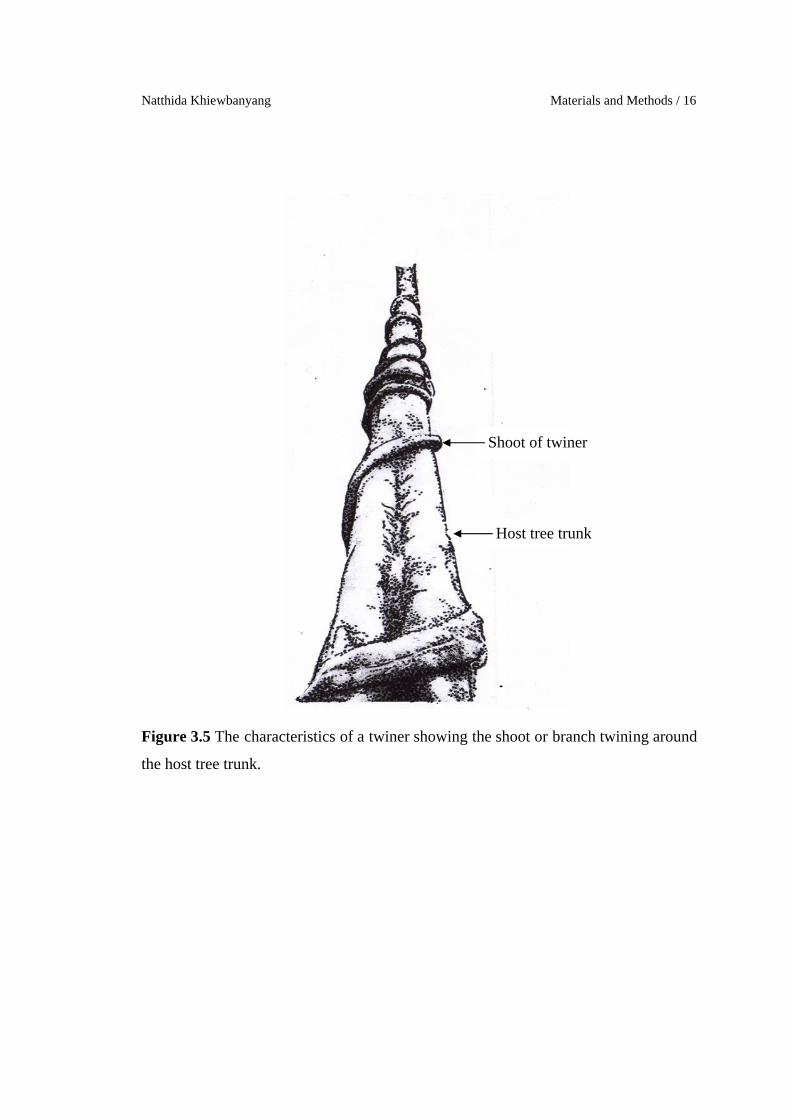
The climbing strategies for each liana stem were classified to five major
categories, which were tendril climbers, root climbers, twiners, scramblers, and hook
climbers as described in Chittibabu & Parthasarathy (2001); Dewalt et al. (2000),
Isnard & Silk (2009); Padaki et al. (2000); Putz (1984); Senbeta et al. (2005) (Figure
3.3). Other climbing strategies that were not included in these categories were also
described and recorded if found in the plot. All lianas encountered were photographed
for species confirmation.
Tendril climbers refer to modified leaves, leaflets or stipules, which twine
around a support (Chittibabu & Parthasarathy, 2001; Putz, 1984). Tendril grasp only
small diameter supports.
Root climbers refer to adventitious roots or aerial roots, adhered to host
trees and growing into cracks and crevices of the bark (Putz, 1984; Padaki et al.,
2000).
Twiners refer to apical portion of the shoots, branches or petioles, which
twine around the host trees (Dewalt et al., 2000; Isnard & Silk, 2009).
Scramblers refer to sprawlers, which lean on but do not closely attach to
host trees (Chittibabu & Parthasarathy, 2001; Putz, 1984).
Hook climbers refer to an organ such as recurved spines, hooks, or thorns,
which sprawl and attach to host trees (Isnard & Silk, 2009; Pakadi et al., 2000).

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 15
Figure 3.3 The characteristics of a tendril showing the stem that is modified into an
organ that grasp a host tree.
Figure 3.4 The characteristics of a hook showing the recurved spines used for
sprawling and attaching to a host tree.
Modified stem of tendril
Recurved spine ofhook climber

Natthida Khiewbanyang Materials and Methods / 16
Figure 3.5 The characteristics of a twiner showing the shoot or branch twining around
the host tree trunk.
Shoot of twiner
Host tree trunk

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 17
Figure 3.6 The characteristics of adventitious roots showing attaching to supporting
host tree.
Adventitious roots of root climber
Host tree trunk

Natthida Khiewbanyang Materials and Methods / 18
Figure 3.7 The characteristics of scramblers showing sprawling branches.

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 19
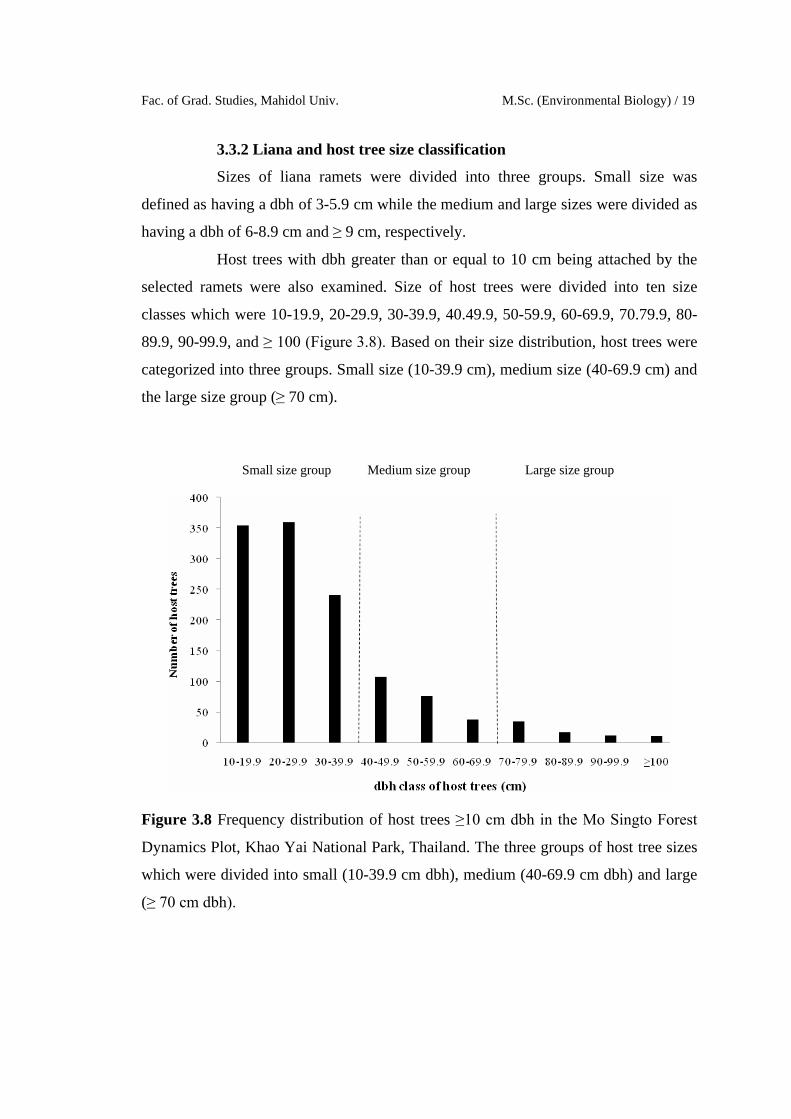
3.3.2 Liana and host tree size classification
Sizes of liana ramets were divided into three groups. Small size was
defined as having a dbh of 3-5.9 cm while the medium and large sizes were divided as
having a dbh of 6-8.9 cm and ≥ 9 cm, respectively.
Host trees with dbh greater than or equal to 10 cm being attached by the
selected ramets were also examined. Size of host trees were divided into ten size
classes which were 10-19.9, 20-29.9, 30-39.9, 40.49.9, 50-59.9, 60-69.9, 70.79.9, 80-
89.9, 90-99.9, and ≥ 100 (Figure 3.8). Based on their size distribution, host trees were
categorized into three groups. Small size (10-39.9 cm), medium size (40-69.9 cm) and
the large size group (≥ 70 cm).
Figure 3.8 Frequency distribution of host trees ≥10 cm dbh in the Mo Singto Forest
Dynamics Plot, Khao Yai National Park, Thailand. The three groups of host tree sizes
which were divided into small (10-39.9 cm dbh), medium (40-69.9 cm dbh) and large
(≥ 70 cm dbh).
Small size group Medium size group Large size group

Natthida Khiewbanyang Materials and Methods / 20
3.3.3 Bark texture classification
Bark texture varied from rough to smooth, depending on characteristics
including lenticels, branch scars, and wounds. Lichens and mosses were not
considered as a part of bark texture. Bark texture of hosts (all of them are associated
with ramets) were classified into smooth (S), slightly rough (SR) and rough (R)
(Campbell & Newbery, 1993; van der Heijden et al., 2008). All host trees were
photographed for confirmation of their bark characteristics by comparing with the
photographs and references as below.
Smooth bark or unbroken bark is the surface being smooth and glossy to
dull area. Generally these have a very thin outer bark and flat (Figure 3.9 a-b).
Slightly rough bark is the surface being any combination of small
dimples, shallow fissures and lenticels visible (Figure 3.10 c-f).
Rough bark is the surface with (g) scales or plates, (h) deeply fissures or
cracks, (i) deeply furrows or ridges, and (j) vertical strips (Figure 3.4 g-j).

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 21
Figure 3.9 Variations in characteristics of the smooth bark texture of host trees (a) and
(b).
a
b

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 23
Figure 3.11 Variations in characteristics of the rough bark texture of host trees (g)-(j)
g h
ji

Natthida Khiewbanyang Materials and Methods / 24
3.4 Data analyses
3.4.1 The climbing strategies of lianas
A type of climbing strategy (twiners, scramblers, adventitious roots,
tendrils, hooks and others if any) exhibited in each ramet was recorded.
The proportion of each climbing strategy was calculated by taking the
number of individual ramets with a given climbing strategy divided by the total
number of individual ramets of all climbing strategy.
Number of individual ramets for each climbing strategy
Total number of individual ramets
In terms of number of species, the proportion was calculated by taking the
number of ramet species with a given climbing strategy divided by the total number of
ramet species of all climbing strategy.
Number of ramet species for each climbing strategy
Total number of individual ramet species
To determine whether liana ramets demonstrate a selection for particular
host tree species, the number of individual trees observed as hosts was compared with
the number of individuals expected to be hosts using the 2 test (Chittibabu &
Parthasarathy, 2001). The average number of individual trees was calculated to
provide the expected proportion of liana colonization. The observed number of trees
hosting different number of lianas was calculated for each tree species.
3.4.2 Relationships between liana size and host tree size, liana climbing
strategies and host tree size, and liana climbing strategies and bark texture
Two different aspects of climbing strategies of lianas were examined. First,
simple linear regression analysis was conducted in order to clarify trends in the
relationship between liana ramet and host tree sizes. The dbh sizes of liana ramets and
host trees were plotted against each other. Second, the relationship between liana size
Proportion =
Proportion =

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 25
of each climbing strategy and host tree size were separately determined. The sizes
were plotted against one another with host tree dbh as the independent variable and
ramet dbh as the dependent variable.
The relationship between ramet size and host tree bark texture was
determined by plotting the number of ramets according to their size groups in a bar
graph. Each bar represented the number of ramet in an assigned size and separated into
different shades based on the types of host tree bark texture the ramet climbed. The
relationship between liana climbing strategies and host tree bark texture was examined
by plotting the number of ramets based on their size as bars. Different shades on a bar
were the number of ramets that use hosts of different bark texture. The preference of
hosts with particular bark texture was tested by using chi-square test based on the
availability of hosts.
The 2 and simple linear regression were analyzed by the PASW Statistics
for Windows, Version 18.0. Chicago: SPSS Inc. USA. All syntheses were test with
statistically significant relationships (p<0.05).

Natthida Khiewbanyang Literature Review / 4
CHAPTER II
LITERATURE REVIEW
2.1 Lianas
Climbing plants are classified into four fundamental categories (Gentry,
1991). The first category is comprised of herbaceous epiphytes, hemiepiphytes and all
herbaceous species that climb host trees by attaching their adventitious roots to host
tree trunks, e.g., Peteris pellusida and Asplenium nidus. The second category of these
climbers is woody hemiepiphytes and stranglers that begin life as epiphytic seedlings
by being rooted after reaching the ground (e.g. Ficus sagittata). Some species begin as
terrestrial climbers and use their adventitious root systems and/or losing contact for
climbing. The third category of woody climber plants is vines that are long and thin-
stemmed climbers, sometimes are referred to herbaceous vines. Many examples of
vines are found in the morning glory family Convolvulaceae. Vines emerge as
terrestrial seedling and are capable of growing in disturb habitats or forest edges. The
last category is lianas that have long and thick stems. Similar to vines, lianas emerge
as terrestrial seedling but are also able to grow in mature forests. They are also called
liane, brush rope, and woody vine (Putz, 1984). There are several common lianas in
tropical forests such as Tetracera indica and Gnetum montanum. Lianas also include
the climbing palms (the rattans).
Lianas exhibit a variety of shapes and sizes. They have evolutionary
evolved in diverse taxa such as gymnosperms, palms and monocotyledons (Cai et al.,
2009). In general, lianas have long and wide vessels that ensure efficient water
transport throughout their long stems (Cai et al., 2009; Isnard & Silk, 2009). They are
rooted in the ground but need structural support for their long and narrow stems (Putz,
1984; Putz & Chai, 1987). Lianas can reproduce by seeds or some portion of stem
such as sprouting, which enable them to colonize host trees (Nabe-Nielsen & Hall,
2002). Young lianas may not depend on host trees for structural support as the growth
rate is slow. The flexible stems of lianas allow them to grow without support up to
Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 5
about 1.5 m in height. As they approach their maturity, rapid growth rate occurs, and
their stems may need host trees to ascend the canopy. Without host trees for structural
support, lianas can still continue their growth but at much slow rates (Putz 1984).
Lianas can grow long distances from the initial rooting point, and so long-distance
cronal is a general method of liana colonization (Nabe-Nielsen & Hall, 2002; Yorke et
al., 2013). Lianas are shade tolerant plants waiting suitable light conditions (Malizia &
Grau, 2006; Schnitzer & Bongers, 2002) and supports (Campanello et al., 2007; Carse
et al., 2000; Malizia & Grau, 2006; Ladwig & Meiners, 2010). Upon reaching host
trees, lianas increase their growth rates, sizes and reproductive outputs. Lianas are able
to survive when they fall from their host trees (Peñalosa, 1984 but see Putz, 1984).
Lianas can produce physiologically independent ramets making liana genets very long
lived. A capacity of liana genets to survive by reproducing asexually when they are
unable to regenerate sexually may also enable them to survive in a wider range of
habitats than do other plants.
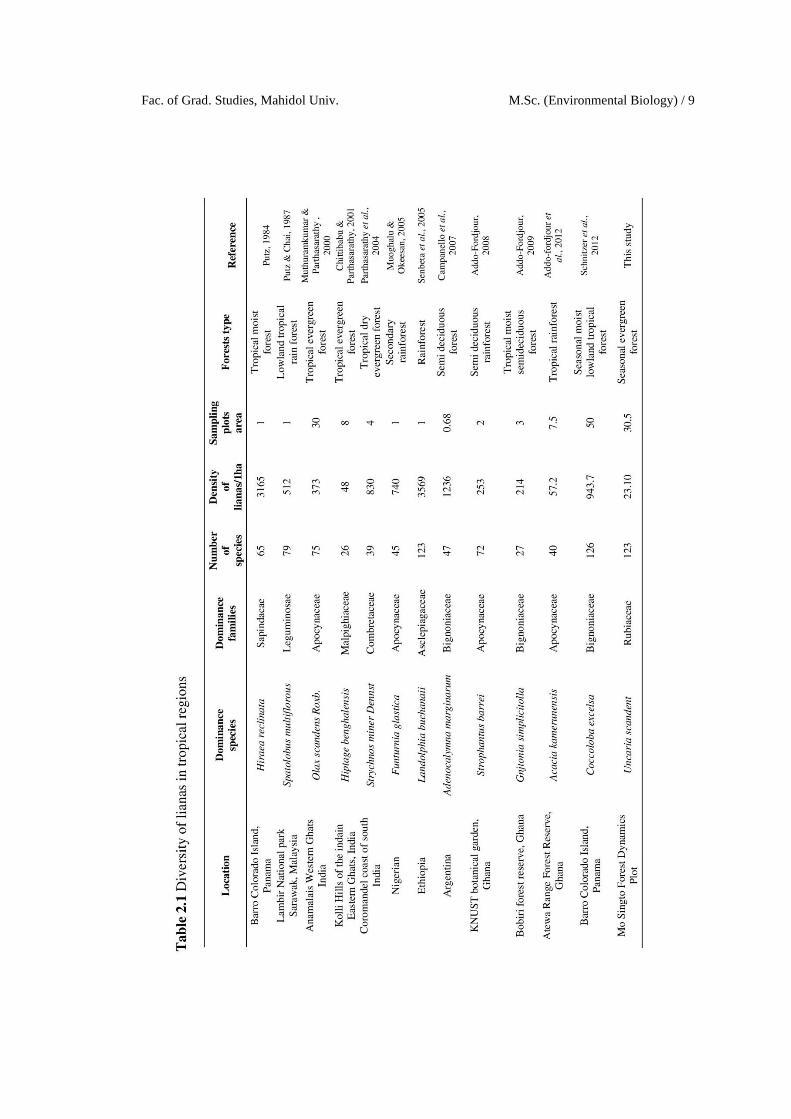
2.2 Liana abundance and diversity
Lianas diversity throughout the world had differences in terms of overall
abundance and richness. In the temperate and subtropical forests, lianas are common
in many areas but their abundance and richness are lower than those in tropical forests
(Durigon et al., 2014; Schnitzer & Bongers, 2002). In tropical forests, liana
abundance, diversity, and taxonomic composition are relatively high among tropical
regions especially in Africa (Senbeta et al., 2005). Many similar studies in different
tropical forests have reported similar results (e.g. carried out in studies Addo-Fordjour,
2008; Muoghalu & Okeesan, 2005; Muthuramkumar & Parthasarathy, 2000; Putz,
1984; Putz & Chai, 1987; Reddy & Parthasarathy, 2003; Schnitzer et al., 2012).
Although lianas are abundant common in many temperate forests, abundance,
diversity and taxonomic composition is low compared with tropical forests (Schnitzer
et al., 2000). A review of liana diversity inventories in various tropical forest sites is
presented in Table 2.1.
Natthida Khiewbanyang Literature Review / 6
2.3 The roles and functions of lianas
Lianas not only form a community component but also play important
roles in several aspects of forest dynamics such as suppressing tree regeneration
(Schnitzer & Carson, 2010), increasing tree mortality (Ingwell et al., 2010), serving as
food sources for many animals such as hornbills, monkeys and gibbons (Kitamura et
al., 2004; Kunz & Linsenmair, 2010; Vogel et al., 2009), providing habitat corridors
for arboreal animals such as gibbons and monkeys Schnitzers & Bongers, 2002) and
binding tree causing the stability of the individual trees (Schnitzers & Bongers, 2002).
The influence of lianas on forest dynamics may result from their interactions with host
trees. Despite many advantages of lianas, forest structures may be modified if liana
diversity and abundance change (DeWalt et al., 2000). In areas where lianas become
abundant, they may negatively impact tree species in gaps and disturbance areas
(Mohandass et al., 2015). For instance, lianas can obstruct tree growth and
regeneration by competing for space (Schnitzer et al., 2000) and causing mechanical
damages such as removing bark, shoot and bud of host trees (Putz, 1984). However,
this negative effect benefits pioneer tree species by promoting growth, recruitment and
survival of these trees (Schnitzer & Carson, 2010).
2.4 Liana climbing strategies
Most lianas climb their hosts using different climbing strategies and
require suitable structural supports to move upward. Climbing plants also show great
diversity in their climbing strategies. Liana climbing strategies have been classified
into tendril climbers, root climbers, twiners, scramblers and hook climbers
(Anbarashan & Parthasarathy, 2013; Campanello et al., 2007; Chittibabu &
Parthasarathy, 2001; DeWalt et al., 2000; Putz, 1984; Senbeta et al., 2005).
Twiners are portions of shoots, branches, or petioles that twine or coil
around supports. Twiners often climb in one direction, and show a predisposition to
turn to the right (Burnham & Revilla-Minaya, 2011). The twiner strategy is efficient
for ascending small host trees. This mode of attachment requires close contact with
surface. The main stem twines around support and is replaced by lateral branches.

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 7
Lianas develop an extensive root system. Root climbers use an
adventitious root to attach to the bark of host trees. This mode of attachment requires
close contact with surface (Putz, 1984). Tendril climbers are the modified leaves,
leaflets, inflorescence, flower peduncle or stipules twine around a support (Chittibabu
& Parthasarathy, 2001, Putz, 1984). At their young state, tendrils are relatively straight
and sensible to touch. Rapid growth occurs when tendrils come into contact with host
trees. Tendril climbers can grasp small supports. They are more suitable to gaps and
forest edges where small supports are usually more common than in forest interiors.
Hook climbers are the passive climbing strategy (Pakadi et al., 2000). This
strategy uses an organ such as recurved spines, hooks, thorns, or outgrowth as a
prickle to attach to the surface of host trees (Putz, 1984). Scramblers or sprawlers lean
on but do not become closely attach to host trees (Putz, 1984). Sometimes, the
scramblers use leader shoots to climb to the host trellis and other shoots to climb
upward to the higher level of host trees.
These various climbing strategies not only form structural components in
any tropical forests and subtropical forests but also indicate different succession stages
of forests. Generally, tendril climbers are the early succession species whereas twiners
and root climbers are an indication of late succession species (DeWalt et al., 2000).
Several studies suggested that high proportions in species richness of twiners may
indicate that the forests may approach to a relatively late state of succession (Cai et al.,
2009; Chittibabu & Parthasarathy, 2001; DeWalt et al., 2000).
2.5 Liana climbing strategies and their limiting factors
The presence of suitability of host trees is an important factor influencing
liana access to the canopy. Most lianas climb their hosts using different climbing
strategies and require suitable structural supports to move upward. The studies in
several tropical and subtropical forests revealed that host trees characteristics may be
more important than environmental factors in determining the success of liana
colonization (Addo-Fordjour et al., 2009; van der Heijden & Phillips, 2008). Host
trees may differ in their suitability for liana colonization due to specific features such
as bark characteristics (Tally et al., 1996) or general factors such as trunk sizes

Natthida Khiewbanyang Literature Review / 8
(Jiménez-castillo & Lusk, 2009; Nabe-Nielsen & Hall, 2002), trunk length
(Campanello et al., 2007). However, the suitability of support also varies with the
strategies by which lianas due to biomechanical constraints (Carrasco-Urra & Gianoli,
2009).
Host tree sizes related with tree age in that older trees, which are usually
large (Nabe-Nielsen & Hall, 2002), tend to be colonized by many liana stems (Pérez-
Salicrup & Meijere, 2005). Several studies have found more lianas associated with
larger trees. Addo-Fordjour (2008) suggested that trees with larger sizes supported
larger lianas than small host trees. The abundance of lianas per tree tended to increase
when increasing sizes of host trees (Campanello et al., 2007). Larger trunk diameters
were also less susceptible to liana colonization due to biomechanical limitations of
liana climbing abilities on larger trees (Putz & Chai, 1987). However, larger trees
carry more lianas than trees with smaller sizes (Nabe-Nielsen, 2001, Préses-Silcarb
2005). These might be because trees with larger trunk sizes might have experienced
longer exposure time to lianas infestation than tree small trees (Alvira et.al., 2004;
Préses-Silcarb, 2005).
It has been found that the limit of trunk sizes could vary with the types of
climbing strategies in tropical forests (Putz, 1984). For example, tendril climbers can
use smaller support than stem twiners (DeWalt et al., 2000). On the other hand,
adhesive root and hook climbers can use much larger supports (Cai et al., 2009).
Climbing mechanism often dictates which host trees lianas can climb (Pénalosa, 1982
but see Ludwig & Mainer, 2010). Host tree species may differ substantially in liana
load, especially among tropical canopy trees (Addo-Fordjour et al., 2009; Putz &
Chai, 1987).
Host tree bark texture has been reported to be important in determining the
presence of different climbing strategies (Chittibabu & Parthasarathy, 2001; Muñoz et
al., 2003). Smoothness of bark appears to be the feature of host trees with low liana
colonization or that are completely liana free (Campanello et al., 2007). Campanello et
al. (2007) suggested that host tree species with low liana load tended to have smooth
bark. For example, 70% of palm (species name) that had very smooth bark and large
leave are free of lianas (Campanello et al., 2007).
Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 9

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 1
CHAPTER I
INTRODUCTION
1.1 Introduction
Lianas are woody climbing plants commonly found in most tropical forests
reaching the highest species diversity in tropical rain forests, evergreen forests, and
moist deciduous forests (Addo-Fordjour et al.,2009; Chittibabu & Parthasarathy, 2001,
Putz, 1984; Schnitzer & Bongers, 2002; Schnitzer, 2005; Senbeta et al., 2005).
However, several studies have suggested that environmental factors influence liana
distribution and abundance (DeWalt et al., 2010; Hu et al., 2010; Nabe-Nielsen & Hall
2002). Some studies have found that environmental factors may not directly influence
the liana distribution and abundance but rather the biotic factors such as the
availability of host trees and their characteristics could be related to liana abundance
(van der Heijden & Phillips, 2009; Nabe-Nielsen, 2001).
Because lianas rely on host trees for structural supports, a variety of
characteristics of host trees is important for the success of liana colonization. Most
lianas climb their hosts using different climbing strategies and require suitable
structural supports to move upward. Lianas employing the twiner and tendril strategies
appear to use hosts of small and medium host sizes (Nabe-Nielsen, 2001, Senbeta et
al., 2005), while lianas with adventitious roots and hooks require host trees that are
large enough to support their weight (Putz 1984). Moreover, host tree bark texture has
also been reported to be important in determining liana and host trees climbing success
(Muthuramkumar & Parthasarathy, 2001). Rough bark texture provides attachment
points for climbing better than trees with smooth bark texture (Campbell & Newbery,
1993; Muthuramkumar & Parthasarathy, 2001). Therefore, different characteristics of
host trees influence the presence of certain climbing strategy of lianas and may also be
related to liana diversity and abundance, which also influence forest dynamics.
Areas of biodiversity hot spot such as tropical forests in Thailand are also
rich in liana diversity. The Mo Singto Forest Dynamics Plot, which is a network of the

Natthida Khiewbanyang Introduction / 2
Center for Tropical Forest Science (CTFS), shows great liana diversity. The
relationship between liana and their limiting factors had previously been studied by
Lertpanich & Brockelman (2003). This study suggested that environmental factors did
not much determine the overall density and diversity of lianas in this plot. Host trees
with the diameter at breast height (dbh) less than 70 cm were highly colonized by
many liana ramets from different species. Therefore, biotic factors such as the
availability of host trees and their characteristics were probably related to liana
climbing success but these factors had yet been investigated. If the climbing strategies
of lianas were related to host tree characteristics, the following questions were
addressed.
1. What were the proportions of all liana climbing strategies in the Mo
Singto Forest Dynamics Plot?
2. Did different liana climbing strategies show selections for different
sizes and different bark textures of host trees?
1.2 Research Hypotheses
1.2.1 The diverse climbing strategies of lianas could be found at the Mo
Singto Forest Dynamics Plot and the proportion of lianas abundance with different
climbing strategies varied with types of supports and structures of the forests. It was
expected that twiner would be the dominant climbing strategy in the studied area
because twiners could generally use supports in the forest with high abundance of
small and medium trees.
1.2.2 Different liana climbing strategies were expected to select for
particular sizes and bark textures of host trees.
1.2.2.1 Twiners and tendril climbers were able to climb on
small sizes whereas lianas using other climbing strategies, such as the scramblers,
hook and adventitious root climbers could use larger host tree size.
1.2.2.2 Bark textures could also influence the presence of
lianas on host trees in that lianas preferred rough bark to smooth bark because liana

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 3
stem directly attach to host bark. Roughness of the bark promoted more areas that
lianas can climb host trees than smoothness of the bark.
1.3 Research Objectives
1.3.1 To record and classify climbing strategies of lianas in Mo Singto
Forest Dynamics Plot
1.3.2 To determine the proportions of different liana climbing strategies
1.3.3 To determine the relationships of different liana climbing strategies
on different sizes and bark textures of host trees

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 41
CHAPTER V
DISCUSSION
5.1 Lianas and host tree abundance
Liana diversity and abundance vary among sites throughout the tropical
forests (Putz, 1984; Schnitzer & Bongers, 2002; Schnitzer, 2012). Species richness of
lianas recorded in this study (123 species of 35 families found in 30.5 ha) was
relatively high when compared with some studies carried out in tropics. For instance,
Muthuramkumar & Parthasarathy (2000) recorded 75 species of lianas in a 30 ha plot
at Varagariar India, whereas Muoghalu & Okeesan (2005) recorded 49 species of
lianas in 20.5 ha plot at rainforest, Nigeria. Therefore, 123 liana species recorded in
this study is fairly high, considering the fact that only liana whose dbh of greater than
or equal to 3 cm were counted. Only the study conducted in the evergreen forest,
Southwest China, did identify the higher number of liana species (147 liana species in
15 ha plot) than the number obtained from this study (Cai, 2009).
Most liana ramets use some host trees such as S. sigun, N. melliferum, and
I. chevalieri, quite frequently. This could be due to the high density of these host tree
species in the study plot. Nevertheless, these trees only accounted for approximately
20% of all trees used by lianas. Thus, high abundance was not the prime choice for
structural supports. Other characteristics of host trees, such as trunk size and bark
texture, may influence variations of liana climbing strategies in forests as revealed by
several studies (Addo-Fordjour et al., 2009; Heijden et al., 2008; Muthuramkumar &
Parthasarathy, 2001; Nabe-Nielsen, 2001; Ludwig & Meiners, 2010).
5.2 Lianas climbing strategies
The Mo Singto Forest Dynamics Plot harbored high diversity of liana
species that exhibited various climbing strategies. The sampling liana ramets exhibited
five different climbing strategies with the twiner strategy as the most common

Natthida Khiewbanyang Discussion / 42
strategy. This finding was consistent with previous studies of tropical forests in that
the majority of liana species exhibited the twiner strategy (Chittibabu & Parthasarathy,
2001; DeWalt et al., 2000; Ghollasimood et al., 2012, Parthasarathy et al., 2004; Putz,
1984). For instance, in the Afromontane rain forests in Ethiopia, Senbeta et al. (2005)
reported that twiner was the dominant climbing strategy. This was similar to the
studies in evergreen forests in India (Parthasarathy et al., 2004), the dipterocarp forest
in Lambia National Park in Malaysia (Putz & Chai, 1987), tropical forests in China
(Cai et al., 2009), and the semi-deciduous rain forest in Ghana (Addo-Fordjour et al.,
2008).
However, some studies in subtropical forests in China reported that the
adventitious root strategy was the most common climbing method (Yuan et al., 2009)
while the tendril strategy was common in the dry evergreen forest (Parthasarathy et
al., 2004) and subtropical Atlantic forests (Campanello et al., 2007). Thus, climbing
strategies of liana species could vary in different areas. These diverse climbing
strategies could be a result of variations in liana species diversity. Although most liana
species usually exhibited one type of climbing strategies, they may use different
strategies in different regions. For instance, species in the genus Dalbergia found in
Afromontane rain forests in Ethiopia employed the twining strategy to climb host trees
(Senbeta et al., 2005). However, some species in this genus at the Mo Singto Plot
instead exhibited the tendril strategy. Another case in the coastal hill forest in
Malaysia reported that Tetracera indica employed the twiner strategy (Ghollasimood
et al., 2012), but this species at the Mo Singto Plot used the combination of twiner and
scrambler strategies. The fact that lianas used varieties of climbing strategies to climb
the forest canopy could be a result of adaptation (Putz, 1984; Senbeta et al., 2005).
It had been hypothesized that the twiner and tendril strategies were more
frequently found than the scrambler strategy in most tropical forests (Addo-Fordjour,
2008; Anbarashan & Parthasarathy, 2013; Cai 2009; Santos et al., 2009). Some studies
suggested that the scrambler strategy was not an effective method (Gentry 1991, Pual
& Yavitt, 2011; Schröder et al., 2013). The twiners tended to use leader shoots to
twine around their host trees and then used the replacing branches to climb upward if
they failed to find a support (Gerwing, 2004). However, twining ramets were
sometimes unable to maintain tensional forces and then fell of the host trees (Silk &

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 43
Holbrook, 2005). Putz (1984) suggested that slipping down the trunks was
occasionally observed. Based on the observation in this study, some liana ramets
frequently began their climbing by using the twiner strategy and then used the
scrambler strategy to climb on trellis of host trees to reach the canopy. The
combination of two strategies may help lianas to remain longer on the trunk of host
trees.
In addition, various climbing strategies not only form structural
components in any tropical forests and subtropical forests but also indicate different
succession stages of forest (Letcher & Chazdon, 2012). The different forms of
climbing habits might be related to differences in forest structure, dynamics, and age
(DeWalt et al., 2000, Putz & Chai, 1987). The tendril strategy was usually common in
forests with high density of small trees, which were characteristics of an early
succession stage while stem and branch twiners were be more common in later
successional forests (DeWalt et al., 2000). Twiners were able to climb small to
medium host tree trunks (Nabe-Nielsen, 2001; Senbeta et al., 2005). This was an
obvious advantage to twiners when the small supports were limited and climb more
successfully trees in the late succession stage of forest (Cai, 2009; Putz & Chai, 1987)
Based on this suggestion, the high proportion of liana ramets employing the twiner
strategy may indicate that the Mo Singto Forest may be reaching the late state of forest
succession. However, this assumption is tentative and needs to be tested.
Comparisons among tropical forest regions revealed that the lianas in this
seasonal evergreen forest exhibited as diverse climbing strategies as seen in other
regions. The dominant climbing strategy in an area may be caused in part by
differences in availability of suitable supports. Since the exhibition of liana climbing
strategies do not depend on species of host trees. It was noticed that liana ramets had
no preference for particular species of host trees because climbing strategies were a
result of adaptation.

Natthida Khiewbanyang Discussion / 44
5.3 Liana and host tree relationships
5.3.1 The relationship between liana ramets and host tree sizes
This study found that the relationships between lianas and their host tree
sizes were not random. The positive relationship indicated that small ramets tended to
use small hosts trees. As these ramets became larger, they also used larger hosts.
Although small ramets could use host trees of any sizes, they were found to climb
small trees more often than larger trees. It is possible that small trees were more
abundant in the area than larger trees. Some studies suggested that some of small
ramets were capable of growing under the low light environment and could only use
small host trees (Nabe-Nielsen, 2001). However, large ramets could not use hosts of
any sizes. They required certain sizes of trees that were able to support their sizes.
In this study, the size of trees being sampled was limited to the minimum
of 10 cm dbh. Nevertheless, liana ramets could potentially use hosts smaller than 10
cm dbh, which were not sampled in this study. Host tree seedlings were also expected
to be important for lianas in addition to small trees. Large host trees with dbh larger
than 70 cm could serve as supports for ramets to reach higher canopy and get direct
sunlight for rapid growth (Malizia & Grau, 2006). Areas with trees whose dbh smaller
than 70 cm usually had high liana density, but this correlation was not statistically
significant (Lertpanich & Brockelman, 2003). Some liana ramets could grow
independently without attaching themselves to host trees, and would climb on the
hosts when they were large. Upright individuals of some species could grow greater
than 2 m tall before requiring any supports that were large enough to support their
weight (Putz, 1984). Therefore, certain sizes of ramets require appropriate host tree
sizes for their successful colonization. Nevertheless, this pattern is not always
followed because some ramets of certain climbing strategies appear to be independent
of host tree sizes.
5.3.2 The relationships between liana climbing strategies and host tree
sizes
The twiner ramets appeared to be associated with host trees with diameter
< 50 cm dbh (Nabe-Nielsen, 2001; Senbeta et al., 2005). However, ramets using

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 45
twiners may be found on large host trees as reported in DeWalt et al. (2000) in that the
relative abundance of stem twiners in a tropical forest increased with forest stand age
(or trunk diameter). These twiners had attached to and grown on their hosts. As their
hosts became larger, the lianas also got larger.
The variation in climbing hosts of different sizes was observed for hook
climbers, but most of them usually used small host trees rather than larger host trees.
In contrast, Putz (1984) suggested that liana ramets employing the hook strategy could
use host trees regardless of their size. It was probable that some hook climbers only
required the presence of either branches or trunks of their hosts. As long as host trees
offered areas for attachment, they would be suitable for hook climbers. Nevertheless,
small branches usually provided better support than large trees because they offered
more surface areas for attachment. Thus, hook climbers tended to use small trees but
could also use trees of any sizes that provided areas for attachment.
Tendril ramets also used small host trees rather than other hosts of larger
sizes, consistent with the previous finding (Leicht-Young et al., 2010; Putz 1984). Due
to the fact that tendril climbers used sensitive organs to attach to their supports, large
host trees were likely too large for direct climbing because a tendril must wrap itself
around the supports. However, because the results from this study indicated that
tendril ramets were independent of host tree size, it would be similar to the hook
climbers that only required surface areas for holding on to the hosts.
A weak correlation between scrambler and host sizes possibly suggested
that these ramets only required structures for leaning. As a result, size of hosts may not
matter. This supported the suggestion that scramblers may depend on the density of
supports and climb more successfully trees in young forest where there are frequently
dense clusters of small trees (Campanello et al., 2007). Scramblers in this study were
found to climb not only their host trees but also other lianas. Similarly, the
combination of twiner and scrambler strategy also showed a weak correlation with
host tree sizes. Small host tree trunk was often used by twiners in the ascending state
but in the canopy scrambling ramets allowed the lianas to extend into the upper
canopy on branches of their host trees. The combination of twiner and scrambler
ramets would be more prevalent in younger forest and this trend was also observed in

Natthida Khiewbanyang Discussion / 46
this study. Therefore, small host trees provided better support for the combination of
twiner and scrambler ramets than less dense and large host trees.
Unlike other climbing strategies, the sizes of most adventitious root
climbers were medium and large. This strategy was found to use medium and large
host trees more often than other climbing strategies (Carrasco-Urra & Gianoli, 2009).
Because adventitious root climbers directly attached to trunks therefore, large host
trees provided more support area for attachment (Tally et al., 1996). Although this was
similar to the requirement of hook and tendril climbers, the adventitious root ramets
were present mostly in medium or large size and required bigger surface areas. Thus,
they were not independent of host tree sizes unlike other organ climbers. Since these
organ climbers appeared to be independent of host tree sizes but still required large
surface areas for attachment, other characteristics of host trees may be important.
5.3.3 The relationships between liana climbing strategies and different
host tree bark textures
Bark textures were related to climbing success with fewer lianas on trees
with smooth bark and more lianas on trees with rough bark that increased surface area.
It is likely that texture of bark help liana to climb successfully.
Although the result of this study revealed the frequent use of hosts with
slightly rough bark texture, it was as expected since most available host trees had
slightly rough bark texture. For instance, the highest percentage (8.66%) of lianas used
Sloenea sigun as hosts might be due to its high abundance and density in this plot.
Illex chavachirii with heavy lianas colonization also had slightly rough bark. These
species of host trees were the most abundant species in the area. Host trees with
smooth bark texture would be low in liana colonization because they lacked
attachment areas while rough and slightly rough bark offered more area. However, the
use of bark textures of lianas may depend on the climbing strategy.
Bark textures were likely to influence liana climbing success (Carse, 2000;
Carsten et al., 2002; Muñoz et al., 2003; Ladwig & Meiners; 2010). Although host
trees in the sampling area had slightly rough bark texture were used by ramets of
various climbing strategies. Some strategies did not often use this type of hosts. The

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 47
results from this study revealed that hook climbers tended to use host trees with the
slightly rough bark because this type of texture offered the attachment areas.
Not all climbing strategies required surface areas for colonization, some
strategies, such as the scramblers, only lean on the hosts with attachment. Host surface
areas as well as bark textures were not important.
Preference of particular bark texture of host trees was also obvious in
ramets using adventitious roots. These ramets showed their particular preference for
hosts with rough bark texture rather than size. Ramets of Rhus radicans used
adventitious roots to climb host trees were associated with trees with rough bark (Tally
et al., 1996). However, it has been suggested that host trees with strongly peeling bark
would be difficult for lianas to colonize using adventitious roots (Jiménez-Castillo &
Lusk, 2009). Rough bark was not stable surfaces for root attachment (Tally et al.
1996). For instance, Choerospondias axillaris, Michelia baillonii, Balakata baccata,
Prunus javanica, and Aphananthe cuspidata which had crack and fissure bark, had a
higher level of colonization by adventitious root ramets than Nauclea orientalis and
Alphonsea boniana, which had scale and vertical strips their bark. Despite this
observation, the roughness of the bark of host trees could promote lianas
establishment. Host trees with smooth bark may help deter liana colonization because
these host trees lack attachment sites.
The success of ramets of different climbing strategies may be explained by
differences in availability and suitability of physical supports. Comparisons among
tropical forest regions revealed that the lianas in this seasonal evergreen forest
exhibited diverse climbing strategies as seen in other regions. The study added to the
evidence that the presence and abundance of liana ramets with different climbing
strategies could be associated with size and bark texture of host trees not host species
in Mo Singto Forest. Host tree size was an important factor related to liana abundance,
as small host trees could facilitate liana colonization and provide a better support for
lianas. However, large host trees may become the necessary support by providing
canopy access.
The use of host trees for climbing supports also depend on the bark
textures of host trees. Rough bark of host trees was important characteristic
determining liana colonization. In addition, this unique morphology and habit of lianas

Natthida Khiewbanyang Discussion / 48
could explain for their distribution across forest succession but it is still unclear and
requires further investigation.
Natthida Khiewbanyang Results / 26
CHAPTER IV
RESULTS
4.1 Liana and host tree abundance
A total of 1,560 liana ramets examined in this study belonged to 52
common species distributing in 23 families. The ten most abundant liana species were
Uncaria scandens, Uncaria macrophylla, Tetracera indica, Premna flavescens,
Sphenodesme pentandra, Sabia limoniacea, Tetrastigma laoticum, Chonemorpha
fragrans, Caelospermum truncatum, Spatholobus harmandii (Table 4.1). The size of
liana ramets that reached host trees varied considerably ranging from 3 cm to 19 cm
dbh. The number of liana ramets per individual host tree ranged from 1-6 with average
of 1.27 ± 0.29 ramets.
These liana ramets attached to 1,251 host trees belonging to 120 species in
25 families (Appendix A). Of these 1,251 host trees, the top ten most abundant species
were Ilex chevalieri, Sloanea sigun, Mastixia pentandra, Symplocos cochinchinensis,
Dipterocarpus gracilis, Nephelium melliferum, Gironniera nervosa, Gonocaryum
lobbianum, Knema elegan, and Cinnamomum subavenium. Although liana ramets
appeared to show host tree species preference (2 = 3681.86, df = 119, p < 0.05), the
majority host tree species being used as structural supports were actually the most
abundant species in the study area (Table 4.2). For example, S. sigun had the highest
proportion (8.66%) of trees being held the lianas. Other host tree species also often
being used for supports were N. melliferum (7.77%), and I. chevalieri (5.91%).

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 27
Table 4.1 List of liana species and climbing strategies of 52 common species with dbh
≥ 3 cm in 30.5 ha plot. AD = Adventitious root; HO = Hook; SC = Scrambler; TD =
Tendril; TW = Twiner; TS = Twiner and Scrambler
Family Species nameNumber of
individuals in 30.5 ha plot
Density/haClimbing strategy
Rubiaceae Uncaria scandens 693 23.10 HO
Rubiaceae Uncaria macrophylla 436 14.53 HO
Dilleniaceae Tetracera indica 300 10.00 TS
Verbenaceae Premna flavescens 264 8.80 TS
Verbenaceae Sphenodesme pentandra 259 8.63 TS
Sabiaceae Sabia limoniacea 258 8.60 TW
Vitaceae Tetrastigma laoticum 219 7.30 TD
Apocynaceae Chonemorpha fragrans 181 6.03 TW
Rubiaceae Caelospermum truncatum 172 5.73 TW
Leguminosae Spatholobus harmandii 165 5.50 TW
Celastraceae Celastrus approximata 163 5.43 TW
Apocynaceae Ichnocarpus polyanthus 160 5.33 AD
Rubiaceae Morinda villosa 151 5.03 TW
Rubiaceae Morinda umbellata 137 4.57 TW
Rubiaceae Uncaria laevigata 131 4.37 HO
Convolvulaceae Neuropeltis racemosa 124 4.13 TW
Celastraceae Salacia chinensis 121 4.03 TS
Leguminosae Dalbergia horrida 119 3.97 TD
Convolvulaceae Erycibe elliptilimba 117 3.90 TWMenispermaceae Diploclisia glaucescens 116 3.87 TS
Melastomataceae Diplectria barbata 115 3.83 SC
Convolvulaceae Erycibe subspicata 110 3.67 TW
Apocynaceae Aganosma harmandiana 105 3.50 TS
Annonaceae Cyathostemma micrantha 98 3.27 TD
Hernandiaceae Illigera pierrei 98 3.27 SC
Apocynaceae Pottsia laxiflora 95 3.17 TW
Rubiaceae Oxyceros longiflora 94 3.13 HO
Vitaceae Tetrastigma aff.pyriforme 93 3.10 SC
Connaraceae Roureopsis stenopetala 91 3.03 SC
Annonaceae Desmos dumosus 86 2.87 SC
Annonaceae Fissistigma oblongum 84 2.80 TS
Rhamnaceae Ventilago denticulata 82 2.73 TS
Gnetaceae Gnetum montanum 79 2.63 TS
Dilleniaceae Tetracera loureiri 77 2.57 TW
Annonaceae Uvaria cordata 60 2.00 TD

Natthida Khiewbanyang Results / 28
Table 4.1 List of liana species and climbing strategies of 52 common species with dbh
≥ 3 cm in 30.5 ha plot. AD = Adventitious root; HO = Hook; SC = Scrambler; TD =
Tendril; TW = Twiner; TW, SC = Twiner and Scrambler (Cont.)
Family Species nameNumber of
individuals in 30.5 ha plot
Density/haClimbing strategy
Annonaceae Uvaria lurida 55 1.83 SC
Moraceae Ficus sagittata 54 1.80 TW
Apocynaceae Urceola micrantha 54 1.80 AD
Elaeagnaceae Elaeagnus conferta 53 1.77 SC
Menispermaceae Hypserpa nitida 51 1.70 TS
Euphorbiaceae Phyllanthus reticulatus 50 1.67 SC
Connaraceae Rourea minor 49 1.63 TS
Combretaceae Combretum latifolium 48 1.60 TW
Thymelaeaceae Linostoma pauciflorum 47 1.57 TD
Apocynaceae Melodinus fusiformis 44 1.47 TW
Urticaceae Poikilospermum suaveolens 43 1.43 SC
Leguminosae Mucuna macrocarpa 40 1.33 TW
Piperaceae Piper ribesoides 39 1.30 TS
Piperaceae Piper retrofractum 36 1.20 TD
Leguminosae Dalbergia velutina 36 1.20 SC
Gnetaceae Gnetum macrostachyum 33 1.10 TS
Apocynaceae Strophanthus caudatus 32 1.07 TS

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 29
Table 4.2 The top ten most abundance species of host trees ≥ 10 cm dbh in the Mo
Singto Forest Dynamics Plot, KhaoYai National Park, Thailand.
4.2 Liana climbing strategies
From all lianas sample recorded, the most common climbing strategy was
the twiner strategy, which was found in 480 liana ramets from 16 liana species
followed by the scrambler strategy seen in 330 liana ramets from 10 species. The
tendril strategy, which was the third most common strategy, was found in 150 liana
ramets from six species while the hook strategy, which was less common, was present
in 120 liana ramets from four species. The least common strategy in the study area was
the adventitious root found in 60 ramets from two species (Table 4.3). Most liana
species exhibited one type of climbing strategies (2 = 544.615, df = 5, p < 0.05). For
example, all species in the family Convolvulaceae used the twiner strategy to climb up
host tree canopy. Most species of lianas (57.14%) in the family Apocynaceae only
used the twiner strategy. In the subfamily Papilionoideae (family Leguminosae), a few
species, all ramets in the genus Dalbergia, used the tendril strategy to climb host trees.
All species in the genus Uncaria used the hook strategy. However, there was an
exception in that some liana species employed the combination of twiner and
scrambler strategies. The liana species that employed this climbing combination
included Aganosma harmandiana, Diploclisia glaucescens, Fissistigma oblongum,
Gnetum macrostachyum, Gnetum montanum, Hypserpa nitida, Piper ribesoides,
Tree speciesNumber of individuals
Number of host tree climbed by lianas
% Liana climbing
Ilex chevalieri 1021 73 5.83
Sloanea sigun 1008 107 8.55
Mastixia pentandra 768 56 4.48
Symplocos cochinchinensis 768 55 4.39
Dipterocarpus gracilis 757 56 4.48
Nephelium melliferum 735 96 7.67
Gironniera nervosa 708 50 4.05
Gonocaryum lobbianum 590 41 3.32
Knema elegan 479 39 3.16
Cinnamomum subavenium 474 35 2.83

Natthida Khiewbanyang Results / 30
Premna flavescens, Rourea minor, Salacia chinensis, Sphenodesme pentandra,
Strophanthus caudatus, Tetracera indica, and Ventilago denticulata.
Table 4.3 The number of liana species and the proportion of individual ramets with
various climbing strategies in 30.5 ha seasonal evergreen forest, Mo Singto Forest
Dynamics Plot.
Climbing strategyNumber of individual
rametsProportion (%)
Number of species
Proportion (%)
Single climbing strategy
Twiner 480 30.77 16 30.77
Scrambler 330 21.15 10 19.23
Tendril 150 9.61 6 11.54
Hook 120 7.70 4 7.69
Adventitious root
Combination of two strategies
60 3.85 2 3.85
Twiner and Scrambler 420 26.92 14 26.92
Total 1560 100 52 100

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 31
4.3 Liana and host tree relationships
4.3.1 Relationships between liana ramet sizes and host tree sizes
Host trees and ramet sizes appeared to be correlate (r2 = 0.166, p < 0.05)
(Figure 4.1). Host trees with larger dbh tended to support liana ramets with larger dbh.
Small host trees could support small ramets. Medium host trees were found to be
related with medium size of ramets. Large ramets tended to use medium and large host
trees. The reverse was not true because large host trees could support all sizes of liana
ramets as small ramets less than 6 cm dbh were present on some of the large host trees.
Although a general trend showed the positive correlation between host
trees and liana ramet sizes, different size group of host trees did not equally provide
structural supports for the ramets. The majority of the lianas used small host trees
particularly of 20-30 cm dbh size. The abundance of liana ramets slightly decreased
when increasing host tree diameter and distinctively decreased at the medium host tree
size class (40-70 cm dbh).
Some ramets did not show any correlation with host tree sizes but rather
host tree bark texture. Host trees with slightly rough bark were usually used by liana
ramets of every size (Figure 4.2). However, patterns of some relationships were
detected under certain conditions, especially for different climbing strategies.

Natthida Khiewbanyang Results / 32
Figure 4.1 The relationship between liana and host tree sizes with a regression line
and r2 = 0.166, p < 0.05.
Figure 4.2 The percentage of liana ramets on three types of bark textures (smooth.
slightly rough and rough), for each of three size classes.

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 33
4.3.2 Relationships between liana climbing strategies and host tree
sizes
There was a positive correlation between liana climbing strategies and host
tree sizes. The results showed that the number of ramets using different climbing
strategies appeared to use certain sizes of host trees.
Ramets employing the twiner strategy showed a strong positive correlation
with host tree sizes. The sizes of these twiners increased with increasing host tree size
(r2 = 0.457, p < 0.05) (Figure 4.3). Most twiners were small and mostly use small host
(Figure 4.9a). Small twiners could twine around small host trees. Both medium and
large twiners were usually found on medium host trees and some of medium and large
twiners could also use large host trees. However, the abundance of twining ramets
decreased with increasing size of host trees. Observed values in each class of host tree
sizes showed that the twiners appeared to use certain sizes of host trees (2 = 15.27, df
= 2, p < 0.05). Although small host trees were the most abundant size of host trees, the
twiners showed more frequent use of small hosts than usual.
The sizes of hook climbers showed a weak correlation with host tree sizes
(r2 = 0.175, p < 0.05) (Figure 4.4). The majority of liana ramets using this climbing
strategy were small. These small ramets tended to use small host trees. However, small
ramets were found to use medium and large host trees as well. The abundance of liana
ramets in each class of host tree sizes showed that the hook climbers were found to use
small host trees but they also used hosts of other sizes as available (Figure 4.9b).
There was a weak positive correlation between ramets using the tendril
strategy and host tree size (r2 = 0.208, p < 0.05) (Figure 4.5). The majority of these
tendril climbers were also small and usually found on small host trees (Figure 4.9c).
Medium ramets appeared to use medium host trees as well as large host trees.
However, the data showed that in fact these ramets were independent of host tree sizes
(2=2.926, df = 2, p<0.05).
The correlation between size of ramets employing the scrambler strategy
and size of host trees also had a weak correlation (r2 = 0.069, p<0.05) (Figure 4.6).
Most of these ramets were small size. Although most of them were found on small
host trees, they appeared to be independent of host tree sizes (Figure 4.10d).

Natthida Khiewbanyang Results / 34
The similar situation was found in ramets that used the combination of
twiner and scrambler strategies to climb host trees. The size of these ramets was
weakly dependent on host tree sizes (r2 = 0.066, df = 2, p < 0.05) (Figure 4.7).
Although most of these ramets were small, they were able to attach to wide range of
host sizes (Figure 4.10e).
A weak correlation between ramets using the adventitious root strategy
and host tree sizes was found (r2 = 0.121, df = 2, p < 0.05) (Figure 4.8). Unlike other
ramets of other climbing strategies, ramets with this climbing strategy were mostly
medium in size. The majority of these ramets tended to use medium host trees (Figure
4.10f). Observed values in each class of host tree sizes showed that the adventitious
root climbers appeared to use medium size hosts more often than expected (2=15.27,
df = 2, p < 0.05).

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 35
Figure 4.3 The relationship between liana size and host tree size of the twiner strategy
with a regression line and r2 = 0.457, p < 0.05.
Figure 4.4 The relationship between liana size and host tree size of the hook strategy
with a regression line and r2 = 0.175, p < 0.05.

Natthida Khiewbanyang Results / 36
Figure 4.5 The relationship between liana size and host tree size of the tendril strategy
with a regression line and r2 = 0.208, p < 0.05.
Figure 4.6 The relationship between lianas diameter and host tree size of the
scrambler strategy with significant regression line at r2 = 0.069, p < 0.05.

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 37
Figure 4.7 The relationship between liana size and host tree size of the combination of
twiner and scrambler strategy with a regression line and r2 = 0.066, p < 0.05.
Figure 4.8 The relationship between liana size and host tree size of the adventitious
root strategy with significant regression line at r2 = 0.121, p < 0.05.

Natthida Khiewbanyang Results / 38
Figure 4.9 The host tree diameter size ≥10 cm dbh used by liana ramets of ≥3 cm dbh
at Mo Singto Forest Dynamics plot, Khao Yai National Park Thailand: (a) ramets
showing the twiner strategy, (b) the hook strategy, and (c) the tendril strategy.
(a)
(b)
(c)

Fac. of Grad. Studies, Mahidol Univ. M.Sc. (Environmental Biology) / 39
Figure 4.10 The host tree diameter size ≥10 cm dbh used by liana ramets of ≥3 cm
dbh at Mo Singto Forest Dynamics plot, Khao Yai National Park Thailand: (d) ramets
showing the scrambler strategy, (e) the combination of twiner and scrambler strategy,
and (f) the adventitious root strategy.
(d)
(e)
(f)
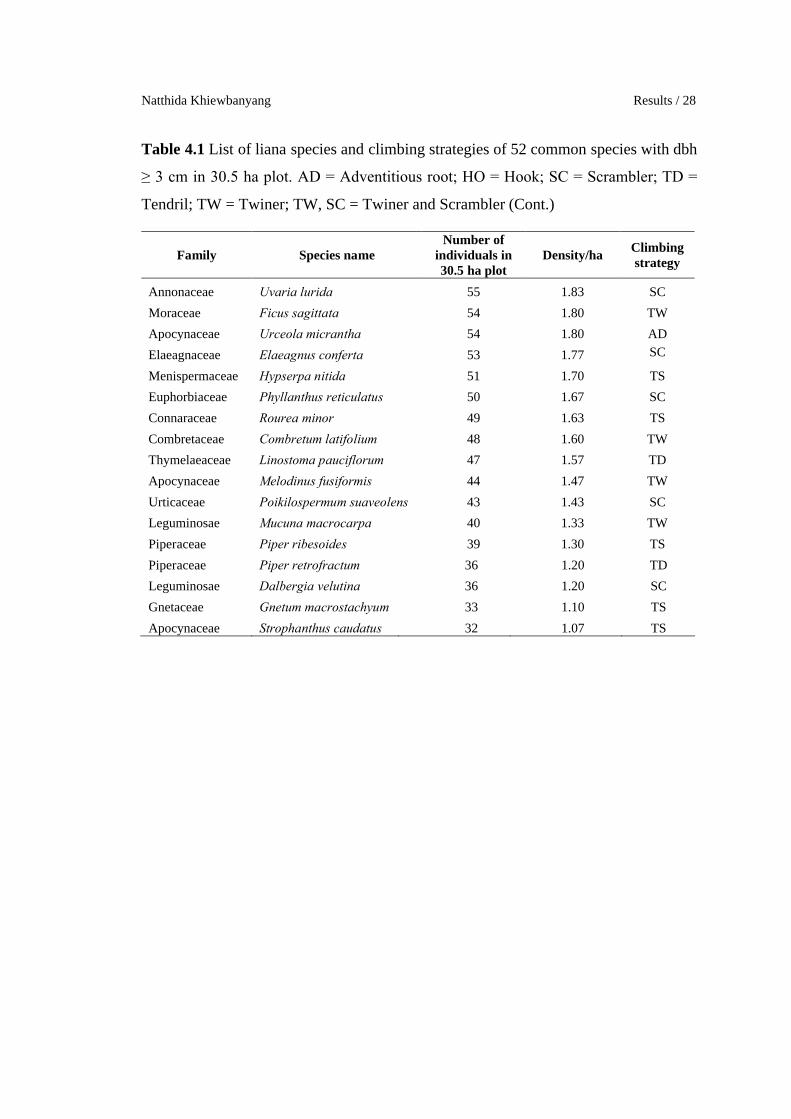
Natthida Khiewbanyang Results / 40
4.3.3 Relationships between liana climbing strategies and different host tree bark
textures
There were proportional differences between the three bark types (smooth,
slightly rough and rough) used by lianas. Ramets regardless of their climbing
strategies tended to use hosts with slightly rough bark rather than rough or smooth
bark (Table 4.4). Ramets mainly used host trees with slightly rough bark (61.54%) and
less frequently used other bark types; only 32.24% of hosts with rough bark and 6.22%
of hosts with smooth bark.
Liana ramets that used different climbing strategies were found to be
correlated with particular types of bark texture of host trees (Table 4.4). Host trees
with slightly rough bark were being used with the highest percentage by the number of
individual ramets. However, the 2 test verified that hook climbers tended to use host
trees with slightly rough bark of host trees more often than expected (2=11.52, df = 2,
p < 0.05). On the other hand, adventitious root climbers required host trees with rough
bark (2= 8.32, df = 2, p < 0.05). In contrast, ramets employing the other climbing
strategies were independent of host tree bark textures.
Table 4.4 Percentage of individual ramets and the percentage of individual ramets for
each climbing strategy belonging to different bark textures of host trees. AD =
Adventitious root; HO = Hook; SC = Scrambler; TD = Tendril; TW = Twiner; TS =
Twiner and Scrambler
Percentage of individual ramets for each
climbing strategyHost tree bark
texture
Percentage of
individual rametsTW SC TD HO AD TS
Smooth 6.22 1.85 1.73 0.44 0.12 0.51 1.54
Slightly rough 61.54 19.29 13.71 5.25 6.64 1.73 15.90
Rough 32.24 9.61 5.70 3.91 1.92 1.60 9.49





























