Embed Size (px)
Citation preview

Eur. J . Biochem. 71, 607-612 (1976)
Lipid Requirement of Membrane-Bound 3-Oxosteroid A4 - d5-Isomerase Studies on Beef Adrenocortical Microsomes
Philippe GEYNET, Christine DE PAILLERETS, and Annette ALFSEN
Labordtoire des Etats Lies Moleculaires, Equipe de Recherche 64 d u Centre National de la Recherche Scientifique, Unite d’Enseignement et de Recherches Biomedicale des Saint-Ptres, Universite de Paris V
(Received August 4/0ctober 6, 1976)
The role of phospholipid in the beef adrenal microsomal 3-oxosteroid A4 - A5-isomerase (EC 5.3.1 . I ) has been investigated with the use of phospholipase A to alter the microsomal phospho- lipids. The byproducts of phospholipase A digestion have been removed with a wash solution containing bovine serum albumin. Removal of 80- 85 of the phospholipid leads to loss of 80 - 90 7) of the 3-oxosteroid A4 - A’-isomerase activity. Reconstitution experiments have been performed by introduction of lipid aqueous dispersions in the enzymatic assay. Asolectin, a commercially available preparation of soy phosphatides, is able to stimulate the enzymatic activity but does not restore the 3-oxosteroid A4 - A5-isomerase activity in phospholipase-A-treated membranes. In contrast, the introduction of aqueous dispersions of microsomal total lipid mixtures in the enzymatic assay brings about a complete restoration of the 3-oxosteroid A4 - A’-isomerase activity in the lipid-depleted membranes. It is concluded that the bovine adrenal microsomal 3-oxosteroid A4 - A5-isomerase requires phospholipid(s) to exhibit its full catalytic activity.
The mammalian 3-oxosteroid A - A 5-isomerase concerned with the conversion of A 5-3-oxosteroids to the corresponding conjugated A 4-3-oxosteroids was first described by Talalay and Wang [l]. In mammalian endocrine organs, such as adrenals, ovary and testis this enzyme is always coupled with the NAD-dependent A 5-3P-hydroxysteroid dehydro- genase and is mainly associated with the microsomal fraction of the cell [2-61. During the past twenty years, the ultimate goal of a number of investigators has been the solubilization and purification of this enzyme system. However, until recently [7], no satis- factory purification had been achieved though nearly all the classical methods of solubilization of mem- brane-bound enzymes had been attempted [2,5,8 - 131. In the same time, an abundance of evidence has accumulated indications that the properties of many membrane-bound enzymes were closely dependent on interactions with membrane phospholipids [14- 221. Since the exact effects of the phospholipid environ- ment on the catalytic activity varied in different enzymes [23], an understanding of the relationship between the microsomal 3-oxosteroid A - A 5-is0-
Enzymes. 3-Oxosteroid d - A 5-isomerase (EC 5.3.3.1) ; A 5-3fl- hydroxysteroid dehydrogenase (EC 1.1.1.51); phospholipase A (EC 3.1.1.4).
merase activity and the phospholipid environment might be of importance for elucidating the significance of its membrane location, physiological regulation and mechanism of action. Moreover it was likely that the methods for purifying such a microsomal enzyme should ultimately depend on the interrelations of phospholipids and catalytic activity.
As a first step in resolving some of these problems, we have examined the effect of phospholipase A on the beef adrenal cortex microsomal3-oxosteroid A - A 5-
isomerase activity. We have been interested not only in the question of phospholipase-induced inactiva- tion, but also with a tentative reconstitution of the catalytic activity by two lipid mixtures. Preliminary reports on the phospholipase A treatment of the microsomal3-oxosteroid A - A ’-isomerase have been given [24,25].
MATERIALS AND METHODS
Materials
Silica gel H according to Stahl was from Merck, Darmstadt (FRG). Crude lyophilized Naju nuja snake venom was purchased from Koch-Light Laboratories Ltd (Colnbrook, Buckinghamshire, England). Phenyl- methylsulfonylfluoride (a potent inhibitor of protease)

608 Lipid Requirement of Membrane-Bound 3-Oxosteroid A - A '-Isomerase
was obtained from Serva-Feinbiochemica (Heidel- berg, FRG). Bovine serum albumin Pentex fraction V powder was from Miles Laboratories Inc. (Kankakee, Illinois, U.S.A.). Asolectin was obtained from As- sociated Concentrates, (Woodside, N.Y., U.S.A.). The 5-pregnene-3,20-dione was from Roussel-UCLAF (Romainville, France) and used as supplied (0- 3 %, 3-oxo-A4-steroid). All other reagents were of the highest grade commercially available.
The commercial asolectin has been reported by Richardson et al. [34] to have the following composi- tion : phosphatidylcholine 31 x, phosphatidylethanol- amine 26 x, phosphatidic acid 11 %, cardiolipin 4 %, phosphatidylinositol (or a phosphatide of the same RF) 18 %, unidentified phosphorus-containing material 11 x. Both amphoteric and acid phospholipids are represented in this mixture.
Prepparation of Microsomes
Microsomes were prepared exactly as described in a preceding paper [26] from fresh beef adrenal glands and stored in liquid nitrogen; no significant loss of activity was observed over 6 months. Protein was determined according to Lowry et al. [27] with crystalline ovalbumin as a standard.
Treatment of Microsomes tvith Phospholipase A
40mg of Naja naja snake venom were dissolved in 2 ml of 20 mM acetate buffer, pH 5.0, heated 5 to 10 min in a boiling water-bath, chilled and centrifuged 30 min at 105000 x g [14]. The supernatant was decanted and filtered at room temperature on a column of biogel PIOO equilibrated with 20mM Tris-HCI buffer, pH 7.5, 5 mM in CaC12. Phospholipase A activity was determined by acid production in the presence of cresol red [28]. In terms of the enzymatic assay, the phospholipase was 10-fold purified with a yield of 70%. Purified phospholipase A was stored at - 20 "C in presence of 1 ;<) (w/v) bovine serum albumin and was stable for many months.
Before treatment with phospholipase A, micro- somes (3.5 to 4 mg of protein) were pre-incubated 6 min at 35 "C in presence of 5 mM CaCI2, 0.2 mM phenylmethylsulfonylfluoride and 20 mM Tris-HCI buffer, pH 7.5 (final concentrations). Various amounts of phospholipase A were added to make a final volume of I ml and the reaction was allowed to proceed 30 min at 35 "C. A control sample without phospholipase A was always run simultaneously. Phospholipase A treatment of membranes results in the formation of byproducts which may interfere with lipid requiring enzymes [14,15,20]. They were re- moved by the following washing procedure : the reac- tion was terminated by dilution of the samples with 9 volumes of a cold 1 %, (w/v) bovine serum albumin
solution in 20 mM Tris-HCI buffer, pH 7.5 containing 0.25 M sucrose and 3 mM EDTA. The mixture was centrifuged at 105000 x g for 60 min. The residue was resuspended in 9 vol. of the same wash solution, using a teflon-glass homogenizer and the mixture was recentrifuged as before. This wash procedure was repeated a third time. A final wash in 9 vol. of 0.25 M sucrose was performed to remove excess serum albu- min. The preparation was resuspended in 0.25 M sucrose and stored in liquid nitrogen. All the opera- tions subsequent to the incubation with phospho- lipase A were carried out at 0-6 "C.
Analysis of Phospholipids in Microsomes
In order to determine the alterations in lipid con- tent and composition produced by the action of the phospholipase A, the treated microsomes were ex- tracted by the procedure of Folch et al. [29] modified by Rouser and Fleischer [30]. The phospholipid com- position was analysed by means of two-dimensional thin-layer chromatography on silica gel H, according to Nielsen et ul. [31]. Chloroform/methanoI/water (65/25/4, by vol.) was used in the first dimension and 1-butanol/acetic acid/water (60/20/20, by vol.) in the second. In this system phosphatidylinositol and phosphatidylserine migrated together. For quan- titative determinations, the lipid spots were scraped from the plate, eluted as described by Schiefer and Neuhoff [32] and their phosphorus content deter- mined by the micro-method of Ames and Dubin [33].
The ratio of the total lipid phosphorus content (pg) to the membrane protein content (mg) was defined as the 'bound-phosphorus' expressed in pg phosphorus per mg protein. The value of the 'bound- phosphorus' in the digested membranes was compared to the value found in the controls. The percentage of 'bound-phosphorus', after phospholipase A diges- tion, was taken as a measure of the destruction of the membrane phospholipids.
A queo us Lip id Dispersions
Microsomal total lipid mixtures were extracted under nitrogen from beef adrenal cortex microsomes by the method of Folch et al. [29] modified by Rouser and Fleischer [30]. Non-lipid material was removed by the Folch partition method [29].
Asolectin or microsomal total lipid mixtures were dispersed in a 20 mM Tris/acetic buffer, pH 8.0, 1 mM in EDTA, by ultrasonic irradiation under a nitrogen stream and cooling in an ice-bath. The sonic irradiation was performed by short bursts of 15 s, with intermittent cooling for 1 min, until clarity is obtained. The opalescent mixture was centrifuged at 15 000 x g for 30 min to remove large lipid aggregates and metal particles released from the tip during

P. Geynet, C. de Paillerets, and A. Alfsen 609
sonication. The dispersions were stored at 4 "C, under nitrogen. Thin-layer chromatography did not show any degradation product of the lipids. The phospho- lipid concentration was evaluated from phosphorus determinations [33].
3-0.uoster-oid A 4 - A5-Isomerase Assay
3-Oxosteroid A 4 - A5-isomerase activity was tested either in 20 mM Tris-HCI buffer, pH 8.5, 5 mM in CaC12 or in 0.1 M Tris-HC1 buffer, pH 8.5, 3 mM in EDTA, with the 5-pregnene-3,20-dione (dissolved in dioxane) as substrate. Kinetic measurements were recorded at room temperature either on a Gilford 2400 or on a Cary 118 spectrophotometer. Formation of progesterone was followed at its specific wave-length absorption: 248 nm, E = 17000 M-' . cm-'.
The final reaction volume was 3 ml in l-cm light- path cuvettes. Each reaction mixture contained 100 pl steroid solution (3.3 v/v organic solvent; 5-pregnene- 3,20-dione = 10.6pM or 21.2pM), 0- loop1 enzyme appropriately diluted in assay buffer. When restora- tion of the catalytic activity was attempted, 0-200 pl lipid dispersions were introduced into the assay mixture. In order to allow the lipid to interact with the enzyme, the assay mixture containing all the reactants but steroid was pre-incubated for 10 min before initiation of the reaction. A 10-min pre-incuba- tion time of the lipid and the enzyme was selected to obtain optimal activation.
In all cases two blank cuvettes were necessary for each enzymatic assay: the first one contained all reactants but enzyme and the other one all reactants but steroid. Measurements initiated with substrate were carried out in duplicate. It was verified by thin- layer chromatography that absorbance changes re- corded at 248 nm were indeed due to the formation of progesterone, as described earlier [26]. The activity was estimated from the initial linear change in ab- sorbance after correction for the absorbance changes of the blanks. One enzyme unit is defined as the amount of enzyme which generates 1 nmol product per min. Specific activity is expressed as nmol product formed per min per mg protein.
RESULTS AND DISCUSSION
Efjects of Phospholipase A
The presence of a possible proteolytic activity in the partially purified phospholipase A has been check- ed by treating microsomes with phospholipase A (1 32 mU/mg membrane protein) in presence of 10 mM EDTA and with omission of phenylmethylsulfonyl- fluoride : no calcium-independent proteolytic activity has been detected. However, in order to eliminate a possible interference of any protease contaminant,
experiments have been conducted in presence of 0.2 mM phenylmethylsulfonylfluoride.
Kinetic measurements have been recorded in the 20 mM Tris-HC1 buffer, pH 8.5, 5 mM in CaC12 with the following control experiments : The enzymatic activities of the phospholipase-A-treated membranes have been measured with 0 or 10 min pre-incubation time in the assay cuvette (no lipid introduced); in these conditions no variation of the 5-pregnene- 3,20-dione isomerase activity has been observed in the 10 min preincubation assay, indicating that pos- sible traces of adhering phospholipase A to the treated membranes, were not interfering.
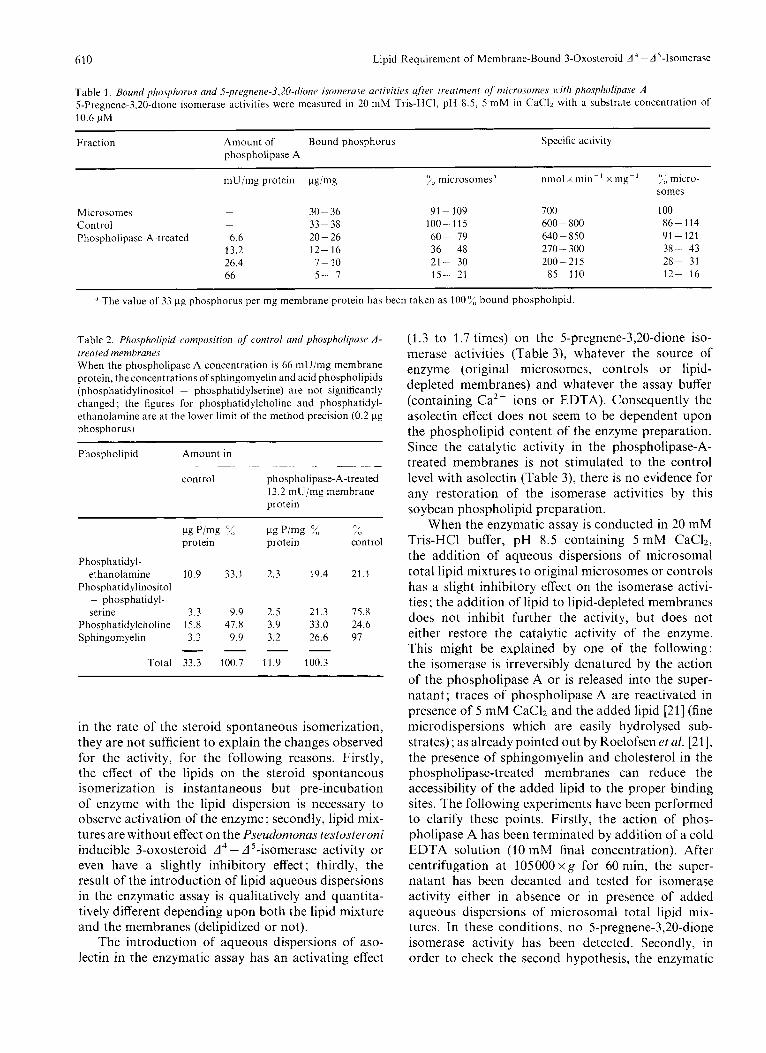
Treatment of the microsomes with phospholipase A influences the catalytic activity of the 3-oxosteroid A - A 5-isomerase, as shown in Table 1. Removal of 80-85%, of the phospholipids leads to 80-90?!, loss of isomerase activity.
Table2 shows the degree of breakdown of each of the four main phospholipid classes brought about by a phospholipase A digestion of 65% of the mem- brane phospholipid content. The phospholipid com- position of the digested membranes shows significantly reduced proportions of phosphatidylethanolamine and phosphatidylcholine. This table does not allow a quantitative appreciation of the phosphatidylserine hydrolysis since phosphatidylinositol and phospha- tidylserine migrate together. It should be noted that the fatty acids and lyso-compounds produced by the phospholipase A treatment are quantitatively removed during the wash procedure (no detectable traces by thin-layer chromatography).
Restoration of5- Pregnene-3,ZO-dione Isomer-ase Activity
Reconstitution experiments are carried out by the addition of aqueous lipid dispersions in the enzymatic assay. The introduction of lipids in the blank cuvette containing all reactants but steroid does not produce a variation in the absorbance at the wavelength of the enzymatic assay (248 nm); in contrast, in the 'blank' containing all the reactants but enzyme, it leads to an increase of the spontaneous rate of isomerization of the 5-pregnene-3,20-dione from 0.03 nmol of product formed per rnin to 0.5 nmol of product formed per min in the presence of 10.6 pM 5-pregnene-3,20-dione and 200 pg phospholipid added per 3 ml assay. Consequently the reported enzymatic activities are always corrected for the spontaneous rate of formation of progesterone. In fact, the intro- duction of lipid in the assay mixture is likely to change the substrate partition between the aqueous phase and the lipid phase, the effective microscopic concentration of the steroid and to modify the critical micelle concentration of the substrate. However, if these changes may be related to the observed increase
610 Lipid Requirement of Membrane-Bound 3-Oxosteroid d - A '-Isomerase
Table 1. Bound phosphorus and S-pre~nene-3,20-~iione isomerasc. acrivities ufier lreafmenl of' microsomes with phospholipase A 5-Pregnene-3,20-dione isomerase activities were measured in 20 mM Tris-HCI, pH 8.5, 5 mM in CaClz with a substrate concentration of 10.6 pM
Fraction Amount of Bound phosphorus Specific activity phospholipase A
mU/mg protein pg/mg :<, microsomesa nmol x min- ' x my- ' :,; micro- s o m a
Microsomes - 30 - 36 91 - 109 700 100
Phospholipase- A-treated 6.6 20-26 60- 79 640 - 850 91 - 121 Control - 33-38 100-115 600 - 800 86- 114
13.2 12- 16 36- 48 270 - 300 38- 43 26.4 7- 10 21- 30 200-215 28- 31 66 5- I 15- 21 85-110 12- 16
-~
The value of 33 pg phosphorus per mg membrane protein has been taken as 100% bound phospholipid.
Table 2. Plinspholipid composition of control and phospholipasr A - treated membranes When the phospholipase A concentration is 66 mU/mg membrane protein, the concentrations of sphingomyelin and acid phospholipids (phosphatidylinositol + phosphatidylserine) are not significantly changed ; the figures for phosphatidylcholine and phosphatidyl- ethanolamine are at the lower limit of the method precision (0.2 pg phosphorus)
Phospholipid Amount in . - - ~ ~.
control phospholipase-A-treated 13.2 mU/mg membrane protein
I-lg P/mg '%, c1g P/mg 2, XI
protein protein control
Phosphatidyl- ethanolamine 10.9 33.1 2.3 19.4 21.1
Phosphatidylinositol + phosphatidyl- serine 3.3 9.9 2.5 21.3 75.8
Phosphatidylcholine 15.8 47.8 3.9 33.0 24.6 Sphingom yelin 3.3 9.9 3.2 26.6 97 - - - -
Total 33.3 100.7 11.9 100.3
in the rate of the steroid spontaneous isomerization, they are not sufficient to explain the changes observed for the activity, for the following reasons. Firstly, the effect of the lipids on the steroid spontaneous isomerization is instantaneous but pre-incubation of enzyme with the lipid dispersion is necessary to observe activation of the enzyme; secondly, lipid mix- tures are without effect on the Pseudomonas testosteroni inducible 3-oxosteroid A 4 - A 5-isomerase activity or even have a slightly irihibitory effect; thirdly, the result of the introduction of lipid aqueous dispersions in the enzymatic assay is qualitatively and quantita- tively different depending upon both the lipid mixture and the membranes (delipidized or not).
The introduction of aqueous dispersions of aso- lectin in the enzymatic assay has an activating effect
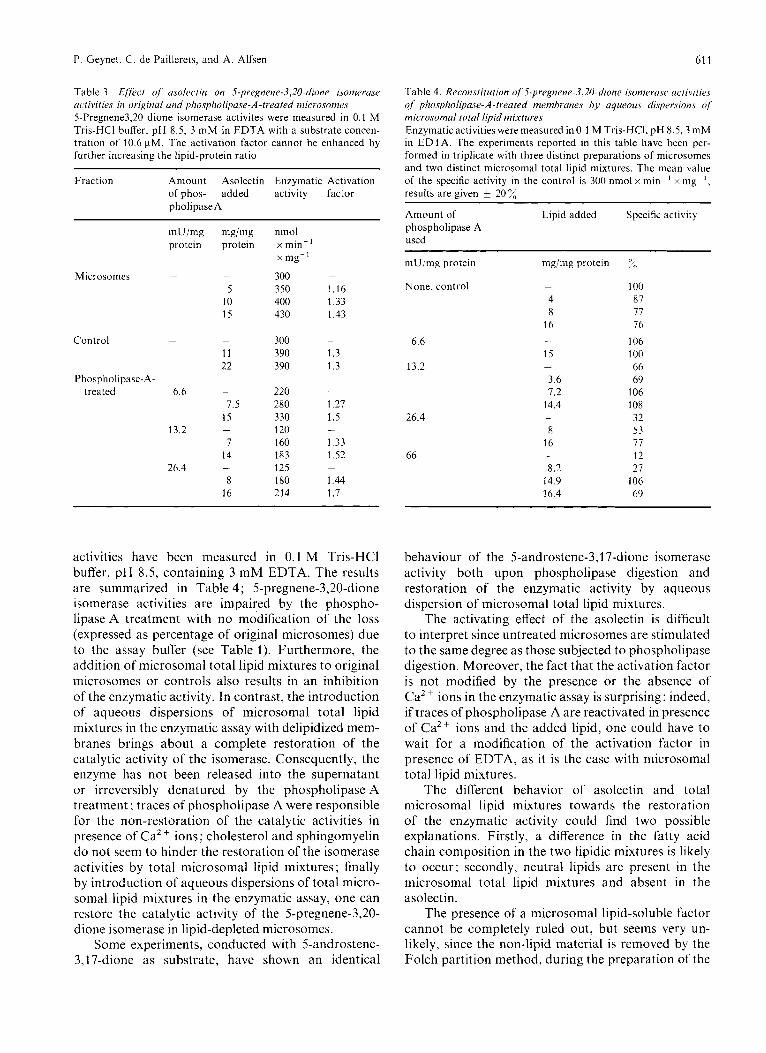
(1.3 to 1.7 times) on the 5-pregnene-3,20-dione iso- merase activities (Table 3), whatever the source of enzyme (original microsomes, controls or lipid- depleted membranes) and whatever the assay buffer (containing Ca2+ ions or EDTA). Consequently the asolectin effect does not seem to be dependent upon the phospholipid content of the enzyme preparation. Since the catalytic activity in the phospho1ipase-A- treated membranes is not stimulated to the control level with asolectin (Table 3), there is no evidence for any restoration of the isomerase activities by this soybean phospholipid preparation.
When the enzymatic assay is conducted in 20 mM Tris-HC1 buffer, pH 8.5 containing 5 mM CaCI2, the addition of aqueous dispersions of microsomal total lipid mixtures to original microsomes or controls has a slight inhibitory effect on the isomerase activi- ties; the addition of lipid to lipid-depleted membranes does not inhibit further the activity, but does not either restore the catalytic activity of the enzyme. This might be explained by one of the following: the isomerase is irreversibly denatured by the action of the phospholipase A or is released into the super- natant; traces of phospholipase A are reactivated in presence of 5 mM CaC12 and the added lipid [21] (fine microdispersions which are easily hydrolysed sub- strates) ; as already pointed out by Roelofsen et aI. [21], the presence of sphingomyelin and cholesterol in the phospholipase-treated membranes can reduce the accessibility of the added lipid to the proper binding sites. The following experiments have been performed to clarify these points. Firstly, the action of phos- pholipase A has been terminated by addition of a cold EDTA solution (10 mM final concentration). After centrifugation at 105000 x g for 60 min, the super- natant has been decanted and tested for isomerase activity either in absence or in presence of added aqueous dispersions of microsomal total lipid mix- tures. In these conditions, no 5-pregnene-3,20-dione isomerase activity has been detected. Secondly, in order to check the second hypothesis, the enzymatic
P. Geynet, C. de Paillerets, and A. Alfsen 61 1
Table 3. Effect of usolectin on S-pregnene-3,20-~li(~ne isomerase activities in original and phospholipase-A-treated microsomes 5-Pregnene3,20 dione isomerase activites were measured in 0.1 M Tris-HCI buKer, pH 8.5, 3 mM in EDTA with a substrate concen- tration of 10.6 pM. The activation factor cannot be enhanced by further increasing the lipid-protein ratio
Fraction Amount Asolectin Enzymatic Activation of phos- added activity factor pholipase A
Microsomcs -
Control -
Phospholipase-A- treated 6.6
13.2
26.4
mg/mg protein
-
5 10 15
-
11 22
-
7.5 15
1 14
8 16
-
-
nmol x min-' x mg-' 300 -
350 1.16 400 1.33 430 1.43
300 -
390 1.3 390 1.3
- 220 280 1.27 3 30 1.5 120 -
160 1.33 183 1.52 125 180 1.44 214 1.7
-
activities have been measured in 0.1 M Tris-HC1 buffer, pH 8.5, containing 3 mM EDTA. The results are summarized in Table 4; 5-pregnene-3,20-dione isomerase activities are impaired by the phospho- lipase A treatment with no mo'dification of the loss (expressed as percentage of original microsomes) due to the assay buffer (see Table 1). Furthermore, the addition of microsomal total lipid mixtures to original microsomes or controls also results in an inhibition of the enzymatic activity. In contrast, the introduction of aqueous dispersions of microsomal total lipid mixtures in the enzymatic assay with delipidized mem- branes brings about a complete restoration of the catalytic activity of the isomerase. Consequently, the enzyme has not been released into the supernatant or irreversibly denatured by the phospholipase A treatment ; traces of phospholipase A were responsible for the non-restoration of the catalytic activities in presence of Ca2 + ions ; cholesterol and sphingomyelin do not seem to hinder the restoration of the isomerase activities by total microsomal lipid mixtures ; finally by introduction of aqueous dispersions of total micro- soma1 lipid mixtures in the enzymatic assay, one can restore the catalytic activity of the 5-pregnene-3,20- dione isomerase in lipid-depleted microsomes.
Some experiments, conducted with 5-androstene- 3,17-dione as substrate, have shown an identical
Table 4. Reconstitution of 5-pregnene-3,20-dione i.sonieru.se activities .f phospholipase-A-treated membranes by aqueous dispecsions of microsomal total lipid mixtures Enzymatic activities were measured in 0.1 M Tris-HCI, pH 8.5,3 mM in EDTA. The experiments reported in this table have been per- formed in triplicate with three distinct preparations of microsomes and two distinct microsomal total lipid mixtures. The mean value of the specific activity in the control is 300 nmol x min-' x my-', results are given 20 X
~~ ~
Amount of Lipid added Specific activity phospholipase A used
mU/mg protein mg/mg protein y<)
None, control - 100 4 87 8 77
16 16 106
15 100 66
3.6 69 7.2 106
14.4 108 32
8 53 16 77
12 8.2 21
14.9 106 16.4 69
- 6.6
13.2 -
26.4 -
66 -
behaviour of the 5-androstene-3,17-dione isomerase activity both upon phospholipase digestion and restoration of the enzymatic activity by aqueous dispersion of microsomal total lipid mixtures.
The activating effect of the asolectin is difficult to interpret since untreated microsomes are stimulated to the same degree as those subjected to phospholipase digestion. Moreover, the fact that the activation factor is not modified by the presence or the absence of Ca2+ ions in the enzymatic assay is surprising: indeed, if traces of phospholipase A are reactivated in presence of Ca2+ ions and the added lipid, one could have to wait for a modification of the activation factor in presence of EDTA, as it is the case with microsomal total lipid mixtures.
The different behavior of asolectin and total microsomal lipid mixtures towards the restoration of the enzymatic activity could find two possible explanations. Firstly, a difference in the fatty acid chain composition in the two lipidic mixtures is likely to occur; secondly, neutral lipids are present in the microsomal total lipid mixtures and absent in the asolectin.
The presence of a microsomal lipid-soluble factor cannot be completely ruled out, but seems very un- likely, since the non-lipid material is removed by the Folch partition method, during the preparation of the

612 P. Geynet, C. de Paillerets, and A. Alfsen: Lipid Requirement of Membrane-Bound 3-Oxosteroid A 4 - A5-Isomerase
microsomal total lipid mixtures. Moreover, the pres- ence of an unknown product has never been detected by thin-layer chromatography.
In view of the results, one can conclude that the beef adrenal cortex 3-oxosteroid A 4 - A '-isomerase from microsomes requires lipid(s) to exhibit its full catalytic activity. The results strongly suggest that the enzyme is specifically requiring for microsomal lipid(s).
A major consequence of this work is the following: our experiments predict the impossibility of getting a purified, still biologically active beef adrenal cortex microsomal 3-oxosteroid A 4 - A '-isomerase which would be devoid of lipid(s). This is not in contradic- tion with the fact that Ford and Engel [7] have succeeded in purifying the A 5-3/3-hydroxysteroid de- hydrogenase-A '-3-oxosteroid isomerase system from sheep adrenocortical microsomes to near homo- geneity, for the following reasons. The purified ma- terial retains biological activity, but the authors do not exclude a partial inactivation of the enzymes caused by the extraction with the non-ionic detergent TritonXl00; the authors do not give any information about the lipid content of their purified material; though it is likely that the 3-oxosteroid A 4 - A 5 - isomerase from other mammalian adrenocortical mi- crosomes should also be lipid-dependent, the possibility of a different interrelation, between the enzyme and the microsomal lipids, is not excluded.
This work was supported by grants from the C.N.R.S. The authors are most grateful to D r H. Weintraub for donating pure 3-oxosteroid A 4 - A5-isomerase from Pseudomonas testosteroni; we also wish to thank Prof. CI. Burstein for his continuing interest and helpful discussions during the course of this work.
REFERENCES
1. Talalay, P. & Wang, V. S. (1955) Biochim. Biophys. Acta, 18,
2. Beyer, K. F. & Samuels, L. T. (1956) J . Biol. Chc.m. 219,
3. Kruskemper, H. L., Forchielli, E. & Ringold, H. J. (1964)
4. Oleinick, N. L. & Koritz, S. B. (1966) Biochemistry, 5 , 3400-
5. Neville, A. M. & Engel, L. L. (1968) Endocrinology, 83, 864-
300- 301.
69 - 76.
Steroids, 3, 295 - 309.
3405.
872.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18. 19.
20.
21.
22
23 24
25
26 27
28
29
30
31
32
33.
34.
Murota, S., Fenselau, C. C. & Talalay, P. (1971) Steroids, 17,
Ford, H. C. & Engel, L. L. (1974) J . Bid. Chem. 249, 1363-
Cheatum, S. G. & Warren, J . C. (1966) Biochim. Biophys.
Koide, S . S . & Torres, M. T. (1965) Biochim. Biopliys. Acfa,
Hamilton, M. A,, McCune, R. W. & Roberts, S. (1972) J .
Ewald, W., Werbin, H. & Chaikoff, I . L. (1964) Steroids, 4,
Geynet, P., Gallay, J. & Alfsen, A. (1972) Eur. J . Biochem. 31,
Cheatum, S . G., Douville, A. W. & Warren, J . C. (1967) Bio-
Fleischer, S. & Fleischer, B. (1967) Meth0d.s Enzynd. 10,
Duttera, S . M., Byrne, W. L. & Ganoza, M. C. (1968) J . B id .
Burstein, C. , Loyter, A. & Racker, E. (1971) J . Biol. Chem. 246,
Burstein, C. , Kandrach, A. & Racker, E. (1971) J . B id . Chem.
Razin, S . (1972) Biochim. Biophys. Acta, 265, 241 - 296. Vessey, D. A. & Zakim, D. (1972) J . Biol. Chem. 247, 3023-
Meissner, G. & Fleischer, S. (1972) Biochim. Biophys. Acta,
Roelofsen, B. & Van Deenen, L. L. M. (1973) Eur. J . Biochem.
Martonosi, A., Lagwinska, E. & Oliver, M. (1974) Ann. N . Y .
Hallinan, T. (1974) Bioclzem. So<. Trans. 2, 817-821. Geynet, P., Nicot, C. & Alfsen, A. (1973) Ahstr. pap. IXth Znt.
Geynet, P., de Paillerets, C., Nicot, C. & Alfsen, A. (1974)
Geynet, P. (1976) Biochim. Biophys. Actu, in press. Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J .
(1 968) Boehringer Biochemica Catalogue, Phospholipasr A a.s.suy,
Folch, J., Lees, M. & Sloane-Stanley, G . H. (1957) J . Biol.
Rouser, G. & Fleischer, S. (1967) Methods Enzymol. 10, 385-
Nielsen, N. C., Fleischer, S. & McConnell, D. G. (1970) Bio-
Schieffer, H. G. & Neuhoff, V. (1971) Hoppe-Sejder'sZ. Plzysiol.
Ames, B. N. & Dubin, D. T. (1960) J . B i d . Chem. 235, 769-
Richardson, S., Hultin, H. D . & Fleischer, S. (1964) Arch.
25-37.
1369.
Acta, 122, 1 - 13.
105, 115-120.
Endocrinol. 54, 297 - 31 5.
759 - 775.
464-469.
chim. Biophys. Acta, 137, 172- 178.
406-433.
Chem. 243,2216- 2228.
4075 - 4082.
246, 4083 - 4089.
3028.
255, 19 - 33.
40,245 - 257.
Acad. Sci. 227, 549-567.
Congr. Biochem. abstract 5n 14.
J . Steroid Biochem. 5, 310-311.
(1951) J . Biol. Chem. 193, 265-275.
Boehringer GmbH, Mannheim.
Chem. 226,497 - 509.
406.
chim. Biophys. Acta, 211, 10- 19.
Chem. 352,913-926.
775.
Biochem. Biophys. 105,254- 260.
P. Geynet, C. de Paillerets, and A. Alfsen, Laboratoire des Etats Lies Moleculaires, Equipe de Recherche 64 du C.N.R.S., U.E.R. Biomedicale des Saint-PZres, Universiti. de Paris V, 45 Rue des Saints-Peres, F-75270 Paris-Cedex-06, France





























