Embed Size (px)
Citation preview

MECHANISMS CONTROLLING PIGMENT MOVEMENTS WITHIN SWORDTAIL
(XIPHOPHOPRUS HELLERI) ERYTHROPHORES IN PRIMARY CELL CULTURE
JIRO MATWMOTO, YOSHIKO WATANABE and MASATAKA OBIKA
Department of Biology, Keio University, Yokohama 223, Japan
and
MAC E. HADLEY Department of Cellular and Developmental Biology, University of Arizona, Tucson, AZ 85721, U.S.A.
(Received 21 October 1977)
Abstract-l. Pigment organelle (pterinosomes and carotenoid vesicles) movements within isolated sword- tail (Xiphophorus helleri) erythrophores cultured in vitro were studied.
2. Pterinosome and carotenoid vesicles migrate centripetally to a juxtanuclear position or centrifugally into the dendritic processes of erythrophores in response to catecholamines.
3. Catecholamine stimulation of alpha adrenoceptors results in centripetal pigment organelle migra- tion whereas hem adrenoceptors mediate centrifugal movements.
4. Melatonin also induces perinuctear aggregation of pigment otganeiles but by receptor mechanisms separate from adrenoceptors.
5. MSH and ACTH stimulate pigment organelle dispersion. Centrifugal dispersion of erythrophore pigments is also stimulated by cyclic AMP and dibutyryl cyclic AMP.
INTRODUCTION
Rapid intracellular pigment movements are con- sidered to be a dynamic property of integumental chromatophores of poikilotherm vertebrates. Much research has been directed toward elucidation of this phenomenon, mostly employing dermat melano- phores as a model (Parker, 1948; Waring, 1963; Bag- nara & Hadley, 1973). Recently, information has been provided that the yellow and red pigment cells, xan- thophores and erythrophores, found in’ the dermis of some poikilotherms are also capable of translocating their pigment granules in response to specific stimula- tory cues (Smith & Smith, 1935; Bagnara & Hadley, 1969; Sage, 1970; Porter, 1973; Horikawa & Matsu- moto, 1974; Castrucci, 1975; Ozato, 1976, 1977; Byers & Porter, 1976; Junqueira & Farias, 1976).
The pigments of xanthophores and erythrophores are comprised of pteridine and carotenoid derivatives which are, therefore, quite different in chemical properties from melanin, the major pigmentary com- ponent of vertebrate meIanophores (Bagnara & Had- ley, 1973). It is well established that pteridines are localized in specific cytoplasmic organelles called pterinosomes whereas carotenoids exist in association with endoplasmic reticulum and related small vesicles (Matsumoto, 1965; Matsumoto & Obika, 1968; Obika & Matsumoto, 1968). Irrespective of different pigmentary composition, the size and light micro- scopic appearance of pterinosomes of fish erythro- phores are similar to those of melanosomes present in vertebrate melanophores (Matsumoto, 1965; Mat- sumoto & Obika, 1968). We considered it of particu-
lar interest, therefore, to determine whether the mech- anisms controlling pterinosome movements in these pigment cells are similar or not to those of melano- some migration within melanophores. In this study we were concerned with determining the nature of the chemical messengers (hormones, neurotrans- mitters) affecting pigment organelle movements within erythrophores and the receptor mechanisms involved in such in~a~llular translocations. We stud- ied the erythrophores of the teleost fish, Xiphophorus helleri (swordtail), under both in vitro tissue culture and in situ isolated scale preparations.
MATERIALS AND METHODS
Examination of isolated scales
The swordtail fish, Xiphophorus helleri, used in this study were a progeny of an inbred, homogeneously red-colored strain maintained in the authors’ laboratory. One and a half to 3 month old fish having a body length of 2.0-3.0 cm were routinely utiiized. For comparison with cultured materials, the response of in siru erythrophores was exam- ined by use of scales detached from the middorsal surface of fish. Erythrophores of isolated scales aggregate upon treatment with 130 mM potassium chloride solution for about 1 hr after removal, thus suggesting that they are in- nervated in uiuo (Fujii, 1959). Therefore, the effects of vary- ing drugs and hormones on in uiuo erythrophore prep- arations were examined utilizing scales subjected to dener- vation treatment, which involves leaving freshly isolated scales in physiological salt solution (Yamamoto, 1941) for 1) hr or longer at room temperature. Aggregation and dis- persion of cytoplasmic pigments within erythrophores were

510 JIRO ~ATSliMoTO PI d.
observed by placing isolated scales on a glass slide under a light microscope.
Fish were rinsed in 70”., ethanol for 15 set and then twice in Cal’-free sterile physiological salt solution for a few minutes and finally in the same sterile solution sup- plemented with IO”,; fetal calf serum (Grand Island Bio- logical Company). Skin and scales were then detached from the fish using a pair of sharp pointed tweezers and collected in the dissociation medium. In most cases, several litter-mate fish were utilized in each series of experiments. Enzymatic digestion of skin tissues was carried out in two steps. first. solely with collagenase (Type 1, Worthington Biochemical Co.. NJ). and then with collagenase and tryp- sin (D~fco I : 250). Collagenase was used at a concentration of 0.Z; by suspending it in the dissociation medium. After 3Omin incubation with the enzyme, trypsin suspended in a small amount of medium was added so as to make a final concentration of 1.0”;. and the skins or scales were subjected to further digestion for 15 min. All incubations were performed at 25°C with continuous swirling. At the end of the incubation. the tissue suspension was filtered
through a stainless mesh strainer (15Opm aperature) in order to remove undigested scales and tissues. Dissociated cells were spun down at 200g for 3 min and then resus- pended in a small amount of the dissociation medium.
The majority of dissociated erythrophores appeared as individual small red spheres whereas others formed rela- tively large spherical aggregates. Non-pigment cells (present in larger numbers than erythrophores) and tissue debris were included in the suspendion. The sorting and subsequent settling of dissociated erythrophores was car- ried out either by manipulating an individual cell or by use of differential adhesion to a substratum. In the former procedure. the erythrophores scattered in a relatively larger amount of the dissociation medium were collected under a binocular microscope using a microtransfer pipette. In the latter procedure, cell suspensions were prepared so as to contain about 3000 erythrophores per ml of the dissoci- ation medium and then placed. one drop per dish. on to the substrate in the culture medium. Since most erythro- phores anchored firmly on the substratum within I$ hr after plating. the cultures were then washed with moderate agitation in fresh culture medium to remove the less rapidly attaching contaminating non-pigment cells. A reconstituted collagen film and Falcon plates were utilized as substrates. The former was prepared on IOmm glass cover slips marked with an address guide, according to the method described hy Ehrmann & Gcy (1956). Before the seeding of erythrophores. the film thus prepared was rinsed. with several changes, in sterile distilled water for 1 week and then in physiological salt solution for 2 days at 4‘C. Though erythrophores settled on the two different substrates employed. their cell shape established on recon- stituted collagen film appeared to be morphologi~dlly more natural than that on Falcon plates. Little difference was observed between erythrophores settled on reconstituted collagen and those on Falcon plates as to their pigment- migratory response.
The culture medium utilized in the present study was composed of seven parts of L-15 [Leibovib (1963), Grand Island Biochemical Company] solution, one part redis- tilled water, two parts fetal calf serum. 100 units per ml of penicillin (Penicillin G potassium) and 1OOpg per ml of streptomycin (Meiji, Tokyo). Erythrophores were culti- vated in 35 x IOmm style Falcon plastic culture dishes containing 2.5 ml of the culture medium at 25°C using air as a gas phase. The medium was renewed every third day.
Based on preliminary observations that most erythro- phores became fully dendritic on the third to fourth day
after the begining of culture, indicating firm settlement on a substratum, all experimental examinations were made using the 4-day old cultures unless otherwise specified. For experiments in which aggregation and dispersion of pig- ment organelles were repeatedly examined. the erythro- phores cultured on reconstituted collagen were used because of their stable attachment as compared to those on Falcon plates. The address guide marked on a cover slip facilitated identification of specific cells in repeated observations. The changes in response to epinephrine dur- ing culti~tion were examined using the cells cultured di- rectly on Falcon plates.
Since erythrophores in culture exhibited a wide variation in cell shape and since they translocated their pigments fairly rapidly without presenting clear images of an inter- mediate phase, the degree of their pigment aggregation and dispersion could not be adequately expressed by an index as used for melanophores. Therefore, the effects were visually assessed. and based on the time required to reach complete aggregation or dispersion of pigment organelles.
The followmg agents were examined for their possible influences on pigment movements: I-epinephrine (Merck, Darmstadt); ~-norepinephrine (Sankyo, Tokyo); theophyl- line and dibenamine h~ldrochloride (Tokyo Chemical fn- dustry, Osaka); propranolol hydrochIoride (Sumitomo Chemical Industry. Osaka); isoproterenol bitartrate (Winthrop Laboratories, New York); acetylcholine chlor- ide. melatonin, adenosine-3’,5’-cyclic monophosphoric acid, sodium salt (cyclic AMP), N6, Oz-dibutyryl adeno- sine-3’S’-cyclic monophosphoric acid, sodium salt (dibu- tyryl cyclic AMP), guanosine-3’.5’-cyclic monophosphoric acid, sbdium salt (cyclic GMP), porcine adren&orticotro- Die hormone (ACTH). (Sinma Chemical. St. Louis): and ulpha . melandcyte-stim;la%ng hormone (MSH) iCiba Geigy, Base]). The pH of the incubation medium, when changed by addition of the chemicals. was readjusted with a small amount of 0.1 N HCI or NaOH hydroxide. ACTH was treated in alkaline medium over boiling water for 5 min in order to intensify MSH-like activity (Bagnara, 19581.
RESULTS
Erythrophorr morphology in primary cdl culturr
In culture, three types of bright-colored pigment cells were observed which differed in morphology and pigmentation : the first type was large in size, approxi- mately 150 pm in width, and consisted of red pigment granules (pterinosomes) and yellow carotenoid vesi- cles which were located diffusely throughout the cyto- plasm. The second type was smaller in size, about 60pm, and similar in pigmentation to the first type. The third type. similar in size to the second but very few in number, was yellow in color. indicating a com- plete absence of red pterinosomes. The first and second types were tentatively identified as corre- sponding to so-called macro- and micro-erythro- phores, respectively [see terminology of Goodrich et al. (1941)). The third type was apparently identica1 to the xanthophores which are abundant in the yel- low-colored mutant. Because of the overwhelmingly larger numbers present, most all examinations were carried out with the macro-erythrophores.
All erythrophores in the 4-day old cultures had well expanded dendritic shapes similar to those seen in freshly isolated scales placed in physiological salt solution {Fig. la). The morphology of these cultured erythrophores varied greatly. Their shape appeared
Control of pigment movements within swordtail erythrophores 511
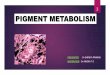
Figs. l-5. Photomicrographs showing swordtail erythrophores in primary cell culture.
Fig. I. (a) General view of a 4-day old culture. Erythrophores have settled on a reconstituted collagen sheet. All erythrophores are in a dispersed state. (b) The same cells treated with 5 x 10~“ M cpmephrine.
All pterinosomes are perinuclearly aggregated. x I 15.
to be more diverse in culture as compared to in situ erythrophores. Within the erythrophores maintained in the normal culture medium, numerous red pterino- somes were distributed homogeneously throughout the entire cytoplasm. Frequently, a central portion of each cell was occupied exclusively with yellow carotenoid pigments as seen in irl situ erythrophores.
Receptors controiling centripetul pigment migration
The administration of epinephrine (5 x 10m4 M) or norepinephrine (5 x 10e4 M) to the cultured erythro- phores brought about rapid centripetal displacement of pterinosomes (Fig. lb). This response was similar to that of erythrophores in situ. Preincubation of cul- tured erythrophores with dibenamine (10e4 M) for 30-50 min completely blocked pigment aggregation
in response to epinephrine and notepinephrine. Iso- proterenol (lo-“-IO-* M) also caused pterinosome aggregation within erythrophores. Acetylcholine (1O-3--1O-s M) was without observable affects on pig- ment movements within erythrophores.
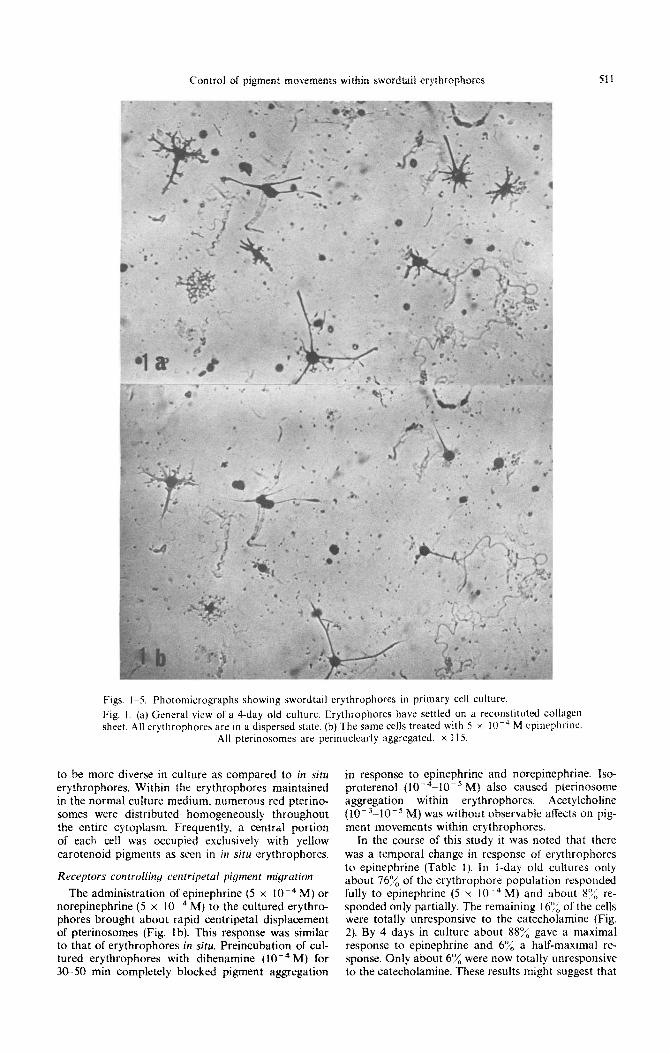
In the course of this study it was noted that there was a temporal change in response of erythrophores to epinephrine (Table I). In l-day old cultures only about 760/;; of the erythrophore population responded fully to epinephrine (5 x 10e4 M) and about H’J< re- sponded only partially. The remaining 16% of the cells were totally unresponsive to the catecholamine (Fig. 2). By 4 days in culture about SSu/;, gave a maximal response to epinephrine and 6”/, a half-maximal re- sponse. Only about 6% were now totally unresponsive to the catecholamine. These resuits might suggest that
512 JIRO MATSUMOTO et ul.
Fig. 2. Appearance of erythrophores showing no response or only half aggregation in response to epinephrine in a l-day old culture. (a) and (c): in the normal medium. (b) and (d): in the presence of epinephrine (5 x 10m4 M). Note the presence of erythrophores insensitive to this hormone (b. arrow) or responding incompletely (d, arrow). These insensitive erythrophores contrast to neighboring cells which exhibit a complete aggregation of pterinosomes in response to epinephrine. (a) and (b). as well
as (c) and (d), indicate the same cells under control and experimental conditions. x 365.
trypsin or other treatments of the cells during prep- tonin did not enhance the response of the erythro- aration for culture may have resulted in destruction phores to this hormone and some pigment cells failed or loss of membrane adrenoceptors. to respond at all.
Melatonin (lo-‘M) also induced aggregation of pterinosomes in the erythrophores both in culture and in situ. This response was not inhibited by dibenamine as was that of norepinephrine and epinephrine. When melatonin ( 10e4 M) was added to 4-day old cultures, there was a variable response to this indoleamine (Table 1). A response of incomplete (half) aggregation was identified by an appearance of a loosely packed perinuclear pigment cluster. In- creased concentrations (up to 5 x lo- 3 M) of mela-
Receptors controlling centrijugul pigment migration
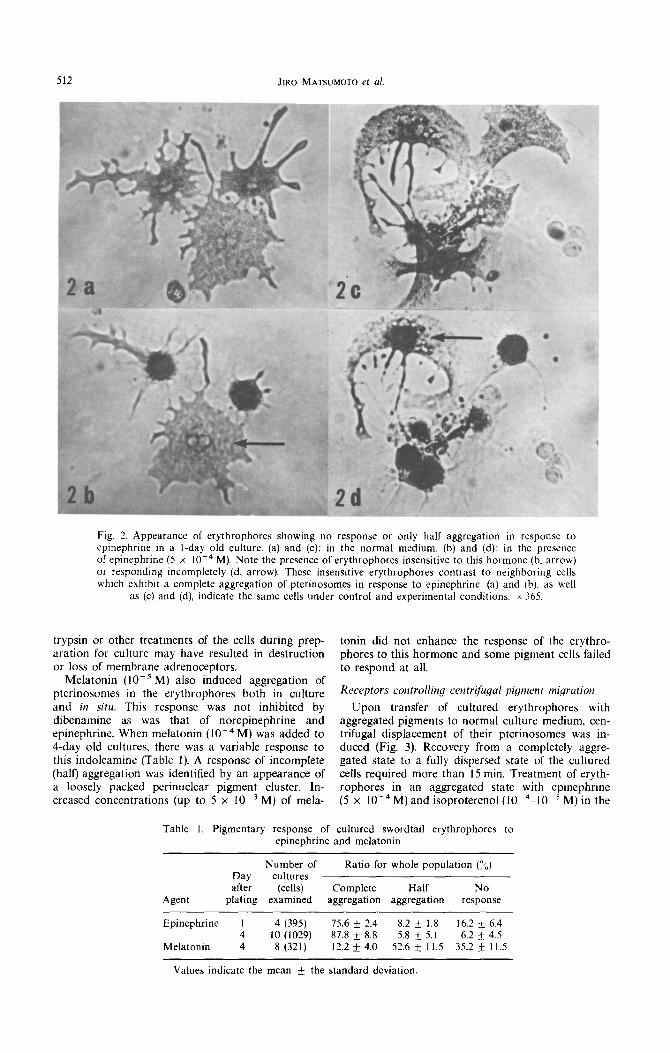
Upon transfer of cultured erythrophores with aggregated pigments to normal culture medium, cen- trifugal displacement of their pterinosomes was in- duced (Fig. 3). Recovery from a completely aggre- gated state to a fully dispersed state of the cultured cells required more than 15 min. Treatment of eryth- rophores in an aggregated state with epinephrine (5 x 10e4M) and isoproterenol (10m4-10-5 M) in the
Table I. Pigmentary response of cultured swordtail erythrophores to epinephrine and melatonin
Agent
Number of Ratio for whole population (yO) Day cultures after (cells) Complete Half No
plating examined aggregation aggregation response
Epinephrine 1 4 (395) 15.6 * 2.4 8.2 f 1.8 16.2 + 6.4 4 10 (1029) 87.8 k 8.8 5.8 rt 5.1 6.2 If: 4.5
Melatonin 4 8 (321) 12.2 + 4.0 52.6 + Il.5 35.2 f 11.5
Values indicate the mean k the standard deviation.
Control of pigment movements within swordtail erythrophores 513
Fig. 3. (a) Erythrophores treated with epinephrine (5 x 10e4M) for IOmin. (b) Ten minutes after the epinephrine-treated cells were transferred into control medium. x 365.
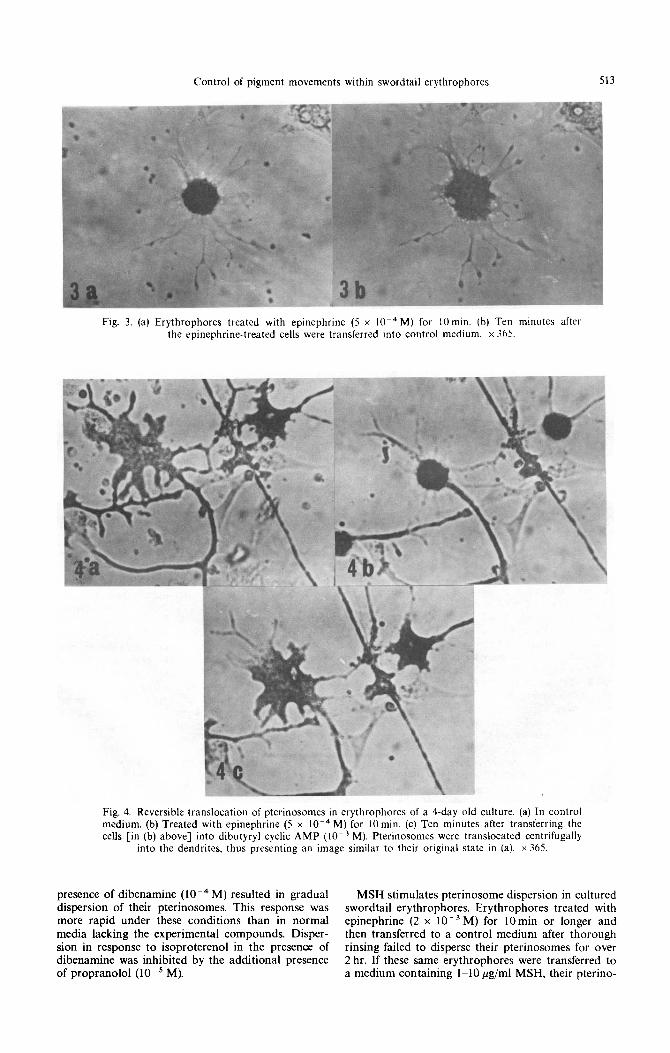
Fig. 4. Reversible translocation of pterinosomes in erythrophores of a Cday old culture. (a) In control medium. (b) Treated with epinephrine (5 x 10m4 M) for 10 min. (c) Ten minutes after transferring the cells [in (b) above] into dibutyryl cyclic AMP (10e3 M). Pterinosomes were translocated centrifugally
into the dendrites. thus presenting an image similar to their original state in (a). x 365.
presence of dibenamine (10m4 M) resulted in gradual MSH stimulates pterinosome dispersion in cultured dispersion of their pterinosomes. This response was swordtail erythrophores. Erythrophores treated with more rapid under these conditions than in normal epinephrine (2 x 10e3 M) for 1Omin or longer and media lacking the experimental compounds. Disper- then transferred to a control medium after thorough sion in response to isoproterenol in the presence of rinsing failed to disperse their pterinosomes for over dibenamine was inhibited by the additional presence 2 hr. If these same erythrophores were transferred to of propranolol (lo-’ M). a medium containing l-10 pg/ml MSH, their pterino-

somes rapidly dispersed. Similar results were obtained using ACTH (alkali-treated, l-10 &ml), theophylhne (10V3 M), cyclic AMP (IO-’ M) and dibutyryl cyclic AMP (10m3 M) (Fig. 4). Cyclic GMP failed to disperse pterinosomes within erythrophores previously aggre- gatcd by epinephrine.
Coinrnon receptors for movements of pterinosomes and
carotrnoid pigments
In culture, there was observed numbers of erythro- phores possessing a noticeable yellow-colored cyto- plasmic matrix. In response to epinephrine (IO-’ M) these erythrophores displayed a unique profile; they were composed of a spherically-shaped aggregate of red pterinosomes in the cell center and surrounded by a deep yellow-colored ring of pigment. Such a pig- mentary scheme was soon changed to a ring-shaped aggregate of red pterinosomes with yellow pigment in the center (Fig. 5). In such a cell, some yellow pigment was retained in the dendrites. Cytochemical tests demonstrated that this yellow pigment was com- pletely soluble in organic solvents such as acetone, petroleum ether, toluene. etc., thus suggesting that it was carotenoid in nature. If these cells were subjected to treatments causing pigment dispersal. such as by addition of theophylline (I Oe3 M), the perinuclear mass of yellow pigment around the pterinosome aggregate disappeared first and subsequently disper- sion of pterinosomes occurred. These observations suggest that carotenoids are controlled by mechan- isms similar to those regulating pterinosome move- ments.
DISCUSSION
The present observations suggest that pigment movements within erythrophores of the teleost fish, X~~~~p~o~~s helleri (swordtail), are controlled by chemical cues as described in the first messenger- second messenger hypothesis of hormone action as originally postulated by Sutherland et al. (1968). The use of cultured chromatophores in the present studies strongly suggest that chemical messengers mediate their effects directly on erythrophores rather than possibly indirectly by other cellular elements present in the skin. These studies clearly indicate that sword- tail integumental erythrophores are capable of rever- sibly translocating their pigment organelles in re- sponse to different stimuli. Since epinephrine and nor- epinephrine were potent pigment- aggregating agents and their action was biocked bv dibenamine, it is probable that swordtail erythrop~ores possess aip~a adrenergic receptors (Ahlquist, 1948; Nickerson & Hohenberg, 1967). The accelerating effect of isoproter- enol on the dispersion of aggregated pigments in the presence of dibenamine and the blockage of such an action by propranolol further indicates that beta adrenergic receptors (Ahlquist, 1948; Nickerson, 1970) are also present in swordtail erythrophores. The pres- ence of ulpha and beta adrenergic receptors in melanophores has similarly been demonstrated in a variety of amphibian (Bagnara & Hadley, 1973. for references) and fresh water fish (Fujii & Miyashita, 1975; Miyashita & Fujii, 1975, Obika, 1976). Little information. however, is available as to the nature of adrenoceptors in fish erythrophores. The present
studies demonstrate that these cellular adrenoceptors are involved in pigment movements of swordtail erythrophores.
Melatonin also induced a perinuclear aggregation of pterinosomes but this effect was not blocked by dibenamine thus suggesting that this indoleamine exerts its action through receptors separate from those of catecholamines which induce a similar physiolo~cal response. The separate nature of these receptors has been demonstrated for fish melano- phores (Obika, 1976; Fujii, personal communication). Not all erythrophores respond to melatonin, however. and this may possibly relate to their pattern of distri- bution in the skin. The possibility exists that in addi- tion to a nervous control regulating rapid color changes, erythrophores in certain areas of the skin are under control by melatonin as are the circadian rhythms of certain melanophore responses in some fish (Reed, 1968; Finnin & Reed, 1973). Wyman (1924a, b), Fain & Hadley (1966). and Nichols rt al.
(1966) noted similar interesting variations in teleost melanophore responses to pineal extracts and meia- tonin, respectively.
MSH and ACTH dispersed pigments within sword- taif erythrophores as did catecholamine stimulating of beta adrenergic receptors. Cyclic AMP and its dibutyryl derivative, but not cyclic GMP, similarly stimulated pigment dispersion. These observations suggest that the chemical messengers used in these studies stimulate pigment dispersion through the in- termediary of the intracellular second messenger, cyc- lic AMP. This is further substantiated by the demon- stration that theophylline, a phosphodiesterase inhibi- tor, which increases intracellular cyclic AMP levels (Butcher & Sutherland, 1962) similarly causes eryth- rophore pigment dispersion.
In summary, these results demonstrate that the control of pigment movements within erythrophores of the teleost, X~~~~p~or~~s helleri, is similar to that of melanosome movements within poikilotherm melanophores as reported by numerous other investi- gators. Adrenoceptors apparently play an important role in the control of rapid pterinosome and caro- tenoid vesicle translocations within erythrophores. Apparently the control of erythrophore pigment movements under normal physiological conditions is mediated by the release of an adrenergic neurotrans- mitter.
~c~~~~~~~~~~~~~~~zr- This work was supported, in part, by a grant from the Fukuzawa Fund, Keio University.
REFERENCES
AHLQUIST R. P. (1948) A study of the adrenotropic recep- tors. Am. J. Physiol. 153, 586600.
BAGNARA J. T. (1958) Hypophyseal control of guanophores in anuran larvae. J. exp. 2001. 137, 2655283.
BAGNARA J. T. & HADLLY M. E. (1969) Control of bright- colored pigment cells of fishes and amphibians. Am. Zool. 9, 465-478.
BAGNARA J. T. & HADLEY M. E. (1973) Chromatophores and Color Change: The Comparative Phq’siolqqy of Ani- mal Pigmentation. Prentice-Hall, Englewood Cliffs.
BUTCHER R. W. & SLITHERLAND E. W. (1962) Adeno- sine-3’S’-phosphate in biological materials. Purification and properties of cyclic-3’,5’-nucleotide phosphodi~ster- ase and use of this enzyme to characterize adeno-
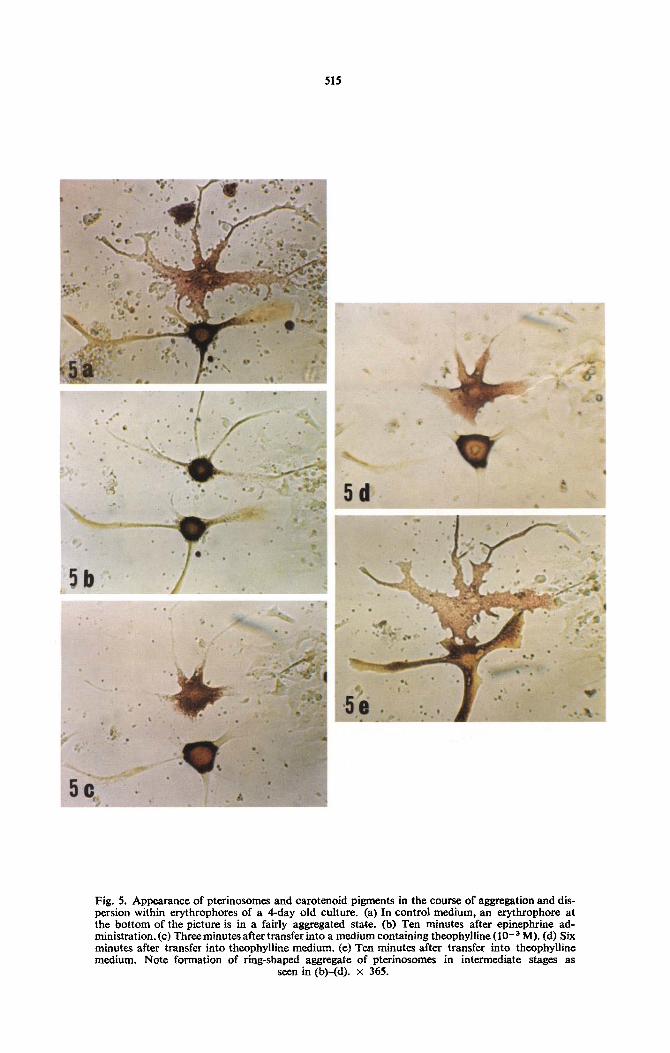
515
Fig. 5. Appearance of pterinosomes and carotenoid pigments in the course of aggregation and dis- persion within erythrophores of a 4-day old culture. (a) In control medium, an erythrophore at the bottom of the picture is in a fairly aggregated state. (b) Ten minutes after epinephrine ad- ministration.(c) Three minutes after transfer into a medium containing theophylline (1O-3 M). (d) Six minutes after transfer into theophylline medium. (e) Ten minutes after transfer into theophylline medium. Note formation of ring-shaped aggregate of pterinosomes in intermediate stages as
seen in (b)-(d). x 365.


Control of pigment movements within swordtail erythrophores 517
sine-3’S’-phosphate in human urine. J. hiol. Chem. 237. 1244-1250.
BYERS H. R. & PORTER K. R. (1976) Pigment migration in cultured erythrophores. J. Cell Biol. 70, 402a.
CASTRUCCI A. M. DE L. (1975) Chromatophores of the teleost Tilapia m~lanopleura. I. Ultrastructure and effect of sodium and potassium on pigment migration. Camp. Biochem. Physiol. !WA, 453456.
EHRMANN R. L. & GEY G. 0. (1956) The growth of cells on a transparent gel of reconstituted rat tail collagen. J. nom. Cancer Inst. 16, 1375-1403.
FAIN W. B. & HADLEY M. E. (1966) In vitro response of melanophores of Fundulus heteroclitus to melatonin. adrenaline. and noradrenaline. Am. Zoo/. 6, 596.
FINNIN B. C. & REED B. L. (1973) A quanta1 bioassay for melatonin. Pigment Cell 1. 180-187.
FUJII R. (1959). Mechanism of ionic action in the melano- phore system of fish. I. Melanophore-concentrating action of potassium and some other ions. Annot. Zoo/. japon. 32. 47-58.
FUJII R. & MIYASHI~A Y. (1975) Receptor mechanism in fish chromatophores. I. Alpha nature of adrenoceptors mediating melanosome aggregation in guppy melano- phores. Comp. Biochem. Physiol. 51. 171-178.
GOODRICH H. B., HILL G. A. & ARRICK M. S. (1941) The chemical identification of gene-controlled pigments in PIatypoecilus and Xiphophorus and comparison with other tropical fish. Genetics 26, 573-556.
HORIKAWA Y. & MATSUMOTO J. (1974) Physiological and ultrastructural bases in the movement of pterinosomes in swordtail erythrophores. Zoo/. Mag. Tokyo 83. 467.
JUNQUEIRA L. C. U. & FARIAS E. C. (1976) Ultrastructure and pigment transport in the xanthophores of 14 species of teleosts. J. Cell Biol. 70, part 2, la.
LEIBOVITZ A. (1963) The growth and maintenance of tissue cell cultures in free gas exchange with the atmosphere. Am. J. Hyg. 78, 173-180.
MATSUMOTO J. (1965) Studies on fine structure and cyto- chemical properties of erythrophores in swordtail, Xiphophorus helleri, with special reference to their pig- ment granules (pterinosomes). J. Cell Biol. 27. 493-504.
MATSUMOTO J. & OBIKA M. (I 968) Morphological and bio- chemical characterization of goldfish erythrophores and their pterinosomes. J. Cell Biol. 39. 233-250.
MIYASHI~A Y. & FUJII R. (1975) Receptor mechanism in fish chromatophores. II. Evidence for beta adrenoceptors mediating melanosome dispersion in guppy melano- phores. Comp. Eiochem. Physiol. SIC, 179-187.
NICHOLS J. C., SCHEEBECK C. & HADLEY M. E. (1966) Com- parative in uiuo response of embryonic, larval. and adult melanophores of Fundulus heteroclitus to melatonin. Am.
Zoo/. 6. 576. NICKERSON M. (1970) Drug inhibiting adrenergic nerves
and structures innervated by them. In The Pharmacologi- cal Basis for Therapeutics (Edited by GOODMAN L. S. and GILMAN A.). 4th edition. pp. 549-585. Macmillan. London.
NICKERSON M. & HOLLENBERG N. K. (1967) Blockage of alpha adrenergic receptors. In Physiological Pharma- cology (Edited by RCI~T W. S. and HOFMANN F. G.). Vol. IV, The Nervous System. Part D. Autonomic Nrr- uous System Drugs. pp. 243-305. Academic Press. New York.
OBIKA M. (1976) An analysis of the mechanism of pigment migration in fish chromatophores. Pigment Cell 3.
254-264.
OBIKA M. & MATSUMOTO J. (1968) Morphological and bio- chemical studies on amphibian bright-colored pigment cells and their pterinosomes. Expl Cc// Res. 52. 646659.
OZATO K. (1976) Pigment dispersion in cultured goldfish erythrophores by dibutyryl adenosine-3’.5’-monophos- phate. Zoo/. May., Tokyo 85. 262-264.
OZA~O K. (1977) Effects of ACTH. adenyl-compounds. and methylxanthines on goldfish erythrophores in culture. Gen. Comp. Endocr. 31. 335-342.
PARKER G. H. (1948) Animal Co/our Changes and Their Neurohumours. Cambridge University Press. Cambridge.
PORTER K. R. (1973) Microtubules in intraL;!I..:!ar :,,,como- tion. In Locomotion of Tissue Cells. Ciha Found. Symp.
14. pp. 149-165. Elsevier. Amsterdam. REED B. L. (1968) The control of circadian pigment changes
in the pencil fish: a proposed role for melatonin. Life Sci. 7. 961-973.
SAC~L M. (19701 Control of prolactin rclcasc and jts role in color change in the teleost. Gi//ichthy,\ nriruhilr,~. J.
uvp. zoo/. 173. 121 127. SMITH D. C. & SMITH M. T. (1935) Obscrvattons on the
color changes and isolated scale erythrophorcs of the squirrel fish. Holoccwtrus asccns~or~i.~ (Osbcck). Bio/. RI,//. 68. 131-139.
SUTHERLAND E. W.. ROBISON G. A. & B~TCHEK R. W. (19681 Some aspects of ‘he biological role of adenosine 3’S’-monophosphate (cyclic AMP). Circulation 37. 279-306.
WARING H. (1963) Color Change Mechanisms of’ Cold Blooded Vertebrates. Academic Press. New York.
WYMAN L. C. (1924a) The reactions of the melanophores of embryonic and larval Fundulus to certain chemical substances. J. exp. Zoo/. 40. 161-180.
WYMAN L. C. (1924b) Blood and nerve as controlling agents in the movements of melanophorcs. J <‘\-p. ZOO/. 39. 73-l 3’.
YAMAMOTO T. (1941) The osmotic properties of the egg of fresh water fish, Ory:ias latipes. J. Far. Sci. Imp, L’nir. Tokyo. Section IV. 5. 461471.
C.&P. 61/3~--1