Embed Size (px)
Citation preview

Mechanisms of tolerance to
Melaleuca alternifolia (tea tree) oil in
Pseudomonas aeruginosa
by
Chelsea Jade Papadopoulos
(nee Longbottom)
BSc (Hons)
This thesis is presented for the degree of
Doctor of Philosophy at The University of Western Australia
2008
Discipline of Microbiology & Immunology
School of Biomedical, Biomolecular & Chemical Sciences
The University of Western Australia,
Crawley, Western Australia, 6009

SUMMARY ............................................................................................................................ I
STATEMENT/DECLARATION .........................................................................................III
ACKNOWLEDGEMENTS ................................................................................................. IV
PRESENTATIONS AND PUBLICATIONS ........................................................................ V
ABBREVIATIONS ............................................................................................................. VI
LIST OF TABLES ............................................................................................................... IX
LIST OF FIGURES ............................................................................................................. XI
1.0 Chapter One – Literature Review ...........................................................................1
1.1 Pseudomonas aeruginosa ...................................................................................... 1
1.1.1 Importance as a pathogen ............................................................................... 1
1.1.2 Resistance to antibacterial agents................................................................... 2
1.1.2.1 Intrinsic resistance mechanisms ................................................................. 2
1.1.2.1.1 Outer membrane ................................................................................... 2
1.1.2.1.2 Lipopolysaccharide .............................................................................. 3
1.1.2.1.3 Efflux.................................................................................................... 4
1.1.2.2 Acquired or adaptive resistance mechanisms ............................................ 7
1.1.2.2.1 Over-expression of efflux pumps ......................................................... 7
1.1.2.2.2 Acquisition, or modified expression, of resistance genes .................... 8
1.1.2.2.3 Decreased expression of porins ............................................................ 8
1.1.2.2.4 Fatty acid changes ................................................................................ 8
1.1.2.2.5 Changes in hydrophobicity .................................................................. 9
1.1.2.3 Antibiotic susceptibility ............................................................................. 9
1.1.2.4 Disinfectant and biocide susceptibility .................................................... 10
1.2 Characteristics of Tea tree oil .............................................................................. 10
1.2.1 Production, composition and physical properties ........................................ 10
1.2.2 Antimicrobial properties .............................................................................. 11
1.2.2.1 Antibacterial properties ............................................................................ 11
1.2.2.2 Antifungal properties ............................................................................... 11
1.2.2.3 Antiviral properties .................................................................................. 12
1.2.3 Other properties ............................................................................................ 12
1.2.3.1 Anti-inflammatory.................................................................................... 12
1.2.3.2 Anti-protozoan ......................................................................................... 13
1.2.3.3 Anti-parasitic ............................................................................................ 13
1.2.3.4 Anti-cancer ............................................................................................... 13
1.2.4 Toxicity ........................................................................................................ 14
1.2.4.1 Oral ........................................................................................................... 14
1.2.4.2 Dermal ...................................................................................................... 14
1.2.4.2.1 Irritant reactions ................................................................................. 14
1.2.4.2.2 Contact allergy ................................................................................... 14
1.2.4.3 Reproductive and developmental toxicity ................................................ 15
1.2.5 Mechanisms of antibacterial action .............................................................. 15
1.3 Pseudomonas and tea tree oil ............................................................................... 16
1.4 Resistance to tea tree oil ....................................................................................... 17
1.5 Research aims....................................................................................................... 17
2.0 Chapter Two- Materials and methods ..................................................................19
2.1 Materials ............................................................................................................... 19

2.1.1 Microorganisms ........................................................................................... 19
2.1.1.1 Efflux mutants, reference isolates and other strains................................. 19
2.1.2 Chemicals, reagents and media .................................................................... 19
2.1.3 Culture media ............................................................................................... 19
2.1.4 Tea tree oil and components ........................................................................ 20
2.1.5 Buffers and other solutions .......................................................................... 20
2.1.5.1 General ..................................................................................................... 21
2.1.5.2 SDS-PAGE ............................................................................................... 22
2.1.5.3 PCR .......................................................................................................... 24
2.1.5.4 PFGE ........................................................................................................ 25
2.1.6 Polyacrylamide gels ..................................................................................... 27
2.1.7 Agarose gels ................................................................................................. 27
2.1.8 PCR primers ................................................................................................. 28
2.1.9 Cellular fatty acid analysis ........................................................................... 28
2.1.10 Disposables and kits ..................................................................................... 29
2.2 Methods ................................................................................................................ 29
2.2.1 Susceptibility testing .................................................................................... 29
2.2.1.1 Antibiotic susceptibility testing – broth microdilution ............................ 29
2.2.1.2 Breakpoint antibiotic testing – agar dilution ............................................ 30
2.2.1.3 Antibiotic susceptibility testing – disc diffusion ...................................... 30
2.2.1.4 Tea tree oil and component MICs ............................................................ 31
2.2.1.5 Metal ion MICs ........................................................................................ 31
2.2.2 Time-kill assays ........................................................................................... 32
2.2.3 LPS analysis ................................................................................................. 32
2.2.3.1 LPS extraction .......................................................................................... 32
2.2.3.2 SDS-PAGE ............................................................................................... 33
2.2.3.3 Silver staining .......................................................................................... 33
2.2.4 Transposon mutagenesis .............................................................................. 34
2.2.4.1 Cell preparation ........................................................................................ 34
2.2.4.2 Electroporation and incubation ................................................................ 35
2.2.4.3 Plating ...................................................................................................... 36
2.2.4.4 Spontaneous resistance to kanamycin ...................................................... 36
2.2.4.5 PCR detection of kanamycin cassette in transposon mutants .................. 37
2.2.4.6 Screening of transposon mutants for TTO susceptibility ......................... 38
2.2.4.7 Identification of disrupted gene in transposon mutants ........................... 39
2.2.4.7.1 Random amplification of transposon ends (RATE) and sequencing . 39
2.2.5 Serial subculture ........................................................................................... 41
2.2.5.1 Susceptibility testing ................................................................................ 42
2.2.5.2 Fatty acid analysis .................................................................................... 42
2.2.5.3 Outer membrane permeability.................................................................. 43
2.2.5.4 Hydrophobicity ........................................................................................ 43
2.2.5.4.1 Flow cytometry .................................................................................. 43
2.2.5.4.2 Microbial adhesion to hydrocarbon (MATH) .................................... 44
2.2.5.4.3 Adhesion to polystyrene beads (APSB) ............................................. 44
2.2.5.5 Pulsed field gel electrophoresis (PFGE) .................................................. 45
2.2.5.5.1 Preparation of bacterial plugs ............................................................ 45

2.2.5.5.2 Washing of plugs ............................................................................... 45
2.2.5.5.3 Restriction digest ................................................................................ 45
2.2.5.5.4 Preparation of plugs for PFGE ........................................................... 46
2.2.5.5.5 PFGE conditions ................................................................................ 46
2.2.5.5.6 Detection of bands.............................................................................. 46
2.2.6 Efflux mutants .............................................................................................. 46
2.2.6.1 Complementation ..................................................................................... 46
2.2.6.2 RT-PCR and Real time RT-PCR.............................................................. 47
2.2.6.2.1 Primer optimization – RT-PCR .......................................................... 48
2.2.6.2.2 RNA extraction .................................................................................. 48
2.2.6.2.3 SYBR® green .................................................................................... 48
2.2.6.3 Checkerboard assays ................................................................................ 49
2.2.6.4 Efflux pump inhibition ............................................................................. 50
3.0 Chapter Three - The Role of Efflux Systems in Tolerance of P. aeruginosa to
TTO and Components ........................................................................................................51
3.1 Introduction .......................................................................................................... 51
3.2 Results .................................................................................................................. 52
3.2.1 Susceptibility of efflux mutants to TTO and components ........................... 52
3.2.1.1 MexAB-OprM .......................................................................................... 52
3.2.1.1.1 MICs ................................................................................................... 52
3.2.1.1.2 Time-kill assays ................................................................................. 52
3.2.1.2 MexCD-OprJ ............................................................................................ 53
3.2.1.2.1 MICs ................................................................................................... 53
3.2.1.2.2 Time-kill curves ................................................................................. 53
3.2.1.3 MexXY ..................................................................................................... 54
3.2.1.3.1 MICs ................................................................................................... 54
3.2.1.4 MexEF-OprN ........................................................................................... 54
3.2.1.5 MexJK ...................................................................................................... 54
3.2.2 Complementation ......................................................................................... 55
3.2.3 Efflux pump inhibition ................................................................................. 56
3.2.4 Checkerboard assays with Tween 80 ........................................................... 57
3.2.5 RT-PCR and Real time RT-PCR.................................................................. 57
3.3 Discussion ............................................................................................................ 59
3.3.1 MexAB-OprM .............................................................................................. 59
3.3.2 MexCD-OprJ ................................................................................................ 60
3.3.3 MexXY ......................................................................................................... 63
3.3.4 MexEF-OprN ............................................................................................... 63
3.3.5 MexJK .......................................................................................................... 64
3.3.6 Other efflux pump systems .......................................................................... 65
3.3.7 Efflux pump inhibitors ................................................................................. 65
3.3.8 RT-PCR and real time RT-PCR ................................................................... 67
3.3.9 Summary ...................................................................................................... 67
4.0 Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in
P. aeruginosa ........................................................................................................................68
4.1 Introduction .......................................................................................................... 68
4.2 Results .................................................................................................................. 70

4.2.1 Susceptibility of rough mutants and parental strains ................................... 70
4.2.1.1 TTO and component MICs ...................................................................... 70
4.2.1.2 Time-kill assays with TTO and components ........................................... 71
4.2.1.2.1 rmlC mutants ...................................................................................... 71
4.2.1.2.2 AK1012 mutant .................................................................................. 72
4.2.1.3 Antibiotics ................................................................................................ 72
4.2.2 SDS-PAGE ................................................................................................... 73
4.3 Discussion ............................................................................................................ 74
4.3.1 Susceptibility to TTO and components ........................................................ 74
4.3.2 Susceptibility to antibiotics and SDS ........................................................... 76
4.3.3 Summary ...................................................................................................... 78
5.0 Chapter Five – Induced resistance to TTO ..........................................................79
5.1 Introduction .......................................................................................................... 79
5.2 Results .................................................................................................................. 80
5.2.1 Serial subculture ........................................................................................... 80
5.2.2 TTO and terpinen-4-ol MICs ....................................................................... 81
5.2.3 PAβN MICs and MBCs ............................................................................... 81
5.2.4 Antibiotic susceptibility ............................................................................... 82
5.2.4.1 Agar dilution ............................................................................................ 82
5.2.4.2 Disc diffusion ........................................................................................... 82
5.2.4.3 Broth microdilution .................................................................................. 83
5.2.5 Metal ion susceptibility ................................................................................ 84
5.2.6 Disinfectant susceptibility ............................................................................ 84
5.2.7 Fatty acid analysis ........................................................................................ 85
5.2.8 Outer membrane permeability...................................................................... 85
5.2.9 Cell surface hydrophobicity ......................................................................... 85
5.2.9.1 Microbial adhesion to hydrocarbon (MATH) .......................................... 86
5.2.9.2 Adhesion to polystyrene beads (APSB) ................................................... 86
5.2.9.3 Flow cytometry ........................................................................................ 86
5.2.10 PFGE ............................................................................................................ 86
5.3 Discussion ............................................................................................................ 86
5.3.1 Selection of resistance to TTO and terpinen-4-ol by serial subculture ........ 86
5.3.1.1 Use of mutS .............................................................................................. 89
5.3.2 Susceptibility of subculture isolates ............................................................. 89
5.3.2.1 TTO and terpinen-4-ol susceptibility ....................................................... 89
5.3.2.1.1 Following subculture in TTO ............................................................. 89
5.3.2.1.2 Following subculture in terpinen-4-ol ................................................ 90
5.3.2.2 Antibiotic susceptibility ........................................................................... 91
5.3.2.2.1 β-Lactam antibiotics ........................................................................... 91
5.3.2.2.2 Quinolones ......................................................................................... 94
5.3.2.2.3 Chloramphenicol ................................................................................ 95
5.3.2.2.4 Aminoglycosides ................................................................................ 95
5.3.2.2.5 Clinical relevance ............................................................................... 96
5.3.2.2.6 Other studies ...................................................................................... 96
5.3.2.3 Metal ion susceptibility ............................................................................ 97
5.3.2.4 Disinfectant susceptibility ........................................................................ 97

5.3.2.5 PAβN ........................................................................................................ 99
5.3.3 Fatty acid analysis ........................................................................................ 99
5.3.4 Outer membrane permeability.................................................................... 100
5.3.5 Hydrophobicity studies .............................................................................. 101
5.3.6 Prolonged use of TTO and antimicrobial resistance .................................. 101
5.3.7 Summary .................................................................................................... 102
6.0 Chapter Six – A library of transposon mutants with altered TTO and terpinen-
4-ol susceptibility ...............................................................................................................104
6.1 Introduction ........................................................................................................ 104
6.2 Results ................................................................................................................ 105
6.2.1 Testing for spontaneous resistance to kanamycin ...................................... 105
6.2.2 Transposon mutagenesis and optimization ................................................ 105
6.2.3 PCR for detection of KanR cassette ............................................................ 106
6.2.4 Control experiment on effect of -80°C storage .......................................... 107
6.2.5 TTO and terpinen-4-ol susceptibilities of mutants..................................... 107
6.2.6 Sequencing and identification of disrupted genes ...................................... 108
6.3 Discussion .......................................................................................................... 109
6.3.1 Transposon mutagenesis ............................................................................ 109
6.3.2 Susceptibility to TTO and terpinen-4-ol .................................................... 110
6.3.3 Sequencing and insertion site identification .............................................. 110
6.3.4 Disrupted genes and regions ...................................................................... 111
6.3.4.1 surA ........................................................................................................ 112
6.3.4.2 Flagellar biogenesis genes...................................................................... 113
6.3.4.2.1 fleN ................................................................................................... 114
6.3.4.2.2 fliC ................................................................................................... 114
6.3.4.2.3 flgB ................................................................................................... 116
6.3.4.2.4 cheZ .................................................................................................. 116
6.3.4.3 rpoN ....................................................................................................... 117
6.3.4.4 hitA ......................................................................................................... 117
6.3.4.5 PA0084 ................................................................................................... 118
6.3.4.6 PA3800 ................................................................................................... 118
6.3.4.7 PA0372 ................................................................................................... 118
6.3.4.8 Region between nusG and rplK ............................................................. 119
6.3.4.9 Region between tyrZ and PA0668.1 ...................................................... 119
6.3.5 Summary .................................................................................................... 119
7.0 Chapter Seven - General discussion ....................................................................121
REFERENCES....................................................................................................................128

I
SUMMARY
Pseudomonas aeruginosa, an important opportunistic pathogen, is resistant to a wide array
of functionally and structurally diverse antimicrobial agents including antibiotics,
disinfectants and biocides. P. aeruginosa is more resistant than other Gram negative
bacteria to tea tree oil (TTO), the essential oil steam distilled from the leaves of Melaleuca
alternifolia and comprised of over 100 terpene hydrocarbon components and their
oxygenated derivatives. TTO is an established topical antimicrobial agent, with
antibacterial, antiviral and antifungal properties.
Intrinsic antimicrobial resistance mechanisms in P. aeruginosa include the low
permeability of the outer membrane and expression of multi-drug efflux pumps. A series of
multi-drug efflux mutants from the resistance-nodulation-cell division family was obtained
and their susceptibility to TTO and several components examined. This demonstrated that
TTO and the components terpinen-4-ol, 1,8-cineole and α-terpineol were substrates of
MexAB-OprM, using both pump deletion mutants and the pump inhibitor Phe-arg β-
naphthylamide dihydrochloride. In complementation studies, the addition of mexAB-oprM
to deletion mutants restored susceptibility to these agents to that of the wild-type,
confirming the role of MexAB-OprM in tolerance to TTO and these three components.
Interplay between the MexAB-OprM and MexCD-OprJ pumps may contribute towards
tolerance of some components of TTO, including 1,8-cineole and α-terpineol. It appears
that TTO, terpinen-4-ol, 1,8-cineole, α-terpineol, ρ-cymene and γ-terpinene are not
substrates of the MexXY and MexJK efflux pumps, though more work is required to
confirm this. A decrease in susceptibility to terpinen-4-ol was noted in a mutant hyper-
expressing MexEF-OprN in conjunction with MexAB-OprM, indicating that terpinen-4-ol
is probably a substrate of MexEF-OprN, while TTO and the other components are not.
Investigation into the role of lipopolysaccharide (LPS) in protection from the antimicrobial
effects of TTO and some components, using a number of LPS mutants of P. aeruginosa,
was undertaken. Mutants with no O-antigen and a truncated LPS core were more
susceptible to TTO, terpinen-4-ol, α-terpineol and 1,8-cineole compared with parental
strains, indicating the significance of a full core in protection from these agents.

II
Serial subculture of several P. aeruginosa strains in TTO and in terpinen-4-ol revealed that
increased tolerance to terpinen-4-ol occurred more readily than for whole TTO. Given that
whole TTO is comprised of a large number of components, thought to have multiple
mechanisms of action, this was not surprising. Low level TTO resistance and reduced
susceptibility to β-lactams, nalidixic acid, chloramphenicol and gentamicin was observed in
P. aeruginosa PAO1 subcultured in TTO, probably due to expression of Mex efflux
systems including MexAB-OprM and MexCD-OprJ. An increase in susceptibility to
ticarcillin and Timentin occurred in PAO1 following serial subculture in terpinen-4-ol.
Susceptibility to ticarcillin has been associated with expression of the MexCD-OprJ system
in P. aeruginosa.
A library of transposon mutants was created to find additional mechanisms by which
P. aeruginosa could tolerate TTO. The library yielded a total of 20 mutants that were more
susceptible than parental strains to TTO and/or terpinen-4-ol. The insertion site of the
transposon was identified in 14 mutants and, in four mutants, this was a gene related to
flagellar biosynthesis. Flagella deficient mutants have previously demonstrated enhanced
susceptibility to the membrane-disrupting surfactant sodium dodecyl sulfate and this echoes
the increased susceptibility to TTO and terpinen-4-ol observed. Three non-sibling surA
mutants were also identified. SurA is involved in the correct folding of outer membrane
proteins, including porins, in Gram negative bacteria: surA mutants of Escherichia coli
have phenotypes that are characteristic of a defective cell envelope, including an increased
susceptibility to hydrophobic agents. The increase in susceptibility to hydrophobic TTO
and terpinen-4-ol in the surA mutants is consistent with this and represents the first report
linking SurA function to antimicrobial resistance in P. aeruginosa.
In conclusion, several Mex efflux systems of P. aeruginosa including MexAB-OprM,
MexCD-OprJ and MexEF-OprN, as well as the LPS core, outer membrane integrity and a
functioning flagella biosynthetic pathway contribute to the tolerance of this organism to
TTO and/or several components.

III
STATEMENT/DECLARATION
Except where duly acknowledged, all work presented in this Thesis was performed by the
PhD candidate.
__________________________
Chelsea Jade Papadopoulos

IV
ACKNOWLEDGEMENTS
I would like to thank my supervisors Christine Carson, Barbara Chang and Thomas Riley
for their guidance throughout this project, for reading numerous chapter drafts and for
helping me realize that I could finish this Thesis. In particular I want to thank Christine, for
without her never-ending patience, enthusiasm and friendship I would not have made it to
the other side and I will forever be grateful to her.
Without the assistance of many people, this project would not have been possible and so I
would like to thank: the Rural Industries Research and Development Corporation for
providing funding; Australian Plantations Pty Ltd for supplying tea tree oil and SNP
Natural Products for supplying terpinen-4-ol; Joseph Lam, Keith Poole, Antonio Oliver,
Herbert Schweizer and Harry Sakellaris for providing the isolates and plasmids used in the
study; Max Aravena-Roman for fatty acid analysis; and Gerry Harnett and Glenys Chidlow
for assistance with all things molecular.
Thanks to the friends and colleagues I shared this journey with, who have helped keep me
sane, both in and out of the lab: Shelly, Krista, Niki, Kate, Shan, Kerry, Trina, Adam,
Avram, Nevada, Syndie, Zoe and Aurelia.
To my wonderful parents and grandmother – thank you for your love and support.
Most importantly, the biggest thank you goes to my husband Mark, for standing by me and
still loving me in the dark times – I could not have made it without you.

V
PRESENTATIONS AND PUBLICATIONS
The following abstracts were accompanied by oral or poster presentations at scientific meetings,
based on work presented in this thesis:
Oral
Papadopoulos CJ, Carson CF, Chang BJ and Riley T. The core lipopolysaccharide of Pseudomonas
aeruginosa and tolerance to tea tree oil. Presented at the Australian Society for Microbiology
Annual Scientific Meeting. Gold Coast, Australia, June 2006.
Poster
Papadopoulos CJ, Carson CF, Chang BJ and Riley TV
The role of the cell envelope and efflux mechanisms of Pseudomonas aeruginosa
in tolerance to tea tree oil and component terpenes. Presented at the American Society for
Microbiology Pseudomonas Conference. Seattle, Washington, USA, August 2007.
Papadopoulos CJ, Carson CF, Chang BJ and Riley TV.
The MexAB-OprM efflux pump plays a role in the tolerance of Pseudomonas aeruginosa to tea tree
oil and terpinen-4-ol. Presented at the Australian Society for Microbiology Annual Scientific
Meeting. Canberra, Australia, September 2005
The following publications were based on the work presented in this thesis:
Peer Reviewed Publications
Papadopoulos CJ, Carson CF, Chang BJ and Riley TV
Role of the MexAB-OprM efflux pump of Pseudomonas aeruginosa in tolerance to tea tree
(Melaleuca alternifolia) oil. Applied and Environmental Microbiology. 2008;74(6):1932-1935
Papadopoulos CJ, Carson CF, BJ Chang and Riley TV
The role of the multi-drug efflux systems of Pseudomonas aeruginosa in tea tree oil tolerance. Peer
Reviewed Poster Presentation, 16th European Congress of Clinical Microbiology and Infectious
Diseases. Nice, France, April 2006.

VI
ABBREVIATIONS
ABC Adenosine triphosphate-binding cassette
AK1012 P. aeruginosa PAO1 AK1012
APSB Adhesion to polystyrene beads
ATP Adenosine triphosphate
BA Blood agar
BHIB Brain heart infusion broth
CCCP Carbonyl cyanide m-chlorophenylhydrazone
CLB Cell lysis buffer
DMSO Dimethyl sulfoxide
EDTA Ethylenediaminetetraacetic acid
EPI Efflux pump inhibitor
FAME Fatty acid methyl esters
h Hour(s)
HRM High resolution melt
LA Luria agar
LA/KAN Luria agar with 500 mg/L kanamycin
LB Luria-Bertani broth
LBA Luria-Bertani agar
LPS Lipopolysaccharide
MATE Multidrug and toxic compound extrusion
MATH Microbial adhesion to hydrocarbons
MFS Major facilitator superfamily
MHA Mueller-Hinton agar
MHB Mueller-Hinton broth
MHB-T Mueller-Hinton broth with 0.002% Tween 80
MIC Minimum inhibitory concentration
min Minute(s)
ms Millisecond(s)
mutS P. aeruginosa PAO1mutS
mutS+20
PAO1mutS subcultured on blood agar only for 20 passages

VII
mutSTTO
PAO1mutS 6% TTO
mutSTTO+10
PAO1mutS 6% TTO plus 10 passages
mutSTTO+20
PAO1mutS 6% TTO plus 20 passages
mutST4ol
PAO1mutS 9% terpinen-4-ol
mutST4ol+10
PAO1mutS 9% terpinen-4-ol plus 10 passages
mutST4ol+20
PAO1mutS 9% terpinen-4-ol plus 20 passages
NA Nutrient agar
NBT Nutrient broth with 0.002% Tween 80
OMP Outer membrane permeability
ORF Open reading frame
PAβN Phe-Arg-β-naphthylamide
PAK P. aeruginosa PAK
PAKrmlC P. aeruginosa PAKrmlC
PAKwbpL P. aeruginosa PAKwbpL
PAO1 P. aeruginosa PAO1
PAO1+20
PAO1 subcultured on blood agar only for 20 passages
PAO1TTO
PAO1 6% TTO
PAO1TTO+10
PAO1 6% TTO plus 10 passages
PAO1TTO+20
PAO1 6% TTO plus 20 passages
PAO1T4ol
PAO1 6.5% terpinen-4-ol
PAO1T4ol+10
PAO1 6.5% terpinen-4-ol plus 10 passages
PAO1T4ol+20
PAO1 6.5% terpinen-4-ol plus 20 passages
PAO1rmlC P. aeruginosa PAO1rmlC
PAO1wbpL P. aeruginosa PAO1wbpL
PBS Phosphate-buffered saline
PFGE Pulsed field gel electrophoresis
PMBN Polymyxin B nonapeptide
pNPP Para-nitro phenyl phosphate
RATE Random amplification of transposon ends
RE Restriction enzyme
RND Resistance-nodulation-cell division

VIII
RT Room temperature
s Second(s)
SDS-PAGE Sodium dodecyl sulfate polyacrylamide gel electrophoresis
SDW Sterile distilled water
SMR Small multidrug resistance
TAE Tris-acetate buffer
TAE/EB TAE plus ethidium bromide
TSBA Trypticase soy blood agar
TTO Tea tree oil
V Volts
10662 P. aeruginosa NCTC 10662

IX
LIST OF TABLES
The page number listed is of the page immediately preceding the table.
Table 1.1 Composition profile for oil of Melaleuca, terpinen-4-ol type p. 10
(tea tree oil)
Table 2.1 P. aeruginosa efflux mutants and parental strains p. 19
Table 2.2 Reference and other strains of bacteria p. 19
Table 2.3 Chemicals, reagents and media p. 19
Table 2.4 Gas chromatography mass spectrometry analysis of TTO p. 20
batch W/E504
Table 2.5 PCR primers used in this study p. 28
Table 2.6 Metal ions and concentrations used in agar dilution p. 31
Table 3.1 TTO and component MICs (%, v/v) by broth p. 52
microdilution for efflux mutants and parent strains
Table 3.2 Initial susceptibility screening of transformants to TTO and p. 55
terpinen-4-ol (%, v/v)
Table 3.3 TTO and component MICs for complemented strains and their p. 55
parent isolates (%, v/v)
Table 3.4 Susceptibility of efflux mutants and parental strain of p. 56
P. aeruginosa to the efflux inhibitor PAβN
Table 3.5 MICs (% v/v) of TTO, terpinen-4-ol and 1,8-cineole against p. 56
P. aeruginosa PAO1 and efflux mutants, with and without
20µg/ml PAβN
Table 4.1 MICs (%, v/v) of TTO and components against of rough p. 70
mutants of P. aeruginosa and parental strains
Table 4.2 MICs of antibiotics and SDS (mg/L) against rough mutants p. 72
of P. aeruginosa and their parental strains
Table 5.1 Concentrations of TTO and terpinen-4-ol reached during serial p. 80
subculture
Table 5.2 TTO and terpinen-4-ol susceptibilities for serial subculture p. 81
isolates derived from P. aeruginosa PAO1

X
Table 5.3 TTO and terpinen-4-ol susceptibilities for serial subculture p. 81
isolates derived from P. aeruginosa PAO1mutS
Table 5.4 Susceptibility of serial subculture isolates of P. aeruginosa to p. 81
PAβN (mg/L)
Table 5.5 Agar dilution antibiotic susceptibility screening results for serial p. 82
subculture isolates of P. aeruginosa
Table 5.6a Broth microdilution antibiotic susceptibilities (mg/L) for serial p. 83
subculture isolates of P. aeruginosa
Table 5.6b Broth microdilution antibiotic susceptibilities (mg/L) for serial p. 83
subculture isolates of P. aeruginosa
Table 5.7 Agar dilution MICs for various metal ions against serial p. 84
subculture isolates of P. aeruginosa
Table 5.8 Broth dilution MICs and MBCs (mg/L) for disinfectants against p. 84
serial subculture isolates of P. aeruginosa
Table 5.9 Average composition and ratios of fatty acids of P. aeruginosa p. 85
PAO1 and serial subculture mutants
Table 5.10 Average composition and ratios of fatty acids of P. aeruginosa p. 85
PAO1muts and serial subculture mutants
Table 6.1 Spontaneous resistance to kanamycin in P. aeruginosa p. 105
NCTC 10662 on Luria agar
Table 6.2 Optimization of electroporation conditions using P. aeruginosa p. 106
PAO1 and pBBR1MCS
Table 6.3 Transposon mutagenesis experiments that yielded p. 106
PCR-confirmed transposon mutants
Table 6.4 MICs of TTO and terpinen-4-ol (% v/v) for transposon mutants p. 107
of P. aeruginosa with altered susceptibility compared to parental
strains
Table 6.5 Susceptibilities of control electroporation isolates of p. 107
P. aeruginosa NCTC 10662 to TTO and terpinen-4-ol (%, v/v)
Table 6.6 Insertion sites of transposon mutants of P. aeruginosa p. 109

XI
LIST OF FIGURES
The page number listed is of the page immediately preceding the figure.
Figure 2.1 Strategy for generating and sequencing EZ::TN™ <KAN-2> p. 34
transposon insertions in the genome of P. aeruginosa
Figure 2.2 Primer binding sites and PCR target product within p. 37
Epicentre EZ::TN™ <KAN-2> sequence
Figure 2.3 Summary of the random amplification of transposon ends p. 39
method for identifying the insertion site in transposon mutants
Figure 3.1 Time-kill curves for P. aeruginosa ML5087 (A, parent), p. 52
K1121 (B, MexAB-OprM-) and K1110 (C, MexAB
+-OprM
-)
incubated with TTO, terpinen-4-ol or MHB-T for 120 min at
35°C and shaking
Figure 3.2 Time-kill curves for ML5087(A, parent), K1115 p. 53
(B, MexAB-OprM- MexCD-OprJ
-) and K1131
(C, MexAB-OprM- MexCD-OprJ
++ [nfxB]) incubated with
TTO, terpinen-4-ol or MHB-T for 120 min at 35°C and shaking
Figure 3.3 Time-kill curves for P. aeruginosa PAO1 (A, parent) and p. 54
K1536 (B, MexAB-OprM+ MexCD-OprJ
++ [nfxB]) incubated
with TTO, terpinen-4-ol or MHB-T for 120 min at 35°C and
shaking
Figure 3.4 Plasmid mini-prep extraction electrophoresis results p. 55
Figure 3.5 Plasmid mini-preps from putative transformants p. 55
Figure 3.6 PCR optimization of Mex primers p. 57
Figure 3.7 PCR optimization of Mex primers p. 57
Figure 3.8 Test SYBR® green TaqGold PCR using first set Mex primers p. 58
Figure 4.1 Structural lipopolysaccharide core model of rough mutants of p. 69
P. aeruginosa serotype O5 (A) and P. aeruginosa serotype O6 (B)
Figure 4.2 Time-kills curves for P. aeruginosa PAO1 and P. aeruginosa p. 71
PAO1rmlC incubated with TTO or MHB-T for up to 120 minutes
at 35°C and shaking

XII
Figure 4.3 Time-kill curves for P. aeruginosa PAO1 and P. aeruginosa p. 71
PAO1rmlC incubated with terpinen-4-ol, 1,8-cineole or MHB-T
for up to 120 minutes at 35°C and shaking
Figure 4.4 Time-kill curves for P. aeruginosa PAKrmlC and p. 71
P. aeruginosa PAK incubated with TTO or MHB-T for up to
120 minutes at 35°C and shaking
Figure 4.5 Time-kill curves for P. aeruginosa PAK and p. 71
P. aeruginosa PAKrmlC incubated with terpinen-4-ol, 1,8-cineole
or MHB-T for up to 120 minutes at 35°C and shaking
Figure 4.6 Time-kill curves for P. aeruginosa PAO1 and p. 72
P. aeruginosa AK1012 incubated with TTO or MHB-T for
up to 120 minutes at 35°C and shaking
Figure 4.7 Time-kill curves for P. aeruginosa PAO1 and p. 72
P. aeruginosa AK1012 incubated with terpinen-4-ol or MHB-T
for up to 120 minutes at 35°C and shaking
Figure 4.8 SDS-PAGE analysis of LPS mutants and parental strains of p. 73
P. aeruginosa
Figure 5.1 Outer membrane permeability of serial subculture mutants and p. 85
parental strains of P. aeruginosa
Figure 5.2 PFGE of serial subculture mutants and parental strains of p. 86
P. aeruginosa digested with SpeI
Figure 6.1 Representative PCR for detection of KanR cassette in transposon p. 106
mutants of P. aeruginosa
Figure 6.2 Detection of kanamycin resistance cassette in transposon p. 107
mutants of P. aeruginosa following storage at -80°C
Figure 6.3 Optimisation of RATE products for transposon mutants p. 108
P114 (a) and P349 (b)
Figure 6.4 Location of transposon mutant insertions relative to p. 109
P. aeruginosa PAO1 chromosome
Figure 6.5 Location of disrupted gene (surA) of transposon mutants p. 112
P114, P151 and P253 within a predicted operon
(red box) on the chromosome of P. aeruginosa

XIII
Figure 6.6 Location of disrupted gene of transposon mutants P331 (fleN) p. 114
and PA165 (cheZ) on the chromosome of P. aeruginosa
Figure 6.7 Location of disrupted gene (fliC) of transposon mutant p. 115
P744 within a predicted operon (red box) on the
chromosome of P. aeruginosa
Figure 6.8 Location of disrupted gene (flgB) of transposon mutant P349 p. 116
within the flg operon (red box) on the chromosome of
P. aeruginosa
Figure 6.9 Location of disrupted gene (rpoN) of transposon mutant P808 p. 117
within the rpoN operon (red box) on the chromosome of
P. aeruginosa
Figure 6.10 Location of disrupted gene (hitA) of transposon mutant PA140 p. 118
within the hitAB operon (red box) on the chromosome of
P. aeruginosa
Figure 6.11 Location of disrupted gene (PA0084) of transposon mutant p. 118
P716 on the chromosome of P. aeruginosa
Figure 6.12 Location of disrupted gene (PA3800) of transposon mutant p. 118
P770 on the chromosome of P. aeruginosa
Figure 6.13 Location of disrupted gene (PA0372) of transposon mutant p. 118
PA172 on the chromosome of P. aeruginosa
Figure 6.14 Transposon insertion site (black arrow) of mutant P546 between p. 119
nusG and rplK, within a predicted operon (red box) in
P. aeruginosa
Figure 6.15 Transposon insertion site (black arrow) of mutant P565 p. 119
between tyrZ and PA0668.1 within the genome of P. aeruginosa

Chapter One – Literature Review
1
1.0 CHAPTER ONE – LITERATURE REVIEW
1.1 Pseudomonas aeruginosa
Pseudomonas aeruginosa is a non-fermenting, aerobic, Gram negative bacillus belonging
to the family Pseudomonadaceae. Cells range from 1-5µm in length and 0.5-1µm in width
and are motile by a single polar flagellum. Fimbriae and pili are also often present
(Palleroni, 1975). Pseudomonads are mesophilic, with optimum temperatures in the range
30-37°C (Forbes et al., 1998).
1.1.1 Importance as a pathogen
Although not common in normal human flora, P. aeruginosa accounts for 11-13.8% of all
nosocomial infections when a microbial cause is identified and is the most common species
isolated clinically from this genus (Kim et al., 2000; Lizioli et al., 2003; Pittet et al., 1999).
An even higher occurrence of nosocomial infection with P. aeruginosa is reported in
intensive care units, with rates of 13.2-22.6% reported (Erbay et al., 2003; Gaynes and
Edwards, 2005; Kim et al., 2000; Lizioli et al., 2003). P. aeruginosa is an opportunistic
pathogen and causes infections of the urinary tract, respiratory tract, gastrointestinal tract
and bones and joints, as well as bacteremia, dermatitis and soft tissue infections.
The ability to grow in nutrient poor conditions means P. aeruginosa is found in a variety of
environmental locations including soil, water and plants. This species is capable of survival
in domestic (hot tubs, whirlpools, wading pools, contact lens solutions) and hospital (sinks,
showers, respiratory equipment, hydrotherapy pools) environments. Because of the
ubiquitous nature of P. aeruginosa, a variety of transmission pathways occur. Rare in
normal flora, P. aeruginosa can be transmitted via ingestion of contaminated food or water;
exposure to contaminated medical devices or solutions; or introduction by penetrating
wounds. Person-to-person transmission is also thought to occur (Forbes et al., 1998).
Although P. aeruginosa is an environmental inhabitant, it is also a prevalent opportunistic
pathogen and takes advantage of compromised host defenses in order to establish infection.

Chapter One – Literature Review
2
Sites for hospital-acquired infections of P. aeruginosa include respiratory tract (cystic
fibrosis patients), urinary tract, wounds, bloodstream, surgical site infections and the central
nervous system. It is particularly problematic for immunocompromised and debilitated
patients, especially cystic fibrosis sufferers. Community-acquired infections include
folliculitis, otitis externa, ocular infections (following trauma), skin and soft tissue
infections and endocarditis (Forbes et al., 1998).
1.1.2 Resistance to antibacterial agents
P. aeruginosa is notorious for its resistance to many antibacterial agents ranging from
conventional antibiotics, such as β-lactams, tetracyclines, chloramphenicol and
fluoroquinolones, to biocides, organic solvents and metals (Cavallo et al., 2000; Jones et
al., 1989; Teitzel and Parsek, 2003). These agents are both structurally and functionally
diverse and consequently there are many mechanisms involved in mediating resistance.
These resistance mechanisms can be broadly defined as either intrinsic, in which pre-
existing characteristics of the bacterial cell facilitate resistance, or acquired, in which
changes to bacteria result in the acquisition of resistance to agents to which they were
formerly susceptible.
1.1.2.1 Intrinsic resistance mechanisms
Intrinsic mechanisms of resistance to antimicrobial agents in P. aeruginosa include the low
permeability of the outer membrane and constitutive expression of efflux pumps and β-
lactamases.
1.1.2.1.1 Outer membrane
Like all Gram negative bacteria, the outer membrane of P. aeruginosa is comprised of a
lipid bilayer composed of phospholipids, lipoproteins, lipopolysaccharide (LPS) and
proteins. The phospholipids and lipoproteins are mainly located in the inner layer while the
LPS is located in the outer layer. An assortment of outer membrane proteins with a variety

Chapter One – Literature Review
3
of cellular growth and metabolism related functions are contained within the outer
membrane.
The major classes of antibiotics used to treat P. aeruginosa need to cross the outer
membrane to reach their targets, e.g. aminoglycosides inhibit protein synthesis via the
ribosome and polymyxins bind to phospholipids in the cytoplasmic membrane (Lambert,
2002). The outer membrane of P. aeruginosa has been shown to be 12 to 100 times less
permeable than that of E. coli (Nikaido, 1986) and thus represents a considerable
penetration barrier to many antibacterial agents (Nikaido and Vaara, 1985). Small
hydrophilic molecules can cross the outer membrane through the porin channels, while
larger molecules are excluded, particularly hydrophobic antibiotics (Lambert, 2002).
1.1.2.1.2 Lipopolysaccharide
LPS is an essential virulence factor for P. aeruginosa (Cryz et al., 1984). The LPS of
P. aeruginosa consists of a hydrophobic lipid A region inserted into the membrane and
attached to a hydrophilic linear polysaccharide region consisting of the core
oligosaccharide and the O-antigen side chain. The core oligosaccharide is negatively
charged and the association of adjacent LPS molecules is stabilized by Mg2+
ions at the
surface of the membrane (Al-Tahhan et al., 2000). The majority of P. aeruginosa strains
can co-express two chemically and antigenically distinct forms of LPS; a serotype-specific
O-antigen containing B-band LPS and a common antigen referred to as A-band LPS. It is
the B-band O antigen structure that is the basis for the classification of the 20 O serotypes
of P. aeruginosa according to the International Antigenic Typing Scheme (Liu et al., 1983;
Liu and Wang, 1990).
In Gram negative bacteria, LPS contributes towards the low outer membrane permeability
observed (Nikaido and Vaara, 1985). It is assumed that the lipid A–core section of the LPS
molecule is the main influence on the barrier function of the outer membrane; mutations
affecting O-antigen synthesis do not make significant changes to the permeability or
integrity of the outer membrane in enteric bacteria (Schnaitman and Klena, 1993). The

Chapter One – Literature Review
4
ionic interactions in the heptose region of the inner core between divalent cations and
negatively-charged phosphates may stabilize the outer membrane through crosslinking of
LPS molecules (Schnaitman and Klena, 1993).
Mutants lacking the O antigen, or O antigen and parts of the outer and/or inner core, are
called rough mutants. In enteric bacteria, the lack of heptose-linked phosphates in „deep
rough‟ mutants resulted in hypersusceptibility to hydrophobic antibiotics and detergents,
less outer membrane proteins, more outer membrane phospholipids and higher levels of cell
lysis (Nikaido and Vaara, 1985; Parker et al., 1992; Schnaitman and Klena, 1993). Rough
mutants of P. aeruginosa have an increased susceptibility to novobiocin and sodium
dodecyl sulphate (Walsh et al., 2000). Rough mutants missing only the O antigen
component (wbpL mutants) have an increase in susceptibility to EDTA, but are not more
susceptible to organic acids and aromatic hydrocarbons than parent strains (Junker et al.,
2001). LPS mutants of P. aeruginosa with no inner core heptose or phosphate have never
been isolated, suggesting the importance of the heptose-linked phosphate for viability (Lam
et al., 2004). Cross-linking of LPS mediated by phosphate moieties and divalent cations is
thought to stabilize the outer membrane of P. aeruginosa, a theory supported by the high
susceptibility of P. aeruginosa to lysis after treatment with divalent cation chelators such as
EDTA (Lam et al., 2004).
1.1.2.1.3 Efflux
Efflux is the pumping of a solute out of a cell (Piddock, 2006). There are five families of
bacterial efflux transporters that can pump out antimicrobials. The largest two are the major
facilitator superfamily (MFS) (Pao et al., 1998) and the adenosine triphosphate (ATP)-
binding cassette (ABC) superfamily (Fath and Kolter, 1993). The remaining three smaller
families are the multidrug and toxic compound extrusion (MATE) family (Brown et al.,
1999), the small multi-drug resistance (SMR) family (Paulsen et al., 1996) and the
resistance-nodulation-cell division (RND) family (Saier Jr et al., 1994). It is the latter, the
RND family, that plays a major role in both intrinsic and acquired resistance of Gram
negative bacteria to a range of antimicrobials and that will be discussed further here.

Chapter One – Literature Review
5
The RND family of efflux pumps usually operates as a tripartite system, with a transporter
protein in the inner membrane, a periplasmic membrane fusion protein and an outer
membrane protein channel. Usually the three genes are organized in an operon, along with
a regulatory gene, though for some systems the outer membrane protein is not located
adjacent to the other genes, e.g. P. aeruginosa mexXY and oprM (Piddock, 2006). RND
pumps require the energy of the proton gradient across the inner membrane in order to
extrude compounds (Lomovskaya et al., 2001).
Efflux systems are also involved in the uptake of essential nutrients and ions and the
excretion of metabolic end products and often have a wide range of substrate specificity.
The genes responsible for efflux are found in both antibiotic-susceptible and antibiotic-
resistant bacteria (Piddock, 2006). In P. aeruginosa an array of RND efflux systems have
been characterized including MexAB-OprM, MexCD-OprJ, MexXY, MexJK, MexEF-
OprN, MexPQ-OpmE, MexMN-OprM and MexGHI-OpmD (Aendekerk et al., 2002; Aires
et al., 1999; Chuanchuen et al., 2002; Kohler et al., 1997; Li et al., 1995; Mima et al.,
2005; Poole et al., 1996). The MexAB-OprM pump is constitutively expressed and
contributes to intrinsic resistance to β-lactams, tetracycline, chloramphenicol and
fluoroquinolones (Li et al., 1995; Zhang et al., 2001), while MexXY (coupled with OprM)
contributes to the intrinsic resistance of P. aeruginosa to aminoglycosides (Aires et al.,
1999). No other RND pumps characterized in P. aeruginosa are constitutively expressed.
Efflux pump inhibition
Intrinsic antibiotic resistance in Gram negative bacteria is largely attributed to multi-drug
resistance efflux pumps but these pumps are also involved in acquired clinical resistance
(Poole, 2007). The clinical benefits of inhibiting bacterial efflux pumps are vast; rendering
normally resistant bacteria susceptible to antibiotics that are rapidly becoming ineffective
forms of treatment due to ever-increasing levels of bacterial resistance. Combination
therapy of an efflux pump inhibitor (EPI) with an appropriate antibiotic should increase
potency, enhance the spectrum of activity and reduce the occurrence of acquired resistance
(Lomovskaya and Bostian, 2006). Much work has been done in the last decade on potential

Chapter One – Literature Review
6
EPIs for bacteria, including P. aeruginosa (Kaatz, 2002; Lomovskaya et al., 2001; Renau et
al., 1999; Stavri et al., 2007). The first commercially available EPI was Phe-Arg-β-
naphthylamide (PAβN), a broad spectrum low molecular weight dipeptide amide which
was designated MC-207,110 (Lomovskaya et al., 2001).
Originally developed against P. aeruginosa, PAβN is active against efflux pumps in
various Gram-negative bacteria, including resistant Klebsiella pneumoniae strains
(Lomovskaya et al., 2001; Lomovskaya and Bostian, 2006; Pages et al., 2005). Active
against the clinically relevant pumps MexAB-OprM, MexCD-OprJ and MexEF-OprN, as
well as the Mex homolog AcrAB-TolC in E. coli, PAβN significantly decreased intrinsic
and acquired resistance to fluoroquinolones in vitro (Lomovskaya et al., 2001). PAβN was
also able to decrease the invasiveness of P. aeruginosa against Madin-Darby canine kidney
epithelial cells, presumably by decreasing the export of invasion determinants (Hirakata et
al., 2007). Not all antibiotic substrates for a given pump are potentiated by PAβN; for
example, PAβN potentiates fluoroquinolones, macrolides/ketolides, oxazolidinones,
chloramphenicol and rifampicin, but not β-lactams or aminoglycosides (Lomovskaya and
Bostian, 2006). It has been demonstrated that PAβN itself is a substrate of Mex efflux
pumps (Warren et al., 2000). Efforts to improve the potency of PAβN identified two basic
moieties that, while essential for activity, had unfavorable pharmacokinetic and
toxicological profiles (Lomovskaya and Bostian, 2006). Although not a clinical option,
PAβN is still a useful research tool for studying the contribution of efflux pumps to
antimicrobial resistance in Gram negative bacteria.
The majority of EPIs that have shown promise in vitro have also shown toxicity in vivo,
rendering them useless in a clinical setting; however, one drug development program has
reached human clinical trial stage. Mpex Pharmaceuticals is currently undertaking phase II
clinical trials with the EPI MP-601,205. The EPI is delivered in aerosol form combined
with ciprofloxacin for respiratory infections in patients with cystic fibrosis and ventilator-
associated pneumonia (Lynch, 2006). Not surprisingly, neither the structure nor mode of
action of MP-601,205 has been disclosed.

Chapter One – Literature Review
7
Carbonyl cyanide m-chlorophenylhydrazone (CCCP) is a protonophore that eliminates the
proton gradient essential for energy dependent efflux systems, such as MexAB-OprM (Li et
al., 1995). CCCP is widely used in vitro to inhibit efflux systems and investigate the
substrate specificity of multi-drug efflux pumps.
1.1.2.2 Acquired or adaptive resistance mechanisms
1.1.2.2.1 Over-expression of efflux pumps
P. aeruginosa may become resistant through mutation in chromosomal genes which
regulate resistance genes, such as efflux operons. Over-expression of efflux pumps may
occur through mutation of repressor genes, or by the presence of an inducing compound
that may or may not be a substrate for the pump (Nikaido, 2005). Although the mexAB-
oprM operon is always transcribed at low but detectable levels, MexAB-OprM is
overproduced following mutation of MexR, the repressor of MexAB-OprM (Adewoye et
al., 2002; Saito et al., 1999). Over-expression of MexAB-OprM in nalB or nalC mutants
produces resistance to nalidixic acid and fluoroquinolones (Nikaido, 2005). Over-
expression of MexCD-OprJ in nfxB mutants significantly increases resistance to
tetracycline, chloramphenicol, quinolones and 4th
generation cephems (Nikaido, 2005).
Over-expression of MexEF-OprN in nfxC mutants confers increased resistance to
quinolones, trimethoprim and chloramphenicol (Nikaido, 2005).
Antibiotics are not the only compounds for which resistance is made possible through the
over-expression of efflux pumps. The overexpression of the mar genes, regulating the
AcrAB efflux pump, increased organic solvent tolerance in several E. coli strains (Asako et
al., 1997; Okusu et al., 1996). The Mar phenotype is induced in E. coli following exposure
to a variety of aromatic chemicalsand facilitated resistance to antibiotics, household
disinfectants and organic solvents (Alekshun and Levy, 1999).

Chapter One – Literature Review
8
1.1.2.2.2 Acquisition, or modified expression, of resistance genes
P. aeruginosa also has the genetic capacity to express a wide range of resistance
mechanisms, which can be acquired from other organisms via plasmids, transposons and
bacteriophages (Lambert, 2002). Plasmid-mediated enzymes can modify and inactivate
aminoglycosides, while over-expression of chromosomal β-lactamases such as AmpC
mediates acquired resistance to β-lactams (Lambert, 2002).
1.1.2.2.3 Decreased expression of porins
The transmembrane water-filled diffusion channels formed by porin proteins allow
hydrophilic agents access to the cytoplasmic membrane and cytoplasm (Nikaido, 1986).
The OprD porin is well characterized as the uptake pathway for hydrophilic carbapenems,
with down-regulation of OprD resulting in resistance to these antibiotics (Quale et al.,
2006). With antipseudomonal carbapenems often the agents of last resort, the emergence of
carbapenem resistance in P. aeruginosa is of concern (Navon-Venezia et al., 2005).
Reduced permeability through diminished expression of OprD and OprF confers resistance
to β-lactams (Kadry, 2003). The loss of porins may increase the level of resistance to
hydrophilic antimicrobials, but may also affect the passage of essential nutrients and thus
survival of the organism in its natural habitat (Nikaido, 2003).
1.1.2.2.4 Fatty acid changes
Modifications of the cell envelope that increase cell membrane rigidity are known
mechanisms of tolerance to organic solvents (Sardessai and Bhosle, 2002). Alterations in
fatty acid composition that decreased the fluidity of the cell membrane allowed
Pseudomonas putida to grow in the presence of the organic solvent toluene (Kim et al.,
2002). An increase in saturated fatty acids was associated with increased tolerance to ο-
xylene in P. putida (Pinkart, 1996). Fatty acid modifications were also observed in
P. aeruginosa adapted to benzalkonium chloride (Guerin-Mechin et al., 1999; Loughlin et
al., 2002) while the change from cis to trans isomerism of unsaturated fatty acids was

Chapter One – Literature Review
9
reported as an adaptation mechanism to environmental stress in P. aeruginosa (Heipieper et
al., 2003).
1.1.2.2.5 Changes in hydrophobicity
Decreased cell surface hydrophobicity is another mechanism of organic solvent tolerance in
bacteria (Sardessai and Bhosle, 2002). Changes in the cell surface hydrophobicity of
organic solvent tolerant E. coli mutants have been reported (Aono and Kobayashi, 1997).
These solvent tolerant mutants had a decrease in cell surface hydrophobicity compared to
parental strains, thought to be related to an increase in LPS content, with no major change
in chemical composition detected.
1.1.2.3 Antibiotic susceptibility
P. aeruginosa is intrinsically resistant to macrolides, tetracyclines, co-trimoxazole, many β-
lactams and most fluoroquinolones. P. aeruginosa is not intrinsically resistant to
carboxypenicillins (ticarcillin), β-lactam/β-lactamase inhibitor combinations, 4th
generation
cephalosporins, some 3rd
generation cephalosporins (cefepime, ceftazidime and
cefoperazone), aminoglycosides (gentamicin, tobramycin and amikacin), monobactams
(aztreonam), some fluoroquinolones (levofloxacin and ciprofloxacin), carbapenems
(imipenem, meropenem and ertapenem) and the polymyxins (colistin) (Driscoll et al.,
2007). However, resistance to any of these antibiotics can develop in P. aeruginosa,
particularly following antibiotic exposure (Driscoll et al., 2007).
The surveillance of the susceptibility profiles of clinical isolates of P. aeruginosa in several
recent studies has indicated an increasing trend of resistance. In a report by the National
Nosocomial Infection Surveillance System on P. aeruginosa isolated from intensive care
units in 2003, resistance to imipenem, fluoroquinolones and 3rd
generation cephalosporins
was 21.1%, 29.5% and 31.9%, respectively (NNIS, 2004).

Chapter One – Literature Review
10
1.1.2.4 Disinfectant and biocide susceptibility
P. aeruginosa is less susceptible to a wide range of disinfectants and biocides and has been
found as a contaminant in antiseptic preparations including benzalkonium chloride and
cetrimide (Lee and Fialkow, 1961; Lowbury, 1951). The susceptibility of P. aeruginosa to
commonly used disinfectants and biocides varies between strains and resistance to infection
control-strength dilutions of benzalkonium chloride, chlorhexidine, cetrimide and phenolic
disinfectants has been documented (reviewed in (Flaherty and Stosor, 2004)).
1.2 Characteristics of Tea tree oil
1.2.1 Production, composition and physical properties
Tea tree oil (TTO) is the essential oil produced from the steam distillation of the leaves of
the Australian native plant Melaleuca alternifolia and an established broad spectrum topical
antimicrobial agent (Carson et al., 2006). Although “tea tree” has been used to describe
members of the Leptospermum, Melaleuca and Neofabricia genera, commercially produced
TTO is regulated by an international standard and is usually derived from M. alternifolia
only. TTO is composed of terpene hydrocarbons, mainly monoterpenes, sesquiterpenes and
their related alcohols and the international standard, ISO 4730 (International Organisation
for Standardisation, 2004), regulates the range of concentrations of 14 of the components.
There are approximately 100 components in TTO, as reported in 1989 by Brophy et al.
following gas chromatography and gas chromatography-mass spectrometry analysis of
more than 800 samples of TTO (Brophy et al., 1989). Table 1.1 outlines the ISO 4730
range for 14 components while Table 2.4 details the concentration of these components in
the batch of TTO used in this study. TTO has low solubility in aqueous solution, but is
miscible with non-polar solvents.

Chapter One – Literature Review
Table 1.1 Composition profile for oil of Melaleuca, terpinen-4-ol type (tea tree
oil)
Component Range (%) Average (%)
Terpinen-4-ol >30 40.1
γ-Terpinene 10-28 23
α-Terpinene 5-13 10.4
1,8-Cineole <15 5.1
Terpinolene 1.5-5 3.1
ρ-Cymene 0.5-12 2.9
α-Pinene 1-6 2.6
α-Terpineol 1.5-8 2.4
δ-Cadinene Tr1-8 1.3
Aromadendrene Tr-7 1.5
Limonene 0.5-4 1.0
Sabinene Tr-3.5 0.2
Globulol Tr-3 0.2
Viridiflorol Tr-1.5 0.1
Legend
(1) Tr = trace.
Adapted from (Brophy et al., 1989) and (International Organisation for
Standardisation, 2004)

Chapter One – Literature Review
11
1.2.2 Antimicrobial properties
1.2.2.1 Antibacterial properties
TTO is well established as an antibacterial agent with in vitro activity against a wide array
of Gram positive and Gram negative bacteria (Carson et al., 2006). The first report of
antibacterial activity was a paper by Penfold in the 1920s that compared the antiseptic
property of crude TTO to that of phenol. The resulting Rideal-Walker coefficient of 11
implied that the TTO used was 11 times more effective than phenol as an antiseptic and
disinfectant (Penfold and Grant, 1925). Modern day in vitro assays utilize either broth or
agar mediums, with broth microdilution assays the most common method used for
determining minimum inhibitory concentrations (MICs).
The MICs for most bacteria range from 0.25 to 1% (v/v), while documented MICs for
P. aeruginosa range from 1 to 8% (v/v) (Altman, 1988; Banes-Marshall et al., 2001;
Papadopoulos et al., 2006). MICs for other bacteria of note include vancomycin resistant
Enterococcus faecium (0.5-1%), Propionibacterium acnes (0.05-0.63%), S. aureus (0.5-
1.25%) and methicillin resistant S. aureus (0.04-0.35%), S. epidermidis (0.45-1.25%) and
E. coli (0.08-2%) (reviewed in (Carson et al., 2006)). The mechanism of antibacterial
action of TTO is discussed in section 1.2.5.
1.2.2.2 Antifungal properties
TTO has activity both in vitro and in vivo against a range of fungi including yeasts,
dermatophytes and other filamentous fungi and has been used to treat conditions including
dandruff and tinea pedis (D'Auria et al., 2001; Griffin and Markham, 2000; Hammer et al.,
2002; Hammer et al., 2004; Nenoff et al., 1996; Satchell et al., 2002a; Satchell et al.
2002b). The majority of studies into the mechanism of antifungal action of TTO have used
Candida albicans as the test organism. Studies have shown that TTO increased cell
permeability and cell fluidity and inhibited cell respiration and germ tube formation (Cox et
al., 2000; Hammer et al., 2000; Hammer et al., 2004).

Chapter One – Literature Review
12
1.2.2.3 Antiviral properties
The antiviral effect of TTO has been demonstrated on a small number of viruses. Several
studies have investigated the in vitro effect of TTO on herpes simplex virus type 1 and 2
during the different stages of viral replication (Minami et al., 2003; Schnitzler et al., 2001).
Both studies concluded that the oil was more effective on free virus. A pilot in vivo study
showed no statistically significant antiviral effects against herpes simplex virus, possibly
due to small group numbers (Carson et al., 2001). Antiviral activity of TTO has been
shown against tobacco mosaic virus following infection of the leaves of Nicotiniana
glutinosa (Bishop, 1995).
1.2.3 Other properties
1.2.3.1 Anti-inflammatory
The production of pro-inflammatory mediators, including IL-1β, IL-8, IL-10, TNFα and
PGE2, by LPS-activated human monocytes in vitro was suppressed by the more water-
soluble components of TTO, terpinen-4-ol and α-terpineol (Hart et al., 2000). The water-
soluble fraction of TTO suppressed the production of superoxide, an inflammatory
mediator, by agonist-activated monocytes but not neutrophils (Brand et al., 2001). Other
contrasting results have shown exposure to TTO can decrease the production of reactive
oxygen species in both stimulated neutrophils and monocytes, but increase production in
non-stimulated neutrophils and monocytes (Caldefie-Chezet et al., 2004).
When examined in vivo, TTO applied topically was shown to regulate the skin swelling
associated with the efferent phase of a contact hypersensitivity response in mice, but did
not affect the number of inflammatory cells (Brand et al., 2002a). A reduction in histamine-
induced skin swelling was also observed in murine ears following topical treatment with
TTO or terpinen-4-ol (Brand et al., 2002b). The topical application of TTO on areas of
histamine-induced wheal and flare in humans also indicated an anti-inflammatory role
(Khalil et al., 2004). Recent studies have shown that TTO administered to agonist-treated
mice via inhalation can modulate the inflammatory response through the hypothalamic-

Chapter One – Literature Review
13
pituitary-adrenal axis and endogenous opioids (Golab et al., 2005; Golab and Skwarlo-
Sonta, 2007).
1.2.3.2 Anti-protozoan
The growth of the protozoa Trypanosoma brucei and Leishmania major was inhibited
following exposure to TTO at 0.5 mg/ml and 403 mg/ml, respectively (Mikus et al., 2000).
Similarly, Trichomonas vaginalis was killed following exposure to 300 mg/ml of TTO
(Viollon et al., 1996).
1.2.3.3 Anti-parasitic
Treatment of the scabies mite, Sarcoptes scabiei var hominis, with TTO or terpinen-4-ol,
following removal from an infected human, was effective at reducing mite survival times
(Walton et al., 2004). Treatment of ocular demodecosis (eyelash mite) in humans by
weekly lid scrub with 50% TTO and daily use of TTO shampoo caused a reduction in
surface inflammatory signs and eradication of the mite in some patients (Gao et al., 2007).
1.2.3.4 Anti-cancer
TTO or terpinen-4-ol treatment caused the apoptosis of human melanoma M14 cells in
vitro, an effect thought to be mediated by the interaction of the oil or component with the
plasma membrane and subsequent re-organization of membrane lipids (Calcabrini et al.,
2004). Further work by the same group using thermodynamic and structural investigations
of lipid monolayers indicated that TTO interacts with the melanoma cell plasma membrane
but does not inhibit the function of the multi-drug resistant drug transporter P-glycoprotein.
Within the monolayers, TTO interacted preferentially with the less-structured dipalmitoyl
phosphatidyl choline “sea” rather than the more ordered lipid rafts (Giordani et al., 2006).

Chapter One – Literature Review
14
1.2.4 Toxicity
1.2.4.1 Oral
Although once described as a “non-poisonous non-irritant antiseptic” (Penfold, 1929), TTO
may produce toxic effects if taken internally, with an LD50 in the rat of 1.9-2.6 ml/kg
(Russell, 1999). There are numerous case reports of toxicity following ingestion of TTO in
volumes ranging from a teaspoon to half a cup of 100% oil, with symptoms ranging from
confusion, appearance of a rash, diarrhoea and feeling “unwell”, to loss of consciousness
leading to coma (Jacobs and Hornfeldt, 1994; Seawright, 1993).
1.2.4.2 Dermal
1.2.4.2.1 Irritant reactions
The irritant capacity of TTO has been investigated in several studies using occlusive patch
testing in human volunteers. The repeated challenge every 24 h for 21 days with TTO
ranging from 5% to 100%, in different formulations, resulted in no irritant reactions in 25
of 28 volunteers, while the remaining three showed allergic reactions (Southwell et al.,
1997). Other studies patch-testing 1% TTO in 20 patients and 10% TTO in 217 patients,
found no irritant reactions (Knight and Hausen, 1994; Veien et al., 2004).
1.2.4.2.2 Contact allergy
Contact allergy is an eczematous, cutaneous reaction to direct contact with an allergen to
which the individual is hypersensitive (Lewis, 1998). Contact dermatitis has been reported
following topical use of TTO in a range of concentrations (Bhushan and Beck, 1997; De
Groot, 1996; Selvaag et al., 1994). The frequency of allergy to TTO in clinical studies
ranged from 0.97% (3 of 309 participants) to 10.7% (3 out of 28 participants) in the study
by Southwell et al. in 1.2.4.2.1 (Aspres and Freeman, 2003; Southwell et al., 1997).

Chapter One – Literature Review
15
1.2.4.3 Reproductive and developmental toxicity
There have been no studies published on the potential of reproductive or developmental
toxicity following exposure to TTO; however, there are a handful of studies that have
looked at components of the oil. Following investigation into the embryofoetoxicity in rats
of the component α-terpinene, a no observed adverse effect level (NOAEL) for dams and
offspring following ingestion of 30 mg/kg body weight was determined (Araujo et al.,
1996). The component β-myrcene was found to have NOAELs of 250 mg/kg body weight
and 300 mg/kg body weight in two studies investigating toxic effects on fertility and
reproduction in the rat (Delgado et al., 1993; Paumgartten et al., 1998). α-Terpinene is
present in TTO at approximately 10% while β-myrcene is present at approximately 0.5%
(Brophy et al., 1989).
A recent report described three pre-pubertal boys who experienced gynecomastia following
the use of commercial personal care products allegedly containing TTO and lavender oil
(Henley et al., 2007), though the constituents of the products were not analysed. The effects
were reversible several months after use of the products ceased. Subsequent in vitro testing
with human cell lines indicated that both oils had weak endocrine disrupting effects
(Henley et al., 2007).
1.2.5 Mechanisms of antibacterial action
The complex and multi-component nature of TTO suggests that the oil may have more than
one mechanism of antibacterial action – multi-component would likely mean multi-action.
Studies with TTO have shown that one of the mechanisms of action involves alteration of
the bacterial membranes (Carson et al., 2002; Cox et al., 1998; Cox et al., 2000; Cox et al.,
2001). Inhibition of respiration and leakage of K+ ions have been observed in E. coli and
S. aureus, providing evidence of damage to the cytoplasmic membrane (Cox et al., 1998;
Cox et al., 2000). Increased permeability of the cytoplasmic membrane of E. coli to the
fluorescent nucleic acid stain, propidium iodide, was also observed (Cox et al., 2000).
Carson et al. (2002) demonstrated significant loss of 260 nm-absorbing material in
S. aureus following exposure to TTO for 60 min. Nucleic acid components absorb

Chapter One – Literature Review
16
predominantly at 260 nm and the loss of 260 nm-absorbing material is a feature in the
action of many antibacterial agents and is indicative of damage to the cytoplasmic
membrane (Joswick et al., 1971; Silver and Wendt, 1967). Sensitivity to sodium chloride
was seen following treatment with TTO or the components 1,8-cineole, terpinen-4-ol and
α-terpineol, while altered cellular morphology including development of mesosomes
following terpinen-4-ol treatment was observed via electron microscopy (Carson et al.,
2002). The effects mentioned above are consistent with a mechanism of action that targets
the function and integrity of bacterial cell membranes. As yet any further mechanisms of
action of TTO have not been reported.
1.3 Pseudomonas and tea tree oil
As well as a high level of resistance to antibiotics, P. aeruginosa is more resistant to TTO
than most other micro-organisms. The outer membrane of P. aeruginosa plays an
important role in the decreased susceptibility to TTO. Permeabilization of the outer
membrane of P. aeruginosa with polymyxin B nonapeptide (PMBN) or EDTA
significantly decreased the MICs of TTO and terpinen-4-ol (Longbottom et al., 2004; Mann
et al., 2000a). Susceptibility to the less-active component γ-terpinene was also increased
following PMBN treatment (Longbottom et al., 2004). The penetration of predominantly
hydrophobic antibiotics is increased by PMBN treatment (Vaara, 1992). The specific
permeabilizing action of PMBN is not known, but it is not associated with the release of
LPS and may involve structural alterations of the outer membrane bilayer (Vaara, 1992).
EDTA weakens the outer membrane by chelating Mg2+
and Ca2+
ions that link LPS
molecules, resulting in the release of LPS from cells and allowing entry of hydrophobic
antibiotics (Vaara, 1992).
As mentioned previously, the multi-drug efflux systems of P. aeruginosa provide resistance
to a wide array of antimicrobials. Treatment with CCCP, known to inhibit energy-
dependent efflux systems in P. aeruginosa (Li et al., 1998b), increased the susceptibility of
P. aeruginosa to TTO and the components terpinen-4-ol, γ-terpinene and α-terpineol (Cox,

Chapter One – Literature Review
17
2007; Longbottom et al., 2004). Intrinsic organic solvent tolerance in P. aeruginosa has
been associated with the MexAB-OprM system (Li et al., 1998b).
1.4 Resistance to tea tree oil
Given the high frequency with which P. aeruginosa develops resistance to antimicrobial
agents, the possibility exists that continuous exposure to TTO may increase baseline levels
of resistance to TTO. Depending on the mechanisms involved, concurrent resistance to
other antimicrobial agents may also arise. The serial subculture of P. aeruginosa with
increasing concentrations of TTO or terpinen-4-ol created tolerised mutants with reduced
susceptibility to both agents, as well as aztreonam and ticarcillin (Longbottom, 2002).
P. aeruginosa NCTC 10662 was grown in concentrations of up to 2 × the MIC of TTO and
3.75 × the MIC of terpinen-4-ol, though the specific mechanism of reduced susceptibility
was not identified (Longbottom, 2002). Habituation to sub-inhibitory concentrations of
TTO for 72 h reduced the susceptibility of S. aureus to a range of clinically relevant
antibiotics (McMahon et al., 2007). Another study, in which S. aureus was serially
subcultured onto solid media containing increasing concentrations of TTO, reported an
increase in the MIC of TTO from 0.25% to 16% (Nelson, 2000). A recent study reported
the inability to create single-step mutants of S. aureus, S. epidermidis or Enterococcus
faecalis following exposure to TTO at concentrations equal to the MIC or greater (Hammer
et al., 2008).
1.5 Research aims
Resistance to TTO in P. aeruginosa is well-documented. Apart from the barrier function of
the outer membrane, other specific mechanisms involved have not been identified.
Therefore the aims of this research project were to:
Determine the role of efflux pumps in resistance to TTO in P. aeruginosa
Examine how the LPS of P. aeruginosa increases tolerance to TTO
Establish whether resistance to TTO or terpinen-4-ol is increased following
repeated exposure and whether this alters susceptibility to other antimicrobials, and
to determine any mechanisms involved

Chapter One – Literature Review
18
Create a library of transposon mutants of P. aeruginosa and characterize any
mutants with altered susceptibility to TTO or terpinen-4-ol

Chapter Two – Materials and Methods
19
2.0 CHAPTER TWO- MATERIALS AND METHODS
2.1 Materials
2.1.1 Microorganisms
2.1.1.1 Efflux mutants, reference isolates and other strains
P. aeruginosa efflux mutants and parental strains were kindly supplied by Professor Keith
Poole (Queens University, Kingston, Canada) and Professor Herbert Schweizer (Colorado
State University, Fort Collins, USA). Professor Poole also supplied E. coli strains.
Reference microorganisms were obtained from the culture collection of the Discipline of
Microbiology and Immunology, The University of Western Australia. Other strains were
supplied by Professor Joseph Lam (University of Guelph, Guelph, Canada), Professor
Antonio Oliver (Hospital Son Dureta, Palma de Mallorca, Spain) and Dr Harry Sakellaris
(The University of Western Australia, Perth). Details of these isolates are shown in Table
2.1 and Table 2.2.
2.1.2 Chemicals, reagents and media
The chemicals, reagents and desiccated media used in this study, along with their
manufacturers, are shown in Table 2.3.
2.1.3 Culture media
Blood agar and BHIB + 20% glycerol (v/v) were supplied by Excel Laboratories, Western
Australia. Other culture media and their suppliers are presented in Table 2.3. Culture media
were prepared following the manufacturer‟s instructions. The media listed below were
prepared as described and autoclaved at 121°C for 15 min and stored at RT.
Ampicillin, chloramphenicol, kanamycin and tetracycline agar plates
Luria agar 15.25 g

Table 2.1 P. aeruginosa efflux mutants and parental strains
Strain Description Efflux phenotype Reference / Source
PAO11 Parent, MexAB-OprM
+ (Hancock and Carey, 1979; Li et al., 1998a)
PAO2002 Mutant MexAB-OprM
- (Schweizer, 1998)
PAO2382 Mutant MexAB-OprM
- MexCD-OprJ
- (Chuanchuen et al., 2001)
PAO238-12 Mutant MexAB-OprM
- MexCD-OprJ
- MexJK
+ (Chuanchuen et al., 2002)
PAO3142 Mutant MexAB-OprM
- MexCD-OprJ
- MexJK
- (Chuanchuen et al., 2002)
K11191 Mutant MexAB-OprM
- (Li et al., 1998a)
K15211 Mutant MexAB-OprM
+ MexCD-OprJ
- (De Kievit et al., 2001)
K15231 Mutant MexA-OprM
+ MexB
- (Hirakata et al., 2002)
K15251 Mutant MexAB-OprM
+ MexXY
- (De Kievit et al., 2001)
K15361 Mutant (nfxB) MexAB-OprM
+ MexCD-OprJ
++ (Hirakata et al., 2002)
3371 Parent, ML5087 MexAB-OprM
+ (Okii et al., 1983)
K11101 Mutant MexAB
+-OprM
- (Li et al., 1998a)
Legend
(1) Strains provided by Keith Poole; (2) Strains provided by Herbert Schweizer. (-) = not expressed, (+) = expressed, (++) = hyperexpressed
Chap
ter Tw
o –
Materia
ls an
d M
ethods

Table 2.1 P. aeruginosa efflux mutants and parental strains (cont.)
Strain Description Efflux phenotype Reference / Source
K11121 Mutant (nalB) MexAB-OprM
++ (Srikumar et al., 1998)
K11151 Mutant MexAB-OprM
- MexCD-OprJ
- (Li et al., 1998a)
K11171 Mutant (nfxC) MexAB-OprM
- MexCD-OprJ
- MexEF-OprN
++ (Li et al., 1998a)
K11211 Mutant MexAB-OprM
- (Srikumar et al., 1997)
K11311 Mutant (nfxB) MexAB-OprM
- MexCD-OprJ
++ (Srikumar et al., 1997)
K12411 PAO2375, Parent MexAB-OprM
+ (Matsumoto et al., 1981)
K12401 Mutant (nfxC) MexAB-OprM
+ MexEF-OprN
++ (Fukuda et al., 1990)
Legend
(1) Strains provided by Keith Poole; (2) Strains provided by Herbert Schweizer. (-) = not expressed, (+) = expressed, (++) = hyperexpressed
Chap
ter Tw
o –
Materia
ls an
d M
ethods

Table 2.2 Reference and other strains of bacteria
Organism Collection/strain number Features Reference/source
P. aeruginosa NCTC 10662; ATCC 25668 Microbiology & Immunology, UWA
P. aeruginosa PAO1 Parent of rough mutants (Hancock and Carey, 1979); Joseph Lam
P. aeruginosa PAO1rmlC Rough (Rahim et al., 2000); Joseph Lam
P. aeruginosa PAO1wbpL Rough (Rocchetta et al., 1998); Joseph Lam
P. aeruginosa AK1012 Rough (Jarrell and Kropinski, 1981); Joseph Lam
P. aeruginosa PAK Parent (Woods et al., 1997); Joseph Lam
P. aeruginosa PAKrmlC Rough (Rahim et al., 2000); Joseph Lam
P. aeruginosa PAKwbpL Rough (Rocchetta et al., 1998); Joseph Lam
P. aeruginosa PAO1 Parent of mutS Antonio Oliver
P. aeruginosa mutS Hypermutable (Oliver et al., 2004); Antonio Oliver
E. coli K340 pRK415 (Keen et al., 1988); Keith Poole
E. coli K1153 pRSP15 (Srikumar et al., 1998); Keith Poole
E. coli K1154 pRSP17 (Srikumar et al., 1998); Keith Poole
E. coli ED8654 Competent cell line (Murray et al., 1977); Microbiology & Immunology, UWA
E. coli M3488 pUC8, AMPR
Microbiology & Immunology, UWA
E. coli DH5α pBBR1MCS, CAPR
(Kovach et al., 1994); Harry Sakellaris
Chap
ter Tw
o –
Materia
ls an
d M
ethods

Chapter Two – Materials and Methods
Table 2.3 Chemicals, reagents and media
Reagent Manufacturer
Acetic acid Labscan Analytical Sciences, Thailand
Acrylamide BDH Chemicals Ltd, England (BDH
England)
Agarose (PCR) Scientifix, Australia
Agarose (PFGE) Cambrex Bio Science Rockland Inc., US
Ammonium persulphate Bio-Rad Laboratories, USA (Bio-Rad)
Ampicillin Sigma Chemical Company, USA (Sigma)
Barium chloride BDH Chemicals Australia Pty Ltd,
Australia (BDH)
Benzalkonium chloride Fluka, Switzerland (Fluka)
Bis (N,N’-Methylene-bis-acrylamide) Bio-Rad
Bovine serum albumin Commonwealth Serum Laboratories,
Australia
Bromophenol blue Sigma
Cadmium acetate Sigma
Chloramphenicol Sigma
Chlorhexidine Sigma-Aldrich Inc, US (Sigma-Aldrich)
Ciprofloxacin Bayer Australia Ltd., Australia
Citric acid BDH
Cobalt (II) chloride hexahydrate Sigma
Dimethyl sulphoxide BDH
Di-sodium hydrogen orthophosphate BDH
DNAse I Roche Diagnostics, Australia
dNTP Invitrogen Life Technologies, USA
(Invitrogen)
Ethidium bromide Sigma
Formaldehyde BDH

Chapter Two – Materials and Methods
Reagent Manufacturer
Gentamicin Sigma
Glycerol BDH
Glycine Sigma
n-hexadecane BDH
Hexadecyltrimethylammonium bromide
(cetrimide)
Sigma
Histidine Sigma-Aldrich
Irgasan (triclosan) Fluka
Iron sulphate heptahydrate Sigma-Aldrich
Kanamycin ICN Biomedicals Inc, US (ICN) and
AG Scientific, US
Lecithin ICN
Lead (II) nitrate Sigma-Aldrich
Luria agar, Miller Becton Dickinson and Company, US (BD)
Luria broth, Miller BD
Luria-Bertani broth BD
Lysozyme Sigma
Manganous sulphate (monohydrate) BDH
β-mercaptoethanol BDH
Mercuric chloride Sigma-Aldrich
Methanol BDH
Molybdenum trioxide BDH England
Moxalactam Sigma
Mueller-Hinton agar Oxoid Ltd, England (Oxoid)
Mueller-Hinton broth Oxoid
Mueller-Hinton broth, cation adjusted BD
Nalidixic acid Sigma
Nickel (II) chloride hexahydrate Sigma-Aldrich

Chapter Two – Materials and Methods
Reagent Manufacturer
Novobiocin Sigma
Nutrient agar Oxoid
Nutrient broth Oxoid
PCR reaction buffer, 10 × Perkin Elmer by Roche Molecular
Systems Inc, USA (Perkin Elmer)
Phe-arg β-naphthylamide dihydrochloride Sigma
Polymyxin B Sigma-Aldrich
Polymyxin E Sigma
Potassium dihydrogen orthophosphate Ajax Finechem, Australia
Silver nitrate Sigma-Aldrich
Sodium arsenate BDH
Sodium chloride BDH
Sodium dodecyl sulphate BDH
Sodium hydroxide BDH
Sodium sulphite BDH
Sodium thiosulphate BDH
Sucrose BDH
TaqGold DNA polymerase Perkin Elmer
TEMED Bio-Rad
Tris base Invitrogen corporation, US
Tryptone soya broth BD
Tween 80
(polyoxyethylene sorbitan mono-oleate)
Sigma-Aldrich
Ultrapure water Fisher Biotec, Australia
Vancomycin Sigma

Chapter Two – Materials and Methods
20
Distilled water 500 ml
Following autoclaving, agar was cooled to 50°C. The appropriate antibiotic was then added
at the following concentrations:
Ampicillin 100 mg/L
Chloramphenicol 50 or 200 mg/L
Kanamycin 500 mg/L
Tetracycline 10 mg/L
Kanamycin broth
Luria broth 7.75 g
Distilled water 500 ml
Kanamycin 500 mg/L
Kanamycin was added to sterilized broth cooled to room temperature.
2.1.4 Tea tree oil and components
TTO (batch W/E504) was provided by Australian Plantations Pty Ltd (Wyrallah,
Australia). Levels of components determined by gas chromatography mass spectrometry
analysis (by the NSW Department of Agriculture, Wollongbar, NSW, Australia) and the
range specified by the international standard (ISO, 2004) are shown in Table 2.4. Terpinen-
4-ol was provided by SNP Natural Products (Sydney, Australia) and was 100% pure. ρ-
Cymene (purity 99%) and γ-terpinene (purity 97%) were purchased from Aldrich Chemical
Company (Milwaukee, USA). Cineole (purity 99%) and α-terpineol (purity 95%) were
purchased from Sigma Chemical Company (St Louis, USA).
2.1.5 Buffers and other solutions
All buffers and solutions were stored at RT unless otherwise stated.

Chapter Two – Materials and Methods
Table 2.4 Gas chromatography mass spectrometry analysis of tea tree oil batch
W/E504
Component Concentration (%) Range specified by ISO1 (%)
Terpinen-4-ol 40.3 >30
γ-Terpinene 19.7 10-28
α-Terpinene 8.6 5-13
1,8-cineole 3.2 Trace-15
Terpinolene 3.2 1.5-5
α-Terpineol 3.1 1.5-8
α-Pinene 2.4 1-6
ρ-Cymene 2.4 0.2-12
Aromadendrene 1.6 Trace-7
δ-Cadinene 1.2 Trace-8
Limonene 1.0 0.5-4
Globulol 0.5 Trace-3
Viridiflorol 0.4 Trace-1.5
Sabinene 0.1 Trace-3.5
Legend
(1) ISO (2004)

Chapter Two – Materials and Methods
21
2.1.5.1 General
Neutralizer
Solution A
Tween 80 7.5 g
Lecithin 0.75 g
Histidine 0.25 g
Potassium dihydrogen orthophosphate 8.5 g
Tryptone soya broth 7.5 g
Distilled water 245 ml
Solution B
Sodium thiosulphate 1.25 g
Distilled water 5 ml
Solution A was heated to boiling prior to autoclaving. Following autoclaving, the solution
was heated to boiling again then cooled to RT while stirring. Solution B was membrane
filter sterilized then added to solution A (at RT).
Phosphate buffered saline
Sodium chloride 8.0 g
Potassium chloride 0.2 g
Di-sodium hydrogen orthophosphate 1.44 g
Potassium dihydrogen orthophosphate 0.24 g
Distilled water 1 L
The pH was adjusted to 7.4 with HCl prior to autoclaving at 121°C for 15 min.
TE buffer
1M Tris pH7.6 1 ml
0.5M EDTA 0.2 ml
Distilled water 98.8 ml
The buffer was autoclaved at 121°C for 15 min.

Chapter Two – Materials and Methods
22
Sucrose (300mM)
Sucrose 51.34 g
Hi-pure water 500 ml
The solution was membrane filter sterilized and stored at 4°C.
2.1.5.2 SDS-PAGE
Stacking gel buffer
Tris base 30 g
SDW 300 ml
The pH was adjusted to 6.8 with 1N HCl. Volume was made up to 500 ml with SDW.
Buffer was stored at 4°C.
Separating gel buffer
Tris base 91 g
SDW 250 ml
The pH was adjusted to 8.8 with concentrated HCl. Volume was made up to 500 ml with
SDW. Buffer was stored at 4°C.
30% Acrylamide-bis
Acrylamide 29.2 g
N, N-methylene-bis-acrylamide 0.8 g
SDW 100 ml
This was stored at 4°C and protected from light.
10% SDS
Sodium dodecyl sulphate 10 g
SDW 100 ml
SDS-PAGE solubilisation buffer

Chapter Two – Materials and Methods
23
Solution A
Tris HCl 0.757 g
SDW 100 ml
The pH was adjusted to 6.8.
Solution B
Glycerol 10 ml
β-mercaptoethanol 5 ml
Bromophenol blue 10 mg
Sodium dodecyl sulphate 3 g
Solution A 50 ml
The solution was stirred until dissolved, then made up to 100 ml total volume with solution
A.
SDS-PAGE Electrophoresis buffer (5×)
Tris base 9 g
Glycine 43.2 g
Sodium dodecyl sulphate 3 g
SDW 600 ml
The pH was adjusted to 8.3 with HCl. Buffer stock was diluted 1-in-5 in SDW for use.
Silver stain fixing solution
Methanol 200 ml
Acetic acid 25 ml
SDW 275 ml
Silver stain oxidizing solution
Sodium sulphite 20 g
SDW 100 ml
Silver stain solution

Chapter Two – Materials and Methods
24
0.1M Sodium hydroxide 28 ml
Ammonia hydroxide 2 ml
20% Silver nitrate 5 ml
The first two components were mixed then the silver nitrate was added drop by drop while
the solution was stirred. The solution was made up to 150 ml with SDW and used within
5 min of preparation.
Silver stain developer solution
Citric acid 12.5 mg
Formaldehyde 125 μl
SDW 250 ml
Silver stain stop solution
Acetic acid 5 ml
SDW 495 ml
2.1.5.3 PCR
50 × Tris-acetate buffer (TAE)
Distilled water 300 ml
Tris base 121 g
Glacial acetic acid 23.6 ml
0.5M EDTA (ph8) 50 ml
Add more distilled water to a final volume of 500 ml. Solution stored at 4°C.
Ethidium bromide, 10 mg/ml
Ethidium bromide 1 g
Distilled water 100 ml
The solution was stirred for several hours until dissolved then stored at 4°C protected from
light.

Chapter Two – Materials and Methods
25
TAE + ethidium bromide (TAE/EB)
Ethidium bromide, 10 mg/ml 1.25 ml
50 × TAE 500 ml
The solution was stored at 4°C.
PCR electrophoresis buffer
50 × TAE 40 ml
Ethidium bromide, 10mg/ml 100 μl
Distilled water was added to a final volume of 2000 ml and the solution stored at 4°C.
6 × Gel loading buffer
Bromophenol blue 1 g
Sucrose 160 g
Distilled water 400 ml
Solution stored at 4°C.
2.1.5.4 PFGE
Cell suspension buffer
2M Tris, pH 8 50 ml
0.5M EDTA, pH 8 200 ml
Hi Pure water to 1 L
TE buffer (TE)
2M Tris pH 8 10 ml
0.5M EDTA pH 8 4 ml
Hi Pure water to 2 L
Cell lysis buffer (CLB)
2M Tris pH 8 25 ml
0.5M EDTA pH 8 100 ml

Chapter Two – Materials and Methods
26
10% Sarcosyl 100 ml
Hi Pure water to 1 L
A volume of 12.5 μl of 20 mg/ml proteinase K was added to 2.5 ml CLB just before use.
2M Tris pH 8
Tris 157.6 g
Hi Pure water 750 ml
The pH was adjusted and the volume made up to 1 L in Hi Pure water.
0.5M EDTA pH 8
EDTA 186.1 g
Hi Pure water 750 ml
The pH was adjusted and the volume made up to 1 L in Hi Pure water.
10% Sarcosyl
N-Lauroylsarcosine 10 g
Hi Pure water 100 ml
50mM Thiourea
Thiourea 1.903 g
Hi Pure water 500 ml
Following autoclaving, the solution was protected from light and stored at RT.
10 × Tris/Borate/EDTA buffer (TBE)
Tris 216 g
Boric acid 110 g
0.5M EDTA 80 ml
Hi Pure water to 2 L
20 mg/ml Proteinase K
Proteinase K 20 mg

Chapter Two – Materials and Methods
27
Hi Pure water 1 ml
The solution was membrane filter sterilized and stored at -20°C.
2.1.6 Polyacrylamide gels
15% Separating gel
Separating gel buffer 2.5 ml
Acrylamide-bis (30%) 5 ml
10% SDS 100 μl
SDW 2.4 ml
APS 50 μl
TEMED 5 μl
4% Stacking gel
Stacking gel buffer 2.5 ml
Acrylamide-bis (30%) 1.3 ml
10% SDS 100 μl
SDW 6.1 ml
APS 50 μl
TEMED 10 μl
2.1.7 Agarose gels
2.5% agarose gel
Agarose 1.75 g
Distilled water 70 ml
TAE/EB 1.4 ml
The agarose and water were mixed in a 200 ml bottle and heated in a 95°C waterbath for
2 h. The TAE/EB was added and the gel poured into a casting tray with one or two 30 well
combs in place. After setting, gels were kept at 4°C and used within a week. If used
immediately gels were refrigerated for at least 30 min prior to use.

Chapter Two – Materials and Methods
28
1% agarose gel
Agarose 0.7 g
Distilled water 70 ml
TAE/EB 1.4 ml
These gels were made in the same way as the 2.5% gels.
2.1.8 PCR primers
All primers used in this study and their suppliers are shown in Table 2.5. All primers were
reconstituted with ultrapure water and stored at -20°C.
2.1.9 Cellular fatty acid analysis
The following reagents were supplied by the Division of Microbiology & Infectious
Diseases, PathWest Laboratory Medicine WA.
Reagent 1 – Saponification reagent
Sodium hydroxide 45 g
Methanol 150 ml
SDW 150 ml
Reagent 2 – Methylation reagent
Hydrochloric acid (6.00 N) 325 ml
Methanol 275 ml
Reagent 3 – Extraction solvent
Hexane 200 ml
Methyl tert-butyl ether 200 ml
Reagent 4 – Base wash
Sodium hydroxide 10.8 g
SDW 900 ml

Chapter Two – Materials and Methods
Table 2.5 PCR primers used in this study 1
Primer Sequence (5’ – 3’) Reference
KAN-2 FP-1 ACCTACAACAAAGCTCTCATCAACC Epicentre
KAN-2 RP-1 GCAATGTAACATCAGAGATTTTGAG Epicentre
EZKANforward ACGGTTTGGTTGATGCGAGTG This study
EZKANreverse GTCCCGTCAAGTCAGCGTAATG This study
Inv-1 ATGGCTCATAACACCCCTTGTATTA (Ducey and Dyer,
2002)
Inv-2 GAACTTTTGCTGAGTTGAAGGATCA (Ducey and Dyer,
2002)
mexAF1 ACCTACGAGGCCGACTACCAGA (Li et al., 2000)
mexAR1 GTTGGTCACCAGGGCGCCTTC (Li et al., 2000)
mexCF1 AGCCAGCAGGACTTCGATACC (Li et al., 2000)
mexCR1 ACGTCGGCGAACTGCAAC (Li et al., 2000)
mexEF1 GTCATCGAACAACCGCTG (Li et al., 2000)
mexER1 GTCGAAGTAGGCGTAGACC (Li et al., 2000)
mexXF1 CATCAGCGAACGCGAGTACAC (Sobel et al., 2003)
mexXR1 CAATTCGCGATGCGGATT (Sobel et al., 2003)
rpsLF1 GCAACTATCAACCAGGCTG (Li et al., 2000)
rpsLR1 GCTGTGCTCTTGCAGGTTGTG (Li et al., 2000)
mexA1 CGA CCA GGC CGT GAG CAA GCA GC (Dumas et al., 2006)
mexA2 GGA GAC CTT CGC CGC GTT GTC GC (Dumas et al., 2006)
mexC1 ATC CGG CAC CGC TGA AGG CTG CG (Dumas et al., 2006)
Legend
(1) All primers supplied by Geneworks

Chapter Two – Materials and Methods
Table 2.5 (continued) PCR primers used in this study 1
Primer Sequence (5’ – 3’) Reference
mexC2 CGG ATC GAG CTG CTG GAT GCG CG (Dumas et al., 2006)
mexE4 CCA GGA CCA GCA CGA ACT TCT TGC (Dumas et al., 2006)
mexE5 CGA CAA CGC CAA GGG CGA GTT
CAC C
(Dumas et al., 2006)
mexX1 TGA AGG CGG CCC TGG ACA TCA GC (Dumas et al., 2006)
mexX2 GAT CTG CTC GAC GCG GGT CAG CG (Dumas et al., 2006)
rpsL-F GCA AGC GCA TGG TCG ACA AGA (Dumas et al., 2006)
rpsL-R CGC TGT GCT CTT GCA GGT TGT GA (Dumas et al., 2006)
Legend
(1) All primers supplied by Geneworks

Chapter Two – Materials and Methods
29
2.1.10 Disposables and kits
Petri dishes (90 mm) and 96-well trays were manufactured by Becton Dickinson (Franklin
Lakes, USA). Electrocuvettes used in electroporation had a 0.2 cm gap and were supplied
by Invitrogen, GeneWorks. The following kits were used: Qiagen QIAamp DNA mini kit;
EPICENTRE® EZ-Tn5™ <KAN-2> Tnp Transposome™ kit and Promega Wizard® Plus
SV Minipreps DNA Purification system.
2.2 Methods
2.2.1 Susceptibility testing
2.2.1.1 Antibiotic susceptibility testing – broth microdilution
MICs for certain antibiotics were determined using broth microdilution in cation-adjusted
MHB according to CLSI guidelines (CLSI, 2006). Briefly, serial doubling dilutions of
antibiotics, at twice the desired final concentration in SDW, were prepared in 100 µl
volumes across the wells of a 96-well tray. The growth method (CLSI, 2006) was used to
prepare cell suspensions, therefore 3 to 5 well isolated colonies of the same morphologic
type from a BA culture were transferred into 5 ml of TSB. The broths were incubated at
35°C with shaking until cultures achieved or exceeded the turbidity of a 0.5 McFarland
standard (usually 2 to 6 h). The turbidity of each culture was adjusted with saline to achieve
a turbidity equivalent to a 0.5 McFarland standard. This suspension was diluted 1:100 with
double strength cation-adjusted MHB, to give approximately 106 cfu/ml. Following
inoculation of the microtitre tray, the final concentration in each well was approximately 5
× 105 cfu/ml. Viable counts on BA were used to confirm the inoculum. Trays were
incubated at 35°C for 24 h. The MIC was the lowest concentration of antibiotic that
resulted in a visibly detectable reduction of inoculum viability. Tests were performed at
least 3 times and a modal value selected.

Chapter Two – Materials and Methods
30
2.2.1.2 Breakpoint antibiotic testing – agar dilution
Susceptibility of some isolates to antibiotics was determined by agar dilution, performed
according to CLSI guidelines (CLSI, 2006) by the Division of Microbiology & Infectious
Diseases of PathWest as part of their routine clinical microbiology service. Agar plates
containing a series of doubling dilutions of a range of antibiotics, based on CLSI
breakpoints (CLSI, 2007), were supplied by Excel. The direct colony suspension method
(CLSI, 2006) was used to prepare cell suspensions, therefore isolates were inoculated onto
BA and incubated at 37°C for 20-24 h. A suspension of each strain equivalent to a 0.5
McFarland standard was made in 0.85% saline. This suspension was then diluted 1:10 with
¼ strength peptone water (Excel) and aliquoted into a seed tray. Pre-dried agar dilution
plates were inoculated with a MAST replicator device (MAST diagnostics, UK) designed
to deliver 104 cfu of inoculum to each spot. Plates were incubated at 35°C and examined at
24 h and each isolate was classified as sensitive or resistant, based on the CLSI breakpoints
for P. aeruginosa.
2.2.1.3 Antibiotic susceptibility testing – disc diffusion
Susceptibility of some isolates to antibiotics was determined by disc diffusion, performed
according to CLSI guidelines (CLSI, 2006) by the Division of Microbiology & Infectious
Diseases of PathWest as part of their routine clinical microbiology service. The growth
method (CLSI, 2006) was used to prepare cell suspensions, therefore 3 to 5 well isolated
colonies of the same morphologic type from a BA culture were transferred into 5ml of
TSB. The broths were incubated at 35°C with shaking until cultures achieved or exceeded
the turbidity of a 0.5 McFarland standard (usually 2 to 6 h). The turbidity of each culture
was adjusted with saline to achieve a turbidity equivalent to a 0.5 McFarland standard.
This suspension was thoroughly swabbed over the surface of a MHA plate in at least two
directions and antibiotic discs were applied within 15 min. Plates were incubated at 35°C
and examined at 24 h. Susceptibilities were determined according to the CLSI guidelines
(CLSI, 2007).

Chapter Two – Materials and Methods
31
2.2.1.4 Tea tree oil and component MICs
The broth microdilution method used was that described by CLSI (CLSI, 2006) with the
following modification: all tests were performed in MHB supplemented with 0.002%
Tween 80 (MHB-T). Serial doubling dilutions of TTO or components were prepared in
100 µl volumes in a 96-well tray over the range 0.03-8% (v/v). The growth method (CLSI,
2006) was used to prepare cell suspensions, therefore 3 to 5 well isolated colonies of the
same morphologic type from a BA culture were transferred into 5ml of TSB. The broths
were incubated at 35°C with shaking until cultures achieved or exceeded the turbidity of a
0.5 McFarland standard (usually 2 to 6 h). The turbidity of each culture was adjusted with
saline to achieve a turbidity equivalent to a 0.5 McFarland standard. This suspension was
diluted 1:100 with double strength MHB-T, to give approximately 106 cfu/ml. Following
inoculation of the microtitre tray, the final concentration in each well was approximately 5
× 105 cfu/ml. Viable counts on BA were used to confirm the inoculum. Trays were
incubated at 35°C for 24 h. The MIC was the lowest concentration of TTO that resulted in a
visibly detectable reduction of inoculum viability. Tests were performed at least 3 times
and a modal value selected. When testing transposon mutants, the method was adjusted to
incorporate 500 mg/L kanamycin into the microtitre tray and NA supplemented with
500 mg/L kanamycin was used for viable counts.
2.2.1.5 Metal ion MICs
Agar dilution was used to determine MICs for a range of metal ions against P. aeruginosa
isolates. The agar plates contained doubling dilutions of the metal compounds (Table 2.6)
in a Mueller-Hinton agar base and were supplied by Excel. The direct colony suspension
method (CLSI, 2006) was used to prepare cell suspensions, therefore the P. aeruginosa
strains were incubated on BA for 20 – 24 h at 37°C. A suspension equivalent to a 0.5
McFarland standard of each strain was made in 0.85% saline. This suspension was then
diluted 1:10 with ¼ strength peptone water (Excel) and aliquoted into a seed tray. Pre-dried
agar dilution plates were inoculated with a MAST replicator device (MAST diagnostics,
UK) designed to deliver 104 cfu of inoculum to each spot. Plates were incubated at 35°C

Chapter Two – Materials and Methods
Table 2.6 Metal ions and concentrations used in agar dilution
Metal ions Form Concentration range
Arsenic Sodium arsenate heptahydrate 0.3-307.2mM
Barium Barium chloride 0.075-76.8mM
Cadmium Cadmium acetate 0.3-9.6mM
Cobalt Cobalt (II) chloride hexahydrate 0.3-9.6mM
Iron Iron sulphate hexahydrate 0.3-9.6mM
Lead Lead (II) nitrate 0.075-4.8mM1
Manganese Manganous sulphate monohydrate 0.3-307.2mM
Mercury Mercuric chloride 0.075-4.8mM
Molybdenum Molybdenum trioxide 0.3-9.6mM1
Nickel Nickel chloride hexahydrate 0.3-38.4mM
Silver Silver nitrate 0.075-4.8mM
Legend
(1) Due to limited solubility this was the highest concentration tested

Chapter Two – Materials and Methods
32
and examined at 24 and 48 h, with the MIC determined as the lowest concentration that
completely inhibited growth. The experiment was done in duplicate.
2.2.2 Time-kill assays
Ten ml of MHB was inoculated with one colony from a 24 h BA culture of P. aeruginosa
NCTC 10662 and incubated for 16-20 h at 35°C with shaking. This culture was diluted
1:100 with MHB-T to yield a 50ml volume of suspension with approximately 107 cfu/ml. A
9.1ml volume of cell suspension was dispensed into a 50 ml flask for each treatment or
control. Five min prior to time 0, 100 µl was removed, serially diluted in PBS and spread-
plated in duplicate onto pre-dried NA. At time 0, treatment/diluent was added to the flask,
the contents mixed for 20 s and 100 µl removed for serial dilution at 0.5 min. When an oil
or component treatment was used, the first 1:10 dilution was made in neutralizer to arrest
the effect of the oil/component. This first dilution was vortexed and left at RT for at least 5
min to allow neutralization to occur. Following this, the remaining dilutions were made in
PBS. Flasks were incubated at 35°C with shaking for the duration of the assay. NA plates
were incubated at 37°C overnight.
2.2.3 LPS analysis
2.2.3.1 LPS extraction
Five ml of MHB was inoculated with one colony from a 24 h BA culture of P. aeruginosa
and incubated for 24 h at 35°C with shaking. One ml of culture was added to a pre-weighed
lockable Eppendorf tube and centrifuged in a microfuge for 1 min at 6000 g. The
supernatant was discarded and the weight of the bacteria determined. Thirty μl of
solubilisation buffer was added for every mg of bacteria and the cells were resuspended.
The tubes were incubated at 100°C for 10 min with occasional vortexing. Thirty μl of
suspension was added to another Eppendorf tube containing 30 μl of 3.3 mg/ml proteinase
K. Following vortexing, tubes were incubated at 60°C for a minimum of 1 h.

Chapter Two – Materials and Methods
33
2.2.3.2 SDS-PAGE
All polyacrylamide gels were prepared and run using the Bio-rad Mini-PROTEAN II
apparatus. Glass plates were washed and dried in air. The separating gel was prepared by
mixing the first four components (Section 2.1.6) and degassing for at least 15 min. APS and
TEMED were added and the gel poured immediately. The gel was overlaid with water-
saturated isobutanol and left to polymerise for at least 20 min. After the gel was set, the
water-saturated isobutanol was poured off and the top of the gel rinsed with distilled water
and gently blotted dry with filter paper. The 4% stacking gel was prepared in the same
manner, layered over the separating gel and embedded with an 11-well comb to form
sample wells. The stacking gel was left to polymerise for 20 min.
Once the gel had set, the comb was removed and the sample wells rinsed in distilled water
and blotted dry with filter paper. The glass plates were clamped into the electrophoresis
apparatus and surrounded by cold SDS-PAGE electrophoresis buffer. Fifteen μl of each
sample was loaded into sample wells and 10µl of All Blue Precision Plus Protein™
Standard (Bio-Rad) added as a marker. Gels were run at 200V until the dye front was near
the end of the gel.
2.2.3.3 Silver staining
Following electrophoresis, the gel was placed in silver stain fixing solution in a square petri
dish and left at RT overnight. The fixing solution was discarded and the gel covered with
oxidizing solution and left shaking at RT for 5 min. The oxidizing solution was removed
and the gel washed three times in distilled water, shaking at RT each time for 10 min.
During the last wash the silver stain was prepared as in 2.1.5. The gel was covered with
stain and left shaking at RT for 10 min. The gel was washed three times as before then
transferred to a clean square petri dish containing silver stain developer. Following gentle
mixing until bands appeared, the developer was discarded and the gel covered in silver
stain stop solution for 10 min. After a further two washes with distilled water, the gel was
scanned.

Chapter Two – Materials and Methods
34
2.2.4 Transposon mutagenesis
A library of transposon mutants was created using the EZ-Tn5™ <KAN-2> Tnp
transposome™ kit (EPICENTRE® Biotechnologies, Madison, USA). The EZ-Tn5™
transposome complex was randomly inserted into the genome of P. aeruginosa by
electroporation (Figure 2.1). Transposon mutants were selected for by kanamycin resistance
encoded by the transposon. Mutants derived from P. aeruginosa 10662 were numbered
with the prefix „PA' and a 3 digit number, while those derived from P. aeruginosa PAO1
were numbered with the prefix „P‟ and a 3 digit number. The library was then screened for
a change in susceptibility to TTO or terpinen-4-ol and the insertion site of any mutant of
interest was determined by random amplification of transposon ends (RATE) and
sequencing.
2.2.4.1 Cell preparation
Several methods of cell preparation for electroporation were used. The method by Choi
et al. (Method 2) was published during this project (2005) and was determined to be more
effective than Method 1 at generating transposon mutants. Method 2 was used for all
subsequent assays.
Method 1 (based on Smith & Iglewski, 1989)
One colony of P. aeruginosa NCTC 10662 or P. aeruginosa PAO1 from a 24 h BA culture
was used to inoculate 20 ml of LB. Following overnight incubation at 35°C with shaking at
150 RPM on an orbital shaker, the entire broth culture was added to 90 ml of fresh LB and
incubated in the same manner until the OD600 reached 1 (approximately 4 h). Cells were
collected by centrifugation at 1000g for 15 min, washed three times in ice cold 300 mM
sucrose solution and resuspended in 1 ml of ice cold sucrose. The suspension was chilled
on ice for 30 min prior to electroporation.
Method 2 (based on Choi et al., 2005)
One colony of P. aeruginosa NCTC 10662 or P. aeruginosa PAO1 from a 24 h BA culture
was used to inoculate 10 ml of LB. Following overnight incubation at 37°C, four 1.5 ml

Chapter Two – Materials and Methods
Figure 2.1 Strategy for generating and sequencing EZ::TN™ <KAN-2> transposon
insertions in the genome of P. aeruginosa.
Adapted from: http://www.epibio.com/pdftechlit/134pl065.pdf

Chapter Two – Materials and Methods
35
volumes of culture were centrifuged in microfuge tubes at RT at 6000 g. Each pellet was
washed twice in 1 ml of 300 mM sucrose at RT. The four pellets were combined in a total
of 100 µl of 300 mM sucrose. The suspension was kept at room temperature until
electroporation.
2.2.4.2 Electroporation and incubation
Optimization
In order to optimize the electroporation conditions and minimize Transposome™ wastage,
control experiments were done. P. aeruginosa PAO1 was electroporated with pBBR1MCS
to optimize voltage, resistance and cell concentration. E. coli ED8654 was electroporated
with pBBR1MCS or pUC8 to test for any malfunction of the electroporator. pBBR1MCS
was extracted from E. coli DH5α, and pUC8 was extracted from E. coli M3488 using the
Wizard® Plus SV Miniprep kit (Promega) according to manufacturer‟s instructions. The
resulting optimal electroporation conditions are described below.
Biorad Gene Pulser II
A 40µl volume of the chilled suspension was added to a pre-chilled 0.2cm electrocuvette
and electroporated in a Biorad Gene Pulser II (exponential decay generator) at 25 µF,
200 Ω and 2.5 kV/cm for approximately 5 ms. This was the control to ensure the
suspension would not arc. Following this, 40 µl of suspension was added to another pre-
chilled electrocuvette and 1 µl of transposome mix from the EZ-Tn5™ <KAN-2> Tnp
Transposome™ Kit was added and the suspension gently mixed. Electroporation was
performed as above. Immediately following the electroporation, 1 ml of fresh LB at RT was
added to the electrocuvette and mixed gently. The entire contents of the electrocuvette were
transferred to a sterile Bijou bottle and incubated at 37°C for 1 h with shaking to facilitate
cell outgrowth.
ElectroSquarePorator™ EMC830
As part of the optimization of electroporation, a second type of electroporator was used.
The ElectroSquarePorator™ EMC830 (BTX, San Diego, USA) is a square wave

Chapter Two – Materials and Methods
36
electroporation generator with a fixed resistance and capacitance that cannot be varied. The
EMC830 was used with the same electrocuvette / transposome conditions as above to
deliver five 99 µs pulses of 2.5 kV at 200 ms intervals. Post-electroporation incubation was
as above.
2.2.4.3 Plating
Twenty five microlitres of neat, 10-1
and 10-2
dilutions of mutagenesis mix were spread-
plated in duplicate onto pre-dried LA plates containing 500 mg/L kanamycin (LA/KAN). A
control treated the same but without the transposome added was also included. Plates were
incubated at 37°C and checked for growth after 24 and 48 h. The remainder of the
mutagenesis mix was stored at 4°C until the outcome of the experiment was determined.
Following incubation of the initial plates and if the experiment was deemed successful,
more kanamycin plates were spread-plated with the mutagenesis mixture in order to obtain
more mutant colonies. Individual colonies were given a reference number and subcultured
onto LA/KAN and incubated for 24-48 h. Cells were harvested from the plates using sterile
swabs and added to 1 ml of BHIB + 20% glycerol in cryotubes before storage at -80°C.
2.2.4.4 Spontaneous resistance to kanamycin
In order to validate this work, it was necessary to determine the degree of spontaneous
resistance to kanamycin in P. aeruginosa. P. aeruginosa cells were prepared as for
transposon mutagenesis with the following changes: following sucrose washes, cells were
resuspended in 1 ml SDW. Forty microlitres of this suspension was added to 1 ml of LB
broth. A 100 ul volume was spread-plated in triplicate onto pre-dried LA containing 300,
400 or 500 mg/L of kanamycin, respectively. Plates were incubated at 37°C and examined
for growth after 24 and 48 h. A viable count on BA was used to determine the inoculum,
which was approximately 108 cfu/ml.

Chapter Two – Materials and Methods
37
2.2.4.5 PCR detection of kanamycin cassette in transposon mutants
To determine whether mutants created from transposon mutagenesis were true mutants or
spontaneously resistant, it was necessary to detect the presence of the Tn903 kanamycin
resistance gene from the EZ-Tn5™ <KAN-2> transposon.
Primer design and storage
The sequence of the EZ-Tn5™ <KAN-2> transposon was used to design forward and
reverse primers that were specific for a 401bp region in the middle of the transposon, using
the primer design software Prime (GCG) (Figure 2.2). Primers were designated
EZKANforward and EZKANreverse. Primers were obtained from GeneWorks Pty Ltd
(Thebarton, Australia) and were resuspended in ultrapure H2O and stored at -20°C.
DNA extraction
Bacterial DNA was extracted from the transposon mutants by the boiling-lysis method.
Several colonies taken from an LA/KAN plate were resuspended in 1 ml DepC H2O in
sterile Eppendorf tubes. Cell suspensions were boiled at 100°C for 15 min and centrifuged
at 9000 g for 3 min. The supernatant was aliquoted into fresh tubes and stored at -20°C
until further use.
Optimization of PCR conditions
PCR conditions were optimized by testing annealing temperatures of 50, 55 and 60°C, with
and without dimethyl sulfoxide (DMSO) and glycerol. Initially a midrange concentration of
2 mM MgCl2 was chosen and achieved good results, therefore this concentration was not
altered.
PCR conditions
A PCR reaction of 20 µl consisted of 8 µl of template DNA, 0.2 µM of EZKANforward
and EZKANreverse primers, 0.2 mM dNTP, 2 mM MgCl2, 1 × PCR reaction buffer and
0.75 U TaqGold polymerase. The following thermocycler conditions were used:
denaturation for 10 min at 95°C, followed by 45 amplification cycles of 30 s at 94°C, 30 s

Chapter Two – Materials and Methods
1 CTGTCTCTTA TACACATCTC AACCATCATC GATGAATTGT GTCTCAAAAT
51 CTCTGATGTT ACATTGCACA AGATAAAAAT ATATCATCAT GAACAATAAA
101 ACTGTCTGCT TACATAAACA GTAATACAAG GGGTGTTATG AGCCATATTC
151 AACGGGAAAC GTCTTGCTCG AGGCCGCGAT TAAATTCCAA CATGGATGCT
201 GATTTATATG GGTATAAATG GGCTCGCGAT AATGTCGGGC AATCAGGTGC
251 GACAATCTAT CGATTGTATG GGAAGCCCGA TGCGCCAGAG TTGTTTCTGA
301 AACATGGCAA AGGTAGCGTT GCCAATGATG TTACAGATGA GATGGTCAGA
351 CTAAACTGGC TGACGGAATT TATGCCTCTT CCGACCATCA AGCATTTTAT
401 CCGTACTCCT GATGATGCAT GGTTACTCAC CACTGCGATC CCCGGAAAAA
451 CAGCATTCCA GGTATTAGAA GAATATCCTG ATTCAGGTGA AAATATTGTT
501 GATGCGCTGG CAGTGTTCCT GCGCCGGTTG CATTCGATTC CTGTTTGTAA
551 TTGTCCTTTT AACAGCGATC GCGTATTTCG TCTCGCTCAG GCGCAATCAC
601 GAATGAATAA CGGTTTGGTT GATGCGAGTG ATTTTGATGA CGAGCGTAAT
651 GGCTGGCCTG TTGAACAAGT CTGGAAAGAA ATGCATAAAC TTTTGCCATT
701 CTCACCGGAT TCAGTCGTCA CTCATGGTGA TTTCTCACTT GATAACCTTA
751 TTTTTGACGA GGGGAAATTA ATAGGTTGTA TTGATGTTGG ACGAGTCGGA
801 ATCGCAGACC GATACCAGGA TCTTGCCATC CTATGGAACT GCCTCGGTGA
851 GTTTTCTCCT TCATTACAGA AACGGCTTTT TCAAAAATAT GGTATTGATA
901 ATCCTGATAT GAATAAATTG CAGTTTCATT TGATGCTCGA TGAGTTTTTC
951 TAATCAGAAT TGGTTAATTG GTTGTAACAC TGGCAGAGCA TTACGCTGAC
1001 TTGACGGGAC GGCGGCTTTG TTGAATAAAT CGAACTTTTG CTGAGTTGAA
1051 GGATCAGATC ACGCATCTTC CCGACAACGC AGACCGTTCC GTGGCAAAGC
1101 AAAAGTTCAA AATCACCAAC TGGTCCACCT ACAACAAAGC TCTCATCAAC
1151 CGTGGCGGGG ATCCTCTAGA GTCGACCTGC AGGCATGCAA GCTTCAGGGT
1201 TGAGATGTGT ATAAGAGACA G
Figure 2.2 Primer binding sites and PCR target product within Epicentre
EZ::TN™ <KAN-2> sequence. 401bp product = red text, forward primer binding site
= yellow, reverse primer binding site = green. Total = 1221 bp.
Sequence adapted from: http://www.epibio.com/pdftechlit/134pl065.pdf

Chapter Two – Materials and Methods
38
at 60°C and a 45 s extension period at 72°C. Finally, samples were incubated at 72°C for
7 min and held at 4°C until required. All PCR reactions were performed in a Gene Amp
PCR System 9700 (Perkin Elmer Applied Biosystems).
Positive control for PCR
In order to optimize the PCR method, a positive control was needed. To prevent sacrificing
transposome mix, DNA from one of the transposon mutants from the first mutagenesis
(PA100) was used in the PCR optimization steps. The PCR product from a positive reaction
was then sequenced by the Molecular Diagnostics Laboratory of PathWest using the
EZKANforward and EZKANreverse primers. Following sequencing, this product was
found to have 100% homology to the 401 bp region of the transposon that the PCR was
designed to amplify. Therefore in all subsequent PCR reactions, PA100 was included as a
positive control.
Agarose gel electrophoresis
PCR products were visualized by gel electrophoresis on a 2.5% agarose gel containing
ethidium bromide. A mixture of approximately 10 µl of PCR product and 2 µl of 6 ×
loading buffer was loaded into each well. The gels were run at 100 V for approximately 40
min. A 123 bp or 100 bp DNA ladder (Invitrogen) was also included in each gel. Bands
were visualized under UV light and photographed. Transposon mutants that did not yield a
positive in this PCR reaction were discarded from further study.
Control confirming kanamycin cassette is present following -80°C storage
Eight transposon mutants were subcultured from -80°C storage onto KAN/LA. Following
DNA extraction as above, DNA was used as a template in the kanamycin cassette detection
PCR to confirm no loss of the cassette during the storage process.
2.2.4.6 Screening of transposon mutants for TTO susceptibility
Transposon mutants confirmed by PCR were tested for susceptibility to TTO and terpinen-
4-ol as in section 2.2.1.3 with the following changes: transposon mutants were tested in the

Chapter Two – Materials and Methods
39
presence of 500 mg/L kanamycin at all times. All mutants were screened for susceptibility
once and those with an MIC that was ¼ or less, or greater than, that of the parent strain
MIC were tested at least once more to confirm.
A control experiment was performed to demonstrate that a change in TTO susceptibility
was not due to the experimental process itself. In this series of experiments, a cell
suspension of P. aeruginosa was prepared, electroporated and incubated overnight as in
2.2.4.1 – 2.2.4.3, with the modification that no Transposome™ was present during
electroporation. One hundred colonies were selected at random and plated out for single
colonies on LA. These isolates were then stored in BHIB/20% glycerol at -80°C. Following
revival from glycerol stocks, MICs for TTO and terpinen-4-ol for the isolates were
determined as described previously.
2.2.4.7 Identification of disrupted gene in transposon mutants
Several different methods were attempted in the identification of the disrupted gene in the
transposon mutant library. The method recommended by EPICENTRE was direct genomic
sequencing (Hoffman and Jendrisak, 1999) however this could not be optimised. A second
method combined restriction digest with EcoR1 and self ligation, with PCR and sequencing
reactions that both used the KAN-2 FP-1 and KAN-2 RP-1 primers (Rawat et al., 2003) but
this was also unsuccessful. Ultimately the random amplification of transposon ends was
employed.
2.2.4.7.1 Random amplification of transposon ends (RATE) and sequencing
RATE is a three-step single-primer PCR protocol (Ducey and Dyer, 2002). The first and
third steps of the PCR were performed at a stringent annealing temperature while the
second step was performed at 30°C. This allowed non-specific amplification of the single-
stranded product generated in the first step. The third step amplified both specific and non-
specific products (Figure 2.3). This method required considerable optimisation from the
published version.

Chapter Two – Materials and Methods
Figure 2.3 Summary of the random amplification of transposon ends method for
identifying the insertion site in transposon mutants. ME = mosaic end. Modified
from Ducey and Dyer (2002).

Chapter Two – Materials and Methods
40
A 50 µl RATE PCR reaction consisted of 10 µl of template DNA, 0.26 µM of either primer
(Inv-1 or Inv-2), 0.2 mM dNTP, either 1.5, 2 or 2.5 mM MgCl2, 0.01% BSA, 1 × reaction
buffer and 2 U TaqGold polymerase. All template DNA was tested with each of six
possible reaction combinations (either primer with each MgCl2 concentration). The
annealing temperature used was 55°C, if this failed to produce a result then the extracts
were also tested at 50 and 60°C. The thermocycler conditions were as follows: denaturation
at 94°C for 10 min; 30 cycles of 30 s at 94°C, 30 s at annealing temperature and 30 s at
72°C; 30 cycles of 30 s at 94°C, 30 s at 30°C and 30 s at 72°C; 30 cycles of 30 s at 94°C,
30 s at annealing temperature and 120 s at 72°C; 7 min at 72°C; and hold at 4°C. RATE
products were electrophoresed in 1% (w/v) agarose at 100 V for 20 min.
The RATE PCR products were purified to remove unused primers and dNTPs using
ExoSAP-IT (USB Corporation, USA) according to the manufacturer‟s instructions. RATE
PCR products were sequenced using ABI BigDye™ terminators v3.1 (Applied Biosystems,
Foster City, USA) in a 10 µl reaction as follows:
Reaction mix (v3.1) 1 µl
5 × buffer (ABI) 1.5 µl
Primer (1 µM) 3.2 µl
DepC 1.8 µl
RATE product 2.5 µl
For RATE products produced with the Inv-1 primer, the KAN-2 RP-1 primer supplied with
the EZ-Tn5 kit was used in sequencing reactions, while those produced with Inv-2 were
sequenced using KAN-2 FP-1. The cycling conditions for sequencing were 45 cycles of
94°C for 10 s, 55°C for 30 s; followed by a hold at 4°C. Following the sequencing reaction,
the product was purified with a Qiagen DyeEx 2.0 Spin Kit (Qiagen, Doncaster, Australia)
according to the manufacturer‟s directions to remove unincorporated dye terminators. The
sequence reaction products were electrophoresed through the capillary of the ABI PRISM
3100 Avant Genetic Analyzer (Applied Biosystems) and a chromatogram produced
indicating nucleotide sequence.

Chapter Two – Materials and Methods
41
Searches for nucleotide homology to the P. aeruginosa PAO1 genome were carried out
using BLASTN. These sequence data were produced by the Pseudomonas genome project
(Stover et al., 2000) and can be obtained from the Pseudomonas genome databaseV2 at
www.pseudomonas.com.
2.2.5 Serial subculture
P. aeruginosa PAO1 (PAO1) and P. aeruginosa PAO1 mutS (mutS) were subcultured in
increasing concentrations of TTO or terpinen-4-ol in nutrient broth supplemented with
0.002% Tween80 (NBT). A 9 ml volume of NBT containing TTO or terpinen-4-ol was
seeded with 1 ml of overnight culture, such that the final concentration of oil/component
was 0.5%. Following incubation at 37°C with shaking for 24 h, a 10 μl volume was
streaked for single colonies on BA, to confirm purity and check for any morphological
changes. The remaining culture was then centrifuged at 1000 g for 15 min. The pellet was
re-suspended in 1 ml NBT and added to a fresh 9 ml aliquot of TTO/NBT or terpinen-4-
ol/NBT. Cells were passaged every 24-48 h and concentrations of TTO or terpinen-4-ol
were increased in a step-wise fashion according to the levels of growth observed. Isolates
were stored at intervals in BHIB/20% glycerol at -80°C.
PAO1, mutS and all tolerised isolates were subcultured twenty times in the absence of TTO
or terpinen-4-ol. Each isolate and parent strain was plated onto BA and incubated at 37°C
for 24 h; this was repeated 20 times. Isolates were stored after ten and 20 subcultures in
BHIB/20% glycerol at -80°C.
Stored isolates from the highest concentration of TTO or terpinen-4-ol reached from each
parental strain, as well as isolates subcultured in the absence of TTO or terpinen-4-ol, were
examined further. Characterization included susceptibility testing, fatty acid analysis and
hydrophobicity studies, as outlined below.

Chapter Two – Materials and Methods
42
2.2.5.1 Susceptibility testing
Serial subculture mutants were tested for susceptibility to TTO and terpinen-4-ol by broth
microdilution according to 2.2.1.3. Broth microdilution, as described by CLSI (CLSI,
2006), was used to determine MICs for several disinfectants against the serial subculture
mutants and parental strains. Briefly, serial doubling dilutions of disinfectants, at twice the
desired final concentration in SDW, were prepared in 100 µl volumes across the wells of a
96-well tray. The growth method (CLSI, 2006) was used to prepare cell suspensions,
therefore 3 to 5 well isolated colonies of the same morphologic type from a BA culture
were transferred into 5 ml of TSB. The broths were incubated at 35°C with shaking until
cultures achieved or exceeded the turbidity of a 0.5 McFarland standard (usually 2 to 6 h).
The turbidity of each culture was adjusted with saline to achieve a turbidity equivalent to a
0.5 McFarland standard. This suspension was diluted 1:100 with double strength MHB, to
give approximately 106 cfu/ml. Following inoculation of the microtitre tray, the final
concentration in each well was approximately 5 × 105 cfu/ml. Viable counts on BA were
used to confirm the inoculum. Trays were incubated at 35°C for 24 h. The MIC was the
lowest concentration of disinfectant that resulted in a visibly detectable reduction of
inoculum viability. Tests were performed at least 3 times and a modal value selected.
2.2.5.2 Fatty acid analysis
Cellular fatty acid analysis of tolerised mutants was performed by Mr Max Aravena-Roman
from the Division of Microbiology & Infectious Diseases, PathWest. Organisms were
grown overnight on BA plates at 37°C and then subcultured onto TSBA and incubated at
28°C for 24 h. Bacterial cells were harvested then killed and lysed by saponification by
addition of 1 ml saponification reagent and heat. This cleaved fatty acids from the cell
lipids. Addition of 2 ml methylation reagent resulted in conversion into fatty acid methyl
esters (FAME) by methylation, increasing the volatility of the fatty acids for the gas
chromatography analysis. The FAMEs were extracted from the aqueous to the organic
phase using 1.25 ml extraction solvent. The organic solvent phase was removed and treated
with 3 ml base wash to remove free fatty acids and residual reagents prior to
chromatographic analysis.

Chapter Two – Materials and Methods
43
2.2.5.3 Outer membrane permeability
The whole cell alkaline phosphatase assay (Wang et al., 2003) was used to assess outer
membrane permeability of serial subculture mutants and parental strains. An overnight
culture of P. aeruginosa in MHB was washed once with fresh broth and used to inoculate a
3 ml volume of fresh MHB to give an OD600 of approximately 0.1. This was incubated at
35°C on a orbital shaker at 150 RPM for 7 h. Cells were recovered by centrifugation at
1000 g, washed with 5 ml ice-cold reaction buffer and resuspended in 5 ml ice-cold
reaction buffer. The final OD600 was recorded. A 0.1 ml volume of the cell suspension was
added to a reaction mixture containing 0.8ml of reaction buffer and 0.1 ml of 0.08 g/ml
para-nitro phenyl phosphate (pNPP). After a 2 min reaction time, a 0.1 ml aliquot of 1 M
KH2PO4 was added to terminate the reaction. The cells were collected by centrifugation and
OD410 of the supernatant was measured. Samples with no substrate were used as controls.
An index of outer membrane permeability was calculated as: OD410/OD600 [S] where [S] =
the pNPP concentration (0.08).
2.2.5.4 Hydrophobicity
2.2.5.4.1 Flow cytometry
Flow cytometry has been used to measure relative cell surface hydrophobicity in yeast
(Colling et al., 2005). A modified version of this method was used in an attempt to measure
the cell surface hydrophobicity of P. aeruginosa. Uniform dyed hydrophobic microspheres,
0.19 µM diameter and dyed Dragon Green, were obtained from Bangs Laboratories
Incorporated, USA. Overnight cultures of PAO1, mutS and their respective serial subculture
mutants were diluted to a range of concentrations for use in optimisation experiments in
both a BD FACSCanto II and a BD FACSCalibur flow cytometer. The method was not
optimised due to a number of problems including the beads binding too quickly, the small
size of the bacterial cells and differentiating between free and bound beads.

Chapter Two – Materials and Methods
44
2.2.5.4.2 Microbial adhesion to hydrocarbon (MATH)
Several different methods of MATH were attempted. In the first method (Aono and
Kobayashi, 1997) a stationary phase culture was centrifuged at 3000 g and the cells
resuspended in cold 0.8% saline to an optical density at 660 nm of 0.6. This absorbance
was referred to as A1. A 3 ml aliquot of suspension was transferred into two glass Bijou
bottles, 0.6 ml of n-octane was added to one bottle (sample) while the other bottle was the
control. Both suspensions were vortexed vigorously for 2 min then allowed to stand at
room temperature for 15 min, to allow separation into n-octane and saline layers. The
OD660 of the saline phase of the sample (A2S) and the control (A2C) was measured. The
MATH value was calculated as a percentage of the decrease in turbidity of the saline phase
using the following equation:
[(A2C – A2S)/A1] × 100
In the second MATH method (Kobayashi et al., 1999) a stationary phase culture was
diluted 1 in 10 in Luria broth then incubated at 35°C with shaking until the OD660 reached
0.6. The culture was washed twice in 0.8% saline and resuspended in 10 ml of 0.8% saline.
A 4 ml aliquot of the suspension was placed in a glass tube and 0.6 ml of n-hexadecane
added. The tube was vortexed for 1 min and left at room temperature for 10 min. The
lower layer was removed using a glass Pasteur pipette and the OD660 was measured. The
MATH value was calculated as:
% = (1-A/0.6) x 100
Where A = the OD660 of the lower layer.
2.2.5.4.3 Adhesion to polystyrene beads (APSB)
This method was based on that of Aono and Kobayashi (Aono and Kobayashi, 1997). A
stationary phase culture was centrifuged at 3000 g and the cells resuspended in cold 0.8%
saline to an optical density at 660 nm of 0.6. 0.2 g of hydrophobic polystyrene SM-2
Biobeads (100/200 mesh; Bio-Rad) were added to 4 ml of the suspension. The mixture was
stirred gently for 30 s at room temperature and filtered through glass wool. An aliquot of
the suspension lacking Biobeads was used as a control. The OD660 value for the filtrate was

Chapter Two – Materials and Methods
45
referred to as A2S (for the sample) and A2C (for the control). The frequency of cell
adherence to the beads, or APSB value, was determined by the following equation:
[(A2C – A2S)/A1] × 100
2.2.5.5 Pulsed field gel electrophoresis (PFGE)
2.2.5.5.1 Preparation of bacterial plugs
Isolates were grown on MH at 37°C for 17 h. A sterile swab was used to transfer colonies
to Bijou bottles containing 3ml CSB, yielding a suspension equivalent to a 5 McFarland
standard, which was held on ice. A 5 ml volume of pre-melted 1.1% Sea Kem® gold
agarose in TE was mixed with 500 μl of pre-warmed 10% SDS and held at 54°C. A 10 μl
aliquot of 20 g/L proteinase K was added to a sterile Eppendorf tube, then 200 μl of cell
suspension and 200 μl plug agarose was added. After gentle mixing, two plugs were poured
into pre-chilled plug moulds. Plugs were chilled for 20 min at 4°C and then pushed from
their moulds into sterile McCartney bottles containing 2.5 ml lysis buffer with 12.5 μl
proteinase K. Plugs were incubated for 2 h at 54°C with shaking.
2.2.5.5.2 Washing of plugs
The lysis buffer was removed and plugs were washed twice for 15 min in 4 ml of warmed
HiPure water, then four times in 2.5 mls of warm TE, while shaking at 50°C.
2.2.5.5.3 Restriction digest
Buffer was aspirated and the plugs were separated into labeled Eppendorf tubes. One set of
plugs was stored in fresh TE at 4°C. The other set was covered with 300 μl of 1 × Buffer B
restriction enzyme (RE) buffer. After 15 min, this was replaced with 300 μl new Buffer B
containing 0.1 g/L BSA and 2 μl (20 units) SpeI. Plugs were incubated overnight at 37°C.
Following incubation, RE buffer mixture was removed and plugs stored in 1 ml TE at 4°C
until use.

Chapter Two – Materials and Methods
46
2.2.5.5.4 Preparation of plugs for PFGE
A 1% agarose gel was prepared in 0.5 × TBE and pre-electrophoresed in 2.5L of 0.5 × TBE
at 14°C for 1.5 h in a contour-clamped homogenous electric field (CHEF) DR-II apparatus
(Biorad, US). Voltage was 200 V and pulse times were 2-54 s. Plugs were cut in half and
one half stored in TE. The other half was placed in 0.5 × TBE for 3 min prior to running.
2.2.5.5.5 PFGE conditions
The half plugs were loaded, along with Salmonella enterica serovar Braenderup H9812
(ATCC® number: BAA-664™) as a standard and the wells sealed with molten agarose and
left to set. The gel was placed into the tank and secured. Five ml of 50 mM thiourea was
added to the tank and the gel was run for 19 h at the same conditions specified for pre-
electrophoresis.
2.2.5.5.6 Detection of bands
The agarose gel was stained in 10 g/L ethidium bromide for 30 min and destained in
distilled water for 2 × 60 min while shaking. Banding patterns were visualized using a UV
transilluminator and a photograph taken. The gel was then scanned using the GelDoc2000
software (Biorad, US).
2.2.6 Efflux mutants
The efflux mutants and parental strains were analyzed for their susceptibility to TTO and
components by broth microdilution and time-kill assays as described in 2.2.1.3 and 2.2.2.
2.2.6.1 Complementation
Complementation of mutants deficient in MexAB-OprM was achieved by transforming
with pRSP17 (pRK415::mexAB-oprM). The plasmids pRK415, pRSP15 and pRSP17 were

Chapter Two – Materials and Methods
47
extracted from E. coli strains K340, K1153 and K1154, respectively, using the Promega
Wizard® Plus SV Minipreps DNA Purification system according to manufacturer‟s
directions. Plasmid extraction was confirmed by electrophoresis of a 10 μl volume of the
mini-prep in a 1% agarose gel at 100 V for 45 min. Cell suspensions of recipient strains of
P. aeruginosa (K1119 and PAO200) were prepared as in 2.2.4.1 (Method 2) and
electroporated with 10 μl of plasmid mini-prep in a Biorad Gene Pulser II as in 2.2.4.2.
Controls for each were electroporated without plasmid present. Following incubation for
2 h at 37°C to allow cell outgrowth, the entire mixture was plated onto an LA plate
containing 10 mg/L tetracycline.
Ten transformants for each recipient strain electroporated with pRK415 were selected and
stored at -80°C. Eighteen transformants for each recipient strain electroporated with
pRSP17 were selected and stored at -80°C. A plasmid mini-prep confirmed the presence of
the respective plasmids in several of these recipients and these recipients were used in the
next stage of experiments. The MICs of TTO and terpinen-4-ol for the transformants and
parental strains were determined by broth microdilution as in 2.2.1.3, with the addition of
10 mg/L of tetracycline required for transformants.
2.2.6.2 RT-PCR and Real time RT-PCR
It was attempted to assess the expression of a range of efflux pump systems in
P. aeruginosa using reverse transcriptase PCR (RT-PCR) and real time RT-PCR. Initially it
was planned to use RT-PCR to determine whether the non-constitutive Mex efflux systems
of P. aeruginosa were being expressed in the presence of TTO and components. However
during the course of the project and after consultation with the authors of several
publications on the subject, it was decided that real time RT-PCR was a better and more
widely accepted, method for this analysis. Due to funding, equipment and time restrictions,
neither method was able to be optimized. The following outlines the methodology used in
the extensive attempts to optimize both RT-PCR and real time RT-PCR.

Chapter Two – Materials and Methods
48
2.2.6.2.1 Primer optimization – RT-PCR
An initial series of primer pairs were selected from the literature, designed to amplify the
first gene in the operon of MexAB-OprM (mexAF1 and mexAR1, product size 252 bp),
MexCD-OprJ (mexCF1 and mexCR1, product size 314 bp), MexEF-OprN (mexEF1 and
mexER1, product size 516 bp) and MexXY (mexXF1 and mexXR1, product size 414 bp).
Primers designed to amplify the constitutively expressed gene rpsL (rpsLF1 and rpsLR1,
product size 220 bp) were also included as a control. Using genomic DNA from PAO1 and
ML5087, a standard TaqGold PCR was used to optimize annealing temperature, MgCl2
concentration and the presence of co-solvents (glycerol and DMSO) for these primer sets.
2.2.6.2.2 RNA extraction
Cells were collected from a 3 ml volume of a mid-late log culture of P. aeruginosa by
centrifugation and the supernatant discarded. The pellet was resuspended in 100 µl TE
buffer with 0.4 g/L lysozyme and incubated at 37°C for 30 min. RNA was extracted from
the sample using a QIAamp® Viral RNA Mini Kit according to manufacturers directions. A
2 µl aliquot of DNase I was added to each 100 µl RNA volume, incubated at 37°C for 15
min and inactivate at 70°C for 15 min. The sample was re-extracted using the Mini Kit to
remove the DNase I, aliquoted and stored at -80°C.
2.2.6.2.3 SYBR® green
After the decision to use real time RT-PCR, a method incorporating SYBR® green was
developed, based on Dumas et al. (2006). SYBR® green binds non-specifically to double-
stranded DNA, therefore it was necessary to confirm that the primer pairs would only give
one reaction product. To save optimization time, an initial PCR using genomic DNA from
PAO1 and ML5087 was run on a Rotor-Gene™ 6000 real time rotary analyzer (Corbett
research), using the primers and cycle conditions previously optimised for RT-PCR and
incorporating a high resolution melt (HRM) analysis at the end of the PCR. The HRM
characterizes DNA samples according to their dissociation behavior as they transition from
double-stranded DNA to single-stranded DNA with increasing temperature: a single peak

Chapter Two – Materials and Methods
49
in a derivative plot using standard HRM analysis software is indicative of a single product
with no primer dimer or non-specific products (Corbett, 2006). The HRM analysis revealed
more than one product for all primer pairs.
The primers used by Dumas et al. (2006) (Table 2.5) were then obtained and optimized as
in 2.2.5.2.1. These primers were initially used in a TaqGold PCR without SYBR® green,
cycled on the Rotor-Gene™ and then run to gel to confirm the positive or negative reaction.
Further optimisation using a HotStar Taq DNA polymerase and master mix conditions
matched to the Qiagen SYBR® green Quantitect RT-PCR Kit was also done. Primer dimer
was a problem with this set of primer pairs and at this stage this avenue of experimentation
was abandoned.
2.2.6.3 Checkerboard assays
Interactions between TTO and Tween 80 were assessed against several efflux pump
mutants using a broth microdilution checkerboard assay. Tween 80 was serially diluted
across columns 1-11 of a 96 well tray to give concentrations ranging from 0.004 – 4% in a
50 μl volume. Row 12 had SDW with no Tween 80 added. TTO dilutions for each row
were prepared in SDW at four times the desired concentration (0.25 – 16%). Fifty μl of
each TTO dilution was added to each row i.e. 16% to row A, 8% to row B etc. Row H had
SDW with no TTO added. Well 12H was used as a growth control well and contained only
SDW and inocula. Inocula were prepared as in 2.2.1.3, however no Tween 80 was added
and double strength MH was used. A 100 μl aliquot of inoculum was added to every well.
Final concentrations of TTO were 0.06-4% and Tween 80 was 0.001 – 1%. Trays were
incubated at 35°C for 24 h.
Fractional inhibitory concentrations (FICs) were determined as follows:
FICA = MICA alone/MICA in combination
FICB = MICB alone/MICB in combination
∑FIC = FICA + FICB
The criteria for interpreting the ∑FICs were as follows:

Chapter Two – Materials and Methods
50
∑FIC < 0.5 = synergy
∑FIC 0.5 – 4 = indifference
∑FIC > 4 = antagonism
2.2.6.4 Efflux pump inhibition
The compound Phe-Arg-ß-naphthylamide (PAβN), a known inhibitor of several efflux
pumps in P. aeruginosa, was added at 20 mg/L to broth microdilution assays using TTO or
terpinen-4-ol.

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
51
3.0 CHAPTER THREE - THE ROLE OF EFFLUX SYSTEMS IN TOLERANCE OF P. aeruginosa TO TTO AND COMPONENTS
3.1 Introduction
The resistance nodulation/cell division class of multi-drug efflux pumps of
P. aeruginosa, in particular the Mex family, has broad substrate specificity and
contributes to the intrinsic and acquired antibiotic resistance of this opportunistic
pathogen. Apart from antibiotics, the broad substrate range of the Mex efflux systems of
P. aeruginosa also includes organic solvents, biocides, dyes and cell signalling
molecules (Chuanchuen et al., 2003; Li et al., 1998a; Li et al., 1998b; Pearson et al.,
1999), but essential oils have not been explored as potential substrates. Previous work
has shown that the activity of TTO and terpinen-4-ol against P. aeruginosa is enhanced
by the protonophore CCCP (Cox, 2007; Longbottom et al., 2004) implicating an ATP-
dependent efflux mechanism. Mutants of E. coli exhibiting the Mar (multiple antibiotic
resistance) phenotype showed slightly reduced susceptibility to α-terpineol, terpinen-4-
ol and linalool (Gustafson et al., 2001). The Mar locus is hypothesized to mediate
resistance by up-regulating the efflux of substrates of the AcrAB-TolC efflux system in
E. coli, including antibiotics, disinfectants and organic solvents (Randall and
Woodward, 2002).
The hypothesis for this chapter of work was that one or more of the Mex efflux systems
of P. aeruginosa contribute to the tolerance of this organism to TTO and components.
A library of genetically-defined efflux mutants of P. aeruginosa was obtained from
international researchers and the susceptibility of these mutants to TTO and components
was determined by broth microdilution and time-kill assays. The efflux pump inhibitor
PAβN was also tested in conjunction with the MIC assays.

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
52
3.2 Results
3.2.1 Susceptibility of efflux mutants to TTO and components
The susceptibility of the library of efflux mutants used in this project was assessed by
broth microdilution and time-kill assays. The MICs of TTO, terpinen-4-ol, 1,8-cineole,
α-terpineol, ρ-cymene and γ-terpinene were determined. Susceptibility to TTO and
terpinen-4-ol was tested by time-kill; these results are listed below according to the
different efflux systems. All isolates tested had MICs for ρ-cymene and γ-terpinene of
>8% (v/v).
3.2.1.1 MexAB-OprM
3.2.1.1.1 MICs
PAO200, K1119 and K1121, all MexAB-OprM- and the MexB
- and OprM
- strains,
K1523 and K1110, respectively, showed an increase in susceptibility to TTO, terpinen-
4-ol, 1,8-cineole and α-terpineol compared with wild type strains (Table 3.1). This
increase ranged from 4 to 8 times for TTO, 4 to >16 times for terpinen-4-ol, >2 to >4
times for 1,8-cineole and 8 to >16 times for α-terpineol.
Hyper-expression of MexAB-OprM (nalB) in K1112 caused no significant change in
susceptibility to TTO and any changes in component susceptibility were beyond the
detection limits of the assay due to component immiscibility at >8% v/v.
3.2.1.1.2 Time-kill assays
In time-kill assays with MexAB-OprM- strain K1121, treatment with 4% TTO (the MIC
of the ML5087 wild type, equal to four times the MIC of K1121) caused a drop in
viability below the limit of detection of the assay (3× 103 cfu/ml) after 20 min, while
only a 90% decrease in viability was seen in ML5087 after 120 min (Figure 3.1). One
quarter of the TTO MIC for K1121 (0.25%) caused death in 90% of cells after 120 min,
compared with growth in ML5087 cells. Treatment with sub-MIC concentrations of
terpinen-4-ol (0.125%, one quarter of the terpinen-4-ol MIC for K1121) killed >90% of

Table 3.1 TTO and component MICs (%, v/v) by broth microdilution for efflux mutants and parent strains
Strain Efflux phenotype TTO Terpinen-4-ol 1,8-Cineole Alpha-
terpineol
PAO1, wild type MexAB-OprM+ 4 2-4 >8 >8
PAO200 MexAB-OprM- 0.5-1 0.25-0.5 2 1
PAO238 MexAB-OprM-
MexCD-OprJ- 0.5 0.25 0.5 <0.125
PAO238-1 MexAB-OprM
- MexCD-OprJ
-
MexJK+
0.5 0.25 0.5 <0.125
PAO314 MexAB-OprM
- MexCD-OprJ
-
MexJK-
0.25 0.25 0.5 <0.125
K1119 MexAB-OprM- 1 0.25-0.5 2 0.5
K1521 MexAB-OprM+ MexCD-OprJ
- 4 2-4 >8 >8
K1523 MexA-OprM+
MexB- 1 0.5 4 1
K1525 MexAB-OprM+
MexXY- 2-4 2 >8 >8
K1536 MexAB-OprM
+ MexCD-
OprJ++
(nfxB) 4 4 >8 >8
Chap
ter Th
ree - The R
ole o
f Efflu
x Syste
ms in
Tolera
nce o
f P. a
erug
ino
sa to
Tea
Tree O
il and
Co
mp
on
ents

Strain Efflux phenotype TTO Terpinen-4-ol 1,8-Cineole Alpha-
terpineol
ML5087, wild type MexAB-OprM+ 4 >8 >8 >8
K1110 MexAB+-OprM
- 1 0.5 2 0.5
K1112 MexAB-OprM++
(nalB) 2 >8 >8 >8
K1115 MexAB-OprM-
MexCD-OprJ- 1 0.25 2 0.25
K1117 MexAB-OprM
- MexCD-OprJ
-
MexEF-OprN++
(nfxC) 1 0.25 2 0.25
K1121 MexAB-OprM- 1 0.5 4 1
K1131 MexAB-OprM
- MexCD-OprJ
++
(nfxB) 1 0.25 2 0.25
K1241, wild type MexAB-OprM+
2 4 >8 >8
K1240 MexAB-OprM
+
MexEF-OprN++
(nfxC) 2 >8 >8 >8
Note: MIC for all strains for γ-terpinene and ρ-cymene was >8% (v/v).
Table 3.1 (continued) TTO and component MICs (%, v/v) by broth microdilution for efflux mutants and parent strains
Chap
ter Th
ree - The R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erug
ino
sa to
Tea
Tree O
il and
Co
mp
on
ents

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120Time (min)
cfu
/ml
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120Time (min)
cfu
/ml
Figure 3.1 Time-kill curves for P. aeruginosa ML5087 (A, parent), K1121 (B,
MexAB-OprM-) and K1110 (C, MexAB
+-OprM
-) incubated with TTO, terpinen-4-
ol or MHB-T for 120 min at 35°C and shaking. Symbols indicate mean ± standard
deviation of at least two replicates. The limit of detection was 3 × 103 cfu/ml.
Legend: Dark blue, MHB-T; orange, 0.25% TTO; light green, 1% TTO; light
blue, 4% TTO; purple, 0.125% terpinen-4-ol; grey, 1% terpinen-4-ol.
A
B
C

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
53
K1121 after 120 min had no significant effect on ML5087. In ML5087, treatment with
1% terpinen-4-ol, less than one eighth of the MIC, caused a modest decrease in viability
of approximately 7.5-fold after 120 min.
A decrease in viability below the limit of detection occurred following treatment with
the MIC of TTO for K1110 (1%) after 10 min, while 0.25% TTO caused death in ~99%
of cells after 120 min (Figure 3.1). Approximately 99% of K1110 cells were killed
following treatment with 0.125% terpinen-4-ol for 120 min.
3.2.1.2 MexCD-OprJ
3.2.1.2.1 MICs
The K1131 isolate, a MexCD-OprJ over-expressing (nfxB) isolate derived from K1121
(MexAB-OprM-), had a similar susceptibility pattern to TTO, terpinen-4-ol, 1,8-cineole,
α-terpineol as K1121 (Table 3.1). The only difference was the decrease in α-terpineol
MIC from 1% to 0.25%. The susceptibility of K1131 was also the same as the MexCD-
OprJ- MexAB-OprM
- strain K1115. The nfxB isolate K1536, that is MexAB-OprM
+ and
hyper-expresses MexCD-OprJ, did not differ in susceptibility from the parent PAO1.
PAO238, a MexCD-OprJ- isolate derived from PAO200 (MexAB-OprM
-), had similar
susceptibility to TTO and terpinen-4-ol as PAO200. In contrast, the MIC for 1,8-cineole
for PAO238 decreased by 4-fold and the α-terpineol decreased by a factor greater than 8
while K1521, a MexCD-OprJ- isolate derived from PAO1, had the same susceptibility
pattern as PAO1.
3.2.1.2.2 Time-kill curves
After 120 min treatment with 0.25% TTO, equal to one quarter the MIC for this efflux
mutant, the viability of K1115 had dropped more than 99.9% to a level just above the
limit of detection of the assay (Figure 3.2). Following 30 min treatment of K1115 with
0.125% terpinen-4-ol (half the MIC), viable cells were no longer detectable with this
assay.

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
Figure 3.2 Time-kill curves for ML5087(A, parent), K1115 (B, MexAB-OprM-
MexCD-OprJ-) and K1131 (C, MexAB-OprM
- MexCD-OprJ
++ [nfxB]) incubated
with TTO, terpinen-4-ol or MHB-T for 120 min at 35°C and shaking. Symbols
indicate mean ± standard deviation of at least two replicates. The limit of detection
was 3 × 103 cfu/ml.
Legend: Dark blue, MHB-T; orange, 0.25% TTO; light green, 1% TTO; light
blue, 4% TTO; purple, 0.125% terpinen-4-ol; grey, 1% terpinen-4-ol.
A
B
C

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
54
The viability of efflux mutant K1131 dropped below the limit of detection after 10 min
treatment with 1% TTO, 30 min treatment with 0.125% terpinen-4-ol and 60 min
treatment with 0.25% TTO (Figure 3.2).
Both PAO1 and K1536 had similar levels of susceptibility to TTO and terpinen-4-ol in
time-kill curves (Figure 3.3). Treatment with 0.25% TTO had comparable levels of
growth to the MHB-T controls for both the wild-type and the mutant strain, while 1%
TTO caused up to 90% cell death after 120 min. Sub-inhibitory levels of terpinen-4-ol
(0.125%) caused little cell death in either strain after 120 min.
3.2.1.3 MexXY
3.2.1.3.1 MICs
K1525, a MexXY- derivative of PAO1, had susceptibility to TTO, terpinen-4-ol, 1,8-
cineole and α-terpineol that was comparable to PAO1 (Table 3.1). The MIC for TTO
and terpinen-4-ol for K1525 was 2%, while for 1,8-cineole and α-terpineol it was >8%.
3.2.1.4 MexEF-OprN
K1240, a MexEF-OprN over-expressing (nfxC) derivative of K1241 (MexAB-OprM+),
was similar in susceptibility to K1241, with the exception of an increase in MIC for
terpinen-4-ol from 4% (K1241) to >8% (Table 3.1). The K1117 isolate, an nfxC
derivative of K1115 (MexCD-OprJ- MexAB-OprM
-), had the same susceptibility profile
as K1115.
3.2.1.5 MexJK
The isolates PAO238-1 (MexAB-OprM-, MexCD-OprJ
-,
MexJK+)
and PAO314
(MexAB-OprM-, MexCD-OprJ
-, MexJK
-) had almost identical susceptibility profiles to
their parental isolate, PAO238 (MexAB-OprM-, MexCD-OprJ
-) (Table 3.1).

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
Figure 3.3 Time-kill curves for P. aeruginosa PAO1 (A, parent) and K1536 (B,
MexAB-OprM+ MexCD-OprJ
++ [nfxB]) incubated with TTO, terpinen-4-ol or
MHB-T for 120 min at 35°C and shaking. Symbols indicate mean ± standard
deviation of at least two replicates. The limit of detection was 3 × 103 cfu/ml.
Legend: Dark blue, MHB-T; orange, 0.25% TTO; light green, 4% TTO; purple,
0.125% terpinen-4-ol.
Chap
ter Fo
ur - T
he R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erugin
osa
to T
ea T
ree Oil a
nd
Co
mp
on
ents
Chap
ter Fo
ur - T
he R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erugin
osa
to T
ea T
ree Oil a
nd
Co
mp
on
ents
Chap
ter Fo
ur - T
he R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erugin
osa
to T
ea T
ree Oil a
nd
Co
mp
on
ents
Chap
ter Fo
ur - T
he R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erugin
osa
to T
ea T
ree Oil a
nd
Co
mp
on
ents
Chap
ter Fo
ur - T
he R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erugin
osa
to T
ea T
ree Oil a
nd
Co
mp
on
ents
A
B

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
55
3.2.2 Complementation
The plasmids pRK415, pRSP15 and pRSP17 were extracted from E. coli strains K340,
K1153 and K1154, respectively (Figure 3.4).
The mutants PAO200 and K1119 (both MexAB-OprM-) were transformed with
pRSP17 (pRK415::mexAB-oprM) with an efficiency of approximately 2.5 × 102 cfu/µg
DNA and 1.7 × 102 cfu/µg DNA, respectively. Eighteen potential transformants for
each recipient were selected (given the numbers K1119-11 to K1119-28 and PAO200-
11 to PAO200-28) and stored in BHIB/glycerol at -80°C. Electroporation of PAO200
and K1119 with pRK415 was also done for control purposes, with an efficiency of
approximately 5 × 105 cfu/µg DNA and 2.8 × 10
4 cfu/µg DNA, respectively (one
transformant for each was selected, numbered K1119-1 and PAO200-1). An initial five
potential transformants from each recipient were chosen for further study and a plasmid
mini-prep prepared of each. K1119-14 was discarded due to a lack of pellet during the
mini-prep and PAO200-13 failed to resuspend properly. The remaining 8 mini-preps
were run on a 1% agar gel, along with a mini-prep of pRSP17 (Figure 3.5).
It appeared from the gel that pRSP17 was present in transformants K1119-11, K1119-
13, K1119-15, PAO200-11 and PAO200-15.
An initial screening test was done to assess the susceptibility of the five transformants
to TTO and terpinen-4-ol (Table 3.2). The TTO and terpinen-4-ol susceptibility of the
transformant PAO200-15 appeared to be restored to wild type levels and this
transformant was selected for further susceptibility profiling. This further testing of
PAO200-15 revealed that the MICs for TTO, terpinen-4-ol, 1,8-cineole and α-terpineol
were the same as those of PAO1 (Table 3.3). The addition of pRK415 alone (in
PAO200-1) did not return the susceptibility to wild-type levels.
Although it was initially planned to use pRSP15 (pRK415::mexCD-oprJ) in
complementation studies, because MexCD-OprJ is not constitutively expressed in
P. aeruginosa, this would only be useful if the expression of the pump could be induced
and detected (see RT-PCR section 3.25 for more details).

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
Figure 3.4 Plasmid mini-prep extraction electrophoresis results.
Ten µl plasmid mixed with 2 µl of 6 × loading buffer and run in a 1% agarose gel
at 80 V for 45 min. Lanes: M, 100 bp marker (each band indicates a further
100 kb, starting from 100 kb for the bottom band); 1, pRK415; 2, pRSP15; 3,
pRSP17.
M 1 2 3
Chap
ter Fo
ur - T
he R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erugin
osa
to T
ea T
ree Oil a
nd
Co
mp
on
ents
M 1 2 3

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
Figure 3.5 Plasmid mini-preps from putative transformants.
Ten µl plasmid mixed with 2 µl of 6 × loading buffer and run in a 1% agarose gel
at 100 V for 40 min. Lanes: M, 100 bp marker (each band indicates a further
100 kb, starting from 100 kb for the bottom band); 1, K1119-11; 2, K1119-12; 3,
K1119-13; 4, K1119-15; 5, PAO200-11; 6, PAO200-12; 7, PAO200-14; 8, PAO200-
15; 9, pRSP17.
M 1 2 3 4 5 6 7 8 9
Chap
ter Fo
ur - T
he R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erugin
osa
to T
ea T
ree Oil a
nd
Co
mp
on
ents

Table 3.2 Initial susceptibility screening of transformants to TTO and terpinen-4-ol (%, v/v)
Isolate Features TTO MIC Terpinen-4-ol
MIC
PAO1 Parent 2-4 2
PAO200 PAO1 ∆mexAB-oprM 0.5 0.5
PAO200-1 PAO200/pRK415 0.5 0.25
PAO200-11 PAO200/pRSP17 (pRK415::mexAB-oprM) 0.5 0.5
PAO200-15 PAO200/pRSP17 (pRK415::mexAB-oprM) 2 4
K1119 PAO1 ∆mexAB-oprM 1 0.5
K1119-1 K1119/pRK415 0.5 1
K1119-11 K1119/pRSP17 (pRK415::mexAB-oprM) 1 2
K1119-13 K1119/pRSP17 (pRK415::mexAB-oprM) 0.5 2
K1119-15 K1119/pRSP17 (pRK415::mexAB-oprM) 1 2
Chap
ter Th
ree - The R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erug
ino
sa to
Tea
Tree O
il and
Co
mp
on
ents

Table 3.3 TTO and component MICs for complemented strains and their parent isolates1 (%, v/v)
Isolate Features TTO MIC Terpinen-4-ol
MIC Cineole MIC
Alpha-terpineol
MIC
PAO1 Parent 2-4 2 >8 >8
PAO200 PAO1 ∆mexAB-oprM 0.5 0.25-0.5 2 1
PAO200-1 PAO200/pRK415 0.5 <0.125 1 0.5
PAO200-15 PAO200/pRSP17
(pRK415::mexAB-oprM) 2 2-4 >8 >8
Legend
(1) Modal value selected from six experiments
Chap
ter Th
ree - The R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erug
ino
sa to
Tea
Tree O
il and
Co
mp
on
ents

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
56
3.2.3 Efflux pump inhibition
Before the efflux pump inhibitor PAβN could be used in broth microdilution assays
with TTO or components, the MIC for PAβN against the efflux mutants had to be
determined. Broth microdilution assays showed the MICs for all mutants and parent
strains tested were 160 or 320 mg/L (Table 3.4.). The concentration of PAβN previously
reported to inhibit efflux pumps was 20 mg/L (Jiang et al., 2006). In order to confirm
the activity of PAβN, the susceptibility of wild-type strains PAO1 and ML5087 to
chloramphenicol and tetracycline, two antibiotics normally effluxed from
P. aeruginosa, was determined in the presence and absence of 20 mg/L PAβN. The
MIC of chloramphenicol was 32 mg/L for PAO1 and 128 mg/L for ML5087; the
addition of PAβN decreased these values to 2 mg/L and 1 mg/L, respectively. Similarly,
the tetracycline MIC was 16 mg/L for both isolates; the addition of PAβN decreased the
MIC to 4 mg/L for PAO1 and 2 mg/L for ML5087. Based on this, 20 mg/L of PAβN
was used in TTO and component MIC assays.
The addition of 20 mg/L of PAβN to MIC broth microdilution assays decreased the
TTO and terpinen-4-ol MICs of PAO1 to levels similar to those in MexAB-OprM-
(PAO200, K1119) and MexB- (K1523) strains (Table 3.5). The addition of PAβN made
little difference to the TTO MIC for PAO200, K1119 and K1523, with at most a 2- to 4-
fold decrease in MIC for K1119. There were further increases in the susceptibility of
these three strains to terpinen-4-ol when PAβN was present, with at least a greater than
4-fold decrease in MIC observed. The presence of PAβN decreased the 1,8-cineole MIC
for PAO1 from >8% to 0.25%, while for K1119 and PAO200 the MIC decreased from
2% to 0.25% (K1523 not done). Similarly, the α-terpineol MIC was decreased in the
presence of PAβN to 0.125% for PAO1 (>64-fold decrease), K1119 (4-fold decrease)
and PAO200 (8-fold decrease) (Table 3.5).
When the MICs for the MexCD-OprJ- isolate K1521 were determined in the presence of
PAβN, the MIC for TTO, α-terpineol, terpinen-4-ol and 1,8-cineole decreased to levels
similar to PAβN-treated PAO1 (Table 3.5). A similar pattern occurred with the nfxB
(MexCD-OprJ hyper-expressing) strain K1536, with PAβN treatment decreasing

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
Table 3.4 Susceptibility of efflux mutants and parental strain of P. aeruginosa to
the efflux inhibitor PAβN
Isolate Phenotype MIC (µg/ml)
PAO1 Parent 320
K1119 PAO1 ∆mexAB-oprM 160
PAO200 PAO1 ∆mexAB-oprM 160
K1521 PAO1 ∆mexCD-oprJ 320
K1523 PAO1 ∆mexB 160
K1525 PAO1 ∆mexXY 320
K1536 PAO1 nfxB 320

Table 3.5 MICs (% v/v) of TTO, terpinen-4-ol and 1,8-cineole against P. aeruginosa PAO1 and efflux mutants, with and without
20 µg/ml PAβN
Isolate Genotype PAβN TTO Terpinen-4-ol 1,8-cineole α-terpineol γ-terpinene ρ-cymene
PAO1 Parent - 4 2-4 >8 >8 >8 >8
+ 0.5 0.25 0.25 0.125 >8 1-2
K1119 PAO1 ∆mexAB-oprM - 1 0.25-0.5 2 0.5 >8 >8
+ 0.25-0.5 <0.125 0.25 0.125 >8 4
PAO200 PAO1 ∆mexAB-oprM - 0.5-1 0.25-0.5 2 1 >8 >8
+ 0.5 <0.125 0.25 0.125 8 2
K1521 PAO1 ∆mexCD-oprJ - 4 2-4 >8 >8 >8 >8
+ 1 0.25 0.125 0.125 >8 4
K1523 PAO1 ∆mexB - 1 0.5 4 1 >8 >8
+ 0.5 <0.125 ND1 ND >8 2
K1525 PAO1 ∆mexXY - 2-4 2 >8 >8 >8 >8
+ 1 <0.125 0.125 0.125 >8 4
K1536 PAO1 nfxB - 8 4 >8 >8 >8 >8
+ 0.5 <0.125 0.125 0.125 >8 2
Legend
(1) ND, not done.
Chap
ter Th
ree - The R
ole o
f Efflu
x System
s in T
olera
nce o
f P. a
erug
ino
sa to
Tea
Tree O
il and
Co
mp
on
ents

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
57
susceptibility to TTO, α-terpineol, terpinen-4-ol and 1,8-cineole to levels similar to
PAβN-treated PAO1 (Table 3.5).
The susceptibility of all isolates tested to ρ-cymene and γ-terpinene alone was beyond
the limits of the broth microdilution assay, with the MIC >8% for all. Susceptibility to
γ-terpinene was not altered by the presence of PAβN in the assay, although any change
may have been beyond the limit of the assay (Table 3.5). The one exception was
PAO200 with a decrease in the MIC detected from >8% to 8%. Susceptibility to
ρ-cymene was increased in all isolates when PAβN was present (Table 3.5). The
ρ-cymene MIC for PAO1, in the presence of PAβN, was 1%, while for PAO200, K1523
and K1536 it was 2% and for K1119, K1521 and K1525 it was 4%.
3.2.4 Checkerboard assays with Tween 80
To determine whether Tween 80 had any effect on the susceptibility of the efflux
mutants to TTO, interactions between TTO and Tween 80 were assessed against a
selection of efflux pump mutants using a broth microdilution checkerboard assay. In all
mutants tested, along with the parent strain PAO1, the MIC for Tween 80 was >1%, the
highest concentration tested in the assay. The TTO MICs were the same no matter what
concentration of Tween 80 was present. Therefore, the sum of fractional inhibitory
concentrations was 2 for all isolates (PAO1, K1119, K1521, K1523 and K1525). This
indicates an “indifferent” result, i.e. neither synergistic nor antagonistic.
3.2.5 RT-PCR and Real time RT-PCR
The initial series of primer pairs chosen for RT-PCR were optimized using standard
PCR techniques (Figures 3.6 and 3.7). It was determined that the optimal conditions for
mexAF1, mexAR1, rpsLF1 and rpsLR1 were 2 mM MgCl2, co-solvents present and a
60°C annealing temperature. For mexCF1, mexCR1, mexEF1, mexER1, mexXF1 and
mexXR1 conditions were 2 mM MgCl2, co-solvents present and a 55°C annealing
temperature. Following a review of the literature RT-PCR was abandoned for real time
RT-PCR using SYBR® green (Dumas et al., 2006).

Chapter Three - The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
50°C 55°C 60°C 60°C + co-solvents
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
50°C 55°C 60°C 60°C + co-solvents
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
50°C 55°C 60°C
1 2 3 4 1 2 3 4 1 2 3 4
45°C + 50°C + 55°C + 60°C + co-solvents co-solvents co-solvents co-solvents
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Figure 3.6 PCR optimization of Mex primers.
(a) RpsL, (b) MexA, (c) MexC. Lanes: 1, PAO1 neat; 2, PAO1 diluted 1 in 10; 3,
ML5087 neat; 4, ML5087 diluted 1 in 10. All products were the expected size.
(a) RpsL
(b) MexA
(c) MexC

Chapter Three - The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
50°C 55°C 60°C
1 2 3 4 1 2 3 4 1 2 3 4
45°C + 50°C + 55°C + 60°C + co-solvents co-solvents co-solvents co-solvents
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
50°C 55°C 60°C
1 2 3 4 1 2 3 4 1 2 3 4
45°C + 50°C + 55°C + 60°C + co-solvents co-solvents co-solvents co-solvents
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Figure 3.7 PCR optimization of Mex primers.
(a) MexE, (b) MexX. Lanes: 1, PAO1 neat; 2, PAO1 diluted 1 in 10; 3, ML5087
neat; 4, ML5087 diluted 1 in 10. All products were the expected size.
(a) MexE
(b) MexX

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
58
In an effort to minimize optimization time, the initial series of RT-PCR primers was
used in a SYBR® green PCR using TaqGold, with HRM analysis. HRM analysis
revealed more than one product for all primer pairs, this was confirmed when the
products were run on a 3% gel (Figure 3.8). Further study with these primers was
abandoned.
A second set of primers, used by Dumas et al. (2006) (see Table 2.5), was obtained and
optimization of annealing temperature and MgCl2 concentration was attempted in a
standard TaqGold PCR in both a Gene Amp PCR System 9700 and a Rotor-Gene™
6000 (results not shown). It appeared that 60°C was the optimum annealing temperature
but a MgCl2 concentration was not determined for all primer sets at this stage. The
Dumas group (Dumas et al., 2006) used a Qiagen SYBR® green Quantitect Kit for RT-
PCR however, due to budget restrictions we were not able to purchase this kit. The kit
combines HotStar Taq DNA polymerase with SYBR® green; we were able to source
some HotStar Taq from another laboratory and used the information on the Qiagen kit
available online to create a PCR master mix. PCR optimisation using the Rotor-Gene™
6000 and the HotStar/SYBR® green mix indicated a 65°C annealing temperature and
the following MgCl2 concentrations: 2.5 mM for mexC1 & mexC2 and mexX1 &
mexX2 and 4 mM for mexA1 & mexA2, mexE4 & mexE5 and rpsL-F & rpsL-R. HRM
analysis indicated that primer dimer was also a problem with these primer pairs.
At this point due to time restrictions this avenue of experimentation was abandoned.

Chapter Three - The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
Figure 3.8 Test SYBR® green TaqGold PCR using first set Mex primers.
Lanes: M = 50 bp marker; 4, 8, 12, 16, 20, 24, 28, 32, 36, 40 = DepC; 1-3 = MexE,
PAO1 neat to 10-2
; 5-7 = MexE, ML5087 neat to 10-2
; 9-11 = MexX, PAO1 neat to
10-2
; 13-15 = MexX, ML5087 neat to 10-2
; 17-19 = MexC, PAO1 neat to 10-2
; 21-23
= MexC, ML5087 neat to 10-2
; 25-27 = RpsL, PAO1 neat to 10-2
; 29-31 = RpsL,
ML5087 neat to 10-2
; 33-35 = MexA, PAO1 neat to 10-2
; 37-39 = MexA, ML5087
neat to 10-2
. Annealing temperature for MexE, MexX and MexC was 60°C;
annealing temperature for RpsL and MexA was 55°C. Mix contained 0.75 u/tube
TaqGold, 4 mM MgCl2, 5% glycerol and 5% DMSO.
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 M
M 29 30 31 32 33 34 35 36 37 38 39 40
50bp
200bp
50bp
200bp

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
59
3.3 Discussion
This investigation into the role of efflux systems in the tolerance of P. aeruginosa to TTO
and some of its components attempted to identify which, if any, of the Mex systems
contribute to this tolerance. The susceptibility of a library of defined Mex mutants was
determined for TTO and a range of components. Complementation was used to confirm the
role of MexAB-OprM and the efflux inhibitor PAβN was used to further investigate the
function of the Mex systems.
3.3.1 MexAB-OprM
MexAB-OprM is the only pump from the Mex family that is constitutively expressed in P.
aeruginosa (Poole, 2001); this facilitated the analysis of the role of MexAB-OprM in
tolerance to TTO and components. Susceptibility to TTO and the components terpinen-4-
ol, 1,8-cineole and α-terpineol was increased in mutants missing the MexAB-OprM pump
or the OprM component of the pump. The hyper-expression of MexAB-OprM did not
further increase the MIC for TTO, suggesting that this was not a mechanism that could lead
to even greater tolerance to TTO in P. aeruginosa. The similar pattern of susceptibility to
TTO and terpinen-4-ol observed in time-kills with K1121 (MexAB-OprM-) and K1110
(MexAB+-OprM
-) indicated that loss of OprM had the same effect as loss of the entire
operon. A similar trend, though with MexB, occurred in MIC assays with K1119 (MexAB-
OprM-) and K1523 (MexA-OprM
+ MexB
-).
Complementation studies showed that addition of the mexAB-oprM operon to a mexAB-
oprM deletion mutant restored the TTO, terpinen-4-ol, 1,8-cineole and α-terpineol
susceptibility to that of the wild type, confirming the role of MexAB-OprM in tolerance to
TTO and these components. The screening assay determined that only one strain, PAO200-
15, was successfully complemented. There are several possible explanations for this. The
plasmid may have been lost during the broth microdilution screening assay. Although
tetracycline was present in the test medium, the presence of toxic TTO or its components
may have affected the stability of the plasmid. If the plasmid stayed in the bacterial

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
60
cytoplasm the mexAB-oprM fragment may have looped out of the plasmid, leaving only
pRK415 without the fragment inserted. While the loss of an insert is proportional to the
plasmid copy number (Bowers et al., 2004), pRK415 is a low copy number plasmid (Keen
et al., 1988) and so less likely to cause failure of complementation in this case.
This is the first report of terpenes as substrates of the MexAB-OprM pump in
P. aeruginosa. The addition of these terpenes to the long list of substrates of MexAB-OprM
is not surprising – the substrates of this pump range in both structure and mechanism of
action and include the organic solvents n-hexane and p-xylene (Li et al., 1998b).
Interplay between the MexAB-OprM and MexCD-OprJ pumps may contribute towards
tolerance of some components of TTO, including 1,8-cineole and α-terpineol. This is
discussed below.
3.3.2 MexCD-OprJ
MexCD-OprJ is not constitutively expressed in P. aeruginosa. Expression of MexCD-OprJ
can be induced by disinfectants such as benzalkonium chloride and chlorhexidine
gluconate, although the antibiotics norfloxacin, tetracycline, chloramphenicol,
streptomycin, erythromycin and carbenicillin are substrates, but not inducers, of MexCD-
OprJ (Morita, 2003).
The concurrent loss of the MexAB-OprM and MexCD-OprJ pumps provided some
evidence for a role for the MexCD-OprJ pump in terpene efflux. In PAO238 (MexAB-
OprM- and MexCD-OprJ
-) the 1,8-cineole MIC was one quarter that of the PAO200
(MexAB-OprM-) strain, while the α-terpineol MIC dropped from 1% to <0.125%. The
TTO and terpinen-4-ol MICs were slightly altered. Similarly, 2-4-fold reductions in MICs
of 1,8-cineole and α-terpineol were seen in K1115 (MexAB-OprM- and MexCD-OprJ
-)
compared to K1121 (MexAB-OprM-). Again, the TTO and terpinen-4-ol MICs remained
almost the same. Whether the loss of MexCD-OprJ alone in a MexAB-OprM+ strain
(K1521) altered the susceptibility to 1,8-cineole, α-terpineol, ρ-cymene or γ-terpinene was

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
61
unable to be determined due to the limit of the assay. But it did not alter susceptibility to
TTO or terpinen-4-ol. It may be that the MexCD-OprJ pump effluxes 1,8-cineole and α-
terpineol more efficiently than terpinen-4-ol or other TTO components since tolerance to
the latter agents did not change. This is interesting as structure ultimately determines
substrate specificity and these components have a similar structure. α-terpineol and
terpinen-4-ol also have very similar aqueous solubilities and octanol water partition
coefficients (Carson et al., 2006) and the same molecular weight.
It has been reported that alterations in the expression of one Mex system can affect the
levels of expression of other pumps, with MexCD-OprJ expression increased with
decreased expression of MexAB-OprM in mutants including K1110, K1119 and K1121 (Li
et al., 2000). This increase appeared to occur at the level of gene expression, although the
mechanism is not yet understood. Hypersusceptibility to multiple antibiotics including
carbenicillin, chloramphenicol and tetracycline, substrates of MexCD-OprJ, was also
observed (Li et al., 2000). Therefore, in the present study it is likely that MexCD-OprJ is
being expressed in the MexAB-OprM- mutants including K1121 and PAO200. The increase
in susceptibility to 1,8-cineole and α-terpineol in the MexAB-OprM- MexCD-OprJ
-
mutants K1115 and PAO238 indicates that 1,8-cineole and α-terpineol are probably
substrates of MexCD-OprJ.
It seems likely that interplay between the MexAB-OprM and MexCD-OprJ pumps may
contribute to tolerance to some components of TTO, including 1,8-cineole and α-terpineol.
On the other hand, results apparently conflicting with this theory were observed as the
MexAB-OprM- MexCD-OprJ
++ (nfxB) isolate K1131 was 4-fold more susceptible to α-
terpineol in MIC assays than the MexAB-OprM- isolate K1121 that it was derived from.
nfxB mutants of P. aeruginosa show resistance to fluoroquinolones and 4th
generation
cephems, but they are hypersusceptible to β-lactams including ordinary cephems, due to the
reduced expression of MexAB-OprM (Gotoh et al., 1998; Masuda et al., 2001). As we
have shown α-terpineol is a substrate of MexAB-OprM, a reduction in the expression of
MexAB-OprM in an nfxB isolate may increase susceptibility to α-terpineol. However,

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
62
K1131 is a ∆mexAB-oprM mutant, so hyperexpression of MexCD-OprJ would not affect
MexAB-OprM expression. In this study K1131 was more susceptible to TTO and terpinen-
4-ol in time-kill assays than K1121, though there was little difference in MIC values for
these mutants. Additionally, the susceptibility of K1131 and K1115 (MexAB-OprM-
MexCD-OprJ-) to TTO, terpinen-4-ol, 1,8-cineole and α-terpineol by MIC was identical.
The only mutant used in this study with a MexAB-OprM+
MexCD-OprJ++
efflux
phenotype, K1536, had an MIC for α-terpineol of >8%, as did the parent PAO1. Any
difference in susceptibility between these two strains was beyond the limit of the assay. In
time-kill curves of PAO1 and K1536 (nfxB) treated with TTO and terpinen-4-ol, K1536
was slightly more susceptible to both agents (Figures 3.6 and 3.7). This may be caused by a
reduction in MexAB-OprM expression due to MexCD-OprJ hyperexpression, as terpinen-
4-ol and TTO are substrates of this pump.
Further work is needed to confirm whether α-terpineol and 1,8-cineole are substrates of
MexCD-OprJ, including susceptibility assays using a larger library of MexCD-OprJ
mutants and gene expression analysis.
Complementation of a MexCD-OprJ- mutant with pRSP15 (pRK415::mexCD-oprJ) was
not done as it was realised that without confirming that expression of MexCD-OprJ was
induced by TTO or the components used in this study, this would be a pointless exercise.
Recently a variety of unrelated membrane damaging agents, many of which were not
MexCD-OprJ substrates, were demonstrated to induce expression of MexCD-OprJ in
P. aeruginosa (Campigotto, 2007). These agents included EDTA, SDS, ethanol and the
organic solvents n-hexane and p-xylene. It was suggested that MexCD-OprJ plays a role in
alleviating cell envelope stress in P. aeruginosa (Campigotto, 2007). Given the
mechanisms of action of the components in TTO includes membrane damage, it is possible
that MexCD-OprJ is induced by the presence of TTO as a stress response and not
necessarily in an attempt to export the oil.

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
63
3.3.3 MexXY
The mexXY operon is induced by exposure to antibiotics that are substrates for this pump,
but only those antibiotics that target the ribosome, such as chloramphenicol, tetracycline,
macrolides and aminoglycosides (Jeannot et al., 2005). Thus not all substrates of MexXY
are also inducers of the pump. The mutation of the repressor gene mexZ also results in
expression of MexXY in P. aeruginosa (Aires et al., 1999).
The MexXY pump usually requires the OprM component to export substrates and so the
loss of the MexAB-OprM pump may also cause a loss of function for MexXY (Masuda et
al., 2000b). There was no change in susceptibility of strain K1525 (lacking the MexXY
components) compared to the wild type strain. K1523, deficient in MexB and the two
MexAB-OprM- strains PAO200 and K1119, had almost identical susceptibilities,
indicating that this change in susceptibility was probably related solely to the loss of the
MexAB-OprM pump and not to a loss of function of MexXY due to loss of the OprM
component. It appears that TTO, terpinen-4-ol, 1,8-cineole, α-terpineol, ρ-cymene and γ-
terpinene are not substrates of MexXY, but experiments determining the expression of
MexXY in the presence of these compounds or an appropriate inducer compound and
subsequent susceptibility assays, are needed to confirm this.
3.3.4 MexEF-OprN
It has been noted that hyper-expression of MexAB-OprM is correlated with a decline in
MexEF-OprN
expression, while loss of MexAB-OprM is associated with increased
expression of MexEF-OprN, indicating an inverse correlation between the expression of
these two efflux pumps (Li et al., 2000). There was no difference in susceptibility to TTO,
terpinen-4-ol, 1,8-cineole or α-terpineol between K1115 (MexAB-OprM- MexCD-OprJ
-)
and K1117 (MexAB-OprM- MexCD-OprJ
- MexEF-OprN
++) in this study. However, the
MIC results with K1240 (>8% compared with 4% in K1241) indicate a role for MexEF-
OprN in tolerance to terpinen-4-ol in conjunction with MexAB-OprM expression. A low
level of MexAB-OprM expression has been reported in many nfxC mutants of P.

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
64
aeruginosa, indicating that the antibiotic resistance profile of these mutants may be a
consequence of MexEF-OprN over-expression coupled with low levels of MexAB-OprM
expression (Maseda et al., 2000). It is possible that the simultaneous expression of MexEF-
OprN and MexAB-OprM resulted in a higher MIC of terpinen-4-ol compared with that
seen for either efflux system alone.
Reported substrates of MexEF-OprN include fluoroquinolones, imipenem and
chloramphenicol (Maseda et al., 2000) but there are no reports of essential oils or their
components as substrates of MexEF-OprN in P. aeruginosa. Further work using a larger
library of nfxC mutants and analysis of the expression of MexEF-OprN in the presence of
terpinen-4-ol and subsequent susceptibility assays, is required to determine whether
terpinen-4-ol is a substrate of MexEF-OprN.
3.3.5 MexJK
An increase in expression of both MexAB-OprM and MexJK was seen in P. aeruginosa
exposed to pentachlorophenol, a membrane-active organochloride (Muller et al., 2007).
There was no difference in susceptibility to TTO or components detected between MexJK-
overproducing strain PAO238-1 (defective in the mexL repressor) and PAO314; both
strains are MexAB-OprM- and MexCD-OprJ
-. MexJK usually requires OprM to export
antibiotics and although once thought to efflux triclosan by functioning as a two component
efflux pump (Chuanchuen et al., 2002) it requires OpmH to export triclosan (Chuanchuen
et al., 2005). Thus the outer membrane protein used in conjunction with MexJK is substrate
dependent. MexJK may require OprM in order to export TTO or its components; on the
other hand, TTO and components may not be substrates of this pump. It would be useful to
compare the susceptibilities of a MexAB-OprM+ MexJK
+ mutant with a MexAB-OprM
-
MexJK+ mutant to TTO and components. Experiments determining the expression of
MexJK in the presence of TTO, terpinen-4-ol, 1,8-cineole, α-terpineol, ρ-cymene and γ-
terpinene, or an appropriate inducer compound and subsequent susceptibility assays, will
provide data on whether these compounds are substrates of this efflux system.

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
65
3.3.6 Other efflux pump systems
During the course of this project, a number of other Mex efflux systems in P. aeruginosa
were reported or characterized by other groups, including MexGHI-OpmD, MexPQ-OpmE,
MexMN-OprM (Aendekerk et al., 2005; Mima et al., 2005). It was not possible to expand
on the library of efflux pump mutants used in this project, due to time constraints and the
considerable cost involved in the international importing of isolates.
The efflux pumps investigated in this project were all members of the RND family. It is
possible that other families of efflux pumps may be involved in tolerance to TTO. PmpM, a
H+-coupled efflux pump of P. aeruginosa from the MATE (multidrug and toxic compound
extrusion) efflux family, has a range of substrates including benzalkonium chloride,
fluoroquinolones and ethidium bromide (He et al., 2004). EmrE, an SMR (small multidrug
resistance) family protein, contributes to intrinsic resistance to ethidium bromide,
acriflavine and aminoglycosides in P. aeruginosa (Li et al., 2003).
3.3.7 Efflux pump inhibitors
The broad spectrum efflux pump inhibitor PAβN was used to investigate the efflux systems
of P. aeruginosa that contribute towards tolerance to TTO and some of its components. To
ensure that the concentration of PAβN used was effective, a control MIC for PAO1 and
ML5087 using tetracycline and chloramphenicol, with and without 20 mg/L PAβN, was
done. For both isolates, the MIC for both antibiotics decreased, changing the classification
of both from resistant to sensitive according to CLSI guidelines (CLSI, 2007) and
confirming that efflux was inhibited.
The decreases in MIC of TTO, terpinen-4-ol, 1,8-cineole, α-terpineol and ρ-cymene for
PAO1 in the presence of PAβN supported the hypothesis that efflux pumps contribute a
large component of the tolerance to TTO and some of its components. The decreases in
MIC observed when PAβN was present in 1,8-cineole, α-terpineol and ρ-cymene MIC

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
66
assays for K1119, PAO200 and K1523, indicate that MexAB-OprM is probably not the
only efflux system contributing to the tolerance of these compounds in P. aeruginosa.
The smallest effect observed following PAβN treatment was in ∆mexAB-oprM mutants
treated with TTO, therefore the majority of resistance to TTO in P. aeruginosa is probably
due to the MexAB-OprM system.
Susceptibility to ρ-cymene was increased for all strains tested in the presence of PAβN.
The increase in susceptibility to ρ-cymene in ∆MexAB-OprM strains when PAβN is
present indicates that ρ-cymene is a substrate of more pumps than just MexAB-OprM or
pumps requiring OprM. It also indicates that ρ-cymene is able to induce expression of other
efflux systems in P. aeruginosa. This potentially applies to 1,8-cineole, α-terpineol,
terpinen-4-ol as well since in ∆MexAB-OprM strains treated with PAβN, their MICs
decreased by a factor of 4 or 8. Therefore these components may be substrates and inducers
of other pumps in P. aeruginosa.
The above interpretation of results depends on the specificity of action of PAβN as an
efflux pump inhibitor. In fact, the specific mechanism of action of PAβN is not established.
It will be important to determine whether PAβN has any permeabilising effects on the cell
membrane, as this would not only potentially impede the function of efflux systems but
also allow entry of hydrophobic compounds, such as TTO and its components, into the cell.
Recent work has suggested that the diterpene totarol is an efflux pump inhibitor in
S. aureus (Smith et al., 2007). It has been suggested that the mechanism of action of totarol
involves the inhibition of energy-coupled respiration (Haraguchi et al., 1996), while
another group‟s work indicated that totarol was able to compromise the integrity of cell
membranes (Micol et al., 2001). If totarol is indeed an inhibitor of efflux pumps, this is not
a particularly specific efflux pump inhibitor, as a lack of energy or structural disruption of
membranes will impede pump function.

Chapter Three – The Role of Efflux Systems in Tolerance of P. aeruginosa to Tea Tree Oil and Components
67
3.3.8 RT-PCR and real time RT-PCR
Unfortunately neither RT-PCR nor real time RT-PCR was able to be optimised during this
project, with the equipment and funding available. The determination of expression levels
of the genes in the various Mex efflux systems examined in this study is the missing piece
of the puzzle. Complementation studies were able to confirm a role for MexAB-OprM in
terpinen-4-ol and TTO tolerance, but this was not possible for the non-constitutively-
expressed efflux systems as we could not confirm their expression in the first place.
Determining expression levels of efflux systems would provide the final proof of the role of
these systems in tolerance to TTO and components in P. aeruginosa.
3.3.9 Summary
The hypothesis of this chapter was that one or more of the Mex efflux systems of
P. aeruginosa contribute to the tolerance of this organism to TTO and components. This
hypothesis has been supported by data demonstrating that components of TTO, including
terpinen-4-ol and ρ-cymene, are substrates of Mex efflux systems in P. aeruginosa, while
α-terpineol and 1,8-cineole may be substrates. The possibility remains that other membrane
pump systems not constitutively expressed, or not examined here, may contribute to the
efflux of terpenes from P. aeruginosa. Nevertheless, this work further extends the already
broad range of known substrates for MexAB-OprM and potentially other Mex systems, to
cyclic monoterpenes.

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
68
4.0 CHAPTER FOUR - THE ROLE OF LIPOPOLYSACCHARIDE IN RESISTANCE TO TTO IN P. AERUGINOSA
4.1 Introduction
Lipopolysaccharide (LPS) is one of the major virulence factors of P. aeruginosa. The LPS
of P. aeruginosa forms an integral part of the outer membrane and its tripartite structure
comprises the outermost domain of the O-specific heteropolysaccharide, which is
connected to the membrane-embedded lipid A by the core oligosaccharide (Dasgupta et al.,
1994). The oligosaccharide core provides a hydrophilic barrier to antibiotics while the
lipid-A forms a hydrophobic barrier (Soares et al., 2008). While small hydrophilic solutes
are able to cross the cell envelope via the porin uptake pathway, the divalent cations
(mainly Mg2+
) crosslinking the multiple phosphate residues of adjacent LPS molecules
work to block the entry of hydrophobic agents into the cell. The low fluidity of the LPS
outer leaflet prevents hydrophobic agents from diffusing through the lipid domains of the
outer membrane (Nikaido, 1986).
Mutations in the bacterial genome affecting the core region can compromise the barrier
function of the LPS and can result in an increased susceptibility to hydrophobic antibiotics
and antimicrobial peptides, as well as a decrease in virulence (Ramjeet et al., 2005; Walsh
et al., 2000). Chemically-induced modification of the outer membrane can also increase
susceptibility to hydrophobic antimicrobial agents. Previous work has shown that treatment
with the divalent cation chelator EDTA or the outer membrane permeabilizer PMBN
increased the susceptibility of P. aeruginosa to TTO and components (Longbottom et al.,
2004). The hypothesis for this chapter of work was that the LPS of P. aeruginosa plays a
role in protection from the antimicrobial effects of TTO and components and that rough
mutants, especially those with an incomplete core, would have increased susceptibility to
these agents.
In this chapter a modest collection of seven rough mutants and parental strains was
obtained and examined for susceptibility to TTO and components. The rough mutants were
derived from two different parental strains, P. aeruginosa PAO1 (PAO1) and P. aeruginosa

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
69
PAK (PAK). An rmlC (PAO1rmlC, PAKrmlC) and wbpL (PAKwbpL, PAO1wbpL) mutant
from each parental strain was studied, as well an AlgC mutant (AK1012) derived from
P. aeruginosa PAO1. The structure of the LPS core in each strain and the positions of the
mutations are shown in Figure 4.1. The rmlC mutants are deficient in L-rhamnose
biosynthesis, producing a complete inner core and a truncated outer core missing the
terminal D-glucose and L-rhamnose residues (Rahim et al., 2000). The wbpL mutants are
deficient in glycosyltransferase, which initiates the synthesis of both A-band and B-band O
antigen. This results in mutants that are A- and B-band deficient but have a full core
(Rocchetta et al., 1998). AlgC deficient mutants, such as AK1012, are deficient in
phosphomannomutase/phosphoglucomutase, essential for synthesis of precursors for the D-
glucose and L-alanine residues of the core, the D-mannose residues required for alginate
production and the D-rhamnose residues of A-band O antigen (Goldberg et al., 1993;
Miller and Lam, 2008). AlgC mutants cannot attach glucose residues to galactosamine in
the LPS core and so are A- and B-band deficient and have a truncated outer core. The order
of roughness in the three types of mutants used in this study, from least rough to most
rough, was wbpL, rmlC and AK1012.

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
Figure 4.1 Structural lipopolysaccharide core model of rough mutants of
P. aeruginosa serotype O5 (A) (Modified from (Sadovskaya et al., 1998; Walsh et
al., 2000)) and P. aeruginosa serotype O6 (B) (Modified from (Masoud et al.,
1995)).
Legend
Ala, alanine; CONH2, carbamoyl group; GalN, galactosamine; Glc, glucose; Hep,
heptose; KDO, 2-keto-3-deoxyoctonic acid; P, phosphate; Rha, rhamnose.
A
B

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
70
4.2 Results
4.2.1 Susceptibility of rough mutants and parental strains
4.2.1.1 TTO and component MICs
The MICs of TTO, terpinen-4-ol, α-terpineol, γ-terpinene, 1,8-cineole and ρ-cymene for the
rough mutants and parent strains are shown in Table 4.1. The rmlC and AK1012 mutants
were generally more susceptible to TTO, terpinen-4-ol, α-terpineol and 1,8-cineole
compared with parental strains. For PAO1rmlC, the TTO and terpinen-4-ol MICs were 1%
and 0.5%, respectively, compared to 4% and 2% for PAO1; similarly for PAKrmlC, the
TTO MIC was 1-2% compared to 4% for PAK, while the terpinen-4-ol MIC was 0.25%
compared to 2-4% for PAK. For AK1012, the MIC of TTO was 2% and the MIC of
terpinen-4-ol was 0.5%. The MIC of α-terpineol was >8% for both PAO1 and PAK, while
for PAO1rmlC, PAKrmlC and AK1012, the MICs were 0.25%, 0.5% and 2%, respectively.
Similarly, the MIC of 1,8-cineole was >8% for both PAO1 and PAK, while for PAO1rmlC,
PAKrmlC and AK1012 the MICs were 2%, 1% and >8%, respectively.
The PAO1wbpL mutant had the same susceptibility as the PAO1 parental strain to all
agents tested. PAKwbpL was more susceptible to α-terpineol and terpinen-4-ol than PAK.
The MIC of terpinen-4-ol was 1%, compared to 2-4% in PAK; the MIC of α-terpineol was
2%, compared to >8% in PAK. Susceptibility of PAKwbpL to the other agents tested by
MIC assay (TTO, 1,8-cineole, ρ-cymene and γ-terpinene) was the same as PAK.
The susceptibility of all isolates to γ-terpinene and ρ-cymene was outside the limits of the
assay due to immiscibility of these components at >8% (v/v).

Table 4.1 MICs (%, v/v) of TTO and components against rough mutants of P. aeruginosa and parental strains
Isolate TTO Terpinen-4-ol α-Terpineol 1,8-Cineole ρ-Cymene γ-Terpinene
PAO1 4 2 >8 >8 >8 >8
PAO1wbpL 4 2 >8 >8 >8 >8
PAO1rmlC 1 0.5 0.25 2 >8 8
AK1012 2 0.5 2 >8 >8 >8
PAK 4 2-4 >8 >8 >8 >8
PAKwbpL 4 1 2 >8 >8 >8
PAKrmlC 1-2 0.25 0.5 1 >8 >8
Chap
ter Fo
ur - T
he R
ole o
f Lip
opo
lysacch
arid
e in R
esistance to
TT
O in
P. a
erugin
osa

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
71
4.2.1.2 Time-kill assays with TTO and components
4.2.1.2.1 rmlC mutants
In time-kill assays with TTO, a 2 log decrease in viability after 120 min was seen with
PAO1 treated with 4% TTO, the MIC for this strain (Figure 4.2). Treatment of PAO1rmlC
with 0.06% TTO, equal to one-sixteenth of the MIC for this rough mutant, caused a
decrease in viability below the limit of detection after 45 min, while treatment with 0.03%
TTO resulted in a 1.5 log decrease in viability after 120 min (Figure 4.2). Treatment of
PAO1rmlC with 0.125% terpinen-4-ol (equal to one-quarter of the MIC) caused a 2.5 log
decrease in viability after 120 min; 0.06% terpinen-4-ol treatment allowed slight growth
over 120 min (Figure 4.3). For the parent strain PAO1, 0.125% terpinen-4-ol (one-sixteenth
of the MIC for this strain) allowed slight growth over 120 min while treatment with 1%
terpinen-4-ol (half the MIC) resulted in a decrease in viability beyond the limit of detection
of the assay after 60 min. Treatment of PAO1rmlC with 0.125% 1,8-cineole (one-sixteenth
of the MIC for this mutant) caused just over a 90% decrease in viability after 120 min
(Figure 4.3), while for the parent strain PAO1, treatment with 1% 1,8-cineole (a maximum
of one-eighth the MIC for this strain) resulted in slight growth after 120 min (Figure 4.3).
Treatment of PAK with 4% TTO, equal to the MIC for that strain, killed 99.9% of cells
after 30 min (Figure 4.4). For the remainder of the 120 min assay, viability remained at a
level just above the limit of detection (3 × 103 cfu/ml). Treatment of PAKrmlC with 4%
TTO, 2-4 times the MIC for that mutant, caused a 2 log decrease in viability after 15 min,
following which the viability dropped below the limit of detection (Figure 4.4). Similar
levels of cell death were observed following treatment of PAK and PAKrmlC with 2%
TTO, with a 2.5 and 3 log decrease in viability, respectively. Slight cell death was observed
in PAKrmlC treated with 0.25% TTO after 120 min, while in PAK growth was observed
with the same treatment. Treatment with 1% terpinen-4-ol, 4 times the MIC for PAKrmlC,
caused a 3 log decrease in viability after 120 min (Figure 4.5). The same concentration
resulted in a 2 log decrease in viability in PAK after 120 min (Figure 4.5). Growth
comparable to the MHB-T control was seen in both PAK and PAKrmlC following
treatment with 0.125% terpinen-4-ol for 120 min. Similarly, treatment with 4% 1,8-cineole

1.00E+02
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
PAO1rmlC MHB-T
PAO1rmlC 0.03%
TTO
PAO1rmlC 0.06%
TTO
PAO1 MHB-T
PAO1 0.25% TTO
PAO1 4% TTO
Figure 4.2 Time-kills curves for P. aeruginosa PAO1 and P. aeruginosa PAO1rmlC incubated with TTO or MHB-T for up to 120 min at
35°C and shaking. Error bars indicate ± standard deviation; limit of detection was 3 × 103 cfu/ml.
Chap
ter Fo
ur - T
he R
ole o
f Lip
opo
lysacch
arid
e in R
esistance to
TT
O in
P. a
erugin
osa

1.00E+02
1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
PAO1rmlC MHB-T
PAO1rmlC 0.06%
Terpinen-4-ol
PAO1rmlC 0.125%
Terpinen-4-ol
PAO1rmlC 0.125%
1,8-cineole
PAO1 MHB-T
PAO1 0.125%
Terpinen-4-ol
PAO1 1%
Terpinen-4-ol
PAO1 1% 1,8-
cineole
Figure 4.3 Time-kill curves for P. aeruginosa PAO1 and P. aeruginosa PAO1rmlC incubated with terpinen-4-ol, 1,8-cineole or MHB-T
for up to 120 min at 35°C and shaking. Error bars indicate ± standard deviation; limit of detection was 3 × 103 cfu/ml.
Chap
ter Fo
ur - T
he R
ole o
f Lip
opo
lysacch
arid
e in R
esistance to
TT
O in
P. a
erugin
osa

1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
PAKrmlC MHB-T
PAKrmlC 0.25%
TTO
PAKrmlC 2% TTO
PAKrmlC 4% TTO
PAK MHB-T
PAK 0.25% TTO
PAK 2% TTO
PAK 4% TTO
Figure 4.4 Time-kill curves for P. aeruginosa PAKrmlC and P. aeruginosa PAK incubated with TTO or MHB-T for up to 120 min at
35°C and shaking. Error bars indicate ± standard deviation; limit of detection was 3 × 103 cfu/ml.
Chap
ter Fo
ur - T
he R
ole o
f Lip
opo
lysacch
arid
e in R
esistance to
TT
O in
P. a
erugin
osa

1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
1.00E+09
0 30 60 90 120
Time (min)
cfu
/ml
PAKrmlC MHB-T
PAKrmlC 0.125%
Terpinen-4-ol
PAKrmlC 1%
Terpinen-4-ol
PAKrmlC 4% 1,8-
cineole
PAK MHB-T
PAK 0.125%
Terpinen-4-ol
PAK 1% Terpinen-
4-ol
PAK 4% 1,8-
cineole
Figure 4.5 Time-kill curves for P. aeruginosa PAK and P. aeruginosa PAKrmlC incubated with terpinen-4-ol, 1,8-cineole or MHB-T for
up to 120 min at 35°C and shaking. Error bars indicate ± standard deviation; limit of detection was 3 × 103 cfu/ml.
Chap
ter Fo
ur - T
he R
ole o
f Lip
opo
lysacch
arid
e in R
esistance to
TT
O in
P. a
erugin
osa

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
72
(4 times the MIC for PAKrmlC and less than half the MIC for PAK, >8%) resulted in
growth comparable to the MHB-T control for PAK and PAKrmlC (Figure 4.5).
4.2.1.2.2 AK1012 mutant
Treatment of AK1012 with 4% TTO, twice the MIC for this mutant, resulted in a 2 log
decrease in viability after 120 min (Figure 4.6). This was comparable to the death seen with
the parent PAO1 when treated with the same concentration of TTO (Figure 4.6). Treatment
with one quarter of the MIC of TTO, 0.5%, caused approximately half a log decrease in the
viability of AK1012 after 120 min; again this was comparable to PAO1. Up to half a log
decrease in viability was seen after 30 min during treatment of AK1012 with 0.125%
terpinen-4-ol (one quarter of the MIC); viability was increased after 120 min back to the
initial levels (Figure 4.7). A 3 log decrease in cell viability of AK1012 occurred after 120
min treatment with the MIC of terpinen-4-ol (0.5%). A similar decrease in viability
occurred in PAO1 cells treated with 1% terpinen-4-ol (half the MIC).
4.2.1.3 Antibiotics
PAO1 and the PAO1-derived rough mutants had similar susceptibility to gentamicin (Table
4.2). The MIC of gentamicin against PAO1rmlC was <0.5 mg/L and for AK1012 1 mg/L,
while for PAO1 the MIC was 1 mg/L. The MIC of gentamicin against PAO1wbpL was
256 mg/L. It was difficult to reproduce a constant value for the MIC of gentamicin against
PAK; therefore a range of 1-128 mg/L was determined. The MIC of gentamicin against
both PAKrmlC and PAKwbpL was 512 mg/L.
Susceptibility to nalidixic acid was decreased in AK1012, with the MIC >256 mg/L
compared to 128 mg/L in PAO1 (Table 4.2). PAO1rmlC was more susceptible, with an
MIC of <1 mg/L, while for PAO1wbpL, the MIC of nalidixic acid was 64 mg/L. For PAK
and PAKwbpL, the MIC of nalidixic acid was 128 mg/L and for PAKrmlC 64 mg/L.

1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
AK1012 MHB-T
AK1012 0.5% TTO
AK1012 4% TTO
PAO1 MHB-T
PAO1 0.5% TTO
PAO1 4% TTO
Figure 4.6 Time-kill curves for P. aeruginosa PAO1 and P. aeruginosa AK1012 incubated with TTO or MHB-T for up to 120 min at
35°C and shaking. Error bars indicate ± standard deviation; limit of detection was 3 × 103 cfu/ml.
Chap
ter Fo
ur - T
he R
ole o
f Lip
opo
lysacch
arid
e in R
esistance to
TT
O in
P. a
erugin
osa

1.00E+03
1.00E+04
1.00E+05
1.00E+06
1.00E+07
1.00E+08
0 30 60 90 120
Time (min)
cfu
/ml
AK1012 MHB-T
AK1012 0.125%
Terpinen-4-ol
AK1012 0.5%
Terpinen-4-ol
PAO1 MHB-T
PAO1 0.125%
Terpinen-4-ol
PAO1 1%
Terpinen-4-ol
Figure 4.7 Time-kill curves for P. aeruginosa PAO1 and P. aeruginosa AK1012 incubated with terpinen-4-ol or MHB-T for up to 120
min at 35°C and shaking. Error bars indicate ± standard deviation; limit of detection was 3 × 103 cfu/ml.
Chap
ter Fo
ur - T
he R
ole o
f Lip
opo
lysacch
arid
e in R
esistance to
TT
O in
P. a
erugin
osa

Table 4.2 MICs of antibiotics and SDS (mg/L) for rough mutants of P. aeruginosa and their parental strains
Isolate Ciprofloxacin Gentamicin Novobiocin Nalidixic Acid Polymyxin B Polymyxin E Vancomycin SDS
PAO1 0.125 1 >512 128 2 2 >1024 >15
PAO1wbpL 0.06 256 512 64 2 2 1024 >15
PAO1rmlC <0.003 <0.5 128 <1 4 8 128 1.9
AK1012 0.5 1 512 >256 2 2 >1024 >15
PAK 0.125 1-128 >512 128 2 2 >1024 >15
PAKwbpL 0.125 512 256 128 2 2 >1024 >15
PAKrmlC 0.125 512 64 64 2 2 512 <0.23
Chap
ter Fo
ur - T
he R
ole o
f Lip
opo
lysacch
arid
e in R
esistance to
TT
O in
P. a
erugin
osa

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
73
The MIC of vancomycin was >1024 mg/L for PAO1, AK1012, PAK and PAKwbpL.
Susceptibility to vancomycin was increased in PAO1rmlC, PAO1wbpL and PAKrmlC with
MICs of 128 mg/L, 1024 mg/L and 512 mg/L, respectively.
The MICs of polymyxin B and polymyxin E were 2 mg/L for all isolates, with the
exception of PAO1rmlC, for which the MICs were 4 mg/L and 8 mg/L, respectively.
PAO1rmlC and PAO1wbpL were more susceptible to ciprofloxacin than PAO1, with MICs
of <0.003 mg/L and 0.06 mg/L, respectively, compared to 0.125 mg/L for PAO1 (Table
4.2). The MIC of ciprofloxacin for AK1012 was 0.5 mg/L. PAK, PAKrmlC and PAKwbpL
had the same ciprofloxacin MIC of 0.125 mg/L.
The MIC of novobiocin for both PAO1 and PAK was >512 mg/L, for PAKrmlC 64 mg/L,
for PAO1rmlC 128 mg/L, for PAKwbpL 256 mg/L and for PAO1wbpL and AK1012
512 mg/L.
The MIC of SDS was >15 g/L for all isolates with the exception of PAO1rmlC (1.9 g/L)
and PAKrmlC (<0.23 g/L).
4.2.2 SDS-PAGE
SDS-PAGE analysis was difficult to optimize due to problems with equipment and
reagents. Figure 4.8 shows the optimized, silver stained, SDS-PAGE gel. Although the
presence of the full A- and B-band O-antigen was difficult to confirm due to the resolution
of gel, the gel photo shows that the parent strains PAO1 and PAK, as well as the wbpL
mutants, have the full core, while the rmlC mutants are missing part of the core. AK1012 is
represented twice, both as a mutant and a revertant. The mutant is missing the O-antigen
and has a partial core, while the revertant appears to have a similar LPS pattern to the
parent, PAO1.

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
Figure 4.8 SDS-PAGE analysis of LPS from rough mutants and parental strains of
P. aeruginosa. Using a 4% separating gel and 15% separating gel, 15 µl of LPS sample
was run at 200 V and silver stained. Lanes: 1, PAO1; 2, PAO1rmlC; 3, PAO1wbpL; 4,
PAK; 5, PAKrmlC; 6, PAKwbpL; 7, AK1012 (revertant); 8, AK1012; M, All Blue
Precision Plus Protein™ Standard (Bio-Rad).
1 2 3 4 5 6 7 8 M
250kD 150kD
100kD 75kD
50kD
37kD
25kD
Core

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
74
4.3 Discussion
4.3.1 Susceptibility to TTO and components
In MIC and time-kill assays, both rmlC mutants, with no O-antigen and a truncated core,
were more susceptible to TTO compared to their parent strains, while the wbpL mutants,
with no O-antigen and a full core, did not show a change in susceptibility. In time-kill
assays, the difference in susceptibility between parent and rmlC mutant was far greater for
PAO1 than PAK. This is surprising as PAO1rmlC and PAKrmlC had similar MICs for
TTO; the reason for this difference is unclear but may stem from the fact that the error bars
for the PAO1rmlC time-kills were much larger than for the PAKrmlC time-kills.
PAO1rmlC and PAKrmlC were at least 4-fold and 8-fold more susceptible, respectively, to
1,8-cineole than their parent strains, while there was no change in susceptibility of the
wbpL mutants. PAO1wbpL also had the same MIC values as PAO1 for terpinen-4-ol and α-
terpineol, while PAKwbpL had decreased MICs for both agents compared to PAK, though
the decrease in MIC was not as great as that seen for PAKrmlC.
The loss of O-antigen alone as seen in wbpL mutants did not affect susceptibility to TTO
and terpinen-4-ol in PAO1wbpL while PAKwbpL was more susceptible to terpinen-4-ol
and also α-terpineol, but not TTO. PAO1 (serotype O5) and PAK (serotype O6) differ in
the outer core region as PAO1 has an additional D-glucose residue (Altman, 1993) (Figure
4.1). Perhaps the additional D-glucose residue provides more protection to PAO1wbpL
from the antimicrobial actions of terpinen-4-ol and α-terpineol. Previously a wbpL mutant
of P. putida was as tolerant as the corresponding parent strain to aromatic hydrocarbons
(Junker et al., 2001), suggesting that the O-antigen is not important in solvent tolerance in
P. putida. It would be interesting to compare the TTO and component susceptibilities of
P. putida wbpL mutants with the P. aeruginosa wbpL mutants in this study.
There was no difference detected in susceptibility to ρ-cymene and γ-terpinene in the rough
mutants examined in this study, with the exception of PAO1rmlC, which had a slightly

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
75
decreased MIC of γ-terpinene compared with PAO1. It is possible that any change in
susceptibility was outside the limits of the MIC assay, e.g. above 8% (v/v). Permeabilizing
the outer membrane of P. aeruginosa with PMBN increased susceptibility to both ρ-
cymene and γ-terpinene (Longbottom et al., 2004; Mann et al., 2000). The mechanism of
action of PMBN has recently been re-investigated in E. coli and comprises outer membrane
permeabilization characterized by the release of periplasmic proteins and LPS (Sahalan and
Dixon, 2008). The full inner core and partial outer core of the rmlC mutants appear to
provide protection from the effects of ρ-cymene and γ-terpinene, though it seems loss of
more of the LPS molecule may increase susceptibility to these components.
Dasgupta et al. (1994) described AK1012 as a leaky mutant, that was obtained by a phage
D3-induced point mutation and was sometimes able to express B-band LPS (Dasgupta et
al., 1994). Personal communication with Professor Joseph Lam indicated this strain had a
high reversion frequency and researchers “often have to recover this mutant from various
stock cultures before finding one that is more stable” (Lam, 2004). The reversion could not
be simply detected by colony morphology, as rough colony morphology is not always
related to defective LPS in P. aeruginosa, unlike the situation for members of the
Enterobacteriaceae (Dasgupta et al., 1994). Hence the reversion of AK1012 was not noted
until SDS-PAGE analysis had been done, following susceptibility testing. It was realized
that the AK1012 mutant in use had reverted and was probably producing B-band O-antigen.
This correlated with results in both the SDS-PAGE gels and the MIC assays. A replacement
AK1012 mutant was acquired and re-tested and the susceptibility results of the replacement
mutant are those presented and discussed within this Thesis.
An increase in susceptibility to terpinen-4-ol and α-terpineol was noted for AK1012, along
with a slight increase in susceptibility to TTO. Yet AK1012, while rougher than
PAO1rmlC, was not more susceptible to TTO and components than PAO1rmlC. This
finding did not fit with the hypothesis for this chapter; it was expected that the rougher
mutants would be the most susceptible and so perhaps reversion or leakiness was a problem
during these assays.

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
76
During the course of this project, Yokota et al. (2007) demonstrated that deep rough
mutants of P. aeruginosa, with no rhamnose or glucose residues, were less hydrophobic
than parental strains and more susceptible to the hydrophobic antibiotics gentamicin and
polymyxin B. Less defective mutants, possessing some neutral sugar residues in their outer
core, were more hydrophobic than parent strains and tended to be less susceptible to
gentamicin and polymyxin B than parent strains (Yokota and Fujii, 2007). The core
structure of one of the deep-rough mutants used by Yokota et al. (2007) appears the same
as that of AK1012 (Rowe and Meadow, 1983), suggesting that the cell surface
hydrophobicity of AK1012 may also be low. Unfortunately a method for determining the
cell surface hydrophobicity of P. aeruginosa was not able to be optimised during this
project (see Chapter 5) and therefore the cell surface hydrophobicity of the rough mutants
used was not determined. If the cell surface of AK1012 is less hydrophobic than PAO1, it
is reasonable to deduce that AK1012 would probably be more susceptible to hydrophobic
compounds such as TTO and terpinen-4-ol. Plesiat et al. (1997) demonstrated that the outer
membrane of AK1012 was more than five times more permeable to the strongly
hydrophobic steroid testosterone than the parent strain (Plesiat et al., 1997). As shown in
Table 4.1, AK1012 was more susceptible to both TTO, terpinen-4-ol and α-terpineol than
PAO1, supporting the theory that the cell surface of AK1012 is less hydrophobic than that
of PAO1.
Overall, these results suggest that a full LPS core is required to protect P. aeruginosa from
the antimicrobial actions of TTO, terpinen-4-ol, α-terpineol and 1,8-cineole.
4.3.2 Susceptibility to antibiotics and SDS
Rough mutants of P. aeruginosa are reported as more susceptible to hydrophobic
antibiotics (Walsh et al., 2000) and, therefore, potentially, hydrophobic TTO. Rough
mutants of P. aeruginosa are also more susceptible to dyes and detergents (Kropinski et al.,
1978). Changes to the LPS have long been implicated in resistance to gentamicin (Shearer
and Legakis, 1985), including an association with rough mutants of P. aeruginosa lacking
O-antigen (Galbraith et al., 1984). The wbpL mutants in this study (lacking O-antigen)

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
77
were both resistant to gentamicin, supporting the findings of Galbraith et al. (1984). Yokota
et al. (2007) recently reported that rough mutants of P. aeruginosa were less susceptible to
both gentamicin and polymyxin B than parent strains, with deep rough mutants more
susceptible than parent strains. This was not reflected in the present study. which indicated
the deep rough mutant, AK1012, was equally susceptible to gentamicin as the parent
PAO1. PAO1rmlC, a rough (but not deep rough) mutant was more susceptible than the
parent to gentamicin, but less susceptible to polymyxin B. PAKrmlC was less susceptible to
gentamicin than PAK. All rough mutants and parent strains had the same MIC for
polymyxin B (2 mg/L) with the exception of PAO1rmlC for which the MIC was 4 mg/L.
The two rmlC mutants in this study were both significantly more susceptible to novobiocin
and SDS than all other mutants and parent strains. PAKwbpL was at least 2-fold more
susceptible to novobiocin than PAK but had the same MIC for SDS (>15 mg/L). The deep
rough mutant AK1012 had similar MICs for these two agents as the parent strain PAO1.
Deep rough mutants of E. coli and Salmonella typhimurium, with incomplete core LPS, are
more susceptible to SDS and hydrophobic novobiocin (Bennett et al., 1981; Moller et al.,
2003). In this study, the deep rough mutant, AK1012, was expected to follow this trend and
be more susceptible to the hydrophobic agents tested. AK1012 has been reported as more
susceptible to nalidixic acid than the parent strain PAO1 (Plesiat et al., 1997), however, this
was also not confirmed. AK1012 was at least 2-fold more resistant to nalidixic acid than
PAO1. As mentioned previously, the AK1012 used in this project may have reverted at
least once during the project. Given the patterns of susceptibility of AK1012 to
hydrophobic antibiotics as well as hydrophobic TTO, it is possible that reversion occurred
again during the MIC assays, though without running an SDS-PAGE gel in conjunction
with every MIC assay it was not possible to confirm this.
PAK, PAKrmlC and PAKwbpL were equally susceptible to ciprofloxacin with MICs of
0.125 mg/L. An interesting pattern of susceptibility to ciprofloxacin occurred with PAO1
and the rough mutants derived from it. PAO1rmlC was at least 40-fold more susceptible to
ciprofloxacin, while AK1012 was 4-fold more resistant. Acquisition of ciprofloxacin
resistance in P. aeruginosa has been associated with defects in lipopolysaccharide synthesis

Chapter Four - The Role of Lipopolysaccharide in Resistance to TTO in P. aeruginosa
78
(Legakis et al., 1989). This was accompanied by increased nalidixic acid MICs (Legakis et
al., 1989), which also occurred in this study with AK1012, with a change in MIC from
128 mg/L in PAO1 to > 256mg/L. Electrophoretic analysis of the LPS from ciprofloxacin
resistant mutants of P. aeruginosa revealed that most had deficiencies in the O-antigen
and/or core region, compared to parent strains (Legakis et al., 1989; Masecar et al., 1990).
A 54kDa outer membrane protein was also expressed in a ciprofloxacin resistant mutant of
P. aeruginosa that was also resistant to several non-quinolone antibiotics (Legakis et al.,
1989). It is not clear why PAO1rmlC was more susceptible to ciprofloxacin when defective
LPS has been associated with resistance to this antibiotic. Also, rough mutants of PAK did
not differ in susceptibility to ciprofloxacin either.
Both rmlC mutants were more susceptible to vancomycin compared to parent strains, with a
minimum 8-fold decrease in MIC for PAO1rmlC and a minimum 2-fold decrease in MIC
for PAKrmlC. Susceptibility to vancomycin has been induced following treatment with
EDTA to permeabilise the outer membrane (Guha et al., 2002). Vancomycin inhibits the
synthesis of peptidoglycan in the cell wall of Gram positive organisms, but due to the size
of the molecule is not able to penetrate the outer membrane of Gram negative organisms
(Nicas, 1999). The lack of a complete LPS core in the rmlC mutants may facilitate the entry
of vancomycin into the cells.
4.3.3 Summary
The hypothesis of this chapter was that the LPS of P. aeruginosa plays a role in protection
from the antimicrobial effects of TTO and components and that the rougher mutants, with
an incomplete core, would be more susceptible to these agents. The data presented here
show that an incomplete LPS core renders P. aeruginosa more susceptible to TTO and
some of its terpene components, including terpinen-4-ol, α-terpineol and 1,8-cineole.

Chapter Five –Induced resistance to TTO
79
5.0 CHAPTER FIVE – INDUCED RESISTANCE TO TTO
5.1 Introduction
One of the well established problems with present day antibiotic therapy is the potential for
resistance to develop in target bacteria, following continual exposure to an antibiotic during
treatment. This development of resistance has resulted in a move towards the exploration of
non-traditional antimicrobials, including biocides, antimicrobial peptides (Cirioni et al.,
2006) and essential oils (Edris, 2007). The likelihood of resistance developing to these
agents following prolonged exposure must also be investigated. Resistant mutants have
been produced by exposure to the phenolic disinfectant pine oil (Moken et al., 1997) which,
like TTO, is comprised of terpene compounds.
The likelihood of resistance to TTO developing has been investigated for Staphylococcus
aureus, including MRSA, Salmonella spp. and Escherichia coli (Nelson, 2000; McMahon
et al., 2007). Nelson (2000) reported that S. aureus isolates subcultured in stepwise fashion
on TTO-containing agar plates had a minimum 4-fold increase in MIC. Habituation to sub-
inhibitory concentrations of TTO for 72 h reduced the susceptibility of S. aureus to a range
of clinically relevant antibiotics (McMahon et al., 2007). However, when the MICs were
compared to the relevant accepted clinical breakpoints for these antibiotics, the isolates
were mostly still considered susceptible.
Although P. aeruginosa already has a higher level of intrinsic tolerance to the antimicrobial
actions of TTO, the aim of this chapter was to investigate whether this level would increase
on continual exposure to increasing concentrations of TTO or terpinen-4-ol. It was of
interest to evaluate any effect this may have on antibiotic, disinfectant or metal ion
susceptibility, or fatty acid composition, cell surface hydrophobicity or outer membrane
permeability. It was predicted that exposure to terpinen-4-ol would decrease susceptibility
to terpinen-4-ol but not TTO as a whole, while exposure to TTO would potentially decrease
susceptibility to terpinen-4-ol but not TTO.

Chapter Five –Induced resistance to TTO
80
5.2 Results
5.2.1 Serial subculture
Two strains of P. aeruginosa were used in this section of work, P. aeruginosa PAO1
(PAO1) and P. aeruginosa PAO1mutS (mutS). The maximum concentration of TTO or
terpinen-4-ol to which each isolate became resistant, along with the number of passages
taken to develop resistance, is shown in Table 5.1. The PAO1 isolate was subcultured to a
maximum concentration of 6% TTO and 6.5% terpinen-4-ol after 46 and 34 passages,
respectively. The mutS isolate was subcultured to a maximum concentration of 6% TTO
and 9% terpinen-4-ol after 45 and 41 passages, respectively. For ease of discussion, the
nomenclature used for these isolates was as follows:
P. aeruginosa PAO1 6% TTO = PAO1TTO
P. aeruginosa PAO1 6.5% terpinen-4-ol = PAO1T4ol
P. aeruginosa PAO1mutS 6% TTO = mutSTTO
P. aeruginosa PAO1mutS 9% terpinen-4-ol = mutST4ol
Once each strain reached the maximum oil/component concentration at which growth
occurred, it was stored at -80°C and also subcultured for a further 20 passages in the
absence of agent on BA and was stored at -80°C after both 10 and 20 passages. The
nomenclature used for these isolates was as follows:
P. aeruginosa PAO1 6% TTO plus 10 passages = PAO1TTO+10
P. aeruginosa PAO1 6% TTO plus 20 passages = PAO1TTO+20
P. aeruginosa PAO1 6.5% terpinen-4-ol plus 10 passages = PAO1T4ol+10
P. aeruginosa PAO1 6.5% terpinen-4-ol plus 20 passages = PAO1T4ol+20
P. aeruginosa PAO1mutS 6% TTO plus 10 passages = mutSTTO+10
P. aeruginosa PAO1mutS 6% TTO plus 20 passages = mutSTTO+20
P. aeruginosa PAO1mutS 9% terpinen-4-ol plus 10 passages = mutST4ol+10
P. aeruginosa PAO1mutS 9% terpinen-4-ol plus 20 passages = mutST4ol+20

Chapter Five –Induced resistance to TTO
Table 5.1 Concentrations of TTO and terpinen-4-ol reached during serial
subculture of P. aeruginosa
Isolate
TTO Terpinen-4-ol
Initial
MIC
(%)
Concentration
(%)
Passages
Initial
MIC
(%)
Concentration
(%)
Passages
PAO1 2 - 4 6 46
2 6.5 34
PAO1mutS 8 6 45
4 9 41

Chapter Five –Induced resistance to TTO
81
The parental strains were also subcultured without oil or component for 20 passages and are
referred to as PAO1+20
and mutS+20
.
During the course of serial subculture, the colonial morphology of several of the isolates
altered when grown on BA. This change in morphology was observed in PAO1 at around
1.5% TTO and 4% terpinen-4-ol and in mutS at around 1.5% TTO and 3% terpinen-4-ol.
Changes observed included colonies that were smaller in size and some that were irregular
in shape. Over the course of the experiments most of the morphologies reverted to that of
the parent strains.
5.2.2 TTO and terpinen-4-ol MICs
The MIC of TTO and terpinen-4-ol was determined for the serial subculture isolates and
parental strains by broth microdilution. Results are shown in Tables 5.2 and 5.3. The MIC
of TTO for PAO1 was 2% to 4%. For PAO1+20
, the MIC of TTO was 4% and for PAO1TTO
8%. The MIC of TTO for the latter isolate dropped following subculture without TTO, to
4% and 2% after 10 and 20 subcultures, respectively. This trend was not seen in the
PAO1T4ol
series of isolates, where the MIC of TTO was 4% for PAO1T4ol
, PAO1T4ol+10
and
PAO1T4ol+20
. The MIC of terpinen-4-ol for the PAO1TTO
isolate was >8%, while for
PAO1TTO+10
and PAO1TTO+20
it was 8%. For the PAO1T4ol
series of isolates, the terpinen-4-
ol MIC was 2%. The MIC of TTO for mutS, mutS+20
, mutSTTO
, mutSTTO+10
and mutSTTO+20
was 8%; for mutST4ol
mutST4ol+10
and mutST4ol+20
it was 4%. The MIC of terpinen-4-ol for
mutS was 4% while for mutS+20
it was 2% and for all other mutS derived isolates it was
>8%.
5.2.3 PAβN MICs and MBCs
The susceptibility of the serial subculture isolates and parental strains to the efflux inhibitor
PAβN was determined by broth microdilution. Results are shown in Table 5.4. The MIC of
PAβN for PAO1, PAO1+20
, mutS and mutS+20
was 320mg/L. The MIC for PAO1TTO
,

Chapter Five –Induced resistance to TTO
Table 5.2 TTO and terpinen-4-ol susceptibilities for serial subculture isolates
derived from P. aeruginosa PAO1
Isolate TTO MIC (%) Terpinen-4-ol MIC (%)
PAO1 2 –4 2
PAO1+20
4 2
PAO1TTO
8 >8
PAO1TTO+10
4 8
PAO1TTO+20
2 8
PAO1T4ol
4 2
PAO1T4ol+10
4 2
PAO1T4ol+20
4 2
Legend
PAO1 = P. aeruginosa PAO1
PAO1+20
= P. aeruginosa PAO1 subcultured on blood agar only for 20 passages
PAO1TTO
= P. aeruginosa PAO1 6% TTO
PAO1TTO+10
= P. aeruginosa PAO1 6% TTO plus 10 passages
PAO1TTO+20
= P. aeruginosa PAO1 6% TTO plus 20 passages
PAO1T4ol
= P. aeruginosa PAO1 6.5% terpinen-4-ol
PAO1T4ol+10
= P. aeruginosa PAO1 6.5% terpinen-4-ol plus 10 passages
PAO1T4ol+20
= P. aeruginosa PAO1 6.5% terpinen-4-ol plus 10 passages

Chapter Five –Induced resistance to TTO
Table 5.3 TTO and terpinen-4-ol susceptibilities for serial subculture isolates
derived from P. aeruginosa PAO1mutS
Isolate TTO MIC (%) Terpinen-4-ol MIC (%)
mutS 8 4
mutS+20
8 2
mutSTTO
8->8 >8
mutSTTO+10
8 >8
mutSTTO+20
8 >8
mutST4ol
4 >8
mutST4ol+10
4 >8
mutST4ol+20
4 >8
Legend
mutS = P. aeruginosa PAO1mutS
mutS+20
= P. aeruginosa PAO1mutS subcultured on blood agar only for 20 passages
mutSTTO =
P. aeruginosa PAO1mutS 6% TTO
mutSTTO+10
= P. aeruginosa PAO1mutS 6% TTO plus 10 passages
mutSTTO+20
= P. aeruginosa PAO1mutS 6% TTO plus 20 passages
mutST4ol
= P. aeruginosa PAO1mutS 9% terpinen-4-ol
mutST4ol+10
= P. aeruginosa PAO1mutS 9% terpinen-4-ol plus 10 passages
mutST4ol+20
= P. aeruginosa PAO1mutS 9% terpinen-4-ol plus 20 passages

Chapter Five –Induced resistance to TTO
Table 5.4 Susceptibility of serial subculture isolates of P. aeruginosa to PAβN
(mg/L)
Isolate MIC MBC
PAO1 320 >320
PAO1+20
320 >320
PAO1TTO
10 >320
PAO1T4ol
<2.5 >320
mutS 320 >320
mutS+20
320 >320
mutSTTO
10 >320
mutST4ol
10 >320
Legend
PAO1 = P. aeruginosa PAO1
PAO1+20
= P. aeruginosa PAO1 subcultured on blood agar only for 20 passages
PAO1TTO
= P. aeruginosa PAO1 6% TTO
PAO1T4ol
= P. aeruginosa PAO1 6.5% terpinen-4-ol
mutS = P. aeruginosa PAO1mutS
mutS+20
= P. aeruginosa PAO1mutS subcultured on blood agar only for 20 passages
mutSTTO =
P. aeruginosa PAO1mutS 6% TTO
mutST4ol
= P. aeruginosa PAO1mutS 9% terpinen-4-ol

Chapter Five –Induced resistance to TTO
82
mutSTTO
and mutST4ol
was 10 mg/L, while for PAO1T4ol
it was <2.5 mg/L. The MBC for all
isolates tested was > 320mg/L.
5.2.4 Antibiotic susceptibility
5.2.4.1 Agar dilution
Agar dilution susceptibility screening using CLSI breakpoints for a range of clinically
relevant antibiotics was performed for the serial subculture isolates and parental strains.
The results are shown in Table 5.5. All isolates were resistant to the following antibiotics at
the stated concentration: cefepime (0.5 mg/L), ceftazidime (0.5 mg/L), ceftriaxone
(0.5 mg/L), ceftriaxone (0.5 mg/L) plus clavulanic acid, cephalothin (32 mg/L),
nitrofurantoin (128 mg/L) and trimethoprim (16 mg/L). All isolates were susceptible to the
following antibiotics at the stated concentration: amikacin (16 mg/L), cefepime (8 and
16 mg/L), ceftazidime (8 mg/L), ciprofloxacin (1 mg/L), gentamicin (4 mg/L), meropenem
(4 mg/L), norfloxacin (4 and 16 mg/L) and tobramycin (2 and 4 mg/L). PAO1TTO
developed resistance to aztreonam (32 mg/L), gentamicin (2 mg/L), ticarcillin (128 mg/L)
and Timentin (128 mg/L ticarcillin plus 2 mg/L clavulanic acid), while PAO1T4ol
developed
susceptibility to ceftriaxone (8 mg/L). The mutST4ol
isolates developed sensitivity to 2 mg/L
gentamicin, while mutSTTO
and mutST4ol
developed resistance to aztreonam, ticarcillin and
Timentin.
5.2.4.2 Disc diffusion
Disc diffusion was also used to test susceptibility of the serial subculture isolates and
parental strains to a range of antibiotics. All isolates were susceptible to the following
antibiotics: ceftazidime (30 µg), norfloxacin (10 µg) and imipenem (10 µg). All isolates
were resistant to the following antibiotics: tetracycline (30 µg), furazolidone (100 µg),
cefpodoxime (10 µg), rifampicin (5 µg), clindamycin (2 µg), fusidic acid (10 µg),
novobiocin (5 µg), cefoxitin (30 µg), vancomycin (30 µg), mupirocin (5 µg), erythromycin

Chapter Five –Induced resistance to TTO
Table 5.5 Agar dilution antibiotic susceptibility screening results for serial
subculture isolates of P. aeruginosa
Isolate
Aztreonam
32mg/L
Ceftriaxone
8mg/L
Gentamicin
2mg/L
Ticarcillin
128mg/L Timentin1
PAO1 S R S S S
PAO1+20
S R S S S
PAO1TTO
R R R R R
PAO1T4ol
S S S S S
mutS S R R S S
mutS+20
S R R S S
mutSTTO
R R R R R
mutST4ol
R R S R R
Legend
(1) 128mg/L ticarcillin plus 2mg/L clavulanic acid
PAO1 = P. aeruginosa PAO1
PAO1+20
= P. aeruginosa PAO1 subcultured on blood agar only for 20 passages
PAO1TTO
= P. aeruginosa PAO1 6% TTO
PAO1T4ol
= P. aeruginosa PAO1 6.5% terpinen-4-ol
mutS = P. aeruginosa PAO1mutS
mutS+20
= P. aeruginosa PAO1mutS subcultured on blood agar only for 20 passages
mutSTTO =
P. aeruginosa PAO1mutS 6% TTO
mutST4ol
= P. aeruginosa PAO1mutS 9% terpinen-4-ol
R = resistant
S = sensitive

Chapter Five –Induced resistance to TTO
83
(15 µg) and penicillin G (10 units). All mutS isolates were resistant to ceftriaxone (30 µg).
PAO1TTO
was resistant to ceftriaxone but PAO1, PAO1+20
and PAO1T4ol
were susceptible.
5.2.4.3 Broth microdilution
Broth microdilution was a third method used to investigate the antibiotic susceptibility
profiles of the serial subculture isolates. These results are shown in Tables 5.6a and 5.6b.
By MICs, the PAO1TTO
isolate was more resistant to aztreonam (4-fold), ceftriaxone (at
least 8-fold), chloramphenicol (4-fold), moxalactam (4-to 8-fold), nalidixic acid (8-fold),
and ticarcillin and Timentin (both 4-fold), compared to the PAO1 parent. PAO1T4ol
appeared to be more susceptible to ceftriaxone (at least 4-fold), moxalactam (4-fold),
ticarcillin and Timentin (both 4-fold).
The PAO1+20
isolate was also less susceptible to moxalactam, however the MIC for this
isolate ranged from 16 mg/L (double that of PAO1) to 128mg/L (16-fold the MIC of
PAO1).
Less variation in susceptibility to antibiotics was seen with the mutS mutants. The mutSTTO
isolate had MICs of moxalactam 2- to 8-fold lower than the mutS parent, while mutS+20
and
mutST4ol
had comparable MICs to the parent.
The MIC of ceftriaxone for PAO1+20
ranged from 16 mg/L (equal to that of PAO1) to
256 mg/L (32-fold that of PAO1). For mutS and mutS+20
, the ceftriaxone MIC was
>256 mg/L, while for mutSTTO
it was 128 mg/L and for mutST4ol
the MIC ranged from 16-
256 mg/L.
Changes in susceptibility to imipenem were more difficult to examine due to a lack of
reproducibility. The general trend observed was that the range of MICs for both PAO1T4ol
and mutST4ol
were lower than for their parent strains (see Table 5.6b).

Table 5.6a Broth microdilution antibiotic susceptibilities (mg/L) for serial subculture isolates of P. aeruginosa
Isolate
Aztreonam Chloramphenicol Moxalactam Nalidixic acid
MIC MBC MIC MBC MIC MBC MIC MBC
PAO1 4 4 64 >512 8 16 - 32 64 256
PAO1+20
4 8 64 >512 16 - 128 16 - 256 64 128
PAO1TTO
16 32 256 >512 32 - 64 128 512 >512
PAO1T4ol
4 4 64 >512 2 4 - 32 128 512
mutS 32 32 128 >512 128 - 256 256 256 512
mutS+20
32 32 64 >512 128 256 256 512
mutSTTO
16 32 256 >512 32 - 64 128 512 >512
mutST4ol
16 32 256 >512 128 - 256 256 512 >512
Chap
ter Five –
Induced
resistan
ce to T
TO

Table 5.6b Broth microdilution antibiotic susceptibilities (mg/L) for serial subculture isolates of P. aeruginosa
Isolate
Ceftriaxone Ticarcillin Timentin
MIC MBC MIC MBC MIC MBC
PAO1 8 - 16 64 16 16 16 16
PAO1+20
8 - 256 64 - >256 16 16 32 16 - 128
PAO1TTO
128 >256 64 128 64 128
PAO1T4ol
2 8 - 16 4 8 4 4
mutS >256 >256 128 128 128 128
mutS+20
>256 >256 128 128 64 128
mutSTTO
128 >256 64 128 64 128
mutST4ol
16 - 256 >256 128 128 64 128
Chap
ter Five –
Induced
resistan
ce to T
TO

Chapter Five –Induced resistance to TTO
84
5.2.5 Metal ion susceptibility
The MICs for a range of metal ions was determined for the serial subculture mutants and
parental strains by agar dilution. The results are shown in Table 5.7. MICs for arsenic were
2- to 4-fold higher in the TTO and terpinen-4-ol subculture isolates compared to parental
strains. PAO1T4ol
and mutST4ol
were more susceptible to molybdenum than all other
isolates, however the extent of this could not be accurately determined as the MIC for all
other isolates was beyond the highest concentration able to be tested in this assay. The
MICs for all other metals tested did not differ by more than 2-fold between parental strains
and subculture isolates.
5.2.6 Disinfectant susceptibility
The MICs of benzalkonium chloride, cetrimide, chlorhexidine and triclosan were
determined for the serial subculture isolates and parental strains by broth microdilution
(Table 5.8). Few significant changes in susceptibility to benzalkonium chloride, cetrimide
or chlorhexidine were seen. PAO1T4ol
had a higher MBC of chlorhexidine although the
MIC was not significantly different and the MIC of cetrimide was also 4-fold lower. The
MIC of chlorhexidine for mutSTTO
was 4-fold lower as was the MBC of cetrimide. mutST4ol
was also more susceptible to chlorhexidine with an MIC of 1mg/L (4-fold lower).
Due to the insolubility of triclosan in aqueous solution, the concentrations tested by broth
microdilution assay were up to 128 mg/L. The MIC of triclosan against the parental strains
was >128 mg/L, while the MIC for all mutants was 128 mg/L. In an attempt to test this
disinfectant at a higher concentration, triclosan also tested by agar dilution. This involved
solubilizing the triclosan in DMSO prior to incorporation into MHA however the triclosan
precipitated out of the agar and further testing was not undertaken.

Table 5.8 Broth dilution MICs and MBCs (mg/L) for disinfectants against serial subculture isolates of P. aeruginosa.
Isolate
Chlorhexidine Cetrimide Benzalkonium chloride
MIC MBC MIC MBC MIC MBC
PAO1 2 8 256 512 64 64
PAO1+20
4 4 256 256 64 64
PAO1TTO
1 4 128 512 64 128
PAO1T4ol
1 32 64 512 64 128
mutS 4 8 256 1024 64 64
mutS+20
4 4 256 512 64 128
mutSTTO
1 4 256 256 128 128
mutST4ol
1 8 256 512 64 128
Chap
ter Five –
Induced
resistan
ce to T
TO

Table 5.7 Agar dilution MICs for various metal ions against serial subculture isolates of P. aeruginosa 1
Isolate
Arsenic
(mM)
Barium
(mM)
Cadmium
(mM)
Cobalt
(mM)
Iron
(mM)
Manganese
(mM)
Mercury
(μM)
Molybdenum
(mM)
PAO1 19.2 4.8 9.6 4.8 9.6 19.2/38.4 9.4 >9.6
PAO1+20
19.2 4.8 9.6 4.8 9.6 19.2/38.4 9.4 >9.6
PAO1TTO
38.4/76.8 2.4 4.8/9.6 4.8 9.6 38.4 9.4 >9.6
PAO1T4ol
38.4 2.4 4.8 2.4/4.8 4.8 19.2 9.4 9.6
mutS 19.2 4.8 9.6 4.8 9.6 38.4 9.4 >9.6
mutS+20
19.2 4.8 9.6 4.8 9.6 38.4 ≤4.7 >9.6
mutSTTO
76.8 2.4 4.8/9.6 4.8 9.6 38.4 9.4 >9.6
mutST4ol
76.8 2.4 4.8 4.8 9.6 38.4 9.4 9.6
1 MICs for silver, nickel and lead for all isolates tested were 37.5µM, 4.8mM and >4.8mM, respectively.
Chap
ter Five –
Induced
resistan
ce to T
TO

Chapter Five –Induced resistance to TTO
85
5.2.7 Fatty acid analysis
Fatty acid analysis was performed on the serial subculture isolates and parental strains
using the MIDI system. Results are shown in Table 5.9 (PAO1) and Table 5.10 (mutS).
Thirty two different fatty acids were identified in the isolates tested. The MIDI system was
unable to distinguish between 18:1 ω6c and 18:1 ω7c; 16:1 ω6c and 16:1 ω7c; 19:1 ω7c
and 19:1 ω6c; 18:2 ω6,9c and 18:0 ANTE; nor 14:0 3OH and 16:1 iso, and so their
combined percentage is presented. A student‟s paired T-test with unequal variance was
used to compare the fatty acid composition of each parental strain and respective tolerised
isolates. No significant differences (P > 0.05) were detected with respect to short chain
fatty acids, long chain fatty acids, cyclic fatty acids, or the ratio of short:long fatty acids.
Similarly, no significant differences were detected for saturated fatty acids, unsaturated
fatty acids and the ratio of saturated:unsaturated fatty acids.
5.2.8 Outer membrane permeability
The whole cell alkaline phosphatase assay was used to determine the index of outer
membrane permeability (OMP) for the serial subculture isolates and parental strains (Figure
5.1). The index of OMP for PAO1T4ol
was increased by approximately 40% compared to
the parent strain. A similar increase was seen for PAO1+20
, while the index for PAO1TTO
was comparable to the parent strain. The indices of OMP for mutSTTO
and mutST4ol
were
increased by approximately 30% and 50%, respectively, compared to mutS while the index
for mutS+20
was increased by approximately 13%. The standard deviation for this assay
ranged from the equivalent of 26% to 64% of the value of the index.
5.2.9 Cell surface hydrophobicity
Several different methods were used in an attempt to determine the cell surface
hydrophobicity of the parent strains and serial subculture isolates.

Chapter Five –Induced resistance to TTO
Table 5.9 Average composition and ratios of fatty acids of P. aeruginosa PAO1 and
serial subculture mutants
Fatty acid
Composition (%) in isolate
PAO1 PAO1+20
PAO1TTO
PAO1T4ol
10:0 0.62 0.80 1.26 0.43
12:0 5.17 5.97 3.27 6.27
13:0 0.07 0.07 0.04 0.06
14:0 0.49 0.60 0.56 0.64
16:0 24.18 22.38 24.78 24.29
16:0 3OH
17:0 0.07 0.07 0.05 0.05
18:0 0.20 0.20 0.16 0.18
19:0
17:0 cyclo 1.10 1.13 2.27 1.79
19:0 cyclo ω7c 1.35 1.21 2.33 2.12
8:0 3OH 0.14 0.15 0.30
10:0 3OH 9.07 10.01 10.85 9.81
11:0 3OH 0.08 0.08 0.09
12:0 2OH 10.05 11.60 11.71 11.02
12:0 3OH 8.94 9.76 8.90 9.58
12:1 3OH 0.73 0.77 0.87 0.95

Chapter Five –Induced resistance to TTO
Fatty acid Composition (%) in isolate
PAO1 PAO1+20
PAO1TTO
PAO1T4ol
17:1 ω8c 0.10 0.08 0.06 0.05
18:1 ω7c 11 methyl
18:1 ω6c/ 18:1 ω7c 27.33 24.53 22.22 24.01
18:1 ω9c 0.17 0.17 0.37
16:1 ω6c/ 16:1 ω7c 10.07 10.32 9.65 8.74
15:1 ω6c
19:1 ω7c/ 19:1 ω6c
15:1 iso F 0.04
18:2 ω6,9c / 18:0
ANTE 0.07 0.07 0.23
14:0 3OH/ 16:1 iso I 0.05 0.06 0.05 0.06
Ratio SAT:UNSAT 1.60 1.78 2.00 1.96
Sum SAT 61.51 64.00 66.53 66.21
Sum UNSAT 38.39 35.87 33.15 33.74
Sum cyclo 2.45 2.34 4.60 3.90
Table 5.9 (continued). Average composition and ratios of fatty acids of P. aeruginosa
PAO1 and serial subculture mutants

Chapter Five –Induced resistance to TTO
Table 5.10 Average composition and ratios of fatty acids of P. aeruginosa
PAO1muts and serial subculture mutants
Fatty acid
Composition (%) in isolate
mutS mutS+20
mutSTTO
mutST4ol
10:0 0.51 0.36 0.46 0.44
12:0 5.00 5.15 5.14 3.10
13:0 0.06 0.05
14:0 0.61 0.55 0.50 0.82
16:0 23.99 24.43 22.54 28.59
16:0 3OH 0.06
17:0 0.16 0.57 0.22 0.31
18:0 0.05 0.26 0.07
19:0 0.05
17:0 cyclo 1.63 1.43 1.44 2.23
19:0 cyclo ω7c 1.61 2.12 1.82 1.96
8:0 3OH 0.19 0.12 0.34 0.09
10:0 3OH 10.12 7.87 12.07 7.74
11:0 3OH 0.07 0.29 0.09
12:0 2OH 11.46 9.30 11.66 11.14
12:0 3OH 9.49 8.51 10.18 8.03
12:1 3OH 0.83 0.64 1.39 0.11

Chapter Five –Induced resistance to TTO
Fatty acid
Composition (%) in isolate
mutS mutS+20
mutSTTO
mutST4ol
17:1 ω8c 0.06 0.42 0.07
18:1 ω7c 11 methyl 0.06
18:1 ω6c/ 18:1 ω7c 23.73 27.20 22.83 23.54
18:1 ω9c 0.27 0.43
16:1 ω6c/ 16:1 ω7c 10.24 10.32 8.71 11.20
15:1 ω6c 0.06
19:1 ω7c/ 19:1 ω6c 0.07
15:1 iso F
18:2 ω6,9c / 18:0
ANTE
0.10 0.23 0.12 0.31
14:0 3OH/ 16:1 iso I 0.08 0.06 0.07
Ratio SAT:UNSAT 1.87 1.57 2.00 1.83
Sum SAT 64.98 60.97 66.56 64.42
Sum UNSAT 34.84 38.63 33.26 35.27
Sum cyclo 3.24 3.55 3.26 4.19
Table 5.10 (continued) Average composition and ratios of fatty acids of P. aeruginosa
PAO1muts and serial subculture mutants

Chapter Five –Induced resistance to TTO
Figure 5.1 Outer membrane permeability of serial subculture mutants and
parental strains of P. aeruginosa
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
1.60
Organism
Rela
tive o
ute
r m
em
bra
ne p
erm
eab
ilit
y
PAO1
PAO1+20
PAO1 TTO
PAO1 T4ol
mutS
mutS +20
mutS TTO
mutS T4ol
Legend
PAO1 = P. aeruginosa PAO1
PAO1+20
= P. aeruginosa PAO1 subcultured on blood agar only for 20 passages
PAO1TTO
= P. aeruginosa PAO1 6% TTO
PAO1T4ol
= P. aeruginosa PAO1 6.5% terpinen-4-ol
mutS = P. aeruginosa PAO1mutS
mutS+20
= P. aeruginosa PAO1mutS subcultured on blood agar only for 20 passages
mutSTTO =
P. aeruginosa PAO1mutS 6% TTO
mutST4ol
= P. aeruginosa PAO1mutS 9% terpinen-4-ol

Chapter Five –Induced resistance to TTO
86
5.2.9.1 Microbial adhesion to hydrocarbon (MATH)
The MATH method was attempted with the parent isolates, however despite trying two
different methods (with various optimisations) and two different organic solvents, results
were preliminary and not reproducible.
5.2.9.2 Adhesion to polystyrene beads (APSB)
The APSB method was also attempted with the parent isolates and again results were
preliminary and not reproducible.
5.2.9.3 Flow cytometry
As mentioned in 2.2.4.4.1, a method using flow cytometry to determine cell surface
hydrophobicity in P. aeruginosa was not able to be optimised. While flow cytometry has
previously been used to successfully determine cell surface hydrophobicity for a range of
yeasts, including Candida and Saccharomyces species (Colling et al., 2005), it has not been
used for bacteria.
5.2.10 PFGE
The serial subculture isolates were compared to parental strains using PFGE. The enzyme
used for digestions was SpeI (Figure 5.2). The isolates were indistinguishable by this
method.
5.3 Discussion
5.3.1 Selection of resistance to TTO and terpinen-4-ol by serial subculture
Serial subculture was used to select several mutants with increased resistance to TTO
and/or terpinen-4-ol. The maximum TTO concentration reached in subculture of both the
PAO1 and mutS derived isolates was 6%, after 46 and 45 passages, respectively. This

Chapter Five –Induced resistance to TTO
Figure 5.2 PFGE of serial subculture mutants and parental strains of P. aeruginosa
digested with SpeI.
Lanes: 1 & 8 = Salmonella enterica serovar Braenderup H9812 (BAA-664™) standard;
band sizes from top of well to bottom in kb: 1135, 668.9, 452.7, 398.4, 336.5, 310.1,
244.4, 216.9, 173.4 (a double), 138.9, 104.5, 78.2, 54.7, 33.3, 28.8, and 20.5. 2 = PAO1;
3 = PAO1TTO
; 4 = PAO1T4ol
; 5 = mutS; 6 = mutSTTO
; 7 = mutS
T4ol.
1 2 3 4 5 6 7 8

Chapter Five –Induced resistance to TTO
87
concentration is equal to 1.5-3-fold and 0.75-fold the TTO MICs of the respective parent
strains. In the case of mutS the maximum concentration reached was less than the MIC for
the parent (see Tables 5.2 and 5.3). There are two possible explanations as to why this is so.
Firstly, the broth microdilution assay used to determine MICs and the serial subculture
assay are two very different systems; the first is a stationary tray with a small volume, the
second is a larger volume that is shaking constantly. Therefore it is likely that the oil or
component is dispersed throughout the broth more thoroughly during serial subculture and
effectively more of the oil or component is able to act on the bacteria. Broth microdilution
has its limitations when used with less water soluble agents like essential oils, however this
method is widely accepted in the literature, particularly when a detergent like Tween 80 is
used to enhance solubility (Carson et al., 1995; Hammer et al., 1996).
The second explanation is related to the vessels used for the experiments: the microdilution
trays were plastic while the serial subculture was done in glass. TTO has been shown to
interact with, and may deform, certain plastics (Rowe, 1999). The MICs of TTO and
terpinen-4-ol against Candida albicans in microdilution trays were two times those
determined by macrodilution in glass bottles (Hammer et al., 2003). The MICs may have
been higher than the maximum subculture value reached because the oil or component was
interacting with the microdilution tray. Alternatively the serial subculture in glass bottles
may have allowed better interaction between microbe and oil, and therefore a lower
maximum concentration was tolerated. Also, the maximum TTO concentration reached for
mutS (6%) was between the MIC (8%) and the next concentration down (4%) – an MIC of
8% indicates that the MIC is higher than 4% but less than or equal to 8%, therefore this
could actually be 6%.
A study on the step-wise induction of resistance to TTO in five isolates of S. aureus
reported three isolates with TTO MICs of 1%, and the remaining two isolates with MICs of
2% and 16%, respectively (Nelson, 2000). The initial TTO MIC for all isolates was 0.25%.
To induce resistance, these isolates were initially exposed to 2.5% TTO and resistant clones
were selected, then plated onto TTO-containing agar plates (0.2-4%), in a step-wise
fashion. The author failed to specify what concentration of TTO was contained in the agar

Chapter Five –Induced resistance to TTO
88
plate that the isolate with the MIC of 16% came from. It is interesting to note the MIC of
16%, as the experience in our laboratory is that, even with the aid of Tween 80, TTO is
very difficult to solubilize at concentrations over 8% (v/v). The increase in MICs reported
in the Nelson study ranged from 4- to 64-fold, but it is difficult to make comparisons to this
study due to the differences in methodology as well as the fundamental issue of a difference
in organism. Attempts by our research group to obtain the isolates created by Nelson were
unsuccessful.
The concentrations of terpinen-4-ol reached in the subcultured isolates were greater than
the MICs for the parental strains (Tables 5.2 and 5.3). The final concentrations reached
were 6.5% and 9% for the PAO1- and mutS-derived isolates, respectively. These
concentrations represent 3.5-fold and 2.25-fold the terpinen-4-ol MICs for the respective
parent strains.
P. aeruginosa seemed better able to adapt to increasing concentrations of terpinen-4-ol than
those of TTO. This suggests that P. aeruginosa is capable of overcoming the mechanism of
action of terpinen-4-ol but not that of TTO. As a single chemical terpinen-4-ol may have
only one mechanism of antimicrobial action while TTO, with close to 100 components,
may have multiple mechanisms. However, the mechanism of action of neither agent is fully
understood.
Terpinen-4-ol did not increase the susceptibility of P. aeruginosa to lysis by detergents
(Cox, 2007), suggesting that it does not act on the outer membrane. On the other hand, pre-
treatment with the outer membrane permeabilizer EDTA increased susceptibility to
terpinen-4-ol (Longbottom et al., 2004). Treatment with 2% terpinen-4-ol for 60 min
resulted in many unidentified electron-sparse inclusions in P. aeruginosa cells viewed by
transmission electron microscopy (Longbottom et al., 2004). Treatment of S. aureus with
terpinen-4-ol caused the leakage of 260-nm-light-absorbing materials and increased
susceptibility to sodium chloride and precipitated the formation of mesosome-like
structures (Carson et al., 2002). The specific mechanism of action of terpinen-4-ol is yet to

Chapter Five –Induced resistance to TTO
89
be elucidated and despite some contrasting reports in the literature, probably involves
disruption of the outer membrane.
5.3.1.1 Use of mutS
MutS is involved in the bacterial mismatch repair system (Smania et al., 2004). The mutS
parent strain was chosen for use in the serial subculture experiments because it is a
hypermutable strain with morphotypes that are often highly antibiotic resistant (Oliver et
al., 2004; Smania et al., 2004). It was thought that the use of this hypermutable strain
would increase the maximum concentration of TTO or terpinen-4-ol that P. aeruginosa was
subcultured in. The mutS strain was already more resistant to both TTO and terpinen-4-ol
than PAO1, but the only notable difference seen from the subculture experiments was that
mutST4ol
was cultured in a maximum of 9% terpinen-4-ol while PAO1T4ol
only reached
6.5% terpinen-4-ol.
5.3.2 Susceptibility of subculture isolates
5.3.2.1 TTO and terpinen-4-ol susceptibility
5.3.2.1.1 Following subculture in TTO
Serial subculture up to a final concentration of 6% TTO resulted in a PAO1-derived isolate
with a TTO MIC of 8%. As the broth microdilution method incorporates serial doubling
dilutions, what this value really indicates is that the MIC was somewhere between 4% and
8%. The MIC for the PAO1 parent was 2-4% therefore this increase in MIC in PAO1TTO
was considered modest. Also, following subculture in the absence of TTO, the MIC for
PAO1TTO
dropped to 4% (10 subcultures) then 2% (20 subcultures). Thus the increase seen
was likely a transient phenotypic change.
The serial subculture up to a final concentration of 6% TTO did not affect the TTO MIC for
mutSTTO
when compared to mutS. There was an increase in terpinen-4-ol MIC for the
mutSTTO
isolate, from 4% in the parent to >8% and no change was seen following

Chapter Five –Induced resistance to TTO
90
subculture in the absence of TTO. The terpinen-4-ol susceptibility pattern matched that of
the PAO1TTO
isolate, with sustained increases in terpinen-4-ol MIC seen in both isolates.
The mutS isolate already had a lower susceptibility to TTO, with an MIC of 8%, so it is not
surprising that growth in 6% TTO did not alter the MIC.
Interestingly, it was the terpinen-4-ol MIC that was altered the most in the PAO1TTO
isolate, going from 2% to >8%, a minimum 4-fold increase. Following subculture without
terpinen-4-ol, the MIC dropped to 8%, which still equates to a 4-fold increase in MIC. The
increase in MIC for terpinen-4-ol following exposure to TTO poses the question – why
would exposure to a complex mixture of over 100 terpene components result in increased
tolerance to this single terpene component? Perhaps this result adds further support to the
notion that terpinen-4-ol is the primary antimicrobial component of TTO or at least that
terpinen-4-ol has a mechanism of action that is fundamental to the action of TTO but may
be shared by other components.
5.3.2.1.2 Following subculture in terpinen-4-ol
When PAO1 was subcultured in the presence of terpinen-4-ol, a different susceptibility
pattern was seen. The terpinen-4-ol MICs did not alter. Even though the PAO1T4ol
isolate
was subcultured in up to 6.5% terpinen-4-ol, the MIC was still 2%. This was somewhat
unexpected – that an isolate with a terpinen-4-ol MIC of 2% could be cultured in 6.5%
terpinen-4-ol, more than three times the concentration that inhibited the isolate. The
differences in test conditions may play a role here; the static 96-well tray versus the shaking
broth. Because P. aeruginosa is an obligate aerobe, the cells will generally gather at the top
of the vessel they are grown in, to allow access to the maximal amount of oxygen. This is
particularly relevant in static vessels, such as microtitre trays, while in a shaking vessel, the
oxygen and therefore the cells would be dispersed throughout the growth medium.
Although terpinen-4-ol has higher water solubility than whole TTO, it will begin to
separate out of solution over time and therefore some separation may occur in a static
microtitre tray. The end result may be that there is a higher concentration of terpinen-4-ol at
the surface, as well as a higher concentration of cells, which leads to more cell death. But

Chapter Five –Induced resistance to TTO
91
when the cells are grown in a shaking broth, oxygen, the component and the cells are more
evenly dispersed throughout the broth and so it is possible that the cells can survive in a
higher concentration of agent. It may be useful to determine TTO and terpinen-4-ol MICs
in a shaking microtitre tray.
The PAO1T4ol
isolate did not have an altered MIC for TTO compared to PAO1. This was
expected as exposure to one component is not likely to decrease susceptibility to the multi-
component TTO if TTO has a different mechanism of action or additional mechanisms.
The mutST4ol
isolate, subcultured in 9% terpinen-4-ol, did not have an altered susceptibility
to TTO when compared to mutS. This also follows the pattern observed with PAO1T4ol
outlined above – exposure to one component was not expected to result in tolerance to
TTO. The mutST4ol
isolate did have an increase in terpinen-4-ol MIC, with the MIC going
from 4% to >8%. This represents at least a doubling of MIC, which was not considered
significant, however it does suggest that tolerance to a single component may occur over
time.
The increased tolerance to observed in the isolates subcultured in terpinen-4-ol was
maintained in the absence of terpinen-4-ol, suggesting that a genotypic change may have
caused this increased tolerance.
5.3.2.2 Antibiotic susceptibility
5.3.2.2.1 β-Lactam antibiotics
In the disc diffusion assays all isolates were deemed sensitive to ceftriaxone, a 3rd
generation cephalosporin, except PAO1TTO
which was resistant. In agar dilution assays all
serial subculture isolates were resistant to 8 mg/L except for PAO1T4ol
which was sensitive.
Other discrepancies between testing methods were also observed. PAO1+20
and mutST4ol
both had MICs for ceftriaxone of 16-256 mg/L by broth microdilution; the CLSI guidelines
set the resistant breakpoint MIC for ceftriaxone as ≥64 mg/L, while 16-32 mg/L is

Chapter Five –Induced resistance to TTO
92
considered intermediate and ≤8 mg/L is susceptible (CLSI, 2007). Therefore the broth
microdilution results span the intermediate to resistant range, yet the disc diffusion results
classify PAO1+20
as susceptible. Despite numerous attempts it was not possible to
determine a consistent MIC value for either isolate. The mutS, mutS+20
and mutSTTO
were
all classified as resistant according to broth microdilution while mutST4ol
was intermediate
to resistant, yet none of these isolates was resistant in the disc diffusion assay. The one
isolate classified as susceptible by agar dilution, PAO1T4ol
, was also susceptible to
ceftriaxone by broth microdilution as well as disc diffusion. Overall the increase in
susceptibility to ceftriaxone in PAO1T4ol
and the decrease in susceptibility in PAO1TTO
appear supported by these results.
Cephalosporins belong to the β-lactam family of antimicrobials and interrupt the synthesis
of the peptidoglycan component of bacterial cell walls by binding to and inhibiting
penicillin-binding proteins (PBPs) in the cytoplasmic membrane, ultimately leading to cell
death (Aoki, 2005). Resistance to ceftriaxone is encoded mainly by the production of some
β-lactam hydrolyzing enzymes and in P. aeruginosa has also been associated with the
MexAB-OprM pump (Li et al., 1995). It seems unlikely that continuous exposure to TTO
would encourage the production of β-lactamases, however it may induce an increase in
expression of MexAB-OprM and thus resistance to ceftriaxone may arise. The reason for
the increase in susceptibility to ceftriaxone observed with PAO1T4ol
is not clear.
The PAO1TTO
isolate was 4- to 8-fold more resistant to moxalactam than PAO1. The
susceptibility of the PAO1+20
isolate was also decreased compared to PAO1, with the MIC
ranging from 2- to 16-fold that of PAO1. The reason for such a range of susceptibilities in
PAO1+20
is not clear, though a similar pattern was observed with ceftriaxone susceptibility
(see above). Moxalactam is an oxacephalosporin, a member of the β-lactam family and a
substrate of the MexAB-OprM efflux system (Masuda et al., 2000a). The apparent decrease
in susceptibility to moxalactam seen in PAO1TTO
may be mediated by an increase in
expression of the MexAB-OprM pump as a response to the presence of TTO.
Interestingly the mutSTTO
isolate was 2- to 8-fold more susceptible to moxalactam than
mutS, while mutS+20
and mutST4ol
had comparable MICs to the parent. Increased

Chapter Five –Induced resistance to TTO
93
susceptibility to moxalactam has been associated with nfxB mutants of P. aeruginosa,
however an increase in susceptibility to imipenem, aztreonam and gentamicin was also
observed (Masuda, 1996) and this pattern was not seen with mutSTTO
. PAO1T4ol
was also
more susceptible to moxalactam, as indicated by a decrease in MIC to ¼ the MIC for
PAO1. As mentioned above, the susceptibility pattern of PAO1T4ol
was similar to that of
nfxB isolates, indicating that PAO1T4ol
may have the expression of MexCD-OprJ induced.
PAO1TTO
was 4-fold more resistant by broth microdilution to ticarcillin and Timentin than
PAO1 and classified resistant to both agents when tested by agar dilution. Ticarcillin is a
carboxypenicillin, a member of the β-lactam family and bactericidal by preventing cross-
linking of peptidoglycan during cell wall synthesis. Timentin is a mixture of ticarcillin plus
clavulanic acid, a β-lactamase inhibitor. Resistance to ticarcillin in P. aeruginosa has been
associated with the overexpression of OprM, and hence related efflux components such as
MexAB or MexXY, the production of acquired β-lactamase and the overexpression of
chromosomally encoded Amp C cephalosporinase (Cavallo et al., 2002). MexAB-OprM-
and MexXY overproducing mutants are very prevalent among clinical strains of
P. aeruginosa with reduced susceptibility to ticarcillin (Hocquet et al., 2007). Of the three
known mechanisms of resistance to ticarcillin, the overexpression of efflux pumps is more
likely to occur in this case. The addition of clavulanic acid did not change the MIC, and
therefore β-lactamases are unlikely to be involved, and there is no obvious advantage to
producing Amp C or β-lactamases to aid in the tolerance to TTO. The ability to pump out
the oil or components of the cell via efflux would be beneficial. On the other hand,
PAO1T4ol
was 4-fold more susceptible to ticarcillin and Timentin than PAO1. Susceptibility
to ticarcillin has been associated with expression of the MexCD-OprJ system in
P. aeruginosa, when hypersusceptibility to both ticarcillin and aminoglycosides was
detected (Llanes, 2004).
In agar dilution assays, PAO1TTO
, mutSTTO
and mutST4ol
were resistant to 32 mg/L
aztreonam while all other isolates were sensitive. In broth microdilution assays the MIC of
aztreonam against PAO1TTO
was increased 4-fold compared to PAO1, while a 2-fold
decrease in the MIC of aztreonam was seen for mutSTTO
and mutST4ol
. Aztreonam is a

Chapter Five –Induced resistance to TTO
94
monobactam and a member of the β-lactam family. Like all β-lactam antibiotics, it is
bactericidal and interferes with cell wall biosynthesis (Thomson and Lister, 2004).
Resistance to aztreonam is often mediated by the MexAB-OprM efflux system (Masuda et
al., 2000a; Quale et al., 2006). P. aeruginosa usually produces low levels of chromosomal
AmpC cephalosporinase, however high level production of AmpC in derepressed mutants
provides resistance to aztreonam and most other β-lactams (Thomson and Lister, 2004). In
PAO1TTO
the resistance is likely to be mediated by an increase in the expression of
MexAB-OprM. The discrepancy in results for the mutS isolates is probably because the
broth microdilution MICs are close to the concentration of aztreonam used in the agar
dilution assays (32 mg/L). The broth microdilution MIC for mutS and mutS+20
was 32 mg/L
while for mutSTTO
and mutST4ol
it was 16 mg/L, a small 2-fold difference. It is feasible that
mutSTTO
and mutST4ol
could be resistant to 32 mg/L in an agar plate during one experiment
but susceptible to 16 mg/L in a microdilution tray in another experiment.
In a recent study, Hocquet et al. (2007) showed that AmpC de-repression in P. aeruginosa
caused a 4- to 16-fold increase in MICs for a range of β-lactams, while levels of efflux-
based resistance were lower. A 2- to 8-fold increase in MIC was seen in MexAB-OprM-
overproducing mutants, but only for ticarcillin and aztreonam. This provides further
support for MexAB-OprM-overproduction causing the modest increases in ticarcillin and
aztreonam MICs observed for PAO1TTO
. Over-expression of MexAB-OprM has been
associated with mutations in the repressor mexR gene (Adewoye et al., 2002; Boutoille et
al., 2004) and also inactivation of nalC (Cao, 2004) or nalD (Sobel, 2005). There are also
roles for quorum sensing molecules that act independently of mexR (Maseda, 2004; Sawada
et al., 2004).
5.3.2.2.2 Quinolones
An 8-fold increase in MIC was detected for nalidixic acid against PAO1TTO
compared with
PAO1. Nalidixic acid is a quinolone antibiotic with a bactericidal action via the inhibition
of DNA synthesis through binding to topoisomerases II and IV (Andriole, 2004).
Resistance to quinolones is mediated by mutations in the target molecules, topoisomerases

Chapter Five –Induced resistance to TTO
95
II and IV, or by efflux, as quinolones are substrates of MexAB-OprM, MexCD-OprJ and
MexXY-OprM (Masuda et al., 2000a). Therefore, induced expression of MexCD-OprJ, or
increased expression of MexAB-OprM or MexXY-OprM, may account for this increase in
resistance to nalidixic acid.
5.3.2.2.3 Chloramphenicol
The MIC of chloramphenicol was 4-fold higher in PAO1TTO
than in the PAO1 parent.
Chloramphenicol is a bacteriostatic antimicrobial that inhibits the peptidyl transferase
activity of the bacterial ribosome, preventing peptide bond formation (Schwarz and White,
2005). Resistance to chloramphenicol may arise by several mechanisms: mutation of the
50S ribosomal subunit, elaboration of chloramphenicol acetyltransferase or via efflux
pumps (Schwarz and White, 2005). It is not likely that mutation of the 50S ribosomal
subunit or the expression of chloramphenicol acetyltransferase would provide an advantage
to P. aeruginosa in the presence of TTO. Therefore the decrease in susceptibility to
chloramphenicol was probably due to efflux. The MexAB-OprM, MexCD-OprJ, MexEF-
OprN and MexXY-OprM efflux systems are all capable of exporting chloramphenicol
when expressed in P. aeruginosa (Li et al., 1995; Maseda et al., 2000; Masuda et al.,
2000a; Masuda, 1996). Adaptation to the disinfectant benzalkonium chloride in E. coli
resulted in increased efflux, as measured by efflux of ethidium bromide (Langsrud et al.,
2004).
5.3.2.2.4 Aminoglycosides
Aminoglycosides bind to the 30S ribosomal subunit, inhibiting protein synthesis, causing
leaky membranes and cell death (Stone, 2003). Gentamicin resistance in P. aeruginosa may
be mediated by expression of MexXY-OprM (Masuda et al., 2000b). Other general
mechanisms of aminoglycoside resistance include alteration of binding sites, reduced
uptake/permeability and aminoglycoside-modifying enzymes (Stone, 2003). PAO1TTO
developed resistance to gentamicin (2 mg/L) following subculture with TTO, which may be

Chapter Five –Induced resistance to TTO
96
related to expression of MexXY-OprM. mutST4ol
developed sensitivity to gentamicin, which
may be due to some alteration of the outer membrane.
5.3.2.2.5 Clinical relevance
When compared to the parental strain PAO1, the serial subculture isolate PAO1TTO
had
increased MICs for eight antibiotics. The increase in MIC was considered clinically
significant if it meant the isolate was classified as resistant according to CLSI guideline
breakpoints (CLSI, 2007), while the parent strain was classified susceptible. This occurred
for ceftriaxone, where the parent was classified as sensitive/intermediate and PAO1TTO
was
classified as resistant. In the case of moxalactam, PAO1 was classified as sensitive while
PAO1TTO
was classified as intermediate/resistant. The classification for aztreonam changed
from susceptible in the parent to intermediate in PAO1TTO
. The change in susceptibility of
PAO1TTO
to these three β-lactam antibiotics would be relevant in a clinical setting, although
a large range of antibiotics from other classes still exist as potential treatment for
P. aeruginosa infections.
Although the MIC for chloramphenicol against PAO1TTO
was considered resistant
(256 mg/L), the parent strain was also resistant (64 mg/L) and so this change would have
no clinical significance.
5.3.2.2.6 Other studies
McMahon et al. (2007) found that S. aureus, E. coli and Salmonella spp. exposed to sub-
lethal concentrations of TTO displayed reduced susceptibility to clinically relevant
antibiotics, compared with control strains (McMahon et al., 2007). The antibiotics tested
included tetracycline, gentamicin and chloramphenicol, although in some cases the increase
in MIC in the TTO-exposed isolates was a doubling or less. The mechanism(s) of decreased
susceptibility were not explored by the authors.

Chapter Five –Induced resistance to TTO
97
Resistance to pine oil, comprised mostly of cyclic hydrocarbons, was associated with
resistance to tetracycline, ampicillin, chloramphenicol and nalidixic acid, and
overexpression of marA, in E. coli mutants (Moken et al., 1997). The multidrug efflux
pump AcrAB was positively regulated in part by marA and susceptibility to pine oil was
increased following deletion of the AcrAB efflux pump or marA (Moken et al., 1997).
5.3.2.3 Metal ion susceptibility
Arsenic was the only metal ion tested for which serial subculture isolates had decreased
susceptibility. Arsenic can exist in multiple oxidation states, with the most common being
arsenite [As (III)] and arsenate [As(V)]. Arsenic is toxic to most microorganisms, however,
resistance to arsenic in pseudomonads has been documented and is associated with the
arsRBCH operon (Patel et al., 2007). Ars-mediated resistance involves the reduction of
arsenate to arsenite via ArsC, a cytoplasmic arsenate reductase. The arsenite is then
extruded by a membrane-associated ArsB efflux pump that is efficient at removing arsenite
as well as antimonite (Parvatiyar et al., 2005). ArsR is a predicted transcriptional regulator
while ArsH is a conserved hypothetical protein of unknown function (Winsor et al., 2005).
The efflux of arsenic in E. coli is driven by ATP, not proton-motive force (Mobley and
Rosen, 1982).
In this work, the MICs for arsenate were 2- to 4-fold higher in the TTO and terpinen-4-ol
subculture isolates compared to parental strains. While this was a modest increase in MIC,
it may be associated with up-regulation of the ars efflux operon in association with
prolonged exposure to TTO and terpinen-4-ol. It is unlikely that TTO and its components
are substrates of this efflux pump, however there may be some association. Any role played
by the ars efflux operon could be confirmed by RT-PCR.
5.3.2.4 Disinfectant susceptibility
Although PAO1T4ol
had an MIC for cetrimide that was ¼ that of PAO1, both strains had the
same MBC. No differences in susceptibility to benzalkonium chloride were noted.

Chapter Five –Induced resistance to TTO
98
Quaternary ammonium compounds such as cetrimide and benzalkonium chloride are
cationic detergents with hydrophilic and lipophilic groups that react with the lipids of
bacterial cell membranes. Membrane surface properties and permeability are thus altered,
causing loss of cell contents and cell death (Ryan, 2004).
Chlorhexidine is a cationic agent which acts by increasing the permeability of cell
membranes (Ryan, 2004). The MBC for chlorhexidine was 4-fold higher for PAO1T4ol
compared with the parent strain, which may indicate alterations in the outer membrane.
The susceptibility of serial subculture isolates to Triclosan was also investigated. Due to the
insolubility of Triclosan, the highest concentration tested by broth microdilution assay was
128 mg/L. The MIC of Triclosan against the parent strains was >128 mg/L, while for all
serial subculture isolates it was 128 mg/L. In an attempt to test this disinfectant at a higher
concentration, Triclosan also tested by agar dilution. This involved solubilizing the
Triclosan in DMSO prior to incorporation into MHA, however the Triclosan precipitated
out of the agar. The plates were tested regardless and the MICs for all isolates tested on
these plates was determined as >1024 mg/L. Given the lower values observed in the broth
microdilution, it was concluded that the precipitation of Triclosan out of the agar had
compromised the validity of the assay and therefore the results were discarded.
Previously reported MICs of Triclosan against PAO1 are >256 mg/L by broth
microdilution (Champlin et al., 2005) and >1024 mg/L by agar dilution (Chuanchuen et al.,
2003). Chuanchuen et al. (2003) used 2-methoxyethanol to dissolve Triclosan for use in
agar dilution. This solvent could not be obtained within a realistic timeframe and therefore
the Triclosan susceptibilities could not be determined with more accuracy. Given the
published MIC values, it is possible that PAO1TTO
, PAO1T4ol
, mutSTTO
and mutST4ol
are
more susceptible to Triclosan by a factor of at least 8, if not more. Triclosan is a phenolic
biocide that has several modes of action. It is bacteriostatic at low concentrations and
bactericidal at high concentrations. It has been shown to inhibit bacterial fatty acid
synthesis at the enoyl-acyl carrier protein reductase step (Heath et al., 1999; Hoang and
Schweizer, 1999) and also cause leakage of potassium ions (Suller and Russell, 2000).

Chapter Five –Induced resistance to TTO
99
5.3.2.5 PAβN
The efflux pump inhibitor PAβN is able to inhibit the clinically relevant Mex efflux
systems of P. aeruginosa (Mesaros et al., 2007). Given the antibiotic susceptibility profiles
of the serial subculture isolates, an increase in efflux was suspected as a mechanism for the
increase in resistance noted. MICs of TTO and terpinen-4-ol for the resistant isolates with
PAβN were expected to indicate a role for efflux. Preliminary experiments to establish the
MIC of PAβN instead demonstrated that these isolates were substantially more susceptible
to PAβN than wild type strains. Campo-Esquisabel et al. (2007) noted that two clinical
isolates of P. aeruginosa were more susceptible to PAβN, with MIC levels similar to those
reported in this study. SDS-PAGE analysis indicated that the major outer membrane
lipoprotein I was missing in both clinical isolates (Campo-Esquisabel et al., 2007). SDS-
PAGE analysis of the serial subculture isolates would be useful in order to determine the
reason for sensitivity to PAβN. The specific mechanism of action of PAβN is not known,
though it is also a substrate of MexAB-OprM, MexCD-OprJ and MexEF-OprN (Warren et
al., 2000).
5.3.3 Fatty acid analysis
The alteration of membrane fatty acid profiles is an adaptive mechanism of bacterial cells
to toxic compounds. The isomerization of cis to trans fatty acids has been suggested as a
biomarker for monitoring stress in P. aeruginosa (Heipieper et al., 2003). The conversion
of unsaturated cytoplasmic membrane fatty acids from the cis to trans form by the cis-trans
isomerase allows cells to adapt to solvent stress (Heipieper et al., 2003).
In this work, no significant differences were detected with respect to short chain fatty acids,
long chain fatty acids, cyclic fatty acids, or the ratio of short:long fatty acids. Similarly, no
significant differences were detected for saturated fatty acids, unsaturated fatty acids and
the ratio of saturated:unsaturated fatty acids. Di Pasqua et al. (2006) reported that
P. fluorescens adapted to the terpenes thymol, carvacrol, limonene, cinnamaldehyde and

Chapter Five –Induced resistance to TTO
100
eugenol did not show substantial changes in fatty acid composition. Interestingly, B. cereus
cells adapted to carvacrol had decreased membrane fluidity mediated by changes in fatty
acid composition The relative amount of iso-C13:0, C14:0 and iso-C15:0 increased while
cis-C16:1 and C18:0 decreased (Ultee et al., 2000). Alterations in fatty acids do not appear
to contribute to the tolerance to TTO and terpinen-4-ol seen in this project.
5.3.4 Outer membrane permeability
A whole-cell alkaline phosphatase assay was used to evaluate OMP. In this assay the
substrate pNPP must diffuse though the outer membrane to reach the periplasmic enzyme
alkaline phosphatase (Martinez et al., 1996). The index of OMP values derived in this assay
were not reproducible, as evidenced by the large standard deviations observed and so no
solid conclusions could be made about any change in the outer membrane permeability of
these isolates.
The induction of resistance of P. aeruginosa PAO1 to several isothiazolone biocides, by
serial subculture in broth and increasing concentrations of biocide, was demonstrated by a
gradual increase in biocide MIC over 11 passages (Winder et al., 2000). The increase in
MIC was accompanied by the loss of T-OMP, an outer membrane protein, which re-
appeared following passage in the absence of biocide. Hydrophilic molecules are able to
diffuse into bacterial cells through outer membrane proteins or porins and so the loss or
modification of porins can be involved in antimicrobial resistance (Pages, 2006).
Decreasing the expression of OprD in P. aeruginosa results in resistance to carbapenems
(Kohler et al., 1999). The ability to survive in high concentrations of terpinen-4-ol, one of
the most water soluble components in TTO, may have been mediated by alteration of
porins in the membrane, though this does not explain why the MIC for PAO1T4ol
was still at
the parental strain level. Investigation of the outer membrane proteins of these isolates may
shed light on whether the structure of the outer membrane is involved.

Chapter Five –Induced resistance to TTO
101
5.3.5 Hydrophobicity studies
Decreases in cell surface hydrophobicity are often reported in cells adapted to organic
solvents (Isken and de Bont, 1998; Weber and de Bont, 1996).therefore it was suspected
that adaptation to TTO may have a similar effect. Studies on cell surface hydrophobicity of
the tolerised isolates were preliminary, but the difficulty in obtaining results is an indication
of the problems faced in studying hydrophobicity in bacterial cells. Experiments using the
commonly used methods of microbial adhesion to hydrocarbon (MATH) assay (Aono and
Kobayashi, 1997; Kobayashi et al., 1999; Rosenberg et al., 1980) and adhesion to
polystyrene beads (Aono and Kobayashi, 1997; Makin and Beveridge, 1996) were
attempted. Despite extensive attempts to optimize these methods, neither proved
reproducible within a realistic timeframe and so both methods were discontinued. The use
of flow cytometry to measure relative cell surface hydrophobicity was also not successful.
This is a more novel method that has been used previously with yeast (Colling et al., 2005)
but not bacteria.
Hydrophobic interaction chromatography is another method that can be used to determine
cell surface hydrophobicity, however due to budget and time constraints this was not
attempted.
5.3.6 Prolonged use of TTO and antimicrobial resistance
This work investigated the likelihood of increased resistance of P. aeruginosa to TTO
occurring following prolonged exposure to an increasing concentration of TTO. The
highest concentration of TTO that both strains could be subcultured in was relatively low
and in the case of mutS was below the MIC. Low level resistance and reduced susceptibility
to β-lactams, nalidixic acid, chloramphenicol and gentamicin observed in PAO1TTO
was
probably linked to expression of Mex efflux systems. A large number and variety of
substrates exists for these efflux pumps (see Chapter 3). The possibility exists that long
term exposure to TTO may induce or increase pump expression in P. aeruginosa and
consequently cause an increase in resistance to other antimicrobials that are substrates of

Chapter Five –Induced resistance to TTO
102
such pumps. Provided a TTO product contained a high concentration of TTO, at least
10% v/v, therapeutic use over a long period of time would probably not contribute to any
increase in resistance to other antimicrobial agents.
5.3.7 Summary
The PAO1TTO
isolate showed a decrease in susceptibility to β-lactams (aztreonam,
moxalactam, ceftriaxone, ticarcillin and Timentin), nalidixic acid, chloramphenicol and
gentamicin, though only aztreonam, moxalactam and ceftriaxone were at clinically
significant levels. Resistance to all but gentamicin can be mediated by the MexAB-OprM
efflux system, while the MexXY-OprM efflux system can generate resistance to
chloramphenicol, nalidixic acid, ticarcillin and Timentin. The most likely mechanism of
this acquired resistance is an increase in expression of the MexAB-OprM efflux system,
coupled with MexXY-OprM. The PAO1T4ol
isolate was more susceptible to moxalactam,
nalidixic acid, imipenem, ticarcillin and Timentin. An increase in susceptibility to all these
agents is associated with hyper-expression of the MexCD-OprJ efflux system, as seen in
nfxB mutants. Therefore, this increase in susceptibility may be mediated by the MexCD-
OprJ system. Investigation of the efflux phenotypes of the serial subculture isolates would
be useful for identification of any change in efflux system expression.
The minimal changes in disinfectant susceptibility detected in the tolerised isolates
indicated that exposure to TTO and terpinen-4-ol did not result in bacterial adaptation(s)
that conferred any advantage with respect to surviving the antibacterial activities of
benzalkonium chloride, cetrimide and chlorhexidine.
It would be useful to determine any fitness costs associated with the TTO-selected and
terpinen-4-ol-selected mutants.
The difference in phenotype seen in the serial subculture isolates indicates that the
adaptation of each strain, PAO1 and mutS, to TTO and terpinen-4-ol, was unique. The
mechanism(s) involved were probably different for each species and each agent however,

Chapter Five –Induced resistance to TTO
103
expression of the Mex efflux systems remains a likely candidate. Changes in the
susceptibility to TTO and terpinen-4-ol in PAO1TTO
appear to be phenotypic, as the
increased tolerance to TTO and T4ol was lost or decreased following subculture in the
absence of TTO. For all other serial subculture isolates, the increased tolerance to TTO and
terpinen-4-ol was maintained in the absence of the TTO or terpinen-4-ol, therefore the
changes were probably genotypic.

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
104
6.0 CHAPTER SIX – A LIBRARY OF TRANSPOSON MUTANTS
WITH ALTERED TTO AND TERPINEN-4-OL
SUSCEPTIBILITY
6.1 Introduction
The previous chapters have investigated putative mechanisms by which P. aeruginosa
gains resistance to the antimicrobial effects of TTO and components. These mechanisms,
namely efflux pumps and the outer membrane barrier function, were chosen for study based
on current understanding of antibiotic and disinfectant resistance mechanisms in
P. aeruginosa. However, it is possible that there are other unidentified mechanisms by
which P. aeruginosa can tolerate the oil and so a genetics-based study was undertaken.
The inactivation of genes and examination of resulting phenotypes is a long established
genetic tool. While targeted mutagenesis can give rise to a greater understanding of the
functions of specific genes, this can be a tedious process. Another method of mutagenesis is
based on the random insertion of pre-formed transposase-transposon complexes, called
„Transposomes‟, into bacterial cells, the subsequent examination of any altered phenotypes
and the sequence-based identification of insertion sites in mutants of interest. The
EZ::TN™ <KAN-2> Tnp Transposome™ kit has been used to generate libraries of
insertion mutants in Gram negative bacteria (Ducey and Dyer, 2002; Wyborn et al., 2004).
The Transposome is a stable complex formed between the hyperactive Tn5 transposase and
a Tn5-derived transposon and contains a kanamycin resistance cassette to enable rapid
selection of transposed cells (Hoffman and Jendrisak, 1999). Direct genomic sequencing
using primers supplied in this kit has been used to identify the insertion site within the
bacterial genomes of Salmonella typhimurium, Proteus vulgaris and Pseudomonas sp.
transposition clones (Hoffman et al., 2000).
Initially, the reference strain P. aeruginosa NCTC 10662 (10662) was selected as the
recipient strain in the mutagenesis experiments, however, following difficulties establishing

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
105
a large library of mutants based on this recipient, P. aeruginosa PAO1 (PAO1) was
substituted.
The objective of this chapter was to create a library of transposon mutants and determine
whether a change in susceptibility to TTO and/or terpinen-4-ol had occurred in any
mutants. Mutants of interest were then further studied and the site of insertion was
identified by sequencing.
6.2 Results
6.2.1 Testing for spontaneous resistance to kanamycin
Preliminary work showed a high level of spontaneous resistance in 10662 to kanamycin in
Luria-Bertani agar (results not shown); therefore the degree of spontaneous resistance to
kanamycin in 10662 was determined for 300, 400 and 500 mg/L of kanamycin in LA
(Table 6.1). The only concentration at which no growth was seen in duplicate experiments
after 24 h incubation was 500 mg/L. A few colonies were present after 48 h. Thus it was
determined that 500 mg/L would be used for all mutagenesis experiments and that
presumed transposon mutants would only be taken from 24 h incubation plates. Similar
testing for PAO1 was not done, however, all transposon mutants were confirmed by the
PCR outlined below.
6.2.2 Transposon mutagenesis and optimization
Initial transposon mutagenesis experiments yielded only one or two mutants and extensive
optimization of the method was required. Two different methods of cell preparation were
used (see 2.2.4.1). The second method (Choi, 2005) was published during the course of this
project and, following modification, yielded the most mutants. It was therefore used to
produce most of the mutants in this study.

Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
Table 6.1 Spontaneous resistance to kanamycin in P. aeruginosa NCTC 10662 on
Luria agar1
Kanamycin
concentration
(µg/ml)
Viable count (cfu/ml) after 24 h Viable count (cfu/ml) after 48 h
Experiment 12
Experiment 23
Experiment 1 Experiment 2
300 1 × 103
3 × 103 3 × 10
3 3 × 10
3
400 0 0.9 × 102 5.1 × 10
2 1.3 × 10
2
500 0 0 2 × 101
2.5 × 101
Legend
(1) Cells prepared as for transposon mutagenesis: overnight culture in Luria broth at
35°C, cells collected and washed three times in 300 mM sucrose then resuspended in
1 ml SDW. Forty microlitres of this suspension was added to 1 ml of LB broth. A
100 ul volume was spread-plated in triplicate onto pre-dried LA containing 300, 400
or 500 µg/ml of kanamycin, respectively. Plates were incubated at 37°C and
examined for growth after 24 h and 48 h. A viable count on BA was used to determine
the inoculum.
(2) Inoculum viable count = 2.98 × 108
cfu/ml
(3) Inoculum viable count = 1.33 × 109
cfu/ml

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
106
The electroporation conditions also required optimization, however, as the expensive
EZ::TN™ kit only contained enough Transposome™ for 10 reactions, it could not be used
as part of the optimization process. Due to the initial low efficiency of mutagenesis, it was
suspected that the Biorad Gene Pulser II was potentially malfunctioning and so a recipient
strain of E. coli (ED8654) was electroporated with pBBR1MCS and also pUC8. This
showed that the electroporator was working, as transformants were produced on the
ampicillin (pUC8, 3.22 × 10-6
cfu/recipient) and chloramphenicol (pBBR1MCS, 6.45 × 10-7
cfu/recipient) plates, respectively. The plasmid pBBR1MCS was electroporated into PAO1
to determine the optimal conditions for electroporation, including voltage, resistance,
length of expression time and cell concentration (Table 6.2). The conditions chosen were
2.5 kV, 1011
cfu/ml initial cell concentration, 200 ohms and 2 h expression time.
A total of 548 putative transposon mutants was generated from multiple experiments. As
mentioned, the use of two different parental strains resulted in the final mutant library. Fifty
three putative mutants were derived from 10662 (given the prefix „PA‟) and 495 were
derived from PAO1 (given the prefix „P‟).
6.2.3 PCR for detection of KanR cassette
In optimizing the PCR designed to detect a 401bp region of the kanamycin resistance
cassette located within the EZ-Tn5™ <KAN-2> transposon, a product of the expected size
was achieved at all three annealing temperatures tested (50, 55 and 60°C), with and without
co-solvents (DMSO and glycerol). The best result was seen at 60°C so this was used as the
annealing temperature and co-solvents were not included for ease of mix making.
Screening of the transposon mutant library confirmed the presence of this cassette in 51 of
the 53 10662 mutants and 486 of the 495 PAO1 mutants (Table 6.3, Figure 6.1). This
represented ~98% of the original library. The 11 PCR negative mutants were not studied
further, leaving a total library of 537 mutants.

Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
Table 6.2 Optimization of electroporation conditions using P. aeruginosa PAO1 and
pBBR1MCS1.
Voltage
(kV)
Initial cell
concentration
(cfu/ml)
Resistance
(ohms)
Number of transformants (cfu/ml)
1 h expression
time
2 h expression
time
1.8 1010
200 1 × 102 4.5 × 10
2
1.8 1011
200 4.45 × 103 1.11 × 10
4
2.0 1010
200 4.5 × 102 1.5 × 10
3
2.0 1011
200 7.95 × 103 1.24 × 10
4
2.5 1010
200 0 7.5 × 102
2.5 1011
200 9.4 × 103 1.66 × 10
4
2.5 1010
100 1.5 × 102 1 × 10
2
2.5 1010
300 2 × 102 1.05 × 10
3
Legend
(1) A negative control with no plasmid was tested for each combination in the table;
all yielded no transformants. Outlined row indicates best optimized conditions.

Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
Table 6.3 Transposon mutagenesis experiments that yielded PCR-confirmed
transposon mutants
Recipient Method Number of
confirmed mutants
Mutants per ng
transposon
10662 11 1 0.17
10662 1 46 7.67
10662 1 1 0.05
10662 1 1 0.1
10662 1 2 0.66
PAO1 1 23 1.15
PAO1 22 12 0.6
PAO1 2 67 3.35
PAO1 2 102 5.1
PAO1 2 93 4.65
PAO1 2 189 9.45
Legend
(1) Cell preparation method 1 (or variation of)
(2) Cell preparation method 2 (or variation of)
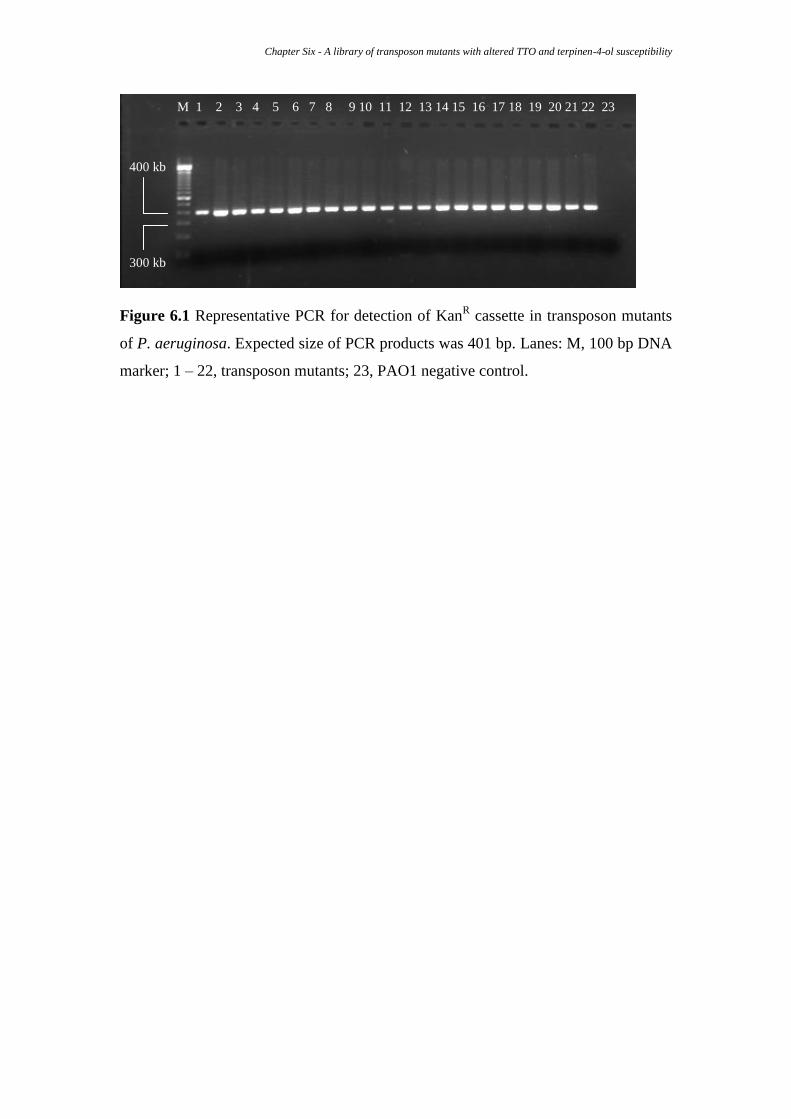
Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
Figure 6.1 Representative PCR for detection of KanR cassette in transposon mutants
of P. aeruginosa. Expected size of PCR products was 401 bp. Lanes: M, 100 bp DNA
marker; 1 – 22, transposon mutants; 23, PAO1 negative control.
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
300 kb
400 kb

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
107
6.2.4 Control experiment on effect of -80°C storage
It was necessary to determine whether the process of storing mutants at -80°C caused any
loss of the kanamycin resistance cassette. Eight mutants were selected at random from the
library and revived from -80°C storage. All eight mutants were positive for the presence of
the kanamycin resistance cassette in the PCR (Figure 6.2).
6.2.5 TTO and terpinen-4-ol susceptibilities of mutants
All transposon mutants in the library were screened for susceptibility to TTO and terpinen-
4-ol. Eight of the 10662 mutants and 12 of the PAO1 mutants had altered MICs for TTO
and/or terpinen-4-ol (Table 6.4). These 20 mutants were considered for further study. The
TTO MICs for the 10662-derived mutants ranged from 0.06-0.5%, while for 10662 the
MICs were 2-4%. Similarly for terpinen-4-ol, the mutant MICs ranged from 0.25-0.5%
while for 10662 the MIC was 2%. For the PAO1-derived mutants, the TTO MICs ranged
from 0.5-4% (PAO1 was 4%) and for terpinen-4-ol the MICs ranged from ≤0.03-1%
(PAO1 was 4%). Three of the PAO1-derived mutants had a TTO MIC equal to or half that
of PAO1, but exhibited an 8-fold reduction in terpinen-4-ol MIC. All other mutants showed
at least a 4-fold reduction in MIC to both TTO and terpinen-4-ol.
To confirm that the changes in susceptibility seen with the above mutants were not an
artifact of the experimental process, a control experiment was performed. Cells of 10662
were prepared and electroporated as in transposon mutagenesis, however, no
Transposome™ was included and the cell suspension was plated on LA. Following storage
of 100 randomly selected isolates at -80°C, the MICs of TTO and terpinen-4-ol for these
isolates were determined (Table 6.5). The majority of MICs were equal to, or one doubling
dilution either side of, the MIC of 10662. Several isolates had a TTO MIC that was higher
than this range. In those isolates the MIC was >8%, which was the limit of detection of the
assay. No isolate had an MIC less than half that of the parent.

Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
Figure 6.2 Detection of kanamycin resistance cassette in transposon mutants of
P. aeruginosa following storage at -80°C. Expected size of PCR products was 401 bp.
Lanes: M, 100 bp DNA marker; 1, PA140; 2, PA151; 3, PA160; 4, PA165; 5, PA172;
6, PA182; 7, PA190; 8, PA201; 9, PA100.
M 1 2 3 4 5 6 7 8 9
300 kb
400 kb

Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
Table 6.4 MICs of TTO and terpinen-4-ol (% v/v) for transposon mutants of
P. aeruginosa with altered susceptibility compared to parental strains.
Strain TTO MIC Terpinen-4-ol MIC
Parent, 10662 2 - 4 2
PA140 0.25 - 0.5 0.5
PA151 0.25 0.5
PA160 0.25 0.5
PA165 0.25 - 0.5 0.5
PA172 0.25 - 0.5 0.5
PA182 0.06 - 0.5 0.25 - 0.5
PA190 0.25 - 0.5 0.25 - 0.5
PA201 0.125- 0.5 0.25
Parent, PAO1 4 4
P114 1 - 2 0.5
P151 1 0.25
P253 2 0.5
P331 1 0.5
P349 1 0.25 - 0.5
P546 4 0.5
P565 2 0.5
P716 0.5 1
P744 1 0.5
P770 1 0.5
P808 1 ≤0.03
P898 0.5 - 2 0.5 - 1

Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
Table 6.5 Susceptibilities of control electroporation isolates of P. aeruginosa NCTC
10662 to TTO and terpinen-4-ol (%, v/v)
Number of isolates TTO MIC Terpinen-4-ol MIC
Parent, 10662 4 2
2 4 1
33 4 2
15 4 4
7 8 2
34 8 4
1 >8 2
7 >8 4
1 >8 8
Legend
Cell suspension of 10662 was prepared, electroporated, and incubated overnight as
per transposon mutagenesis, with the modification that no Transposome™ was
present during electroporation. One hundred colonies were selected at random and
plated out for single colonies on Luria agar. These isolates were then stored in
BHIB/20% glycerol at -80°C. Following revival from glycerol stocks, MICs for TTO
and terpinen-4-ol for the isolates were determined by broth microdilution.

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
108
6.2.6 Sequencing and identification of disrupted genes
Three different methods were used to try sequencing the insertion site in the transposon
mutants. The method suggested by EPICENTRE, the manufacturer of the transposon
mutagenesis kit, was a direct genomic sequencing method. Despite numerous attempts, this
method yielded no sequence. The second method combined restriction digest with EcoR1
and self ligation, with PCR and sequencing reactions that both used the KAN-2 FP-1 and
KAN-2 RP-1 primers. This method also yielded no sequence. The third and final method
used was RATE, the random amplification of transposon ends, based on a three-step single-
primer PCR protocol with either Inv-1 or Inv-2 (Figure 2.3) (Ducey and Dyer, 2002).
The first and third steps of the PCR were performed at a stringent annealing temperature
while the second step was performed at 30°C. This allowed non-specific amplification of
the single-stranded product generated in the first step, while the third step amplified both
specific and non-specific products. The published method did not specify the
concentrations of MgCl2, dNTPs or enzyme used. Although the primers used in this study
were the same as those used by Ducey (2002), the 50°C stringent annealing temperature did
not work for all mutants. Therefore, a lot of optimization was required before the final
method was selected. This final method involved the use of combinations of different
MgCl2 concentrations and different stringent annealing temperatures for each of the two
primers.
The RATE reaction yielded a product or multiple products for each of the 20 mutants in at
least one of the primer/MgCl2/annealing temperature combinations attempted. Figure 6.3
shows the optimization process for two mutants, P114 and P349. Because there was no
specific combination of conditions that gave the best results, each combination was used for
each mutant to increase the likelihood of a specific RATE product.
Despite numerous attempts, only nine mutants were sequenced both upstream and
downstream of the insertion. A further five mutants were sequenced on one side of the
insertion. The remaining six mutants were not able to be sequenced. The results are

Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
50°C 55°C 60°C
1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
(a)
50°C 55°C 60°C
1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
(b)
Figure 6.3 Optimisation of RATE products for transposon mutants P114 (a) and P349
(b). Lanes: 1, Inv-1 and 2.5 mM MgCl2; 2, Inv-1 and 1.5 mM MgCl2; 3, Inv-1 and
2 mM MgCl2; 4, Inv-2 and 2 mM MgCl2; 5, Inv-2 and 1.5 mM MgCl2; 6, Inv-2 and
2.5 mM MgCl2. The red box indicates the RATE products that were able to be
sequenced.

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
109
summarised in Table 6.6 and the locations of the insertions on the PAO1 chromosome are
shown in Figure 6.4. Mutants P114, P151 and P253 have an insertion at different locations
within surA. P331 has an insertion in fleN and P349 has an insertion in flgB. P546 has an
insertion in the intergenic space between the nusG and rplK. P565 has an insertion in the
intergenic space between tyrZ and PA0668.1. P716 has an insertion in the gene designated
PA0084. P744 has an insertion in fliC gene, P770 has an insertion in the gene designated
PA3800 and P808 has an insertion in rpoN. PA140 has an insertion in hitA, PA165 has an
insertion in cheZ and PA172 has an insertion in the gene designated PA0372.
6.3 Discussion
6.3.1 Transposon mutagenesis
Extensive modification and optimization of two different methods (Choi, 2005; Smith,
1989) for transposon mutagenesis was required. The final method used was the modified
version of Choi et al. (2005). Problems were encountered with the first method used when
the system unexpectedly stopped generating mutants. Despite a large amount of re-
optimization and checking of reagents and equipment used, this method did not produce
any more mutants. Hence the second method was identified in the literature and adapted for
this project.
Additional problems were encountered when Luria-Bertani agar was inadvertently
substituted for LA in the kanamycin plates. P. aeruginosa has a higher level of resistance to
kanamycin in Luria-Bertani agar plates and therefore the experiments using this agar had a
lawn of growth on the outgrowth plates. The mistake was eventually identified and when
replaced with LA the lawn of outgrowth was not seen again.
This optimization process was time consuming and also required two EZ::TN™ <KAN-2>
Tnp Transposome™ kits in total, making it quite expensive. Although the kit literature
claimed to yield in the order of 102 mutants per reaction for Pseudomonas, in most
experiments this was not the case. This low yield combined with mutagenesis method

Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
Table 6.6 Insertion sites of transposon mutants of P. aeruginosa
Mutant Primer Sequence adjacent to
insertion (nt) Gene
Location of
disrupted gene (nt)
P114 RP1 652890-653410
surA 652480-653772 FP1 NI
1
P151 RP1 652560-653215
surA 652480-653772 FP1 NI
P253 RP1 653089-653730
surA 652480-653772 FP1 NI
P331 RP1 1583489-1584129
fleN 1583956-1584798 FP1 1584121-1584785
P349 RP1 1163906-1164527
flgB 1164275-1164682 FP1 1164519-1165986
P546 RP1 4783194-4783866 nusG 4783228-4783761
2
FP1 4782716-4783197 rplK 4782680-47831112
P565 RP1 721288-721941 tyrZ 720357-721556
3
FP1 721933-722467 PA0668.1 722096 - 7236313
P716 RP1 101628-102283
PA0084 101778-103274 FP1 102288-102687
P744 RP1 1184010-1184102
fliC 1183058-1184524 FP1 1184094-1184573
P770 RP1 4259491-4260147
PA3800 4259255-4260397 FP1 NI
P808 RP1 4993114-4993664
rpoN 4992870-4994363 FP1 4993659-4994309
PA140 RP1 5257963-5258643
hitA 5257696-5258703 FP1 5258642-5258990
PA165 RP1 1585628-1586321
cheZ 1586034-1586822 FP1 1586313-1586794
PA172 RP1 416637-416964
PA0372 416009-417406 FP1 NI
Legend
(1) NI = Not identified. The following mutants were unable to have either side of the
insertion site identified: P898, PA151, PA160, PA182, PA190 and PA201. (2) and
(3)
insertion site is located between these two genes. nt = nucleotide.
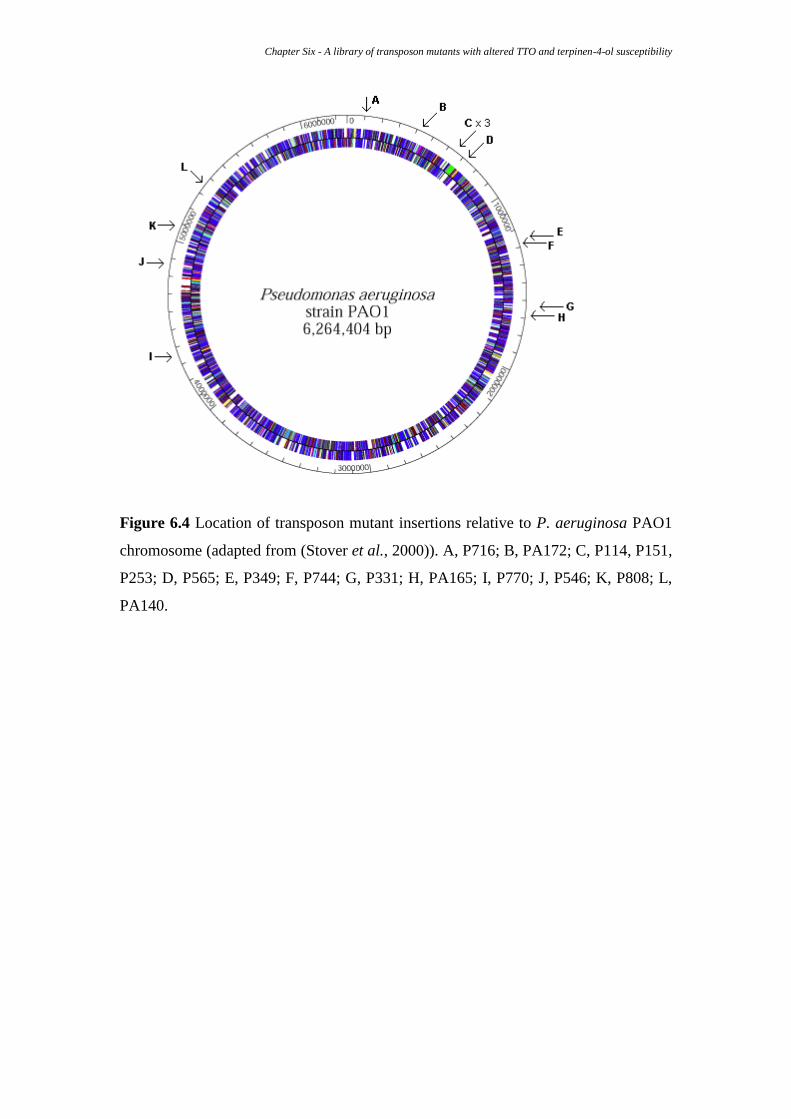
Chapter Six - A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
Figure 6.4 Location of transposon mutant insertions relative to P. aeruginosa PAO1
chromosome (adapted from (Stover et al., 2000)). A, P716; B, PA172; C, P114, P151,
P253; D, P565; E, P349; F, P744; G, P331; H, PA165; I, P770; J, P546; K, P808; L,
PA140.

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
110
problems meant that the entire series of mutagenesis experiments, which had to be
terminated due to time and funding constraints, resulted in a relatively modest library of
537 putative mutants. A near-saturation library of transposon mutants in P. aeruginosa
PAO1 consisted of 30,100 mutants, corresponding to an average of five insertions per gene
(Jacobs et al., 2003). While such a large-scale study was beyond our scope, a library of
2,000 – 5,000 mutants would have improved our study by providing more complete
genome coverage (Cao, 2004).
6.3.2 Susceptibility to TTO and terpinen-4-ol
Although standard susceptibility testing breakpoints are established for a large array of
bacteria and antibiotics in relation to clinical use (CLSI, 2006), the same can not be said for
other antimicrobials such as biocides, disinfectants and essential oils. Therefore, in this
study, a change in susceptibility at least 4-fold higher or lower (i.e. two doubling dilutions)
than the parental strain was considered to be of interest. Twenty mutants with an increased
susceptibility to TTO and/or terpinen-4-ol were identified from the library. This represented
8 out of the 51 10662-derived mutants (15.6%) and 12 out of the 486 PAO1-derived
mutants (2.5%). The TTO MIC of the mutants was decreased from 2-fold to 64-fold
compared to the MIC for parental strains. Similarly for terpinen-4-ol, the MIC for the
mutants was decreased by 4-fold to greater than 128-fold, compared to parental MICs.
P253, P546 and P565 had a TTO MIC the same as or 2-fold less than the parental value,
however, the terpinen-4-ol MIC for these isolates were reduced 8-fold and so they were
selected for further study.
6.3.3 Sequencing and insertion site identification
The EZ::TN™ <KAN-2> Tnp Transposome™ kit states that the primers supplied in the kit
can be used for direct sequencing from genomic DNA (Hoffman et al., 2000). Therefore
initial attempts to sequence the insertion site were based around a direct sequencing method
that involved sequencing directly from 2-3 µg of bacterial DNA, using the primers supplied
in the kit. Despite numerous attempts, this was not successful. An alternative method

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
111
(Rawat et al., 2003) was attempted that included DNA extraction, restriction enzyme
digestion, overnight self-ligation, PCR of the ligation mixture and sequencing using the
primers supplied in the kit; this was also unsuccessful. Finally, another method based on
the random amplification of transposon ends via a three-step single-primer PCR (Ducey
and Dyer, 2002) was used.
A problem with this method was that the third stage of PCR amplifies both the specific and
non-specific products from the second round, yet there was no way to determine which
products were which, until after sequencing was done. The non-specific products simply
did not yield a result when sequencing was attempted. This effectively meant that
approximately half of all PCR products generated using this method were useless.
Interestingly, the original published method used a stringent annealing temperature of
55°C, while in this project a PCR product was obtained at 50, 55 and 60°C and in various
combinations of MgCl2 concentration and either the Inv-1 or Inv-2 primer. There was not
one set of conditions that was considered optimal and therefore such a wide range of
conditions was used. Although the primers used in this project were the same as those used
by Ducey & Dyer (2002), the enzyme and thermocycler used by this group were not
mentioned and so this may account for the differences noted.
6.3.4 Disrupted genes and regions
The disrupted genes that were identified in this study are discussed below.
Complementation studies were not carried out in this work and so the relationship between
the identified genes and the change in susceptibility observed is discussed based on the
function of the gene product and the current published literature in the area. Future work
confirming the role of each disrupted gene in susceptibility to TTO and terpinen-4-ol, by
complementation, is required.

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
112
6.3.4.1 surA
In this study surA was identified as the disrupted gene in the transposon mutants P114,
P151 and P253. The three mutants were all slightly more susceptible to TTO (2- to 4-fold)
and substantially more susceptible to terpinen-4-ol (8- to 16-fold). These surA mutants
arose in separate mutagenesis experiments and therefore were unlikely to be sibling
mutants; this was confirmed by sequencing which showed that the location of the insertion
was different in all three mutants. SurA is a periplasmic chaperone and peptidyl-prolyl
isomerase involved in the correct folding of outer membrane proteins, specifically porins,
in Gram negative bacteria (Bitto and McKay, 2003; Hennecke et al., 2005). The gene surA
was first discovered in E. coli during a screening assay intended to identify genes required
for survival in stationary phase, although impaired survival only occurred under certain
conditions (Tormo et al., 1990). surA mutants of E. coli have phenotypes that are
characteristic of a defective cell envelope, including an increased susceptibility to
hydrophobic agents including SDS, EDTA, rifampicin and crystal violet (Behrens et al.,
2001). Such mutant cells also contain reduced amounts of outer membrane proteins
(Rouviere and Gross, 1996). Recently SurA has been described as the primary chaperone
involved in the periplasmic transfer of outer membrane proteins to the YaeT complex in
E. coli (Sklar et al., 2007). The increased susceptibility of the surA mutants in this study to
both TTO and terpinen-4-ol corresponds with the documented increased susceptibility to
other hydrophobic agents of other surA mutants (Behrens et al., 2001) and also correlates
with previous work showing increased TTO and terpinen-4-ol susceptibility following
compromising of the outer membrane of P. aeruginosa (Longbottom et al., 2004; Mann et
al., 2000).
Operon prediction software (Ermolaeva et al., 2001) revealed that surA is part of an operon
in P. aeruginosa that contains ostA, surA, pdxA, ksgA, an open reading frame (ORF) and
apaH (Figure 6.5). This operon occurs with the same gene order in the E. coli genome and
has been referred to as a superoperon, in which the ORF is apaG (Dartigalongue et al.,
2001; Pease et al., 2002). Located directly upstream of surA in the operon is ostA, a gene
involved in organic solvent tolerance in E. coli (Aono et al., 1994). Disruption of this

Figure 6.5 Location of disrupted gene (surA) of transposon mutants P114, P151 and P253 within a predicted operon (red box) on the
chromosome of P. aeruginosa PAO1 (adapted from http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008). PA0587: conserved
hypothetical protein; PA0588: conserved hypothetical protein; PA0589: conserved hypothetical protein; apaH: bis(5'-nucleosyl)-
tetraphosphatase ApaH; PA0591: conserved hypothetical protein ApaG; pdxA: pyridoxal phosphate biosynthetic protein PdxA; surA: peptidyl-
prolyl cis-trans isomerase SurA; ostA: organic solvent tolerance protein OstA precursor; PA0596: hypothetical protein; PA0597: putative
nucleotidyl transferase; PA0598: hypothetical protein; PA0599: hypothetical protein (Winsor et al., 2005). nt = nucleotide.
PA0591 PA0587 PA0589
PA0598 PA0597
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
113
protein in Helicobacter pylori resulted in altered membrane permeability, sensitivity to
n-hexane and susceptibility to hydrophobic and β-lactam antibiotics (Chiu et al., 2007).
Downstream of surA on the chromosome is pdxA. PdxA is a pyridoxal phosphate
biosynthetic (vitamin B6) protein involved in the biosynthesis of cofactors, prosthetic
groups and carriers (Winsor et al., 2005). surA and pdxA are co-transcribed and insertional
mutations in surA in E. coli resulted in a PdxA- phenotype (Pease et al., 2002; Tormo et al.,
1990). The next gene in this operon is ksgA, encoding an rRNA modification enzyme (16S
rRNA methyltransferase) and co-transcribed with pdxA in E. coli (Pease et al., 2002). The
next gene downstream of ksgA is the ORF PA0591, encoding a conserved hypothetical
protein in P. aeruginosa and also known as apaG (Winsor et al., 2005). This gene has 72%
nucleotide homology with apaG in E. coli and encodes an uncharacterized protein affecting
Mg2+
/Co2+
(Winsor et al., 2005). The final gene in the operon is apaH which has 64%
homology with apaH in E. coli and codes for a diadenosine tetraphosphatase (Winsor et al.,
2005). The function of this operon is unknown, however, it is possible that an insertion in
surA, the second gene in the operon, does have a polar effect and affects transcription of
pdxA and/or other downstream genes, though it is not known how this would contribute to a
change in susceptibility to TTO and terpinen-4-ol.
6.3.4.2 Flagellar biogenesis genes
The single polar flagellum of P. aeruginosa is well established as an important factor for
virulence and colonization (Arora et al., 1998; Montie et al., 1982). Fifty genes assigned to
17 putative operons in the P. aeruginosa PAO1 genome are known or predicted to be
involved in the assembly or regulation of the flagellar organelle, or are linked chemotaxis
genes (Dasgupta et al., 2003). The flagellar genes of P. aeruginosa are clustered in three
non-contiguous regions of the chromosome, regions I, II and III. The fla regulon is
comprised of dedicated flagellar genes encoding proteins that participate in the regulation
of the flagellar transcriptional circuit, including fleQ, fleS, fleR, fliA, flgM and fleN
(Dasgupta et al., 2003). The insertion site in four of the transposon mutants generated in
this study, with increased susceptibility to TTO and/or terpinen-4-ol, was a flagellar-related
gene. In another of the transposon mutants, the disrupted gene was identified as rpoN. The

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
114
alternative sigma factor σ54
, encoded by rpoN, is responsible for regulating the expression
of flagellum (see 6.3.4.3) (Dasgupta et al., 2003).
6.3.4.2.1 fleN
The insertion site in the transposon mutant P331 was identified as fleN. The location of fleN
on the chromosome of PAO1 is shown in Figure 6.6. When compared with its parental
strain, this mutant was 2- to 4-fold more susceptible to TTO and 8-fold more susceptible to
terpinen-4-ol. fleN is involved in controlling the number of flagella and chemotactic ability
in P. aeruginosa. Loss of fleN causes an increase in the number of polar flagella (Dasgupta
et al., 2000). The transcription of flagellar genes in P. aeruginosa is a highly regulated
process, the highest-level positive regulator of which is FleQ (Dasgupta et al., 2003). The
activity of FleQ is posttranslationally inhibited by direct interaction with FleN (Dasgupta
and Ramphal, 2001). It has been suggested that fleN is located in an operon with flhF, co-
transcribed from a promoter upstream of flhF (Dasgupta and Ramphal, 2001). flhF
determines the polar placement site of new flagella in P. putida (Pandza et al., 2000). No
studies on the susceptibility of fleN mutants to antimicrobial agents could be found in
published literature. Any effects downstream following disruption of fleN are unknown.
6.3.4.2.2 fliC
The fliC mutant identified in this study, P744, was more susceptible to both TTO (4-fold)
and terpinen-4-ol (8-fold) than the parental strain. The gene fliC encodes for two
structurally distinct flagellin proteins, designated type-a and type-b (Spangenberg et al.,
1996). The genome of P. aeruginosa PAO1 encodes type-b flagellin while clinical isolates
often express type-a flagellin (Arora et al., 2001).
Segura et al. (2001) demonstrated that fliP mutants of P. putida were more susceptible to
the organic solvent toluene (Segura et al., 2001). FliP is involved in the export of flagellar
proteins. The authors suggest a relationship between an intact flagellar export system and
the transport of efflux proteins that are required for toluene tolerance and are located in the

Figure 6.6 Location of disrupted gene of transposon mutants P331 (fleN) and PA165 (cheZ) on the chromosome of P. aeruginosa PAO1. The
red box indicates a predicted operon with flhF and fleN (Dasgupta and Ramphal, 2001), the green box indicates a predicted operon with flhF,
fliA, cheY and cheZ minus fleN (adapted from http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008). PA1450: conserved
hypothetical protein; PA1451: conserved hypothetical protein; flhA: flagellar biosynthesis protein FlhA; flhF: flagellar biosynthesis protein FlhF;
fleN: flagellar synthesis regulator FleN; fliA: sigma factor FliA; cheY: two-component response regulator CheY; cheZ: chemotaxis protein CheZ;
PA1458: putative two-component sensor; PA1459: putative chemotaxis signal transduction methyltransferase; motC: MotC, involved in flagella
assembly and chemotaxis (Winsor et al., 2005). nt = nucleotide.
cheY
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
an
d terp
inen
-4-o
l suscep
tibility

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
115
periplasmic space and/or the outer membrane (Segura et al., 2001). Similarly, mutants of P.
putida with insertions in the genes encoding flagellar structural proteins FliC, FlgK, FlaG,
FliI and FliH, as well as the transcriptional activator FleQ and the flagellum-specific RNA
polymerase sigma factor FliA, were unable to grow in the presence of toluene and octanol
(Kieboom et al., 2001). These flagella mutants were non-motile and had decreased
expression of the SrpABC organic solvent efflux pump (Kieboom et al., 2001). Given the
suggested role for flagellar proteins and tolerance to organic solvents in P. putida, it is
possible a similar relationship is responsible for the increased susceptibility to TTO and
terpinen-4-ol in the flagella mutants in this study.
Flagellum-deficient fliC mutants of P. aeruginosa are more susceptible to lysis by SDS and
complemented fliC mutants show restored resistance to that of the wild type (Zhang et al.,
2007). An intact flagellum was also required to confer resistance to surfactant protein A-
mediated membrane permeabilization in P. aeruginosa (Zhang et al., 2007). Outer
membrane defects are known to increase the sensitivity of Gram negative bacteria to
detergents like SDS (Vaara, 1992). The increased susceptibility to SDS and surfactant
protein-A, and reduced expression of LPS seen with the loss of flagellum in several P.
aeruginosa mutants, including fliC, indicates that flagella deficient mutants are likely to
have weakened outer membranes (Zhang et al., 2007). It is therefore not surprising that the
fliC mutant P774 was more susceptible to the antimicrobial actions of both TTO and
terpinen-4-ol in this study, considering the mechanism of action of these compounds
involves loss of membrane integrity and function (Carson et al., 2006). Given that SDS is a
substrate of the Mex efflux systems in P. aeruginosa (Srikumar et al., 1998) and the
finding that flagellar mutants of P. putida have altered efflux systems (Kieboom et al.,
2001), perhaps the increase in susceptibility to SDS in flagella mutants of P. aeruginosa is
associated with decreased expression of efflux systems.
The fliC gene is the first gene in transcribed in a predicted operon containing fliD and two
genes encoding hypothetical proteins, flaG and fliS (Figure 6.7) (Ermolaeva et al., 2001).
FliD encodes the flagellar cap protein and is involved in mucin adhesion (Arora et al.,
1998). The insertion in fliC may have polar effects on the transcription of the downstream

Figure 6.7 Location of disrupted gene (fliC) of transposon mutant P744 within a predicted operon (red box) on the chromosome of
P. aeruginosa PAO1 (adapted from http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008). PA1090: hypothetical protein; PA1091:
flagellar glycosyl transferase, FgtA; fliC: flagellin type B, FliC; flaG: hypothetical protein, FlaG; fliD: flagellar capping protein FliD; fliS:
hypothetical protein, FliS; PA1096: hypothetical protein; fleQ: transcriptional regulator FleQ; fleS: two-component sensor; fleR: two-component
response regulator (Winsor et al., 2005). nt = nucleotide.
fliS flaG PA1090 PA1096
fleR
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
116
genes in the operon, including fliD, however, what link if any this has with antimicrobial
susceptibility is not known. If a role exists for flagellar proteins and efflux in
P. aeruginosa, then this may affect susceptibility to a range of antimicrobials including
TTO.
6.3.4.2.3 flgB
The insertion site in the transposon mutant P349 is flgB. FlgB is a flagellar basal-body rod
protein and the first gene transcribed in the large flg operon, comprising flgB, flgC, flgD,
flgE, flgF, flgG, flgH, flgI, flgJ, flgK and flgL (Figure 6.8) (Ermolaeva et al., 2001).
Although no studies of the susceptibility of flgB mutants were identified in the literature, it
is possible that the lack of functional flagella caused by this mutation may affect
susceptibility to membrane active compounds, as seen in fliC mutants. Additionally, as flgB
is the first gene in the operon, this insertion may affect the transcription of the rest of the
operon. Again, effects on efflux proteins may be mediated by the interruption of the flagella
biosynthetic pathway, as discussed above.
6.3.4.2.4 cheZ
The insertion site in the transposon mutant PA165 was identified as cheZ. The location of
cheZ on the chromosome of PAO1 is shown in Figure 6.6. The CheZ phosphatase catalyses
the removal of the phosphoryl group from the signaling molecule, CheY, in E. coli
chemotaxis (Silversmith, 2005) Phosphorylation of CheY signals a switch in flagellar
rotation from counterclockwise to clockwise, switching the swimming phenotype from
smooth to tumbling and cheZ mutants of E. coli tumble excessively (Parkinson et al.,
1983). Mutants of cheZ in P. aeruginosa are fully motile but change direction more often
than their parent strain and form smaller swarms (Masduki et al., 1995). cheZ is the last
gene transcribed in a predicted operon that also contains flhF, fliA and cheY (Ermolaeva et
al., 2001). As the last gene in the operon, the disruption of cheZ is unlikely to affect
transcription of the other genes in the operon. This mutant was more susceptible to both
TTO (4- to 16-fold) and terpinen-4-ol (4-fold) than the parental strain, but the link between

Figure 6.8 Location of disrupted gene (flgB) of transposon mutant P349 within the flg operon (red box) on the chromosome of P. aeruginosa
PAO1 (adapted from http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008). braC: branched-chain amino acid transport protein
BraC; PA1075: hypothetical protein; PA1076: hypothetical protein; flgB: flagellar basal-body rod protein FlgB; flgC: flagellar basal-body rod
protein FlgC; flgD: flagellar basal-body rod modification protein FlgD; flgE: flagellar hook protein FlgE; flgG: flagellar basal-body rod protein
FlgG; flgH: flagellar L-ring protein precursor FlgH; flgI: flagellar P-ring protein precursor FlgI; flgJ: flagellar protein FlgJ; flgK: flagellar hook-
associated protein 1 FlgK; flgL: flagellar hook-associated protein type 3 FlgL; PA1088: hypothetical protein; PA1089: conserved hypothetical
protein (Winsor et al., 2005). nt = nucleotide.
flg C
flg B
PA1075
braC
PA1076
PA1088
PA1089
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
117
the two has not been determined and no data on the antimicrobial susceptibility of cheZ
mutants was identified in the literature.
6.3.4.3 rpoN
The insertion site in the transposon mutant P808 was identified as rpoN. This mutant was
more susceptible to TTO (4-fold) and substantially more susceptible to terpinen-4-ol (>128-
fold) than the parental strain, PAO1. RpoN is an alternate sigma factor, σ54
, that controls
transcription of pili and flagella in P. aeruginosa and so rpoN mutants do not synthesize
flagellin and are non-motile (Totten et al., 1990). As discussed in 6.3.4.2, a lack of
functional flagella can increase susceptibility to antimicrobial agents. An rpoN mutant of
P. putida was more susceptible to organic solvents, mediated by decreased expression of an
efflux system (Kieboom et al., 2001).
RpoN is also involved in the negative control of quorum sensing machinery and has been
implicated in the regulation of virulence in P. aeruginosa (Heurlier et al., 2003; Thompson
et al., 2003). Interestingly, stationary phase rpoN mutants of P. aeruginosa are less
susceptible to quinolones and carbapenems, thought to be mediated partly by increased
production of the siderophore pyoverdine and vqsR gene expression (Viducic et al., 2007).
There are genes downstream of rpoN in P. aeruginosa that, along with rpoN, appear to
comprise an operon (Figure 6.9) (Winsor et al., 2005). These genes are yhbH (encodes
conserved hypothetical protein), pstN (encodes nitrogen regulatory protein), yhbJ (encodes
conserved hypothetical protein) and PA4466 (encodes putative phosphoryl carrier protein)
(Winsor et al., 2005). The role of the rpoN operon has not been elucidated, but it is possible
that disruption of rpoN affects transcription of the other genes in the operon.
6.3.4.4 hitA
The insertion site in mutant PA140 is hitA. HitA is a ferric-iron III binding periplasmic
protein with 64% amino acid similarity to HitA in Haemophilus influenzae (Winsor et al.,

Figure 6.9 Location of disrupted gene (rpoN) of transposon mutant P808 within the rpoN operon (red box) on the chromosome of P. aeruginosa
PAO1 (adapted from http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008). PA4453: conserved hypothetical protein; PA4454:
conserved hypothetical protein; PA4455: putative permease of ABC transporter; PA4456: putative ATP-binding component of ABC transporter;
PA4457: conserved hypothetical protein; PA4458: conserved hypothetical protein; PA4459: conserved hypothetical protein; PA4460: conserved
hypothetical protein; PA4461: putative ATP-binding component of ABC transporter; rpoN: RNA polymerase factor sigma-54; yhbH: conserved
hypothetical protein; pstN: nitrogen regulatory IIA protein, PstN; yhbJ: conserved hypothetical protein; PA4466: putative phosphoryl carrier
protein; PA4467: hypothetical protein; PA4468: superoxide dismutase; PA4469: hypothetical protein; fumC1: fumarate hydratase, FumC;
PA4471: hypothetical protein (Winsor et al., 2005). nt = nucleotide.
yhbH
pstN
yhbJ
PA4466 PA4461
PA4460
PA4459
PA4458
PA4456
PA4455 PA4454
PA4453 PA4468 PA4469
PA4471
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
118
2005). The hitA gene is located on the PAO1 chromosome in an operon with hitB, an iron
transport system permease (Figure 6.10) (Ermolaeva et al., 2001). Bacterial iron transport is
required for the growth of pathogens in a host environment and therefore virulence
(Adhikari et al., 1995). Iron acquisition in P. aeruginosa can occur via the siderophores
pyochelin and pyoverdine; the genome of PAO1 has many homologues of iron-siderophore
receptor genes (Poole and McKay, 2003) and so it is unlikely that an insertion in one iron
transport operon would make a large impact on iron acquisition. Compared to the parental
strain, PA140 is 4- to 16-fold more susceptible to TTO and 4-fold more susceptible to
terpinen-4-ol. Increased susceptibility to antimicrobials has not been associated in the
literature with defects in iron transport.
6.3.4.5 PA0084
The insertion site in mutant P716 is identified as PA0084 (Figure 6.11). This gene codes for
a hypothetical conserved protein in P. aeruginosa, it is uncharacterized and its function is
unknown (Winsor et al., 2005). The link between this disrupted gene and the increased
susceptibility of P716 to TTO and terpinen-4-ol is not known.
6.3.4.6 PA3800
The insertion site in mutant P770 is identified as PA3800 (Figure 6.12). This gene codes for
a hypothetical conserved protein in P. aeruginosa with unknown function. The location of
this gene on the chromosome, between yfgM and yfgK, is the same in E. coli and it is part
of a 15 kb region that is well conserved between PAO1 and E. coli (Winsor et al., 2005).
The link between this disrupted gene and the increased susceptibility of P770 to TTO and
terpinen-4-ol is not known.
6.3.4.7 PA0372
The insertion site in mutant PA172 is identified as PA0372 (Figure 6.13). The product of
this gene is a probable zinc protease (Winsor et al., 2005). Other zinc proteases or zinc

Figure 6.10 Location of disrupted gene (hitA) of transposon mutant PA140 within the hitAB operon (red box) on the chromosome of
P. aeruginosa PAO1 (adapted from http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008). PA4682: hypothetical protein; PA4583:
hypothetical protein; PA4684: hypothetical protein; PA4685: hypothetical protein; PA4686: hypothetical protein; hitA: ferric iron-binding
periplasmic protein HitA; hitB: iron (III)-transport system permease HitB; PA4689: hypothetical protein; PA4690: hypothetical protein;
PA4690.1: 5S ribosomal RNA; PA4690.2: 23S ribosomal RNA (Winsor et al., 2005). nt = nucleotide.
PA4690.2 PA4683
PA4682
PA4685 PA4690.1
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Figure 6.11 Location of disrupted gene (PA0084) of transposon mutant P716 on the chromosome of P. aeruginosa PAO1 (adapted from
http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008).
PA0080 PA0083 PA0085 PA0087
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Figure 6.12 Location of disrupted gene (PA3800) of transposon mutant P770 on the chromosome of P. aeruginosa PAO1 (adapted from
http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008).
PA3796
PA3797
PA3794
PA3793 PA3801 PA3807
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Figure 6.13 Location of disrupted gene (PA0372) of transposon mutant PA172 on the chromosome of P. aeruginosa PAO1 (adapted from
http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008).
PA0367 PA0369
PA0370 PA0377
PA0379 PA0380
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
119
metalloproteases in P. aeruginosa include LasA and LasB, which degrade elastin and are
important for pathogenesis (Gustin et al., 1996). There is no documented link between zinc
proteases and antimicrobial susceptibility and the link between PA0372 and increased
susceptibility to TTO and terpinen-4-ol is not known.
6.3.4.8 Region between nusG and rplK
The insertion site in mutant P546 is the intergenic region between nusG, encoding the
transcription antitermination protein NusG and rplK, encoding the 50S ribosomal protein
L11. These two genes are predicted to be transcribed in an operon (Figure 6.14) (Ermolaeva
et al., 2001) and so the insertion may have polar effects on rplK. P546 was eight times
more susceptible to terpinen-4-ol than the parent strain PAO1, but susceptibility to TTO
was unchanged. There is no clear connection between this operon and the increased
susceptibility to terpinen-4-ol in this mutant.
6.3.4.9 Region between tyrZ and PA0668.1
The insertion site in mutant P565 is the intergenic region between tyrZ, encoding a tyrosyl-
tRNA synthetase 2, and PA0668.1, encoding a 16S ribosomal RNA (Figure 6.15). Neither
tyrZ nor PA0668.1 is predicted to be transcribed in an operon (Ermolaeva et al., 2001).
Similar to P546, P565 was 8-fold more susceptible to terpinen-4-ol than the parent strain
PAO1, while susceptibility to TTO was increased by a factor of two. Again, there is no
clear association between the susceptibility of this mutant and the insertion site.
6.3.5 Summary
A modest library of transposon mutants of P. aeruginosa yielded a total of 20 mutants that
were more susceptible to TTO and/or terpinen-4-ol. The insertion site was identified in 14
of these mutants, while for six mutants the insertion site was not able to be identified in the
course of this project. The disrupted gene in four mutants was related to flagellar
biosynthesis. Since flagellar mutants of P. putida are associated with increased

Figure 6.14 Transposon insertion site (black arrow) of mutant P546 between nusG and rplK, within a predicted operon (red box) in
P. aeruginosa PAO1 (adapted from http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008). rpoB: DNA-directed RNA polymerase
beta chain; PA4270.1: non-coding RNA gene; rplJ: 50S ribosomal protein L7 / L12; rplA: 50S ribosomal protein L1; rplK: 50S ribosomal
protein L11; nusG: transcription antitermination protein NusG; secE: pre-protein translocase subunit SecE; PA4276.1: tRNA; tufB: elongation
factor Tu; PA4277.1: tRNA; PA4277.2: tRNA; PA4277.3: tRNA; PA4278: hypothetical protein; PA4279: hypothetical protein; birA: BirA
bifunctional protein; PA4280.1: 5S ribosomal RNA; PA4280.2: 23S ribosomal RNA (Winsor et al., 2005). nt = nucleotide.
secE nusG rplK
PA4270.1 PA
4276
.1 P
A42
77.2
P
A42
77.3
PA
4277
.1
PA4278
PA4279
PA4280.1
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Figure 6.15 Transposon insertion site (black arrow) of mutant P565 between tyrZ and PA0668.1 within the genome of P. aeruginosa PAO1
(adapted from http://www.ncbi.nlm.nih.gov/sites/entrez, accessed January 2008). PA0660: hypothetical protein; PA0661: conserved hypothetical
protein; argC: N-acetyl-gamma-glutamyl-phosphate reductase; PA0663: hypothetical protein; PA0664: hypothetical protein; PA0665: conserved
hypothetical protein; anmK: anhydro-N-acetylmuramic acid kinase; PA0667: conserved hypothetical protein; tyrZ: tyrosyl-tRNA synthetase 2;
PA0668.1: 16S ribosomal RNA; PA0668.2: tRNA; PA0668.3: tRNA; PA0668.4: 23S ribosomal RNA; PA0668.5: 5S ribosomal RNA; PA0669:
putative DNA polymerase alpha chain (Winsor et al., 2005). nt = nucleotide.
PA0665 PA
0688
.3
PA
0688
.2
PA
0688
.5
PA0669
PA0664 PA0663
PA0661
PA0660
Chap
ter Six - A
libra
ry of tra
nsp
oso
n m
uta
nts w
ith a
ltered T
TO
and
terpin
en-4
-ol su
sceptib
ility

Chapter Six –A library of transposon mutants with altered TTO and terpinen-4-ol susceptibility
120
susceptibility to organic solvents through decreased efflux, it would be interesting to
determine the efflux phenotype of these transposon mutants. Three non-sibling surA
mutants were identified and it is possible that impaired outer membrane folding in these
mutants caused the increase in susceptibility to TTO and terpinen-4-ol.

Chapter Seven –General Discussion
121
7.0 CHAPTER SEVEN - GENERAL DISCUSSION
The antimicrobial properties of TTO and components have been studied since the 1920s,
when Penfold first compared the antiseptic properties of the oil with that of phenol and
implied it was 11-fold more effective (Penfold and Grant, 1925; Penfold, 1929). Eight
decades have passed and TTO is now widely accepted as a broad spectrum topical
antibacterial agent. Other properties of the oil, including antiviral, antifungal, anti-
inflammatory and anti-cancer properties have also been documented.
An exception in the wide spectrum of antibacterial activity is P. aeruginosa, an
opportunistic Gram negative pathogen that is resistant to a wide array of antibiotics and is
less susceptible than other Gram negative bacteria to TTO. P. aeruginosa is often
associated with nosocomial infections and colonizes wounds and burns. Given that TTO is
used topically, an understanding of the mechanisms of tolerance to TTO and its individual
components is essential, as the first step towards potential modification of TTO
preparations for use against P. aeruginosa.
As TTO is a complex mixture of over 100 components, with multiple mechanisms of
action, the use of TTO is unlikely to cause resistance to arise in the same manner as
conventional antimicrobials. However, one study with S. aureus reported resistance to TTO
following habituation, while another reported increases in MICs of several antibiotics
following exposure to sub-inhibitory levels TTO in S. aureus, Salmonella spp. and E. coli
(McMahon et al., 2007; Nelson, 2000). It is critical to address the issue of increased
resistance arising following treatment with TTO, especially any associated increase in
resistance to other antimicrobials and so this was an important part of this thesis.
The work reported in this thesis was an investigation of the mechanisms of tolerance of
P. aeruginosa to TTO and some of its components. This was broken down into four main
areas of investigation, which included the potential roles of efflux systems and of LPS, a
genetics-based attempt to identify unknown mechanisms of tolerance, as well as an
investigation into whether the level of resistance to TTO in P. aeruginosa would increase

Chapter Seven –General Discussion
122
upon repeated exposure to TTO. Both whole TTO as well as several components of the oil
were investigated, including terpinen-4-ol, to which the main antimicrobial activity of TTO
is attributed, and also ρ-cymene, 1,8-cineole, α-terpineol and γ-terpinene. Terpinen-4-ol and
α-terpineol were selected on the basis of their known antimicrobial activity and the fact that
they comprise 40% of TTO. 1,8-Cineole was selected since it has some antimicrobial
activity and its concentration in TTO has been manipulated partly in response to the
misconception that it is a skin irritant (Carson et al., 2006). γ-Terpinene and ρ-cymene were
selected since they constitute up to 30% of TTO.
Of the 5 Mex efflux systems investigated, the MexAB-OprM system provided resistance to
TTO as well as terpinen-4-ol, 1,8-cineole and α-terpineol. This was the first reporting of
terpenes as a substrate of this pump. A potential role exists for MexCD-OprJ in terpene
efflux, though further work is required to confirm this. Terpinen-4-ol was described as a
substrate for MexEF-OprN. TTO and the components tested in this work did not appear to
be substrates of the MexJK system. Due to the problems encountered with RT-PCR,
expression of MexXY could not be confirmed and therefore it could not be conclusively
determined whether TTO and the components tested were substrates of this efflux system.
Initially, γ-terpinene and ρ-cymene did not appear to be substrates of any of the pumps
examined, although it was suspected that their low antimicrobial activity may have meant
that changes to their MICs went undetected above the upper limit of the susceptibility test.
The use of the EPI PAβN showed that ρ-cymene is probably a substrate for several of the
Mex efflux pumps, including MexAB-OprM, though expression work is required to
confirm this. Alternatively, we may not have examined the pumps for which they are
substrates. Given that γ-terpinene has demonstrated activity against P. aeruginosa under
certain circumstances (Longbottom et al., 2004; Mann et al., 2000), the pumps examined in
this project may not have included pumps for which γ-terpinene is a substrate.
Resistance to terpene compounds has previously been associated with an efflux system in
Gram negative bacteria (Moken et al., 1997). Deletion of the acrAB locus in E. coli
increased susceptibility to pine oil, which is mainly comprised of α-terpineol (~65%)
(Moken et al., 1997). Resistance to pine oil in E. coli was associated with resistance to

Chapter Seven –General Discussion
123
multiple antibiotics including tetracycline, ampicillin, chloramphenicol and nalidixic acid.
Similarly, the PAO1TTO
serial subculture mutant created in this study was more resistant to
chloramphenicol and nalidixic acid, amongst other antibiotics, which was probably
mediated by one or more of the Mex efflux systems. Previous studies have shown an
additive effect on drug resistance from the simultaneous expression of Mex efflux systems
in P. aeruginosa (Lee et al., 2000). Although TTO, terpinen-4-ol, 1,8-cineole and α-
terpineol were confirmed as substrates of MexAB-OprM, it is important to determine
whether they are substrates for the other Mex efflux systems discussed above. A more
extensive library of efflux mutants is required for this, in conjunction with real time RT-
PCR.
Flagellar mutants of P. putida have altered expression of efflux systems and are susceptible
to organic solvents (Kieboom et al., 2001) and it has been suggested that an intact flagellar
export system is involved in the transfer of efflux pump components within the bacterial
membrane (Segura et al., 2001). In strains of P. aeruginosa lacking flagella, type III
secretion system gene expression was increased, while the overproduction of type III
secretion system transcriptional activator ExsA caused a decrease in expression of flagella,
indicating an inverse relationship (Soscia et al., 2007). Interestingly, it has been reported
that the overexpression of MexCD-OprJ or MexEF-OprN in P. aeruginosa impaired type
III secretion, due to a lack of expression of genes belonging to the type III secretion regulon
(Linares, 2005).
The disrupted gene in several of the TTO-susceptible transposon mutants generated in this
project was involved in flagellar biosynthesis. Treatment with sub-lethal concentrations of
carvacrol, one of the major antimicrobial components in the essential oils of oregano and
thyme, inhibited the production of flagellin synthesis in E. coli, while production of heat
shock protein 60 was induced (Burt et al., 2007). Exposure to sub-inhibitory concentrations
of piperacillin/tazobactam caused a significant decrease in flagellum-mediated swimming in
P. aeruginosa (Fonseca et al., 2004) and while treatment with sub-MIC concentrations of
mupirocin reduced flagellin expression and flagella formation in P. aeruginosa (Horii et
al., 2003). It would be of interest to investigate flagellin synthesis in both P. aeruginosa

Chapter Seven –General Discussion
124
and E. coli following treatment with sub-lethal concentrations of terpinen-4-ol and other
components of TTO and also to explore the relationship between flagella and expression of
efflux systems in P. aeruginosa. Zhang et al. (2007) suggested that flagellar mutants have a
reduction in LPS expression and subsequently a reduction in the stability of the outer
membrane, causing an increase in susceptibility to agents such as SDS. It would be
interesting to examine the relationship between flagella and LPS expression, and
susceptibility to TTO, in P. aeruginosa.
Another disrupted gene identified in three of the transposon mutants created in this study
was surA. In E. coli, SurA facilitates the maturation of outer membrane porins and is a
periplasmic chaperone (Bitto and McKay, 2003; Hennecke et al., 2005). Assembly of TolC,
an outer membrane protein in E. coli involved in antibiotic efflux, is not affected by the loss
of SurA (Werner et al., 2003). However, it is possible that SurA is required as a chaperone
of outer membrane proteins belonging to efflux systems in P. aeruginosa, and therefore the
disruption of surA may lead to a decrease in efflux, and hence an increase in susceptibility
to the substrates of said efflux system. This could be explored by examining the efflux
status of the three surA mutants from this study.
The creation of a library of transposon mutants in this project was more difficult and took
much longer than anticipated. The EZ::TN <KAN-2> Transposome kit was selected for this
process because it presented a straight-forward method for generating a mutant library, in
conjunction with the ability to easily identify the interrupted gene(s). Literature associated
with the kit indicated the user could expect to generate approximately 102 mutants per
reaction from a Pseudomonas sp. However, this was not the case. Each kit allowed for 10
reactions and it soon became apparent after the first mutagenesis yielded only one mutant,
that optimisation was needed. It was not viable to use the actual kit when optimizing the
different parts of the assay, including the method of cell preparation as well as the
electroporation parameters. Optimization was done using several different plasmids and
recipient strains of both E. coli and P. aeruginosa. While not ideal, this avenue of
optimization did enhance the effectiveness of the mutagenesis and allowed expansion of the
mutant library to a total of 537 confirmed mutants. In hindsight it would probably have

Chapter Seven –General Discussion
125
ended up being faster and cheaper to have used a non-kit-based mutagenesis protocol with
Tn5.
In a recent study, Shapira & Mimran (2007) screened a library of 20,000 random
transposon mutants of E. coli for altered susceptibility to the essential oil thymol (Shapira
and Mimran, 2007). Interestingly, the EZ::TN<R6Kγori/KAN-2> transposon was used to
create the library. This is similar to the EZ::TN<KAN-2> transposon but features the R6Kγ
origin of replication which aids in the recovery of the sequence flanking the transposon
insertion site. Despite the large size of the library, Shapira & Mimran only identified 4
mutants with increased susceptibility to thymol, of which 3 were mutated in the same gene
and 5 mutants that were more resistant to thymol. The gene that was identified as disrupted
in 3 of the mutants more susceptible to thymol was rfaQ, the product of which is a glycosyl
transferase involved in LPS-core biosynthesis (Shapira and Mimran, 2007). The MIC of
thymol against the three mutants was decreased by 14.6% compared to the wild type. A
slight decrease in the MIC of menthol and carvacrol was also reported. These results are
worthy of note given the findings in Chapter Four, i.e. that a full LPS core is required for
tolerance to TTO and terpinen-4-ol.
Although the transposon mutant library discussed in this thesis was small compared to
other published transposon mutant libraries from P. aeruginosa, there was still a total of 20
isolates with altered susceptibility to TTO and/or terpinen-4-ol, with some interesting
results discovered upon identification of the disrupted genes. Complementation of all
transposon mutants with altered susceptibilities created in this study is required to
demonstrate that the changes in susceptibility were directly related to the loss of function of
the identified genes. Any fitness costs associated with the transposon mutants also needs to
be investigated.
Although not explored in this thesis, the possibility exists that P. aeruginosa is able to
tolerate such high concentrations of TTO through the process of biodegradation. Members
of the Pseudomonas genus, particularly P. putida, possess the capability of biodegrading
aromatic hydrocarbons following their transportation across the cell membrane (Hearn et

Chapter Seven –General Discussion
126
al., 2008). P. fluorescens was able to biodegrade α-pinene, dissolved in organic solvents
within a bacterial two liquid phase bioreactor (Munoz et al., 2008).α-Pinene usually
comprises around 1-6% (v/v) in TTO, and was present at 2.4% (v/v) in the batch of TTO
used in this thesis. Pseudomonas pseudoalcaligenes KF707 was able to utilize ρ-cymene or
α-terpinene as a sole carbon source and both terpenes enhanced survival in the presence of
polychlorinated biphenyl (Oh et al., 2003). The batch of TTO used in this thesis contained
2.4% ρ-cymene and 8.6% α-terpinene. It would be interesting to determine whether
P. aeruginosa can use any of the terpene components in TTO as a sole carbon source, and
whether catabolism or degradation of the components of TTO is another mechanism for
tolerance.
The key achievements of this thesis can be summarised as follows;
a) Components of TTO, including terpinen-4-ol, 1,8-cineole and α-terpineol, were
identified as substrates of the MexAB-OprM efflux system.
b) Terpinen-4-ol was identified as a possible substrate of MexEF-OprN.
c) Transposon mutants with insertions in flagellum biogenesis genes and a gene
responsible for correct folding of outer membrane proteins, surA, were created and
were more susceptible to TTO and terpinen-4-ol.
d) A complete LPS core was shown to be required to protect P. aeruginosa from the
antibacterial actions of TTO and terpinen-4-ol.
e) When P. aeruginosa was repeatedly subcultured in the presence of TTO, an
increase in resistance to a range of antibiotics was detected, though only resistance
to aztreonam, moxalactam and ceftriaxone was at clinically significant levels.
f) Repeated subculture in TTO did not significantly increase tolerance to TTO.
P. aeruginosa is a significant opportunistic pathogen that is intrinsically more resistant to
TTO than most other bacteria. Previous studies attributed this to the outer membrane. The
work described in this thesis has expanded the knowledge of the mechanisms of tolerance

Chapter Seven –General Discussion
127
to TTO to include the MexAB-OprM efflux system, a complete LPS core, and flagellum
biogenesis.

References
128
REFERENCES
Adewoye, L., Sutherland, A., Srikumar, R., and Poole, K. (2002) The mexR repressor of the mexAB-oprM
multidrug efflux operon in Pseudomonas aeruginosa: characterization of mutations compromising
activity. J. Bacteriol. 184: 4308-4312.
Adhikari, P., Kirby, S.D., Nowalk, A.J., Veraldi, K.L., Schryvers, A.B., and Mietzner, T.A. (1995)
Biochemical characterization of a Haemophilus influenzae periplasmic iron transport operon. J. Biol.
Chem. 270: 25142-25149.
Aendekerk, S., Ghysels, B., Cornelis, P., and Baysse, C. (2002) Characterization of a new efflux pump,
MexGHI-OpmD, from Pseudomonas aeruginosa that confers resistance to vanadium. Microbiol.
148: 2371-2381.
Aendekerk, S., Diggle, S.P., Song, Z., Hoiby, N., Cornelis, P., Williams, P., and Camara, M. (2005) The
MexGHI-OpmD multidrug efflux pump controls growth, antibiotic susceptibility and virulence in
Pseudomonas aeruginosa via 4-quinolone-dependent cell-to-cell communication. Microbiol. 151:
1113-1125.
Aires, J.R., Kohler, T., Nikaido, H., and Plesiat, P. (1999) Involvement of an active efflux system in the
natural resistance of Pseudomonas aeruginosa to aminoglycosides. Antimicrob. Agents Chemother.
43: 2624-2628.
Al-Tahhan, R.A., Sandrin, T.R., Bodour, A.A., and Maier, R.M. (2000) Rhamnolipid-induced removal of
lipopolysaccharide from Pseudomonas aeruginosa: effect on cell surface properties and interaction
with hydrophobic substrates. Appl. Environ. Microbiol. 66: 3262-3268.
Alekshun, M.N., and Levy, S.B. (1999) The mar regulon: multiple resistance to antibiotics and other toxic
chemicals. Trends Microbiol. 7: 410-413.
Altman, E., I. Sadovskaya, J. S. Lam, and J. C. Richards. (1993) Structural analysis of Pseudomonas
aeruginosa IATS O5 lipopolysaccharide core oligosaccharide. Fourth International Symposium on
Pseudomonas: Biotechnology and Molecular Biology. Session I, Abstr. 29.
Altman, P.M. (1988) Australian tea tree oil. The Australian Journal of Pharmacy 69: 276-278.
Andriole, V.T. (2004) Quinolones. In Treatment of infectious diseases. Gorbach, S.L., Bartlett, J.G. and
Blacklow, N.R. (eds): Lippincott, Williams and Wilkins.
Aoki, F.Y. (2005) Chapter 45 - Principles of antimicrobial therapy and the clinical pharmacology of
antimicrobial drugs. In Principles of critical care. Schmidt, G.A., Hall, J.B. and Wood, L.D.H. (eds):
McGraw-Hill Professional.
Aono, R., Negishi, T., and Nakajima, H. (1994) Cloning of organic solvent tolerance gene ostA that
determines n-hexane tolerance level in Escherichia coli. Appl. Environ. Microbiol. 60: 4624-4626.
Aono, R., and Kobayashi, H. (1997) Cell surface properties of organic solvent-tolerant mutants of Escherichia
coli K-12. Appl. Environ. Microbiol. 63: 3637-3642.
Araujo, I.B., Souza, C.A., De-Carvalho, R.R., Kuriyama, S.N., Rodrigues, R.P., Vollmer, R.S., Alves, E.N.,
and Paumgartten, F.J. (1996) Study of the embryofoetotoxicity of alpha-terpinene in the rat. Food
Chem. Toxicol. 34: 477-482.
Arora, S.K., Ritchings, B.W., Almira, E.C., Lory, S., and Ramphal, R. (1998) The Pseudomonas aeruginosa
flagellar cap protein, FliD, is responsible for mucin adhesion. Infect. Immun. 66: 1000-1007.
Arora, S.K., Bangera, M., Lory, S., and Ramphal, R. (2001) A genomic island in Pseudomonas aeruginosa
carries the determinants of flagellin glycosylation. Proc. Natl. Acad. Sci. U S A 98: 9342-9347.
Asako, H., Nakajima, H., Kobayashi, K., Kobayashi, M., and Aono, R. (1997) Organic solvent tolerance and
antibiotic resistance increased by overexpression of marA in Escherichia coli. Appl Environ
Microbiol 63: 1428-1433.

References
129
Aspres, N., and Freeman, S. (2003) Predictive testing for irritancy and allergenicity of tea tree oil in normal
human subjects. Exogenous Dermatol. 2: 258-261.
Banes-Marshall, L., Cawley, P., and Phillips, C.A. (2001) In vitro activity of Melaleuca alternifolia (tea tree)
oil against bacterial and Candida spp. isolates from clinical specimens. Br. J. Biomed. Sci. 58: 139-
145.
Behrens, S., Maier, R., de Cock, H., Schmid, F.X., and Gross, C.A. (2001) The SurA periplasmic PPIase
lacking its parvulin domains functions in vivo and has chaperone activity. Embo. J. 20: 285-294.
Bennett, G.M., Seaver, A., and Calcott, P.H. (1981) Effect of defined lipopolysaccharide core defects on
resistance of Salmonella typhimurium to freezing and thawing and other stresses. Appl. Environ.
Microbiol. 42: 843-849.
Bhushan, M., and Beck, M.H. (1997) Allergic contact dermatitis from tea tree oil in a wart paint. Contact
Derm. 36: 117-118.
Bishop, C.D. (1995) Antiviral activity of the essential oil of Melaleuca alternifolia (Maiden & Betche) Cheel
(Tea Tree) against tobacco mosaic virus. J. Ess. Oil Res. 7: 641-644.
Bitto, E., and McKay, D.B. (2003) The periplasmic molecular chaperone protein SurA binds a peptide motif
that is characteristic of integral outer membrane proteins. J. Biol. Chem. 278: 49316-49322.
Boutoille, D., Corvec, S., Caroff, N., Giraudeau, C., Espaze, E., Caillon, J., Plesiat, P., and Reynaud, A.
(2004) Detection of an IS21 insertion sequence in the mexR gene of Pseudomonas aeruginosa
increasing [beta]-lactam resistance. FEMS Micro. Lett. 230: 143-146.
Bowers, L.M., Lapoint, K., Anthony, L., Pluciennik, A., and Filutowicz, M. (2004) Bacterial expression
system with tightly regulated gene expression and plasmid copy number. Gene 340: 11-18.
Brand, C., Ferrante, A., Prager, R.H., Riley, T.V., Carson, C.F., Finlay-Jones, J.J., and Hart, P.H. (2001) The
water soluble components of the essential oil of Melaleuca alternifolia (tea tree oil), suppress the
production of superoxide by human monocytes, but not neutrophils, activated in vitro. Inflamm. Res.
50: 213-219.
Brand, C., Grimbaldeston, M.A., Gamble, J.R., Drew, J., Finlay-Jones, J.J., and Hart, P.H. (2002a) Tea tree
oil reduces the swelling associated with the efferent phase of a contact hypersensitivity response.
Inflamm. Res. 51: 236-244.
Brand, C., Townley, S.L., Finlay-Jones, J.J., and Hart, P.H. (2002b) Tea tree oil reduces histamine-induced
oedema in murine ears. Inflamm. Res. 51: 283-289.
Brophy, J.J., Davies, N.W., Southwell, I.A., Stiff, I.A., and Williams, L.R. (1989) Gas chromatographic
quality control for oil of Melaleuca terpinen-4-ol type (Australian tea tree). J. Agric. Food Chem. 37:
1330-1335.
Brown, M.H., Paulsen, I.T., and Skurray, R.A. (1999) The multidrug efflux protein NorM is a prototype of a
new family of transporters. Mol. Microbiol. 31: 394-395.
Burt, S.A., van der Zee, R., Koets, A.P., de Graaff, A.M., van Knapen, F., Gaastra, W., Haagsman, H.P., and
Veldhuizen, E.J. (2007) Carvacrol induces heat shock protein 60 and inhibits synthesis of flagellin in
Escherichia coli O157:H7. Appl. Environ. Microbiol. 73: 4484-4490.
Calcabrini, A., Stringaro, A., Toccacieli, L., Meschini, S., Marra, M., Colone, M., Salvatore, G., Mondello, F.,
Arancia, G., and Molinari, A. (2004) Terpinen-4-ol, the main component of Melaleuca alternifolia
(Tea Tree) oil inhibits the in vitro growth of human melanoma cells. J. Invest. Derm. 122: 349-360.
Caldefie-Chezet, F., Guerry, M., Chalchat, J.C., Fusillier, C., Vasson, M.P., and Guillot, J. (2004) Anti-
inflammatory effects of Melaleuca alternifolia essential oil on human polymorphonuclear
neutrophils and monocytes. Free Radic. Res. 38: 805-811.

References
130
Campigotto, A.J. (2007) Influence of membrane-damaging agents and the sigma factor AlgU on the induction
of the MexCD-OprJ efflux system of Pseudomonas aeruginosa. In Microbiology & Immunology.
Vol. Masters of Science Ontario: Queen's University, pp. 145.
Campo-Esquisabel, A.B., Marti, S., Sierra, J.M., Martinez-Martinez, L., and Vila, J. (2007) Altered outer
membrane protein profiles of Pseudomonas aeruginosa highly susceptible to the efflux pump
inhibitor Phe-Arg-beta-naphthlamide. In 17th European Congress of Clinical Microbiology and
Infectious Diseases. Vol. P881 Munich, Germany.
Cao, L., Srikumar, R., and Poole, K. (2004) MexAB-OprM hyperexpression in NalC-type multidrug-resistant
Pseudomonas aeruginosa: identification and characterization of the nalC gene encoding a repressor
of PA3720-PA3719. Mol. microbiol. 53: 1423-1436.
Carson, C.F., Hammer, K.A., and Riley, T.V. (1995) Broth micro-dilution method for determining the
susceptibility of Escherichia coli and Staphylococcus aureus to the essential oil of Melaleuca
alternifolia (tea tree oil). Microbios 82: 181-185.
Carson, C.F., Ashton, L., Dry, L., Smith, D.W., and Riley, T.V. (2001) Melaleuca alternifolia (tea tree) oil
gel (6%) for the treatment of recurrent herpes labialis. J. Antimicrob. Chemother. 48: 450-451.
Carson, C.F., Mee, B.J., and Riley, T.V. (2002) Mechanism of action of Melaleuca alternifolia (tea tree) oil
on Staphylococcus aureus determined by time-kill, lysis, leakage, and salt tolerance assays and
electron microscopy. Antimicrob. Agents Chemother. 46: 1914-1920.
Carson, C.F., Hammer, K.A., and Riley, T.V. (2006) Melaleuca alternifolia (Tea Tree) oil: a review of
antimicrobial and other medicinal properties. Clin. Microbiol. Rev. 19: 50-62.
Cavallo, J.D., Fabre, R., Leblanc, F., Nicolas-Chanoine, M.H., and Thabaut, A. (2000) Antibiotic
susceptibility and mechanisms of beta-lactam resistance in 1310 strains of Pseudomonas aeruginosa:
a French multicentre study (1996). J. Antimicrob. Chemother. 46: 133-136.
Cavallo, J.D., Plesiat, P., Couetdic, G., Leblanc, F., and Fabre, R. (2002) Mechanisms of beta-lactam
resistance in Pseudomonas aeruginosa: prevalence of OprM-overproducing strains in a French
multicentre study (1997). J Antimicrob Chemother 50: 1039-1043.
Champlin, F.R., Ellison, M.L., Bullard, J.W., and Conrad, R.S. (2005) Effect of outer membrane
permeabilisation on intrinsic resistance to low triclosan levels in Pseudomonas aeruginosa. Int. J.
Antimicrob. Agents 26: 159-164.
Chiu, H.C., Lin, T.L., and Wang, J.T. (2007) Identification and characterization of an organic solvent
tolerance gene in Helicobacter pylori. Helicobacter 12: 74-81.
Choi, K.H., Kumar, A., and Schweizer, H. P. (2005) A 10 min method for preparation of highly
electrocompetent Pseudomonas aeruginosa cells: application for DNA fragment transfer between
chromosomes and plasmid transformation. J. Microbiol. Methods 64: 391-397.
Chuanchuen, R., Beinlich, K., Hoang, T.T., Becher, A., Karkhoff-Schweizer, R.R., and Schweizer, H.P.
(2001) Cross-resistance between triclosan and antibiotics in Pseudomonas aeruginosa is mediated by
multidrug efflux pumps: exposure of a susceptible mutant strain to triclosan selects nfxB mutants
overexpressing MexCD-OprJ. Antimicrob. Agents Chemother. 45: 428-432.
Chuanchuen, R., Narasaki, C.T., and Schweizer, H.P. (2002) The MexJK efflux pump of Pseudomonas
aeruginosa requires OprM for antibiotic efflux but not for efflux of triclosan. J. Bacteriol. 184:
5036-5044.
Chuanchuen, R., Karkhoff-Schweizer, R.R., and Schweizer, H.P. (2003) High-level triclosan resistance in
Pseudomonas aeruginosa is solely a result of efflux. Am. J. Infect. Control 31: 124-127.
Chuanchuen, R., Murata, T., Gotoh, N., and Schweizer, H.P. (2005) Substrate-dependent utilization of OprM
or OpmH by the Pseudomonas aeruginosa MexJK Efflux Pump. Antimicrob. Agents Chemother. 49:
2133-2136.

References
131
Cirioni, O., Giacometti, A., Silvestri, C., Della Vittoria, A., Licci, A., Riva, A., and Scalise, G. (2006) In vitro
activities of tritrpticin alone and in combination with other antimicrobial agents against
Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 50: 3923-3925.
CLSI (2006) Clinical and Laboratory Standards Institute Methods for dilution antimicrobial susceptibility
tests for bacteria that grow aerobically. Villanova, PA, USA.
CLSI (2007) Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial
Susceptibility Testing: Seventh Informational Supplement. Wayne, Pennsylvania, USA.
Colling, L., Carter, R.N., Essmann, M., and Larsen, B. (2005) Evaluation of relative yeast cell surface
hydrophobicity measured by flow cytometry. Infect. Dis. Obstet. Gynecol. 13: 43-48.
Corbett (2006) High resolution melt assay design and analysis: CorProtocol™ 6000-1-July06. In Rotor-
GeneTM
6000 real time rotary analyzer.
Cox, S.D., Gustafson, J.E., Mann, C.M., Markham, J.L., Liew, Y.C., Hartland, R.P., Bell, H.C., Warmington,
J.R., and Wyllie, S.G. (1998) Tea tree oil causes K+ leakage and inhibits respiration in Escherichia
coli. Lett. Appl. Microbiol. 26: 355-358.
Cox, S.D., Mann, C.M., Markham, J.L., Bell, H.C., Gustafson, J.E., Warmington, J.R., and Wyllie, S.G.
(2000) The mode of antimicrobial action of the essential oil of Melaleuca alternifolia (tea tree oil). J.
Appl. Microbiol. 88: 170-175.
Cox, S.D., Mann, C.M., Markham, J.L., Gustafson, J.E., Warmington, J.R., and Wyllie, S.G. (2001)
Determining the antimicrobial actions of tea tree oil. Molecules 6: 87-91.
Cox, S.D., Markham, L. J (2007) Susceptibility and intrinsic tolerance of Pseudomonas aeruginosa to
selected plant volatile compounds. J. Appl. Microbiol. 103: 930-936.
Cryz, S.J., Jr., Pitt, T.L., Furer, E., and Germanier, R. (1984) Role of lipopolysaccharide in virulence of
Pseudomonas aeruginosa. Infect. Immun. 44: 508-513.
D'Auria, F.D., Laino, L., Strippoli, V., Tecca, M., Salvatore, G., Battinelli, L., and Mazzanti, G. (2001) In
vitro activity of tea tree oil against Candida albicans mycelial conversion and other pathogenic
fungi. J. Chemother.. 13: 377-383.
Dartigalongue, C., Missiakas, D., and Raina, S. (2001) Characterization of the Escherichia coli sigma E
regulon. J. Biol. Chem. 276: 20866-20875.
Dasgupta, N., Arora, S.K., and Ramphal, R. (2000) fleN, a gene that regulates flagellar number in
Pseudomonas aeruginosa. J. Bacteriol. 182: 357-364.
Dasgupta, N., and Ramphal, R. (2001) Interaction of the antiactivator FleN with the transcriptional activator
FleQ regulates flagellar number in Pseudomonas aeruginosa. J. Bacteriol. 183: 6636-6644.
Dasgupta, N., Wolfgang, M.C., Goodman, A.L., Arora, S.K., Jyot, J., Lory, S., and Ramphal, R. (2003) A
four-tiered transcriptional regulatory circuit controls flagellar biogenesis in Pseudomonas
aeruginosa. Mol. Microbiol. 50: 809-824.
Dasgupta, T., de Kievit, T.R., Masoud, H., Altman, E., Richards, J.C., Sadovskaya, I., Speert, D.P., and Lam,
J.S. (1994) Characterization of lipopolysaccharide-deficient mutants of Pseudomonas aeruginosa
derived from serotypes O3, O5, and O6. Infect. Immun. 62: 809-817.
De Groot, A.C. (1996) Airborne allergic contact dermatitis from tea tree oil. Contact Derm. 35: 304-305.
De Kievit, T.R., Parkins, M.D., Gillis, R.J., Srikumar, R., Ceri, H., Poole, K., Iglewski, B.H., and Storey,
D.G. (2001) Multidrug efflux pumps: expression patterns and contribution to antibiotic resistance in
Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 45: 1761-1770.
Delgado, I.F., Nogueira, A.C., Souza, C.A., Costa, A.M., Figueiredo, L.H., Mattos, A.P., Chahoud, I., and
Paumgartten, F.J. (1993) Peri- and postnatal developmental toxicity of beta-myrcene in the rat. Food
Chem. Toxicol. 31: 623-628.

References
132
Di Pasqua, R., Hoskins, N., Betts, G., and Mauriello, G. (2006) Changes in membrane fatty acids composition
of microbial cells induced by addiction of thymol, carvacrol, limonene, cinnamaldehyde, and
eugenol in the growing media. J. Agric. Food Chem. 54: 2745-2749.
Driscoll, J.A., Brody, S.L., and Kollef, M.H. (2007) The epidemiology, pathogenesis and treatment of
Pseudomonas aeruginosa infections. Drugs 67: 351-368.
Ducey, T.F., and Dyer, D.W. (2002) Rapid identification of EZ::TNTM
transposon insertion sites in the
genome of Neisseria gonorrhoeae. Epicentre Forum 9: 6-7.
Dumas, J.L., van Delden, C., Perron, K., and Kohler, T. (2006) Analysis of antibiotic resistance gene
expression in Pseudomonas aeruginosa by quantitative real-time-PCR. FEMS Microbiol. Lett. 254:
217-225.
Edris, A.E. (2007) Pharmaceutical and therapeutic potentials of essential oils and their individual volatile
constituents: a review. Phytother Res 21: 308-323.
Erbay, H., Yalcin, A.N., Serin, S., Turgut, H., Tomatir, E., Cetin, B., and Zencir, M. (2003) Nosocomial
infections in intensive care unit in a Turkish university hospital: a 2-year survey. Intensive Care
Med. 29: 1482-1488.
Ermolaeva, M.D., White, O., and Salzberg, S.L. (2001) Prediction of operons in microbial genomes. Nucleic
Acids Res. 29: 1216-1221.
Fath, M., and Kolter, R. (1993) ABC transporters: bacterial exporters. Microbiol. Rev. 57: 995-1017.
Flaherty, J., and Stosor, V. (2004) Chapter 34: Non-fermentative Gram negative bacilli. Section V:
Epidemiology and prevention of nosocomial infections caused by specific pathogens. In Hospital
Epidemiology and Infection Control. Mayhall, C. (ed): Lippincott Williams & Wilkins.
Fonseca, A.P., Extremina, C., Fonseca, A.F., and Sousa, J.C. (2004) Effect of subinhibitory concentration of
piperacillin/tazobactam on Pseudomonas aeruginosa. J. Med. Microbiol. 53: 903-910.
Forbes, B.A., Sahm, D.F., and Weissfeld, A.S. (1998) Pseudomonas, Burkholderia, and similar organisms. In
Bailey and Scott's diagnostic microbiology. Roche, J. (ed). St Louis: Mosby Inc., pp. 447-461.
Fukuda, H., Hosaka, M., Hirai, K., and Iyobe, S. (1990) New norfloxacin resistance gene in Pseudomonas
aeruginosa PAO. Antimicrob. Agents Chemother. 34: 1757-1761.
Galbraith, L., Wilkinson, S.G., Legakis, N.J., Genimata, V., Katsorchis, T.A., and Rietschel, E.T. (1984)
Structural alterations in the envelope of a gentamicin-resistant rough mutant of Pseudomonas
aeruginosa. Ann. Microbiol. (Paris) 135B: 121-136.
Gao, Y.Y., Di Pascuale, M.A., Elizondo, A., and Tseng, S.C. (2007) Clinical treatment of ocular demodecosis
by lid scrub with tea tree oil. Cornea 26: 136-143.
Gaynes, R., and Edwards, J.R. (2005) Overview of nosocomial infections caused by gram-negative bacilli.
Clin. Infect. Dis. 41: 848-854.
Giordani, C., Molinari, A., Toccacieli, L., Calcabrini, A., Stringaro, A., Chistolini, P., Arancia, G., and
Diociaiuti, M. (2006) Interaction of tea tree oil with model and cellular membranes. J. Med. Chem.
49: 4581-4588.
Golab, M., Burdzenia, O., Majewski, P., and Skwarlo-Sonta, K. (2005) Tea tree oil inhalations modify
immunity in mice. J. Appl. Biomed. 3: 101-108.
Golab, M., and Skwarlo-Sonta, K. (2007) Mechanisms involved in the anti-inflammatory action of inhaled tea
tree oil in mice. Exp. Biol. Med. 232: 420-426.
Goldberg, J.B., Hatano, K., and Pier, G.B. (1993) Synthesis of lipopolysaccharide O side chains by
Pseudomonas aeruginosa PAO1 requires the enzyme phosphomannomutase. J. Bacteriol. 175: 1605-
1611.

References
133
Gotoh, N., Tsujimoto, H., Tsuda, M., Okamoto, K., Nomura, A., Wada, T., Nakahashi, M., and Nishino, T.
(1998) Characterization of the MexC-MexD-OprJ multidrug efflux system in DeltamexA-mexB-
oprM mutants of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 42: 1938-1943.
Griffin, S.G., and Markham, J.L. (2000) An agar dilution method for the determination of the minimum
inhibitory concentration of essential oils. J. Ess. Oil Res. 12: 249-255.
Guerin-Mechin, L., Dubois-Brissonnet, F., Heyd, B., and Leveau, J.Y. (1999) Specific variations of fatty acid
composition of Pseudomonas aeruginosa ATCC 15442 induced by quaternary ammonium
compounds and relation with resistance to bactericidal activity. J. Appl. Microbiol. 87: 735-742.
Guha, A., Choudhury, A., Unni, B.G., and Roy, M.K. (2002) Effect of outer-membrane permeabilizers on the
activity of antibiotics and plant extracts against Pseudomonas aeruginosa. Folia Microbiol. (Praha)
47: 379-384.
Gustafson, J.E., Cox, S.D., Liew, Y.C., Wyllie, S.G., and Warmington, J.R. (2001) The bacterial multiple
antibiotic resistant (Mar) phenotype leads to increased tolerance to tea tree oil. Pathology 33: 211-
215.
Gustin, J.K., Kessler, E., and Ohman, D.E. (1996) A substitution at His-120 in the LasA protease of
Pseudomonas aeruginosa blocks enzymatic activity without affecting propeptide processing or
extracellular secretion. J. Bacteriol. 178: 6608-6617.
Hammer, K.A., Carson, C.F., and Riley, T.V. (1996) Susceptibility of transient and commensal skin flora to
the essential oil of Melaleuca alternifolia (tea tree oil). Amer. J. Infect. Cont. 24: 186-189.
Hammer, K.A., Carson, C.F., and Riley, T.V. (2000) Melaleuca alternifolia (tea tree) oil inhibits germ tube
formation by Candida albicans. Med. Mycol. 38: 355-362.
Hammer, K.A., Carson, C.F., and Riley, T.V. (2002) In vitro activity of Melaleuca alternifolia (tea tree) oil
against dermatophytes and other filamentous fungi. J. Antimicrob. Chemother. 50: 195-199.
Hammer, K.A., Carson, C.F., and Riley, T.V. (2003) Antifungal activity of the components of Melaleuca
alternifolia (tea tree) oil. J. Appl. Microbiol. 95: 853-860.
Hammer, K.A., Carson, C.F., and Riley, T.V. (2004) Antifungal effects of Melaleuca alternifolia (tea tree) oil
and its components on Candida albicans, Candida glabrata and Saccharomyces cerevisiae. J.
Antimicrob. Chemother. 53: 1081-1085.
Hammer, K.A., Carson, C.F., and Riley, T.V. (2008) Frequencies of resistance to Melaleuca alternifolia (tea
tree) oil and rifampicin in Staphylococcus aureus, Staphylococcus epidermidis and Enterococcus
faecalis. Int. J. Antimicrob. Agents 32: 170-173.
Hancock, R.E., and Carey, A.M. (1979) Outer membrane of Pseudomonas aeruginosa: heat- 2-
mercaptoethanol-modifiable proteins. J. Bacteriol. 140: 902-910.
Haraguchi, H., Oike, S., Muroi, H., and Kubo, I. (1996) Mode of antibacterial action of totarol, a diterpene
from Podocarpus nagi. Planta Med. 62: 122-125.
Hart, P.H., Brand, C., Carson, C.F., Riley, T.V., Prager, R.H., and Finlay-Jones, J.J. (2000) Terpinen-4-ol, the
main component of the essential oil of Melaleuca alternifolia (tea tree oil), suppresses inflammatory
mediator production by activated human monocytes. Inflamm. Res. 49: 619-626.
He, G.X., Kuroda, T., Mima, T., Morita, Y., Mizushima, T., and Tsuchiya, T. (2004) An H(+)-coupled
multidrug efflux pump, PmpM, a member of the MATE family of transporters, from Pseudomonas
aeruginosa. J. Bacteriol. 186: 262-265.
Hearn, E.M., Patel, D.R., and van den Berg, B. (2008) Outer-membrane transport of aromatic hydrocarbons as
a first step in biodegradation. Proc. Natl. Acad. Sci. U S A 105: 8601-8606.
Heath, R.J., Rubin, J.R., Holland, D.R., Zhang, E., Snow, M.E., and Rock, C.O. (1999) Mechanism of
triclosan inhibition of bacterial fatty acid synthesis. J. Biol. Chem. 274: 11110-11114.

References
134
Heipieper, H.J., Meinhardt, F., and Segura, A. (2003) The cis-trans isomerase of unsaturated fatty acids in
Pseudomonas and Vibrio: biochemistry, molecular biology and physiological function of a unique
stress adaptive mechanism. FEMS Microbiol. Lett. 229: 1-7.
Henley, D.V., Lipson, N., Korach, K.S., and Bloch, C.A. (2007) Prepubertal gynecomastia linked to lavender
and tea tree oils. N. Engl. J. Med. 356: 479-485.
Hennecke, G., Nolte, J., Volkmer-Engert, R., Schneider-Mergener, J., and Behrens, S. (2005) The periplasmic
chaperone SurA exploits two features characteristic of integral outer membrane proteins for selective
substrate recognition. J. Biol. Chem. 280: 23540-23548.
Heurlier, K., Denervaud, V., Pessi, G., Reimmann, C., and Haas, D. (2003) Negative control of quorum
sensing by RpoN (sigma54) in Pseudomonas aeruginosa PAO1. J. Bacteriol. 185: 2227-2235.
Hirakata, Y., Srikumar, R., Poole, K., Gotoh, N., Suematsu, T., Kohno, S., Kamihira, S., Hancock, R.E., and
Speert, D.P. (2002) Multidrug efflux systems play an important role in the invasiveness of
Pseudomonas aeruginosa. J. Exp. Med. 196: 109-118.
Hirakata, Y., Kondo, A., Hoshino, K., Miyazaki, T., Kurihara, S., Izumikawa, K., Seki, M., Yanagihara, K.,
Miyazaki, S., Kamihira, S., and Kohno, S. (2007) Efflux pump inhibitors reduce the invasiveness of
Pseudomonas aeruginosa. In 17th European Congress of Clinical Microbiology and Infectious
Diseases Vol. P817 Munich, Germany.
Hoang, T.T., and Schweizer, H.P. (1999) Characterization of Pseudomonas aeruginosa enoyl-acyl carrier
protein reductase (FabI): a target for the antimicrobial triclosan and its role in acylated homoserine
lactone synthesis. J. Bacteriol. 181: 5489-5497.
Hocquet, D., Roussel-Delvallez, M., Cavallo, J.D., and Plesiat, P. (2007) MexAB-OprM- and MexXY-
overproducing mutants are very prevalent among clinical strains of Pseudomonas aeruginosa with
reduced susceptibility to ticarcillin. Antimicrob. Agents Chemother. 51: 1582-1583.
Hoffman, L.M., and Jendrisak, J. (1999) Use of EZ::TNTM
TransposomesTM
for genetic analysis and direct
sequencing of bacterial genomic DNA. Epicentre Forum 6: 1-4.
Hoffman, L.M., Jendrisak, J.J., Meis, R.J., Goryshin, I.Y., and Reznikof, S.W. (2000) Transposome
insertional mutagenesis and direct sequencing of microbial genomes. Genetica 108: 19-24.
Horii, T., Morita, M., Muramatsu, H., Muranaka, Y., Kanno, T., and Maekawa, M. (2003) Effects of
mupirocin at subinhibitory concentrations on flagella formation in Pseudomonas aeruginosa and
Proteus mirabilis. J. Antimicrob. Chemother. 51: 1175-1179.
International Organisation for Standardisation (2004) ISO 4730:2004, Oil of Melaleuca, terpinen-4-ol type
(tea tree oil). Geneva, Switzerland: International Organisation for Standardisation.
Isken, S., and de Bont, J.A. (1998) Bacteria tolerant to organic solvents. Extremophiles 2: 229-238.
Jacobs, M.A., Alwood, A., Thaipisuttikul, I., Spencer, D., Haugen, E., Ernst, S., Will, O., Kaul, R., Raymond,
C., Levy, R., Chun-Rong, L., Guenthner, D., Bovee, D., Olson, M.V., and Manoil, C. (2003)
Comprehensive transposon mutant library of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U S A
100: 14339-14344.
Jacobs, M.R., and Hornfeldt, C.S. (1994) Melaleuca Oil Poisoning. J. Toxicol. - Clin. Toxicol. 32: 461-464.
Jarrell, K.F., and Kropinski, A.M. (1981) Isolation and characterization of a bacteriophage specific for the
lipopolysaccharide of rough derivatives of Pseudomonas aeruginosa strain PAO. J. Virol. 38: 529-
538.
Jeannot, K., Sobel, M.L., El Garch, F., Poole, K., and Plesiat, P. (2005) Induction of the MexXY efflux pump
in Pseudomonas aeruginosa is dependent on drug-ribosome interaction. J. Bacteriol. 187: 5341-
5346.

References
135
Jiang, X., Zhang, Z., Li, M., Zhou, D., Ruan, F., and Lu, Y. (2006) Detection of extended-spectrum beta-
lactamases in clinical isolates of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 50:
2990-2995.
Jones, M.V., Herd, T.M., and Christie, H.J. (1989) Resistance of Pseudomonas aeruginosa to amphoteric and
quaternary ammonium biocides. Microbios 58: 49-61.
Joswick, H.L., Corner, T.R., Silvernale, J.N., and Gerhardt, P. (1971) Antimicrobial actions of
hexachlorophene: release of cytoplasmic materials. J. Bacteriol. 108: 492-500.
Junker, F., Rodriguez-Herva, J.J., Duque, E., Ramos-Gonzalez, M.I., Llamas, M., and Ramos, J.L. (2001) A
WbpL mutant of Pseudomonas putida DOT-T1E strain, which lacks the O-antigenic side chain of
lipopolysaccharides, is tolerant to organic solvent shocks. Extremophiles 5: 93-99.
Kaatz, G.W. (2002) Inhibition of bacterial efflux pumps: a new strategy to combat increasing antimicrobial
agent resistance. Expert Opin. Emerg. Drugs. 7: 223-233.
Kadry, A.A. (2003) Lack of efflux mechanism in a clinical isolate of Pseudomonas aeruginosa highly
resistant to beta-lactams and imipenem. Folia Microbiol. (Praha). 48: 529-533.
Keen, N.T., Tamaki, S., Kobayashi, D., and Trollinger, D. (1988) Improved broad-host-range plasmids for
DNA cloning in gram-negative bacteria. Gene. 70: 191-197.
Khalil, Z., Pearce, A.L., Satkunanathan, N., Storer, E., Finlay-Jones, J.J., and Hart, P.H. (2004) Regulation of
wheal and flare by tea tree oil: complementary human and rodent studies. J. Invest. Dermatol. 123:
683-690.
Kieboom, J., Bruinenberg, R., Keizer-Gunnink, I., and de Bont, J.A. (2001) Transposon mutations in the
flagella biosynthetic pathway of the solvent-tolerant Pseudomonas putida S12 result in a decreased
expression of solvent efflux genes. FEMS Microbiol. Lett. 198: 117-122.
Kim, I.S., Shim, J.H., and Suh, Y.T. (2002) Changes in membrane fluidity and fatty acid composition of
Pseudomonas putida CN-T19 in response to toluene. Biosci. Biotechnol. Biochem. 66: 1945-1950.
Kim, J.M., Park, E.S., Jeong, J.S., Kim, K.M., Kim, J.M., Oh, H.S., Yoon, S.W., Chang, H.S., Chang, K.H.,
Lee, S.I., Lee, M.S., Song, J.H., Kang, M.W., Park, S.C., Choe, K.W., and Pai, C.H. (2000)
Multicenter surveillance study for nosocomial infections in major hospitals in Korea. Nosocomial
infection surveillance committee of the Korean society for nosocomial infection control. Am. J.
Infect. Control 28: 454-458.
Knight, T.E., and Hausen, B.M. (1994) Melaleuca oil (tea tree oil) dermatitis. J. Amer. Acad. Dermatol. 30:
423-427.
Kobayashi, H., Takami, H., Hirayama, H., Kobata, K., Usami, R., and Horikoshi, K. (1999) Outer membrane
changes in a toluene-sensitive mutant of toluene-tolerant Pseudomonas putida IH-2000. J. Bacteriol.
181: 4493-4498.
Kohler, T., Michea-Hamzehpour, M., Henze, U., Gotoh, N., Curty, L.K., and Pechere, J.C. (1997)
Characterization of MexE-MexF-OprN, a positively regulated multidrug efflux system of
Pseudomonas aeruginosa. Mol. Microbiol. 23: 345-354.
Kohler, T., Michea-Hamzehpour, M., Epp, S.F., and Pechere, J.C. (1999) Carbapenem activities against
Pseudomonas aeruginosa: respective contributions of OprD and efflux systems. Antimicrob. Agents
Chemother. 43: 424-427.
Kovach, M.E., Phillips, R.W., Elzer, P.H., Roop, R.M., 2nd, and Peterson, K.M. (1994) pBBR1MCS: a
broad-host-range cloning vector. Biotechniques. 16: 800-802.
Kropinski, A.M., Chan, L., and Milazzo, F.H. (1978) Susceptibility of lipopolysaccharide-defective mutants
of Pseudomonas aeruginosa strain PAO to dyes, detergents, and antibiotics. Antimicrob. Agents
Chemother. 13: 494-499.
Lam, J.S. (2004) Personal communication.

References
136
Lam, J.S., Matewish, M.J., and Poon, K.K.H. (2004) Lipopolysaccharides of Pseudomonas aeruginosa. In
Pseudomonas: Biosynthesis of Macromolecules and Molecular Metabolism. Ramos, J.L. (ed):
Springer.
Lambert, P.A. (2002) Mechanisms of antibiotic resistance in Pseudomonas aeruginosa. J. R. Soc. Med. 95
Suppl 41: 22-26.
Langsrud, S., Sundheim, G., and Holck, A.L. (2004) Cross-resistance to antibiotics of Escherichia coli
adapted to benzalkonium chloride or exposed to stress-inducers. J. Appl. Microbiol. 96: 201-208.
Lee, A., Mao, W., Warren, M.S., Mistry, A., Hoshino, K., Okumura, R., Ishida, H., and Lomovskaya, O.
(2000) Interplay between efflux pumps may provide either additive or multiplicative effects on drug
resistance. J. Bacteriol. 182: 3142-3150.
Lee, J.C., and Fialkow, P.J. (1961) Benzalkonium chloride-source of hospital infection with gram-negative
bacteria. J. Amer. Med. Ass. 177: 708-710.
Legakis, N.J., Tzouvelekis, L.S., Makris, A., and Kotsifaki, H. (1989) Outer membrane alterations in
multiresistant mutants of Pseudomonas aeruginosa selected by ciprofloxacin. Antimicrob. Agents
Chemother. 33: 124-127.
Lewis, R., (ed) (1998) Lewis' dictionary of toxicology.: CRC press.
Li, X.Z., Nikaido, H., and Poole, K. (1995) Role of mexA-mexB-oprM in antibiotic efflux in Pseudomonas
aeruginosa. Antimicrob. Agents Chemother. 39: 1948-1953.
Li, X.Z., Zhang, L., Srikumar, R., and Poole, K. (1998a) Beta-lactamase inhibitors are substrates for the
multidrug efflux pumps of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 42: 399-403.
Li, X.Z., Zhang, L., and Poole, K. (1998b) Role of the multidrug efflux systems of Pseudomonas aeruginosa
in organic solvent tolerance. J. Bacteriol. 180: 2987-2991.
Li, X.Z., Barre, N., and Poole, K. (2000) Influence of the MexA-MexB-OprM multidrug efflux system on
expression of the MexC-MexD-OprJ and MexE-MexF-OprN multidrug efflux systems in
Pseudomonas aeruginosa. J. Antimicrob. Chemother. 46: 885-893.
Li, X.Z., Poole, K., and Nikaido, H. (2003) Contributions of MexAB-OprM and an EmrE homolog to intrinsic
resistance of Pseudomonas aeruginosa to aminoglycosides and dyes. Antimicrob. Agents Chemother.
47: 27-33.
Linares, J.F., Lopez, J. A., Camafeita, E., Albar J. P., Rojo, F., and Martinez, J. L. (2005) Overexpression of
the multidrug efflux pumps MexCD-OprJ and MexEF-OprN is associated with a reduction of type
III secretion in Pseudomonas aeruginosa. J. Bacteriol. 187: 1384-1391.
Liu, P.V., Matsumoto, H., Kusama, H., and Bergan, T. (1983) Survey of heat-stable, major somatic antigens
of Pseudomonas aeruginosa. Int. J. Syst. Bacteriol. 33: 256-264.
Liu, P.V., and Wang, S. (1990) Three new major somatic antigens of Pseudomonas aeruginosa. J. Clin.
Microbiol. 28: 922-925.
Lizioli, A., Privitera, G., Alliata, E., Antonietta Banfi, E.M., Boselli, L., Panceri, M.L., Perna, M.C., Porretta,
A.D., Santini, M.G., and Carreri, V. (2003) Prevalence of nosocomial infections in Italy: result from
the Lombardy survey in 2000. J. Hosp. Infect. 54: 141-148.
Llanes, C., Hocquet, D., Vogne, C., Benali-Baitich, D., Neuwirth, C., and Plesiat, P. (2004) Clinical strains of
Pseudomonas aeruginosa overproducing MexAB-OprM and MexXY pumps simultaneously.
Antimicrob. Agents Chemother. 48: 1797-1802.
Lomovskaya, O., Warren, M.S., Lee, A., Galazzo, J., Fronko, R., Lee, M., Blais, J., Cho, D., Chamberland, S.,
Renau, T., Leger, R., Hecker, S., Watkins, W., Hoshino, K., Ishida, H., and Lee, V.J. (2001)
Identification and characterization of inhibitors of multidrug resistance efflux pumps in
Pseudomonas aeruginosa: novel agents for combination therapy. Antimicrob. Agents Chemother. 45:
105-116.

References
137
Lomovskaya, O., and Bostian, K.A. (2006) Practical applications and feasibility of efflux pump inhibitors in
the clinic--A vision for applied use. Biochem. Pharmacol. 71: 910-918.
Longbottom, C.J. (2002) Characterisation of the resistance to tea tree oil in Pseudomonas. Microbiology.
Bachelor of Science (Honours) Perth: The University of Western Australia.
Longbottom, C.J., Carson, C.F., Hammer, K.A., Mee, B.J., and Riley, T.V. (2004) Tolerance of Pseudomonas
aeruginosa to Melaleuca alternifolia (tea tree) oil is associated with the outer membrane and energy-
dependent cellular processes. J. Antimicrob. Chemother. 54: 386-392.
Loughlin, M.F., Jones, M.V., and Lambert, P.A. (2002) Pseudomonas aeruginosa cells adapted to
benzalkonium chloride show resistance to other membrane-active agents but not to clinically
relevant antibiotics. J. Antimicrob. Chemother. 49: 631-639.
Lowbury, E.J. (1951) Contamination of cetrimide and other fluids with Pseudomonas pyocyanea. Br. J. Ind.
Med. 8: 22-25.
Lynch, A.S. (2006) Efflux systems in bacterial pathogens: an opportunity for therapeutic intervention? An
industry view. Biochem. Pharmacol. 71: 949-956.
Makin, S.A., and Beveridge, T.J. (1996) The influence of A-band and B-band lipopolysaccharide on the
surface characteristics and adhesion of Pseudomonas aeruginosa to surfaces. Microbiol. 142: 299-
307.
Mann, C.M., Cox, S.D., and Markham, J.L. (2000) The outer membrane of Pseudomonas aeruginosa NCTC
6749 contributes to its tolerance to the essential oil of Melaleuca alternifolia (tea tree oil). Lett. Appl.
Microbiol. 30: 294-297.
Martinez, M.B., Flickinger, M.C., and Nelsestuen, G.L. (1996) Accurate kinetic modeling of alkaline
phosphatase in the Escherichia coli periplasm: implications for enzyme properties and substrate
diffusion. Biochem. 35: 1179-1186.
Masduki, A., Nakamura, J., Ohga, T., Umezaki, R., Kato, J., and Ohtake, H. (1995) Isolation and
characterization of chemotaxis mutants and genes of Pseudomonas aeruginosa. J. Bacteriol. 177:
948-952.
Masecar, B.L., Celesk, R.A., and Robillard, N.J. (1990) Analysis of acquired ciprofloxacin resistance in a
clinical strain of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 34: 281-286.
Maseda, H., Yoneyama, H., and Nakae, T. (2000) Assignment of the substrate-selective subunits of the
MexEF-OprN multidrug efflux pump of Pseudomonas aeruginosa. Antimicrob. Agents Chemother.
44: 658-664.
Maseda, H., Sawada, I., Saito, K., Uchiyama, H., Nakae, T. and Nomura, N. (2004) Enhancement of the
mexAB-oprM efflux pump expression by a quorum-sensing autoinducer and its cancellation by a
regulator, MexT, of the mexEF-oprN efflux pump operon in Pseudomonas aeruginosa. Antimicrob.
Agents Chemother. 48: 1320-1328.
Masoud, H., Sadovskaya, I., de Kievit, T., Altman, E., Richards, J., and Lam, J. (1995) Structural elucidation
of the lipopolysaccharide core region of the O- chain-deficient mutant strain A28 from Pseudomonas
aeruginosa serotype 06 (International Antigenic Typing Scheme). J. Bacteriol. 177: 6718-6726.
Masuda, N., Sakagawa, E., Ohya, S., Gotoh, N., Tsujimoto, H., and Nishino, T. (2000a) Substrate specificities
of MexAB-OprM, MexCD-OprJ, and MexXY-oprM efflux pumps in Pseudomonas aeruginosa.
Antimicrob. Agents Chemother. 44: 3322-3327.
Masuda, N., Sakagawa, E., Ohya, S., Gotoh, N., Tsujimoto, H., and Nishino, T. (2000b) Contribution of the
MexX-MexY-OprM efflux system to intrinsic resistance in Pseudomonas aeruginosa. Antimicrob.
Agents Chemother. 44: 2242-2246.
Masuda, N., Sakagawa, E., Ohya, S., Gotoh, N., and Nishino, T. (2001) Hypersusceptibility of the
Pseudomonas aeruginosa nfxB mutant to beta-lactams due to reduced expression of the ampC beta-
lactamase. Antimicrob. Agents Chemother. 45: 1284-1286.

References
138
Masuda, N., Gotoh, N., Ohya, S., and Nishino, T. (1996) Quantitative correlation between susceptibility and
OprJ production in NfxB mutants of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 40:
909-913.
Matsumoto, H., Nakazawa, T., Ohta, S., and Terawaki, Y. (1981) Chromosomal locations of catA, pobA, dcu
and chu genes in Pseudomonas aeruginosa. Genet. Res. 38: 251-266.
McMahon, M.A., Blair, I.S., Moore, J.E., and McDowell, D.A. (2007) Habituation to sub-lethal
concentrations of tea tree oil (Melaleuca alternifolia) is associated with reduced susceptibility to
antibiotics in human pathogens. J. Antimicrob. Chemother. 59: 125-127.
Mesaros, N., Glupczynski, Y., Avrain, L., Caceres, N.E., Tulkens, P.M., and Van Bambeke, F. (2007) A
combined phenotypic and genotypic method for the detection of Mex efflux pumps in Pseudomonas
aeruginosa. J. Antimicrob. Chemother. 59: 378-386.
Micol, V., Mateo, C.R., Shapiro, S., Aranda, F.J., and Villalain, J. (2001) Effects of (+)-totarol, a diterpenoid
antibacterial agent, on phospholipid model membranes. Biochim. Biophys. Acta 1511: 281-290.
Mikus, J., Harkenthal, M., Steverding, D., and Reichling, J. (2000) In vitro effect of essential oils and isolated
mono- and sesquiterpenes on Leishmania major and Trypanosoma brucei. Planta Medica 66: 366-
368.
Miller, W.L., and Lam, J.S. (2008) Molecular Biology of Cell-Surface Polysaccharides in Pseudomonas
aeruginosa: from Gene to Protein Function. In Pseudomonas: Genomics and Molecular Biology.
Cornelis, P. (ed): Horizon Scientific Press.
Mima, T., Sekiya, H., Mizushima, T., Kuroda, T., and Tsuchiya, T. (2005) Gene cloning and properties of the
RND-type multidrug efflux pumps MexPQ-OpmE and MexMN-OprM from Pseudomonas
aeruginosa. Microbiol. Immunol. 49: 999-1002.
Minami, M., Kita, M., Nakaya, T., Yamamoto, T., Kuriyama, H., and Imanishi, J. (2003) The inhibitory effect
of essential oils on herpes simplex virus type-1 replication in vitro. Microbiol. Immunol. 47: 681-
684.
Mobley, H.L., and Rosen, B.P. (1982) Energetics of plasmid-mediated arsenate resistance in Escherichia coli.
Proc. Natl. Acad. Sci. U S A 79: 6119-6122.
Moken, M.C., McMurry, L.M., and Levy, S.B. (1997) Selection of multiple-antibiotic-resistant (mar) mutants
of Escherichia coli by using the disinfectant pine oil: roles of the mar and acrAB loci. Antimicrob.
Agents Chemother. 41: 2770-2772.
Moller, A.K., Leatham, M.P., Conway, T., Nuijten, P.J., de Haan, L.A., Krogfelt, K.A., and Cohen, P.S.
(2003) An Escherichia coli MG1655 lipopolysaccharide deep-rough core mutant grows and survives
in mouse cecal mucus but fails to colonize the mouse large intestine. Infect. Immun. 71: 2142-2152.
Montie, T.C., Doyle-Huntzinger, D., Craven, R.C., and Holder, I.A. (1982) Loss of virulence associated with
absence of flagellum in an isogenic mutant of Pseudomonas aeruginosa in the burned-mouse model.
Infect. Immun. 38: 1296-1298.
Morita, Y., Murata, T., Mima, T., Shiota, S., Kuroda, T., Mizushima, T., Gotoh, N., Nishino, T. and Tsuchiya,
T. (2003) Induction of mexCD-oprJ operon for multidrug efflux pump by disinfectants in wild-type
Pseudomonas aeruginosa. J. Antimicrob. Chemother. 51: 991-994.
Muller, J.F., Stevens, A.M., Craig, J., and Love, N.G. (2007) Transcriptome analysis reveals that multidrug
efflux genes are upregulated to protect Pseudomonas aeruginosa from pentachlorophenol stress.
Appl. Environ. Microbiol. 73: 4550-4558.
Munoz, R., Chambaud, M., Bordel, S., and Villaverde, S. (2008) A systematic selection of the non-aqueous
phase in a bacterial two liquid phase bioreactor treating alpha-pinene. Appl. Microbiol. Biotechnol.
79: 33-41.
Murray, N.E., Brammar, W.J., and Murray, K. (1977) Lambdoid phages that simplify the recovery of in vitro
recombinants. Mol. Gen. Genet. 150: 53-61.

References
139
Navon-Venezia, S., Ben-Ami, R., and Carmeli, Y. (2005) Update on Pseudomonas aeruginosa and
Acinetobacter baumannii infections in the healthcare setting. Curr. Opin. Infect. Dis. 18: 306-313.
Nelson, R.R. (2000) Selection of resistance to the essential oil of Melaleuca alternifolia in Staphylococcus
aureus. J Antimicrob Chemother 45: 549-550.
Nenoff, P., Haustein, U.F., and Brandt, W. (1996) Antifungal activity of the essential oil of Melaleuca
alternifolia (tea tree oil) against pathogenic fungi in vitro. Skin Pharmacol. 9: 388-394.
Nicas, T.I. (1999) Biochemical mode of action of vancomycin and history of global clinical vancomycin use.
In Intersci. Conf. Antimicrob. Agents Chemother.
Nikaido, H., and Vaara, M. (1985) Molecular basis of bacterial outer membrane permeability. Microbiol. Rev.
49: 1-32.
Nikaido, H. (2003) Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol.
Rev. 67: 593-656.
Nikaido, H. (2005) Chapter 19: Role, structure, and function of multidrug efflux pumps in Gram negative
bacteria. In Frontiers in antimicrobial resistance: a tribute to Stuart B Levy. White, D.G., Alekshun,
M.N., McDermott, G. and Levy, S.B. (eds): ASM press.
Nikaido, H., Hancock, REW (1986) Outer membrane permeability in Pseudomonas aeruginosa. In The
Bacteria. Sokatch, J. (ed). Orlando: Academic press, pp. 145-193.
NNIS (2004) National Nosocomial Infections Surveillance (NNIS) System Report, data summary from
January 1992 through June 2004, issued October 2004. Am. J. Infect. Control 32: 470-485.
Oh, E., Koh, S., Kim, E., Ahn, Y., and So, J. (2003) Plant terpenes enhance survivability of polychlorinated
biphenyl (PCB) degrading Pseudomonas pseudoalcaligenes KF707 labeled with gfp in microcosms
contaminated with PCB. J. Microbiol. Biotechnol. 13: 463-468.
Okii, M., Iyobe, S., and Mitsuhashi, S. (1983) Mapping of the gene specifying aminoglycoside 3'-
phosphotransferase II on the Pseudomonas aeruginosa chromosome. J Bacteriol 155: 643-649.
Okusu, H., Ma, D., and Nikaido, H. (1996) AcrAB efflux pump plays a major role in the antibiotic resistance
phenotype of Escherichia coli multiple-antibiotic-resistance (Mar) mutants. J. Bacteriol. 178: 306-
308.
Oliver, A., Levin, B.R., Juan, C., Baquero, F., and Blazquez, J. (2004) Hypermutation and the preexistence of
antibiotic-resistant Pseudomonas aeruginosa mutants: implications for susceptibility testing and
treatment of chronic infections. Antimicrob. Agents Chemother. 48: 4226-4233.
Pages, J.M., Masi, M., and Barbe, J. (2005) Inhibitors of efflux pumps in Gram-negative bacteria. Trends
Mol. Med. 11: 382-389.
Pages, J.M. (2006) Role of bacterial porins in antibiotic susceptibility of Gram-negative bacteria. In Bacterial
and eukaryotic porins: structure, function and mechanism. Benz, R. (ed). Weinheim: Wiley VCH,
pp. 41-59.
Palleroni, N.J. (1975) General properties and taxonomy of the genus Pseudomonas. In Genetics and
biochemistry of Pseudomonas. Clark, P.H. and Richmond, M.H. (eds). London: John Wiley and
Sons, pp. 1-36.
Pandza, S., Baetens, M., Park, C.H., Au, T., Keyhan, M., and Matin, A. (2000) The G-protein FlhF has a role
in polar flagellar placement and general stress response induction in Pseudomonas putida. Mol.
Microbiol. 36: 414-423.
Pao, S.S., Paulsen, I.T., and Jr, M.H.S. (1998) Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 62: 1-
34.
Papadopoulos, C.J., Carson, C.F., Hammer, K.A., and Riley, T.V. (2006) Susceptibility of pseudomonads to
Melaleuca alternifolia (tea tree) oil and components. J. Antimicrob. Chemother. 58: 449-451.

References
140
Parker, C.T., Kloser, A.W., Schnaitman, C.A., Stein, M.A., Gottesman, S., and Gibson, B.W. (1992) Role of
the rfaG and rfaP genes in determining the lipopolysaccharide core structure and cell surface
properties of Escherichia coli K-12. J. Bacteriol. 174: 2525-2538.
Parkinson, J.S., Parker, S.R., Talbert, P.B., and Houts, S.E. (1983) Interactions between chemotaxis genes and
flagellar genes in Escherichia coli. J. Bacteriol. 155: 265-274.
Parvatiyar, K., Alsabbagh, E.M., Ochsner, U.A., Stegemeyer, M.A., Smulian, A.G., Hwang, S.H., Jackson,
C.R., McDermott, T.R., and Hassett, D.J. (2005) Global analysis of cellular factors and responses
involved in Pseudomonas aeruginosa resistance to arsenite. J. Bacteriol. 187: 4853-4864.
Patel, P.C., Goulhen, F., Boothman, C., Gault, A.G., Charnock, J.M., Kalia, K., and Lloyd, J.R. (2007)
Arsenate detoxification in a Pseudomonad hypertolerant to arsenic. Arch. Microbiol. 187: 171-183.
Paulsen, I.T., Skurray, R.A., Tam, R., Saier Jr, M.H., Turner, R.J., Weiner, J.H., Goldberg, E. B. and Grinius,
L.L. (1996) The SMR family: a novel family of multidrug efflux proteins involved with the efflux of
lipophilic drugs. Mol. Microbiol. 19: 1167-1175.
Paumgartten, F.J., De-Carvalho, R.R., Souza, C.A., Madi, K., and Chahoud, I. (1998) Study of the effects of
beta-myrcene on rat fertility and general reproductive performance. Braz. J. Med. Biol. Res. 31: 955-
965.
Pearson, J.P., Van Delden, C., and Iglewski, B.H. (1999) Active efflux and diffusion are involved in transport
of Pseudomonas aeruginosa cell-to-cell signals. J. Bacteriol. 181: 1203-1210.
Pease, A.J., Roa, B.R., Luo, W., and Winkler, M.E. (2002) Positive growth rate-dependent regulation of the
pdxA, ksgA, and pdxB genes of Escherichia coli K-12. J. Bacteriol. 184: 1359-1369.
Penfold, A.R., and Grant, R. (1925) The germicidal values of some Australian essential oils and their pure
constituents, together with those for some essential oil isolates, and synthetics, part III. Journal of the
Royal Society of New South Wales 59: 346-349.
Penfold, A.R. (1929) Notes on products. The perfumery and essential oil record: 155-156.
Piddock, L.J. (2006) Clinically relevant chromosomally encoded multidrug resistance efflux pumps in
bacteria. Clin. Microbiol. Rev. 19: 382-402.
Pinkart, H., Wolfram, J. W., Rogers, R., and White, D. (1996) Cell envelope changes in solvent-tolerant and
solvent-sensitive Pseudomonas putida strains following exposure to o- xylene. Appl. Environ.
Microbiol. 62: 1129-1132.
Pittet, D., Harbarth, S., Ruef, C., Francioli, P., Sudre, P., Petignat, C., Trampuz, A., and Widmer, A. (1999)
Prevalence and risk factors for nosocomial infections in four university hospitals in Switzerland.
Infect. Control Hosp. Epidemiol. 20: 37-42.
Plesiat, P., Aires, J.R., Godard, C., and Kohler, T. (1997) Use of steroids to monitor alterations in the outer
membrane of Pseudomonas aeruginosa. J. Bacteriol. 179: 7004-7010.
Poole, K., Gotoh, N., Tsujimoto, H., Zhao, Q., Wada, A., Yamasaki, T., Neshat, S., Yamagishi, J., Li, X.Z.,
and Nishino, T. (1996) Overexpression of the mexC-mexD-oprJ efflux operon in nfxB-type
multidrug-resistant strains of Pseudomonas aeruginosa. Mol. Microbiol. 21: 713-724.
Poole, K. (2001) Multidrug efflux pumps and antimicrobial resistance in Pseudomonas aeruginosa and
related organisms. J. Mol. Microbiol. Biotechnol. 3: 255-264.
Poole, K., and McKay, G.A. (2003) Iron acquisition and its control in Pseudomonas aeruginosa: many roads
lead to Rome. Front. Biosci. 8: d661-686.
Poole, K. (2007) Efflux pumps as antimicrobial resistance mechanisms. Ann. Med. 39: 162-176.
Quale, J., Bratu, S., Gupta, J., and Landman, D. (2006) Interplay of efflux system, ampC, and oprD
expression in carbapenem resistance of Pseudomonas aeruginosa clinical isolates. Antimicrob.
Agents Chemother. 50: 1633-1641.

References
141
Rahim, R., Burrows, L.L., Monteiro, M.A., Perry, M.B., and Lam, J.S. (2000) Involvement of the rml locus in
core oligosaccharide and O polysaccharide assembly in Pseudomonas aeruginosa. Microbiol. 146:
2803-2814.
Ramjeet, M., Deslandes, V., St. Michael, F., Cox, A.D., Kobisch, M., Gottschalk, M., and Jacques, M. (2005)
Truncation of the lipopolysaccharide outer core affects susceptibility to antimicrobial peptides and
virulence of Actinobacillus pleuropneumoniae Serotype 1. J. Biol. Chem. 280: 39104-39114.
Randall, L.P., and Woodward, M.J. (2002) The multiple antibiotic resistance (mar) locus and its significance.
Res. Vet. Sci. 72: 87-93.
Rawat, M., Heys, J., and Av-Gay, Y. (2003) Identification and characterization of a diamide sensitive mutant
of Mycobacterium smegmatis. FEMS Microbiol. Lett. 220: 161-169.
Renau, T.E., Leger, R., Flamme, E.M., Sangalang, J., She, M.W., Yen, R., Gannon, C.L., Griffith, D.,
Chamberland, S., Lomovskaya, O., Hecker, S.J., Lee, V.J., Ohta, T., and Nakayama, K. (1999)
Inhibitors of efflux pumps in Pseudomonas aeruginosa potentiate the activity of the fluoroquinolone
antibacterial levofloxacin. J. Med. Chem. 42: 4928-4931.
Rocchetta, H.L., Burrows, L.L., Pacan, J.C., and Lam, J.S. (1998) Three rhamnosyltransferases responsible
for assembly of the A-band D-rhamnan polysaccharide in Pseudomonas aeruginosa: a fourth
transferase, WbpL, is required for the initiation of both A-band and B-band lipopolysaccharide
synthesis. Mol. Microbiol. 28: 1103-1119.
Rosenberg, M., Gutnick, G., and Rosenberg, E. (1980) Adherence of bacteria to hydrocarbons: A simple
method for measuring cell-surface hydrophobicity. FEMS Micro. Lett. 9: 29-33.
Rouviere, P.E., and Gross, C.A. (1996) SurA, a periplasmic protein with peptidyl-prolyl isomerase activity,
participates in the assembly of outer membrane porins. Genes Dev. 10: 3170-3182.
Rowe, J.S. (1999) Formulating for effect. In Tea tree: the genus Melaleuca. Southwell, I. and Lowe, R. (eds).
Amsterdam, The Netherlands: Harwood Academic Publishers, pp. 207–212.
Rowe, P.S., and Meadow, P.M. (1983) Structure of the core oligosaccharide from the lipopolysaccharide of
Pseudomonas aeruginosa PAC1R and its defective mutants. Eur. J. Biochem. 132: 329-337.
Russell, M. (1999) Toxicology of tea tree oil. In Tea tree: the genus Melaleuca. Southwell, I. and Lowe, R.
(eds). Amsterdam, The Netherlands: Harwood Academic Publishers, pp. 191-201.
Ryan, K.J. (2004) Sterilization and disinfection. In Sherris medical microbiology: an introduction to
infectious disease. Ryan, K.J. and Ray, C.G. (eds): McGraw-Hill professional.
Sadovskaya, I., Brisson, J.-R., Lam, J.S., Richards, J.C., and Altman, E. (1998) Structural elucidation of the
lipopolysaccharide core regions of the wild-type strain PAO1 and O-chain-deficient mutant strains
AK1401 and AK1012 from Pseudomonas aeruginosa serotype O5. Eur. J. Biochem. 255: 673-684.
Sahalan, A.Z., and Dixon, R.A. (2008) Role of the cell envelope in the antibacterial activities of polymyxin B
and polymyxin B nonapeptide against Escherichia coli. Int. J. Antimicrob. Agents 31: 224-227.
Saier Jr, M.H., Tam. R., Reizer, A., and Reizer, J. (1994) Two novel families of bacterial membrane proteins
concerned with nodulation, cell division and transport. Mol. Microbiol. 11: 841-847.
Saito, K., Yoneyama, H., and Nakae, T. (1999) nalB-type mutations causing the overexpression of the
MexAB-OprM efflux pump are located in the mexR gene of the Pseudomonas aeruginosa
chromosome. FEMS Microbiol. Lett. 179: 67-72.
Sardessai, Y., and Bhosle, S. (2002) Tolerance of bacteria to organic solvents. Res. Microbiol. 153: 263-268.
Satchell, A.C., Saurajen, A., Bell, C., and Barnetson, R.S. (2002a) Treatment of dandruff with 5% tea tree oil
shampoo. J. Amer. Acad. Dermatol. 47: 852-855.
Satchell, A.C., Saurajen, A., Bell, C., and Barnetson, R.S. (2002b) Treatment of interdigital tinea pedis with
25% and 50% tea tree oil solution: a randomized, placebo-controlled, blinded study. Australas. J.
Dermatol. 43: 175-178.

References
142
Sawada, I., Maseda, H., Nakae, T., Uchiyama, H., and Nomura, N. (2004) A quorum-sensing autoinducer
enhances the mexAB-oprM efflux-pump expression without the MexR-mediated regulation in
Pseudomonas aeruginosa. Microbiol. Immunol. 48: 435-439.
Schnaitman, C.A., and Klena, J.D. (1993) Genetics of lipopolysaccharide biosynthesis in enteric bacteria.
Microbiol. Rev. 57: 655-682.
Schnitzler, P., Schön, K., and Reichling, J. (2001) Antiviral activity of Australian tea tree oil and eucalyptus
oil against herpes simplex virus in cell culture. Pharmazie 56: 343-347.
Schwarz, S., and White, D.G. (2005) Chapter 9: Phenicol resistance. In Frontiers in antimicrobial resistance:
a tribute to Stuart B Levy. White, D.G., Alekshun, M.N., McDermott, P.F. and Levy, S.B. (eds):
ASM press.
Schweizer, H.P. (1998) Intrinsic resistance to inhibitors of fatty acid biosynthesis in Pseudomonas aeruginosa
is due to efflux: application of a novel technique for generation of unmarked chromosomal mutations
for the study of efflux systems. Antimicrob. Agents Chemother. 42: 394-398.
Seawright, A. (1993) Tea tree oil poisoning - Comment. Med. J. Aust. 159: 831.
Segura, A., Duque, E., Hurtado, A., and Ramos, J.L. (2001) Mutations in genes involved in the flagellar
export apparatus of the solvent-tolerant Pseudomonas putida DOT-T1E strain impair motility and
lead to hypersensitivity to toluene shocks. J. Bacteriol. 183: 4127-4133.
Selvaag, E., Eriksen, B., and Thune, P. (1994) Contact allergy due to tea tree oil and cross-sensitization to
colophony. Contact Derm. 31: 124-125.
Shapira, R., and Mimran, E. (2007) Isolation and characterization of Escherichia coli mutants exhibiting
altered response to thymol. Microb. Drug Resist. 13: 157-165.
Shearer, B.G., and Legakis, N.J. (1985) Pseudomonas aeruginosa: evidence for the involvement of
lipopolysaccharide in determining outer membrane permeability to carbenicillin and gentamicin. J.
Infect. Dis. 152: 351-355.
Silver, S., and Wendt, L. (1967) Mechanism of action of phenethyl alcohol: breakdown of the cellular
permeability barrier. J. Bacteriol. 93: 560-566.
Silversmith, R.E. (2005) High mobility of carboxyl-terminal region of bacterial chemotaxis phosphatase CheZ
Is diminished upon binding divalent cation or CheY-P substrate. Biochem. 44: 7768-7776.
Sklar, J.G., Wu, T., Kahne, D., and Silhavy, T.J. (2007) Defining the roles of the periplasmic chaperones
SurA, Skp, and DegP in Escherichia coli. Genes Dev. 21: 2473-2484.
Smania, A.M., Segura, I., Pezza, R.J., Becerra, C., Albesa, I., and Argarana, C.E. (2004) Emergence of
phenotypic variants upon mismatch repair disruption in Pseudomonas aeruginosa. Microbiol. 150:
1327-1338.
Smith, A.W., and Iglewski, B. H (1989) Transformation of Pseudomonas aeruginosa by electroporation.
Nucl. Acids Res. 17: 10509.
Smith, E.C., Kaatz, G.W., Seo, S.M., Wareham, N., Williamson, E.M., and Gibbons, S. (2007) The phenolic
diterpene totarol inhibits multidrug efflux pump activity in Staphylococcus aureus. Antimicrob.
Agents Chemother.
Soares, T.A., Straatsma, T.P., and Lins, R.D. (2008) Influence of the B-band O-antigen chain in the structure
and electrostatics of the lipopolysaccharide membrane of Pseudomonas aeruginosa. J. Braz. Chem.
Soc. 19: 312-320.
Sobel, M.L., McKay, G.A., and Poole, K. (2003) Contribution of the MexXY multidrug transporter to
aminoglycoside resistance in Pseudomonas aeruginosa clinical isolates. Antimicrob. Agents
Chemother. 47: 3202-3207.

References
143
Sobel, M.L., Hocquet, D., Cao, L., Plesiat, P, and Poole, K. (2005) Mutations in PA3574 (nalD) lead to
increased MexAB-OprM expression and multidrug resistance in laboratory and clinical isolates of
Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 49: 1782-1786.
Soscia, C., Hachani, A., Bernadac, A., Filloux, A., and Bleves, S. (2007) Cross talk between type III secretion
and flagellar assembly systems in Pseudomonas aeruginosa. J. Bacteriol. 189: 3124-3132.
Southwell, I.A., Freeman, S., and Rubel, D. (1997) Skin irritancy of tea tree oil. J. Ess. Oil Res. 9: 47-52.
Spangenberg, C., Heuer, T., Burger, C., and Tummler, B. (1996) Genetic diversity of flagellins of
Pseudomonas aeruginosa. FEBS Lett. 396: 213-217.
Srikumar, R., Li, X.Z., and Poole, K. (1997) Inner membrane efflux components are responsible for beta-
lactam specificity of multidrug efflux pumps in Pseudomonas aeruginosa. J. Bacteriol. 179: 7875-
7881.
Srikumar, R., Kon, T., Gotoh, N., and Poole, K. (1998) Expression of Pseudomonas aeruginosa multidrug
efflux pumps MexA-MexB-OprM and MexC-MexD-OprJ in a multidrug-sensitive Escherichia coli
strain. Antimicrob. Agents Chemother. 42: 65-71.
Stavri, M., Piddock, L.J.V., and Gibbons, S. (2007) Bacterial efflux pump inhibitors from natural sources. J.
Antimicrob. Chemother.: 59: 1247-1260.
Stone, J. (2003) Antimicrobial therapy. In Fundamentals of anaesthesia. Pinnock, C.A., Lin, T. and Smith, T.
(eds): Cambridge University Press.
Stover, C.K., Pham, X.Q., Erwin, A.L., Mizoguchi, S.D., Warrener, P., Hickey, M.J., Brinkman, F.S.,
Hufnagle, W.O., Kowalik, D.J., Lagrou, M., Garber, R.L., Goltry, L., Tolentino, E., Westbrock-
Wadman, S., Yuan, Y., Brody, L.L., Coulter, S.N., Folger, K.R., Kas, A., Larbig, K., Lim, R., Smith,
K., Spencer, D., Wong, G.K., Wu, Z., Paulsen, I.T., Reizer, J., Saier, M.H., Hancock, R.E., Lory, S.,
and Olson, M.V. (2000) Complete genome sequence of Pseudomonas aeruginosa PA01, an
opportunistic pathogen. Nature. 406: 959-964.
Suller, M.T., and Russell, A.D. (2000) Triclosan and antibiotic resistance in Staphylococcus aureus. J.
Antimicrob. Chemother. 46: 11-18.
Teitzel, G.M., and Parsek, M.R. (2003) Heavy metal resistance of biofilm and planktonic Pseudomonas
aeruginosa. Appl. Environ. Microbiol. 69: 2313-2320.
Thompson, L.S., Webb, J.S., Rice, S.A., and Kjelleberg, S. (2003) The alternative sigma factor RpoN
regulates the quorum sensing gene rhlI in Pseudomonas aeruginosa. FEMS Microbiol. Lett. 220:
187-195.
Thomson, K.S., and Lister, P.D. (2004) Other beta-lactam antibiotics. In Treatment of infectious diseases.
Gorbach, S.L., Bartlett, J.G. and Blacklow, N.R. (eds): Lippincott, Williams and Wilkins.
Tormo, A., Almiron, M., and Kolter, R. (1990) surA, an Escherichia coli gene essential for survival in
stationary phase. J. Bacteriol. 172: 4339-4347.
Totten, P.A., Lara, J.C., and Lory, S. (1990) The rpoN gene product of Pseudomonas aeruginosa is required
for expression of diverse genes, including the flagellin gene. J. Bacteriol. 172: 389-396.
Ultee, A., Kets, E.P., Alberda, M., Hoekstra, F.A., and Smid, E.J. (2000) Adaptation of the food-borne
pathogen Bacillus cereus to carvacrol. Arch. Microbiol. 174: 233-238.
Vaara, M. (1992) Agents that increase the permeability of the outer membrane. Microbiol. Rev. 56: 395-411.
Veien, N.K., Rosner, K., and Skovgaard, G.L. (2004) Is tea tree oil an important contact allergen? Cont.
Derm. 50: 378-379.
Viducic, D., Ono, T., Murakami, K., Katakami, M., Susilowati, H., and Miyake, Y. (2007) rpoN gene of
Pseudomonas aeruginosa alters its susceptibility to quinolones and carbapenems. Antimicrob. Agents
Chemother. 51: 1455-1462.

References
144
Viollon, C., Mandin, D., and Chaumont, J.P. (1996) Activities antagonistes, in vitro, de quelques huiles
essentielles et de composes naturels volatils viv a vis de la croissance de Trichomonas vaginalis.
Fitoterapia 63: 279-281.
Walsh, A.G., Matewish, M.J., Burrows, L.L., Monteiro, M.A., Perry, M.B., and Lam, J.S. (2000)
Lipopolysaccharide core phosphates are required for viability and intrinsic drug resistance in
Pseudomonas aeruginosa. Mol. Microbiol. 35: 718-727.
Walton, S.F., McKinnon, M., Pizzutto, S., Dougall, A., Williams, E., and Currie, B.J. (2004) Acaricidal
activity of Melaleuca alternifolia (tea tree) oil: in vitro sensitivity of Sarcoptes scabiei var hominis
to terpinen-4-ol. Arch. Dermatol. 140: 563-566.
Wang, Y., Ha, U., Zeng, L., and Jin, S. (2003) Regulation of membrane permeability by a two-component
regulatory system in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 47: 95-101.
Warren, M.S., Lee, J.C., Lee, A., Hoshino, K., Ishida, H., and Lomovskaya, O. (2000) An inhibitor of the
Mex pumps from Pseudomonas aeruginosa is also a substrate of these pumps. In Intersci. Conf.
Antimicrob. Agents Chemother. 40: 207.
Weber, F.J., and de Bont, J.A. (1996) Adaptation mechanisms of microorganisms to the toxic effects of
organic solvents on membranes. Biochim. Biophys. Acta 1286: 225-245.
Werner, J., Augustus, A.M., and Misra, R. (2003) Assembly of TolC, a structurally unique and
multifunctional outer membrane protein of Escherichia coli K-12. J. Bacteriol. 185: 6540-6547.
Winder, C.L., Al-Adham, I.S., Abdel Malek, S.M., Buultjens, T.E., Horrocks, A.J., and Collier, P.J. (2000)
Outer membrane protein shifts in biocide-resistant Pseudomonas aeruginosa PAO1. J. Appl.
Microbiol. 89: 289-295.
Winsor, G.L., Lo, R., Sui, S.J., Ung, K.S., Huang, S., Cheng, D., Ching, W.K., Hancock, R.E., and Brinkman,
F.S. (2005) Pseudomonas aeruginosa Genome database and PseudoCAP: facilitating community-
based, continually updated, genome annotation. Nucleic Acids Res. 33(Database issue): D338-343.
Woods, D.E., Lam, J.S., Paranchych, W., Speert, D.P., Campbell, M., and Godfrey, A.J. (1997) Correlation of
Pseudomonas aeruginosa virulence factors from clinical and environmental isolates with
pathogenicity in the neutropenic mouse. Can. J. Microbiol. 43: 541-551.
Wyborn, N.R., Clark, A., Roberts, R.E., Jamieson, S.J., Tzokov, S., Bullough, P.A., Stillman, T.J., Artymiuk,
P.J., Galen, J.E., Zhao, L., Levine, M.M., and Green, J. (2004) Properties of haemolysin E (HlyE)
from a pathogenic Escherichia coli avian isolate and studies of HlyE export. Microbiol. 150: 1495-
1505.
Yokota, S., and Fujii, N. (2007) Contributions of the lipopolysaccharide outer core oligosaccharide region on
the cell surface properties of Pseudomonas aeruginosa. Comp. Immunol. Microbiol. Infect. Dis. 30:
97-109.
Zhang, L., Li, X.-Z., and Poole, K. (2001) Fluoroquinolone susceptibilities of efflux-mediated multidrug-
resistant Pseudomonas aeruginosa, Stenotrophomonas maltophilia and Burkholderia cepacia. J.
Antimicrob. Chemother. 48: 549-552.
Zhang, S., McCormack, F.X., Levesque, R.C., O'Toole, G.A., and Lau, G.W. (2007) The flagellum of
Pseudomonas aeruginosa is required for resistance to clearance by surfactant protein A. PLoS ONE
2: e564.












![Teebaumöl – Melaleuca alternifolia · 2018-02-15 · Gesundheitsbrief 43 [23] Ihre Gesundheit ist unser Anliegen... seit 30 Jahren Seite 1 von 2 Teebaumöl – Melaleuca alternifolia](https://img.pdfslide.net/doc/110x75/5f4bd57a57b87f3abd371c1b/teebauml-a-melaleuca-alternifolia-2018-02-15-gesundheitsbrief-43-23-ihre.jpg)
















