Embed Size (px)
Citation preview

Microbial Colonization of an In SituSediment Cap and Correlation toStratified Redox ZonesD A V I D W . H I M M E L H E B E R , † , ⊥
S A R A H . T H O M A S , † , ‡
F R A N K E . L O F F L E R , † , ‡
M A R T I A L T A I L L E F E R T , § A N DJ O S E P H B . H U G H E S * , † , |
School of Civil and Environmental Engineering, GeorgiaInstitute of Technology, Atlanta, Georgia 30332, School ofBiology, Georgia Institute of Technology, Atlanta, Georgia30332, School of Earth and Atmospheric Sciences, GeorgiaInstitute of Technology, Atlanta, Georgia 30322, and School ofMaterial Science and Engineering, Georgia Institute ofTechnology, Atlanta, Georgia 30332
Received July 2, 2008. Revised manuscript received October21, 2008. Accepted October 23, 2008.
In situ capping is a management technique for contaminatedsediments involving the placement of clean material at thesediment-water interface. This work combined porewatergeochemicalprofilingwithquantitativemicrobialdatato investigatethe intrinsic microbial colonization of a sand cap. Geochemicalcharacterization using voltammetric microelectrodes indicatedverticalstratificationofbiogeochemicalprocesseswithinacappedsediment column. Following dissection of the column, quantitativereal-time PCR (qPCR) enumerated microbial populationswithineachdiscreteredoxzoneandwasaccompaniedbyterminal-restriction fragment length polymorphism (T-RFLP) to elucidategeneral community shifts. Bacteria and Archaea were presentwithin the cap according to qPCR, with higher concentrationsgenerally observed in the underlying sediment. Iron-reducingpopulations were detected and quantified using newly designedqPCR primer pairs for Anaeromyxobacter spp. and Shewanellaspp. and published primer sets for δ-Proteobacteria andGeobacteracea. Results confirmed geochemical measurementsindicating that microbial Fe(III) reduction was a major processin the overlying cap. Genes encoding microbial sulfate reduction(dsrA) and methanogenesis (mcrA) were also present withinthe cap but were more prevalent in the sediment. Canonicalcorrespondence analysis of terminal-restriction fragment lengthpolymorphism (T-RFLP) patterns verified that spatial changesin bacterial community composition were significantly correlatedtodepthandFe2+andMn2+concentrationgradients.Cumulatively,results demonstrate that microorganisms indigenous to aquaticsedimentscolonizedtheoverlyingcaptoformcomplexcommunitiesmirroring redox stratification. Implications of capping forbiogeochemical cycling, contaminant fate and transport, andremedial design are discussed.
IntroductionContaminated aquatic sediments have adverse effects onaquatic life and limit recreational and economic uses,especially in urban and industrial settings (1). In situ cappinghas emerged as an option for managing contaminatedsediments and involves placing a layer of clean fill materialat the sediment-water interface to prevent contaminantrelease and contact with benthic macrofauna and overlyingsurface water. Traditionally, clean sand has been employedfor capping and can considerably delay contaminant break-through when diffusive transport processes dominate (2).Recent efforts have focused on active capping technologies,in which sediment caps are augmented with constituents tosequester or transform contaminants during transportthrough the cap (3-5). A recent study (6) demonstrated thatemplacing an in situ cap at the sediment-water interfaceinduces an upward, vertical shift of geochemical species intothe overlying cap while simultaneously conserving redoxstratification. One issue that has not been addressed,however, is if active microbial populations also shift fromthe sediment to colonize the overlying cap.
Intrinsic microbial colonization of an in situ sedimentcap will directly impact biogeochemical cycling and affectcontaminant fate and transport. Biogeochemical cycling atthe sediment-water interface of aquatic sediments can becomplex and subject to multiple site-specific variables (e.g.,organic carbon input, hydrodynamic flow patterns). Undis-turbed sediments below oxic surface waters typically possessan aerobic layer extending millimeters to centimeters intothe sediment bed (7) inhabited by an abundant and diversecommunity of heterotrophic microorganisms due to thepresence of oxygen and accessible organic matter (8). Beneaththis aerobic layer a gradient of anaerobic biogeochemicalprocesses exist, stratified according to the thermodynamicyields of their respective terminal electron accepting pro-cesses (9). Microbial composition and functionality con-tribute to this redox stratification and influence contaminantbioattenuation mechanisms within sediments. For example,metal sequestration (10) and the microbial transformationsof petroleum hydrocarbons (11), chlorinated solvents (12),polychlorinated biphenyls (PCBs) (13), and polycyclic aro-matic hydrocarbons (PAHs) (14) have all been documentedwithin aquatic sediments under specific redox conditions.
Studies described herein assess the ability and degree towhich microorganisms indigenous to sediments inhabit andcolonize an overlying sand cap. Spatial distributions of targetpopulations implicated in metal reduction, sulfate reduction,and methanogenesis are examined using quantitative real-time PCR (qPCR) and compared against porewater geochem-ical profiles. Bacterial community composition is monitoredwith terminal-restriction fragment length polymorphism (T-RFLP) analysis. Results from the qPCR and T-RFLP studies,coupled with high-resolution geochemical characterization,provide further insight into biogeochemical processes andcontaminant fate following in situ capping of aquaticsediments.
Materials and MethodsCapped Sediment Column. A laboratory-scale sedimentcolumn was constructed to monitor biogeochemical changesinduced by capping with a clean sand. Details of sedimentcolumn construction have been reported previously (6).Briefly, wet sediment from the Anacostia River, Washington,DC containing PCBs, PAHs, metals, and organic pesticides(3) was packed to a depth of 7 cm in a glass chromatography
* Corresponding author phone: (404) 894-2201; fax: (404) 894-2278; e-mail: [email protected].
† School of Civil and Environmental Engineering.‡ School of Biology.§ School of Earth and Atmospheric Sciences.| School of Material Science and Engineering.⊥ Present Address: Geosyntec Consultants, Columbia, MD 21046.
Environ. Sci. Technol. 2009, 43, 66–74
66 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 43, NO. 1, 2009 10.1021/es801834e CCC: $40.75 2009 American Chemical SocietyPublished on Web 11/25/2008

column (20 cm long, 5.0 cm i.d.; Spectrum Chromatography)fitted with a Teflon bottom end plate. The sediment wasthen capped with 7 cm of dry, autoclaved ASTM C-33 concretesand (U.S. Silica). Overlying water was maintained at 4 cmabove the cap-water interface with a constant head reservoirand consisted of deionized water (>18.2 Ω) containing onlysodium chloride (1.26 mM) to match the salinity of theAnacostia sediment porewater. The capped sediment columnwas incubated under aerobic conditions in the dark at roomtemperature with static (diffusive) conditions for 13 weeks,then upflow (advective) conditions for 16 weeks. Duringupflow conditions, a syringe pump provided anoxic influentsolution to the bottom of the column at a flow rate of 2.0 cm3
hr-1 (seepage velocity ) 4.4 cm day-1; Darcy velocity ) 2.4cm day-1). The influent was equivalent to the overlying watersolution but sparged for 30 min with nitrogen gas (Airgas)to remove dissolved oxygen. No substrates or growth factorswere supplied with the influent solution.
Porewater Geochemical Profiles. Voltammetric micro-electrodes were used to analyze aqueous geochemical species,with details of the measurements provided elsewhere (6).Briefly, all voltammetric measurements were performed usinga platinum counter electrode, an Ag/AgCl reference electrode,and an Au/Hg solid-state microelectrode fabricated asdescribed by Brendel and Luther (15). The Au/Hg micro-electrodes were capable of simultaneously measuring O2(aq),H2O2, Fe2+, Mn2+, S2O3
2-, ΣH2S [)H2S + HS- + S2- + S0 +Sx
2-], organic-FeIII(aq), and FexSy(aq) species (15-17). Each Au/
Hg microelectrode was calibrated for dissolved oxygen bylinear sweep voltammetry then for manganese by cathodicsquare wave voltammetry (CSWV). Linear sweep voltammetry(LSV) was exclusively used to measure dissolved oxygen (MDL∼4 µM), while cathodic square wave voltammetry (CSWV)was used to quantify Mn2+ (MDL ∼15 µM), Fe2+ (MDL ∼25µM), and ΣH2S (MDL ∼0.2 µM) Concentrations of Fe2+andΣH2S were determined from manganese calibration curvesfollowing the pilot ion method (15). The sensitivities of theorganic-FeIII
(aq) complexes have yet to be determined due inpart to the number of natural ligands present in sediments,and their concentrations are therefore reported in units ofcurrent intensity (nA) (16). Similarly, the stoichiometry ofFexSy (aq) is unknown and current intensities (nA) are reportedfor these species (17). All voltammetric analyses wereperformed with a computer-operated DLK-100A potentiostatand a micromanipulator capable of vertical movements atthe submillimeter scale (Analytical Instrument Systems, Inc.).Voltammograms were processed with a Matlab-based pro-gram (18). Multiple porewater geochemical profiles wereobtained during static and upflow conditions, with a finalprofile obtained 16 weeks after the onset of upflow conditions(29 weeks of total incubation; 71 pore volumes flushed) (6).
Column Dissection. The capped sediment column wasdissected for microbiological analyses immediately after thefinal geochemical profiling at 16 weeks of upflow conditions.Overlying surface water was removed from the column witha needle and syringe under aseptic conditions. A core samplewas obtained from the center of the column using a zerocontamination soil sampler (JMC Soil Samplers), with thecore sample encased in a PETG copolyester liner (30 cmlength; 2.0 cm i.d.). The core was frozen at -20 °C thenextruded and dissected under aseptic conditions. DNA wasextracted from the solid and aqueous phases of eachdissection zone, with detailed methodology provided in theSupporting Information. Notably, the DNA extraction pro-cedure was also applied to autoclaved, dry ASTM C-33 sandand the extract accompanied all measurements as a controlto quantify microbial populations in the sand prior to capping.
The first column dissection was made 10 mm below thecap-water interface, with successive dissections at constantintervals until the sediment-cap interface was reached
(Figure 1). Seven samples were obtained from the sand capmaterial (y ) 0 to -70 mm) and four samples were obtainedfollowing dissection of the underlying sediment (y )-70 to-140 mm). The samples were immediately placed into sterile,15 mL centrifuge tubes and stored at-20 °C. Dissected zoneswere labeled sequentially beginning with Zone 1 at thecap-water interface and ending with Zone 11 in the sediment(Figure 1).
Primer Design. qPCR primer pairs for Shewanella spp.and Anaeromyxobacter spp. were designed with the OligoDesign and Analysis Tools (www.idtdna.com) based on thenearly complete 16S rRNA gene sequences available inGenBank. The 16S rRNA gene sequences were aligned usingthe MegAlign program of the Lasergene software package(DNA Star, Inc.). The sequences of the selected primers areprovided in Table 1. The reverse primer for Shewanella spp.,She220R, is the reverse complement of She211f, a traditionalPCR primer (19). The Anaeromyxobacter spp.-targeted prim-ers were obtained using Applied Biosystems Primer ExpressTM
software. A 200 bp section of the 16S rRNA gene sequencecontaining an Anaeromyxobacter-specific region was identi-fied by analysis of the alignment and entered into the Primer
FIGURE 1. Porewater geochemical characterization of AnacostiaRiver sediment capped with ASTM C-33 concrete sand after 16weeks of upflow conditions. Values for organic-FeIII
(aq) andFexSy (aq) complexes are reported as current intensity and plottedalong the top x-axis. The concentrations of O2, Fe2+, Mn2+, andΣH2S (µM) are referenced to the lower x-axis. Dissectionlocations are indicated by solid gray lines, with numberscorresponding to dissection zones. The cap-water interface islocated at y ) 0 mm. The sediment-cap interface is located at y )-70 mm.
VOL. 43, NO. 1, 2009 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 67

TABL
E1.
Cond
ition
san
dPr
imer
sEm
ploy
eddu
ring
Quan
titat
ive
Real
-Tim
ePC
RAn
alys
is
targ
etgr
oup
qPCR
chem
istr
ypr
imer
/pro
bena
me
sequ
ence
stan
dard
sre
fere
nces
a
tota
lB
acte
ria
Taq
Man
Bac
1055
YF
5′-A
TG
GY
TG
TC
GT
CA
GC
T-3
′G
eob
acte
rlo
vley
iS
Z20
,52
Bac
1392
R5′
-AC
GG
GC
GG
TG
TG
TA
C-3
′B
ac11
15P
rob
e5′
-FA
M-C
AA
CG
AG
CG
CA
AC
CC
-TA
MR
A-3
′to
talA
rch
aea
Taq
Man
AR
CH
1-13
69F
5′-C
GG
TG
AA
TA
CG
TC
CC
TG
C-3
′M
eth
ano
cocc
us
mar
ipal
ud
is21
,53
AR
CH
2-13
69F
5′-C
GG
TG
AA
TA
TG
CC
CC
TG
C-3
′P
RO
K15
41R
5′-A
AG
GA
GG
TG
AT
CC
TG
CC
GC
A-3
′T
M13
89F
5′-F
AM
-CT
TG
TA
CA
CA
CC
GC
CC
GT
C-T
AM
RA
-3′
δ-P
rote
ob
acte
riab
Taq
Man
361F
5′-A
AG
CC
TG
AC
GC
AS
CA
A-3
′G
eob
acte
rb
emid
jien
sis
2268
5R5′
-AT
CT
AC
GG
AT
TT
CA
CT
CC
TA
CA
C-3
′E
ub
15′
-FA
M-G
TA
TT
AC
CG
CG
GN
TG
CT
GG
C-T
AM
RA
-3′
An
aero
myx
ob
acte
rsp
p.
SY
BR
Gre
enI
Am
yx-3
99F
5′-G
CA
AC
GC
CG
CG
TG
TG
T-3
′A
nae
rom
yxo
bac
ter
deh
alo
gen
ans
str.
2CP
-Cth
isst
ud
yA
myx
-466
R5′
-TC
CC
TC
GC
GA
CA
GT
GC
TT
-3′
Geo
bac
tera
cea
spp
.S
YB
RG
reen
IG
eo56
4F5′
-CA
AG
TC
GT
AC
GA
GA
AA
CA
TA
TC
-3′
Geo
bac
ter
lovl
eyiS
Z42
,54
Geo
840R
5′-G
AA
GA
GG
AT
CG
TC
TT
TC
CA
CG
A-3
′S
hew
anel
lae
spp
.S
YB
RG
reen
IS
he
120F
5′-G
CC
TA
GG
GA
TC
TG
CC
CA
GT
CG
-3′
Sh
ewan
ella
on
eid
ensi
sM
R1
this
stu
dy
Sh
e22
0R5′
-CT
AG
GT
TC
AT
CC
AA
TC
GC
G-3
′D
ehal
oco
cco
ides
spp
.T
aqM
anD
hc1
200F
5′-C
TG
GA
GC
TA
AT
CC
CC
AA
AG
CT
-3′
Deh
alo
cocc
oid
essp
.str
.FL2
(55)
Dh
c127
1R5′
-CA
AC
TT
CA
TG
CA
GG
CG
GG
-3′
Dh
c124
0Pro
be
5′-F
AM
-TC
CT
CA
GT
TC
GG
AT
TG
CA
GG
CT
GA
A-T
AM
RA
-3′
dsr
Ag
ene
SY
BR
Gre
enI
dsr
-1f
5′-A
CS
CA
CT
GG
AA
GC
AC
G-3
′D
esu
lfo
vib
rio
vulg
aris
sub
sp.v
ulg
aris
Hild
enb
oro
ug
h(2
3)d
sr-5
00r
5′-C
GG
TG
MA
GY
TC
RT
CC
TG
-3′
mcr
Ag
ene
SY
BR
Gre
enI
ME
1fw
d5′
-GC
MA
TG
CA
RA
TH
GG
WA
TG
TC
-3′
Met
han
osa
rcin
am
azei
(23)
ME
3re
v5′
-TG
TG
TG
AA
WC
CK
AC
DC
CA
CC
-3′
aO
rig
inal
liter
atu
reso
urc
esfo
rp
rim
er/p
rob
en
ames
and
seq
uen
ces.
bP
erfo
rmed
by
Mic
rob
ial
Insi
gh
ts,
Inc.
,R
ock
ford
,T
N.
68 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 43, NO. 1, 2009

ExpressTM program to find a short region with a consistentmelting temperature. BLAST analysis suggested primerspecificity to the target sequences.
Quantitative Real-Time PCR (qPCR) Analysis. Targetorganisms, reaction chemistries, primer sequences, andstandards for all qPCR analyses are summarized in Table 1.Total Bacteria were quantified as described by Ritalahti et al.(20). Total Archaea were quantified using primers andmethods described by Da Silva et al. (21), with genomic DNAextracted from Methanococcus maripaludis (ATCC 43000) asa standard. Quantification of δ-Proteobacteria was performedby Microbial Insights, Inc. using primers and conditionsdescribed by Stults et al. (22). The qPCR reaction mixturesfor Anaeromyxobacter spp.-, Geobacter spp.-, and Shewanellaspp.-targeted analysis contained 15 µL of Power SYBR GreenPCR master mix (Applied Biosystems), 300 nM of each primer,and 3 µL of template DNA (10.8-90.0 ng of DNA) in a totalreaction volume of 30 µL. The qPCR temperature programwas as follows: 2 min at 50 °C, 10 min at 95 °C, then 40 cyclesof denaturation at 95 °C for 15 s and annealing at 60 °C for1 min, with fluorescence measured after each annealing step.The dsrA gene was quantified according to Wilms et al. (23),with the exception that only 2.5 µL of template DNA wasutilized instead of 10 µL. The standard for dsrA calibrationwas genomic DNA from Desulfovibrio vulgaris (ATCC 29579;GenBank accession number NC_002937), whose genomecontains one copy of the dsrA gene. The mcrA gene wasquantified as described (23), except that the reverse primerwas modified such that a W (A or T) replaced S (C or G) atthe ninth position (Table 1) to fully complement the originalprimers designed by Hales et al. (24). The standard for mcrAcalibration was genomic DNA from Methanosarcina mazei(ATCC BAA-159; GenBank accession number NC_003901),whose genome contains one copy of the mcrA gene. Furtherdetails on qPCR procedures are provided in the SupportingInformation.
Terminal-Restriction Fragment Length Polymorphism(T-RFLP) Analysis. Dissection zones 1, 2, 3, 4, 6, 8, and 10exhibited unique geochemical characteristics and wereselected for T-RFLP analysis, following previously describedprocedures (25) described briefly in the Supporting Informa-tion. The restriction enzymes utilized were HhaI, MspI, andRsaI. Anaeromyxobacter dehalogenans strain 2CP-C DNA wasdigested with each digestion enzyme to ensure that experi-mentally derived T-RFLP patterns matched predicted in silicorestriction fragments (see the Supporting Information).
Data Analysis. T-RFLP profiles were analyzed to elucidatestatistically significant information regarding bacterial com-munity composition and diversity among dissection zones(26). Canonical correspondence analysis (CCA) was per-formed on Zones 1, 2, 3, 4, 6, 8, and 10 to examine potentialrelationships between bacterial community patterns andenvironmental variables (27) (CANOCO 4.5 software, Mi-crocomputer Power) with significance defined at p < 0.05.Quantitative environmental variables tested were averagedepth and concentrations of Mn2+, Fe2+, organic-FeIII
(aq), andFexSy(aq), measured during the voltammetric analysis. Resultswere summarized in ordination diagrams (CanoDraw 4.0).
Three additional, independent species diversity indiceswere calculated from normalized HhaI T-RFLP fragmentpatterns to analyze microbial diversity and composition. Thefirst indicator was species richness (S), equivalent to theamount of unique T-RFs in each zone (28). The secondindicator was Shannon’s diversity index (H), an estimate ofdiversity accounting for both species abundance and even-ness, calculated using StatsDirect Statistical Software (Stats-Direct, Ltd.). Typical values are between 1.5 and 3.5 withhigher values indicating a greater degree of diversity. Thethird indicator was the Smith and Wilson evenness value(Evar.), recommended by Blackwood et al. (29) to examine
species parity from T-RFLP data, ranging from 0 (skeweddistribution) to 1 (even distribution) (30). Phylogeneticanalysis using a web-based, in silico process was alsoperformed to identify dominant community members in eachdissection zone and is described in the Supporting Informa-tion.
ResultsPorewater Geochemical Profile. Voltammetric analysis ofporewaters indicate that multiple biogeochemical processeswere spatially distributed within the capped sediment column(Figure 1). Oxygen was present in the overlying water atrelatively constant concentrations before decreasing tonondetect concentrations only 2.8 mm below the cap-waterinterface. A region dominated by manganese reduction wasobserved below the aerobic portion of the column, evidencedby peaking Mn2+ concentrations (0 to -24.0 mm). This wasfollowed by a region dominated by Fe(III) reduction, indicatedby measured increases in Fe2+ and organic-FeIII
(aq) levels.Ferrous iron decreased to nondetect concentrations at-87.0mm due to the formation of dissolved iron sulfide complexeswithin the sediment. Sulfate reduction within the cap andsediment may have been utilized for FexSy(aq) formation (6).In general, the capped sediment column exhibited stratifiedredox zones distributed according to the thermodynamicyield of terminal electron accepting processes and encom-passed cap and sediment material. Spatial and temporaldevelopment of the stratified biogeochemical processes isthe focus of previous work (6) which includes more detaileddiscussions of the porewater profile.
Material from the dissection zones were characterized byvarious biogeochemical processes according to the porewatergeochemical profile (Figure 1). Zone 1 included oxygen andmanganese reduction; Zone 2 exhibited manganese reduc-tion; Zones 3 and 4 displayed iron and manganese reduction;Zone 5 included Fe(III) reduction; Zones 6 through 8displayed Fe(III) reduction and FexSy(aq) formation; and Zones9 through 11 included Fe(III) reduction and FexSy(aq) formationbut no dissolved, free ferrous iron.
Target Microbial Populations. qPCR analyses confirmedthat microbial populations were present and quantifiable ineach dissected zone of the sediment and overlying sand cap(Figure 2). Bacterial cell numbers in the cap generallydecreased in each subsequent zone (Figure 2A), from 5.2 ×108 cells per mL porewater in Zone 1 to 1.7 × 106 cells permL porewater in Zone 6, before an increase in Zone 7 directlyabove the sediment-cap interface. Bacterial cell titers wererelatively constant within the sediment (Zones 8-10), withobserved mean values between 5.9 × 108 and 6.8 × 108 cellsper mL porewater. Neither Bacteria nor Archaea weredetected in DNA extracted from dry, autoclaved ASTM C-33sand (Figure 2).
Bacteria associated with dissimilatory metal reductionand potential contaminant biotransformations were detectedin each zone of the sediment and the overlying sand cap(Figure 2A). δ-Proteobacteria were detected throughout thecolumn at levels similar to those of total Bacteria. Geobacterspp., Anaeromyxobacter spp., and Shewanella spp., were eachdetected in all zones. Geobacter cell titers in the cap rangedfrom 105 to 106 cells per mL porewater before peaking in thesediment (Zone 9) at 6.7 × 107 cells per mL porewater.Anaeromyxobacter numbers were most prominent in theupper cap layers (9.9 × 105 in Zone 1) and in the sediment(1.8 × 107 in Zone 9). The presence of Shewanella species,suspected to be present following detection of an organic-FeIII
(aq) complex observed previously in association with thisorganism (6, 31), were most abundant in the upper cap layers(1.0 × 106 in Zone 1), then stabilized to mean values between104 and 105 cell per mL of porewater before increasing slightlyin the sediment. In Zone 1, dissimilatory metal- and iron-
VOL. 43, NO. 1, 2009 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 69
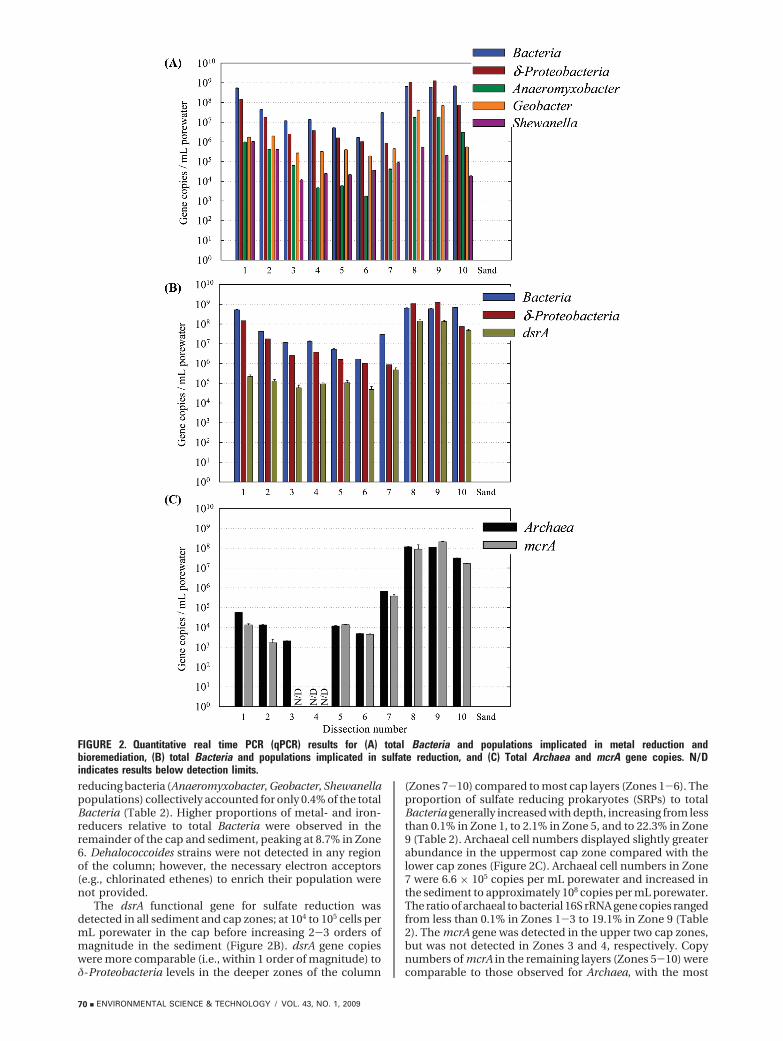
reducing bacteria (Anaeromyxobacter, Geobacter, Shewanellapopulations) collectively accounted for only 0.4% of the totalBacteria (Table 2). Higher proportions of metal- and iron-reducers relative to total Bacteria were observed in theremainder of the cap and sediment, peaking at 8.7% in Zone6. Dehalococcoides strains were not detected in any regionof the column; however, the necessary electron acceptors(e.g., chlorinated ethenes) to enrich their population werenot provided.
The dsrA functional gene for sulfate reduction wasdetected in all sediment and cap zones; at 104 to 105 cells permL porewater in the cap before increasing 2-3 orders ofmagnitude in the sediment (Figure 2B). dsrA gene copieswere more comparable (i.e., within 1 order of magnitude) toδ-Proteobacteria levels in the deeper zones of the column
(Zones 7-10) compared to most cap layers (Zones 1-6). Theproportion of sulfate reducing prokaryotes (SRPs) to totalBacteria generally increased with depth, increasing from lessthan 0.1% in Zone 1, to 2.1% in Zone 5, and to 22.3% in Zone9 (Table 2). Archaeal cell numbers displayed slightly greaterabundance in the uppermost cap zone compared with thelower cap zones (Figure 2C). Archaeal cell numbers in Zone7 were 6.6 × 105 copies per mL porewater and increased inthe sediment to approximately 108 copies per mL porewater.The ratio of archaeal to bacterial 16S rRNA gene copies rangedfrom less than 0.1% in Zones 1-3 to 19.1% in Zone 9 (Table2). The mcrA gene was detected in the upper two cap zones,but was not detected in Zones 3 and 4, respectively. Copynumbers of mcrA in the remaining layers (Zones 5-10) werecomparable to those observed for Archaea, with the most
FIGURE 2. Quantitative real time PCR (qPCR) results for (A) total Bacteria and populations implicated in metal reduction andbioremediation, (B) total Bacteria and populations implicated in sulfate reduction, and (C) Total Archaea and mcrA gene copies. N/Dindicates results below detection limits.
70 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 43, NO. 1, 2009

mcrA gene copies detected in the sediment (2.1 × 108 inZone 9). The proportion of mcrA gene copies to Archaea 16SrRNA gene copies ranged from 12.5% in Zone 2 to greaterthan 100% in Zone 9 (Table 2). A modified mcr gene hasrecently been identified in enrichments of anaerobic methaneoxidizers (32) and may be involved in reverse methanogenesisunder anoxic conditions (33, 34). However, the mcrA geneis conserved among methanogenic Archaea, which are a morecommonly encountered environmental population.
Bacterial Community Analysis. Analysis of T-RFLP pat-terns via CCA testing revealed that changes in bacterialcommunity composition among dissection zones could besignificantly correlated to environmental variables. Averagedepth best explained changes in the T-RFLP fingerprints andwas the only variable independently related to the data (p) 0.015). The T-RFLP data were explained with the greatestdegree of confidence (p ) 0.008), when average depth, Fe2+,and Mn2+ concentrations (µM) were all considered, sug-gesting these three variables collectively influenced com-munity composition (Figure 3). Therefore, based on the CCAresults, the hypothesis of no relation between organism andenvironment is rejected due to statistically significant speciescomposition changes correlated to environmental variables.
Indices of bacterial richness and evenness were uniquebut generally constant across all zones analyzed (Table 3).Most species richness values ranged from 95 to 132, indicatinga complex community. Previous studies estimating richnessfrom T-RFLP data classified values between 45 and 72 ashaving a high degree of complexity in community structure(28). The maximum species richness was observed in Zone1 (S ) 132), followed by Zones 8 and 10 (S ) 122 for eachzone). A moderate, positive correlation between bacterialnumbers and species richness (R2 ) 0.75) was observed suchthat the zones with the highest richness also had the highestqPCR bacterial cell titers. Notably, Zone 6 had very low speciesrichness (S ) 21) and modifications to the T-RFLP experi-mental protocol and replicate T-RFLP analyses yielded similarresults. Shannon index values were between 3.45 and 4.00for most zones, with Zone 6 being the exception (H ) 2.13).A modest correlation was detected between qPCR bacterialnumbers and diversity (R2 ) 0.66). Smith-Wilson estimatesof evenness suggested that species abundances within mostzones were relatively similar, except for Zone 1 which showedskewed abundance values (Evar. ) 0.23) (i.e., certain popula-tions were dominant relative to the community). In silicophylogenetic assignment of T-RFLP peaks at the phylum-
class level provided additional insight into communitycomposition and shifts among dissection zones and can befound in the Supporting Information.
DiscussionThe microbial colonization of a laboratory-scale in situsediment cap was demonstrated using an integrated, culture-independent approach of voltammetry-based porewatergeochemical measurements, qPCR to enumerate targetmicrobial populations, and T-RFLP to examine changes inbacterial community structure. qPCR analysis confirmed thatBacteria and Archaea inhabited the cap in numbers ap-proaching those in the underlying sediment. Microorganismscolonizing the overlying cap originated from the sediment,since PCR analysis did not detect microorganisms associatedwith the sand prior to cap emplacement. Statistical analysisof T-RFLP data with CCA demonstrated that changes incommunity patterns were significantly related to environ-
TABLE 2. Proportion of Groups of Metal Reducers, SulfateReducers, And Methanogens to Total Microorganisms for EachDissection Zone, Calculated from qPCR Resultsa
dissectionzone
proportionof metal-reducersb
to Bacteria
proportionof dsrA toBacteria
proportionof Archaeato Bacteria
proportionof mcrA to
Archaea
1 0.4 <0.1 <0.1 23.42 4.0 0.3 <0.1 12.53 2.7 0.5 <0.1 N/A4 4.1 0.7 N/A c N/A5 6.3 2.1 0.2 >1006 8.7 2.9 0.3 92.57 1.2 1.6 2.2 57.98 3.8 22.2 18.5 75.99 6.9 22.3 19.1 >10010 0.2 6.7 4.7 52.2
a All values are reported in percentages of gene copiesto gene copies. The sediment-cap interface is locatedbetween Zones 7 and 8. b Metal- and iron- reducing bac-teria, defined as the cumulative gene copies of Anaeromyxo-bacter, Geobacter, and Shewanella. c Calculation not able tobe performed due to quantities below detection limits.
FIGURE 3. Ordination diagram based on canonical correspo-ndence analysis of T-RFLP community profiles from dissectionregions 1, 2, 3, 4, 8, and 10. The coordinates of the site points arethe scores of each individual site on the two synthetic axes.Arrows indicate quantitative environmental variables and point inthe direction of maximum change in the value of the associatedvariable. The eigenvalues of axis 1 (horizontal) and axis 2(vertical) are 0.696 and 0.497, respectively, and represent thepercentage of community variance explained by extracted,synthetic gradients.
TABLE 3. Indices of Bacterial Community Structure Calculatedfrom HhaI T-RFLP Fragment Patterns at Each DissectionRegion Analyzed
dissectionzone richness (S )
Shannondiversity (H )
Smith-Wilsonevenness
(Evar.)
1 132 3.87 ( 0.0023a 0.232 95 3.45 ( 0.0025 0.433 105 3.63 ( 0.0037 0.484 93 3.52 ( 0.0052 0.516 21 2.13 ( 0.0049 0.408 122 4.00 ( 0.0033 0.5310 122 3.73 ( 0.0029 0.45
a Estimated value ( one standard deviation.
VOL. 43, NO. 1, 2009 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 71

mental variables (depth, Fe2+, and Mn2+), suggesting thatmicrobial data collected in these studies did not reflect onlythe elution of planktonic microorganisms from the sediment.If this were the case, then environmental variables wouldnot adequately explain spatial changes in bacterial com-munities. Although the elution of organisms may havecontributed to the microbial numbers observed in the cap,the results demonstrate that microbial populations inhabitedthe overlying cap and formed distinct communities accordingto depth and redox gradients.
Metal- and iron- reducing populations (δ-Proteobacteria,Anaeromyxobacter spp., Geobacter spp., and Shewanella spp.)were detected in each zone of the cap and sediment,confirming geochemical data suggesting that microbial Fe(III)reduction was a major biogeochemical process (6). In Zone1, Anaeromyxobacter, Geobacter, Shewanella populationsaccounted for only 0.4% of the total Bacteria (Table 3),consistent with observed geochemical data indicating oxygenreduction was occurring. Zones 2-9 displayed higher pro-portions of Anaeromyxobacter, Geobacter, Shewanella popu-lations relative to total Bacteria, agreeing with the geochem-ical data that implied metal reduction in these zones.Geobacter consistently outnumbered Shewanella and Anaer-omyxobacter within the cap. Interestingly, Anaeromyxobacterlevels were low in the cap but exceeded Geobacter numbersin the sediment. Anaeromyxobacter and Shewanella cell titerswere higher in the upper zones relative to the rest of the cap,possibly due to their roles in oxygen and manganesereduction. Geobacter spp. were also detected in Zone 1 andcan remain viable under aerobic conditions (35) and/orparticipate in manganese reduction (36).
The proportion of SRPs compared to total Bacteriain-creased 3-4 orders of magnitude in the sediment comparedtothe cap (Table 2), complementing porewater geochemicalprofiles suggesting sulfate reduction in the sediment(Figure 1 (6)). The detection of the dsrA gene in the capregions means that microbial sulfate reduction may havesupplied the sulfide for FexSy(aq) formation in the cap, withsulfide possibly also contributing to manganese oxidereduction in the upper cap (y ) -20 mm (37)). Thedetection of dsrA in the more oxidized regions of the cap(Zones 1 and 2) is not unfounded since certain SRPswithstand exposure to oxygen (38). The detection of themcrA gene in the lower portion of the cap suggests thepresence of methanogenic Archaea, demonstrating thatthe entire range of microbial metabolic processes (oxygenreduction to methanogenesis) were spatially distributedwithin the cap. Thus, a diverse range of microorganisms,spanning the spectrum of microbially catalyzed redoxreactions, were colonized in the cap after 29 weeks ofemplacement. This colonization most probably contrib-uted to redox zone evolution within the system, in parallelwith abiotic reactions discussed in detail previously (6)(e.g., Mn(IV) reduction by Fe(II)). The microbial coloniza-tion and development of biogeochemical zones observedwithin the cap provides additional insight into microbialcommunity response following physical and geochemicalperturbations to the sedimentary environment. Microbialpopulations appear to sense, respond, and adapt to redoxchanges at the sediment-water interface to create strati-fied, functional communities in short time periods. Similarobservations have been made in natural systems such asflooded tropical soils (39).
Finally, intrinsic microbial colonization of an in situ capcan impact contaminant fate and transport within suchengineered systems. Organisms colonizing the overlying capduring this study included Anaeromyxobacter, Geobacter, andShewanella spp., with members of these groups capable oftransforming multiple classes of environmental contaminantsincluding radionuclides (40), petroleum hydrocarbons (41)
and chlorinated ethenes (42). In previous experiments,actively dechlorinating Dehalococcoides eluted from enrichedAnacostia River sediment under upflow conditions (43),suggesting they may also populate a sediment cap undersuitable conditions. This is significant considering Dehalo-coccoides have been linked to the reduction of chlorinatedethenes (44), PCBs (45), polybrominated diphenyl ethers (46),chlorophenols (47), and dioxins (48) among other chlorinatedorganics (48, 49). Despite previous detection and quantifica-tion of Dehalococcoides strains in Anacostia sediment (43),Dehalococcoides were not detected in this study due to a lackof chlorinated organics provided as electron acceptors. Inthe aerobic region of the cap, the presence of a diversebacterial community suggests that aerobic PAH mineraliza-tion (14) may occur if oxygen penetration is sufficient. Also,methanotrophic bacteria have been linked to the cometabolictransformation of organic contaminants (50) and weretentatively identified through T-RFLP analysis to be presentat the cap-water interface (Supporting Information TableS.2). Importantly, rapid microbial colonization providescapping with an intrinsic, secondary level of treatment (i.e.,biotransformations) in addition to physical isolation of thesediment contaminants. The natural colonization of the capmaterial demonstrated here suggests that enhanced microbialcontaminant degradation in the cap is feasible, thus openingup new avenues for innovative cap design (e.g., cappingcombined with biostimulation) and active capping tech-nologies. Alternatively, the presence of microorganismswithin subaqueous sediment caps can also have negativeimplications that should be considered. Biogenic gas pro-duction (e.g., methanogenesis) within the sediment and capcould lead to contaminant partitioning and transport in thegas phase (51), physical instability of the cap (3), andformation and release of greenhouse gases to the atmosphere.Additional research is needed to optimize cap design andemplacement to fully exploit microbial activity in caps andto ensure long-term cap stability and performance.
AcknowledgmentsWe thank Rebecca Daprato and Laura Guest for helpfuldiscussions and assistance with T-RFLP analysis, ClaribelCruz-Garcia for helpful discussions regarding qPCR analysis,and John Fortner for assistance and advice during columndissection. Funding for this research was provided by theHazardous Substance Research Center-South and Southwestand a fellowship to D.W.H from the Georgia Institute ofTechnology.
Supporting Information AvailableTabular qPCR data (Table S.1) and additional T-RFLP analysesincluding representative data (Figure S.1), positive controldata (Figure S.2), and in silico phylogenetic assignments(Table S.2). This material is available free of charge via theInternet at http://pubs.acs.org.
Literature Cited(1) The Incidence and Severity of Sediment Contamination in Surface
Waters of the United States, National Sediment Quality Survey,2nd ed.; U.S. Environmental Protection Agency: Washington,DC, 2004.
(2) Thoma, G. J.; Reible, D. D.; Valsaraj, K. T.; Thibodeaux, L. J.Efficiency of capping contaminated sediments in situ. 2.Mathematics of diffusion adsorption in the capping layer.Environ. Sci. Technol. 1993, 27 (12), 2412–2419.
(3) Reible, D. D.; Lambert, D.; Constant, D. W.; Mutch, R. D.; Zhu,Y. Active capping demonstration in the Anacostia River,Washington, DC Remediation 2007, 17 (1), 39-53.
(4) McDonough, K. M.; Murphy, P.; Olsta, J.; Zhu, Y. W.; Reible, D.;Lowry, G. V. Development and placement of a sorbent-amendedthin layer sediment cap in the Anacostia River. Soil Sed. Contam.2007, 16 (3), 313–322.
72 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 43, NO. 1, 2009

(5) McDonough, K. M. F. J.; Lowry, G. Adsorption of polychlorinatedbiphenyls to activated carbon: equilibrium isotherms and apreliminary assessment of the effect of dissolved organic matterand biofilm loadings. Water Res. 2008, 42 (3), 575–584.
(6) Himmelheber, D. W.; Taillefert, M.; Pennell, K. D.; Hughes, J. B.Spatial and temporal evolution of biogeochemical processesfollowing in situ capping of contaminated sediments. Environ.Sci. Technol. 2008, 42, 4113–4120.
(7) Jorgensen, B. B.; Revsbech, N. P. Diffusive boundary-layers andthe oxygen-uptake of sediments and detritus. Limnol. Ocean.1985, 30 (1), 111–122.
(8) Findlay, S.; Sobczak, W. K. Microbial Communities in HyporheicSediments. In Streams and Ground Waters; Jones, J. B.;Mulholland, P. J. Eds.; Academic Press: Sand Diego, CA, 2000.
(9) Zehnder, A. J. B.; Stumm, W. Geochemistry and biogeochemistryof anaerobic habitats. In Biology of Anaerobic Microorganisms;Zehnder, A. J. B., Ed.; Wiley: New York, 1988.
(10) Gadd, G. M. Microbial influence on metal mobility andapplication for bioremediation. Geoderma 2004, 122 (2-4), 109–119.
(11) Phelps, C. D.; Young, L. Y. Anaerobic biodegradation of BTEXand gasoline in various aquatic sediments. Biodegradation 1999,10 (1), 15–25.
(12) Conant, B.; Cherry, J. A.; Gillham, R. W. A PCE groundwaterplume discharging to a river: Influence of the streambed andnear-river zone on contaminant distributions. J. Contam. Hydrol.2004, 73 (1-4), 249–279.
(13) Magar, V. S.; Johnson, G. W.; Brenner, R. C.; Quensen, J. F.;Foote, E. A.; Durell, G.; Ickes, J. A.; Peven-McCarthy, C. Long-term recovery of PCB-contaminated sediments at the LakeHartwell Superfund site: PCB dechlorination. 1. End-membercharacterization. Environ. Sci. Technol. 2005, 39 (10), 3538–3547.
(14) Samanta, S. K.; Singh, O. V.; Jain, R. K. Polycyclic aromatichydrocarbons: Environmental pollution and bioremediation.Trends Biotechnol. 2002, 20 (6), 243–248.
(15) Brendel, P. J.; Luther, G. W. Development of a gold amalgamvoltammetric microelectrode for the determination of dissolvedFe, Mn, O2, and S(-II) in porewaters of marine and fresh-watersediments. Environ. Sci. Technol. 1995, 29 (3), 751–761.
(16) Taillefert, M.; Bono, A. B.; Luther, G. W. Reactivity of freshlyformed Fe(III) in synthetic solutions and (pore)waters: Volta-mmetric evidence of an aging process. Environ. Sci. Technol.2000, 34 (11), 2169–2177.
(17) Theberge, S. M.; Luther, G. W. Determination of the electro-chemical properties of a soluble aqueous FeS species presentin sulfidic solutions. Aquat. Geochem. 1997, 3 (3), 191–211.
(18) Bristow, G.; Taillefert, M. VOLTINT: A Matlab-based programfor semi-automated processing of geochemical data acquiredby voltammetry. Comp. Geosci. 2008, 34 (2), 153–162.
(19) Todorova, S. G.; Costello, A. M. Design of Shewanella-specific16S rRNA primers and application to analysis of Shewanella ina minerotrophic wetland. Environ. Microb. 2006, 8 (3), 426–432.
(20) Ritalahti, K. M.; Amos, B. K.; Sung, Y.; Wu, Q. Z.; Koenigsberg,S. S.; Loffler, F. E. Quantitative PCR targeting 16S rRNA andreductive dehalogenase genes simultaneously monitors multipleDehalococcoides strains. Appl. Environ. Microb. 2006, 72 (4),2765–2774.
(21) Da Silva, M. L. B.; Alvarez, P. J. J. Enhanced anaerobicbiodegradation of benzene-toluene-ethylbenzene-xylene-etha-nol mixtures in bioaugmented aquifer columns. Appl. Environ.Microbiol. 2004, 70 (8), 4720–4726.
(22) Stults, J. R.; Snoeyenbos-West, O.; Methe, B.; Lovley, D. R.;Chandler, D. P. Application of the 5 ′ fluorogenic exonucleaseassay (TaqMan) for quantitative ribosomal DNA and rRNAanalysis in sediments. Appl. Environ. Microbiol. 2001, 67 (6),2781–2789.
(23) Wilms, R.; Sass, H.; Kopke, B.; Cypionka, H.; Engelen, B. Methaneand sulfate profiles within the subsurface of a tidal flat arereflected by the distribution of sulfate-reducing bacteria andmethanogenic archaea. FEMS Microbiol. Ecol. 2007, 59 (3), 611–621.
(24) Hales, B. A.; Edwards, C.; Ritchie, D. A.; Hall, G.; Pickup, R. W.;Saunders, J. R. Isolation and identification of methanogen-specific DNA from blanket bog feat by PCR amplification andsequence analysis. Appl. Environ. Microbiol. 1996, 62 (2), 668–675.
(25) Daprato, R. C.; Loffler, F. E.; Hughes, J. B. Comparative analysisof three tetrachloroethene to ethene halorespiring consortiasuggests functional redundancy. Environ. Sci. Technol. 2007,41 (7), 2261–2269.
(26) Kitts, C. Terminal restriction fragment patterns: A tool forcomparing microbial communities and assessing communitydynamics. Curr. Issues. Intest. Microbiol. 2001, 2 (1), 17–25.
(27) Ter Braak, C. J. F.; Verdonschot, P. F. M. Canonical cor-respondence-analysis and related multivariate methods inaquatic ecology. Aquat. Sci. 1995, 57 (3), 255–289.
(28) Liu, W. T.; Marsh, T. L.; Cheng, H.; Forney, L. J. Characterizationof microbial diversity by determining terminal restrictionfragment length polymorphisms of genes encoding 16S rRNA.Appl. Environ. Microbiol. 1997, 63 (11), 4516–4522.
(29) Blackwood, C. B.; Hudleston, D.; Zak, D. R.; Buyer, J. S.Interpreting ecological diversity indices applied to terminalrestriction fragment length polymorphism data: Insights fromsimulated microbial communities. Appl. Environ. Microbiol.2007, 73 (16), 5276–5283.
(30) Smith, B.; Wilson, J. B. A consumer’s guide to evenness indices.Oikos 1996, 76 (1), 70–82.
(31) Taillefert, M.; Beckler, J. S.; Carey, E.; Burns, J. L.; Fennessey,C. M.; DiChristina, T. D. Shewanella putrefaciens produces anFe(III)-solubilizing organic ligand during anaerobic respirationon insoluble Fe(III) oxides. J Inorg. Biochem. 2007, 101 (11-12),1760–1767.
(32) Friedrich, M. W. Methyl-coenzyme m reductase genes: Uniquefunctional markers for methanogenic and anaerobic methane-oxidizing archaea. Environ. Microbiol. 2005, 397, 428–442.
(33) Hallam, S. J.; Girguis, P. R.; Preston, C. M.; Richardson, P. M.;DeLong, E. F. Identification of methyl coenzyme-M reductaseA (mcrA) genes associated with methane-oxidizing archaea.Appl. Environ. Microbiol. 2003, 69 (9), 5483–5491.
(34) Hallam, S. J.; Putnam, N.; Preston, C. M.; Detter, J. C.; Rokhsar,D.; Richardson, P. M.; DeLong, E. F. Reverse methanogenesis:Testing the hypothesis with environmental genomics. Science.2004, 305 (5689), 1457–1462.
(35) Lin, W. C.; Coppi, M. V.; Lovley, D. R. Geobacter sulfurreducenscan grow with oxygen as a terminal electron acceptor. Appl.Environ. Microbiol. 2004, 70 (4), 2525–2528.
(36) Lovley, D. R.; Holmes, D. E.; Nevin, K. P. Dissimilatory Fe(III)and Mn(IV) reduction. Adv. Microbiol. Phys. 2004, 49, 219–286.
(37) Burdige, D. J.; Nealson, K. H. Chemical and microbiologicalstudies of sulfide-mediated manganese reduction. Geomicrobiol.J. 1986, 4 (4), 361–387.
(38) Mogensen, G. L.; Kjeldsen, K. U.; Ingvorsen, K. Desulfovibrioaerotolerans sp. nov., an oxygen tolerant sulphate reducingbacterium isolated from activated sludge. Anaerobe. 2005, 11(6), 339–349.
(39) Pett-Ridge, J.; Firestone, M. K. Redox fluctuation structuresmicrobial communities in a wet tropical soil. Appl. Environ.Microb. 2005, 71 (11), 6998–7007.
(40) Anderson, R. T.; Vrionis, H. A.; Ortiz-Bernad, I.; Resch, C. T.;Long, P. E.; Dayvault, R.; Karp, K.; Marutzky, S.; Metzler, D. R.;Peacock, A.; White, D. C.; Lowe, M.; Lovley, D. R. Stimulatingthe in situ activity of Geobacter species to remove uranium fromthe groundwater of a uranium-contaminated aquifer. Appl.Environ. Microbiol. 2003, 69 (10), 5884–5891.
(41) Anderson, R. T.; Rooney-Varga, J. N.; Gaw, C. V.; Lovley, D. R.Anaerobic benzene oxidation in the Fe(III) reduction zone ofpetroleum contaminated aquifers. Environ. Sci. Technol. 1998,32 (9), 1222–1229.
(42) Sung, Y.; Fletcher, K. F.; Ritalaliti, K. M.; Apkarian, R. P.; Ramos-Hernandez, N.; Sanford, R. A.; Mesbah, N. M.; Loffler, F. E.Geobacter lovleyi sp. nov. strain SZ, a novel metal-reducing andtetrachloroethene-dechlorinating bacterium. Appl. Environ.Microb. 2006, 72 (4), 2775–2782.
(43) Himmelheber, D. W.; Pennell, Kurt D.; Hughes, J. B. Naturalattenuation processes during in situ capping. Environ. Sci.Technol. 2007, 41 (15), 5306–5313.
(44) Loffler, F. E.; Edwards, E. A. Harnessing microbial activities forenvironmental cleanup. Curr. Opin. Biotechnol. 2006, 17 (3),274–284.
(45) Bedard, D. L.; Ritalahti, K. M.; Loffler, F. E. The Dehalococcoidespopulation in sediment-free mixed cultures metabolicallydechlorinates the commercial polychlorinated biphenyl mixtureAroclor 1260. Appl. Environ. Microbiol. 2007, 73, 2513–2521.
(46) He, J. Z.; Robrock, K. R.; Alvarez-Cohen, L. Microbial reductivedebromination of polybrominated diphenyl ethers (PBDEs).Environ. Sci. Technol. 2006, 40 (14), 4429–4434.
(47) Adrian, L.; Hansen, S. K.; Fung, J. M.; Gorisch, H.; Zinder, S. H.Growth of Dehalococcoides strains with chlorophenols aselectron acceptors. Environ. Sci. Technol. 2007, 41, 2318–2323.
VOL. 43, NO. 1, 2009 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 73

(48) Fennell, D. E.; Nijenhuis, I.; Wilson, S. F.; Zinder, S. H.; Haggblom,M. M. Dehalococcoides ethenogenes strain 195 reductivelydechlorinates diverse chlorinated aromatic pollutants. Environ.Sci. Technol. 2004, 38 (7), 2075–2081.
(49) Adrian, L.; Szewzyk, U.; Wecke, J.; Gorisch, H. Bacterialdehalorespiration with chlorinated benzenes. Nature. 2000, 408(6812), 580–583.
(50) Dolan, M. E.; Mccarthy, P. L. Methanotrophic chloroethenetransformation capacities and 1,1-dichloroethene transforma-tion product toxicity. Environ. Sci. Technol. 1995, 29 (11), 2741–2747.
(51) Yuan, Q. Z.; Valsaraj, K. T.; Reible, D. D.; Willson, C. S. Alaboratory study of sediment and contaminant release duringgas ebullition. J. Air Waste Manage. Assoc. 2007, 57 (9), 1103–1111.
(52) Harms, G.; Layton, A. C.; Dionisi, H. M.; Gregory, I. R.; Garrett,V. M.; Hawkins, S. A.; Robinson, K. G.; Sayler, G. S. Real-timePCR quantification of nitrifying bacteria in a municipal waste-
water treatment plant. Environ. Sci. Technol. 2003, 37 (2), 343–351.
(53) Suzuki, M. T.; Taylor, L. T.; DeLong, E. F. Quantitative analysisof small-subunit rRNA genes in mixed microbial populationsvia 5 ′-nuclease assays. Appl. Environ. Microbiol. 2000, 66 (11),4605–4614.
(54) Cummings, D. E.; Snoeyenbos-West, O. L.; Newby, D. T.;Niggemyer, A. M.; Lovley, D. R.; Achenbach, L. A.; Rosenzweig,R. F. Diversity of Geobacteraceae species inhabiting metal-polluted freshwater lake sediments ascertained by 16S rDNAanalyses. Microbiol. Ecol. 2003, 46 (2), 257–269.
(55) He, J. Z.; Ritalahti, K. M.; Aiello, M. R.; Loffler, F. E. Completedetoxification of vinyl chloride by an anaerobic enrichmentculture and identification of the reductively dechlorinatingpopulation as a Dehalococcoides species. Appl. Environ. Mi-crobiol. 2003, 69 (2), 996–1003.
ES801834E
74 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 43, NO. 1, 2009











![[5] Enrichment, Cultivation, and Detection of Reductively ...web.utk.edu/~microlab/LoefflerLab/Publications_files/Löffler et... · (Maymo´‐Gatell et al., 1997) were introduced](https://img.pdfslide.net/doc/110x75/5a7135e67f8b9a98538cb270/5-enrichment-cultivation-and-detection-of-reductively-webutkedumicrolabloefflerlabpublicationsfilesloffler.jpg)

















