Embed Size (px)
Citation preview

PRIMARY RESEARCH PAPER
Microhabitat influence on chironomid community structureand stable isotope signatures in West Greenland lakes
Nina S. Reuss • Ladislav Hamerlık • Gaute Velle •
Anders Michelsen • Ole Pedersen •
Klaus P. Brodersen
Received: 5 July 2013 / Revised: 6 February 2014 / Accepted: 7 February 2014
� Springer International Publishing Switzerland 2014
Abstract Most functional feeding types are repre-
sented within the species rich group of aquatic
chironomids. Thus, we hypothesized that different
lake types and microhabitats within lakes would (1)
host specific chironomid communities and (2) that the
individual communities would show specific d13C
stable isotope signatures reflecting the prevailing
origin of food source. To test our hypotheses, five
lakes in southwest Greenland were investigated at a
high taxonomic resolution and with detailed informa-
tion on d13C signature of the chironomids and of
individual microhabitats (macrophytes, sediment,
stones, and profundal). We found that there was a
significant difference in d13C between the chironomid
assemblages of freshwater lakes and oligosaline lakes,
while assemblages of the littoral microhabitats did not
differ significantly. The d13C of chironomids reflected
the wide variety of habitat signals, particularly in the
freshwater lakes. Our results indicate that many
chironomid taxa are ubiquitous and are found in
several microhabitats, suggesting that they can adjust
their feeding strategy according to the habitat. The
implication is that chironomid assemblage composition
has only limited use as indicator of littoral microhabitats
in the Arctic. On the other hand, the d13C signature of
fossil chironomids might have a potential as indicator of
microhabitats in freshwater lakes.
Keywords Arctic � Lake types � Habitats �Stable d13C isotopes � Chironomidae
Introduction
Chironomids are an important component of Arctic
lake ecosystems, often considered the dominant
primary consumers (Danks, 1981; Chetelat et al.,Handling editor: Sonja Stendera
N. S. Reuss (&) � L. Hamerlık � O. Pedersen �K. P. Brodersen
Freshwater Biological Laboratory, Department of
Biology, University of Copenhagen, 2100 Copenhagen,
Denmark
e-mail: [email protected]
L. Hamerlık
Faculty of Natural Sciences, Matthias Belius University,
Banska Bystrica, Slovakia
G. Velle
Uni Environment, Uni Research, Bergen, Norway
G. Velle
Department of Biology, University of Bergen, Bergen,
Norway
A. Michelsen
Terrestrial Ecology Section, Department of Biology,
University of Copenhagen, Copenhagen, Denmark
A. Michelsen
Center for Permafrost, University of Copenhagen,
Copenhagen, Denmark
123
Hydrobiologia
DOI 10.1007/s10750-014-1821-9

2010) that can occupy several trophic levels (Pinder,
1995; Reuss et al., 2013b). Chironomids are among the
most flexible aquatic organisms regarding the range of
occupied habitat types. They occur in every type of
freshwater; moreover, they colonize saline, marine,
and terrestrial habitats. Substratum type, along with
food quality and quantity, has been identified as an
important factor influencing the qualitative and quan-
titative structure of the chironomid communities of
lakes (Lindegaard, 1992).
Some chironomid species show strong preference
for a certain type of substratum, however, many
species are versatile in utilizing different substrate
types (Pinder, 1986). The basic substrate types, such as
hard rock, soft sediment, aquatic plants, and sub-
merged wood, are usually dominated by different
subfamilies (Pinder, 1980). Submerged macrophytes
and hard substrates provide a firm substratum in a
well-oxygenated environment and allow growth of
diatoms as an important food source and thus represent
similar ecological conditions (Tokeshi & Pinder,
1985). The littoral regions of lakes usually support
larger and more diverse populations of benthic inver-
tebrates than profundal areas (Moore, 1981), mainly
due to higher habitat complexity (Tolonen et al., 2001;
Brodersen et al., 2001b). Similarly, there is generally a
positive relationship between macrophytes and chi-
ronomid diversity and abundance as a result of an
increased surface area on macrophytes available for
invertebrates, stabilizing effects on the substratum
(Moore, 1981), and since macrophytes are a potential
food source (Pinder, 1986).
Stable isotopes of carbon (d13C) is an important
tool for identifying carbon flow in freshwater ecosys-
tems. The carbon isotopic signal is relatively stable
during trophic transfer and is, therefore, a good source
indicator while other stable isotope signals, like the
nitrogen isotopes (d15N), are subject to substantial
trophic fractionation (Peterson & Fry, 1987; Vander
Zanden & Rasmussen, 1999). Phytoplankton usually
show a depleted d13C signal compared to benthic algae
as the benthic algae are affected by carbon limitation
due to boundary layer conditions (France, 1995;
Hecky & Hesslein, 1995). Particulate organic matter
(POM) also show a depleted d13C signal (Chetelat
et al., 2010), while profundal sediments can be further
depleted by the use of respired CO2 (Peterson & Fry,
1987; Vander Zanden & Rasmussen, 1999). Investi-
gations of the origin of food for zooplankton,
zoobenthos, and higher trophic levels in boreal and
Arctic lakes have been able to differentiate between
use of littoral benthic sources, phytoplankton, and
allochthonous sources based on their isotopic signals
(Karlsson & Bystrom, 2005; Rautio & Vincent, 2007;
Chetelat et al., 2010; Premke et al., 2010).
We have studied the effects of habitat- and food
availability on the chironomid assemblages of low
Arctic lakes through detailed taxonomic and stable
isotope analyses of individual chironomid taxa of
microhabitats in different lake types. The microhab-
itats in the current study were further characterized by
pigment and microscopy analyses. Chironomids and
microhabitats were sampled by a SCUBA diver to
ensure a level of accuracy that is rare in such studies.
The study lakes are situated in the Kangerlussuaq
region, southwest Greenland, an area of limited
anthropogenic impact and a high number of lakes
differing in both physical and chemical properties. We
hypothesized that different lake types and microhab-
itats within the lakes would constitute specific chiron-
omid communities. In addition, the d13C isotopic
information of individual chironomid taxa and their
habitats were expected to reveal important informa-
tion of their food sources and if some habitats, i.e.,
macrophytes, are used as a direct food source or
primarily as shelter or substrate for the chironomids.
The ecological information of the chironomids
obtained in this study, with detailed knowledge of
habitat community structure and food sources, is
expected to be valuable in the understanding of aquatic
lake ecosystems in general, as a stepping stone for
contemporary ecological lake studies and in the
evaluation of paleolimnological records based on
analyses of assemblage structure of subfossil chiron-
omid head capsules.
Materials and methods
Study site
Two freshwater lakes, two oligosaline lakes, and a
lake receiving direct glacial input were sampled in an
attempt to maximize the number of chironomid taxa
and microhabitat types (Table 1; Fig. 1). Only one of
the lakes, Lake Ferguson, is known to contain fish. All
lakes are from the Kangerlussuaq area, southwest
Greenland. The Kangerlussuaq area is characterized as
Hydrobiologia
123

low Arctic, with annual average precipitation around
140 mm and average July temperature of 10.7�C
(Brodersen & Anderson, 2002). Both fresh- and
oligosaline lakes in this area have been targeted for
paleolimnological studies and have shown to be
sensitive to environmental changes (e.g., McGowan
et al., 2003; Anderson et al., 2008; Heggen et al.,
2010). These studies have suggested that some lakes
change between fresh- and oligosaline types over the
Holocene (McGowan et al., 2008; Reuss et al., 2013a).
Sampling
Microhabitats and chironomids were sampled in July
2009 by a SCUBA diver carefully collecting material
from individual microhabitats. The specific microhab-
itats included macrophytes, stones, and soft sediment
at various depths (Fig. 1). Profundal sediments were
collected with an Ekman grab at a maximum depth of
20 m. Temperature, conductivity, O2, and pH were
measured by a YSI-650 Multiparameter Water Quality
Sonde at the lake surface. Conductivity measurements
of the oligosaline lakes were obtained from Brodersen
& Anderson (2002) constituting an average of mea-
surements conducted on several occasions between
1996 and 2000. Surface water samples were collected
in 5 l bottles for analyses of particulate organic matter
(POM). Surface water was also collected in rinsed
50-ml plastic bottles for total phosphorus (TP) and
analyzed according to Strickland & Parsons (1968).
Habitats and food items
Individual habitats and possible food items including
macrophytes (Potamogeton filiformis, Calliergon
giganteum, Scorpidium scorpioides, Chara globular-
is, and Cladophora sp.), stone scrapes, sediment, and
planktonic POM were collected in triplicate for
characterization by isotope and pigment analyses.
Stones were scraped with a scalpel and the
scrapes transferred to 20-ml plastic vials in the field,
while macrophytes were rinsed in filtered lake water in
the lab before transferring to 20-ml plastic vials.
Sediments were collected by ‘‘mini-Kajaks’’ (50-ml
syringes with top cut off) and the surface *1 cm was
transferred to 20 ml plastic vials in the field. Profundal
sediments were sampled directly from the surface
*1 cm in the grab sampler. All samples were stored
frozen and subsequently freeze-dried before analyses,
thereby minimizing exposure to light and heat. POM
was collected on pre-combusted GF-F filters by
filtering of 0.2–2 l surface water on return to the
laboratory on the field day. Filters were blotted on
tissue paper, folded and wrapped in aluminum foil,
and stored frozen for pigment analyses or dried at
60�C for isotope analyses. In addition, material of the
dominating terrestrial vegetation types of the catch-
ments including Betula nana, Salix glauca, Rhodo-
dendron tomentosum, Vaccinium sp., Poaceae, and
litter (3–5 samples from the lake catchments) were
collected in paper bags and dried for minimum 24 h at
60�C for isotope analyses.
Samples for pigment analyses were freeze-dried
shortly before extraction and homogenized with a
spatula. 0.1–0.5 g stone scrape and sediment or
0.04–0.08 g plant material were extracted in 5 ml
cold 80:15:5% HPLC grade acetone:methanol:milli-Q
water spiked with an internal standard (Sudan II). The
mixture was sonicated, extracted in the freezer over
night, filtered (0.45 lm), diluted with milli-Q water,
Table 1 Physical and chemical characteristics of the investigated lakes
# Name Type Latitude
(N)
Longitude
(W)
Altitude
(m a.s.l.)
Area
(ha)
Max. depth
(m)
Cond.
(lS cm-1)
pH TP
(lg l-1)
Secci
(m)
1 Lake Ferguson Fresh 66.97898 50.69489 66 750 [80 40 8.3 b.d. 8.5
2 Great Salt lake/SS17 Oligo 66.99000 50.59833 204 31 11 2,798b 9.5 22 5.5
3 Glacier Lake 1a Silty 67.13521 50.12833 335 40 22 5 7.8 33 n.a.
4 Cube lakea/SS903 Fresh 67.12800 50.17470 335 37 32 115 8.4 2 9
5 Lake Braya/SS04 Oligo 66.98830 51.04670 175 73 23 2,636b 9.3 7 5.5
Abbreviations a.s.l., above sea level; b.d., below detection; fresh, freshwater; n.a., not available; oligo, oligosalinea Unofficial names, SS number refers to numbers used in previous publications (Brodersen & Anderson, 2002; Reuss et al., 2013b)b Conductivity measurements based on average measures from 1996 to 2000 (Brodersen & Anderson, 2002)
Hydrobiologia
123

and analyzed by high performance liquid chromatog-
raphy (HPLC) on a Shimadzu Prominence HPLC
equipped with an on-line photodiode array detector
(SPD-M20A PDA), as described by Reuss & Conley
(2005). Identification and quantification of the pig-
ments were conducted as a combination of retention
time and absorption spectra compared to standards
obtained from DHI, Hørsholm, Denmark and pub-
lished absorption spectra (Jeffrey et al., 1997). Filter
samples were extracted in only 3 ml extraction solvent
and not diluted due to small pigment concentrations
and retained water on the filter. For further character-
ization of the habitats, microalgae were sampled from
stone scrapes and in the surface water by use of a
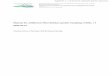
Fig. 1 Cube Lake/SS903
(a), southwest Greenland
and five examples of micro
habitats (b–f) therein. The
upper littoral in Cube Lake
was dominated by mosses,
Calliergon giganteum (b),
followed by Potamogegon
spp. (c) with depth. In
between the Potamogeton,
small patches of Chara
(d) and exposed stone
surfaces (e) occurred. Note
the numerous chironomid
tubes on the surface of the
stone (e). On the deeper
slopes of micro habitats c–e,
patches of bare, soft
sediment (f) appeared.
Photos by the authors
Hydrobiologia
123

20 lm net, and fixed in Lugol. Herbaria were prepared
of each macrophyte for subsequent identification.
Chironomids
Chironomids from stones, sediment, and macrophytes
were sampled with a 200-lm hand net, while the
profundal was sampled using an Ekman grab. The
collected chironomid larvae were allowed to defecate in
filtered lake water for at least 24 h in the refrigerator
before identification, enumeration, and sub-sampling
(preferably minimum three individuals, range 1–22).
Specimens were dried for minimum 24 h at 60�C and
stored at room temperature until preparation for isotope
analyses. Whole individuals or heads of chironomid
larvae (1–6 individuals) from each taxonomic group
were fixed in 70% ethanol and subsequently mounted
for further identification. Identification of the larvae was
supported by collection and identification of pupal
exuviae from the lakes. Pre-imaginal stages of chiron-
omids were identified according to standard manuals
(Wiederholm, 1983, 1986; Langton, 1991; Brooks et al.,
2007).
Isotope analyses
Isotope analyses of dried samples were conducted, as
described in Reuss et al. (2013b). In short, dry samples
for isotope analyses of terrestrial and aquatic plant
material were ground in a ball mill. Sediment, stone
scrapes, and animals were homogenized using a
mortar or spoon or the entire sample was used
depending on sample size. Animals and terrestrial
material were weighted in tin capsules. Lake sediment,
stone scrapes, and aquatic plant material were placed
in open silver capsules and acid-fumed overnight in a
desiccator with a beaker of concentrated HCl to
remove inorganic carbon. Filters with POM were
similarly fumed before packing into tin capsules.
Analyses of d13C, d15N, C%, and N% were conducted
on an isoprime isotope ratio mass spectrometer
(Micromass-GV Instruments, Manchester, UK) cou-
pled to a Eurovector CN elemental analyzer (Milan,
Italy) at the Department of Biology, University of
Copenhagen, Denmark. C/N ratios are expressed as
weight ratio, and the isotope data are presented in the
d-notation; d = [Rsample/Rstd - 1] * 1,000 where
R signifies the 13C/12C and 15N/14N ratios in the
samples and VPDB and AIR standards, respectively.
Reproducibility was better than ±0.1% for d13C and
±0.2% d15N. The d15N data are not included in the
current paper but have been addressed in a previous
paper (Reuss et al. 2013b). A few chironomid isotopic
samples were excluded from further analyses due to
low sample weight, which can bias the isotopic signal.
Data analyses
For statistical analyses, square root transformed rela-
tive abundance of chironomid taxa was used. The
distribution of species and habitat data was examined
by detrended correspondence analysis (DCA) in order
to determine the first axis gradient length. A gradient
length of 4.45 standard deviations units of turnover
(SD) indicated that it was appropriate to use unimodal
response functions. Canonical correspondence ana-
lysis (CCA) was employed to discover significant
variables driving the distribution of taxa in both lake
types together and separately. In constrained analysis
of the individual lake types, the following variables
were tested: stones, sediments, macrophytes, Chloro-
phyll-a, depth, TP, pH, and conductivity. Habitat types
were tested as binary dummy variables and are
depicted as centroids in the ordination diagram.
Additional variables were tested in the full dataset,
including lake type (freshwater, oligosaline), altitude,
temperature, and Secchi-depth. Forward selection
with 499 random permutations was used to test the
significance of the variables. All ordinations were
performed in CANOCO 5 (ter Braak & Smilauer,
2012). In this version of CANOCO, there are
improvements to eliminate the drawbacks of forward
selection identified by Blanchet et al. (2008). Due to
their unique taxonomical composition, samples from
Glacier Lake 1 and the two profundal samples were
removed from the analyses. In addition, the moss
sample from Lake Ferguson only contained four
specimens and was removed from further analyses.
Analysis of similarity (ANOSIM, Clarke & Green,
1988) was performed to test the significance of the a
priori defined groups of lake types (freshwater and
oligosaline), as well as habitat types (stones, macro-
phytes, and sediments). Similarity percentage analysis
(SIMPER) was used to determine the contribution of
each species to the observed similarity within each
group. SIMPER allows identification of the most
important taxa for creating the observed pattern of
similarity within a defined group. Both SIMPER and
Hydrobiologia
123

ANOSIM were performed using the Community
Analysis Package 3.0 (Saeby & Henderson, 2004).
Spearman rank order correlation was performed
between the average d13C signal of all chironomids
within a microhabitat and the average d13C signal of
the microhabitat. In order to compile a combined d13C
signal for fresh and oligosaline lakes, the d13C signal
for the individual chironomid taxa was normalized as
the distance from the average littoral chironomid
signal for each individual lake. The Glacier Lake 1
chironomids were excluded from this analysis as a
normalization of the values would not be valid with
only stone samples from this lake. In order to test for
differences in the normalized d13C chironomid signal
among microhabitat types (profundal, macrophytes,
sediments, and stones), One way analysis of variance
followed by all pairwise multiple comparison proce-
dures (Holm–Sidak method) was used for the fresh-
water lakes, while Kruskal–Wallis one way analysis of
variance on ranks was used for the oligosaline lakes.
All correlation and variance analyses were conducted
using SigmaPlot for Windows version 11.0.
A simple two end-member mixing model, com-
monly used for quantification of contributions of
two food sources (e.g., Chetelat et al., 2010; Premke
et al., 2010), was used to find the contribution of
benthic algae in the diet of the chironomids in the
different microhabitats. Here, stone scrapes (STO)
represented benthic algae and pelagic POM repre-
sented a combination of pelagic and terrestrial
contribution such that
% benthic algae ¼ d13CCHI � d13CPOM
d13CSTO � d13CPOM
� 100 ð1Þ
where d13CCHI indicates the average chironomid d13C
signal of the microhabitat of interest. These calcula-
tions were only performed for the freshwater habitats
as the variation in d13C values was too limited in the
oligosaline systems. No correction for fractionation or
lipid content was conducted.
Results
Taxonomic composition
Our sampling of 24 microhabitats in five lakes yielded
a total of 2,741 chironomid specimens belonging to 23
taxa (Table 2). Twenty-two taxa (1,395 specimens)
were recorded in the three freshwater lakes (including
Glacier Lake 1) and 10 taxa (1,346 specimens) in the
two oligosaline lakes. Nine taxa were found in both
lake types, and only Tanytarsus gracilentus was
unique for oligosaline lakes, albeit at a low abundance.
The Orthocladiinae subfamily dominated both in
numbers of taxa and in abundances (Fig. 2). In the
freshwater lakes, the second most abundant subfamily
was Chironominae (especially the tribe Chironomini),
while Tanypodinae was most abundant in the oligos-
aline lakes. Moreover, while the tribe Tanytarsini
represented a substantial part of the chironomid
community of freshwater lakes (four taxa, 13.5% of
total abundance), it was negligible in the oligosaline
lakes (one taxon, 0.5% abundance). Characteristic
taxa for freshwater lakes included Dicrotendipes
modestus, Psectrocladius limbatellus group A, and
Arctopelopia melanosoma (together accounting for
67% of the total abundance). Psectrocladius sordidel-
lus, P. barbimanus groups, and Ablabesmyia pulchri-
pennis dominated (sum *80%) in the oligosaline
lakes.
The first two axes of the DCA explained 34.3% of
the variance of the biological data and distinguished
the chironomids in the oligosaline habitats from the
freshwater habitats (not shown, first axis gradient
length 4.45 SD). The freshwater habitats represent a
greater heterogeneity (first axis gradient length 3.92
SD) than the oligosaline habitats (first axis gradient
length 2.00 SD). The separation of the two lake types
was also clear from the CCA. Here, the isolated
clusters of oligosaline and freshwater lakes were
grouped along the conductivity gradient, which was
the strongest explanatory variable (Fig. 3a). ANOSIM
confirmed a significant difference in the chironomid
assemblages of freshwater and oligosaline lakes
(P = 0.002). Therefore, freshwater and oligosaline
lakes were treated separately in all further analyses.
Forward selection in CCA revealed that depth and soft
sediment substrate were significant for the chironomid
community structure in freshwater lakes (Fig. 3b, c).
These two variables explained 34.8% of the variability
of the community data. The soft sediment samples
were distinguished from the other habitat types and
grouped toward the left top part of the ordination
diagram (Fig. 3b). Stone- and macrophyte samples
were mostly overlapping and located in the middle- to
right part of the diagram along the depth gradient. In
Hydrobiologia
123

Ta
ble
2T
ota
lca
rbo
n,
nit
rog
en,
and
iso
top
icco
mp
osi
tio
no
fin
div
idu
alch
iro
no
mid
sin
clu
din
gth
eir
ob
serv
edh
abit
ats
(in
dic
ated
by
1if
pre
sen
tat
the
hab
itat
,S
edse
dim
ent,
Sto
sto
nes
,C
ha
Ch
ara
,M
os
mo
ss,
Po
tP
ota
mo
get
on
,C
laC
lad
op
ho
ra)
Nam
eaC
od
eS
ub
fam
ily
/
trib
e
Fre
qu
ency
(%)c
%C
%N
C/
N
d13C
±S
E(n
ot
no
rmal
ized
)H
abit
at
nd
Fre
shw
ater
nd
Oli
go
sali
ne
Sed
Sto
Ch
aM
os
Po
tC
la
Arc
top
elo
pia
mel
an
oso
ma
Arc
mel
Tan
yp
od
inae
25
55
.89
.85
.71
1-
23
.0±
2.1
11
11
Arc
top
elo
pia
mel
an
oso
ma
(yo
un
g
inst
ar)
Arc
mel
(y)
Tan
yp
od
inae
52
.91
0.9
4.9
7-
26
.1±
0.9
Ab
lab
esm
yia
pu
lch
rip
enn
isA
bl
pu
lT
any
po
din
ae4
65
2.1
10
.35
.13
-2
5.1
±0
.21
8-
29
.5±
0.4
11
11
1
Pro
cla
diu
s(H
.)cf
.p
ara
gre
tis
Pro
par
Tan
yp
od
inae
46
54
.78
.86
.31
0-
18
.9±
1.2
12
-3
0.7
±0
.41
11
11
Pro
cla
diu
scf
.p
ara
gre
tis
(yo
un
g
inst
ar)
Pro
par
(y)
Tan
yp
od
inae
51
.69
.95
.23
-3
0.3
±0
.0
Cri
coto
pu
s(C
.)sp
.C
risp
.O
rth
ocl
adii
nae
21
48
.21
0.1
4.8
6-
16
.0±
0.6
2-
27
.3±
0.2
11
Het
ero
tris
socl
ad
ius
ma
rcid
us
gr.
Het
mar
Ort
ho
clad
iin
ae8
54
.58
.36
.63
-1
8.5
±0
.51
1
Het
ero
tris
socl
ad
ius
sub
pil
osu
sg
r.*
Het
sub
Ort
ho
clad
iin
ae8
52
.57
.96
.76
-2
1.6
±3
.41
Hyd
rob
aen
us
cf.
fusi
styl
us
Hy
dfu
sO
rth
ocl
adii
nae
45
3.9
7.9
6.8
3-
24
.8±
0.2
1
Pse
ctro
cla
diu
sb
arb
ima
nu
sg
r.P
seb
arO
rth
ocl
adii
nae
38
54
.88
.76
.43
-2
7.1
±0
.31
8-
29
.6±
0.1
11
11
11
Pse
ctro
cla
diu
sli
mb
ate
llu
sg
r.A
Pse
sorA
Ort
ho
clad
iin
ae2
14
9.3
8.7
5.6
9-
18
.9±
1.7
11
1
Pse
ctro
cla
diu
sli
mb
ate
llu
sg
r.B
Pse
sorB
Ort
ho
clad
iin
ae3
34
8.2
9.3
5.3
13
-1
9.0
±0
.99
-2
6.6
±0
.31
11
Pse
ctro
cla
diu
sso
rdid
ellu
sg
r.P
roso
rO
rth
ocl
adii
nae
58
52
.79
.15
.96
-2
4.1
±0
.83
0-
28
.2±
0.1
11
11
11
Ort
ho
cla
dii
na
eu
nid
enti
fied
Ort
un
idO
rth
ocl
adii
nae
13
56
.49
.26
.26
-3
0.6
±0
.51
11
Ch
iro
no
mu
sa
nth
raci
us
gr.
Ch
ian
tC
hir
on
om
ini
17
40
.48
.05
.06
-2
2.6
±1
.61
11
Ch
iro
no
mu
scf
.h
yper
bo
reu
sC
hi
hy
bC
hir
on
om
ini
50
50
.89
.75
.36
-2
2.1
±2
.31
2-
26
.4±
0.6
11
11
1
Ch
iro
no
mu
sh
yper
bo
reu
s(y
ou
ng
inst
ar)
Ch
isp
.
(y)
Ch
iro
no
min
i5
1.9
11
.44
.53
-2
6.1
±0
.2
Dic
rote
nd
ipes
mo
des
tus
Dic
mo
dC
hir
on
om
ini
50
52
.39
.95
.41
7-
16
.4±
1.2
3-
25
.9±
0.2
11
11
Pa
racl
ad
op
elm
an
igri
tula
gr.
*P
arn
igC
hir
on
om
ini
17
57
.29
.46
.15
-2
2.9
±3
.31
Po
lyp
edil
um
laet
um
gr.
Po
lla
eC
hir
on
om
ini
85
0.2
10
.64
.73
-1
5.9
±0
.71
Mic
rop
sect
rasp
p.b
, *M
icsp
.T
any
tars
ini
21
52
.78
.16
.59
-1
7.7
±0
.51
Ta
nyt
ars
us
spp
.bT
ansp
.T
any
tars
ini
33
51
.28
.06
.56
-2
2.1
±1
.31
11
11
aA
few
tax
aw
ere
excl
ud
edfr
om
the
tab
led
ue
tom
issi
ng
iso
top
icsa
mp
les
(Mic
rop
sect
rasp
.C
fro
mfr
esh
wat
erla
kes
and
Ta
nyt
ars
us
gra
cile
ntu
san
dC
hir
on
om
us
an
thra
cin
us
fro
mo
lig
osa
lin
ela
kes
)b
Incl
ud
essp
ecie
sA
and
B
*In
clu
des
pro
fun
dal
spec
imen
sfr
om
Lak
eF
erg
uso
n(2
0m
)c
Fre
qu
ency
of
occ
urr
ence
inal
l2
4h
abit
atsa
mp
les
tak
end
Nu
mb
ero
fsa
mp
les
anal
yze
dfo
ris
oto
pes
Hydrobiologia
123

the oligosaline lakes, none of the tested environmental
variables was significant.
Characteristic species of each habitat type in the
two lake types are listed in Table 3. No significant
differences in the chironomid composition, abun-
dance, and taxa number were found among different
microhabitat types in the full dataset (including five
lakes), or in the freshwater and oligosaline datasets
tested separately. On the other hand, the assemblage
structure of chironomids dwelling on different sedi-
ment types differed in the two lake types (Fig. 2).
Stones supported the lowest taxa richness in the
freshwater lakes (ten taxa), with Orthocladiinae
dominating (60% of taxa). In sediment- and macro-
phyte habitats, both including 14 taxa, the distribution
of subfamilies was relatively even with members of
Chironominae slightly more abundant. However, the
abundance of Orthocladiinae and Chironominae was
similar for each habitat type, Orthocladiinae being
slightly more prevalent in sediment and on macro-
phytes and Chironominae more prevalent on stones. In
the oligosaline lakes, stones were taxonomically the
richest microhabitat (ten taxa) with subfamilies evenly
represented. However, the numerical prevalence of
Orthocladiinae in all oligosaline microhabitats was
striking and accounted for 59–80% of the total
a b
c d
e f
Fig. 2 Structure of the
chironomid assemblages.
Number of taxa (a) and
relative abundance (b) of the
subfamilies and tribes in
freshwater and oligosaline
lakes. Number of taxa
(c) and relative abundance
(d) of subfamilies and tribes
on the microhabitat types in
the freshwater lakes.
Number of taxa (e) and
relative abundance (f) of the
subfamilies and tribes on
microhabitat types in the
oligosaline lakes. Total
relative abundance was
calculated as the average of
relative abundance of
subfamilies in each sample
Hydrobiologia
123

abundance. Macrophytes supported the highest abun-
dance of the Tanypodinae in both lake types (up to
30%) with Arctopelopia melanosoma as the most
typical taxa dwelling on macrophytes in freshwater
lakes and Ablabesmyia pulchripennis (along with
Psectrocladius sordidellus group) in oligosaline lakes.
Conversely, stones included the lowest proportion of
Tanypodinae in both lake types.
Isotopic composition of habitats and chironomids
The habitats within the freshwater lakes showed large
differences in the d13C values, while significantly less
variability was found within the oligosaline lakes
(Table 4). Only the d13C signal of Potamogeton
filiformis showed distinct enriched values in the
oligosaline lakes. The d13C signal of the terrestrial
vegetation showed generally depleted values compa-
rable to the POM samples. There was a positive
relationship (r2 = 0.618, P \ 0.001, n = 22, Fig. 4)
between the average d13C signal of the chironomid
assemblages and their microhabitat. However, large
differences were observed for the individual micro-
habitats, and particularly in the macrophytes (Fig. 5).
The contribution of benthic algae to the chironomid
diet at the different microhabitats, based on a two end-
member model of the d13C signal in the freshwater
lakes, varied between 73% on the stones to 0% at the
profundal site. Moss (11%), Chara (31%), and sedi-
ment (61%) were intermediate.
In the freshwater lakes, chironomids from stones
showed the highest normalized d13C values and
profundal chironomids showed the lowest d13C values
(P \ 0.05, Fig. 6), while chironomids from macro-
phytes and sediments were intermediate. No signifi-
cant difference was found between the normalized
chironomid signal from stones, sediments or macro-
phytes in the oligosaline lakes. Due to the general
a
b
c
Fig. 3 Canonical correspondence analyses (CCA) biplots of
microhabitats (a, b) and taxa (c) of the chironomid community
composition with the significant environmental variables in the
complete dataset (a) and freshwater lakes (b, c) (excluding
Glacier Lake 1 and profundal sites). The two variables
(sediment and depth) explained 37.4% of the overall variability
of biological data in the freshwater lakes. Because sediment
(SED) represents a binary dummy variable, it is pictured as a
centroid and not as a vector. a Open circles freshwater, solid
circles oligosaline lakes. Codes of the microhabitats: lake
number—habitat type and depth (rounded to nearest meter).
Lake numbers as in Table 1. b Open circles stones (STO), solid
squares soft sediment (SED), gray diamonds macrophytes
(MOS = moss, CHA = Chara). Codes as in (a). c Chironomid
taxa codes as in Table 2
b
Hydrobiologia
123

difference in chironomid species composition between
the freshwater and oligosaline lakes, a composite
figure of each lake type was created with taxa specific
d13C values normalized to the distance from the mean
littoral d13C of the lake (Fig. 7). The non-normalized
d13C signal for the individual chironomid taxa in the
different lake types is presented in Table 2.
Primary producers of the habitats
Pigment analyses and microscopic investigation of the
microhabitats and POM samples showed that the
primary producers in the water column were diverse.
Particularly, the freshwater lakes included specimens
and pigments characteristic of chrysophytes, green
algae, and dinoflagellates (Table 4). The stone scrapes
contained a mix of diatoms, cyanobacteria, and green
algae, the latter dominating in the oligosaline lakes.
The presence of okenone in the profundal samples
from the oligosaline lakes formed an important
distinction compared to the freshwater lakes. Okenone
is a pigment of purple sulfur bacteria, indicating
hypoxic or anoxic conditions within the photic zone
(Pfennig, 1989). The pigment concentration in the
profundal of freshwater lakes was relatively low.
Discussion
There was a significant difference between the taxo-
nomic composition of chironomid communities in
freshwater lakes and in oligosaline lakes. Contrary to
our expectations, however, no significant differences
were found among communities of the littoral micro-
habitats. The d13C isotopic signal of the chironomids
reflected the wide variety of the habitat signals. In the
freshwater lakes, the chironomid signals reflected the
range in habitat isotopic signals from 13C-enriched
stone scrapes to 13C-depleted macrophytes and deep
sediments. Despite the lack of chironomid community
differences among the microhabitats, the normalized
chironomid d13C isotopic signal was significantly
different in the freshwater microhabitats, indicating an
extended ability of the chironomid community to
adapt their feeding strategy to their habitat. In other
words, ‘‘you are what you eat’’ applies also to
chironomids.
Taxonomic composition
Most chironomid taxa were found both in freshwater
and oligosaline lakes. However, the proportions of the
dominating taxa were different in the two lake types.
In freshwater lakes, soft sediment had a slightly
different assemblage composition than stones and
macrophytes, while in the oligosaline lakes, all
microhabitat types shared uniform chironomid assem-
blages. The glacially influenced lake had a unique
taxonomic composition compared to the other lakes
studied. The Orthocladiinae dominated all sediment
types of both lake types, except for the stone habitat in
the freshwater lakes, where Chironominae were
slightly more abundant. This is surprising since
Orthocladiinae often dominates rock and gravel sub-
strata along with Diamesinae, while Chironominae
and Tanypodinae are characteristic for fine sediments
(Pinder, 1980). In the freshwater lakes, depth and
occurrence of soft sediment were significant factors in
shaping the chironomid community structure. Sub-
strate type is known to have a significant influence on
chironomid assemblages within lakes (Pinder, 1980),
and in our study the soft sediment apparently repre-
sented a different microhabitat type as opposed to firm
surfaces, such as stones and macrophytes. Depth
Table 3 Characteristic chironomid taxa for different habitat types in freshwater and oligosaline lakes
Freshwater Oligosaline
Stones Macrophytes Sediment Stones Macrophytes Sediment
Dic mod Arc mel Pse lim_B Pse sor Pse sor Pse sor
Pse lim_A Dic mod Dic mod Pse bar Abl pul Pse bar
Arc mel Pse lim_A Pro par Abl pul Chi hyp Dic mod
– Chi hyp – – – –
– Pse sor – – – –
For taxon codes, see Table 2. Only taxa with more than 10% contribution to similarity are shown (SIMPER)
Hydrobiologia
123

Ta
ble
4M
icro
-hab
itat
char
acte
rist
ics,
iso
top
es,
pig
men
ts,
and
mic
rosc
op
y(e
xcl
ud
ing
Gla
cier
Lak
e1
)
Hab
itat
/up
lan
dC
od
e%
C%
NC
/Nd1
3C
Ran
ge
Pig
men
tsa
Mic
rosc
op
yb
Fre
shw
ater
Par
ticu
late
org
anic
mat
ter
(pel
agic
)
PO
M–
––
-2
8.3
(-3
0.8
to-
24
.8)
Th
in(f
uco
xan
thin
,
lute
in,
zeax
anth
in)
Ch
ryso
ph
yte
s,d
ino
-flag
ella
tes,
gre
enal
gae
Ch
ara
glo
bu
lari
sC
HA
29
.52
.31
3.0
-1
9.4
(-2
3.9
to-
15
.6)
Lu
tein
–
Mo
sses
cM
OS
29
.41
.42
2.0
-2
8.9
(-3
3.0
to-
24
.8)
Lu
tein
–
Sto
ne
scra
pe
ST
O1
7.4
1.1
15
.7-
11
.4(-
13
.5to
-8
.8)
Fu
cox
anth
in/d
iad
ino
xan
thin
Dia
tom
s,cy
an
ob
act
eria
,
gre
enal
gae
Sed
imen
tS
ED
3.3
0.3
11
.1-
20
.9(-
23
.7to
-1
7.4
)F
uco
xa
nth
in,
lute
in,
zeax
anth
in–
Pro
fun
dal
PR
O3
.10
.31
1.4
-2
6.2
(-2
7.0
to-
25
.5)
Th
insa
mp
les
–
Oli
go
sali
ne
Par
ticu
late
org
anic
mat
ter
(pel
agic
)
PO
M–
––
-3
0.0
(-3
5.1
to-
28
.2)
Th
in(l
ute
in,
fuco
xan
thin
,
allo
xan
thin
,ze
axan
thin
)
Gre
enal
gae
Po
tam
og
eto
nfi
lifo
rmis
PO
T3
7.9
2.1
19
.4-
18
.9(-
20
.7to
-1
6.6
)L
ute
in–
Cla
do
ph
ora
sp.
CL
A3
4.7
2.8
12
.2-
26
.3(-
26
.4to
-2
6.1
)L
ute
in–
Sto
ne
scra
pe
ST
O1
8.6
2.0
10
.2-
27
.0(-
27
.4to
-2
6.8
)F
uco
xa
nth
in,
lute
in,
zeax
anth
in/e
chin
eno
ne
Gre
ena
lga
e,d
iato
ms,
cyan
ob
acte
ria
Sed
imen
tS
ED
9.0
0.9
10
.8-
27
.5(-
28
.6to
-2
6.6
)F
uco
xa
nth
in,
lute
in,
zeax
anth
in–
Pro
fun
dal
PR
O7
.80
.71
0.6
-2
7.3
(-2
8.5
to-
26
.2)
Ok
eno
ne,
lute
in,
zeax
anth
in,
dia
tox
anth
in
–
Ter
rest
rial
Bet
ula
na
na
BE
T4
9.3
2.1
24
.3-
29
.1( -
30
.1to
-2
8.2
)–
–
Mo
sses
MO
S4
5.7
0.9
54
.5-
26
.4(-
28
.3to
-5
.0)
––
Po
acea
eP
OA
46
.51
.63
1.0
-2
6.9
(-2
7.9
to-
26
.0)
––
Rh
od
od
end
ron
sp.
RH
O5
1.5
1.9
26
.9-
27
.2(-
28
.5to
-2
5.4
)–
–
Sa
lix
gla
uca
SA
L5
0.2
2.6
19
.9-
28
.3(-
28
.8to
-2
7.3
)–
–
Va
ccin
ium
sp.
VA
C4
9.7
1.7
29
.3-
29
.5(-
30
.7to
-2
7.8
)–
–
Lit
ter
LIT
22
.70
.82
7.8
-2
8.3
(-2
9.1
to-
27
.5)
––
aD
om
inat
ing
caro
ten
oid
s:fu
cox
anth
in/d
iad
ino
xan
thin
/dia
tox
anth
in=
dia
tom
s/ch
ryso
ph
yte
s,lu
tein
=g
reen
alg
ae/h
igh
erp
lan
ts,
zeax
anth
in/e
chin
eno
ne
=cy
ano
bac
teri
a,
allo
xan
thin
=cr
yp
top
hy
tes,
ok
eno
ne
=p
urp
lesu
lfu
rb
acte
ria,
mo
stab
un
dan
tin
bo
ldb
Do
min
atin
gg
rou
ps
asse
ssed
by
mic
rosc
op
yin
spec
tio
n,
mo
stab
un
dan
tin
bo
ldc
Th
em
oss
esin
clu
de
Sco
rpid
ium
sco
rpio
ides
(Lak
eF
erg
uso
n)
and
Ca
llie
rgo
ng
iga
nte
um
(Cu
be
lak
e)
Hydrobiologia
123

remained of key importance, even when excluding the
profundal samples from the analyses. This suggests
that water depth, due to the strong relationship with
other environmental variables that are crucial for the
larval development of chironomids, such as macro-
phyte and algal growth, water temperature, oxygen
concentration, substrate or food quality, plays an
important role for the distribution of littoral and
sublittoral taxa (e.g., Heiri, 2004; Kurek & Cwynar,
2009; Velle et al., 2012b). Several studies have
highlighted the community structure of subfossil-
and living chironomid assemblages in the Kangerlus-
suaq region. In studies of chironomid exuviae
(Brodersen et al., 2001a) and subfossil assemblages
(Brodersen & Anderson, 2002) it was shown that
nutrients, lake depth, and salinity (conductivity) were
important shaping environmental factors. Further
studies of physiology along the salinity gradient have
also found a significant relationship between the
larvae ability to regulate the oxygen uptake (oxyreg-
ulatory capacity) and the composition of the chiron-
omid assemblage (Brodersen et al., 2004; Brodersen,
2007). The living chironomid assemblages from the
five lakes in this study do not deviate from the earlier
observations. Most lakes in the region are dimictic
with summer stratification from the end of June until
mid-August, and some of the oligosaline lakes are
meromictic (Brodersen & Anderson, 2000). Conse-
quently, samples taken below the thermocline of the
oligosaline lakes ([10 m, Great Salt Lake and Lake
Braya) will among other factors also be influenced by
oxygen depletion by the end of the stratification
season.
The chironomid fauna was significantly different in
the freshwater lakes and in the oligosaline lakes.
Different community structure of the two lake types
has previously been recognized in southwest Green-
land (Brodersen & Anderson, 2002). However, no
unique halophilic chironomid species were observed
in the oligosaline lakes, and similarly to findings of
Williams (1991), the assemblages are merely com-
prised of freshwater species tolerant to increased
salinity. An important feature of the assemblage
structure in the oligosaline lakes compared to fresh-
water lakes was, however, the substantial decrease of
the proportion of Tanytarsini in the oligosaline lakes.
A similar finding has been reported in Tibetan lakes
(Zhang et al., 2007). The considerably lower number
of taxa recorded in the oligosaline lakes compared to
the freshwater lakes is in accordance with the signif-
icant decrease of richness with increasing conductivity
found in other studies (Brodersen, 2007; Zhang et al.,
2007). The observed biodiversity decline could be
caused by tolerance limits of key aquatic macrophytes
that create a range of specialized microhabitats (Wil-
liams et al., 1990; Verschuren et al., 2000). However,
we sampled similar numbers of macrophyte micro-
habitats in each lake type, suggesting that the
decreased abundance of macrophytes is not a likely
reason for the lower diversity in the oligosaline lakes.
Brodersen (2007) argues that the low diversity along
with the lack of the true oligotrophic taxa in the
Fig. 4 Correlation between the d13C of the individual micro-
habitats and their chironomids (average ± SE). The line
indicates the 1:1 line. Circled samples are the Potamogeton
samples with high d13C of the plant and lower values of the
inhabiting chironomids
Fig. 5 Difference between the mean chironomid d13 isotopic
signal and their habitat signal. Microhabitats include macro-
phytes, sediments, and stones. Profundal and moss samples from
Lake Ferguson have been excluded
Hydrobiologia
123

profundal of the oligosaline lakes is a result of higher
productivity, incomplete mixing (due to the presence
of the pycnocline) and in turn low levels of
hypolimnetic oxygen. Higher TP and lower transpar-
ency compared to freshwater lakes indicate slightly
higher productivity of the oligosaline lakes. However,
Fig. 6 d13C of the individual chironomid taxa normalized to
the average littoral chironomid community signal on the
different habitat types of the freshwater and oligosaline lakes
(excluding Glacier Lake 1). The letters indicate significant
difference (P \ 0.05). The horizontal line median; box 25th–
75th percentiles; whiskers 10th and 90th percentiles; solid dots
outlying points
Fig. 7 d13C of the individual chironomid taxa (d13Cchi)
normalized to the average littoral chironomid community
(d13Clittoral chi) signal of the freshwater and oligosaline lakes
(excluding Glacier Lake 1). The horizontal line median; box
25th–75th percentiles; whiskers 10th and 90th percentiles; solid
dots outlying points. The number of habitats where the
individual taxa has been sampled for isotope analyses is
included as bars in the bottom of the panels (right axis)
Hydrobiologia
123

all the littoral habitats in the present study were
located above the thermocline (data not shown), and
thus oxygen depletion during stratification is not likely
to influence the chironomid assemblages and cause a
declined diversity. Because of the lack of obvious
indicator taxa for oligosaline conditions, we suggest
that an assemblage including low diversity combined
with a low abundance or lack of Tanytarsini can be
used as an indicator of increased salinity in paleoeco-
logical records.
Hydrobaenus cf. fusistylus was unique to Glacier
Lake 1 in our study, where it was dominating the
chironomid assemblage. H. fusistylus is common in
lakes and streams with direct glacier influence of the
Kangerlussuaq area (L. Hamerlik, unpublished data)
and could potentially serve as an indicator for past
glacial input.
Diet of the chironomid community
The d13C isotopic signal of the microhabitats was
reflected by the chironomid community signal, how-
ever, some differences were observed in all micro-
habitats. The observed differences between the
microhabitat and the average chironomid signal are
larger than could be expected based on fractionation
between food item and consumer, which often is
reported as under 1% (Peterson & Fry, 1987; Post,
2002). This suggests that the chironomid community
also obtain food items from outside the microhabitat or
selectively feed on particular components of the
microhabitat. The discrepancy between the d13C
isotopic signal of the chironomids and their micro-
habitat was particularly conspicuous for the Potamog-
eton habitats in the oligosaline lakes. Potamogeton
showed high d13C isotopic signals, possibly due to
incorporation of HCO3- (Peterson & Fry, 1987;
Madsen & Sand-Jensen, 1991), which was not
reflected in the chironomid community. This suggests
that the macrophytes may primarily act as a refuge or
substratum for the chironomids. This was expected
based on previous knowledge of the chironomid
feeding preferences where the macrophytes rarely
constitute a direct food source (Pinder, 1986; Berg,
1995). In general, there was little epiphyton on the
Potamogeton that could contribute as a food source,
and epiphyton on other macrophytes was either
limited or not possible to separate for isotopic
examination. Only Chara epiphytes in Lake Ferguson
were abundant enough to be analyzed separately, and
the discrepancy in the isotopic signal between the
average chironomid community and the cleaned
Chara was 2.4% while the difference was 0.1%between chironomids and the collected epiphytes.
This indicates that the chironomids preferably eat the
epiphytes, if present. The depleted values of chiron-
omids inhabiting most of the macrophyte microhab-
itats indicate a filtering strategy including pelagic
POM in the diet. For a filtrator, it makes sense to dwell
on a macrophyte as the water higher in the water
column will be less polluted from flocculating sedi-
ments. The terrestrial vegetation and litter around the
lakes that could act as a source of allochthonous
organic matter generally showed low d13C isotopic
signals close to the POM signal, suggesting that
inclusion of terrestrial or pelagic sources in the diet
would result in a lower d13C isotopic signal. The lower
d13C isotopic signal in some chironomids compared to
the POM (particularly in the oligosaline lakes) could
be caused by selective feeding on the pelagic algae
component of the POM that could be depleted in 13C
compared to the detrital component (Rautio &
Vincent, 2007). On the other hand, selective feeding
on benthic algae could result in a higher d13C isotopic
signal since benthic algae have relatively high d13C
values (France, 1995), in accordance with observa-
tions of sediment samples in the freshwater lakes in
this study. This pattern of enriched zoobenthos in the
littoral zone has also been observed in boreal lakes
from Sweden and attributed to selective feeding of
epipelic algae (Premke et al., 2010). Selective feeding
on parts of the benthic mats by the benthic feeders
including chironomids was also suggested by Rautio
& Vincent (2007).
The food source on stones, as identified by
pigments and microscopy, consisted of diatoms, green
algae, and cyanobacteria both in the freshwater, and
oligosaline lakes. The algae groups were also present
on the soft sediment. The composition of the benthic
algae communities are consistent with what has
previously been found in Arctic lakes, while cyano-
bacteria often dominate in microbial mats in Arctic
lakes (Vadeboncoeur et al., 2003; Bonilla et al., 2005;
Rautio & Vincent, 2007). Cohesive microbial mats
were not observed in any of the investigated habitats
and may not be an attractive food source, as has been
shown for zooplankton (Rautio & Vincent, 2007). The
observed benthic algae community in this study,
Hydrobiologia
123

particularly the diatoms, seems like an attractive food
source, as has been shown in studies including gut
analyses (Tokeshi & Pinder, 1985; Pinder, 1992;
Chetelat et al., 2010). This is in line with the mixing
model results from the freshwater lakes based on the
average d13C signal of the chironomid community of
the habitats and the average habitat signal. These
showed 73 and 61% benthic algae contribution to the
diet of chironomids on stones and sediment, respec-
tively. However, d13C from stone scrapes was used as
a proxy for benthic algae but consists of a mix of algae
and dead organic material, suggesting that the pro-
portion of benthic algae could be overestimated as a
‘clean’ algae signal may be even higher. The d13C
signal of POM from pelagic/terrestrial organic matter
is also mixed, but seems like a reliable end-member in
these systems. Microscopic investigations of the
planktonic algae showed dominance of chrysophytes
in the freshwater lakes, while green algae dominated
in the oligosaline lakes, consistent with what has been
found elsewhere in the Arctic (Bonilla et al., 2005,
2009). The contribution of benthic algae in the diet of
profundal chironomids seems negligible according to
the mixing model results. A combination of autoch-
thonous and allochthonous compounds including both
primary and secondary organisms from the water
column and sediment likely constitutes the diet of
these chironomids. While the net tows constitute a
spot measurement of the plankton community, pro-
fundal sediments of the oligosaline lakes confirmed
general dominance of green algae, as well as hypoxic
or anoxic conditions in these lakes. Low oxygen
conditions are suggested by the presence of the
pigment okenone from phototrophic purple sulfur
bacteria (Pfennig, 1989; McGowan et al., 2008; Reuss
et al., 2013a).
The d13C isotopic signal of the chironomids could
have been affected by factors unrelated to the micro-
habitats or diet, e.g., the lipid content of the chiron-
omids, which can lead to a decreased d13C isotopic
signal (Kling et al., 1992; Post et al., 2007). However,
no significant difference was observed in the d13C
isotopic signal with or without lipid removal for
zooplankton in a study from Arctic and Subarctic
Canada (Rautio & Vincent, 2007). In addition, feeding
on methane-oxidizing bacteria has been shown to
cause a d13C depleted signal of chironomids in the
tribe Chironomini (Jones et al., 2008; van Hardenb-
roek et al., 2012).
Habitat influence
Due to the differences in the d13C isotopic values and
the variation within the fresh versus the oligosaline
lakes, the two lake types were treated separately in the
analyses of microhabitats. A common problem when
comparing isotopic signal between lakes, or even
within the same lake types, is differences in the
absolute isotopic values (Vander Zanden & Rasmus-
sen, 1999; Post, 2002; Bade et al., 2004). Therefore,
the d13C isotopic values of the individual chironomid
samples were normalized to the average littoral
chironomid community signal of the specific lake.
This procedure accounts for the different off-set in the
isotopic values among lakes, while retaining the
absolute differences among habitats. As found for
the chironomids in the freshwater lakes in this study,
there is usually a clear difference in the d13C signal of
the littoral compared to the profundal or pelagic
sources, and this has been central in previous studies of
carbon sources for different animal groups of boreal to
Arctic lake ecosystems (Karlsson & Bystrom, 2005;
Rautio & Vincent, 2007; Chetelat et al., 2010; Premke
et al., 2010). Within the littoral zone, the stone scrapes,
where benthic algae are expected to constitute the
highest proportion of the organic matter, have the
highest d13C values. The lower d13C values found in
the sediment reflect the input of other sources of
organic matter, including POM from the water column
and terrestrial material. The observed difference in the
d13C signal of the chironomids could therefore be a
useful proxy of benthic algal production. However, the
difference between littoral and profundal is stronger
than the within-littoral habitat differences. In the
oligosaline lakes, the d13C values were more similar
among habitats compared to the freshwater lakes,
possibly due to extensive availability of recycled CO2
that cause a depleted d13C isotopic signal (Peterson &
Fry, 1987). A shift in the contribution from allochth-
onous sources versus benthic production due to
changes in the light conditions caused by the DOC
content could also affect the d13C signal (Premke
et al., 2010).
In order to gain more insight into the chironomid
community, we investigated the normalized d13C
isotopic signal of the individual taxa from the two
lake types. Normalizing the isotopic signal and
lumping data from lakes with different depth and size
can influence the results. However, the good
Hydrobiologia
123

correspondence of the assemblage structure within the
freshwater and oligosaline lakes indicates that this
generalization is valid for the littoral habitats. Both in
the freshwater and oligosaline lakes, a gradual change
in normalized d13C was observed and with some taxa
having large variability. This gradual distribution of
the d13C signal from the chironomid taxa is likely
caused by the strong influence of the habitats from
where the individual chironomids were sampled and
also that many of the chironomid taxa were ubiquitous
and found in several habitats. The only clearly lower
signal was found for the chironomid specimens
collected in the profundal (20 m) of the Lake Ferguson
embayment. The wide distribution of the chironomid
taxa in the microhabitats together with the observed
differences in their isotopic signal implies that several
chironomid taxa are generalist consumers that can
switch among habitats and among diets.
The range of the d13C signature from chironomids
in the freshwater lakes was generally higher than the
range in the oligosaline lakes. Most of the taxa with the
broadest signature range, such as Arctopelopia cf.
melanosoma, Psectrocladius limbatellus sp. A, P.
limbatellus sp. B, Procladius cf. paragretis, and
Dicrotendipes modestus were also the most abundant
taxa that were found in most microhabitat types. Some
of those taxa are versatile in feeding habits (e.g.,
Dicrotendipes; Moog, 1995), yet others are specialists
(e.g., Arctopelopia). This suggests that the broad range
in the isotopic signatures is not explained by the
feeding variability of single specimen but rather by the
range in habitat types occupied by many specimens. In
the oligosaline lakes, the isotopic signature ranges
were much smaller even though the average occupa-
tion rate was considerably higher than in freshwater
lakes (53 and 27%, respectively). However, the low
d13C variability of oligosaline microhabitats is
reflected in the low d13C signal variability of the
chironomid taxa. Despite the variability within the
individual chironomid taxa, the overall pattern of
chironomids, particularly in the freshwater lakes, may
be valuable in evaluating their preference of food
items and possibly reconstruction of availability of
benthic algae production.
Perspectives for ecology and paleolimnology
Chironomids have gained a strong position as indica-
tors of long-term climate change studies due to their
wide distribution and broad ecological requirements,
combined with good preservation of their chitinous
head capsules in the lake sediment (Battarbee, 2000).
However, several studies have shown limited consis-
tency among temperature reconstructions based on
subfossil chironomid assemblages (Velle et al., 2005,
2012a) and other indicators (Bigler et al., 2002; Rosen
et al., 2003; Velle et al., 2010a) on Holocene time
scales. These differences were likely due to confound-
ing effects associated with ecological mechanisms and
in-lake processes, i.e., food and habitat availability
(Brodersen & Quinlan, 2006). Ecological information
from studies, such as the current, is therefore needed to
understand the mechanisms controlling the chirono-
mids and improve the information that can be
extracted from paleolimnological studies (Saros,
2009; Velle et al., 2010b; Eggermont & Heiri, 2012).
A weighted average of the d13C isotopic values of
chironomid assemblages in sediment samples could
potentially be used as a proxy for ‘‘littoral benthic
production.’’ Such information would be useful in
combination with paleoenvironmental inferences from
other proxies to help disentangle inconsistencies in
paleoenvironmental reconstructions, in addition to
providing important ecological information on the
lake. A higher than average d13C isotopic value would
indicate more use of benthic algae while lower than
average values would indicate a larger contribution to
the diet from pelagic and/or terrestrial sources.
However, due to the overwhelming influence on the
isotopic signal of profundal specimens, it would be
necessary to make an estimate of the contribution of
profundal versus littoral species in long-core samples
before constructing and interpreting a weighted aver-
age isotope signal.
Studies, such as the present, also help disentangle
the complicated and at times unknown microhabitat—
food source interactions of particular taxa and high-
light the apparent ability of chironomids to adapt to the
available food source. Existing information on chir-
onomids often includes family/subfamily resolution
and such a coarse resolution can bias food-web studies
wherever chironomids constitute a major part of the
aquatic biota (e.g., Lindegaard, 1992), such as in the
Arctic.
Acknowledgements This work was supported by a STENO-
Grant from The Danish Council for Independent Research—
Natural Sciences to N. S. Reuss. We also thank the Danish
Hydrobiologia
123

National Research Foundation for supporting the activities
within the Center for Permafrost (CENPERM DNRF100). We
are grateful to two anonymous reviewers whose comments to a
previous version considerably improved the paper.
References
Anderson, N. J., K. P. Brodersen, D. B. Ryves, S. McGowan, L.
S. Johansson, E. Jeppesen & M. J. Leng, 2008. Climate
versus in-lake processes as controls on the development of
community structure in a low-Arctic lake (South-West
Greenland). Ecosystems 11: 307–324.
Bade, D. L., S. R. Carpenter, J. J. Cole, P. C. Hanson & R.
H. Hesslein, 2004. Controls of d13C-DIC in lakes: geo-
chemistry, lake metabolism, and morphometry. Limnology
and Oceanography 49: 1160–1172.
Battarbee, R. W., 2000. Palaeolimnological approaches to cli-
mate change, with special regard to the biological record.
Quaternary Science Reviews 19: 107–124.
Berg, M. B., 1995. Larval food and feeding behavior. In Ar-
mitage, P. D., P. S. Cranston & L. C. V. Pinder (eds), The
Chironomidae: Biology and Ecology of Non-biting Mid-
ges. Chapman and Hall, London: 136–168.
Bigler, C., I. Larocque, S. M. Peglar, H. J. B. Birks & R. I. Hall,
2002. Quantitative multiproxy assessment of long-term
patterns of holocene environmental change from a small
lake near Abisko, Northern Sweden. Holocene 12:
481–496.
Blanchet, F. G., P. Legendre & D. Borcard, 2008. Forward
selection of explanatory variables. Ecology 89:
2623–2632.
Bonilla, S., V. Villeneuve & W. F. Vincent, 2005. Benthic and
planktonic algal communities in a high Arctic Lake: pig-
ment structure and contrasting responses to nutrient
enrichment. Journal of Phycology 41: 1120–1130.
Bonilla, S., M. Rautio & W. F. Vincent, 2009. Phytoplankton
and phytobenthos pigment strategies: implications for algal
survival in the changing Arctic. Polar Biology 32:
1293–1303.
Brodersen, K. P., 2007. Chironomids (Diptera) from sub-saline
lakes in West Greenland: diversity, assemblage structure
and respiratory adaptation. In Andersen, T. (ed.), Contri-
butions to the Systematics and Ecology of Aquatic Diptera
– A Tribute to Ole A. Sæther. The Caddis Press, Columbus:
61–68.
Brodersen, K. P. & N. J. Anderson, 2000. Subfossil insect
remains (Chironomidae) and lake-water temperature
inference in the Sisimiut-Kangerlussuaq region, Southern
West Greenland. Geology of Greenland Survey Bulletin
186: 78–82.
Brodersen, K. P. & N. J. Anderson, 2002. Distrubution of
chironomids (Diptera) in low Arctic West Greenland lakes:
trophic conditions, temperature and environmental recon-
struction. Freshwater Biology 47: 1137–1157.
Brodersen, K. P. & R. Quinlan, 2006. Midges as palaeoindica-
tors of lake productivity, eutrophication and hypolimnetic
oxygen. Quaternary Science Reviews 25: 1995–2012.
Brodersen, K. P., C. Lindegaard & N. J. Anderson, 2001a.
Holocene temperature and environmental reconstruction
from lake sediments in the Søndre Strømfjord region,
Southern West Greenland. Geology of Greenland Survey
Bulletin 189: 59–64.
Brodersen, K. P., B. V. Odgaard, O. Vestergaard & N.
J. Anderson, 2001b. Chironomid stratigraphy in the shal-
low and eutrophic Lake Sobygaard, Denmark: chirono-
mid–macrophyte co-occurrence. Freshwater Biology 46:
253–267.
Brodersen, K. P., O. Pedersen, C. Lindegaard & K. Hamburger,
2004. Chironomids (Diptera) and oxy-regulatory capacity:
an experimental approach to paleolimnological interpre-
tation. Limnology and Oceanography 49: 1549–1559.
Brooks, S. J., P. G. Langdon & O. Heiri, 2007. The Identification
and Use of Palaearctic Chironomidae Larvae in Palaeo-
ecology. QRA Technical Guide No. 10. Quaternary
Research Association, London.
Chetelat, J., L. Cloutier & M. Amyot, 2010. Carbon sources for
lake food webs in the Canadian High Arctic and other
regions of Arctic North America. Polar Biology 33:
1111–1123.
Clarke, K. R. & R. H. Green, 1988. Statistical design and ana-
lysis for a biological effects study. Marine Ecology-Pro-
gress Series 46: 213–226.
Danks, H. V., 1981. Arctic arthropods: a review of systematics
and ecology with particular reference to the North Amer-
ican fauna. Entomological Society of Canada, Ottawa.
Eggermont, H. & O. Heiri, 2012. The chironomid-temperature
relationship: expression in nature and palaeoenvironmental
implications. Biological Reviews 87: 430–456.
France, R. L., 1995. Differentiation between littoral and pelagic
food webs in lakes using stable carbon isotopes. Limnology
and Oceanography 40: 1310–1313.
Hecky, R. E. & R. H. Hesslein, 1995. Contributions of benthic
algae to lake food webs as revealed by stable isotope
analysis. Journal of the North American Benthological
Society 14: 631–653.
Heggen, M. P., H. H. Birks & N. Anderson, 2010. Long-term
ecosystem dynamics of a small lake and its catchment in
west Greenland. Holocene 20: 1207–1222.
Heiri, O., 2004. Within-lake variability of subfossil chironomid
assemblages in shallow Norwegian lakes. Journal of Pa-
leolimnology 32: 67–84.
Jeffrey, S. W., R. F. C. Mantoura & T. Bjørnland, 1997. Data for
the identification of 47 key phytoplankton pigments. In
Jeffrey, S. W., R. F. C. Mantoura & S. W. Wright (eds),
Phytoplankton Pigments in Oceanography. UNESCO,
Paris: 447–559.
Jones, R. I., C. E. Carter, A. Kelly, S. Ward, D. J. Kelly & J.
Grey, 2008. Widespread contribution of methane-cycle
bacteria to the diets of lake profundal chironomid larvae.
Ecology 89: 857–864.
Karlsson, J. & P. Bystrom, 2005. Littoral energy mobilization
dominates energy supply for top consumers in subarctic
lakes. Limnology and Oceanography 50: 538–543.
Kling, G. W., B. Fry & W. J. Obrien, 1992. Stable isotopes and
planktonic trophic structure in Arctic Lakes. Ecology 73:
561–566.
Kurek, J. & L. C. Cwynar, 2009. Effects of within-lake gradients
on the distribution of fossil chironomids from maar lakes in
western Alaska: implications for environmental recon-
structions. Hydrobiologia 623: 37–52.
Hydrobiologia
123

Langton, P. H., 1991. A key to pupal exuviae of West Palae-
arctic Chironomidae. Private Publication, Huntingdon.
Lindegaard, C., 1992. Zoobenthos ecology of Thingvallavatn –
vertical-distribution, abundance, population-dynamics and
production. Oikos 64: 257–304.
Madsen, T. V. & K. Sand-Jensen, 1991. Photosynthetic carbon
assimilation in aquatic macrophytes. Aquatic Botany 41:
5–40.
McGowan, S., D. B. Ryves & N. J. Anderson, 2003. Holocene
records of effective precipitation in West Greenland. The
Holocene 13: 239–249.
McGowan, S., R. K. Juhler & N. J. Anderson, 2008. Autotrophic
response to lake age, conductivity and temperature in two
West Greenland lakes. Journal of Paleolimnology 39:
301–317.
Moog, O. 1995. Fauna Aquatica Austriaca. A Comprehensive
Species Inventory of Austrian Aquatic Organisms with
Ecological Notes. Wasserwirtschaftskataster, Bundesmin-
isterium fur Land- und Forstwirtschaft, Wien.
Moore, J. W., 1981. Factors influencing the species composi-
tion, distribution and abundance of benthic invertebrates in
the profundal zone of a eutrophic Northern Lake. Hydro-
biologia 83: 505–510.
Peterson, B. J. & B. Fry, 1987. Stable isotopes in ecosystem
studies. Annual Review of Ecology and Systematics 18:
293–320.
Pfennig, N., 1989. Ecology of phototrophic purple and green
sulfur bacteria. In Schlegel, H. G. & B. Bowien (eds),
Autotrophic Bacteria. Science Tech Publishing, Madison:
97–116.
Pinder, L. C. V., 1980. Spatial distribution of Chironomidae in
an English chalk stream. In Murray, D. A. (ed.), Chiro-
nomidae. Ecology, Systematics, Cytology and Physiology.
Proceedings of the VIIth international Chironomid sym-
posium. Pergamon Press, Oxford: 153–161.
Pinder, L. C. V., 1986. Biology of freshwater Chironomidae.
Annual Review of Entomology 31: 1–23.
Pinder, L. C. V., 1992. Biology of epiphytic Chironomidae
(Diptera, Nematocera) in chalk streams. Hydrobiologia
248: 39–51.
Pinder, L. C. V., 1995. The habitats of chironomid larvae. In
Armitage, P. D., P. S. Cranston & L. C. V. Pinder (eds), The
Chironomidae: biology and ecology of non-biting midges.
Chapman and Hall, London: 107–135.
Post, D. M., 2002. Using stable isotopes to estimate trophic
position: models, methods, and assumptions. Ecology 83:
703–718.
Post, D. M., C. A. Layman, D. Arrington, G. Takimoto, J.
Quattrochi & C. G. Montana, 2007. Getting to the fat of the
matter: models, methods and assumptions for dealing with
lipids in stable isotope analyses. Oecologia 152: 179–189.
Premke, K., J. Karlsson, K. Steger, C. Gudasz, E. von Wachen-
feldt & L. J. Tranvik, 2010. Stable isotope analysis of ben-
thic fauna and their food sources in boreal lakes. Journal of
the North American Benthological Society 29: 1339–1348.
Rautio, M. & W. F. Vincent, 2007. Isotopic analysis of the
source of organic carbon for zooplankton in shallow Sub-
arctic and Arctic waters. Ecography 30: 77–87.
Reuss, N. & D. J. Conley, 2005. Effects of sediment storage
conditions on pigment analyses. Limnology and Ocean-
ography: Methods 3: 477–487.
Reuss, N. S., N. J. Anderson, S. C. Fritz & G. L. Simpson, 2013a.
Responses of microbial phototrophs to late-Holocene
environmental forcing of lakes in south-west Greenland.
Freshwater Biology 58: 690–704.
Reuss, N. S., L. Hamerlik, G. Velle, A. Michelsen, O. Pedersen
& K. P. Brodersen, 2013b. Stable isotopes reveal that
chironomids occupy several trophic levels within West
Greenland lakes: implications for food web studies. Lim-
nology and Oceanography 58: 1023–1034.
Rosen, P., U. Segerstrom, L. Eriksson & I. Renberg, 2003. Do
diatom, chironomid, and pollen records consistently infer
Holocene July air temperature? A comparison using sedi-
ment cores from four alpine lakes in northern Sweden.
Arctic Antarctic and Alpine Research 35: 279–290.
Saeby, R. M. H. & P. A. Henderson, 2004. Commuity Analysis
Package 3.0. Pisces Conservation Ltd, Lymington.
Saros, J., 2009. Integrating neo- and paleolimnological
approaches to refine interpretations of environmental
change. Journal of Paleolimnology 41: 243–252.
Strickland, J. D. H. & T. R. Parsons, 1968. A Practical Hand-
book of Seawater Analysis. Bulletin No. 167. Fisheries
Research Board of Canada, Ottawa.
ter Braak, C. J. F. & P. Smilauer, 2012. Canoco Reference
Manual and User’s Guide: Software for Ordination, Ver-
sion 5.0. Microcomputer Power, Ithaca.
Tokeshi, M. & L. C. V. Pinder, 1985. Microhabitats of stream
invertebrates on 2 submersed macrophytes with contrasting
leaf morphology. Holarctic Ecology 8: 313–319.
Tolonen, K. T., H. Hamalainen, I. J. Holopainen & J.
Karjalainen, 2001. Influences of habitat type and envi-
ronmental variables on littoral macroinvertebrate com-
munities in a large lake system. Archiv fur
Hydrobiologie 152: 39–67.
Vadeboncoeur, Y., E. Jeppesen, M. J. Vander Zanden, H.
H. Schierup, K. Christoffersen & D. M. Lodge, 2003. From
Greenland to Green Lakes: cultural eutrophication and the
loss of benthic pathways in lakes. Limnology and Ocean-
ography 48: 1408–1418.
van Hardenbroek, M., A. Lotter, D. Bastviken, N. Duc & O.
Heiri, 2012. Relationship between d13C of chironomid
remains and methane flux in Swedish lakes. Freshwater
Biology 57: 166–177.
Vander Zanden, M. J. & J. B. Rasmussen, 1999. Primary con-
sumer d13C and d15N and the trophic position of aquatic
consumers. Ecology 80: 1395–1404.
Velle, G., S. J. Brooks, H. J. B. Birks & E. Willassen, 2005.
Chironomids as a tool for inferring Holocene climate: an
assessment based on six sites in southern Scandinavia.
Quaternary Science Reviews 24: 1429–1462.
Velle, G., A. E. Bjune, J. Larsen & H. J. B. Birks, 2010a.
Holocene climate and environmental history of Bru-
rskardstjørni, a lake in the catchemnt of Øvre Heimdals-
vatn, south-central Norway. Hydrobiologia 642: 13–34.
Velle, G., K. P. Brodersen, H. J. B. Birks & E. Willassen, 2010b.
Midges as quantitative temperature indicator species: les-
sons for palaeoecology. Holocene 20: 989–1002.
Velle, G., K. P. Brodersen, H. J. B. Birks & E. Willassen, 2012a.
Inconsistent results should not be overlooked: a reply to
Brooks et al. 2012. The Holocene 22: 1501–1508.
Velle, G., R. J. Telford, O. Heiri, J. Kurek & H. Birks, 2012b.
Testing intra-site transfer functions: an example using
Hydrobiologia
123

chironomids and water depth. Journal of Paleolimnology
48: 545–558.
Verschuren, D., J. Tibby, K. Sabbe & N. Roberts, 2000. Effects
of depth, salinity, and substrate on the invertebrate com-
munity of a fluctuating tropical lake. Ecology 81: 164–182.
Wiederholm, T., 1983. Chironomidae of the Holarctic region
keys and diagnoses. Part I: larvae. Entomologica Scandi-
navica Supplements 19: 457.
Wiederholm, T., 1986. Chironomidae of the Holarctic region
keys and diagnoses. Part II: pupae. Entomologica scandi-
navica Supplements 28: 147–298.
Williams, W. D., 1991. Comments on the so-called salt lakes of
Greenland. Hydrobiologia 210: 67–74.
Williams, W. D., A. J. Boulton & R. G. Taaffe, 1990. Salinity as
a determinant of salt lake fauna – a question of scale.
Hydrobiologia 197: 257–266.
Zhang, E., R. Jones, A. Bedford, P. Langdon & H. Tang, 2007. A
chironomid-based salinity inference model from lakes on
the Tibetan Plateau. Journal of Paleolimnology 38:
477–491.
Hydrobiologia
123