Embed Size (px)
Citation preview

Molecular and Cellular Pathobiology
miR-21 Induces Myofibroblast Differentiation and Promotesthe Malignant Progression of Breast Phyllodes Tumors
Chang Gong1,2, Yan Nie1,2, Shaohua Qu1,2, Jian-You Liao1, Xiuying Cui1,2, Herui Yao3, Yunjie Zeng4,Fengxi Su1,2, Erwei Song1,2, and Qiang Liu1,2
AbstractPhyllodes tumors of breast, even histologically diagnosed as benign, can recur locally and have metastatic
potential. Histologic markers only have limited value in predicting the clinical behavior of phyllodes tumors. Itremains unknown what drives the malignant progression of phyllodes tumors. We found that the expressionof myofibroblast markers, a-smooth muscle actin (a-SMA), fibroblast activation protein (FAP), and stromalcell–derived factor-1 (SDF-1), is progressively increased in the malignant progression of phyllodes tumors.Microarray showed that miR-21 was one of the most significantly upregulated microRNAs in malignantphyllodes tumors compared with benign phyllodes tumors. In addition, increased miR-21 expression wasprimarily localized to a-SMA–positive myofibroblasts. More importantly, a-SMA and miR-21 are independentpredictors of recurrence and metastasis, with their predictive value of recurrence better than histologicgrading. Furthermore, miR-21 mimics promoted, whereas miR-21 antisense oligos inhibited, the expression ofa-SMA, FAP, and SDF-1, as well as the proliferation and invasion of primary stromal cells of phyllodes tumors.The ability of miR-21 to induce myofibroblast differentiation was mediated by its regulation on Smad7 andPTEN, which regulate the migration and proliferation, respectively. In breast phyllodes tumor xenografts,miR-21 accelerated tumor growth, induced myofibroblast differentiation, and promoted metastasis. This studysuggests an important role of myofibroblast differentiation in the malignant progression of phyllodes tumorsthat is driven by increased miR-21. Cancer Res; 74(16); 4341–52. �2014 AACR.
IntroductionPhyllodes tumors of the breast are typically large and fast
growing tumors that accounts for up to 1% of all breastneoplasms (1). Although many phyllodes tumors tend tobehave in a benign manner, the clinical outcome of phyllodestumors is hard to predict, with frequent local relapse andsometimes distant metastasis. Current approach to preventrecurrence and metastasis is surgical resection with widemargin, given that adjuvant chemotherapy or radiotherapy is
not effective against phyllodes tumors (2). However, even withwide surgical resection, local recurrence rate is still as high as8% to 36% (3). Furthermore, recurrent phyllodes tumorsshowed a progression toward more malignant phenotype (4)with the acquisition of new genetic changes (5). It was reportedthat 22% of phyllodes tumors that have undergone malignanttransformation give rise to hematogenous metastasis (6). Itremains unclear what drives malignant transformation ofphyllodes tumors and existing biologic markers only have alimited value in predicting prognosis.
Phyllodes tumors, composed of an epithelial and a cellularstromal component, arefibroepithelial tumors that fall into thedisease spectrum between fibroadenoma and fibrosarcoma(7). Although all forms of phyllodes tumors are regarded ashaving malignant potential, phyllodes tumors can be histolog-ically classified as benign, borderline, ormalignant on the basisof stromal cellularity, mitotic activity of stromal cells, stromalnuclear atypia, stromal overgrowth, and type of border (infil-trating or pushing). Their potentially recurring and metasta-sizing behavior is attributed to the characteristics of stromalcells, mainly fibroblasts.
Fibroblasts are highly heterogeneous, and those isolatedfrom different sites reflect a substantial topographic diversity(8). A normal fibroblast can acquire an "activated" phenotype,which expresses a-smooth muscle actin (a-SMA) and is sonamed "myofibroblasts." Numerous growth factors, chemo-kines, and extracellular matrix (ECM)–degrading proteases
1Guangdong Provincial Key Laboratory of Malignant Tumor Epigeneticsand Gene Regulation, SunYat-Sen Memorial Hospital, SunYat-Sen Uni-versity, Guangzhou, People's Republic of China. 2Breast Tumor Center,SunYat-Sen Memorial Hospital, SunYat-Sen University, Guangzhou, Peo-ple's Republic of China. 3Department of Oncology, SunYat-Sen MemorialHospital, SunYat-Sen University, Guangzhou, People's Republic of China.4Department of Pathology, SunYat-Sen Memorial Hospital, SunYat-SenUniversity, Guangzhou, People's Republic of China.
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
C. Gong and Y. Nie contributed equally to this article.
Corresponding Authors: Qiang Liu or Erwei Song, Breast Tumor Center,SunYat-SenMemorial Hospital, SunYat-SenUniversity, 107YanjiangWestRoad, Guangzhou, 510120, People's Republic of China. Phone: 86-20-81332603; Fax: 86-20-81332853; E-mail: [email protected]; [email protected]
doi: 10.1158/0008-5472.CAN-14-0125
�2014 American Association for Cancer Research.
CancerResearch
www.aacrjournals.org 4341
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125

have been shown to mediate the activation of fibroblasts (9).Myofibroblasts are found in the stroma of many cancers,including breast cancer, colorectal cancer, and melanoma(10–12). Myofibroblasts in epithelial cancers have an increasedproliferative activity (9) and can promote cancer invasion (13).It was reported that myofibroblast differentiation also exists inthe stromal cells of some phyllodes tumors (14). However,whether the fibroblasts–myofibroblasts transition (FMT) playsa role in the malignant transformation of phyllodes tumor andwhether myofibroblast can be a prognostic marker of phyl-lodes tumors are not known yet.
microRNAs (miRNA) are noncoding RNAs that regulate geneexpression and can be master regulators of many fundamentalbiologic processes, including embryogenesiss (15) and organdevelopment (16). Our previous studies also showed thatmiRNAs play an important role in the differentiation of cancerstem cells and stromal cells (17, 18). In this study, we examinedthe role of myofibroblasts in the malignancy of phyllodestumors and investigated whether miRNA plays a role in FMT.
Patients and MethodsPatients and tissue samples
Breast phyllodes tumor samples were obtained from 268female patients with 167 benign, 36 borderline, and 65 malig-nant phyllodes tumors in the Breast TumorCenter, SunYat-SenMemorial Hospital, Sun Yat-Sen University (Guangzhou, Peo-ple's Republic of China), from January 2000 to June 2011. Thepatients were followed up for 8 to 148 months (median follow-up, 112 months). Pathologic diagnosis, as well as mitoses andstromal overgrowth status, was confirmed by two pathologistsindependently. Fresh phyllodes tumor samples were obtainedwithin 20 minutes after resection and were snap-frozen inliquid nitrogen for miRNA assay. The remaining tissues werefixed and embedded for studies of histology, immunohis-tochemistry (IHC), and in situ hybridization (ISH). All sampleswere collectedwith informed consent according to the InternalReview and Ethics Boards of SunYat-Sen Memorial Hospital.
ISH and data analysismiR-21 expression was examined by ISH on the formalin-
fixed and paraffin-embedded sections of breast phyllodestumors. Briefly, after dewaxing and rehydration, samples weredigested with proteinase K, fixed again in 4% paraformalde-hyde, hybridizedwith the 50-digoxin–labeled LNATM-modifiedmiR-21 probe (20 nmol/L, 38102–01; Exiqon) at 51�Covernight,and then incubated overnight at 4�C with anti-digoxin mono-clonal antibody (1:1,000; 11093274910; Roche Applied Science).After being stained with nitrobluetetrazolium/5-bromo-4-chloro-3-indolyl phosphate solution in the dark, the sectionswere mounted and evaluated. To determine the colocalizationof miR-21 and a-SMA, FITC-conjugated anti-digoxin mono-clonal antibody (1:200; 1120774191; Roche Applied Science) incombination with 594-conjugated goat antibodies againstmouse (1:200; A11037; Invitrogen) were used in the hybridiza-tion assays. The sections were then examined by confocalmicroscopy. The staining scores were determined on the basisof both the intensity and proportion of positive cells in 10
random fields under �400 magnification as described before(19). The staining index (SI) was calculated as follows: SI ¼staining intensity � proportion of positively stained cells. A SIscore of 4was used as a cutoff value based on the distribution offrequency of SI score formiR-21 expression and the expressionlevels of miR-21were defined as high (SI > 4) or low (SI � 4).
Separation and culture primary stromal cells frombreast phyllodes tumors
Normal breast stromal cells were extracted from the breaststroma of four samples obtained by reduction mammoplasty.Stromal cells from fibroadenoma and phyllodes tumors (8fibroadenoma, 8 benign, and 8 malignant phyllodes tumors)were isolated from tumors obtained by lumpectomies ormastectomies. Briefly, the samples were mechanically disag-gregated and digested with collagenase type III (1 mg/mL;Boehringer Mannheim) and hyaluronidase (125 U/mL; Sigma)at 37�Cwith agitation for 12 to 18 hours inDulbecco'sModifiedEagle Medium with 10% fetal calf serum. The dissociatedtissues were incubated, followed by the centrifugation at250� g for 5minutes. The pelletwas resuspended. For isolatingprimary culturedfibroblasts, isolated cells abovewere followedby differential sedimentation, plating, and growth in highserum media conditions that select for fibroblast growth.Fibroblast was then expanded and stored when cells under-went two to three population doublings within total 8 to 10days after tissue dissociation. We used fibroblasts passaged forup to five population doublings for subsequent experiments, tominimize clonal selection and culture stress that could occurduring extended tissue culture.
Animal experimentAll procedures of animal experiments were approved by the
Animal Care andUse Committee of Sun Yat-SenUniversity andconformed to the legal mandates and national guidelines forthe care and maintenance of laboratory animals. Breast phyl-lodes tumors stromal cells (1 � 107) mixed with Matrigel inequal volume were inoculated into themammary fat pads of 6-week-old female nude mice. When the xenografts were palpa-ble (around 0.5 cm in diameter), Lipofectamine alone (5 mL) orwith lin4/miR-21minics (15 mg/injection) or lin4/miR-21 anti-sense oligonucleotides (ASO; 20mg/injection) was injected intothe tumor twice a week. Tumor growth was evaluated bymonitoring tumor volume (TV¼ length� width2� 0.5) every3 days for 8 weeks. The animals were sacrificed when thexenografts reached 1.5 cm in diameter. Tumor xenografts aswell as the livers and lungs of mice were harvested for furtherevaluation. Paraffin sections (4 mm) of the harvested livers andlungs were stained with hematoxylin and eosin (H&E) forhistologic assessment; RNA and protein were extracted fromthe tumors for qRT-PCR and Western blot analysis.
Statistical analysisThe in vitro data were depicted as mean � SD of three
independent experiments performed in triplicate. All statisti-cal analyses were performed using SPSS 16.0 statistical soft-ware package (SPSS). Student t test and one-way ANOVA wereused to compare the markers of myofibroblasts and miR-21
Gong et al.
Cancer Res; 74(16) August 15, 2014 Cancer Research4342
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125

expression levels between the phyllodes tumors with differenttumor grades, whereas x2 test was used to analyze the rela-tionship between a-SMA, miR-21 expression, and clinicopath-ologic status. Kaplan–Meier curves and log-rank test were usedto compare the local recurrence-free survival (LRFS) andoverall survival (OS) in different patient groups. Spearmanorder correlations were used to measure the associationbetween different variables. Receiver operator characteristic(ROC) curves were constructed by plotting sensitivity versus(1 � specificity), and the areas under the curves (AUC) werecalculatedwith the Hanley andMcNeil method. In all cases, P <0.05 was considered statistically significant.
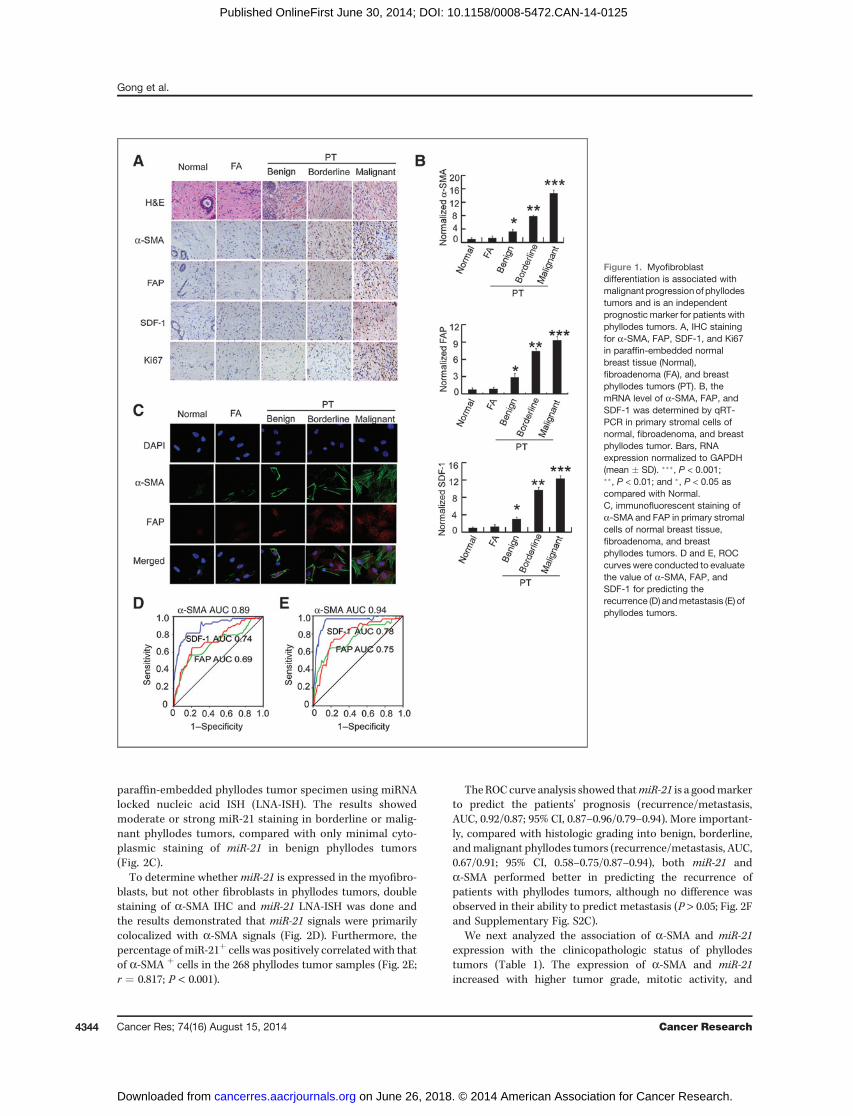
ResultsMyofibroblast differentiation is associated withmalignant progression of phyllodes tumors and is anindependent prognostic marker for phyllodes tumorpatientsTo investigate whethermyofibroblast differentiation is asso-
ciated with malignant progression of phyllodes tumors, weexamined the presence of myofibroblasts in 268 phyllodestumor samples, including 167 benign, 36 borderline, and 65malignant phyllodes tumors. Normal breast tissue and fibroa-denomawere used as control. First, we tested the expression ofKi67 and myofibroblast markers, including a-SMA, fibroblastactivation protein (FAP), and stromal cell–derived factor-1(SDF-1), in paraffin-embedded phyllodes tumor samples byIHC. Similar to previous studies (20–22), the expression of Ki67increased significantly with phyllodes tumors grade (Fig. 1A).Interestingly, the expression of a-SMA, FAP, and SDF-1 wasalso progressively increased from normal breast tissue andfibroadenoma to benign, borderline, and malignant phyllodestumors (Fig. 1A).To confirm the myofibroblast markers are overexpressed in
the stromal cells of phyllodes tumors, we measured the mRNAand protein levels of a-SMA, FAP, and SDF-1 in the primarystromal cells isolated from normal breast tissue, fibroade-noma, benign, borderline, and malignant phyllodes tumorswith eight cases per group. Using qRT-PCR, we found that themRNA levels of a-SMA, FAP, and SDF-1 were progressivelyincreased from normal breast tissue and fibroadenoma tobenign, borderline, and malignant phyllodes tumors (Fig.1B). Immunofluorescent (IF) staining (Fig. 1C) and Westernblot analysis (Supplementary Fig. S1A) also showed the similarresults at the protein levels ofa-SMA, FAP, SDF-1. ELISA assay,which measured the secretary SDF-1 protein in the medium ofstromal cells, showed that the SDF-1 level in malignant phyl-lodes tumors is significantly higher than that in benign andborderline phyllodes tumors (Supplementary Fig. S1B).Together, these results indicate that myofibroblast differenti-ation is associatedwith themalignant progression of phyllodestumors.Next, we tested whether themyofibroblast markers could be
of prognostic value for patients with phyllodes tumors. The 268patients with phyllodes tumors were followed up for 8 to 148months (median follow-up, 112months). During the follow-up,49 cases were diagnosed with recurrence, including 18 in
benign group, 9 in borderline group, and 22 in malignantgroup. In addition, 31 cases were diagnosed with metastasis,with 3 in borderline group and 28 in malignant group.
The efficacy ofa-SMA, FAP, and SDF-1 to predict recurrenceof phyllodes tumors was calculated by ROC curve, a commonlyused tool to evaluate the value of diagnostic markers (23). TheROC curve analysis showed that a-SMA performed better inpredicting the patients' prognosis [recurrence/metastasis,AUC, 0.89/0.94; 95% confidence interval (CI), 0.83–0.94/0.89–0.98] than FAP (recurrence/metastasis, AUC, 0.69/0.75; 95% CI,0.60–0.78/0.66–0.85) and SDF-1 (recurrence/metastasis, AUC,0.74/0.78; 95% CI, 0.66–0.82/0.70–0.87; Fig. 1D and E). BecauseAUC values higher than 0.8 are believed to represent gooddiscrimination (24), these results suggest that the levels ofa-SMA could be used to predict the prognosis of phyllodestumors.
SDF-1 expression levels are upregulated duringmalignant progression of phyllodes tumors
It has been reported that miRNAs are important regula-tors of key biologic processes, including differentiation (18).To investigate whether miRNA plays a role in the myofibro-blast differentiation of phyllodes tumors, we used miRNAarray to compare miRNA profiles between two benign andtwo malignant phyllodes tumors. The criteria used to screendifferentially expressed miRNAs between benign and malig-nant phyllodes tumors were fold change >3.0 or <0.33,normalized data �1 in all samples, and the expression isconsistently increased or decreased in both malignant phyl-lodes tumors than two benign phyllodes tumors. Among the1,285 human miRNAs in the array, 18 miRNAs were upre-gulated and three were downregulated in malignant phyl-lodes tumors compared with benign phyllodes tumors (Fig.2A). Among these differentially expressed miRNAs, threemiRNAs, including miR-21, miR-130b, and miR-92a, caughtour attention because these miRNAs have been reported tomodulate both differentiation and malignant progression ofcancer (25–30).
The microarray results of miR-21, miR-130b, and miR-92awere confirmed with qRT-PCR in stromal cells isolated fromeight cases each of benign and malignant phyllodes tumors.Nevertheless, the expression of miR-130b or miR-92a was alsosignificantly upregulated (Supplementary Fig. S2A) or down-regulated (Supplementary Fig. S2B) in the stromal cells offibroadenoma, respectively. Because fibroadenoma is still abenign disease and has very low chance of becoming malig-nant (31), the results indicate that these two miRNAs may notplay a significant role in the malignant transformation ofphyllodes tumors. However, the expression of miR-21 wassimilar between normal breast tissue and fibroadenoma, butwas significantly upregulated by 4.3-, 11.1-, and 20.6-fold inbenign, borderline, and malignant phyllodes tumors, respec-tively (Fig. 2B; P < 0.001). The progressive increase ofmiR-21 inmalignant progression of phyllodes tumors suggests thatmiR-21 may regulate the malignant progression of phyllodestumors and its myofibroblast differentiation.
To confirm the findings of miR-21 expression in moreclinical samples, we examined miR-21 expression in the 268
miR-21 Promotes Malignancy of Phyllodes Tumors
www.aacrjournals.org Cancer Res; 74(16) August 15, 2014 4343
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125
paraffin-embedded phyllodes tumor specimen using miRNAlocked nucleic acid ISH (LNA-ISH). The results showedmoderate or strong miR-21 staining in borderline or malig-nant phyllodes tumors, compared with only minimal cyto-plasmic staining of miR-21 in benign phyllodes tumors(Fig. 2C).
To determine whethermiR-21 is expressed in the myofibro-blasts, but not other fibroblasts in phyllodes tumors, doublestaining of a-SMA IHC and miR-21 LNA-ISH was done andthe results demonstrated that miR-21 signals were primarilycolocalized with a-SMA signals (Fig. 2D). Furthermore, thepercentage of miR-21þ cells was positively correlated with thatof a-SMA þ cells in the 268 phyllodes tumor samples (Fig. 2E;r ¼ 0.817; P < 0.001).
TheROCcurve analysis showed thatmiR-21 is a goodmarkerto predict the patients' prognosis (recurrence/metastasis,AUC, 0.92/0.87; 95% CI, 0.87–0.96/0.79–0.94). More important-ly, compared with histologic grading into benign, borderline,andmalignant phyllodes tumors (recurrence/metastasis, AUC,0.67/0.91; 95% CI, 0.58–0.75/0.87–0.94), both miR-21 anda-SMA performed better in predicting the recurrence ofpatients with phyllodes tumors, although no difference wasobserved in their ability to predict metastasis (P > 0.05; Fig. 2Fand Supplementary Fig. S2C).
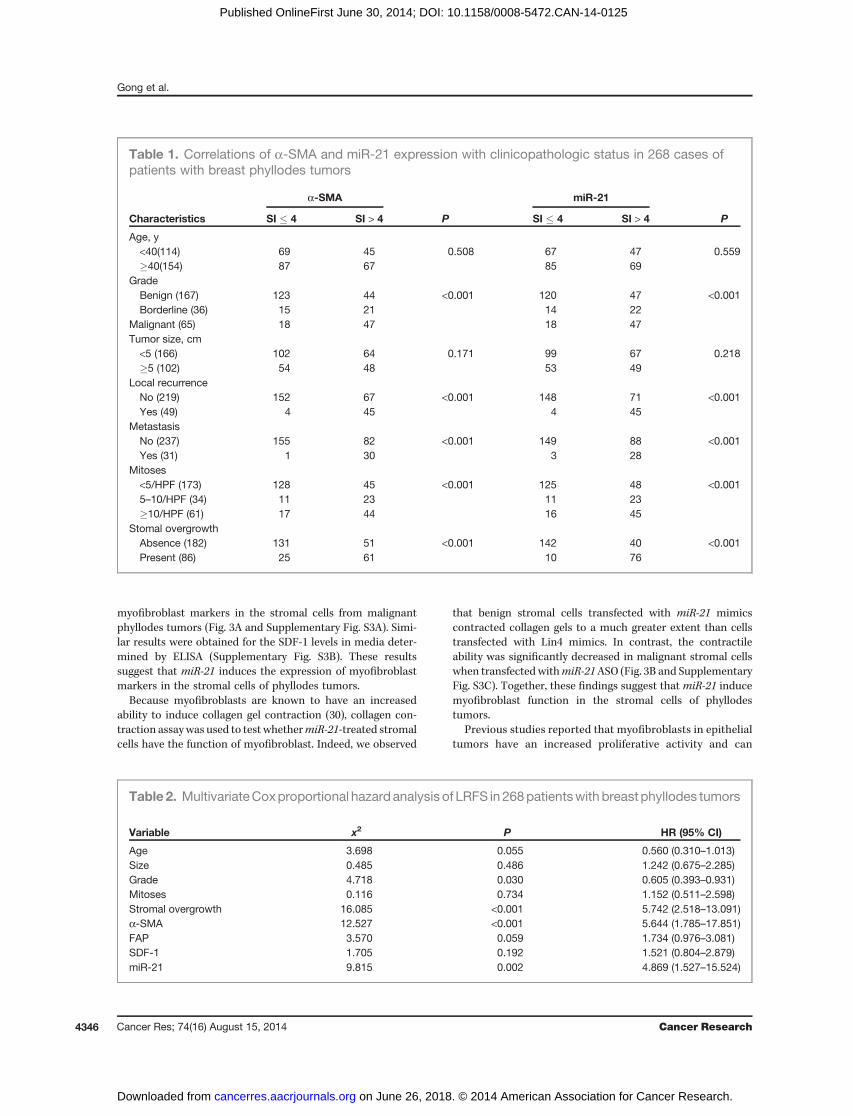
We next analyzed the association of a-SMA and miR-21expression with the clinicopathologic status of phyllodestumors (Table 1). The expression of a-SMA and miR-21increased with higher tumor grade, mitotic activity, and
Figure 1. Myofibroblastdifferentiation is associated withmalignant progressionof phyllodestumors and is an independentprognostic marker for patients withphyllodes tumors. A, IHC stainingfor a-SMA, FAP, SDF-1, and Ki67in paraffin-embedded normalbreast tissue (Normal),fibroadenoma (FA), and breastphyllodes tumors (PT). B, themRNA level of a-SMA, FAP, andSDF-1 was determined by qRT-PCR in primary stromal cells ofnormal, fibroadenoma, and breastphyllodes tumor. Bars, RNAexpression normalized to GAPDH(mean � SD). ���, P < 0.001;��, P < 0.01; and �, P < 0.05 ascompared with Normal.C, immunofluorescent staining ofa-SMA and FAP in primary stromalcells of normal breast tissue,fibroadenoma, and breastphyllodes tumors. D and E, ROCcurves were conducted to evaluatethe value of a-SMA, FAP, andSDF-1 for predicting therecurrence (D) andmetastasis (E) ofphyllodes tumors.
Gong et al.
Cancer Res; 74(16) August 15, 2014 Cancer Research4344
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125
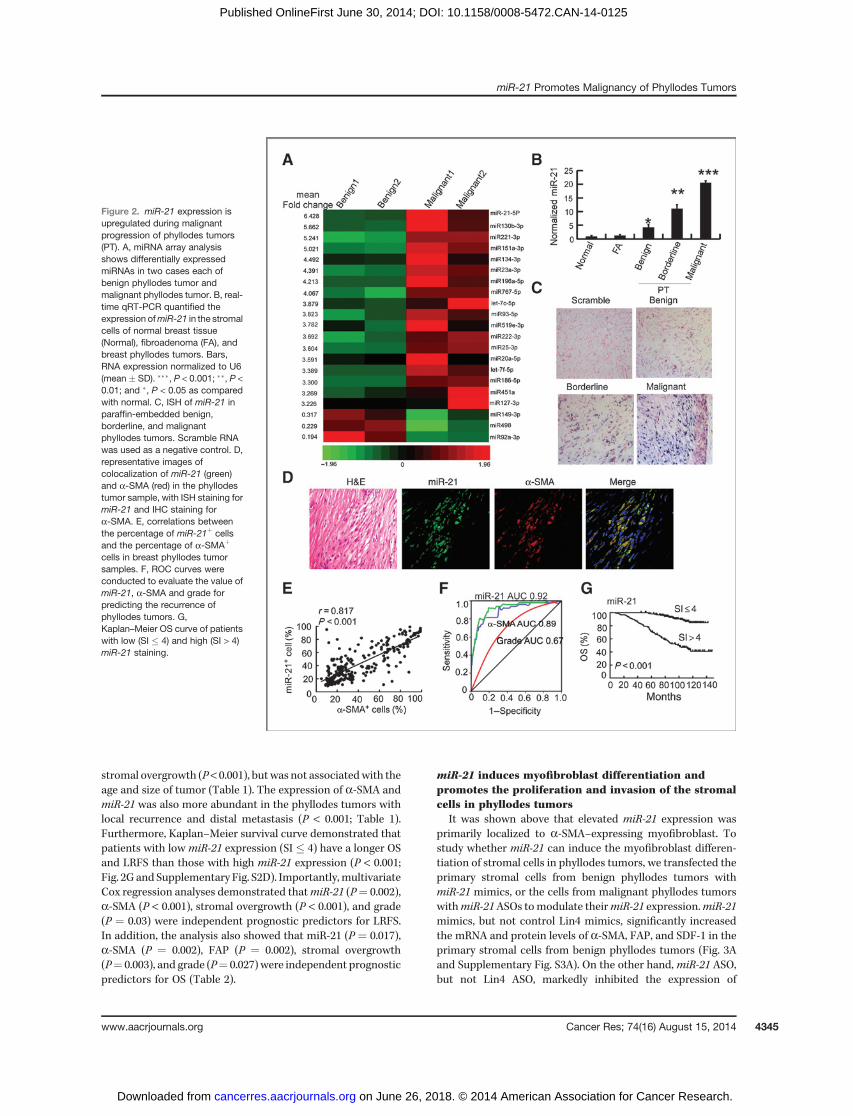
stromal overgrowth (P< 0.001), butwas not associatedwith theage and size of tumor (Table 1). The expression of a-SMA andmiR-21 was also more abundant in the phyllodes tumors withlocal recurrence and distal metastasis (P < 0.001; Table 1).Furthermore, Kaplan–Meier survival curve demonstrated thatpatients with low miR-21 expression (SI � 4) have a longer OSand LRFS than those with high miR-21 expression (P < 0.001;Fig. 2G and Supplementary Fig. S2D). Importantly,multivariateCox regression analyses demonstrated thatmiR-21 (P¼ 0.002),a-SMA (P < 0.001), stromal overgrowth (P < 0.001), and grade(P ¼ 0.03) were independent prognostic predictors for LRFS.In addition, the analysis also showed that miR-21 (P ¼ 0.017),a-SMA (P ¼ 0.002), FAP (P ¼ 0.002), stromal overgrowth(P¼ 0.003), and grade (P¼ 0.027) were independent prognosticpredictors for OS (Table 2).
miR-21 induces myofibroblast differentiation andpromotes the proliferation and invasion of the stromalcells in phyllodes tumors
It was shown above that elevated miR-21 expression wasprimarily localized to a-SMA–expressing myofibroblast. Tostudy whether miR-21 can induce the myofibroblast differen-tiation of stromal cells in phyllodes tumors, we transfected theprimary stromal cells from benign phyllodes tumors withmiR-21 mimics, or the cells from malignant phyllodes tumorswithmiR-21ASOs tomodulate theirmiR-21 expression.miR-21mimics, but not control Lin4 mimics, significantly increasedthe mRNA and protein levels of a-SMA, FAP, and SDF-1 in theprimary stromal cells from benign phyllodes tumors (Fig. 3Aand Supplementary Fig. S3A). On the other hand, miR-21 ASO,but not Lin4 ASO, markedly inhibited the expression of
Figure 2. miR-21 expression isupregulated during malignantprogression of phyllodes tumors(PT). A, miRNA array analysisshows differentially expressedmiRNAs in two cases each ofbenign phyllodes tumor andmalignant phyllodes tumor. B, real-time qRT-PCR quantified theexpression ofmiR-21 in the stromalcells of normal breast tissue(Normal), fibroadenoma (FA), andbreast phyllodes tumors. Bars,RNA expression normalized to U6(mean � SD). ���, P < 0.001; ��, P <0.01; and �, P < 0.05 as comparedwith normal. C, ISH of miR-21 inparaffin-embedded benign,borderline, and malignantphyllodes tumors. Scramble RNAwas used as a negative control. D,representative images ofcolocalization of miR-21 (green)and a-SMA (red) in the phyllodestumor sample, with ISH staining formiR-21 and IHC staining fora-SMA. E, correlations betweenthe percentage of miR-21þ cellsand the percentage of a-SMAþ
cells in breast phyllodes tumorsamples. F, ROC curves wereconducted to evaluate the value ofmiR-21, a-SMA and grade forpredicting the recurrence ofphyllodes tumors. G,Kaplan–Meier OS curve of patientswith low (SI � 4) and high (SI > 4)miR-21 staining.
miR-21 Promotes Malignancy of Phyllodes Tumors
www.aacrjournals.org Cancer Res; 74(16) August 15, 2014 4345
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125
myofibroblast markers in the stromal cells from malignantphyllodes tumors (Fig. 3A and Supplementary Fig. S3A). Simi-lar results were obtained for the SDF-1 levels in media deter-mined by ELISA (Supplementary Fig. S3B). These resultssuggest that miR-21 induces the expression of myofibroblastmarkers in the stromal cells of phyllodes tumors.
Because myofibroblasts are known to have an increasedability to induce collagen gel contraction (30), collagen con-traction assaywas used to test whethermiR-21-treated stromalcells have the function of myofibroblast. Indeed, we observed
that benign stromal cells transfected with miR-21 mimicscontracted collagen gels to a much greater extent than cellstransfected with Lin4 mimics. In contrast, the contractileability was significantly decreased in malignant stromal cellswhen transfectedwithmiR-21ASO (Fig. 3B and SupplementaryFig. S3C). Together, these findings suggest that miR-21 inducemyofibroblast function in the stromal cells of phyllodestumors.
Previous studies reported that myofibroblasts in epithelialtumors have an increased proliferative activity and can
Table 1. Correlations of a-SMA and miR-21 expression with clinicopathologic status in 268 cases ofpatients with breast phyllodes tumors
a-SMA miR-21
Characteristics SI � 4 SI > 4 P SI � 4 SI > 4 P
Age, y<40(114) 69 45 0.508 67 47 0.559�40(154) 87 67 85 69
GradeBenign (167) 123 44 <0.001 120 47 <0.001Borderline (36) 15 21 14 22
Malignant (65) 18 47 18 47Tumor size, cm<5 (166) 102 64 0.171 99 67 0.218�5 (102) 54 48 53 49
Local recurrenceNo (219) 152 67 <0.001 148 71 <0.001Yes (49) 4 45 4 45
MetastasisNo (237) 155 82 <0.001 149 88 <0.001Yes (31) 1 30 3 28
Mitoses<5/HPF (173) 128 45 <0.001 125 48 <0.0015–10/HPF (34) 11 23 11 23�10/HPF (61) 17 44 16 45
Stomal overgrowthAbsence (182) 131 51 <0.001 142 40 <0.001Present (86) 25 61 10 76
Table 2. MultivariateCoxproportional hazardanalysis of LRFS in 268patientswith breast phyllodes tumors
Variable x2 P HR (95% CI)
Age 3.698 0.055 0.560 (0.310–1.013)Size 0.485 0.486 1.242 (0.675–2.285)Grade 4.718 0.030 0.605 (0.393–0.931)Mitoses 0.116 0.734 1.152 (0.511–2.598)Stromal overgrowth 16.085 <0.001 5.742 (2.518–13.091)a-SMA 12.527 <0.001 5.644 (1.785–17.851)FAP 3.570 0.059 1.734 (0.976–3.081)SDF-1 1.705 0.192 1.521 (0.804–2.879)miR-21 9.815 0.002 4.869 (1.527–15.524)
Gong et al.
Cancer Res; 74(16) August 15, 2014 Cancer Research4346
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125
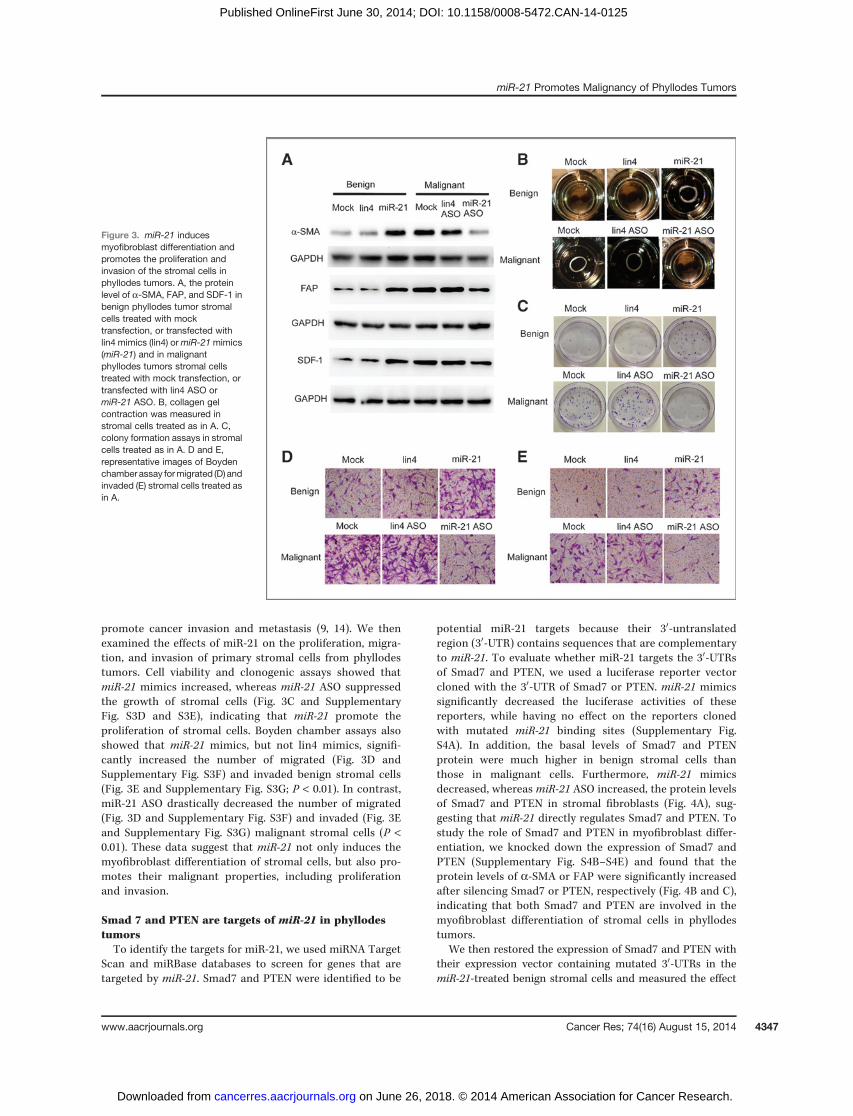
promote cancer invasion and metastasis (9, 14). We thenexamined the effects of miR-21 on the proliferation, migra-tion, and invasion of primary stromal cells from phyllodestumors. Cell viability and clonogenic assays showed thatmiR-21 mimics increased, whereas miR-21 ASO suppressedthe growth of stromal cells (Fig. 3C and SupplementaryFig. S3D and S3E), indicating that miR-21 promote theproliferation of stromal cells. Boyden chamber assays alsoshowed that miR-21 mimics, but not lin4 mimics, signifi-cantly increased the number of migrated (Fig. 3D andSupplementary Fig. S3F) and invaded benign stromal cells(Fig. 3E and Supplementary Fig. S3G; P < 0.01). In contrast,miR-21 ASO drastically decreased the number of migrated(Fig. 3D and Supplementary Fig. S3F) and invaded (Fig. 3Eand Supplementary Fig. S3G) malignant stromal cells (P <0.01). These data suggest that miR-21 not only induces themyofibroblast differentiation of stromal cells, but also pro-motes their malignant properties, including proliferationand invasion.
Smad 7 and PTEN are targets of miR-21 in phyllodestumorsTo identify the targets for miR-21, we used miRNA Target
Scan and miRBase databases to screen for genes that aretargeted by miR-21. Smad7 and PTEN were identified to be
potential miR-21 targets because their 30-untranslatedregion (30-UTR) contains sequences that are complementaryto miR-21. To evaluate whether miR-21 targets the 30-UTRsof Smad7 and PTEN, we used a luciferase reporter vectorcloned with the 30-UTR of Smad7 or PTEN. miR-21 mimicssignificantly decreased the luciferase activities of thesereporters, while having no effect on the reporters clonedwith mutated miR-21 binding sites (Supplementary Fig.S4A). In addition, the basal levels of Smad7 and PTENprotein were much higher in benign stromal cells thanthose in malignant cells. Furthermore, miR-21 mimicsdecreased, whereas miR-21 ASO increased, the protein levelsof Smad7 and PTEN in stromal fibroblasts (Fig. 4A), sug-gesting that miR-21 directly regulates Smad7 and PTEN. Tostudy the role of Smad7 and PTEN in myofibroblast differ-entiation, we knocked down the expression of Smad7 andPTEN (Supplementary Fig. S4B–S4E) and found that theprotein levels of a-SMA or FAP were significantly increasedafter silencing Smad7 or PTEN, respectively (Fig. 4B and C),indicating that both Smad7 and PTEN are involved in themyofibroblast differentiation of stromal cells in phyllodestumors.
We then restored the expression of Smad7 and PTEN withtheir expression vector containing mutated 30-UTRs in themiR-21-treated benign stromal cells and measured the effect
Figure 3. miR-21 inducesmyofibroblast differentiation andpromotes the proliferation andinvasion of the stromal cells inphyllodes tumors. A, the proteinlevel of a-SMA, FAP, and SDF-1 inbenign phyllodes tumor stromalcells treated with mocktransfection, or transfected withlin4 mimics (lin4) ormiR-21mimics(miR-21) and in malignantphyllodes tumors stromal cellstreated with mock transfection, ortransfected with lin4 ASO ormiR-21 ASO. B, collagen gelcontraction was measured instromal cells treated as in A. C,colony formation assays in stromalcells treated as in A. D and E,representative images of Boydenchamber assay formigrated (D) andinvaded (E) stromal cells treated asin A.
miR-21 Promotes Malignancy of Phyllodes Tumors
www.aacrjournals.org Cancer Res; 74(16) August 15, 2014 4347
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125
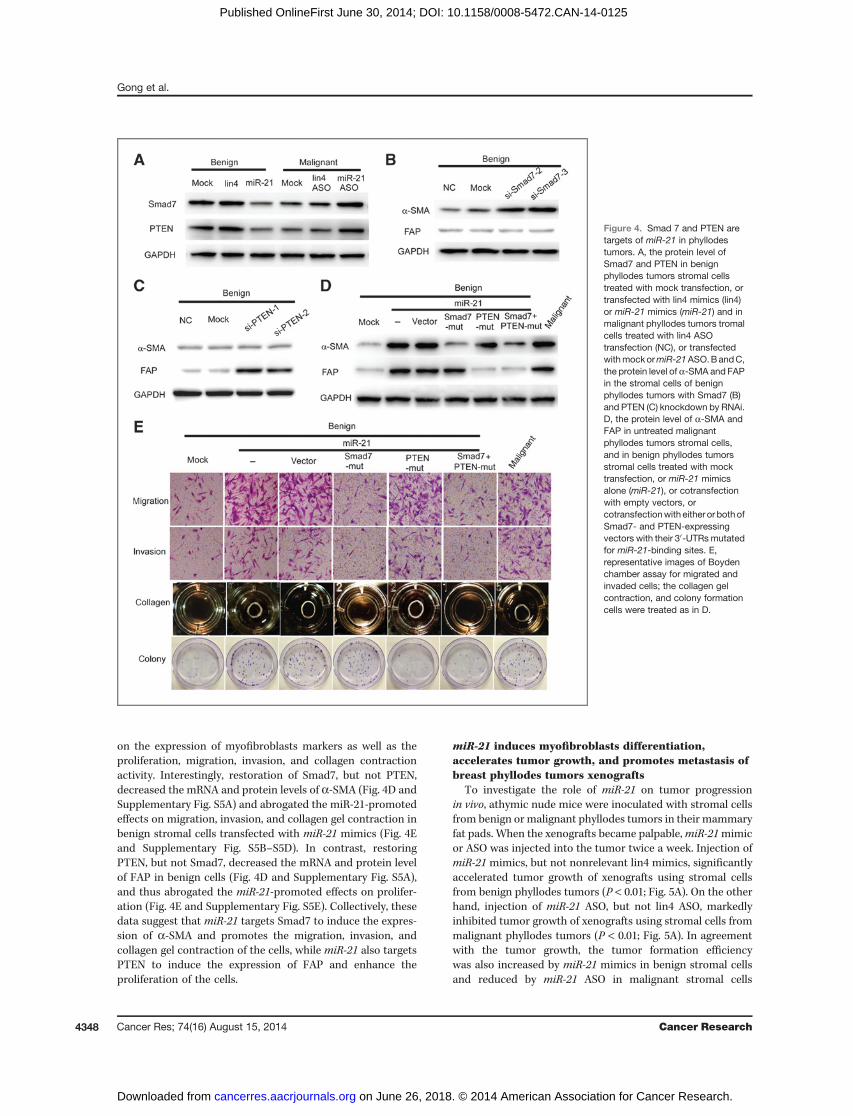
on the expression of myofibroblasts markers as well as theproliferation, migration, invasion, and collagen contractionactivity. Interestingly, restoration of Smad7, but not PTEN,decreased the mRNA and protein levels of a-SMA (Fig. 4D andSupplementary Fig. S5A) and abrogated the miR-21-promotedeffects on migration, invasion, and collagen gel contraction inbenign stromal cells transfected with miR-21 mimics (Fig. 4Eand Supplementary Fig. S5B–S5D). In contrast, restoringPTEN, but not Smad7, decreased the mRNA and protein levelof FAP in benign cells (Fig. 4D and Supplementary Fig. S5A),and thus abrogated the miR-21-promoted effects on prolifer-ation (Fig. 4E and Supplementary Fig. S5E). Collectively, thesedata suggest that miR-21 targets Smad7 to induce the expres-sion of a-SMA and promotes the migration, invasion, andcollagen gel contraction of the cells, while miR-21 also targetsPTEN to induce the expression of FAP and enhance theproliferation of the cells.
miR-21 induces myofibroblasts differentiation,accelerates tumor growth, and promotes metastasis ofbreast phyllodes tumors xenografts
To investigate the role of miR-21 on tumor progressionin vivo, athymic nude mice were inoculated with stromal cellsfrom benign or malignant phyllodes tumors in their mammaryfat pads. When the xenografts became palpable,miR-21mimicor ASO was injected into the tumor twice a week. Injection ofmiR-21 mimics, but not nonrelevant lin4 mimics, significantlyaccelerated tumor growth of xenografts using stromal cellsfrom benign phyllodes tumors (P < 0.01; Fig. 5A). On the otherhand, injection of miR-21 ASO, but not lin4 ASO, markedlyinhibited tumor growth of xenografts using stromal cells frommalignant phyllodes tumors (P < 0.01; Fig. 5A). In agreementwith the tumor growth, the tumor formation efficiencywas also increased by miR-21 mimics in benign stromal cellsand reduced by miR-21 ASO in malignant stromal cells
Figure 4. Smad 7 and PTEN aretargets of miR-21 in phyllodestumors. A, the protein level ofSmad7 and PTEN in benignphyllodes tumors stromal cellstreated with mock transfection, ortransfected with lin4 mimics (lin4)or miR-21 mimics (miR-21) and inmalignant phyllodes tumors tromalcells treated with lin4 ASOtransfection (NC), or transfectedwithmockormiR-21ASO.BandC,the protein level of a-SMA and FAPin the stromal cells of benignphyllodes tumors with Smad7 (B)and PTEN (C) knockdown by RNAi.D, the protein level of a-SMA andFAP in untreated malignantphyllodes tumors stromal cells,and in benign phyllodes tumorsstromal cells treated with mocktransfection, or miR-21 mimicsalone (miR-21), or cotransfectionwith empty vectors, orcotransfectionwith either or bothofSmad7- and PTEN-expressingvectors with their 30-UTRsmutatedfor miR-21-binding sites. E,representative images of Boydenchamber assay for migrated andinvaded cells; the collagen gelcontraction, and colony formationcells were treated as in D.
Gong et al.
Cancer Res; 74(16) August 15, 2014 Cancer Research4348
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125
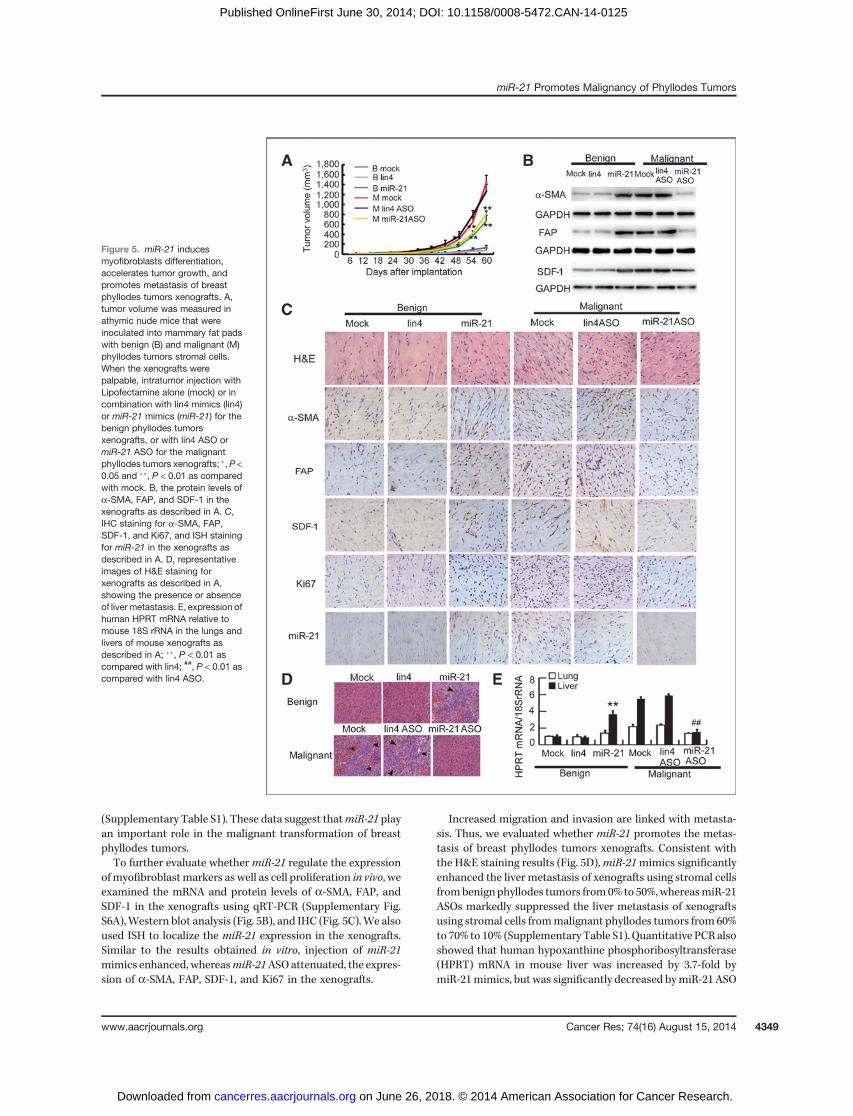
(Supplementary Table S1). These data suggest thatmiR-21 playan important role in the malignant transformation of breastphyllodes tumors.To further evaluate whether miR-21 regulate the expression
ofmyofibroblastmarkers as well as cell proliferation in vivo, weexamined the mRNA and protein levels of a-SMA, FAP, andSDF-1 in the xenografts using qRT-PCR (Supplementary Fig.S6A),Western blot analysis (Fig. 5B), and IHC (Fig. 5C).We alsoused ISH to localize the miR-21 expression in the xenografts.Similar to the results obtained in vitro, injection of miR-21mimics enhanced, whereasmiR-21ASOattenuated, the expres-sion of a-SMA, FAP, SDF-1, and Ki67 in the xenografts.
Increased migration and invasion are linked with metasta-sis. Thus, we evaluated whether miR-21 promotes the metas-tasis of breast phyllodes tumors xenografts. Consistent withthe H&E staining results (Fig. 5D),miR-21mimics significantlyenhanced the liver metastasis of xenografts using stromal cellsfrombenign phyllodes tumors from0% to 50%,whereasmiR-21ASOs markedly suppressed the liver metastasis of xenograftsusing stromal cells frommalignant phyllodes tumors from 60%to 70% to 10% (Supplementary Table S1). Quantitative PCRalsoshowed that human hypoxanthine phosphoribosyltransferase(HPRT) mRNA in mouse liver was increased by 3.7-fold bymiR-21mimics, but was significantly decreased bymiR-21 ASO
Figure 5. miR-21 inducesmyofibroblasts differentiation,accelerates tumor growth, andpromotes metastasis of breastphyllodes tumors xenografts. A,tumor volume was measured inathymic nude mice that wereinoculated into mammary fat padswith benign (B) and malignant (M)phyllodes tumors stromal cells.When the xenografts werepalpable, intratumor injection withLipofectamine alone (mock) or incombination with lin4 mimics (lin4)or miR-21 mimics (miR-21) for thebenign phyllodes tumorsxenografts, or with lin4 ASO ormiR-21 ASO for the malignantphyllodes tumors xenografts; �,P <0.05 and ��, P < 0.01 as comparedwith mock. B, the protein levels ofa-SMA, FAP, and SDF-1 in thexenografts as described in A. C,IHC staining for a-SMA, FAP,SDF-1, and Ki67, and ISH stainingfor miR-21 in the xenografts asdescribed in A. D, representativeimages of H&E staining forxenografts as described in A,showing the presence or absenceof liver metastasis. E, expression ofhuman HPRT mRNA relative tomouse 18S rRNA in the lungs andlivers of mouse xenografts asdescribed in A; ��, P < 0.01 ascompared with lin4; ##, P < 0.01 ascompared with lin4 ASO.
miR-21 Promotes Malignancy of Phyllodes Tumors
www.aacrjournals.org Cancer Res; 74(16) August 15, 2014 4349
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125

(Fig. 5E). However, there was no significant lung metastasisobserved in the mice according to histologic examination,human HPRT mRNA level (Fig. 5E), and wet weight of lung(Supplementary Fig. S6B). Together, these results indicate thatmiR-21 promotes myofibroblast differentiation, tumor forma-tion as well as liver metastasis in breast phyllodes tumorxenografts.
DiscussionIn this study, we have found that myofibroblast differenti-
ation, driven by the upregulation of miR-21, is progressivelyincreased during the malignant progression of phyllodestumors in breast. Furthermore, a-SMA and miR-21 can serveas independent prognostic markers of phyllodes tumors, withtheir predictive value better than histologic classification.
It has been well established that the prognostic value ofhistologicmarkers in phyllodes tumors is not as good as that incancer. Even benign phyllodes tumors can have malignantpotential and a significant portion of them can recur locally. Itwas reported that the local recurrence rates of patients withbenign, borderline, and malignant phyllodes tumors aftersurgery were 21% (111 of 540), 46% (18 of 39), and 65% (26of 40), respectively (3). In our study, the local recurrence ratesare 11% (18 of 167), 25% (9 of 36), and 34% (22 of 65) for thepatients with benign, borderline, and malignant phyllodestumors correspondingly. Importantly, the local recurrencerates for patients with low a-SMA or low miR-21 expressionare 2.6% (4 of 156) or 2.6% (4 of 152), whereas the ones forpatients with high a-SMA or high miR-21 expression are 40%(45 of 112) or 39% (45 of 116). Furthermore, the ROC curveanalysis showed that both miR-21 (AUC, 0.92) and a-SMA(AUC, 0.89) performed better in predicting the recurrence thanhistologic grade (AUC, 0.67), suggesting that miR-21 anda-SMA could serve as novel molecular markers to predict therecurrence of phyllodes tumors.
Previous studies reported that stromal myofibroblasts areimportant promoters of tumor growth and progression inmultiple cancer types, but myofibroblast is hardly reportedto be a direct tumor-initiating component of tumors. Fibro-blast is a major component of phyllodes tumors and therecurring or metastasizing behavior of phyllodes tumors isdetermined by the properties of fibroblasts. Here, we show, forthe first time, that myofibroblast is the major malignantcomponent of phyllodes tumors. FMT, driven by upregulatedmiR-21, underlies the malignant transformation of phyllodestumors. Inhibition of miR-21 reversed the FMT and decreasedthe malignancy of phyllodes tumors, suggesting that miR-21could be a therapeutic target in phyllodes tumors.
miR-21 has been reported to be an important oncomir inmany types of cancer (32–34).Most of the studies so far focusedon the increased expression ofmiR-21 in cancer cells or tumormixture. However, it was reported thatmiR-21 expression waspredominantly seen in cancer-associated fibroblast-like cellsin breast cancer, with no difference in expression levelsbetween low- and high-grade cancers (35). It was also shownthat miR-21 expression in esophageal squamous-cell carcino-ma was mainly localized in the cytoplasm of stromal cells
adjacent to malignant cells (36) Thus, it is possible thatmyofibroblast transition, driven by highmiR-21 in the stromalcells, could also be an important tumor-promoting event inmany epithelial cancers. On the other hand, breast cancer cellscan directly induce the expression of myofibroblast markers infibroblasts (37), suggesting that both primary tumor andmyofibroblasts can work together to promote tumorigenesis.Highly expressed miR-21 could be shuttled between cancercells and fibroblasts in the form of secreted exosomes to playtheir function in respective cells.
Besides its tumor-promoting role, miR-21 is also impli-cated in drug resistance (38) as well as radiation resistance(39) of cancer cells. It is possible that high miR-21 inphyllodes tumors is not only important for the malignantprogression of phyllodes tumors, but also may be responsi-ble for its poor response to chemotherapy and radiotherapy.Antagonizing miR-21 in phyllodes tumors may resensitizetumors to chemotherapy or radiotherapy, in addition to thedecreased malignancy.
PTEN is a well-established target of miR-21. Our studydemonstrated that miR-21 inhibited PTEN expression, andthus induced FAP expression. FAP, a member of the serineprotease family, has been shown to support tumor growth andproliferation (40). A recent study also showed that miR-21induced lung fibrosis by targeting Smad7 and mediatingpulmonary fibroblasts differentiation into myofibroblasts(30). Our results demonstrated that miR-21 induced FMTand a-SMA expression via the downregulation of Smad7.Together, our data indicate that miR-21 downregulatesPTEN and Smad-7, which then upregulates the expression ofFAP and a-SMA that are responsible for the enhanced prolife-ration and invasiveness of phyllodes tumors respectively.
In summary, our study suggests an important role ofFMT in the malignant transformation of phyllodes tumorsthat are driven by increased miR-21. Inhibition of miR-21may be a novel approach in treating phyllodes tumors.a-SMA andmiR-21 are independent predictors of recurrencefor phyllodes tumors.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: C. Gong, Y. Nie, E. Song, Q. LiuDevelopment of methodology: Y. Nie, S. QuAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): C. Gong, Y. Nie, X. Cui, H. YaoAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): C. Gong, Y. Nie, S. Qu, J.-Y. Liao, H. Yao, F. Su, Q. LiuWriting, review, and/or revision of the manuscript: C. Gong, Y. Nie,E. Song, Q. LiuAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): Y. Zeng, Q. LiuStudy supervision: E. Song, Q. Liu
Grant SupportThis work was supported by grants from 973 (2010CB912800 and
2011CB504203) Projects from Ministry of Science and Technology of China; theNatural Science Foundation of China (81230060, 81261140373, 81272893,81172524, 81372817, 81272894, and 81072178); Science Foundation ofGuangdong Province (S2012030006287 and 2012J2200092); Special Project onNew Drug Innovation of Sun Yat-sen University; Sun Yat-sen University TrainingProject (11ykpy28, 12ykpy32, and 13ykzd14); Elite Yong Scholars Program of Sun
Gong et al.
Cancer Res; 74(16) August 15, 2014 Cancer Research4350
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125

Yat-sen Memorial Hospital (Y201401); Translational medicine public platformof Guangdong Province (4202037); Grant KLB09001 from the Key Laboratory ofMalignant Tumor Gene Regulation and Target Therapy of Guangdong HigherEducation Institutes, SunYat-Sen University and Grant [2013]163 from KeyLaboratory of Malignant Tumor Molecular Mechanism and Translational Med-icine of Guangzhou Bureau of Science and Information Technology.
The costs of publication of this article were defrayed in part by the paymentof page charges. This article must therefore be hereby marked advertisementin accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received January 18, 2014; revised April 22, 2014; accepted May 21, 2014;published OnlineFirst June 30, 2014.
References1. Pimiento JM, Gadgil PV, Santillan AA, Lee MC, Esposito NN, Kiluk JV,
et al. Phyllodes tumors: race-related differences. J Am Coll Surg2011;213:537–42.
2. Chaney AW, Pollack A, McNeeseMD, Zagars GK, Pisters PW, PollockRE, et al. Primary treatment of cystosarcoma phyllodes of the breast.Cancer 2000;89:1502–11.
3. Barth RJ, Jr. Histologic features predict local recurrence after breastconserving therapy of phyllodes tumors. Breast Cancer Res Treat1999;57:291–5.
4. ShpitzB,BomsteinY,SternbergA,KleinE, TiomkinV,KaufmanA, et al.Immunoreactivity of p53, Ki-67, and c-erbB-2 in phyllodes tumors ofthe breast in correlation with clinical and morphologic features. J SurgOncol 2002;79:86–92.
5. Jones AM, Mitter R, Springall R, Graham T, Winter E, Gillett C, et al.A comprehensive genetic profile of phyllodes tumours of the breastdetects important mutations, intra-tumoral genetic heterogeneityand new genetic changes on recurrence. J Pathol 2008;214:533–44.
6. Azzopardi JG, Ahmed A, Millis RR. Problems in breast pathology.Major Probl Pathol 1979;11:i-xvi, 1–466.
7. Khan SA, Badve S. Phyllodes tumors of the breast. Curr Treat OptionsOncol 2001;2:139–47.
8. Chang HY, Chi JT, Dudoit S, Bondre C, van de RijnM, Botstein D, et al.Diversity, topographic differentiation, and positionalmemory in humanfibroblasts. Proc Natl Acad Sci U S A 2002;99:12877–82.
9. Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer 2006;6:392–401.
10. Ronnov-Jessen L, PetersenOW,Koteliansky VE,BissellMJ. The originof the myofibroblasts in breast cancer. Recapitulation of tumor envi-ronment in culture unravels diversity and implicates converted fibro-blasts and recruited smooth muscle cells. J Clin Invest 1995;95:859–73.
11. Lieubeau B, Garrigue L, Barbieux I, Meflah K, Gregoire M. The role oftransforming growth factor beta 1 in the fibroblastic reaction associ-ated with rat colorectal tumor development. Cancer Res 1994;54:6526–32.
12. Tsukamoto H, Mishima Y, Hayashibe K, Sasase A. Alpha-smoothmuscle actin expression in tumor and stromal cells of benign andmalignant human pigment cell tumors. J Invest Dermatol 1992;98:116–20.
13. De Wever O, Mareel M. Role of tissue stroma in cancer cell invasion.J Pathol 2003;200:429–47.
14. Aranda FI, Laforga JB, Lopez JI. Phyllodes tumor of the breast.An immunohistochemical study of 28 cases with special atten-tion to the role of myofibroblasts. Pathol Res Pract 1994;190:474–81.
15. Wienholds E, Koudijs MJ, van Eeden FJ, Cuppen E, Plasterk RH. ThemicroRNA-producing enzyme Dicer1 is essential for zebrafish devel-opment. Nat Genet 2003;35:217–8.
16. Yi R,O'Carroll D, Pasolli HA, ZhangZ,Dietrich FS, TarakhovskyA, et al.Morphogenesis in skin is governed by discrete sets of differentiallyexpressed microRNAs. Nat Genet 2006;38:356–62.
17. Yu F, Yao H, Zhu P, Zhang X, Pan Q, Gong C, et al. let-7 regulates selfrenewal and tumorigenicity of breast cancer cells. Cell 2007;131:1109–23.
18. Tang W, Yu F, Yao H, Cui X, Jiao Y, Lin L, et al. miR-27a regulatesendothelial differentiation of breast cancer stem like cells. Oncogene2014;33:2629–38.
19. Nie Y, Liu X, Qu S, Song E, Zou H, Gong C. Long non-codingRNA HOTAIR is an independent prognostic marker for nasopha-
ryngeal carcinoma progression and survival. Cancer Sci 2013;104:458–64.
20. Esposito NN, Mohan D, Brufsky A, Lin Y, Kapali M, Dabbs DJ.Phyllodes tumor: a clinicopathologic and immunohisto-chemical study of 30 cases. Arch Pathol Lab Med 2006;130:1516–21.
21. Noronha Y, Raza A, Hutchins B, Chase D, Garberoglio C, Chu P, et al.CD34, CD117, and Ki-67 expression in phyllodes tumor of the breast:an immunohistochemical study of 33 cases. Int J Surg Pathol 2010;19:152–8.
22. Kucuk U, Bayol U, Pala EE, Cumurcu S. Importance of P53, Ki-67expression in the differential diagnosis of benign/malignantphyllodes tumors of the breast. Indian J Pathol Microbiol 2013;56:129–34.
23. ZweigMH,CampbellG.Receiver-operatingcharacteristic (ROC)plots:a fundamental evaluation tool in clinical medicine. Clin Chem 1993;39:561–77.
24. Engel JM, Junger A, Hartmann B, Little S, Schnobel R, Mann V, et al.Performance and customization of 4 prognostic models for postop-erative onset of nausea and vomiting in ear, nose, and throat surgery.J Clin Anesth 2006;18:256–63.
25. Wu Q, Yang Z, Wang F, Hu S, Yang L, Shi Y, et al. MiR-19b/20a/92aregulates the self-renewal and proliferation of gastric cancer stemcells. J Cell Sci 2013;126:4220–9.
26. Ohyagi-Hara C, Sawada K, Kamiura S, Tomita Y, Isobe A, HashimotoK, et al. miR-92a inhibits peritoneal dissemination of ovarian cancercells by inhibiting integrin alpha5 expression. Am J Pathol 2013;182:1876–89.
27. Gong X, Zhang K, Wang Y, Wang J, Cui Y, Li S, et al. MicroRNA-130btargets Fmr1 and regulates embryonic neural progenitor cell prolifer-ation and differentiation. Biochem Biophys Res Commun 2013;439:493–500.
28. MaS, TangKH,ChanYP, LeeTK, KwanPS,CastilhoA, et al.miR-130bPromotes CD133(þ) liver tumor-initiating cell growth and self-renewalvia tumor protein 53-induced nuclear protein 1. Cell Stem Cell2010;7:694–707.
29. Mishra S, Deng JJ, Gowda PS, Rao MK, Lin CL, Chen CL, et al.Androgen receptor and microRNA-21 axis downregulates transform-ing growth factor beta receptor II (TGFBR2) expression in prostatecancer. Oncogene. 2013 Sep 16. [Epub ahead of print].
30. LiuG, Friggeri A, YangY,Milosevic J, DingQ, Thannickal VJ, et al.miR-21 mediates fibrogenic activation of pulmonary fibroblasts and lungfibrosis. J Exp Med 2010;207:1589–97.
31. WeinsteinD, StranoS,CohenP, FieldsS,Gomori JM,Degani H.Breastfibroadenoma: mapping of pathophysiologic features with three-time-point, contrast-enhanced MR imaging—pilot study. Radiology 1999;210:233–40.
32. Toiyama Y, Takahashi M, Hur K, Nagasaka T, Tanaka K, Inoue Y, et al.Serum miR-21 as a diagnostic and prognostic biomarker in colorectalcancer. J Natl Cancer Inst 2013;105:849–59.
33. Hede K. MicroRNAs as onco-miRs, drivers of cancer. J Natl CancerInst 2010;102:1306–8.
34. Tahiri A, Leivonen SK, Luders T, Steinfeld I, Ragle Aure M, Geisler J,et al. Deregulation of cancer-related miRNAs is a common event inboth benign and malignant human breast tumors. Carcinogenesis2014;35:76–85.
35. Rask L, Balslev E, Jorgensen S, Eriksen J, Flyger H, Moller S, et al.High expression of miR-21 in tumor stroma correlates withincreased cancer cell proliferation in human breast cancer. APMIS2011;119:663–73.
miR-21 Promotes Malignancy of Phyllodes Tumors
www.aacrjournals.org Cancer Res; 74(16) August 15, 2014 4351
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125

36. Nouraee N, Van Roosbroeck K, Vasei M, Semnani S, Samaei NM,Naghshvar F, et al. Expression, tissue distribution and function ofmiR-21 in esophageal squamous cell carcinoma. PLoS ONE 2013;8:e73009.
37. Martinez-Outschoorn UE, Pavlides S, Whitaker-Menezes D, DaumerKM, Milliman JN, Chiavarina B, et al. Tumor cells induce the cancerassociated fibroblast phenotype via caveolin-1 degradation: implica-tions for breast cancer and DCIS therapy with autophagy inhibitors.Cell Cycle 2010;9:2423–33.
38. Hong L,HanY, ZhangY, ZhangH, ZhaoQ,WuK, et al.MicroRNA-21: atherapeutic target for reversing drug resistance in cancer. Expert OpinTher Targets 2013;17:1073–80.
39. Anastasov N, Hofig I, Vasconcellos IG, Rappl K, Braselmann H,Ludyga N, et al. Radiation resistance due to high expression of miR-21 and G2/M checkpoint arrest in breast cancer cells. Radiat Oncol2012;7:206.
40. Liu R, Li H, Liu L, Yu J, Ren X. Fibroblast activation protein: a potentialtherapeutic target in cancer. Cancer Biol Ther 2012;13:123–9.
Cancer Res; 74(16) August 15, 2014 Cancer Research4352
Gong et al.
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125

2014;74:4341-4352. Published OnlineFirst June 30, 2014.Cancer Res Chang Gong, Yan Nie, Shaohua Qu, et al. Malignant Progression of Breast Phyllodes Tumors
Induces Myofibroblast Differentiation and Promotes themiR-21
Updated version
10.1158/0008-5472.CAN-14-0125doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2014/07/21/0008-5472.CAN-14-0125.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/74/16/4341.full#ref-list-1
This article cites 38 articles, 5 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/74/16/4341.full#related-urls
This article has been cited by 2 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/74/16/4341To request permission to re-use all or part of this article, use this link
on June 26, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst June 30, 2014; DOI: 10.1158/0008-5472.CAN-14-0125




























