Embed Size (px)
Citation preview

Module d’ouverture :
« Biotechnologies en 10 leçons »

Les Enzymes
Professeur D. CombesINSA, Toulouse

Les enzymes
Les enzymes
Production des enzymes
Purification des enzymes
Immobilisation des enzymes

1783 Spallanzani, viande "liquéfiée" par le sucgastrique des faucons.1833 première découverte d'une enzyme : Payen etPersoz.1858 à 1871, Pasteur observe les changementschimiques qui se produisent lors de lafermentation.1878 : Kühne propose le mot ENZYME qui signifie"dans la levure".1897, Bertrand : coenzymes.
Historique

fin des années 20 : les enzymes sont des protéinesannées 40 et 50, des centaines de nouvelles enzymes sont découvertes, purifiées, cristallisées : voies métaboliques clefs élucidées, mécanismes d'action et de régulation des enzymes étudiés. 1960 : 1ère enzyme (la Ribonucléase) séquencée1969 : Ribonucléase synthétisée par voie chimique.de 1969 à nos jours : Génie Génétique
Historique

1 - Oxydoréductases : réactions d'oxydo-réduction.
2 - Transférases :transfert d'un groupe d'une molécule àune autre.
3 - Hydrolases : coupure hydrolytique des liaisons C-O, C-N, C-C
4 - Lyases : coupure des liaisons C-C, C-O et C-N par élimination, en formant des = liaisons ou des cycles
5 - Isomérases : changements structuraux dans une molécule
6 - Ligases : liaison entre deux molécules, couplée àl'hydrolyse d'une liaison Phosphate de l'ATP
Classification

Transférasestransfert d'un groupe spécifique d'une molécule à une autre
Dextrane Saccharase = E.C. 2.4.1.5Sacch + Dexn (1,6 Glu) D-fruct. + (Dex)n+1
Lévane Saccharase = E.C. 2.4.1.10Sacch + Levn (2,6 Fruct) D-glu + (Lev)n+1
Saccharose Phosphorylase = E.C. 2.4.1.7Sacch + P D-fructose + G-1P
Hexokinase = E.C. 2.7.1.1ATP + Hexose ADP + Hexose 6-P
Exemples

IsomérasesChangements géométriques ou structuraux dans unemolécule :glucose-isomérase (E.C. 5.3.1.5) Xylose Isomérase
glucose fructose
Exemples
Ligases
liaison entre deux molécules, couplée à l'hydrolyse d'une liaison Phosphate de l'ATP :
E.C. 6.4.1.1 Pyruvate carboxylase
ATP+Pyruvate+HCO3- ADP+ PO4-+Oxaloacétate

Enzymes / Catalyseurs
Site actif
Liaison tridimensionnelle
Flexibilité du site actif
Efficacité catalytique
Notion de Catalyse
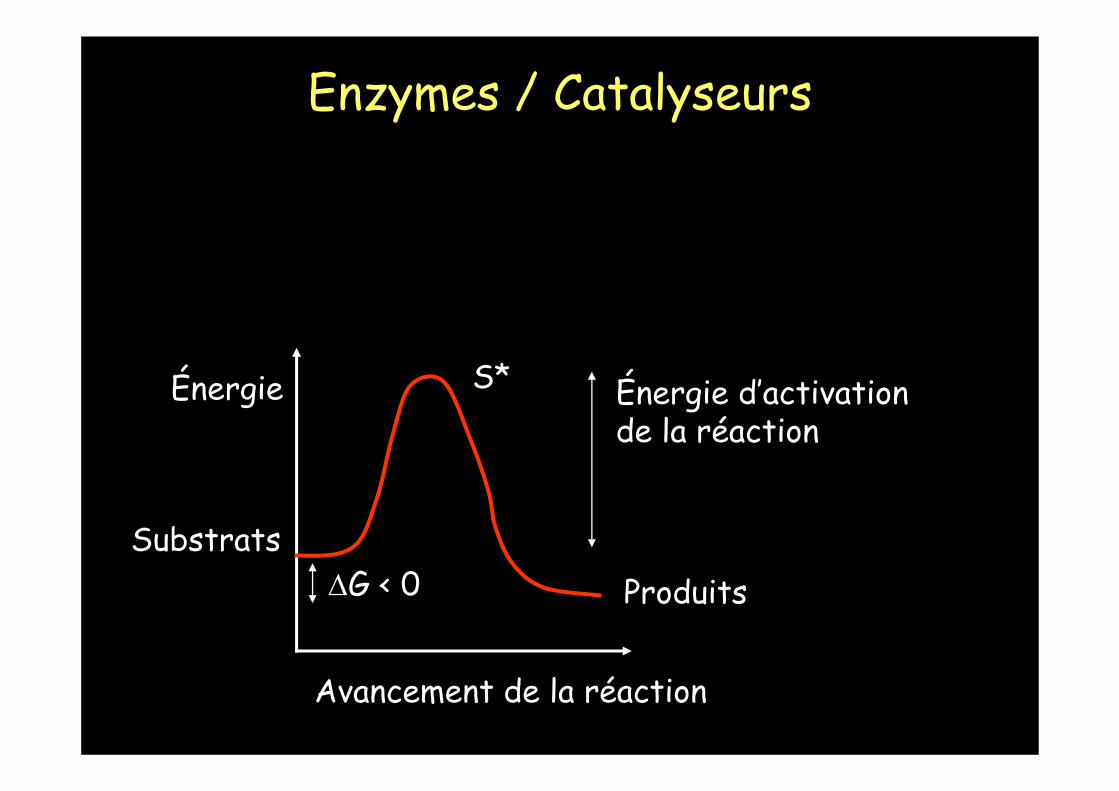
Énergie
Avancement de la réaction
Énergie d’activationde la réaction
ΔG < 0Substrats
Produits
S*
Enzymes / Catalyseurs

Il existe un état de transition S*
Comment accélérer la réaction detransformation de S en P ?
-élever la température
- diminuer l'énergie d'activation
Enzymes / Catalyseurs
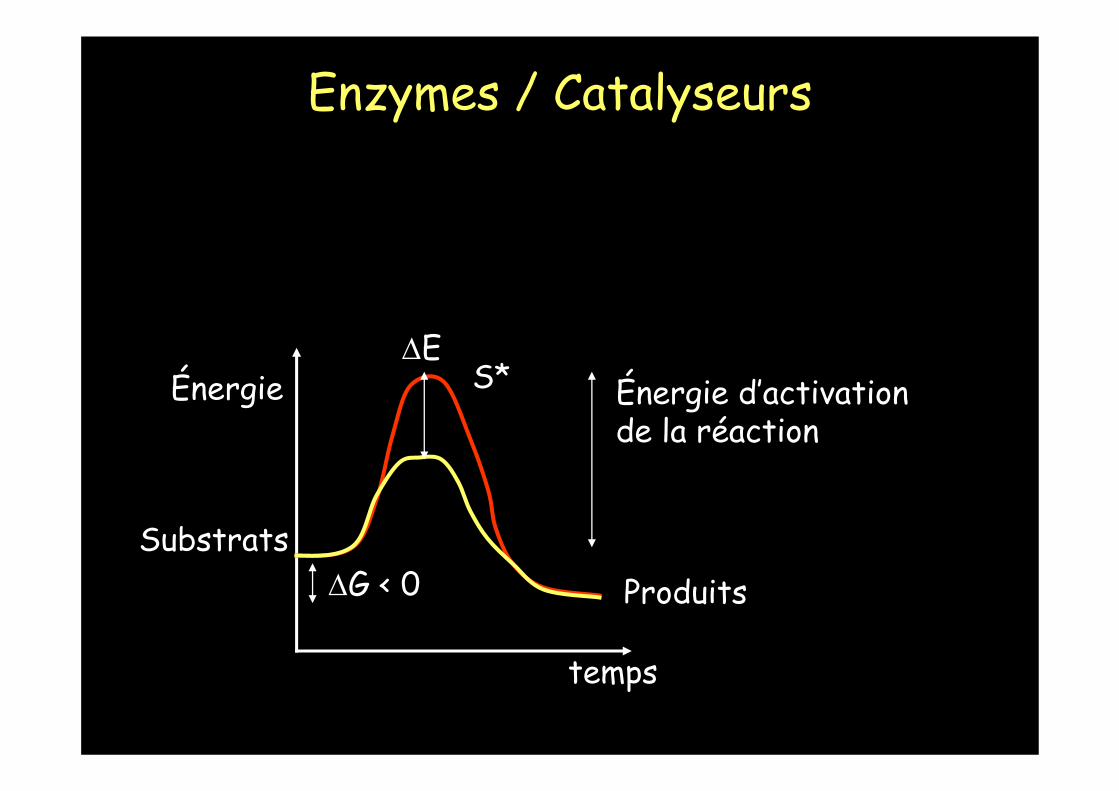
Énergie
temps
Énergie d’activationde la réaction
ΔG < 0
ΔE
SubstratsProduits
S*
Enzymes / Catalyseurs

Les enzymes diminuent sélectivement l'énergie d'activation d'une réaction donnée.
La température ambiante est alors suffisante pour qu'un nombre important de S P.
Les enzymes sont des catalyseurs qui augmentent la vitesse des réactions chimiques sans être consommées
Enzymes / Catalyseurs

- région complémentaire en taille, forme et nature chimique du substrat
- il y a seulement une douzaine d'acides aminés autour du site actif avec seulement 2 ou 3 qui sont directement impliqués.
Site actif

A B
C
enzyme
Site actif
Site actif

Les enzymes accélèrent les vitesses de réaction en diminuant l'énergie d'activation :
- les réactions enzymatiques se déroulent suivant un mécanisme de réactions organiques (acide-base, nucléophile, électrophile) pour lesquelles l'enzyme fournit les groupes catalytiques
- les facteurs de proximité et d'orientation sont prépondérants. Sans enzyme, les rencontres de deux molécules se font au hasard : ce n'est plus le cas ici
- certaines liaisons du substrat sont tendues par l'enzyme
Efficacité catalytique des enzymes

Observations :
– la vitesse initiale de réaction est directement proportionnelle à la concentration de la préparation enzymatique
– cette vitesse augmente de manière non linéaire avec la concentration du substrat
– elle atteint une vitesse maximale
Cinétique d’enzymes à un substrat : Michaelis-Menten

Hypothèses– 1- L'enzyme est un catalyseur– 2 - L'enzyme et le substrat réagissent
rapidement pour former un complexe enzyme-substrat
– 3 - Un seul substrat et un seul complexe enzyme-substrat sont impliqués et le complexe enzyme-substrat se brise pour donner directement l'enzyme libre et le produit
– 4 - L'enzyme, le substrat et le complexe enzyme-substrat sont en équilibre. De plus la vitesse de dissociation de ES en E + S est beaucoup plus rapide que la vitesse de coupure de ES pour former E + P
Michaelis-Menten

- 5 - La concentration du substrat est beaucoup plus élevée que celle de l'enzyme ; ainsi la formation du complexe ES n'altère pas la valeur de la concentration de S
– 6 - La vitesse globale de la réaction est limitée par la coupure du complexe ES pour former l'enzyme libre et le produit
– 7 - La vitesse est mesurée dans les premiers instants de la réaction de telle manière que la réaction inverse n'est pas significative.
Michaelis-Menten
EQUATION DE MICHAELIS-MENTEN

VM [S]V = KM + [S]
Michaelis-Menten : équation

Vi
[S]
VM
VM/2
KM
Michaelis-Menten : traitement graphique

•Novozyme (Danemark) : 50%
•DSM (Hollande) : 20%
•Genencor International (USA) : 15%
•Solvay-Miles (USA) : 5%
• Amano, Nagase (Japon)
Les producteurs d’enzymes

millions de $
1000
500
250
750
01970 1980 19901966 2000
1250
Evolution des ventesmondiales d’ enzymes

1970 1980 19901966
Réflexe négatif des consommateurs :problème d'allergiecausé par les poudres.
Les enzymes sont incluses dans des granules en fibre de cellulose
Le marché des enzymes dans les détergents

Détergents32 %
Amidon
15 %Textile
10 %
Produits laitiers
14 %
CosmétiqueAlcoolBière jus de fruitsnourriture animalediagnosticschimieboulangeriearômesgraissespapiercuir
Autres 29 %
Répartition des ventes d’enzymes en 1995
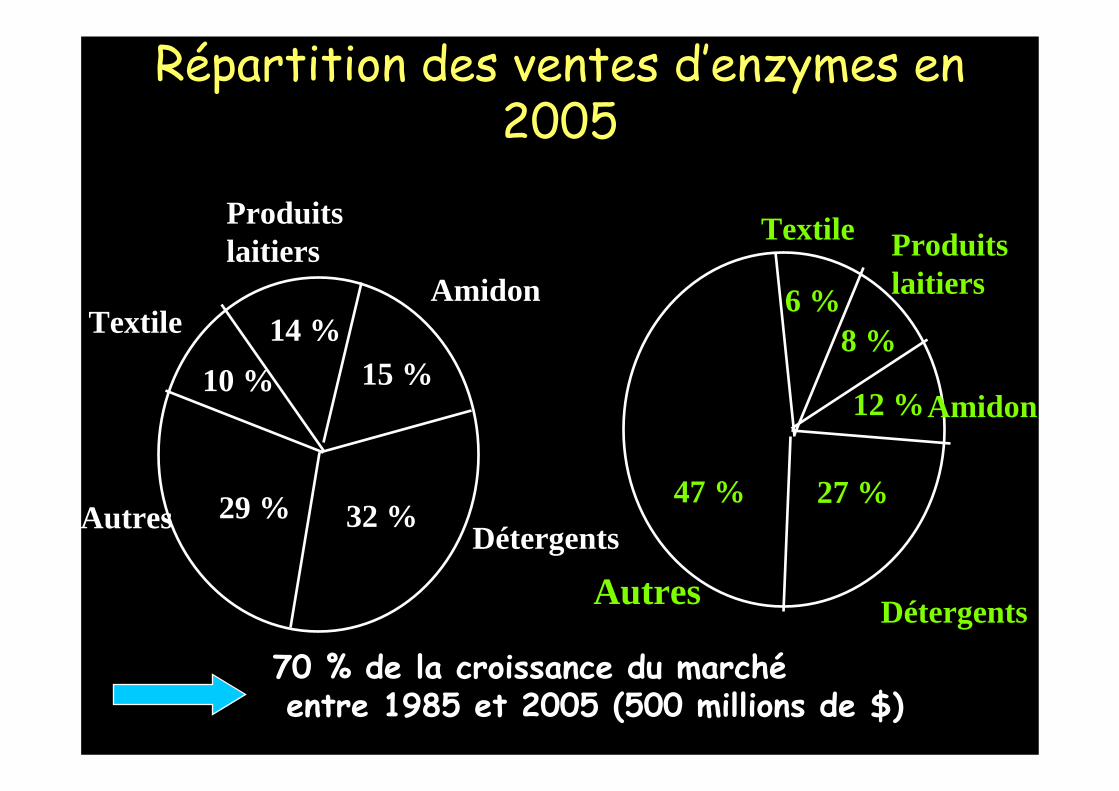
Détergents32 %
Amidon
15 %Textile
10 %
Produits laitiers
14 %
Autres 29 %
Détergents
27 %
Amidon12 %
Textile
6 %
Produits laitiers
8 %
Autres
47 %
70 % de la croissance du marchéentre 1985 et 2005 (500 millions de $)
Répartition des ventes d’enzymes en 2005

• 75 % sont des enzymes catalysant des hydrolyses,utilisées pour dépolymériser des substances naturelles.
• 40 % : Protéase (produits laitiers, détergents).• Carbohydrolase (industries de l'amidon et dutextile, boulangerie, alcools).
•Plusieurs milliers d'enzymes ont été identifiées• en 1983 : 30 enzymes produites à grande échelle.• en 1995 : 60 enzymes produites à grande échelle.
Types d’enzymes
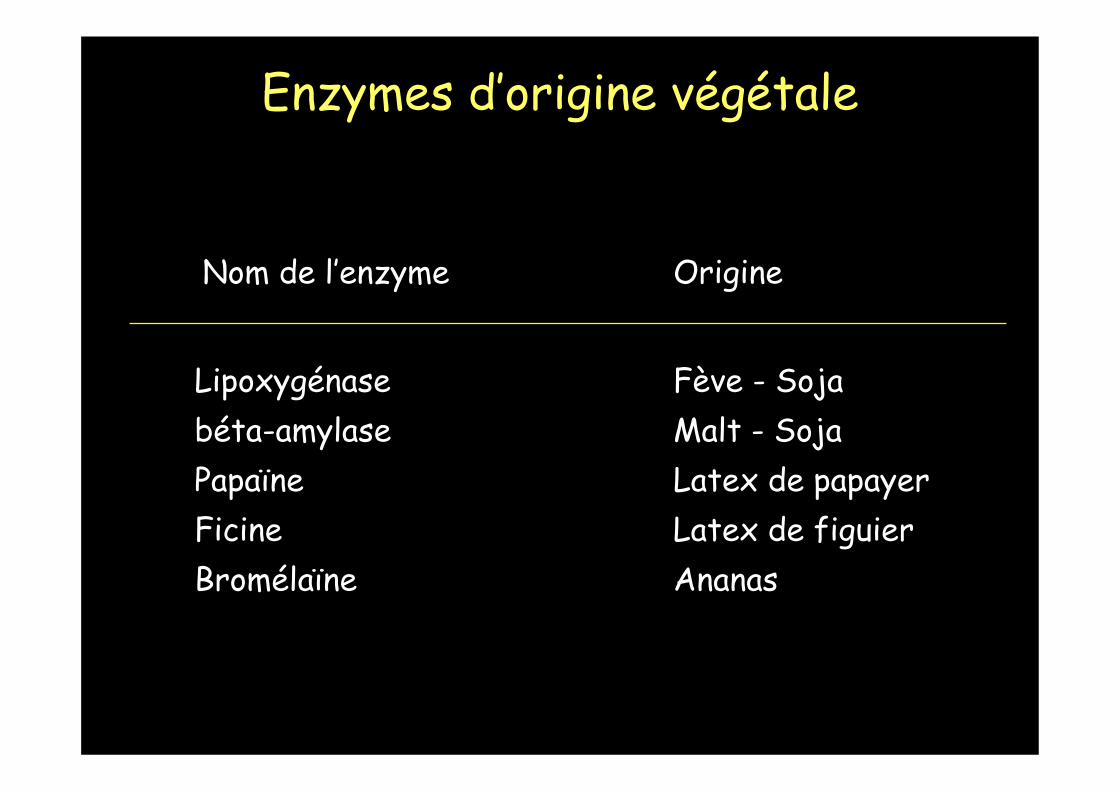
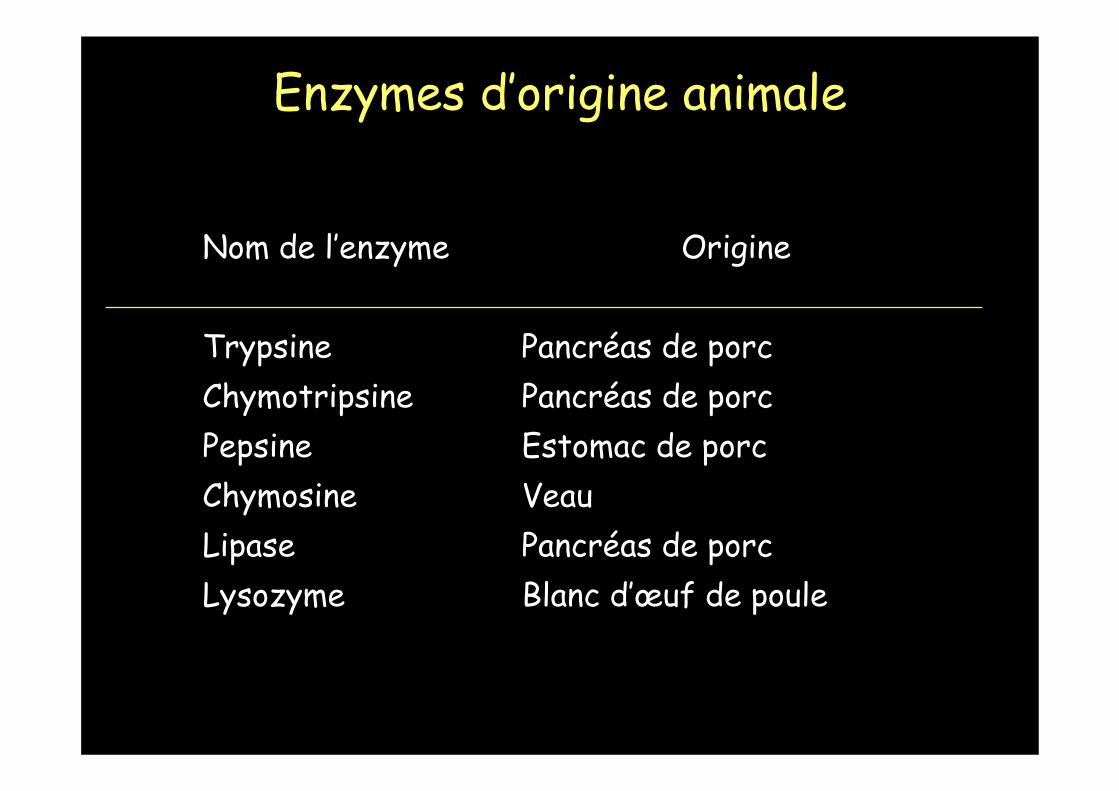
Nom de l’enzyme Origine
Lipoxygénase Fève - Sojabéta-amylase Malt - SojaPapaïne Latex de papayerFicine Latex de figuierBromélaïne Ananas
Enzymes d’origine végétale
Nom de l’enzyme Origine
Trypsine Pancréas de porcChymotripsine Pancréas de porcPepsine Estomac de porcChymosine VeauLipase Pancréas de porcLysozyme Blanc d’œuf de poule
Enzymes d’origine animale

1991 1993 1995Native 77 15 8
ADN recombinant 16 71 67
Ingénierie des protéines 7 14 25
Les sources d’enzymes

•indépendance des contraintes saisonnières et géographiques•utilisation de matières premières bon marché•utilisation d’installations industrielles polyvalentes•rendements de production pouvant faire l’objet d’amélioration.•lourdeur des investissements•coût énergétique•nécessité d’un personnel qualifié•risques de contamination, pollution
Production par fermentation

Endo Glucose oxydaseCatalase
Exo alpha-amylaseAmyloglucosidaseMaltaseCellulasePectinaseBéta-glucanaseInulinaseLipaseLactaseProtéase acide
Enzymes produites par A. niger

•Culture dans un milieu induisant la production efficace del'enzyme.•Retrait de la biomasse, extraction et purification del'enzyme, addition de conservateurs, immobilisation.•Isolement d'un micro-organisme naturel sécrétant uneenzyme d'intérêt.•Le plus souvent la fermentation se fait dans une cuveagitée (jusqu’à 200 m3).•Energie d’agitation élevée (5 à 10 CV /m3).• Système de refroidissement important.•Contrôle de la T°, du pH, de la pO2, des mousses.
Technologie de base

• Génie génétique : transfert de gènes entre micro-organismes.Clonage du gène d'une enzyme provenant d'un organismerare chez un organisme permettant une productionindustrielle efficace.
•Ingénierie des protéines : production d'enzymes surmesure.
•Purification : production sous forme de cristaux purs.
Apport des Biotechnologies modernes

Obtenir en quantité suffisante une protéine « pure »tout en préservant ses propriétés physiologiques
(structure et activité).
Purification des enzymes
Source Extraction Purification
•Dosage de la quantité produite
•Contrôle de la pureté
•Contrôle de l’activité
• Stockage

1 - Obtention de la source de la protéine : biopsie d’organes, collecte de plantes, cultures de cellules, ...
2 - Lyse des cellules : lyse mécanique, lyse chimique ou biochimique.
3 - Séparation des composants insolubles : centrifugation ou filtration.
Extraction
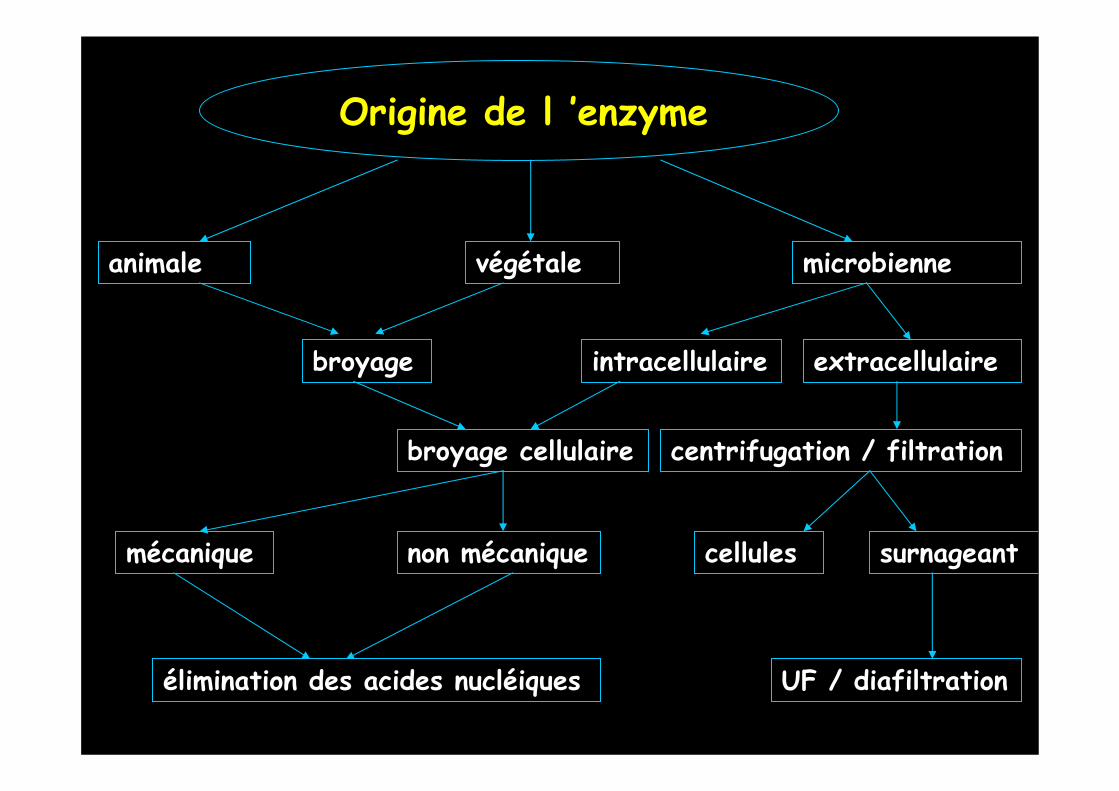
Origine de l ’enzyme
animale végétale microbienne
broyage intracellulaire extracellulaire
centrifugation / filtrationbroyage cellulaire
UF / diafiltration
cellules surnageantnon mécaniquemécanique
élimination des acides nucléiques

Techniques basées sur les propriétés de solubilité
Dénaturation thermique
Précipitation par ajout de sels
Ajout de solvant
Procédés membranaires
Les méthodes de chromatographie
Techniques de purification

Procédé Membrane Force motrice
Filtration poreuse gradient de pression
hydrostatique
Ultrafiltration poreuse gradient de pression
hydrostatique
Osmose inverse non poreuse gradient de pression
hydrostatique
Dialyse poreuse gradient de potentiel
chimique
Electrodialyse poreuse différence de potentiel
électrostatique
Les procédés membranaires

Propriétés des protéines
Hydrophobicité ActivitéCharge
échange d’ionshydrophobe
Taille
exclusion affinité
Méthodes de chromatographie
Principales méthodes de purification des protéines par chromatographie

1 - Le gène de la protéine n’est pas cloné : seules les méthodes classiques de purification peuvent être utilisées.
2 - Le gène de la protéine est cloné : les méthodes classiques peuvent être utilisées mais aussi le gène de la protéine peut être modifié de telle sorte que la protéine soit exprimée sous forme de protéine de fusion.
Protéine naturelle et protéine de fusion
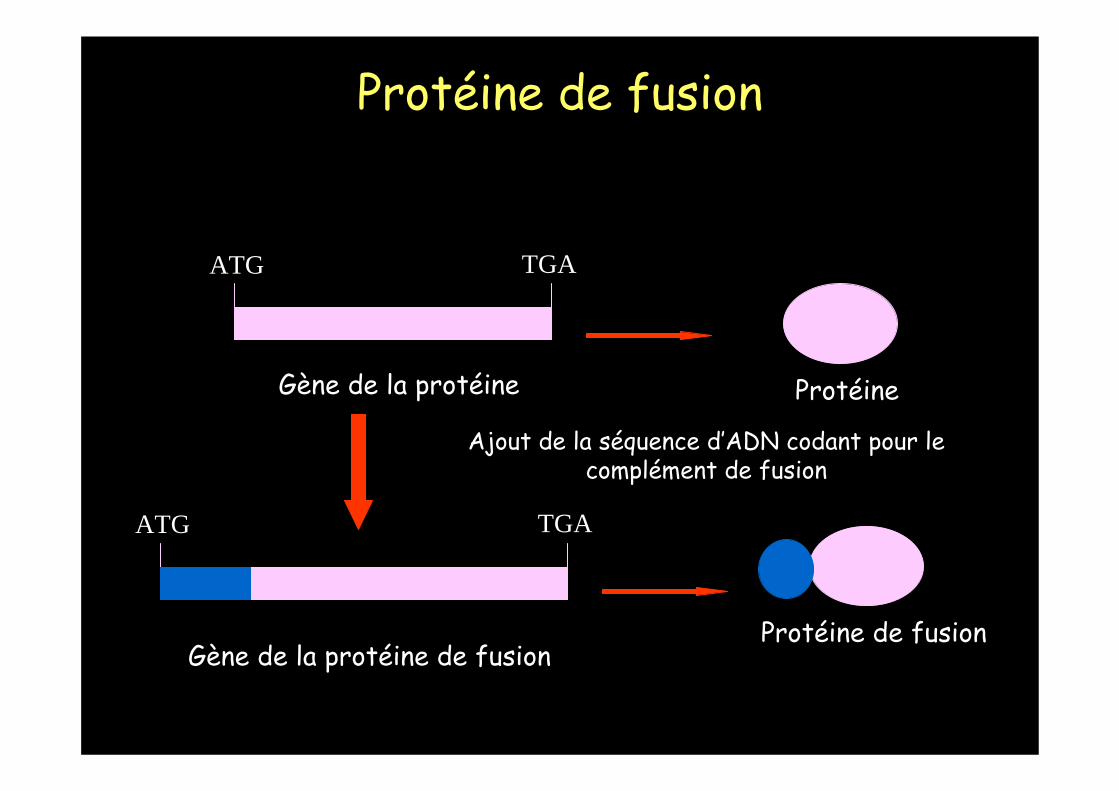
Protéine
ATG TGA
Gène de la protéine
Protéine de fusion
TGAATG
Gène de la protéine de fusion
Ajout de la séquence d’ADN codant pour lecomplément de fusion
Protéine de fusion

•Dialyse, diafiltration
•Concentration
•Détermination de la quantité de protéine
•Évaluation de la pureté
•Mesure de l’activité
•Stockage : conserver une protéine tout en préservant son activité (sous forme lyophilisée,conservation à 4°C, à -20°C, à -70°C, ajout dans le tampon de stockage de composés qui stabilisent la protéine (polyols, sels, agents réducteurs, ...)).
Fin de la purification

Introduction
Méthodes d’immobilisation– inclusion– adsorption– liaison covalente– Réticulation
Conséquences de l’immobilisation
Immobilisation des enzymes
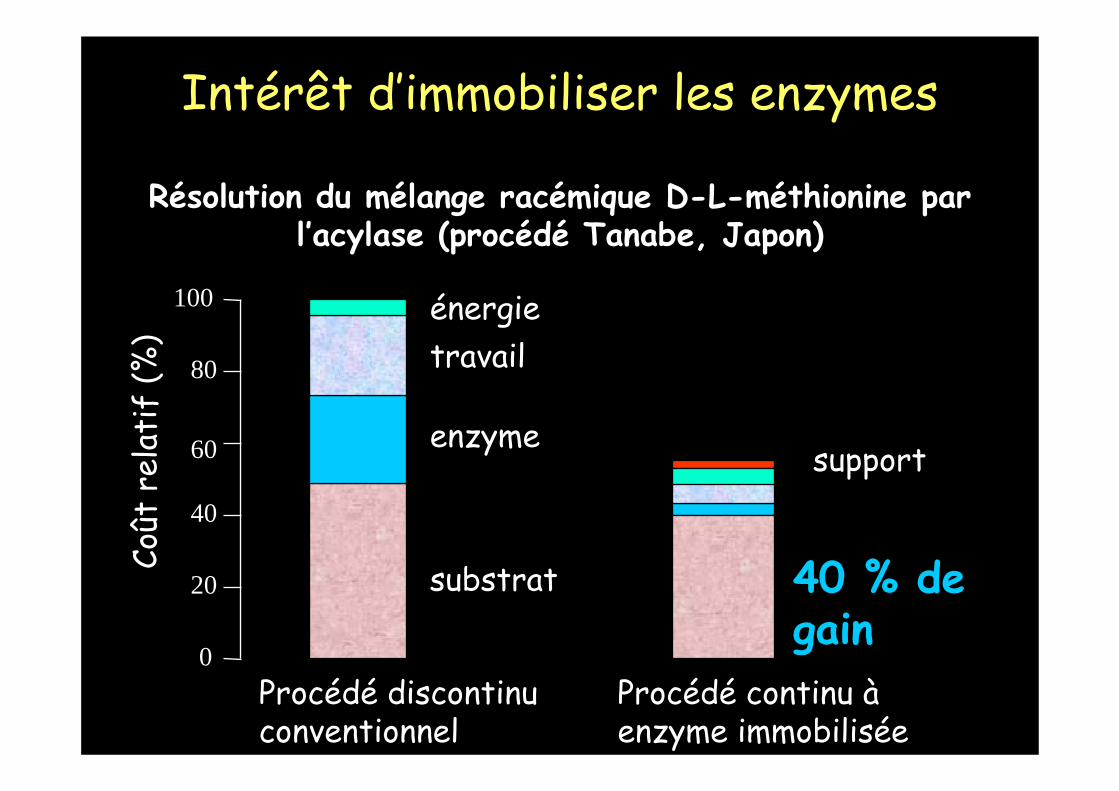
Résolution du mélange racémique D-L-méthionine par l’acylase (procédé Tanabe, Japon)
0
100
60
80
40
20
Coût
rel
atif
(%)
Procédé discontinu conventionnel
substrat
enzyme
travailénergie
40 % de gain
support
Procédé continu àenzyme immobilisée
Intérêt d’immobiliser les enzymes

Les enzymes sont retenues à l’intérieur d’une matrice poreuse
EE
E
E E
EE
Inclusion dans un gel
E
E
E
E
Inclusion dans une fibre
EE
E
EE
E
EE
E
Inclusion dans une micro-capsule
L’inclusion
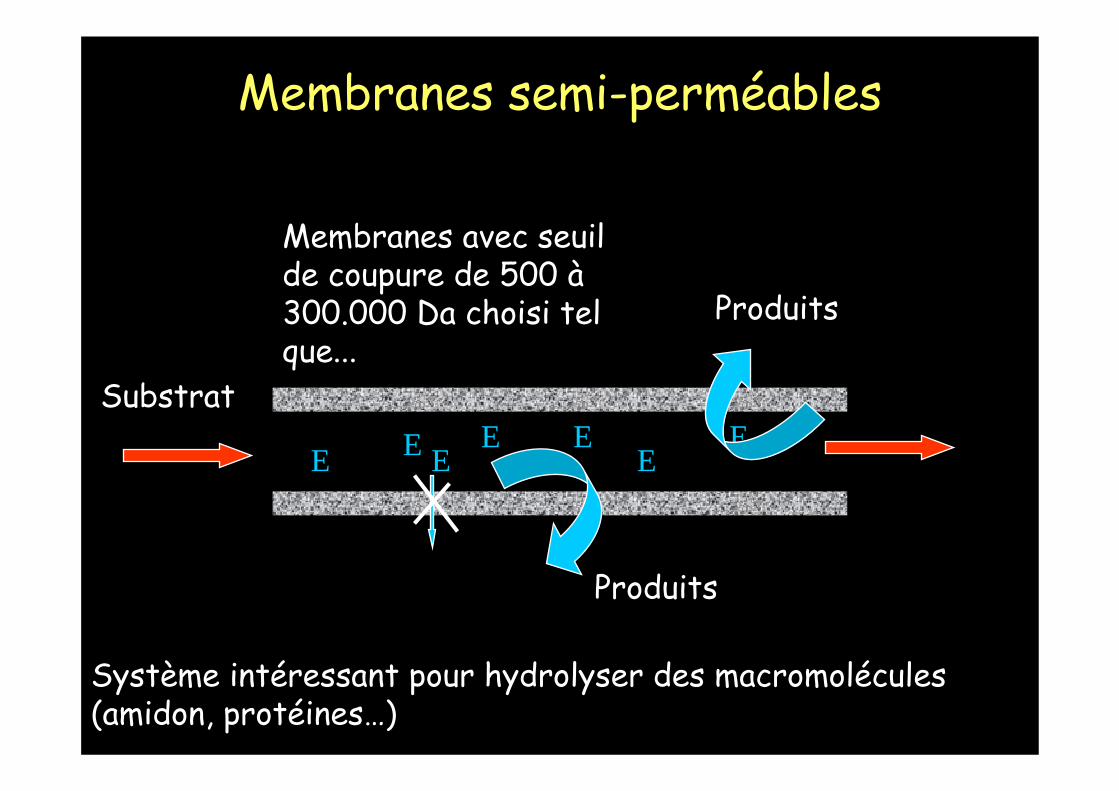
SubstratE E
EEE
EE
Membranes avec seuil de coupure de 500 à300.000 Da choisi tel que...
Produits
Produits
Système intéressant pour hydrolyser des macromolécules (amidon, protéines…)
Membranes semi-perméables

Les réactions de polymérisation ou de gélification sont bien maîtrisées
L’inclusion permet en une seule étape d’immobiliser la totalité de la masse d ’enzyme
Il n’existe aucun lien entre l’enzyme et le polymère : peu de risque de dénaturation
Applications à tout type d’enzymes
Les solvants utilisés et les valeurs de pH extrêmes peuvent s’avérer dénaturant.
Problèmes de diffusion au travers des gels.
Mauvaise tenue mécanique des gels.
Problèmes d’encombrement stérique.
L’inclusion

Les enzymes sont retenues à la surface d’un support insolublepar établissement d’interactions secondaires entre les groupesfonctionnels de l ’enzyme et du support.
+
Immobilisation d’enzymes par adsorption

interactions de Van der Waals :électrostatiques entre atomes ou molécules
interactions hydrophobesliaisons ioniques :
les résidus d’acides aminés basiques et acides sontionisés (+ et -) respectivement au voisinage de pH 7) d’où lapossibilité d’échange d’ions.
liaisons hydrogènes :polarisation des liaisons entre un atome d’hydrogène et
un atome plus électronégatif (O ou N)
Types d’interaction

Très grande facilité de mise en œuvre
Possibilité de régénérer le support par désorption et remplacement de l’enzyme.
- procédé Tanabe pour résolution du mélange racémique D,L méthionine par l’acylase : le support DEAE Sephadex a un prix non négligeable : même support utilisépendant 5 ans.
Risque important de désorption
Problèmes stériques
Mauvaise orientation de l’enzyme : perte d’activité
L’adsorption
X
X
XX
activation
A
A
AA
+
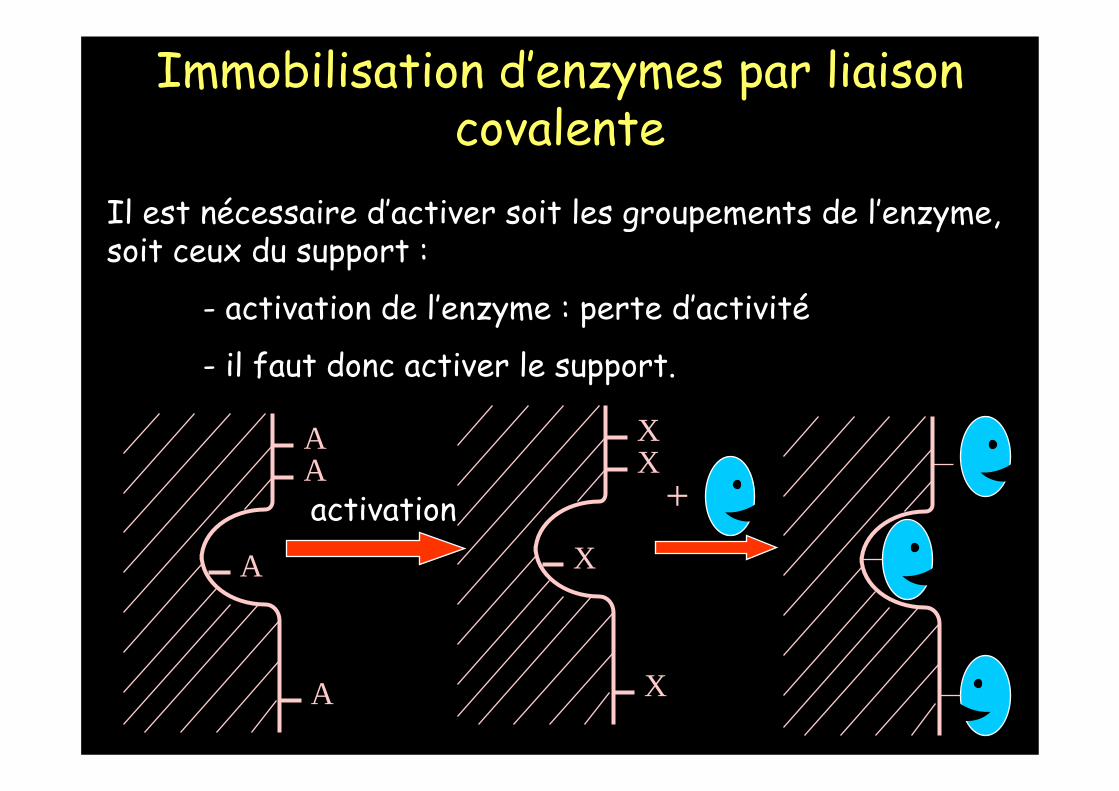
Immobilisation d’enzymes par liaison covalente
Il est nécessaire d’activer soit les groupements de l’enzyme, soit ceux du support :
- activation de l’enzyme : perte d’activité
- il faut donc activer le support.

Très grande solidité de la liaison enzyme-support
Grande variété de supports et de méthodes
Rigidification de la structure tridimensionnelle : meilleure stabilité
Complexité de la méthode.
Modification de la structure de l’enzyme : perte d’activité
Impossible de prévoir le rendement de greffage
Supports présentant de mauvaises propriétés mécaniques
La liaison covalente


























![[Philip R. Amidon] Philostorgius Church History ((Bookos.org)](https://img.pdfslide.net/doc/110x75/55cf9dab550346d033aea3e3/philip-r-amidon-philostorgius-church-history-bookosorg.jpg)


