Embed Size (px)
Citation preview

Sistemi za kontrolu kvaliteta proteina –
molekularni šaperoni i proteazom

Kako protein nakon sinteze postaje
funkcionalan? • Proces ekspresije gena nije završen prevođenjem
informacije sadržane u iRNK u redosled ak polipeptidnog lanca.
• Nekovalntne interakcije i kovalentne modifikacije su odgovorne za “sazrevanje” funkcionalnog proteina.

Kako protein nakon sinteze postaje
funkcionalan?
• Većina promena koje mora da pretrpi
novosintetisani polipeptid je uslovljena
nekovalentnim interakcijama:
– uvijanje u jedinstvenu
trodimenzionalnu konformaciju.
– vezivanje kofaktora neophodnih za
aktivnost.
– neretko, protein-protein interakcija
kojom ostvaruje svoju funkciju.

Kako protein nakon sinteze postaje
funkcionalan?
• Mnogi proteini moraju da pretrpe i post-translacione kovalentne modifikacije na tačno određenim ak.
• Najčešće kovalentne modifikacije su glikozilacija i fosforilacija, mada je poznato preko 100 različitih tipova kovalentnih modifikacija.
• Informacija neophodna za sve nabrojane korake maturacije funkcionalnog proteina sadržana je u redosladu ak polipeptidnog lanca koji se sintetiše na ribozomima.

Prostorno uvijanje proteina
je jedno od nerešenih pitanja
nauke

Ko-translaciono prostorno uvijanje
polipeptida • Proces prostornog uvijanja rastućeg polipeptida odvija
se uporedo sa njegovom sintezom.
• U trenutku kada se novosintetisani polipeptid oslobađa od ribozoma on stiče najveći deo svoje konformacije, ali ona nije finalna.

Ko-translaciono prostorno uvijanje
polipeptida • Tokom evolucije, sekvenca proteina nije bila selektovana
samo za konformaciju koju može da stekne već i za sposobnost brzog prostornog uvijanja u trenutku dok rastući polipeptid izlazi iz ribozoma.
• Levinathal paradox

Formiranje trodimenzionalne
strukture odvija se kroz dve faze

Formiranje trodimenzionalne
strukture odvija se kroz dve faze • Brza faza - Polipeptid stiče grubu tercijalnu strukturu još
tokom izlaska iz ribozoma.
• Za sintezu polipeptida prosečne veličine potrebno je nekoliko minuta, a u roku od nekoliko sekundi stiče kompaktnu strukturu koja sadrži većinu finalne sekundarne strukture ( helikse i ploče) uređene na način koji je grubo sličan finalnoj tercijalnoj konformaciji.

Izlivena globula
• Nastala otvorena i fleksibilna
struktura polipeptida naziva
se izlivena (eng. molten)
globula.
• U odnosu na finalnu
trodimenzionalnu strukturu
izlivena globula je otvorena i
manje uređena.

Formiranje trodimenzionalne
strukture odvija se kroz dve faze
• Izlivena globula je početna tačka za
relativno spor proces prostornog
podešavanja što za rezultat ima sticanje
finalne trodimenzionalne konformacije
proteina.
• Fino podešavanje bočnih ostataka ak
čime protein stiče finalnu prostornu
konformaciju predstavlja sporu fazu
formiranja trodimentionalne strukture
proteina.

Kontrola kvaliteta proteina
• Protein koji ima izložen hidrofobni region znatne veličine na svojoj površini je obično abnormalan.
• Hidrofobni regioni formiraju intermedijere - proteinske agregate – štetne za ćeliju.

Kontrola kvaliteta proteina
• Ćelija je razvila mehanizme za kontrolu kvalitata proteina koji prepoznaju i otklanjaju proteine sa izloženim hidrofobnim regionima.
• Više od 30% novosintetisanih polipeptidnih lanaca biva selektovano za brzu razgradnju kao rezultat delovanja sistema za kontrolu kvaliteta proteina.

Ćelijski mehanizmi za kontrolu
kvaliteta proteina • Neki proteini spontano stiču
pravilnu trodimenzionalnu konformaciju.
• Prostorno uvijanje mnogih proteina je efikasnije uz pomoć specijalne klase proteina nazvanih molekularni šaperoni.
• Kada pokušaji ponovnog uvijanja proteina ne uspeju nepravilno uvijeni proteini se potpuno razgrađuju u proteazomu.

Uloga molekularnih šaperona
a. Pomažu proteinima da se prostorno uviju.
b. Tokom procesa prostornog uvijanja štite proteine od protein protein interakcije sa drugim proteinima.
c. Odmotavaju proteinske agregate
d. Odmotavaju proteine koji su nepavilno uvijeni pre procesa njihove degradacije.
e. Transport proteina
f. Druge uloge

Molekularni šaperoni
• Proteini toplotnog šoka (hsp) čiji se nivo sinteze drastično povećava nakon kratkog izlaganja ćelije povišenim temperaturama (42C).
• Eukariotske ćelije poseduju dve glavne familije molekularnih šaperona koje pomažu proteinima da se prostorno uviju
• Hsp60 i hsp70
• Razlikuju se po mehanizmu delovanja i vremenskom periodu (u životu proteina) kada deluju.

Proteini hsp60 i hsp70
• Zajedničke osobine:
• Pokazuju afinitet za izložene hidrofobne
nizove pogrešno uvijenih proteina.
• Hidrolizuju ATP.
• Funkciju pomaganja proteinskog foldinga
ostvaruju sa malim setom asociranih
proteina (partner proteini).

Proteini hsp70
• Deluju rano u životu proteina i prepoznaju niz od
približno 7 hidrofobnih ak na površini proteina.
• Za ciljni protein se prvo vezuje pomoćni protein Hsp40, a
zatim monomer Hsp70 u kompleksu sa ATP-om (Hsp70-
ATP).

• Protein Hsp70 ima ATP-aznu aktivnost i vrši hidrolizu
ATP-a u ADP, podležući konformacionoj promeni koja
mu omogućava da se čvršće veže za ciljni protein.
• Faktor za razmenu nukleotida, protein GrpE, uklanja
nastali ADP, što uzrokuje da protein Hsp40 prvo napusti
formirani kompleks, a zatim disocira i Hsp70.
• Ponovljeni ciklusi vezivanja i oslobađanja proteina
Hsp40 i kompleksa Hsp70-ATP pružaju šansu ciljnom
polipeptidu da ponovo „proba“ da zauzme ispravnu
konformaciju.

Proteini hsp60
• Proteini hsp60 formiraju velike oligomerne strukture u
čiju izolovanu unutrašnjost ulazi ciljni protein. Poznati su
i kao šaperonini, a njihovi pomoćni proteini kao ko-
šaperonini.

Proteini hsp60
• Deluju kasnije u životu proteina i takođe prepoznaje
hidrofobne regione ak na površini proteina.
• Proteini ove familije formiraju oligomernu strukturu u
obliku bureta, koja u trenutku kada obavlja svoju funkciju
stupa u interakciju sa pomoćnim proteinom, čija je
struktura u obliku kape.
•
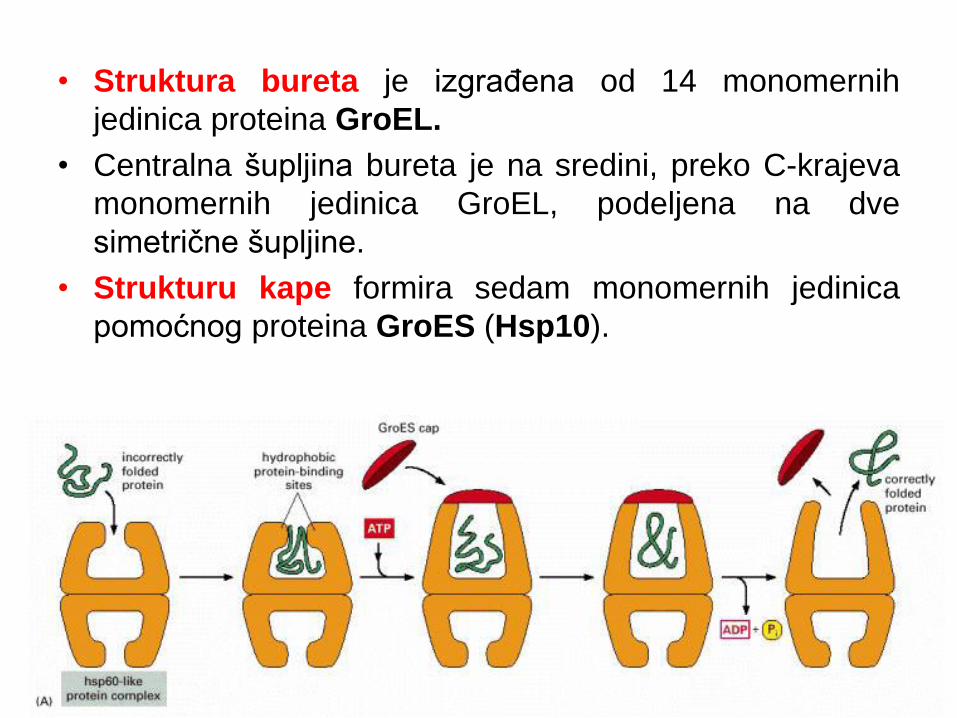
• Struktura bureta je izgrađena od 14 monomernih
jedinica proteina GroEL.
• Centralna šupljina bureta je na sredini, preko C-krajeva
monomernih jedinica GroEL, podeljena na dve
simetrične šupljine.
• Strukturu kape formira sedam monomernih jedinica
pomoćnog proteina GroES (Hsp10).

Proteini hsp60
• U svakom ciklusu samo jedna polovina simetričnog
bureta, odnosno jedan prsten, stupa u interakciju sa
ciljnim proteinom.
• Prsten koji je aktivan i asociran sa kapom označava se
kao proksimalni, dok se onaj koji je u tom trenutku
neaktivan označava kao distalni.

Razgradnja proteina
• Kada je protein određen za degradaciju, nema povratka što obezbedjuje da nema ni parcijalno degradovanih proteina koji bi mogli interferirati sa biološkim procesima.
• U ćeliji postoje dva glavna puta destrukcije proteina:
a) Razgradnja proteazama u lizozomima.
b) Proteoliza ubikvitin-proteaznim putem.

Proteazom – mašinerija za degradaciju
proteina
• Krajnja mašinerija za razgradnju proteina u eukariotskim ćelijama je proteazom.
• Proteazomi su ATP zavisni proteinski kompleks lokalizovani i u citoplazmi i u jedru.
• Proteazom se sastoji iz 20S centralnog šupljeg cilindra (jezgra proteazoma) čija su oba kraja asocirana sa 19S kapama.

Proteazom – mašinerija za degradaciju
proteina
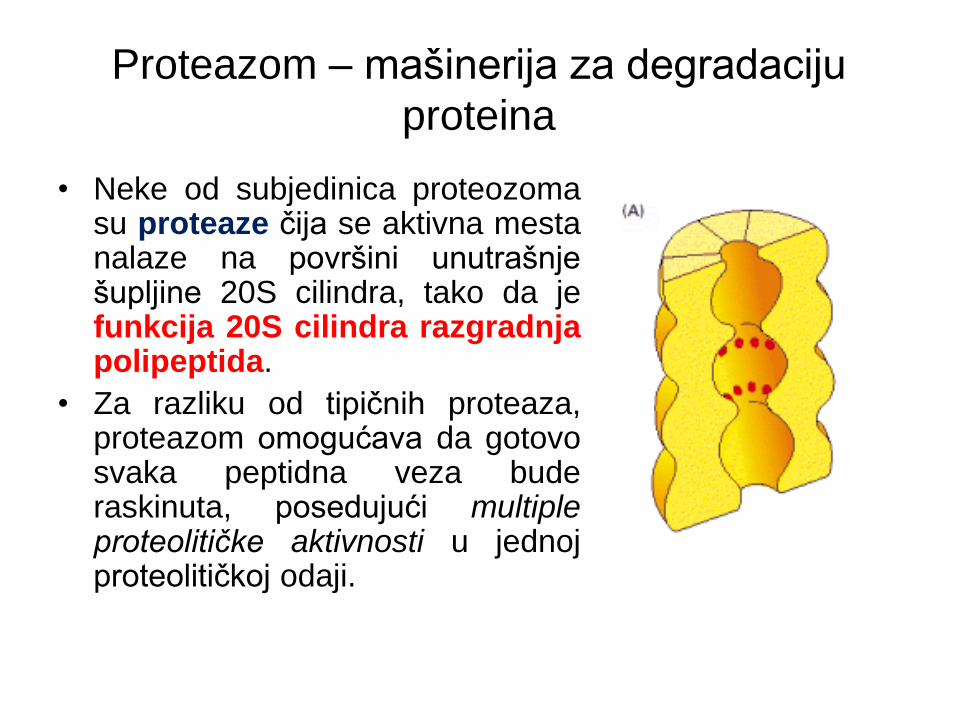
• Struktura 20S centralnog šupljeg cilindra je visoko
konzervirana od kvasca do čoveka i sastoji se od 4
međusobno naslojena prstena.
• Svaki prsten je sačinjen od 7 subjedinica koje se dele
u dve grupe: α subjedinice koje čine spoljašnja dva
prstena i β subjedinice unutrašnja dva prstena.
Proteazom – mašinerija za degradaciju
proteina
• Neke od subjedinica proteozoma su proteaze čija se aktivna mesta nalaze na površini unutrašnje šupljine 20S cilindra, tako da je funkcija 20S cilindra razgradnja polipeptida.
• Za razliku od tipičnih proteaza, proteazom omogućava da gotovo svaka peptidna veza bude raskinuta, posedujući multiple proteolitičke aktivnosti u jednoj proteolitičkoj odaji.

Proteazom – mašinerija za razgradnju
proteina
• 19S kape selektivno vezuje proteine markirane za razgradnju i funkcionišu kao regulatorne kapije na ulazu u unutrašnjost cilindra.
• ATP-azne subjedinice kape odmotavaju proteine koji treba da se razgrade i ubacuju ih u jezgro proteazoma.

Proteini se obeležavaju za
razgradnju
• Sa nekoliko izuzetaka, proteazom deluje na proteine koji
su obeleženi za razgradnju preko kovalentno vezanog
većeg broja kopija malog proteina (76 ak) koji se naziva
ubikvitin.
• Da bi se konjugovao sa ciljnim proteinom, ubikvitin prvo
mora biti aktiviran.

Proteini se obeležavaju za
razgradnju
• Ubikvitin se aktivira za konjugaciju sa ciljnim proteinima
preko vezivanja za ATP-zavisni proteina E1, koji se
naziva ubikvitin aktivirajući enzim.

Proteini se obeležavaju za
razgradnju
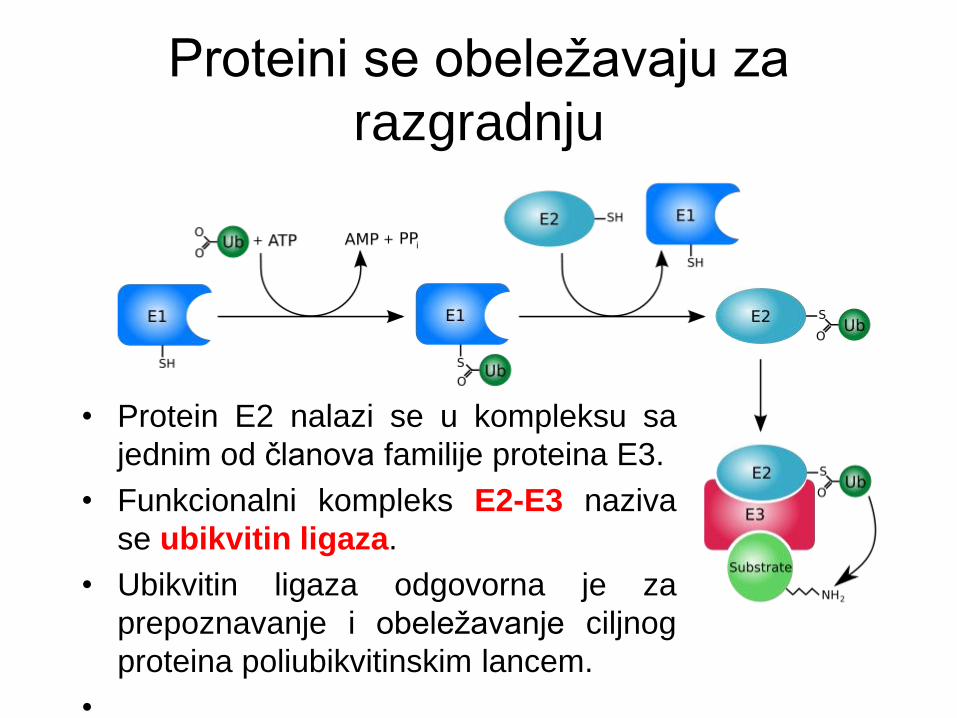
• Aktivirani ubikvitin se zatim prebacuje sa (cisteinskog
ostatka) proteina E1 na (cisteinski ostatak) proteina E2,
koji se naziva ubikvitin-konjugujući enzim.
Proteini se obeležavaju za
razgradnju
• Protein E2 nalazi se u kompleksu sa
jednim od članova familije proteina E3.
• Funkcionalni kompleks E2-E3 naziva
se ubikvitin ligaza.
• Ubikvitin ligaza odgovorna je za
prepoznavanje i obeležavanje ciljnog
proteina poliubikvitinskim lancem.
•

Proteini se obeležavaju za
razgradnju
• Jednom kada je proteinski substrat mono-ubikvitiran,
poliubikvitinski lanac se formira kroz iste ubikvitin
konjugacione kaskade, pri čemu je karboksilna grupa
ubikvitina kovalentno vezana za Lys reziduu ubikvitina
koji je već konjugovan za proteinski substrat.
• Multiubikvitinski lanac na ciljnom proteinu je ono što
prepoznaju kape proteazoma.

Ubikvitin aktivirajući enzim E1 ili UBA
• Jedan enzim E1 je odgovoran za aktivaciju ubikvitina.
• Kod kvasaca je samo jedan funkcionalni enzim UBA,
UBA1.
• Delecija gena UBA1 kod kvasca je letalna.

Ubikvitin konjugujući enzim E2 ili UBC
• Svi enzimi E2 (a njih je na desetine, najmanje 13 E2
kod kvasca, a oko 30 kod sisara) sadrže konzervirano
oko 130 ak dugačko katalitičko jezgro, domen UBC.

Ubikvitin ligaze E3
• E3 ligaze odnosno kompleksi ligaza prepoznaju
specifične motive substrata i katalizuju transfer ubikvitina
na proteinski substrat.
• U ćelijama sisara postoji nekoliko stotina različitih proteina E3, koji formiraju komplekse sa specifičnim proteinima E2.

Ubikvitin-proteazomski sistem
• Ubikvitin-proteozomski sistem sastoji se od mnogo
različitih proteolitičkih puteva, koji imaju protein E1 na
početku i proteazom na kraju puta, a razlikuju se u E2-
E3 ubikvitin ligazama i pomoćnim faktorima.
• Različite ubikvitin-ligaze prepoznaju različite signale za
razgradnju i time deluju na različite proteine.

Uloge ubikvitin-proteozomski
sistem • Prepoznaju i eliminišu proteine koji
imaju pogrešnu trodimenzionalnu strukturu ili druge abnormalnosti.
• Postoje i proteini koji se sve vreme brzo degradaju. To su kratkoživeći proteini čija se kontrola funkcije ostvaruje preko regulacije njihove razgradnje.
• Uslovno kratkoživeći proteini su proteini koji su metabolički stabilni pod određenim uslovima, dok su pod drugim uslovima nestabilni.

Kontrola regulisane razgradnje
proteina • U ćeliji postoji čitav niz mehanizama kojima se kontroliše
regulisana razgradanja proteina,
• Dva opšta puta indukovane degradacije proteina:
• Aktivacija kompleksa ubikvitin ligaze (E2-E3).
• Aktivacija signala za razgradnju.

Regulisana razgradnja proteina preko
aktivacije ubikvitin ligaze
• Aktivacija E2-E3 kompleksa ostvaruje se:
– Fosforilacijom specifičnog ak ostatka.
– Alosteričkom aktivacijom vezivanjem nekog malog molekula (liganda) ili dodavanjem proteinske subjedinice.

Regulisana razgradnja proteina preko
aktivacije signala za razgradnju
• Načini aktivacije signala za degradaciju su različiti:
• Fosforilacijom se otkriva “sakriven” signal za degradaciju.
• Regulisanom disocijacijom proteinske subjedinice.
• Kidanjem određene peptidne veze, čime se formira novi N-kraj kojeg specifičan protein E3 prepoznaje kao destabilišući N-kraj (signal za razgradnju).

Kontrola proteina preko regulacije
njihove razgradnje
• Proteazom, procesivna
mašinerija za degradaciju
proteina je odgovoran za
različite unutarćelijske
aktivnosti:
• Uklanjanja pogrešno savijenih
proteina;
• Regulacije ćelijskog ciklusa
razgradnjom ciklina;
• U stečenom imunskom
odgovoru obradom peptida
antigena, itd.





























